
Radioprotective efficacy of tocopherol succinate is mediated through granulocyte-colony stimulating...
12
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Radioprotective efficacy of tocopherol succinate is mediated through granulocyte-colony stimulating...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Radioprotective efficacy of tocopherol succinate is mediatedthrough granulocyte-colony stimulating factor
Pankaj K. Singh a, Stephen Y. Wise a, Elizabeth J. Ducey a, Darren S. Brown a, Vijay K. Singh a,b,⇑a Radiation Countermeasures Program, Armed Forces Radiobiology Research Institute, Bethesda, MD 20889-5603, USAb Department of Radiation Biology, F. Edward Hébert School of Medicine, Uniformed Services University of the Health Sciences, Bethesda, MD, USA
a r t i c l e i n f o
Article history:Received 19 March 2011Received in revised form 14 July 2011Accepted 5 August 2011Available online 30 August 2011
Keywords:ApoptosisGastrointestinal injuryc-RadiationMiceTocopherol succinate
a b s t r a c t
The purpose of this study was to elucidate the role of granulocyte colony-stimulating factor (G-CSF)induced by a-tocopherol succinate (TS) in protecting mice from total-body irradiation. CD2F1 mice wereinjected with a radioprotective dose of TS and the levels of cytokine in serum induced by TS were deter-mined by multiplex Luminex. Neutralization of G-CSF was accomplished by administration of a G-CSFantibody and confirmed by cytokine analysis. The role of G-CSF on gastrointestinal tissue protectionafforded by TS after irradiation (11 Gy, 0.6 Gy/min of 60Co c-radiation) was determined by analysis ofjejunum histopathology for crypt, villi, mitotic figures, apoptosis, and cell proliferation. Our results dem-onstrate that TS protected mice against high doses of radiation-induced gastrointestinal damage and TSalso induced very high levels of G-CSF and keratinocyte-derived chemokine (KC) production in peripheralblood 24 h after subcutaneous administration. When TS-injected mice were administered a neutralizingantibody to G-CSF, there was complete neutralization of G-CSF in circulating blood, and the protectiveeffect of TS was significantly abrogated by G-CSF antibody. Histopathology of jejunum from TS-injectedand irradiated mice demonstrated protection of gastrointestinal tissue, yet the protection was abrogatedby administration of a G-CSF antibody. In conclusion, our current study suggests that induction of G-CSFresulting from TS administration is responsible for protection from 60Co c-radiation injury.
Published by Elsevier Ltd.
1. Introduction
Uncontrolled radiation exposure may result from a release ofradioactivity from small sources, transportation accidents, nuclearpower plant accidents, nuclear weapons, and terrorist attacksusing a radiological dispersal device. A terrorist detonation of a nu-clear or other radiological weapons would lead to a mass-casualtyscenario involving victims with varying degrees of exposure to ion-izing radiation [1,2]. At present there are no preventive measuresavailable to protect from the hazards of radiation exposure. Theproblem has become more acute in recent times due to the unpre-dictable nature of terrorist attacks. Radiation hazards can varyfrom late-life pathologies to acute mortality [3,4]. At doses above1 Gy in humans, the hematopoietic system is compromised, lead-ing to decreases in white blood cell counts and increased suscepti-bility to infection. At doses above 8 Gy, injury to thegastrointestinal system becomes serious and contributes to mor-tality. The acute consequences of exposures between 1 and 8 Gyare termed the ‘‘hematopoietic syndrome,’’ while the acute effects
following exposure doses of about 8 to 30 Gy are known as the‘‘gastrointestinal (GI) syndrome.’’ Individuals receiving radiationdoses that nevertheless result in the hematopoietic and GI syn-dromes are more likely to be amenable to pharmacological inter-vention. Therefore, these two syndromes are the specific targetsfor the development of novel therapeutics. GI tract injury is a majorcause of death and radiation-induced severe health impairments ormorbidity. The GI syndrome is characterized by massive apoptoticcell death in the intestinal epithelium, followed by disintegrationof the intestinal wall and death from fluid imbalance, intestinalbleeding, and sepsis.
Shielding is the best protection against the harmful effects ofionizing radiation. An alternative to shielding are pharmaceuticalradiation countermeasures. However, to date there are no safeand effective drugs available for protection against radiation dam-age outside the clinic [5]. Over several decades, many agents havebeen investigated as countermeasures for radiation exposure butnone have been approved for use. Hence, the development of anon-toxic radiation countermeasure is vital. Natural compoundshave been evaluated as radioprotectants and they seem to showtheir effects through antioxidant and immunostimulant activities[5–7]. Tocols, commonly known as vitamin E, consist of four toc-opherols (a, b, c, d) and four tocotrienols (a, b, c, d) [8]. Alpha-tocopherol is a powerful biological antioxidant that also helps
1043-4666/$ - see front matter Published by Elsevier Ltd.doi:10.1016/j.cyto.2011.08.016
⇑ Corresponding author at: Radiation Countermeasures Program, Armed ForcesRadiobiology Research Institute, 8901 Wisconsin Ave, Bethesda, MD 20889-5603,USA. Tel.: +1 301 295 2347; fax: +1 301 295 6503.
E-mail address: [email protected] (V.K. Singh).
Cytokine 56 (2011) 411–421
Contents lists available at SciVerse ScienceDirect
Cytokine
journal homepage: www.elsevier .com/locate / issn/10434666

Author's personal copy
protect cells against the adverse effects of free radicals [9,10].These fat-soluble vitamins intercept free radicals, prevent a chainreaction of lipid destruction, and protect molecules and tissuesfrom oxidative damage by radiation. Alpha-tocopherol has beenthe focus of research because it is the predominant form in humanand animal tissues [8]. The other tocopherols and tocotrienols havebeen receiving attention recently because of their unique biologicaleffects in nutrition and health [11–15].
Granulocyte-colony stimulating factor (G-CSF) is a glycoproteinsecreted by macrophages, fibroblasts, and endothelial cells [16,17].G-CSF is particularly useful for treatment of radiation victims, pro-vided that residual uninjured hematopoietic stem progenitor cellspersist [18]. G-CSF has Food and Drug Administration marketingapproval for the management of treatment-associated neutropeniabut there is no marketing approval for the management of radia-tion-induced marrow aplasia [19].
