
Serum Granulocyte Colony Stimulating Factor and Alzheimer's Disease

Epidermal Overexpression of Granulocyte-MacrophageColony-Stimulating Factor Induces Both KeratinocyteProliferation and Apoptosis1
Kai Breuhahn,2 Amrit Mann,2 Gabriele Muller,Arnd Wilhelmi, Peter Schirmacher, Alexander Enk,and Manfred Blessing3
SFB-432, I. Medical Department [K. B., A. M., M. B.], Department ofDermatology [G. M., A. E.], Boehringer Ingelheim Research Group, SFB-311, I. Medical Department [A. W., M. B.], and Institute of Pathology[P. S.], Johannes Gutenberg University, D-55131 Mainz, Germany
AbstractGranulocyte-macrophage colony-stimulating factor (GM-CSF) is released by keratinocytes in sizeable amountsonly under pathological conditions, e.g., after topicalapplication of a tumor promoter, in atopic dermatitis (AD),and after wounding. To study the biological function ofthis cytokine release, we generated transgenic mice thatconstitutively overexpress GM-CSF in the epidermis. Anincrease in the numbers of mast cells and Langerhanscells (LCs) in transgenics versus nontransgenic controlswas observed but no severe inflammation. This isconsistent with a central role of this cytokine in thedevelopment and maturation of LCs. Mitotic activity inthe epidermis of transgenic mice was elevated, butepidermal thickness and differentiation were normal.Homeostasis is maintained by an increase of apoptosis inthe epidermis. We describe the differential expression ofregulators of apoptosis and discuss a potentialmechanism for this novel proapoptotic activity of GM-CSF on keratinocytes. Both stimulation of proliferationand promotion of apoptosis are of great relevance totumorigenesis. The latter may be a means of removingdamaged cells after genotoxic stress or injury.
IntroductionGM-CSF4 is a cytokine regulating proliferation, differentia-tion, and survival of hematopoetic cells (1–3). Besides he-
matopoetic cells, other cell types including keratinocyteshave been identified, both as sources and as targets forGM-CSF (4–7). The release of this cytokine by keratinocytesis triggered by disturbances in skin homeostasis such astumor promotion or wounding (4–6, 8). Because GM-CSFhas been shown to stimulate keratinocyte proliferation aswell as immune cell proliferation and maturation, it had beenproposed to mediate both the proliferative burst in keratino-cytes as well as inflammation after disturbances of the ho-meostasis in the skin. In previous studies, keratinocyte hy-perplasia and hyperproliferation were induced by GM-CSFand accompanied by the induction of the stress keratin pair6/16, indicative of the alternative or regenerative pathway ofkeratinocyte differentiation (5, 9, 10). In the dermis, neutro-phil and macrophage infiltration and fibrosis were observed(10). However, because these studies involved the applica-tion of GM-CSF by injections or the infection with a replica-tion-deficient adenoviral vector, it is not clear whether theseeffects resulted from GM-CSF alone or from synergistic ef-fects with these secondary stimuli. Also, the significance ofkeratinocyte-derived GM-CSF could not be addressed inthese studies because they involved either systemic levels ofGM-CSF or intradermal expression of this cytokine (5, 10).
In regard to the skin, GM-CSF is also particularly importantfor the generation and maturation of epidermal LCs as well asthe survival of macrophages (11, 12). In this context, a causalrole of GM-CSF overproduction in the establishment andchronicity of lesions in AD patients has been discussed (7,12). In addition to increased tissue levels of IL-3, IL-4, IL-5,IL-10, and IL-13, AD lesions display highly elevated levels ofGM-CSF and an increase in dendritic cells as well as infil-tration of T cells, macrophages, and monocytes (13).
Besides proliferation and differentiation, apoptosis is an-other critical parameter affecting homeostasis and tumori-genesis. Apoptosis occurs at high frequencies during certainphases of the hair cycle in follicular cells as well as in inter-follicular keratinocytes in response to genotoxic stress (14,15). The latter may be a mechanism to remove geneticallydamaged cells and therefore prevent the formation of malig-nancies (14). GM-CSF has been shown to prevent apoptosisin many immune cells. This activity is mediated through theinduction of antiapoptotic factors like Bcl-2 via the b-chain ofthe GM-CSF receptor (3, 12, 16). However, there are noreports on the effects of GM-CSF on keratinocyte apoptosisin vivo.
Apoptosis is a complex event and is tightly regulated.Members of the Bcl-2 family of proteins are crucial factorsregulating programmed cell death. Among those are theantiapoptotic factors Bcl-2, Bcl-xL, and A1 and the proapop-totic factors Bcl-xs, Bad, Bak, and Bax (17–20). Apoptosis ispromoted or inhibited by the formation of specific ho-
Received 9/15/99; revised 12/3/99; accepted 1/5/00.The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indi-cate this fact.1 This work was funded by Grant SFB-432/B1 from the Deutsche Forsch-ungsgemeinschaft and a grant from the Boehringer Ingelheim Foundation.2 These two authors contributed equally to this work.3 To whom requests for reprints should be addressed, at Johannes Gu-tenberg University, SFB-311, I. Medical Department, Obere ZahlbacherStrasse 63, D-55131 Mainz, Germany. Phone: 49-6131-3933357; Fax:49-6131-3933364; E-mail: [email protected] The abbreviations used are: GM-CSF, granulocyte-macrophage colony-stimulating factor; LC, Langerhans cell; AD, atopic dermatitis; IL, interleu-kin; RT-PCR, reverse transcription-PCR; wt, wild type; BrdUrd, bromode-oxyuridine; TUNEL, terminal deoxynucleotidyltransferase-mediated nickend labeling; FACS, fluorescence-activated cell sorter; BMP, bone mor-phogenic protein.
111Vol. 11, 111–121, February 2000 Cell Growth & Differentiation

modimers or heterodimers between family members (17–19).Thus, the balance between different members of the Bcl-2family controls cell survival and cell death. Expression ofBcl-2 in the epidermis is restricted to the basal layer and hairfollicles (21, 22). Conversely, Bcl-xL is predominantly seen insuprabasal layers, whereas Bax is weakly expressed in alllayers (23, 24). Strong epidermal expression has been re-ported recently for Bak (25). In addition to these intracellularregulators of programmed cell death, keratinocytes havealso been shown to express Fas, a major receptor for apop-totic signals. Moreover, expression of Fas is strongly up-regulated in keratinocytes upon treatment with IFN-g. Thus,besides differential expression of members of the Bcl-2family, regulation of Fas expression via IFN-g may be amajor mechanism for controlling apoptosis in keratinocytes(26, 27).
To analyze the effect of constitutive release of GM-CSF bykeratinocytes without additional stimuli on skin architectureas well as keratinocyte proliferation, differentiation, and ap-optosis, we generated transgenic mice expressing GM-CSFin the skin under the control of a keratin 5-based expressionvector. We observed an increase in the number of LCs andmast cells in the skin of these transgenics that is compatiblewith the proposed role of GM-CSF in the perpetuation andchronicity of AD lesions. However, no significant infiltrationby other inflammatory cells, such as T cells or macrophages,was seen. Both mitotic and apoptotic frequencies were dou-bled for basal interfollicular keratinocytes in these mice, re-sulting in the maintenance of regular epidermal thicknessand the standard pathway of keratinocyte differentiation.This novel proapoptotic activity of GM-CSF may be medi-ated through an induction of Fas by IFN-g and changes in theexpression of Bcl-2 family members. Apoptosis induced byGM-CSF release from keratinocytes may be a means toeliminate damaged keratinocytes after genotoxic stress.
ResultsGeneration of Transgenic Mice. Transgenic mice weregenerated by pronuclear injection of the keratin 5-basedexpression vector for GM-CSF (Fig. 1). Three independenttransgenic founder animals with autosomal integration of thetransgene were obtained. All were fertile and producedtransgenic offspring at the expected Mendelian frequency.The resulting transgenic lines were designated according tothe guidelines set by the Institute for Laboratory Animal
Research (Washington, DC) as TgN(K5mGMCSF)1 Mbl,TgN(K5mGMCSF)2 Mbl, and TgN(K5mGMCSF)3 Mbl. Forsimplicity, these lines are being abbreviated hereafter as Tg1,Tg2, and Tg3, respectively. Animals of all three lines dis-played no obvious gross abnormalities. The density of hairfollicles and the outer appearance of skin and fur were nor-mal. A deficit in total body weight of ;10% in comparison tocontrols was noted during the first 2–3 months of the lifespan. However, this difference vanished at later time points,and neither life expectancy nor fertility was compromised.
Expression of the Transgene. Tissue specificity of theexpression of the transgene was determined by RT-PCRanalysis using RNAs extracted from various tissues as tem-plates. Expression of the GM-CSF transgene was demon-strated in the skin of both lines Tg1 and Tg2 (Fig. 2). Addi-tionally, a weak signal was obtained in heart of line Tg2 (Fig.2B). Expression of the transgene was undetectable by thismethod in kidney, lung including trachea, liver, brain, andspleen in both transgenic lines.