Recently, we have demonstrated that TS protects mice againsthematopoietic syndrome-producing doses of ionizing radiation, re-duces neutropenia and monocytopenia, and has a dose reductionfactor of 1.28 [20]. It modulates the expression of antioxidant en-zymes, inhibits the expression of oncogenes, and increases col-ony-forming unit-spleen (CFU-S) numbers and bone marrowcellularity in irradiated mice [21]. TS protects mice against hema-topoietic syndrome doses of ionizing radiation by inducing G-CSFand the protective effects of TS can be abrogated by the use of aneutralizing antibody to G-CSF [22]. TS injection mobilized stemcells to the peripheral circulation which mitigated radiation injurywhen transfused to irradiated mice [23]. In this study our objectivewas to further investigate the role of G-CSF in the radioprotectiveefficacy of TS in mice exposed to higher doses of 60Co c -radiationthat can induce GI syndrome. We demonstrate that TS protectedmice against high doses of radiation that cause GI syndrome,induced significant levels of several cytokines, and that administra-tion of a G-CSF antibody completely neutralized TS-stimulated G-CSF in peripheral blood. G-CSF antibody administration abrogatedGI protection afforded by TS in mice exposed to high radiationdoses of 60Co c -radiation.
2. Materials and methods
2.1. Mice
Male 6–8 week-old CD2F1 mice were purchased (Harlan, India-napolis, IN, USA) and housed (8 per cage) in an air-conditionedfacility accredited by the Association for Assessment and Accredi-tation of Laboratory Animal Care International. All mice were keptin rooms with a 12 h light/dark cycle. The mouse holding room wasmaintained at 21 ± 2 �C with 10–15 hourly cycles of fresh air and arelative humidity of 50 ± 10%. Upon arrival, the mice were held inquarantine for 1 week. A microbiological examination of represen-tative samples ensured the absence of Pseudomonas aeruginosa.Mice were provided certified rodent rations (Harlan Teklad RodentDiet, Harlan Teklad, WI, USA) and acidified water (HCl, pH = 2.5–2.8) ad libitum. All animal procedures were performed accordingto a protocol approved by the Armed Forces Radiobiology ResearchInstitute’s Institutional Animal Care and Use Committee. Researchwas conducted according to the Guide for the Care and Use ofLaboratory Animals prepared by the Institute of Laboratory AnimalResources, National Research Council, US National Academy ofSciences.
2.2. Drug preparation and administration
TS (Sigma–Aldrich, St. Louis, MO, USA) was administered as asuspension as described earlier [23]. For a 400 mg/kg dose
(10 mg for a 25-g mouse), 1000 mg of TS were dispersed in8.35 ml PEG-400 and 0.5 ml emulsifier for a total volume of10 ml. For vehicle control, the same composition of emulsifierand PEG-400 was used. TS was administered subcutaneously (sc)in a volume of 0.1 ml using a 23-gauge needle. Monoclonal anti-mouse G-CSF antibody (8.21 mg/ml in PBS) was purchased fromR&D Systems Inc., (Minneapolis, MN, USA). Antibody was dilutedto 3000 lg/ml in D-PBS and administered in a volume of 0.2 mlD-PBS using a 23-gauge needle.
2.3. Irradiation
Mice were placed in well-ventilated Plexiglas boxes compart-mentalized to accommodate eight non-anaesthetized mice perbox and exposed to bilateral irradiation in the AFRRI cobalt-60facility at a dose rate of 0.6 Gy/min. After irradiation, mice were re-turned to their cages and monitored. Sham-irradiated mice weretreated in exactly the same way as the irradiated animals exceptthat the cobalt-60 rods were not raised from the pool of shieldingwater. Radiation dosimetry has been described earlier [23].
2.4. G-CSF neutralization
An antibody to neutralize G-CSF was used to determinewhether TS-induced G-CSF is a key factor for the survival of irradi-ated TS-treated mice. Mice were administered TS (400 mg/kg), sc,24 h before blood harvest. The TS-treated mice then received eitherthe G-CSF antibody (0.2 ml, 600 lg/mouse) or the isotype control(0.2 ml, 600 lg/mouse) intraperitoneally 8 h after TS administra-tion. A monoclonal anti-mouse G-CSF antibody (8.21 mg/ml inphosphate buffered saline: PBS) and rat IgG1 isotype control(8.90 mg/ml in PBS) were purchased (R&D Systems Inc., MN,USA). The antibody was diluted to 3000 lg/ml in PBS and adminis-tered in a volume of 0.2 ml PBS using a 23-gauge needle. Bloodsamples were collected 16 h after G-CSF antibody injection.
2.5. Blood collection and Luminex analysis of cytokines
Blood was collected from anesthetized (Isoflurane, Abbott Lab-oratories, Chicago, IL, USA) mice via the inferior vena cava with a23-gauge needle. After collection, blood was transferred to CapiJectserum separator tubes (3T-MG; Terumo Medical Corp., Elkton, MD,USA), allowed to clot for 30 min, and centrifuged at 400g for10 min. The serum was collected and stored at �70 �C until used.
The Luminex protocol is a sandwich immunoassay systememploying beads. This novel method allows the simultaneousdetection of different cytokines in the Luminex-200 (LuminexCorp., Austin, TX), a dual-laser flow analyzer. Mouse serum sam-ples were analyzed for interleukin-1b (IL-1b), IL-6, IL-10, IL-12(p70), G-CSF, granulocyte macrophage-colony stimulating factor(GM-CSF), keratinocyte-derived chemokine (KC), and tumor necro-sis factor-a (TNF-a). A cytokine analysis kit was custom ordered(M200003JZX, Bio-Rad Inc., Hercules, CA) and included all neces-sary reagents for analysis. In brief, cytokine antibody-conjugatedbeads were added to each well of a flat-bottom 96-well plate(Bio-Rad Inc.). Serum samples were diluted 1:4 with the provideddiluent and pipetted into the wells, incubated and then washedusing a Bio-Plex Pro wash station (Bio-Rad Inc.). Then the diluteddetection antibody was added. After incubation for 1 h, the platewas washed again using the Bio-Plex Pro wash station to removeexcess detection antibody, and streptavidin–phycoerythrin wasadded. After final incubation and washing, the fluorochrome boundto magnetic beads was quantified in the Luminex-200, which wascalibrated using calibration microspheres. The median fluores-cence intensity of fluorochrome-conjugated antibody bound toindividual microspheres was derived from flow analysis of 50
412 P.K. Singh et al. / Cytokine 56 (2011) 411–421

Author's personal copy
microspheres per region. Cytokine quantification was plotted viastandard curves. The intensity of the fluorescence was directly pro-portional to the concentration of cytokine. Calculations were per-formed via Bio-Plex Manager software, version 5.0 (Bio-Rad Inc.).
2.6. Histopathology of jejunum for crypt enumeration
Jejunum was the primary tissue used for histopathologic analy-sis. Jejuna from TS-treated and control mice collected at differenttimes after 11 Gy irradiation were immersion fixed in a 20:1 vol-ume of fixative (Z-FIX�, Anatech Ltd., Battle Creek, MI, USA) to tis-sue for at least 24 h and up to 7 days. Fixed jejunum were routinelyprocessed using progressive dehydration of tissues in ethanol,cleaning in xylene, paraffin vacuum-infiltration, and paraffinembedding. Paraffin 4 lm cross sections of jejunum were stainedwith hematoxylin and eosin (H&E), and observed under a NikonEclipse TS100 microscope (Melville, NY, USA) equipped with theRetiga 2000R Q imaging camera (Surrey, BC, Canada). The cryptmicrocolony survival assay was performed as described by Withersand Elkind [24]. The circumference of a transverse cross-section ofthe intestine was used as a unit. The crypts of Lieberkühn (intesti-nal glands) were considered viable if they contained at least 10 epi-thelial cells (either columnar enterocytes or goblet cells), a lumenand at least one Paneth cell. The number of surviving crypts wascounted in each circumference. Six circumferences were scoredper mouse and 4 mice were used to generate each data point.
Five villi were measured in each circumference. The number ofvilli was scored in each circumference. The mitotic figures werecounted in high power fields (HPF), six HPF were scored per crosssection. Eight mice, six sections per mouse were used to generateeach data point.
2.7. Terminal deoxynucleotidyl transferase dUTP nick end labeling(TUNEL) assay
The paraffin sections of jejunum were used to detect frag-mented DNA in their nuclei to enumerate apoptotic cells. The jeju-num sections were stained with a TUNEL apoptosis detection kit(GenScript USA, Inc., Piscataway, NJ, USA) following the manufac-turer’s protocol and method described by Paries et al. [25]. Thefluorescein-labeled nucleotides bind with the DNA 30-OH endsusing terminal deoxynucleotidyl transferase. Cells with fluores-cence were observed under a Zeiss Axio Observer D1 fluorescencemicroscope with Axiovision 4.7 software (Göttingen, Germany).The number of apoptotic cells was counted in HPF. Eight HPF werescored in each section, six sections per mouse, and eight mice wereused to generate each data point.
2.8. Evaluation of cell proliferation by detecting Ki-67
To evaluate the G-CSF-mediated proliferative efficacy of TS, Ki-67 positive cells were evaluated in G-CSF antibody- or isotype-in-jected mice exposed to radiation. Jejunum samples from G-CSFantibody-treated and control mice were collected on days 4 and8 after 11 Gy irradiation, and immunohistochemistry was per-formed as described earlier [26,27]. The paraffin-embedded sec-tions of jejunum were used to detect the Ki-67. The jejunumsections were deparaffinized with xylene, dehydrated in 100% eth-anol and rehydrated by sequentially immersing the slides throughgraded ethanol washes (95%, 90%, 80% and 70%) for 3 min each atroom temperature. The endogenous peroxidase activity wasblocked by incubating slides for 10 min in 3% hydrogen peroxidein PBS. The antigen was retrieved by using retrievagen (BD Biosci-ences, San Jose, CA, USA). After a wash in PBS, sections were put inblocking solution (1X tris buffered saline with 0.1% tween-20 with3% goat serum) for 45 min at 37 �C and incubated with Ki-67
monoclonal mouse antibody (1:40; Dako, Glostrup, Denmark) for1 h in a humidified chamber. Slides were incubated with biotinyl-ated secondary antibody (BD Biosciences, San Jose, CA, USA) for45 min, and washed in PBS with 0.1% Tween-20. Ki-67 positivecells were detected by applying streptavidin-HRP (BD Biosciences,San Jose, CA, USA) for 30 min at room temperature and adding100 ll of DAB chromogen (BD Biosciences, San Jose, CA, USA).The slides were counter-stained in hematoxylin for 45 s, rinsedin water, dehydrated through four changes of alcohol (95%, 95%,100%, 100%), cleared in xylene, and mounted with mounting med-ium (Histomount; National Diagnostics, Charlotte, NC, USA). Ki-67positive cells were counted using a Nikon Eclipse TS100 micro-scope (Melville, NY, USA) equipped with the Retiga 2000R Q imag-ing camera (Surrey, BC, Canada). The number of Ki-67 positive cellswas counted in each crypt. Ten to twelve crypts were scored ineach section, six sections per mouse, and four mice were used togenerate each data point.
2.9. Statistical analysis
Means with standard error or percentage were reported, ifapplicable. Analysis of variance (ANOVA) was used to detect signif-icant differences among groups and over a period of time for a par-ticular group. If significance was found then a pair-wisecomparison via the Tukey–Kramer procedure was used to identifydifferences between specific groups. For survival data, a log-ranktest was used to compare survival curves. Fisher’s exact test wasused to compare 30 day survival with a Bonferroni correction usedto control the type I error if multiple comparison was used. The sig-nificance level was set at 5% for each test. All statistical tests weretwo-sided. Statistical software (PC SAS or SPSS) was used for statis-tical analyses.
3. Results
3.1. Radioprotective efficacy of TS
We demonstrated earlier that TS protects mice against a 9.2 Gy(LD90/30) dose of radiation inducing hematopoietic syndrome[20,22]. Here we studied the efficacy of TS against doses of radia-tion that caused GI syndrome. When mice were administered TS(400 mg/kg) and irradiated 24 h later with 11 Gy of 60Co c -radia-
0
20
40
60
80
100
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30
Surv
ival
(%)
Post-irradiation (d)
TS
Vehicle
TS + G-CSF ab
*
Fig. 1. Radioprotective efficacy of TS when administered 24 h prior to 60Co c-irradiation. CD2F1 male mice were injected sc with 400 mg/kg TS or vehicle andirradiated 24 h later with 11 Gy 60Co c-radiation (n = 24). G-CSF antibody wasinjected 16 h before irradiation (8 h after drug administration). Survival wasmonitored for 30 days. Mice treated with TS were significantly protected, withrespect to vehicle or TS plus G-CSF antibody-injected groups (⁄p < 0.001).
P.K. Singh et al. / Cytokine 56 (2011) 411–421 413

Author's personal copy
tion (LD60/10), 30 day survival protection was achieved, as shownin Fig. 1. The data in Fig. 1 was derived from two experiments, eachof which employed 12 mice in each group. TS protected 22 miceout of 24 (92%) while all 24 control mice receiving vehicle and ex-posed to 11 Gy of 60Co gamma-radiation died by day 13 after irra-diation. Deaths in the control group began on day 4 afterirradiation with 11 Gy. Two mice of the TS-treated group died ondays 13 and 15 (one each). There was a significant difference be-tween two groups in comparing survival curves. The differenceamong survivors between TS and vehicle groups was significant(p < 0.001, Fig. 1). The protective effect of TS was significantly abro-gated by G-CSF antibody administration. All mice receiving TS plusG-CSF antibody and exposed to 11 Gy of 60Co gamma-radiationdied by day 14 after irradiation. A significant difference in survivalwas also observed between the TS plus G-CSF antibody-treatedmice compared to only TS-treated mice (p < 0.001).