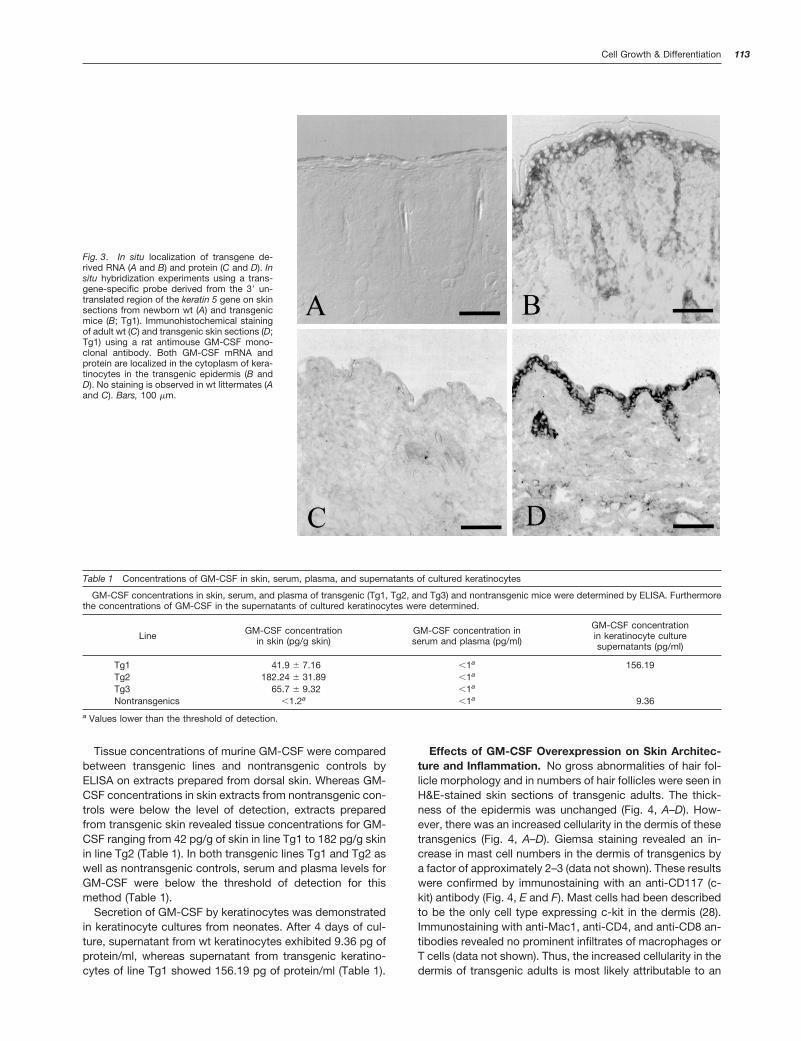
Cell type-specific expression of the GM-CSF transgene inskin was verified by in situ hybridization experiments andimmunohistochemistry on skin sections (Fig. 3). In situ hy-bridization experiments using a transgene-specific antisensecRNA probe revealed an accumulation of transgene-derivedtranscripts in interfollicular keratinocytes as well as follicularcells in the skin of transgenic neonates, whereas no specificsignal was seen in skin sections from nontransgenic controlneonates (Fig. 3, A and B). The corresponding sense cRNAprobe produced no specific signals in skin sections of controlmice or of transgenic mice (data not shown). By immunohis-tochemistry using a GM-CSF-specific antibody, no signifi-cant staining was observed in normal adult skin (Fig. 3C). Bycontrast, strong cytoplasmic staining was observed in inter-follicular keratinocytes as well as follicular cells in the skin oftransgenic adults (Fig. 3D). Control experiments with sec-ondary antibody alone resulted in no staining in skin sectionsof control mice or of transgenic mice (data not shown).
Fig. 1. Expression vector for GM-CSF. A cDNA coding for murine GM-CSF was brought under the control of the bovine keratin 5 promoter.Downstream of the cDNA the splice and polyadenylation signals derivedfrom the bovine keratin 5 gene were included. B, BamHI; E, EcoRI; S, SalI;X, XhoI.
Fig. 2. Expression of the transgene-derived RNA. RT-PCR analysis withtotal RNAs from kidney (Lanes 1), lung (Lanes 2), heart (Lanes 3), liver(Lanes 4), skin (Lanes 5), brain (Lanes 6), and spleen (Lanes 7) wasperformed. Lane 8 (A) shows genomic DNA from transgenic mice con-taining an intron sequence. Mice of line Tg1 (A) expressed the transgeneonly in the skin (A, Lane 5), whereas Tg2 transgenics (B) showed a signalin skin (B, Lane 5) and also in the heart (B, Lane 3). Nontransgenic miceexhibit no expression of transgene-specific transcripts (data not shown).BMP-6 was used as a control for the RT-PCR reaction (A and B).
112 GM-CSF Transgenic Mice
Tissue concentrations of murine GM-CSF were comparedbetween transgenic lines and nontransgenic controls byELISA on extracts prepared from dorsal skin. Whereas GM-CSF concentrations in skin extracts from nontransgenic con-trols were below the level of detection, extracts preparedfrom transgenic skin revealed tissue concentrations for GM-CSF ranging from 42 pg/g of skin in line Tg1 to 182 pg/g skinin line Tg2 (Table 1). In both transgenic lines Tg1 and Tg2 aswell as nontransgenic controls, serum and plasma levels forGM-CSF were below the threshold of detection for thismethod (Table 1).
Secretion of GM-CSF by keratinocytes was demonstratedin keratinocyte cultures from neonates. After 4 days of cul-ture, supernatant from wt keratinocytes exhibited 9.36 pg ofprotein/ml, whereas supernatant from transgenic keratino-cytes of line Tg1 showed 156.19 pg of protein/ml (Table 1).
Effects of GM-CSF Overexpression on Skin Architec-ture and Inflammation. No gross abnormalities of hair fol-licle morphology and in numbers of hair follicles were seen inH&E-stained skin sections of transgenic adults. The thick-ness of the epidermis was unchanged (Fig. 4, A–D). How-ever, there was an increased cellularity in the dermis of thesetransgenics (Fig. 4, A–D). Giemsa staining revealed an in-crease in mast cell numbers in the dermis of transgenics bya factor of approximately 2–3 (data not shown). These resultswere confirmed by immunostaining with an anti-CD117 (c-kit) antibody (Fig. 4, E and F). Mast cells had been describedto be the only cell type expressing c-kit in the dermis (28).Immunostaining with anti-Mac1, anti-CD4, and anti-CD8 an-tibodies revealed no prominent infiltrates of macrophages orT cells (data not shown). Thus, the increased cellularity in thedermis of transgenic adults is most likely attributable to an
Fig. 3. In situ localization of transgene de-rived RNA (A and B) and protein (C and D). Insitu hybridization experiments using a trans-gene-specific probe derived from the 39 un-translated region of the keratin 5 gene on skinsections from newborn wt (A) and transgenicmice (B; Tg1). Immunohistochemical stainingof adult wt (C) and transgenic skin sections (D;Tg1) using a rat antimouse GM-CSF mono-clonal antibody. Both GM-CSF mRNA andprotein are localized in the cytoplasm of kera-tinocytes in the transgenic epidermis (B andD). No staining is observed in wt littermates (Aand C). Bars, 100 mm.
Table 1 Concentrations of GM-CSF in skin, serum, plasma, and supernatants of cultured keratinocytes
GM-CSF concentrations in skin, serum, and plasma of transgenic (Tg1, Tg2, and Tg3) and nontransgenic mice were determined by ELISA. Furthermorethe concentrations of GM-CSF in the supernatants of cultured keratinocytes were determined.
Line GM-CSF concentrationin skin (pg/g skin)
GM-CSF concentration inserum and plasma (pg/ml)
GM-CSF concentrationin keratinocyte culturesupernatants (pg/ml)
Tg1 41.9 6 7.16 ,1a 156.19Tg2 182.24 6 31.89 ,1a
Tg3 65.7 6 9.32 ,1a
Nontransgenics ,1.2a ,1a 9.36
a Values lower than the threshold of detection.