3.2. Induction of cytokines by TS in mice as determined by Luminexassay
Our ongoing studies with several radiation countermeasureshave shown a relationship between survival efficacy and an in-crease in cytokines in the circulating blood. We therefore wantedto determine if the observed survival benefit afforded by TS aftertotal-body irradiation is associated with increased levels of
cytokines in circulating blood. The dose response and time courseof cytokines, induced by TS, in circulating blood was determined.Mice were injected sc with TS at doses of 0, 50, 100, 200, 400,and 800 mg/kg. Blood samples for serum were collected 2, 4, 8,12, 24, 36, and 48 h after drug or vehicle administration. MultiplexLuminex assay was performed for IL-1b, IL-12(p70), G-CSF, GM-CSF, KC, and TNF-a.
As shown in Fig. 2, when mice were treated with TS we ob-served a significant induction of all cytokines tested. Maximumlevels of G-CSF as well as KC were induced 24 h after TS (200,400, and 800 mg/kg) administration. Forty-eight hours after TS(200, 400, and 800 mg/kg), maximum levels of IL-12 (p70) wereobserved. For IL-1b and TNF-a, maximum levels were observedat 12 h after TS injection. As the drug dose increased, the levelsof cytokines induced increased over time. IL-1 b, GM-CSF, IL-12(p70), and TNF-a were induced by TS but to a much lesser extentcompared to G-CSF or KC (Fig. 2). We also evaluated additionalcytokines but did not achieve significant levels (data notpresented).
3.3. Neutralization of TS-stimulated G-CSF by administration of aG-CSF antibody
We have reported the abrogation of the radioprotective efficacyof TS by administration of an exogenous G-CSF antibody to TS-trea
0
3000
6000
9000
12000
15000
2 h 4 h 8 h 12 h 24 h 36 h 48 h
Conc
entr
atio
n (p
g/m
l)
Time after drug administration
G-CSF * ***
*
*
*
*
**
0
100
200
300
400
2 h 4 h 8 h 12 h 24 h 36 h 48 h
Conc
entr
atio
n (p
g/m
l)
Time after drug administration
IL-1 β *
**
*
*
****
*
0
50
100
150
200
250
2 h 4 h 8 h 12 h 24 h 36 h 48 h
Con
cent
ratio
n (p
g/m
l)
Time after drug administration
IL-12 (p70)0 50100 200400 800
***
**** *
mg/kg
*
0
50
100
150
200
2 h 4 h 8 h 12 h 24 h 36 h 48 h
Conc
entr
atio
n (p
g/m
l)
Time after drug administration
GM-CSF*****
*
*
**
*
0
50
100
150
200
2 h 4 h 8 h 12 h 24 h 36 h 48 h
Conc
entr
atio
n (p
g/m
l)
Time after drug administration
TNF-α
***
**** *
**
0
4000
8000
12000
16000
2 h 4 h 8 h 12 h 24 h 36 h 48 h
Conc
entr
atio
n (p
g/m
l)
Time after drug administration
KC
*
*
*
*
**
Fig. 2. Time course and dose response of serum levels of cytokines after sc administration of TS. Six groups of CD2F1 mice were injected with various doses (0, 50, 100, 200,400, or 800 mg/kg) of TS. Blood samples were collected 2, 4, 8, 12, 24, 36, or 48 h after TS administration. Serum samples were analyzed by Luminex assay for cytokines. Errorbars indicate the standard error of the mean (SEM) for n = 8 of two independent experiments. ⁄ Denotes significant difference compared to vehicle (0 mg/kg TS) control group.
414 P.K. Singh et al. / Cytokine 56 (2011) 411–421
Author's personal copy
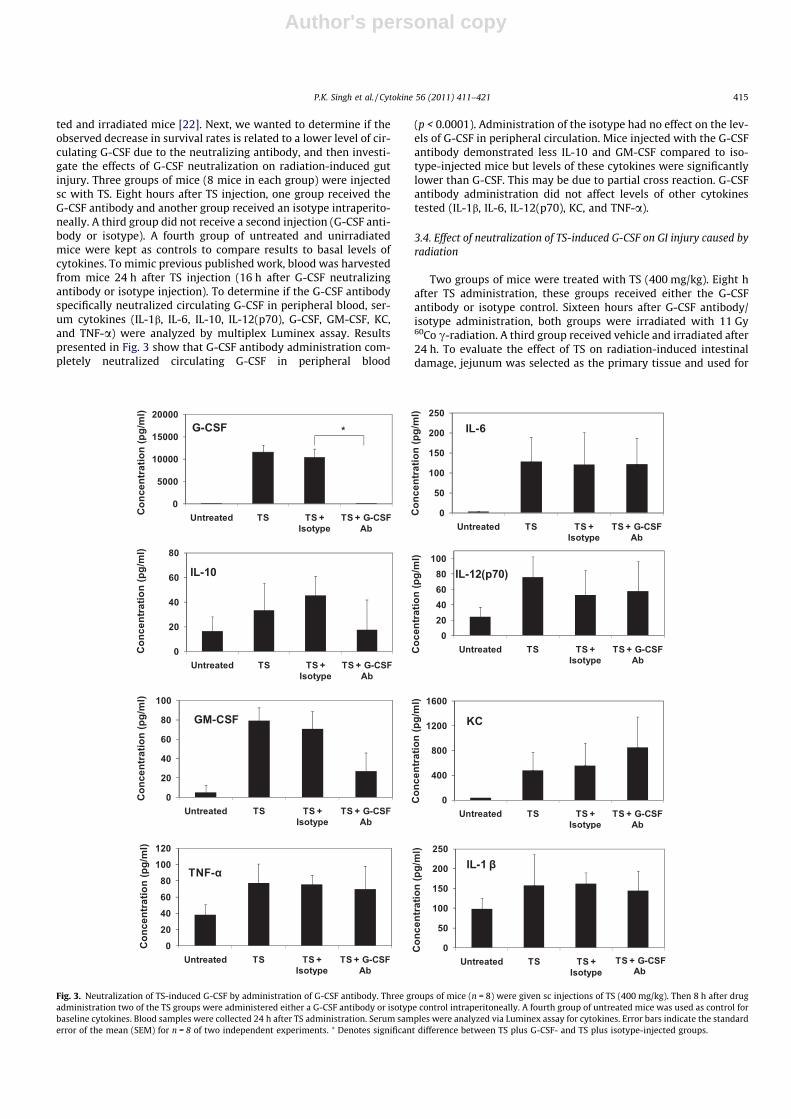
ted and irradiated mice [22]. Next, we wanted to determine if theobserved decrease in survival rates is related to a lower level of cir-culating G-CSF due to the neutralizing antibody, and then investi-gate the effects of G-CSF neutralization on radiation-induced gutinjury. Three groups of mice (8 mice in each group) were injectedsc with TS. Eight hours after TS injection, one group received theG-CSF antibody and another group received an isotype intraperito-neally. A third group did not receive a second injection (G-CSF anti-body or isotype). A fourth group of untreated and unirradiatedmice were kept as controls to compare results to basal levels ofcytokines. To mimic previous published work, blood was harvestedfrom mice 24 h after TS injection (16 h after G-CSF neutralizingantibody or isotype injection). To determine if the G-CSF antibodyspecifically neutralized circulating G-CSF in peripheral blood, ser-um cytokines (IL-1b, IL-6, IL-10, IL-12(p70), G-CSF, GM-CSF, KC,and TNF-a) were analyzed by multiplex Luminex assay. Resultspresented in Fig. 3 show that G-CSF antibody administration com-pletely neutralized circulating G-CSF in peripheral blood
(p < 0.0001). Administration of the isotype had no effect on the lev-els of G-CSF in peripheral circulation. Mice injected with the G-CSFantibody demonstrated less IL-10 and GM-CSF compared to iso-type-injected mice but levels of these cytokines were significantlylower than G-CSF. This may be due to partial cross reaction. G-CSFantibody administration did not affect levels of other cytokinestested (IL-1b, IL-6, IL-12(p70), KC, and TNF-a).
3.4. Effect of neutralization of TS-induced G-CSF on GI injury caused byradiation
Two groups of mice were treated with TS (400 mg/kg). Eight hafter TS administration, these groups received either the G-CSFantibody or isotype control. Sixteen hours after G-CSF antibody/isotype administration, both groups were irradiated with 11 Gy60Co c-radiation. A third group received vehicle and irradiated after24 h. To evaluate the effect of TS on radiation-induced intestinaldamage, jejunum was selected as the primary tissue and used for
0
50
100
150
200
250
Untreated TS TS + Isotype
TS + G-CSF Ab
Con
cent
ratio
n (p
g/m
l)IL-6
0
20
40
60
80
Untreated TS TS + Isotype
TS + G-CSF Ab
Con
cent
ratio
n (p
g/m
l)
IL-10
020406080
100
Untreated TS TS + Isotype
TS + G-CSF Ab
Coc
entra
tion
(pg/
ml)
IL-12(p70)
0
20
40
60
80
100
Untreated TS TS + Isotype
TS + G-CSF Ab
Con
cent
ratio
n (p
g/m
l)
GM-CSF
0
400
800
1200
1600
Untreated TS TS + Isotype
TS + G-CSF
TS + G-CSF
Ab
Con
cent
ratio
n (p
g/m
l)
KC
020406080
100120
Untreated TS TS + Isotype
TS + G-CSF Ab
Con
cent
ratio
n (p
g/m
l)
TNF-α
0
50
100
150
200
250
Untreated TS TS + Isotype Ab
Conc
entr
atio
n (p
g/m
l)
IL-1 β
0
5000
10000
15000
20000
Untreated TS TS + Isotype
TS + G-CSF Ab
Conc
entr
atio
n (p
g/m
l)
G-CSF *
Fig. 3. Neutralization of TS-induced G-CSF by administration of G-CSF antibody. Three groups of mice (n = 8) were given sc injections of TS (400 mg/kg). Then 8 h after drugadministration two of the TS groups were administered either a G-CSF antibody or isotype control intraperitoneally. A fourth group of untreated mice was used as control forbaseline cytokines. Blood samples were collected 24 h after TS administration. Serum samples were analyzed via Luminex assay for cytokines. Error bars indicate the standarderror of the mean (SEM) for n = 8 of two independent experiments. ⁄ Denotes significant difference between TS plus G-CSF- and TS plus isotype-injected groups.
P.K. Singh et al. / Cytokine 56 (2011) 411–421 415

Author's personal copy
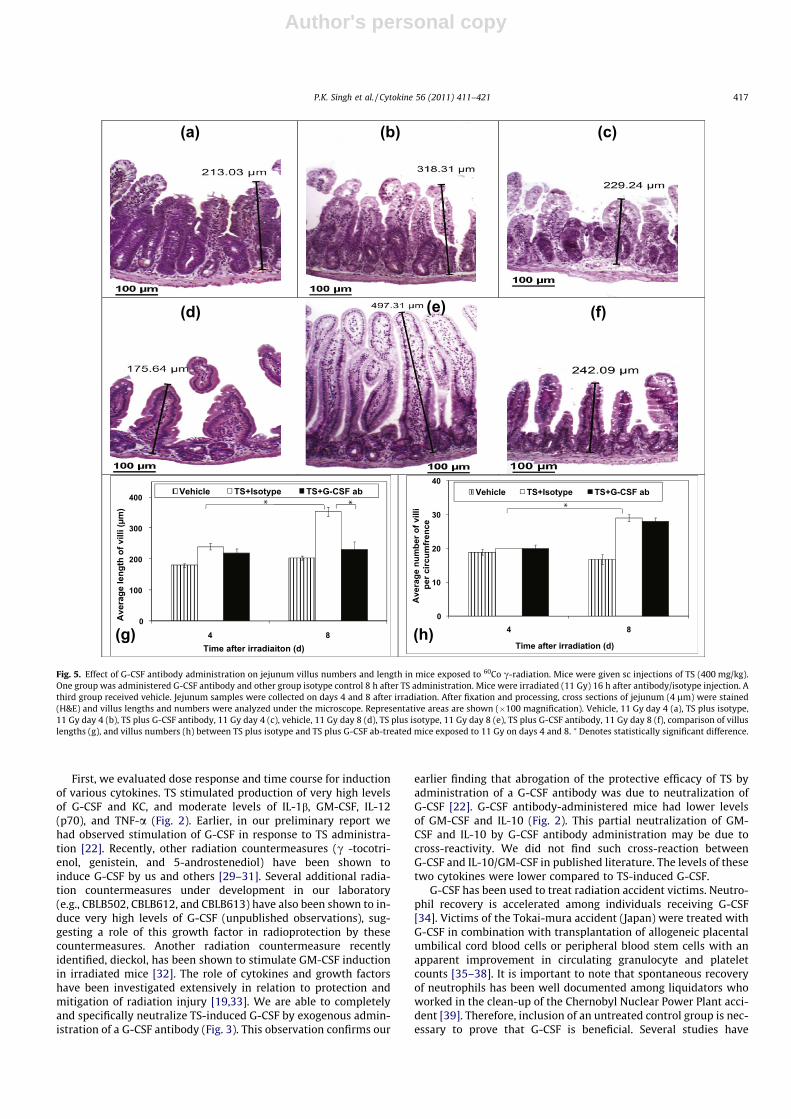
histopathological analysis. Jejunum samples were harvested ondays 4 and 8 after irradiation for histopathology and analyzed for(a) crypt number, (b) villus numbers and length, (c) mitotic figures,(d) apoptosis, and (e) cell proliferation.