113Cell Growth & Differentiation
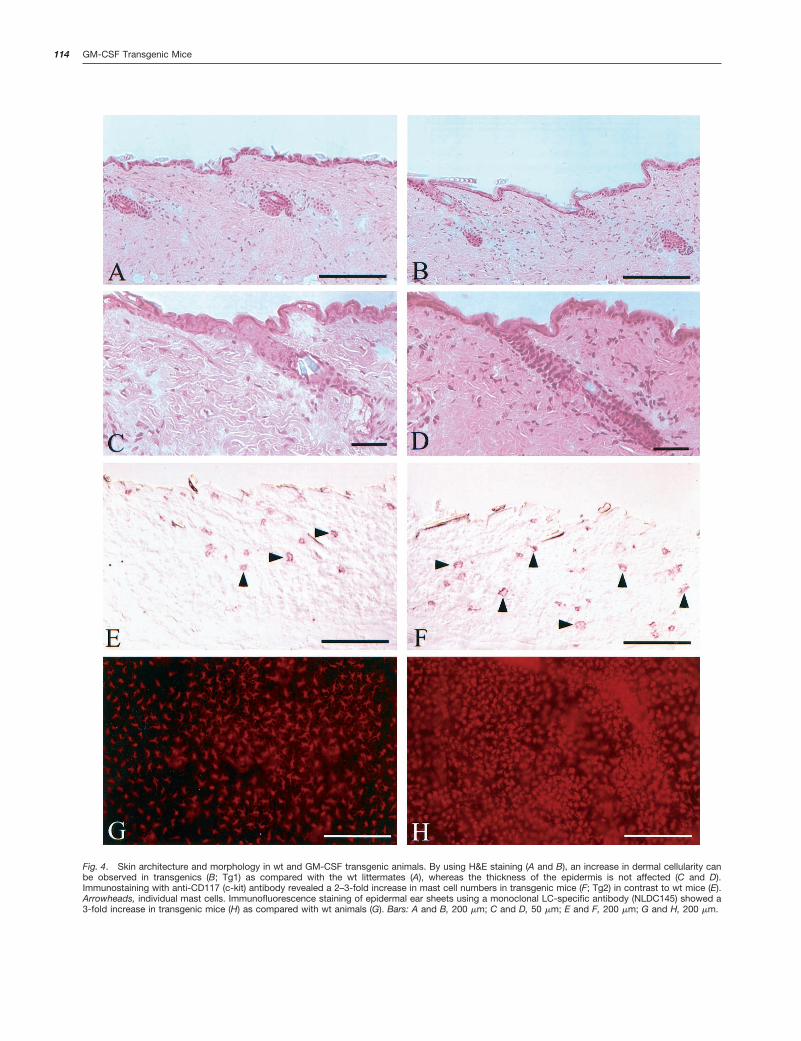
Fig. 4. Skin architecture and morphology in wt and GM-CSF transgenic animals. By using H&E staining (A and B), an increase in dermal cellularity canbe observed in transgenics (B; Tg1) as compared with the wt littermates (A), whereas the thickness of the epidermis is not affected (C and D).Immunostaining with anti-CD117 (c-kit) antibody revealed a 2–3-fold increase in mast cell numbers in transgenic mice (F; Tg2) in contrast to wt mice (E).Arrowheads, individual mast cells. Immunofluorescence staining of epidermal ear sheets using a monoclonal LC-specific antibody (NLDC145) showed a3-fold increase in transgenic mice (H) as compared with wt animals (G). Bars: A and B, 200 mm; C and D, 50 mm; E and F, 200 mm; G and H, 200 mm.
114 GM-CSF Transgenic Mice

increased number of resident dermal cells, such as fibro-blasts, and increased numbers of mast cells.
Because GM-CSF had been described as a factor promot-ing generation and maturation of epidermal LCs, we pre-pared epidermal sheets from the ears of normal controls andtransgenic mice of lines Tg1 and Tg2. Immunostaining withanti-NLDC145 antibody, a dendritic cell-specific marker (29,30), revealed an increase in LCs in the epidermal sheetsprepared from the two transgenic lines in comparison tonontransgenic controls (Fig. 4, G and H). Although it hadbeen described previously that injection of GM-CSF in themouse skin evokes inflammation, a robust inflammatory in-filtrate was not seen in transgenic skin. However, a differencein the cytokine milieu was noted. Extracts from transgenicdorsal skin revealed an increase in the tissue concentrationof IFN-g as compared with nontransgenic controls (Table 2).By contrast, IL-12 and IL-1b levels were unchanged, pointingtoward a nonactivated state of the epidermal LCs (Table 2).
Effects of GM-CSF Overexpression on Proliferation,Differentiation, and Apoptosis of Keratinocytes. GM-CSF had been reported to stimulate keratinocyte prolifer-ation in culture (4, 31). To investigate whether GM-CSFpromotes keratinocyte proliferation in vivo, we performedBrdUrd labeling experiments and determined the epider-mal mitotic indices in two transgenic lines, Tg1 and Tg2,as well as in nontransgenic control littermates. To identifybasal keratinocytes, we counterstained the specimenswith an anti-keratin 5 antibody. BrdUrd labeling experi-ments revealed an increase in numbers of S-phase nucleiin interfollicular keratinocytes of GM-CSF overexpressingtransgenic mice (Fig. 5, A and B). Whereas two S-phasenuclei per 100 total cells were found in the basal layer ofcontrol epidermis, four and six labeled nuclei per 100 totalcells were found in the basal layers of mice of lines Tg1and Tg2, respectively (Table 3).
Apoptosis in the epidermis was analyzed by double label-ing experiments using the TUNEL assay for the detection ofapoptotic cells and an anti-keratin 5 staining for the identi-fication of keratinocytes. An increase of stained apoptotickeratinocytes in the epidermis of transgenic mice versusnontransgenic controls was noticed (Fig. 5, C and D). Forquantification, apoptotic keratinocytes only in the basal layerof interfollicular epidermis were counted because the normalprocess of keratinocyte differentiation that takes place in
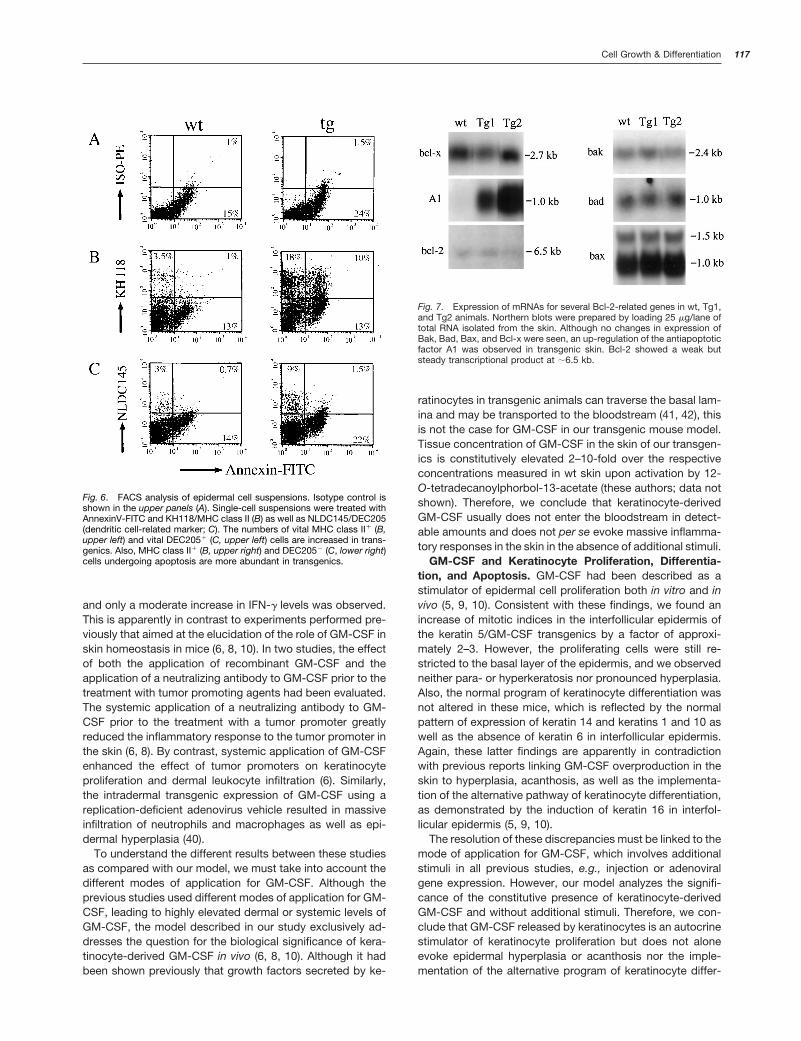
suprabasal epidermal layers bears some resemblance tocertain apoptotic changes. We found that the frequency ofapoptotic basal keratinocytes in normal interfollicular epider-mis was 0.31 per 100 keratinocytes (Table 3). By contrast,this frequency is doubled in transgenic mice to 0.66 apop-totic keratinocytes per 100 keratinocytes (Table 3). The in-crease of apoptosis in keratinocytes was corroborated bydouble-label FACS analysis on epidermal cells stained withannexin-V and a LC-specific antibody (NLDC145) as well asan antibody to MHC class II (KH118) molecules (Fig. 6). Thereis an increase in transgenics versus nontransgenics of MHCclass II-positive, nondendritic epidermal cells undergoingapoptosis (Fig. 6, B and C). These cells are most likelykeratinocytes, because up-regulation of MHC class II mole-cules had been shown on keratinocytes from these trans-genic mice.5
To see how this increase in the frequency of basal kerati-nocyte apoptosis is conveyed, we analyzed the expressionof several genes relevant to apoptosis and cell survival byNorthern blotting experiments using RNAs extracted fromcontrol skin and skin from mice of lines Tg1 and Tg2 (Fig. 7).To exclude the influence of the hair cycle on the expressionof proapoptotic or antiapoptotic genes, RNA samples wereprepared from age-matched animals. No changes were ob-served for the factors Bcl-2, Bcl-x, Bak, Bax, and Bad at theRNA level. However, a strong increase in transgenics versusnontransgenics was seen for the antiapoptotic factor A1 (Fig.7). 18S and 28S RNA as well as b-actin were used as loadingcontrols. No differences were observed in intensity (data notshown).
Analysis of keratinocyte differentiation markers as keratins1, 10, and 14 and especially of the hyperproliferative markerkeratin 6 in interfollicular epidermis by immunohistochemis-try and Western blotting revealed no differences in expres-sion of these markers between transgenic epidermis andnontransgenic control epidermis (data not shown).
DiscussionGM-CSF is a cytokine that is normally not released by kera-tinocytes. However, under pathological conditions such asafter wounding, during tumor promotion, or in inflammatoryskin diseases such as AD, keratinocytes secrete GM-CSF (4,7, 8, 31). GM-CSF has been described to affect a broadspectrum of different cell types including T cells, monocytes,neutrophils, eosinophils, epidermal LCs, as well as melano-cytes and keratinocytes (4, 5, 11, 12, 32, 33). Therefore, thecontribution of keratinocyte-derived GM-CSF to these path-ological conditions appears to be complex. To investigatethe effect of GM-CSF release by keratinocytes in an in vivosituation, we generated transgenic mice overexpressing mu-rine GM-CSF in the epidermis under the control of a consti-tutively active keratin 5 promoter-based expression vector(34).
5 G. Muller, M. Blessing, A. Mann, K. Breuhahn, K. Steinbrink, C. Szalma,J. Bergmann, J. Knop, and A. E. Enk. Mice overexpressing GM-CSF inskin are resistant to tolerance induction by UV-light or low-zone applica-tion of contact allergens, manuscript in preparation.
Table 2 Cytokine concentrations in skin
IFN-g, IL-12, and IL-1b concentrations in skin extracts of wt and trans-genic mice were determined by ELISA. An increase of IFN-g level wasobserved in transgenic mice as compared with wt animals. IL-1b concen-trations were high in skin extracts but revealed no significant differencesbetween wt and transgenic animals. The mean values and SD from threeto six animals are figured.
Line
IFN-gconcentration
in skin(pg/g skin)
IL-12concentration
in skin(pg/g skin)
IL-1bconcentration
in skin(pg/g skin)
Tg1 82.79 6 2.35 12.92 6 11.48 232.13 6 23.7Tg2 97.29 6 19.79 11.64 6 10.18 210.75 6 17.28Nontransgenics 41.28 6 9.26 11.28 6 9.29 237.84 6 8.17
115Cell Growth & Differentiation

Phenotype and Skin Architecture of Keratin 5/GM-CSFTransgenic Mice. The transgenic mice were healthy andfertile and displayed elevated levels of GM-CSF in the skin.Secretion of this cytokine by keratinocytes was demon-strated in culture experiments. The bioactivity of keratino-cyte-derived GM-CSF had been demonstrated previously(4). These transgenics did not develop accumulations ofmacrophages, blindness, or a fatal syndrome of tissue dam-age that had been described previously in transgenic micecarrying multiple copies of the GM-CSF gene (35). This dif-ference probably results from the local restriction of GM-CSF
overproduction in our transgenics, which do not show ele-vated serum levels for this cytokine. By contrast, the micecarrying multiple copies of the GM-CSF gene do exhibitmeasurable levels of GM-CSF in the serum, peritoneal cav-ities, and pleural cavities, thereby evoking systemic effects(36, 37). Histological examination as well as immunohisto-chemistry of skin sections revealed no gross abnormalities inthe skin of our transgenics. We did not observe significantepidermal thickening or hyperplasia. There was an increasein epidermal LC numbers and dermal mast cells. The formerconfirms the bioactivity of keratinocyte-derived GM-CSF be-cause it is consistent with the already described effects ofGM-CSF on the generation and maturation of cells belongingto the dendritic cell lineages and may also contribute to thechronicity of lesions in AD (2, 7, 11, 38). An increase in mastcell numbers has also been described in lesional skin of ADpatients (39). However, because the GM-CSF transgenicmice do not spontaneously develop AD-like lesions, otherfactors must be crucial to the development of this disease.
Besides the increased number of mast cells, no prominentinflammatory reaction was observed. There was no infiltra-tion of T cells, macrophages, neutrophils, or eosinophils inthe dermal compartment of our transgenics. Consistent withthese findings, there was no elevation of tissue levels forIL-1b, IL-2, IL-4, and IL-12 in the skin of these transgenics,
Fig. 5. Epidermal proliferation and apoptosis in the skin of adult transgenic mice (Tg1) as determined by BrdUrd labeling experiments (A and B) and TUNELstaining (C and D). S-phase nuclei in the basal layer of the epidermis of wt (A) and transgenic littermates (B) were analyzed and counted using fluorescencemicroscopy. Arrowheads, S-phase nuclei. Apoptosis in wt and transgenic animals was assessed in both control (C) and transgenic (D; Tg1) mice using theFITC-tagged TUNEL method. Arrowheads, apoptotic cells in the basal layer. A murine anti-keratin 14 antibody and a CyTM3-labeled secondary antibodywere used for counterstaining the basal layer of the epidermis in both cases. Bars, 200 mm.
Table 3 Proliferative and apoptotic frequencies
Proliferative and apoptotic frequencies in interfollicular keratinocytes ofwt and transgenic littermates are shown. For the quantification of mitoticindices, at least 3000 cells in the basal layer of the epidermis were countedwith a minimum of three animals from each line. For the determination ofapoptotic frequencies, a minimum of 1600 basal cells with specimensfrom at least three animals were counted after TUNEL reaction and anti-keratin 14 staining (P , 0.05).
Line Mitotic indices, labelednuclei per 100 cells
Apoptotic cells, labelednuclei per 100 nuclei
Tg1 4.04 6 0.36 0.67Tg2 6.10 6 1.55 0.66Nontransgenics 2.15 6 0.32 0.31
116 GM-CSF Transgenic Mice
and only a moderate increase in IFN-g levels was observed.This is apparently in contrast to experiments performed pre-viously that aimed at the elucidation of the role of GM-CSF inskin homeostasis in mice (6, 8, 10). In two studies, the effectof both the application of recombinant GM-CSF and theapplication of a neutralizing antibody to GM-CSF prior to thetreatment with tumor promoting agents had been evaluated.The systemic application of a neutralizing antibody to GM-CSF prior to the treatment with a tumor promoter greatlyreduced the inflammatory response to the tumor promoter inthe skin (6, 8). By contrast, systemic application of GM-CSFenhanced the effect of tumor promoters on keratinocyteproliferation and dermal leukocyte infiltration (6). Similarly,the intradermal transgenic expression of GM-CSF using areplication-deficient adenovirus vehicle resulted in massiveinfiltration of neutrophils and macrophages as well as epi-dermal hyperplasia (40).
To understand the different results between these studiesas compared with our model, we must take into account thedifferent modes of application for GM-CSF. Although theprevious studies used different modes of application for GM-CSF, leading to highly elevated dermal or systemic levels ofGM-CSF, the model described in our study exclusively ad-dresses the question for the biological significance of kera-tinocyte-derived GM-CSF in vivo (6, 8, 10). Although it hadbeen shown previously that growth factors secreted by ke-
ratinocytes in transgenic animals can traverse the basal lam-ina and may be transported to the bloodstream (41, 42), thisis not the case for GM-CSF in our transgenic mouse model.Tissue concentration of GM-CSF in the skin of our transgen-ics is constitutively elevated 2–10-fold over the respectiveconcentrations measured in wt skin upon activation by 12-O-tetradecanoylphorbol-13-acetate (these authors; data notshown). Therefore, we conclude that keratinocyte-derivedGM-CSF usually does not enter the bloodstream in detect-able amounts and does not per se evoke massive inflamma-tory responses in the skin in the absence of additional stimuli.
GM-CSF and Keratinocyte Proliferation, Differentia-tion, and Apoptosis. GM-CSF had been described as astimulator of epidermal cell proliferation both in vitro and invivo (5, 9, 10). Consistent with these findings, we found anincrease of mitotic indices in the interfollicular epidermis ofthe keratin 5/GM-CSF transgenics by a factor of approxi-mately 2–3. However, the proliferating cells were still re-stricted to the basal layer of the epidermis, and we observedneither para- or hyperkeratosis nor pronounced hyperplasia.Also, the normal program of keratinocyte differentiation wasnot altered in these mice, which is reflected by the normalpattern of expression of keratin 14 and keratins 1 and 10 aswell as the absence of keratin 6 in interfollicular epidermis.Again, these latter findings are apparently in contradictionwith previous reports linking GM-CSF overproduction in theskin to hyperplasia, acanthosis, as well as the implementa-tion of the alternative pathway of keratinocyte differentiation,as demonstrated by the induction of keratin 16 in interfol-licular epidermis (5, 9, 10).