Photomicrographs shown in Fig. 4 demonstrated crypts in threegroups on days 4 and 8. On day 8 after irradiation, isotype-injectedmice had higher numbers of crypts compared to a day 4 sample ofthe same treatment group (p < 0.005) (Fig. 4g). Similar increaseswere observed for villus length (p < 0.005) (Fig. 5g) and villus num-bers (p < 0.0001) and (Fig. 5h). Such increases for crypts and villiwere not observed in the G-CSF antibody-injected groups. On day8 after irradiation, isotype-injected mice had longer villi comparedto G-CSF antibody-injected mice (p < 0.005) (Fig. 5g). We have alsopresented the results of vehicle control group to demonstrate theextent of attenuation by G-CSF antibody.
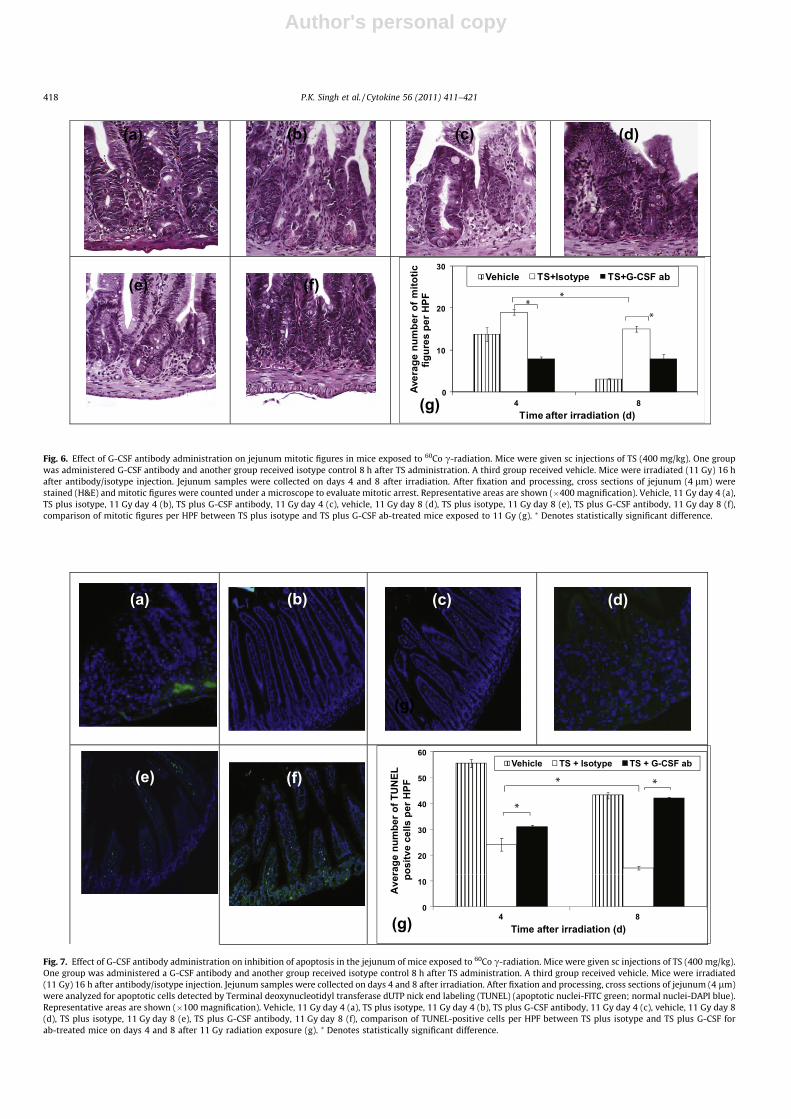
Histopathology of jejunum also was scored for mitotic figures(Fig. 6). There was a significantly higher number of mitotic figuresin isotype control mice compared to G-CSF antibody-injected miceon days 4 (p < 0.0001) and 8 (p < 0.0001) (Fig. 6g). The number ofmitotic figures decreased in isotype-injected mice on day 8 com-pared to day 4 (p < 0.005). There was no change in vehicle groupon days 4 and 8.
3.5. Effect of neutralization of TS induced G-CSF on radiation-inducedapoptosis in jejunum
To evaluate the effect of a G-CSF antibody on radiation-inducedapoptosis in TS-treated mice, jejunum samples were collected fromboth groups of mice (TS-treated and those injected with G-CSFantibody or isotype) on days 4 and 8 after irradiation. Histologyslides were stained with the TUNEL assay to detect apoptotic cells(Fig. 7) as described above. Significantly higher numbers of TUNEL-positive cells per HPF were observed on days 4 (p < 0.05) and 8(p < 0.0001) in G-CSF antibody-injected mice compared toisotype-injected mice. This result suggests an increase of apoptotic
cells in response to irradiation in G-CSF antibody-injected mice.The isotype-injected mice had lower numbers of apoptotic cellson day 8 compared to day 4 (p < 0.02). Vehicle-treated group hadhigher number of apoptotic cells compared to TS plus isotype-trea-ted group.
3.6. Effect of neutralization of TS-induced G-CSF on regeneration ofcrypt cells in irradiated mice
To evaluate the effect of TS-induced G-CSF on crypt cell regen-erating potential, Ki-67 assays were carried out with jejunum sam-ples of TS-treated and irradiated mice receiving either the G-CSFantibody or isotype (Fig. 8). Higher numbers of Ki-67-positive cellsper crypt were observed in TS-treated mice receiving isotype com-pared to mice receiving the G-CSF antibody on days 4 (p < 0.0001)and 8 (p < 0.0001) (Fig. 8g). Unlike G-CSF antibody-injected mice,isotype-injected mice had higher numbers of proliferating cellson day 8 compared to day 4, suggesting an increase of proliferatingcells over time in the absence of the G-CSF antibody (p < 0.0001).Vehicle treated group was comparable to the group receiving TSand G-CSF antibody.
4. Discussion and conclusion
We have observed that TS protects CD2F1 mice against a radia-tion dose of 11 Gy (capable of inducing GI syndrome). Recently, an-other tocol (c-tocotrienol) also has been shown to protect micefrom radiation-induced GI injury supporting our results [28]. Wereported earlier that the radioprotective efficacy of TS in micereceiving 9.2 Gy radiation can be abrogated by G-CSF antibodyadministration [22]. Our interest was to evaluate whether admin-istration of a G-CSF antibody abrogates the radioprotective efficacyof TS on GI tissue in mice irradiated with higher doses of radiationthat induce GI damage. The protective effect of TS against 11 Gywas significantly abrogated by G-CSF antibody.