The resolution of these discrepancies must be linked to themode of application for GM-CSF, which involves additionalstimuli in all previous studies, e.g., injection or adenoviralgene expression. However, our model analyzes the signifi-cance of the constitutive presence of keratinocyte-derivedGM-CSF and without additional stimuli. Therefore, we con-clude that GM-CSF released by keratinocytes is an autocrinestimulator of keratinocyte proliferation but does not aloneevoke epidermal hyperplasia or acanthosis nor the imple-mentation of the alternative program of keratinocyte differ-
Fig. 6. FACS analysis of epidermal cell suspensions. Isotype control isshown in the upper panels (A). Single-cell suspensions were treated withAnnexinV-FITC and KH118/MHC class II (B) as well as NLDC145/DEC205(dendritic cell-related marker; C). The numbers of vital MHC class II1 (B,upper left) and vital DEC2051 (C, upper left) cells are increased in trans-genics. Also, MHC class II1 (B, upper right) and DEC2052 (C, lower right)cells undergoing apoptosis are more abundant in transgenics.
Fig. 7. Expression of mRNAs for several Bcl-2-related genes in wt, Tg1,and Tg2 animals. Northern blots were prepared by loading 25 mg/lane oftotal RNA isolated from the skin. Although no changes in expression ofBak, Bad, Bax, and Bcl-x were seen, an up-regulation of the antiapoptoticfactor A1 was observed in transgenic skin. Bcl-2 showed a weak butsteady transcriptional product at ;6.5 kb.
117Cell Growth & Differentiation

entiation that is characterized by the induction of keratins 6and 16.
Because hyperproliferation in interfollicular keratinocytesappeared to be in contradiction to the lack of significantalterations of epidermal architecture and thickness, we won-dered about the fate of the epidermal cells. Therefore, weanalyzed apoptosis by the TUNEL assay and FACS analysisfor annexin-V. Both the methods revealed an increase innumbers of apoptotic keratinocytes in the epidermis of thekeratin 5/GM-CSF transgenic mice. Thus, we propose thatstandard epidermal thickness is maintained in the transgenicanimals by increased rates of apoptosis. However, short-ened transit times of keratinocytes may also contribute to themaintenance of normal homeostasis (43, 44).
Bcl-2 family members, important regulators of apoptosis(17–19, 45, 46), are differentially expressed during the proc-ess of terminal differentiation of keratinocytes. Substantialepidermal expression of Bcl-2, Bcl-x, Bad, Bax, and Bak innormal epidermis had been demonstrated before (21–25).However, we do not see prominent changes in the levels ofexpression of these factors in the skin of our keratin 5/GM-CSF transgenic mice. Instead, we see a massive induction ofa novel regulator of programmed cell death, A1, which ishardly detectable in normal skin. Therefore, we conclude thatthe increase in apoptotic frequency in interfollicular epider-mis of our transgenic mice does not result from a modulationof expression of the abundant regulators such as Bcl-2,Bcl-x, Bad, Bax, and Bak. Also, the induction of the normallyinfrequent Bcl-2 family member A1 is not an explanation forthe increased frequency of apoptosis, because A1 has beendescribed solely as antiapoptotic (47). Therefore, a dominantmechanism must be involved rendering keratinocytes in ourGM-CSF transgenics more susceptible to apoptosis. Be-cause GM-CSF has been reported to prevent apoptosis in avariety of cell types (3, 12, 16), this apoptosis-promotingmechanism in our transgenics may be indirect via the mod-erate elevation of IFN-g tissue levels observed in our trans-genics. It had been shown that IFN-g can promote apoptosisin cultured keratinocytes by inducing the expression of pre-desquamin and Fas (26, 27, 48). Whether this is the case inour transgenics is currently under investigation. We suggestthat GM-CSF may play an important role in epithelial carci-nogenesis because of its immunomodulating effect in thecutaneous system on one hand and because of its effects oncell proliferation, differentiation, and programmed cell deathof keratinocytes after genotoxic stress.
Materials and MethodsGeneration of Transgenic Mice. The cDNA coding for murine GM-CSF(49) was inserted by blunt-end ligation into the end-filled SalI site of thekeratin 5 expression vector (34, 50). The expression cassette was excisedby KpnI digestion. The 10-kb construct-DNA was gel purified and used forpronuclear microinjection of fertilized eggs of mice of strain FVB/N, es-sentially as described (51). Offspring were biopsied at ears or tails andanalyzed by PCR using a bovine keratin 5-specific primer (59-ATG AAGACA GCG TTT GCA CCC-39; position 1122–1142; GenBank accession no.Z32746; Ref. 52) and a murine GM-CSF cDNA-specific oligonucleotide(59-CTG GCT GTC ATG TTC AAG GCG-39; position 218–198; GenBankaccession no. X05906; Ref. 49) or by Southern blot analysis using a bovinekeratin 5-specific probe (52).
Keratinocyte Culture. Keratinocytes were derived from newbornmice and cultured as described previously (34). Aliquots of the superna-tants from 4-day-old cultures were used for ELISA to determine cytokineproduction.
RT-PCR Analysis. RT-PCR analysis was performed using total cellu-lar RNA from kidney, lung, heart, liver, dorsal skin, brain, and spleen. TotalRNA was isolated using peqGOLD RNA Pure (Peqlab Biotechnologie,Erlangen, Germany). Reverse transcription was accomplished using aRT-PCR kit according to the manufacturer’s instructions (Stratagene, LaJolla, CA). Two hundred ng of total RNA were used as template for eachassay, and PCR was performed using Taq-DNA polymerase (BoehringerMannheim, Mannheim, Germany) in 35 cycles (94°C for 60 s; 61°C for60 s; and 72°C for 60 s). Primer pairs for both endogenous BMP-6-specificRNA and transgene-derived RNA were flanking introns. Endogenous mu-rine mRNA for BMP-6 was used as a standard as described previously(34, 53). The primer pair used to detect transgene-derived mRNA con-sisted of primer GM-exo-1 (59-CTA CCA GAC ATA CTG CCC CCC-39;position 222–242; GenBank accession no. X05906; Ref. 49), specific forthe murine GM-CSF cDNA, and of the reverse primer GM-exo-2 (59-GAGTAG AAG CTG CTA CTG CCG-39; position 384–363; GenBank accessionno. K03535) specific for the bovine keratin 5 splice site, as deduced fromthe sequence of the corresponding cDNA (54). The primers produced a364-bp diagnostic band at an annealing temperature of 60°C when cDNAsfrom appropriate transgenic tissues were used as templates.
In Situ Hybridization. The transgene-specific cDNA probe derivedfrom the splice and polyadenylation site of the bovine keratin 5 gene wasgenerated by PCR. The primer pair consisted of primer K5-1 (59-tatagaattc AGC AAG TTA CTG CCT CCC-39; position 558–576; GenBankaccession no. K03535) and of primer K5-2 (59-tata aagctt ACC AGG CTAGAG ACT GGG-39; position 736–717; GenBank accession no. K03535;Ref. 55), with EcoRI and HindIII restriction sites added at the 59 end.Fragments were cloned into pGEM-4Z (Promega Corp., Heidelberg, Ger-many), and digoxigenin-labeled sense and antisense RNAs were gener-ated using in vitro transcription with SP6- and T7-RNA polymerasesaccording to the supplier’s protocol (Boehringer Mannheim). The prepa-ration of skin cryosections and stainings were performed as describedpreviously (56).
BrdUrd Labeling. The number of nuclei in S-phase of the cell cyclewas determined by BrdUrd labeling experiments using the In Situ CellProliferation Kit, FLUOS (Boehringer Mannheim). Mice received injectionsof 30 mg of BrdUrd i.p. per gram of body weight and were sacrificed aftera labeling period of 2 h. Fixation and processing of the samples wereperformed according to the manufacturer’s instructions. The ratio of la-beled nuclei in the basal layer of interfollicular epidermis to unlabeled cellswas determined. For each experiment, values were obtained from at leastthree age- and sex-matched animals.
Cytokine Determinations. Blood samples were obtained by cardiacpuncture using 22-gauge needles. One half of the blood was mixed with50 ml of 0.5 M EDTA for plasma preparation, and the other half was left atroom temperature for 2 h for serum collection. After centrifugation at 6000rpm (Sorvall; FA-Micro) for 10 min, the supernatants were used for ELISA.For skin extracts, mice were sacrificed, and 2-cm2 skin samples weretaken from the shaved mid-dorsum. The tissue was pulverized underliquid nitrogen, weighed, and suspended in 600 ml of PBS. After theaddition of 1 mg of protease inhibitor phenylmethylsulfonyl fluoride (SigmaChemical Co., Deisenhofen, Germany), the samples were mixed inten-sively and centrifuged for 30 min with 12,000 rpm (Sorvall; FA-Micro) at4°C. The supernatants were used for ELISA. IFN-g, IL-2, IL-4, IL-12, IL-1b,and GM-CSF levels in serum, plasma, and skin extracts were evaluatedusing cytokine-specific ELISA kits (R&D Systems, Inc., Minneapolis, MN),according to the manufacturer’s instructions.