0
20
40
60
80
100
120
84Time after irradiation (d)
Vehicle TS + Isotype TS + G-CSF ab
Ave
rage
num
ber o
f cry
pts
per
circ
umfr
ence
(a) (b) (c) (d)
(e) (f)
(g)
Fig. 4. Effect of G-CSF antibody administration on jejunum crypt number in mice exposed to 60Co c-radiation. Mice were given sc injections of TS (400 mg/kg) or vehicle. OneTS-injected group was administered a G-CSF antibody and another TS-injected group received an isotype control 8 h after TS administration. A third group received vehicle.Mice were irradiated (11 Gy) 16 h after antibody/isotype injection. Jejunum samples were collected on days 4 and 8 after irradiation. After fixation and processing, crosssections of jejunum (4 lm) were stained (H&E) and crypts were counted under a microscope. Representative areas are shown (�100 magnification). Vehicle, 11 Gy day 4 (a),TS plus isotype, 11 Gy day 4 (b), TS plus G-CSF antibody, 11 Gy day 4 (c), vehicle, 11 Gy day 8 (d), TS plus isotype, 11 Gy day 8 (e), TS plus G-CSF antibody, 11 Gy day 8 (f),comparison of crypt numbers between TS plus isotype and TS plus G-CSF ab-treated mice exposed to 11 Gy on day 4 and 8 (g). ⁄ Denotes statistically significant difference.
416 P.K. Singh et al. / Cytokine 56 (2011) 411–421
Author's personal copy
First, we evaluated dose response and time course for inductionof various cytokines. TS stimulated production of very high levelsof G-CSF and KC, and moderate levels of IL-1b, GM-CSF, IL-12(p70), and TNF-a (Fig. 2). Earlier, in our preliminary report wehad observed stimulation of G-CSF in response to TS administra-tion [22]. Recently, other radiation countermeasures (c -tocotri-enol, genistein, and 5-androstenediol) have been shown toinduce G-CSF by us and others [29–31]. Several additional radia-tion countermeasures under development in our laboratory(e.g., CBLB502, CBLB612, and CBLB613) have also been shown to in-duce very high levels of G-CSF (unpublished observations), sug-gesting a role of this growth factor in radioprotection by thesecountermeasures. Another radiation countermeasure recentlyidentified, dieckol, has been shown to stimulate GM-CSF inductionin irradiated mice [32]. The role of cytokines and growth factorshave been investigated extensively in relation to protection andmitigation of radiation injury [19,33]. We are able to completelyand specifically neutralize TS-induced G-CSF by exogenous admin-istration of a G-CSF antibody (Fig. 3). This observation confirms our
earlier finding that abrogation of the protective efficacy of TS byadministration of a G-CSF antibody was due to neutralization ofG-CSF [22]. G-CSF antibody-administered mice had lower levelsof GM-CSF and IL-10 (Fig. 2). This partial neutralization of GM-CSF and IL-10 by G-CSF antibody administration may be due tocross-reactivity. We did not find such cross-reaction betweenG-CSF and IL-10/GM-CSF in published literature. The levels of thesetwo cytokines were lower compared to TS-induced G-CSF.
G-CSF has been used to treat radiation accident victims. Neutro-phil recovery is accelerated among individuals receiving G-CSF[34]. Victims of the Tokai-mura accident (Japan) were treated withG-CSF in combination with transplantation of allogeneic placentalumbilical cord blood cells or peripheral blood stem cells with anapparent improvement in circulating granulocyte and plateletcounts [35–38]. It is important to note that spontaneous recoveryof neutrophils has been well documented among liquidators whoworked in the clean-up of the Chernobyl Nuclear Power Plant acci-dent [39]. Therefore, inclusion of an untreated control group is nec-essary to prove that G-CSF is beneficial. Several studies have
0
100
200
300
400
84
Ave
rage
leng
th o
f vill
i (µm
)
Time after irradiaiton (d)
Vehicle TS+Isotype TS+G-CSF ab
0
10
20
30
40
84
Ave
rage
num
ber o
f vill
ipe
r circ
umfr
ence
Time after irradiation (d)
Vehicle TS+Isotype TS+G-CSF ab
(a) (b) (c)
(d) (e) (f)
(g) (h)
Fig. 5. Effect of G-CSF antibody administration on jejunum villus numbers and length in mice exposed to 60Co c-radiation. Mice were given sc injections of TS (400 mg/kg).One group was administered G-CSF antibody and other group isotype control 8 h after TS administration. Mice were irradiated (11 Gy) 16 h after antibody/isotype injection. Athird group received vehicle. Jejunum samples were collected on days 4 and 8 after irradiation. After fixation and processing, cross sections of jejunum (4 lm) were stained(H&E) and villus lengths and numbers were analyzed under the microscope. Representative areas are shown (�100 magnification). Vehicle, 11 Gy day 4 (a), TS plus isotype,11 Gy day 4 (b), TS plus G-CSF antibody, 11 Gy day 4 (c), vehicle, 11 Gy day 8 (d), TS plus isotype, 11 Gy day 8 (e), TS plus G-CSF antibody, 11 Gy day 8 (f), comparison of villuslengths (g), and villus numbers (h) between TS plus isotype and TS plus G-CSF ab-treated mice exposed to 11 Gy on days 4 and 8. ⁄ Denotes statistically significant difference.
P.K. Singh et al. / Cytokine 56 (2011) 411–421 417
Author's personal copy
0
10
20
30
84Av
erag
e nu
mbe
r of m
itotic
fig
ures
per
HPF
Time after irradiation (d)
Vehicle TS+Isotype TS+G-CSF ab
(a) (b) (c) (d)
(g)
(e) (f)
Fig. 6. Effect of G-CSF antibody administration on jejunum mitotic figures in mice exposed to 60Co c-radiation. Mice were given sc injections of TS (400 mg/kg). One groupwas administered G-CSF antibody and another group received isotype control 8 h after TS administration. A third group received vehicle. Mice were irradiated (11 Gy) 16 hafter antibody/isotype injection. Jejunum samples were collected on days 4 and 8 after irradiation. After fixation and processing, cross sections of jejunum (4 lm) werestained (H&E) and mitotic figures were counted under a microscope to evaluate mitotic arrest. Representative areas are shown (�400 magnification). Vehicle, 11 Gy day 4 (a),TS plus isotype, 11 Gy day 4 (b), TS plus G-CSF antibody, 11 Gy day 4 (c), vehicle, 11 Gy day 8 (d), TS plus isotype, 11 Gy day 8 (e), TS plus G-CSF antibody, 11 Gy day 8 (f),comparison of mitotic figures per HPF between TS plus isotype and TS plus G-CSF ab-treated mice exposed to 11 Gy (g). ⁄ Denotes statistically significant difference.