Immunohistology. Mouse skin samples from age- and sex-matchedanimals were embedded in tissue freezing medium (Jung; Leica-Instru-ments, Nussloch, Germany), frozen immediately in liquid nitrogen, andstored at 280°C. Cryosections were cut at 5-mm thickness and fixed in4% paraformaldehyde in PBS for 20 min at room temperature or withacetone at 4°C for 15 min. After preblocking with 1% BSA, 5% serum,0.5% Tween 20, and 100 mM MgCl2 in 10 mM Tris-HCl (pH 7.4) for 30 minat room temperature, the primary antibody was applied in the blockingsolution at 4°C overnight. The secondary antibody incubation was per-formed at 37°C for 60 min. Signal detection was carried out using NBT/X-Phosphate or Fast Red (Boehringer Mannheim) as substrate. Endoge-
118 GM-CSF Transgenic Mice

nous alkaline phosphatase activity was inhibited by addition of 1 mM
levamisol to the substrate solution. For fluorescence analysis, secondaryantibodies conjugated with FITC and CyTM3 were used.
For the preparation of epidermal ear sheets and immunostaining, micewere sacrificed, and ears were removed. After dividing the ear in dorsaland ventral halves, the s.c. fat and cartilage were removed with the helpof tweezers. The ear halves were incubated in 5 ml of 0.5 M NH4SCNsolution in 0.1 M Na2HPO4, 0.1 M KH2PO4 buffer (pH 6.8) for 20 min at37°C. Epidermis was separated from dermis and washed carefully in PBS.Fixation was carried out in ice-cold acetone for 40–45 min. After gentlywashing three times in PBS, the epidermal sheets were incubated with theprimary antibody for 1 h at 4°C. After washing three times in PBS, thesecondary antibody was added for 1 h at 4°C. The epidermis was spreadout on a glass slide and examined under a fluorescence microscope (57).
Preparation of Epidermal Cell Suspensions for FACS Analysis.Mice were sacrificed, and ears were removed. After disinfection with 70%ethanol, ears were split into dorsal and ventral halves using fine tweezers.After removal of s.c. fat and cartilage, ear halves were incubated in 0.5%Trypsin/0.2 mM EDTA for 20 min at 37°C. The digestion reaction wasstopped by the addition of excess FCS. The epidermis was removed fromthe dermis, and single-cell suspension was obtained by pressing theepidermis through a nylon mesh (58).
Flow Cytometric Analysis. After washing the cells twice with PBS,indirect immunofluorescence staining was performed. The cells were in-cubated with each primary monoclonal antibody diluted to the optimalconcentration for immunostaining at 4°C for 20 min. After two washes withcold medium (PBS, 2% FCS, and 0.5 mM EDTA), cells were incubated withphycoerythrin- and FITC-conjugated secondary antibodies at 4°C for 20min and washed twice in PBS. About 5,000–10,000 cells were collectedby flow cytometry using a FACScan (Becton Dickinson, Mountain View,CA). Debris and dead cells were gated out, and live cells were plottedusing the FACScan Lysis II software (Becton Dickinson).
Antibodies. The following antibodies at the mentioned dilutions wereused for immunohistochemistry in this study: rat monoclonal antibodyCD11b (1:200; PharMingen), CD4 (1:50; PharMingen; FITC-conjugated),CD8a (1:50; PharMingen; FITC-conjugated), DEC205 (NLDC145, BMA),CD117 (1:50; PharMingen), cytokeratin 10 (1:1600; Sigma), GM-CSF (1:50; PharMingen), and rabbit polyclonal antibody against murine cytokera-tin 6 (59) and cytokeratin 14 (34).
Secondary antibodies used in different dilutions in this study wereCyTM3-conjugated donkey antimouse IgG (Dianova), CyTM3-conjugatedgoat antirat IgG (Dianova), AP-conjugated rabbit antirat IgG (Sigma), andAP-conjugated sheep antirabbit IgG (Boehringer Mannheim).
Antibodies used for flow cytometric analysis in this study were asfollows: mouse monoclonal antibody against murine MHC class II (KH118;I-Aq; PharMingen), DEC205 (NLDC145, BMA), and AnnexinV-FITC(PharMingen). Isotype-matched controls were obtained from PharMingen(San Diego, CA).
Quantification of Apoptosis. Detection and quantification of pro-grammed cell death in the epidermis were performed using In Situ CellDeath Detection Kit, FLUOS (Boehringer Mannheim). Fresh cryosectionsfrom different mouse lines were treated as recommended by the supplier,counterstained for cytokeratin 14, and analyzed by fluorescence micros-copy. The number of apoptotic cells in the basal epidermal layer wasdetermined and related to 100 total basal cell nuclei (49,6-diamidino-2-phenylindole). The number of nuclei per visualized field was calculatedby an average value obtained from 50 fields. Nuclei were counted from atleast three age- and sex-matched animals of each line.
Histology. For histopathological diagnosis, skin specimens were fixedovernight in neutral buffered 4% formalin, processed for paraffin embed-ding, sectioned, and stained routinely with H&E. For mast cell detection,skin cryosections were stained with May-Grunwald/Giemsa stain. Forquantification of mast cells, at least three age-matched animals of eachline were analyzed.
Northern Blot Analysis. Total RNA from wt and transgenic skin wasisolated using peqGOLD RNA Pure (Peqlab Biotechnologie). Twenty-fivemg of RNA were electrophoresed on 1% agarose formaldehyde gels andsubsequently blotted onto nylon membranes (Hybond N, Amersham,Braunschweig, Germany). After hybridization with 32P-labeled cDNAprobes, the filters were processed at high stringency as described (60).
The probes were generated by PCR (35 cycles; 94°C for 30s; 58°C for45 s; and 72°C for 55 s) with spleen cDNA serving as template and using
the following primer pairs: A1 (A1–1: 59-GCC TCC AGA TAT GAT TAGGG-39; position 28–48; GenBank accession no. L16462) and (A1–2: 59-CTG ATA ACC ATT CTC GTG GG-39; position 698–678; GenBank ac-cession no. L16462; Ref. 47); b-actin (act-1: 59-GTG GGC CGC TCT AGGCAC CA-39; position 183–203; GenBank accession no. X03672) and(act-2: 59-CTG TGC CCA TCT ACG AGG GCT A-39; position 578–565;GenBank accession no. X03672; Ref. 61); Bad (Bad-1: 59-CAG GGA GGTGTC ATT AAC CC-39; position 378–398; GenBank accession no. L37296)and (Bad-2: 59-CCA GGA CTG GAT AAT GCG CG-39; position 1041–1021;GenBank accession no. L37296; Ref. 17); Bak (Bak-1: 59-ATT CAG GTGACA AGT GAC GG-39; position 212–232; GenBank accession no. Y13231)and (Bak-2: 59-GAG AGA GGT TTA GTC CAG CC-39; position 940–920;GenBank accession no. Y13231; Ref. 62); Bax (Bax-1: 59-ATG GAC GGGTCC GGG GAG CA-39; position 1–21; GenBank accession no. L22472)and (Bax-4: 59-CTT CTT CCA GAT GGT GAG CG-39; position 571–551;GenBank accession no. L22472; Ref. 63); Bcl-2 (Bcl2–1: 59-AGA ACAGGG TAT GAT AAC CG-39; position 1836–1856; GenBank accession no.M16506) and (Bcl2–2: 59-TAT CCT GGA TCC AGG TGT GC-39; position2386–2366; GenBank accession no. M16506; Ref. 64); Bcl-x (Bcl-x-1:59-GCT AAA CAC AGA GCA GAC CC-39; position 166–186; GenBankaccession no. L35049) and (Bcl-x-2: 59-GAG ATC CAC AAA AGT GTCCC-39; position 825–805; GenBank accession no. L35049; Ref. 65). AllPCR products were verified by sequence analysis.
AcknowledgmentsWe are indebted to Karin Nicol and Martina Protschka for excellent tech-nical assistance and Prof. Meyer zum Buschenfelde and Prof. Galle forconstant encouragement and support. We thank R. Kastelein (DNAX) forproviding us with a cDNA coding for murine GM-CSF.
References1. Nishijima, I., Nakahata, T., Hirabayashi, Y., Inoue, T., Kurata, H., Miya-jima, A., Hayashi, N., Iwakura, Y., Arai, K., and Yokota, T. A humanGM-CSF receptor expressed in transgenic mice stimulates proliferationand differentiation of hemopoietic progenitors to all lineages in responseto human GM-CSF. Mol. Biol. Cell, 6: 497–508, 1995.
2. Caux, C., Vanbervliet, B., Massacrier, C., Dezutter-Dambuyant, C., deSaint-Vis, B., Jacquet, C., Yoneda, K., Imamura, S., Schmitt, D., andBanchereau, J. CD341 hematopoietic progenitors from human cordblood differentiate along two independent dendritic cell pathways in re-sponse to GM-CSF1TNFa. J. Exp. Med., 184: 695–706, 1996.
3. Dibbert, B., Daigle, I., Braun, D., Schranz, C., Weber, M., Blaser, K.,Zangemeister-Wittke, U., Akbar, A. N., and Simon, H. U. Role for Bcl-xLin delayed eosinophil apoptosis mediated by granulocyte- macrophagecolony-stimulating factor and interleukin-5. Blood, 92: 778–783, 1998.
4. Kupper, T. S., Lee, F., Coleman, D., Chodakewitz, J., Flood, P., andHorowitz, M. Keratinocyte derived T-cell growth factor (KTGF) is identicalto granulocyte macrophage colony stimulating factor (GM-CSF). J. Inves-tig. Dermatol., 91: 185–188, 1988.
5. Braunstein, S., Kaplan, G., Gottlieb, A. B., Schwartz, M., Walsh, G.,Abalos, R. M., Fajardo, T. T., Guido, L. S., and Krueger, J. G. GM-CSFactivates regenerative epidermal growth and stimulates keratinocyte pro-liferation in human skin in vivo. J. Investig. Dermatol., 103: 601–604, 1994.
6. Robertson, F. M., Bijur, G. N., Oberyszyn, A. S., Pellegrini, A. E., Boros,L. G., Sabourin, C. L., and Oberyszyn, T. M. Granulocyte-macrophagecolony stimulating factor gene expression and function during tumorpromotion. Carcinogenesis (Lond.), 15: 1017–1029, 1994.
7. Pastore, S., Fanales-Belasio, E., Albanesi, C., Chinni, L. M., Giannetti,A., and Girolomoni, G. Granulocyte macrophage colony-stimulating factoris overproduced by keratinocytes in atopic dermatitis. Implications forsustained dendritic cell activation in the skin. J. Clin. Investig., 99: 3009–3017, 1997.
8. Vasunia, K. B., Miller, M. L., Puga, A., and Baxter, C. S. Granulocyte-macrophage colony-stimulating factor (GM-CSF) is expressed in mouseskin in response to tumor-promoting agents and modulates dermal in-flammation and epidermal dark cell numbers. Carcinogenesis (Lond.), 15:653–660, 1994.
119Cell Growth & Differentiation

9. Kaplan, G., Walsh, G., Guido, L. S., Meyn, P., Burkhardt, R. A., Abalos,R. M., Barker, J., Frindt, P. A., Fajardo, T. T., Celona, R., et al. Novelresponses of human skin to intradermal recombinant granulocyte/macro-phage-colony-stimulating factor: Langerhans cell recruitment, keratino-cyte growth, and enhanced wound healing. J. Exp. Med., 175: 1717–1728,1992.
10. Xing, Z., Gauldie, J., Tremblay, G. M., Hewlett, B. R., and Addison, C.Intradermal transgenic expression of granulocyte-macrophage colony-stimulating factor induces neutrophilia, epidermal hyperplasia, Langer-hans’ cell/macrophage accumulation, and dermal fibrosis. Lab. Investig.,77: 615–622, 1997.
11. Caux, C., Dezutter-Dambuyant, C., Schmitt, D., and Banchereau, J.GM-CSF and TNF-a cooperate in the generation of dendritic Langerhanscells. Nature (Lond.), 360: 258–261, 1992.
12. Bratton, D. L., Hamid, Q., Boguniewicz, M., Doherty, D. E., Kailey,J. M., and Leung, D. Y. Granulocyte macrophage colony-stimulating fac-tor contributes to enhanced monocyte survival in chronic atopic derma-titis. J. Clin. Investig., 95: 211–218, 1995.
13. Leung, D. Y. Atopic dermatitis: immunobiology and treatment withimmune modulators. Clin. Exp. Immunol., 107: 25–30, 1997.
14. Rodriguez-Villanueva, J., Greenhalgh, D., Wang, X. J., Bundman, D.,Cho, S., Delehedde, M., Roop, D., and McDonnell, T. J. Human keratin-1.bcl-2 transgenic mice aberrantly express keratin 6, exhibit reducedsensitivity to keratinocyte cell death induction, and are susceptible to skintumor formation. Oncogene, 16: 853–863, 1998.
15. Lindner, G., Botchkarev, V. A., Botchkareva, N. V., Ling, G., van derVeen, C., and Paus, R. Analysis of apoptosis during hair follicle regression(catagen) [see comments]. Am. J. Pathol., 151: 1601–1617, 1997.
16. Kinoshita, T., Yokota, T., Arai, K., and Miyajima, A. Suppression ofapoptotic death in hematopoietic cells by signalling through the IL-3/GM-CSF receptors. EMBO J., 14: 266–275, 1995.
17. Yang, E., Zha, J., Jockel, J., Boise, L. H., Thompson, C. B., andKorsmeyer, S. J. Bad, a heterodimeric partner for Bcl-XL and Bcl-2,displaces Bax and promotes cell death. Cell, 80: 285–291, 1995.
18. Sedlak, T. W., Oltvai, Z. N., Yang, E., Wang, K., Boise, L. H., Thomp-son, C. B., and Korsmeyer, S. J. Multiple Bcl-2 family members demon-strate selective dimerizations with Bax. Proc. Natl. Acad. Sci. USA, 92:7834–7838, 1995.
19. Zha, J., Harada, H., Yang, E., Jockel, J., and Korsmeyer, S. J. Serinephosphorylation of death agonist BAD in response to survival factorresults in binding to 14-3-3 not BCL-X(L) [see comments]. Cell, 87: 619–628, 1996.
20. Pena, J. C., Fuchs, E., and Thompson, C. B. Bcl-x expression influ-ences keratinocyte cell survival but not terminal differentiation. CellGrowth Differ., 8: 619–629, 1997.
21. Hockenbery, D. M., Zutter, M., Hickey, W., Nahm, M., and Korsmeyer,S. J. BCL2 protein is topographically restricted in tissues characterized byapoptotic cell death. Proc. Natl. Acad. Sci. USA, 88: 6961–6965, 1991.
22. Stenn, K. S., Lawrence, L., Veis, D., Korsmeyer, S., and Seiberg, M.Expression of the bcl-2 protooncogene in the cycling adult mouse hairfollicle. J. Investig. Dermatol., 103: 107–111, 1994.
23. Krajewski, S., Krajewska, M., Shabaik, A., Miyashita, T., Wang, H. G.,and Reed, J. C. Immunohistochemical determination of in vivo distributionof Bax, a dominant inhibitor of Bcl-2. Am. J. Pathol., 145: 1323–1336,1994.
24. Krajewski, S., Krajewska, M., Shabaik, A., Wang, H. G., Irie, S., Fong,L., and Reed, J. C. Immunohistochemical analysis of in vivo patterns ofBcl-X expression. Cancer Res., 54: 5501–5507, 1994.
25. Krajewski, S., Krajewska, M., and Reed, J. C. Immunohistochemicalanalysis of in vivo patterns of Bak expression, a proapoptotic member ofthe Bcl-2 protein family. Cancer Res., 56: 2849–2855, 1996.
26. Sayama, K., Yonehara, S., Watanabe, Y., and Miki, Y. Expression ofFas antigen on keratinocytes in vivo and induction of apoptosis in culturedkeratinocytes. J. Investig. Dermatol., 103: 330–334, 1994.
27. Brysk, M. M., Selvanayagam, P., Arany, I., Brysk, H., Tyring, S. K., andRajaraman, S. Induction of apoptotic nuclei by interferon-g and by pre-desquamin in cultured keratinocytes. J. Interferon Cytokine Res., 15:1029–1035, 1995.
28. Takahashi, H., Saitoh, K., Kishi, H., and Parsons, P. G. Immunohis-tochemical localisation of stem cell factor (SCF) with comparison of itsreceptor c-Kit proto-oncogene product (c-KIT) in melanocytic tumours.Virchows Arch., 427: 283–288, 1995.
29. Breel, M., Laman, J., and Kraal, G. Murine hybrid cell lines expressingthe NLDC-145 dendritic cell determinant. Immunobiology, 178: 167–176,1988.
30. Swiggard, W. J., Mirza, A., Nussenzweig, M. C., and Steinman, R. M.DEC-205, a 205-kDa protein abundant on mouse dendritic cells andthymic epithelium that is detected by the monoclonal antibody NLDC-145:purification, characterization, and N-terminal amino acid sequence. CellImmunol., 165: 302–311, 1995.
31. Hancock, G. E., Kaplan, G., and Cohn, Z. A. Keratinocyte growthregulation by the products of immune cells. J. Exp. Med., 168: 1395–1402,1988.
32. Noso, N., Sticherling, M., Bartels, J., Mallet, A. I., Christophers, E.,and Schroder, J. M. Identification of an N-terminally truncated form of thechemokine RANTES and granulocyte-macrophage colony-stimulatingfactor as major eosinophil attractants released by cytokine-stimulateddermal fibroblasts. J. Immunol., 156: 1946–1953, 1996.
33. Imokawa, G., Yada, Y., Kimura, M., and Morisaki, N. Granulocyte/macrophage colony-stimulating factor is an intrinsic keratinocyte-derivedgrowth factor for human melanocytes in UVA-induced melanosis. Bio-chem. J., 313: 625–631, 1996.
34. Amendt, C., Schirmacher, P., Weber, H., and Blessing, M. Expressionof a dominant negative type II TGF-b receptor in mouse skin results in anincrease in carcinoma incidence and an acceleration of carcinoma devel-opment. Oncogene, 17: 25–34, 1998.
35. Lang, R. A., Metcalf, D., Cuthbertson, R. A., Lyons, I., Stanley, E.,Kelso, A., Kannourakis, G., Williamson, D. J., Klintworth, G. K., Gonda,T. J., et al. Transgenic mice expressing a hemopoietic growth factor gene(GM-CSF) develop accumulations of macrophages, blindness, and a fatalsyndrome of tissue damage. Cell, 51: 675–686, 1987.
36. Gearing, A. J., Metcalf, D., Moore, J. G., and Nicola, N. A. Elevatedlevels of GM-CSF and IL-1 in the serum, peritoneal and pleural cavities ofGM-CSF transgenic mice. Immunology, 67: 216–220, 1989.
37. Metcalf, D., Elliott, M. J., and Nicola, N. A. The excess numbers ofperitoneal macrophages in granulocyte-macrophage colony-stimulatingfactor transgenic mice are generated by local proliferation. J. Exp. Med.,175: 877–884, 1992.
38. Herbst, B., Kohler, G., Mackensen, A., Veelken, H., and Lindemann, A.GM-CSF promotes differentiation of a precursor cell of monocytes andLangerhans-type dendritic cells from CD341 haemopoietic progenitorcells. Br. J. Haematol., 101: 231–241, 1998.
39. Damsgaard, T. E., Olesen, A. B., Sorensen, F. B., Thestrup-Pedersen,K., and Schiotz, P. O. Mast cells and atopic dermatitis. Stereologicalquantification of mast cells in atopic dermatitis and normal human skin.Arch. Dermatol. Res., 289: 256–260, 1997.
40. Xing, Z., Tremblay, G. M., Sime, P. J., and Gauldie, J. Overexpressionof granulocyte-macrophage colony-stimulating factor induces pulmonarygranulation tissue formation and fibrosis by induction of transforminggrowth factor-b1 and myofibroblast accumulation. Am. J. Pathol., 150:209–222, 1997.
41. Cheng, J., Turksen, K., Yu, Q. C., Schreiber, H., Teng, M., and Fuchs,E. Cachexia and graft-vs.-host-disease-type skin changes in keratin pro-moter-driven TNF a transgenic mice. Genes Dev., 6: 1444–1456, 1992.
42. Wang, X., Zinkel, S., Polonsky, K., and Fuchs, E. Transgenic studieswith a keratin promoter-driven growth hormone transgene: prospects forgene therapy. Proc. Natl. Acad. Sci. USA, 94: 219–226, 1997.
43. Potten, C. S. Epidermal transit times. Br. J. Dermatol., 93: 649–658,1975.
44. Potten, C. S., Hume, W. J., and Parkinson, E. K. Migration and mitosisin the epidermis. Br. J. Dermatol., 111: 695–699, 1984.
45. Tao, W., Kurschner, C., and Morgan, J. I. Bcl-xS and Bad potentiatethe death suppressing activities of Bcl-xL, Bcl-2, and A1 in yeast. J. Biol.Chem., 273: 23704–23708, 1998.
120 GM-CSF Transgenic Mice

46. Tao, W., Kurschner, C., and Morgan, J. I. Modulation of cell death inyeast by the Bcl-2 family of proteins. J. Biol. Chem., 272: 15547–15552,1997.
47. Lin, E. Y., Orlofsky, A., Berger, M. S., and Prystowsky, M. B. Char-acterization of A1, a novel hemopoietic-specific early-response gene withsequence similarity to bcl-2. J. Immunol., 151: 1979–1988, 1993.
48. Takahashi, H., Kobayashi, H., Hashimoto, Y., Matsuo, S., and Iizuka,H. Interferon-g-dependent stimulation of Fas antigen in SV40- trans-formed human keratinocytes: modulation of the apoptotic process byprotein kinase C. J. Investig. Dermatol., 105: 810–815, 1995.
49. Gough, N. M., Gough, J., Metcalf, D., Kelso, A., Grail, D., Nicola, N. A.,Burgess, A. W., and Dunn, A. R. Molecular cloning of cDNA encoding amurine haematopoietic growth regulator, granulocyte-macrophage col-ony stimulating factor. Nature (Lond.), 309: 763–767, 1984.
50. Blessing, M., Nanney, L. B., King, L. E., Jones, C. M., and Hogan, B. L.Transgenic mice as a model to study the role of TGF-b-related moleculesin hair follicles. Genes Dev., 7: 204–215, 1993.
51. Hogan, B. L. M., Costatini, F., and Lacy, E. Manipulating the MouseEmbryo. A Laboratory Manual, pp. 153–197. Cold Spring Harbor, NY:Cold Spring Harbor Laboratory, 1986.
52. Casatorres, J., Navarro, J. M., Blessing, M., and Jorcano, J. L. Anal-ysis of the control of expression and tissue specificity of the keratin 5gene, characteristic of basal keratinocytes. Fundamental role of an AP-1element. J. Biol. Chem., 269: 20489–20496, 1994.
53. Gitelman, S. E., Kobrin, M., Lee, A., Fet, V., Lyons, K., Hogan, B. L. M.,and Derynck, R. Structure and sequence of the mouse Bmp6 gene. Calcif.Tissue Int., 60: 283–290, 1997.
54. Jorcano, J. L., Franz, J. K., and Franke, W. W. Amino acid sequencediversity between bovine epidermal cytokeratin polypeptides of the basic(type II) subfamily as determined from cDNA clones. Differentiation, 28:155–163, 1984.
55. Jorcano, J. L., Rieger, M., Franz, J. K., Schiller, D. L., Moll, R., andFranke, W. W. Identification of two types of keratin polypeptides within theacidic cytokeratin subfamily I. J. Mol. Biol., 179: 257–281, 1984.
56. Kaiser, S., Schirmacher, P., Philipp, A., Protschka, M., Moll, I., Nicol,K., and Blessing, M. Induction of bone morphogenetic protein-6 in skin
wounds. Delayed reepithelialization and scar formation in BMP-6 overex-pressing transgenic mice. J. Investig. Dermatol., 111: 1145–1152, 1998.
57. Howells, G. L., and Williams, D. M. The sequential demonstration ofnon-specific esterase reactivity Ia antigen and Thy-1 antigen in murineepidermal sheets. Arch. Oral Biol., 32: 679–683, 1987.
58. Becker, D., Reske-Kunz, A. B., Knop, J., and Reske, K. Biochemicalproperties of MHC class II molecules endogenously synthesized andexpressed by mouse Langerhans cells. Eur. J. Immunol., 21: 1213–1220,1991.
59. Blessing, M., Schirmacher, P., and Kaiser, S. Overexpression of bonemorphogenetic protein-6 (BMP-6) in the epidermis of transgenic mice:inhibition or stimulation of proliferation depending on the pattern of trans-gene expression and formation of psoriatic lesions. J. Cell Biol., 135:227–239, 1996.
60. Church, G. M., and Gilbert, W. Genomic sequencing. Proc. Natl.Acad. Sci. USA, 81: 1991–1995, 1984.
61. Tokunaga, K., Taniguchi, H., Yoda, K., Shimizu, M., and Sakiyama, S.Nucleotide sequence of a full-length cDNA for mouse cytoskeletal b-actinmRNA. Nucleic Acids Res., 14: 2829, 1986.
62. Ulrich, E., Kauffmann-Zeh, A., Hueber, A. O., Williamson, J., Chit-tenden, T., Ma, A., and Evan, G. Gene structure, cDNA sequence, andexpression of murine Bak, a proapoptotic Bcl-2 family member. Genom-ics, 44: 195–200, 1997.
63. Oltvai, Z. N., Milliman, C. L., and Korsmeyer, S. J. Bcl-2 heterodimer-izes in vivo with a conserved homolog, Bax, that accelerates programmedcell death. Cell, 74: 609–619, 1993.
64. Negrini, M., Silini, E., Kozak, C., Tsujimoto, Y., and Croce, C. M.Molecular analysis of mbcl-2: structure and expression of the murine genehomologous to the human gene involved in follicular lymphoma. Cell, 49:455–463, 1987.
65. Gonzalez-Garcia, M., Perez-Ballestero, R., Ding, L., Duan, L., Boise,L. H., Thompson, C. B., and Nunez, G. bcl-XL is the major bcl-x mRNAform expressed during murine development and its product localizes tomitochondria. Development (Camb.), 120: 3033–3042, 1994.
121Cell Growth & Differentiation
Copyright © 2022 FDOKUMEN




