0
10
20
30
40
50
60
84
Ave
rage
num
ber o
f TU
NEL
po
sitv
e ce
lls p
er H
PF
Time after irradiation (d)
Vehicle TS + Isotype TS + G-CSF ab
(g)
(a) (b) (c) (d)
(e) (f)(e)
(g)
Fig. 7. Effect of G-CSF antibody administration on inhibition of apoptosis in the jejunum of mice exposed to 60Co c-radiation. Mice were given sc injections of TS (400 mg/kg).One group was administered a G-CSF antibody and another group received isotype control 8 h after TS administration. A third group received vehicle. Mice were irradiated(11 Gy) 16 h after antibody/isotype injection. Jejunum samples were collected on days 4 and 8 after irradiation. After fixation and processing, cross sections of jejunum (4 lm)were analyzed for apoptotic cells detected by Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) (apoptotic nuclei-FITC green; normal nuclei-DAPI blue).Representative areas are shown (�100 magnification). Vehicle, 11 Gy day 4 (a), TS plus isotype, 11 Gy day 4 (b), TS plus G-CSF antibody, 11 Gy day 4 (c), vehicle, 11 Gy day 8(d), TS plus isotype, 11 Gy day 8 (e), TS plus G-CSF antibody, 11 Gy day 8 (f), comparison of TUNEL-positive cells per HPF between TS plus isotype and TS plus G-CSF forab-treated mice on days 4 and 8 after 11 Gy radiation exposure (g). ⁄ Denotes statistically significant difference.
418 P.K. Singh et al. / Cytokine 56 (2011) 411–421

Author's personal copy
demonstrated that G-CSF significantly shorten the period of neu-tropenia in irradiated rhesus macaques, canines, and mice [40–43]. Our study supports the previously observed beneficial effectsof G-CSF in protecting against radiation injury. G-CSF has been in-cluded in radiation stockpiles developed by the US Centers for Dis-ease Control and Prevention and by the World Health Organizationfor use during a nuclear disaster.
The above observations prompted us to further investigate therole of TS-induced G-CSF in radioprotection. We demonstrated thatthe radioprotective efficacy of TS in the GI of mice is abrogated by aG-CSF neutralizing antibody, suggesting that TS-induced G-CSFplays an important role in radioprotection offered by TS. TS-in-jected mice receiving a G-CSF antibody displayed greater GI injury(based on crypt, villi, mitotic figures, apoptotic cells, and cryptregeneration) compared to mice receiving an isotype. TS-treatedmice receiving isotype had higher numbers of crypts, villi, mitoticfigures, and Ki-67 positive cells compared to TS-treated and G-CSFantibody-injected mice. In addition, TS-treated mice receiving iso-type had lower numbers of apoptotic cells (TUNEL positive cells)compared to TS-treated and G-CSF antibody-injected mice, sug-gesting the abrogation of the radioprotective efficacy of TS by aG-CSF antibody. In other words, TS decreases radiation-inducedapoptosis and this effect is G-CSF mediated. Administration ofG-CSF antibody reverses the positive effect of TS described above.
It is important to note that numbers of surviving crypts afterexposure to various doses of radiation depends on the strain ofmice as radiosensitivity of different strains varies. There are pub-lished reports demonstrating that the crypt number per circumfer-ence was less than 20 at 3.5 days after irradiation at 8–12 Gy inICR, C3H/HeJ, and BDF1 mice [44–46]. There are also reports withother strains of mice (CD2F1, C57BL/6 and DDO/Lee mice)
demonstrating significantly higher number of crypts when ex-posed to 7–15 Gy of radiation [47–49]. Yoon et al. reported 83and 49 crypts per circumference in DDO/Lee mice exposed to 9and 15 Gy, respectively [49]. In our laboratory, we have consis-tently observed approximately 70 crypts per circumference inCD2F1 and C57BL/6 mice (unpublished observation) exposed to11 Gy 60Co gamma-radiation.
We have detected Ki-67, a nuclear protein expressed in allphases of the cell cycle except the resting phase, by immunohisto-chemistry using Ki-67 monoclonal mouse antibody to score prolif-erating cells. Quantitative comparison of bromodeoxyuridine(BrdU) and Ki-67-positive cells showed 50% higher numbers ofthe latter in earlier studies [26,27]. This is expected because BrdUcan be incorporated into DNA only during the S-phase of the mito-tic process, whereas Ki-67 is expressed for its whole duration.
The mechanism of the radioprotection by TS through G-CSF isnot well understood. Currently our laboratory is investigating therole of TS-induced G-CSF in radioprotection using in vivo andin vitro (CD34+ and human osteoblast cells) models. We have ob-served that use of G-CSF antibody exacerbates the effects of radia-tion exposure, and G-CSF antibody-treated mice die earlier thanuntreated mice (unpublished observation). This may be due tothe neutralization of G-CSF induced in response to irradiationwhich further suggests the beneficial role of G-CSF in radiation in-jury. Recently, we have reported that radiation exposure inducesG-CSF production in mice [30]. TS may be activating the inductionof nuclear factor-jB which may be responsible for induction ofG-CSF. We are trying to abrogate the effects of TS on survival andG-CSF induction by using small interfering RNA to inhibit NF-jBgene expression in CD34+ cells. G-CSF is known to activate severalsignaling pathways [50], promote survival [51,52], proliferation
0
5
10
15
20
25
30
8yaD4yaD
Ave
rage
num
ber o
f Ki-6
7 po
sitv
e ce
lls p
er c
rypt
Time after irradiation (d)
Veh + 11 Gy TS + Isotype TS + GCSF ab
*
*
(b)(a) (c) (d)
(e) (f)
(g)
Fig. 8. Effect of G-CSF antibody administration on regeneration of crypt cells in mice exposed to 60Co c-radiation. Mice were given sc injections of TS (400 mg/kg). One groupwas administered G-CSF antibody and another group received isotype control 8 h after TS administration. Mice were irradiated (11 Gy) 16 h after antibody/isotype injection.A third group received vehicle. Jejunum samples were collected on days 4 and 8 after irradiation. After fixation and processing, cross sections of jejunum (4 lm) wereanalyzed for Ki-67 by immunohistochemistry. Representative areas are shown (�200 magnification). Vehicle, 11 Gy day 4 (a), TS plus isotype, 11 Gy day 4 (b), TS plus G-CSFantibody, vehicle, 11 Gy day 8 (c), vehicle, 11 Gy day 8 (d), TS plus isotype, 11 Gy day 8 (e), TS plus G-CSF ab, 11 Gy day 8 (f), comparison of Ki-67 positive cells/crypt betweenTS plus isotype and TS plus G-CSF for ab-treated mice exposed to 11 Gy, days 4 and 8 (g). ⁄ Denotes statistically significant difference.
P.K. Singh et al. / Cytokine 56 (2011) 411–421 419

Author's personal copy
[53], differentiation [54], and mobilization of hematopoietic stemand progenitor cells [55–57]. G-CSF has also been reported to inhi-bit apoptosis [41,58–60].
Results presented in this communication support the positionthat TS is a promising radiation countermeasure and potent stim-ulator of G-CSF, and its radioprotective efficacy is mediatedthrough G-CSF. It will be interesting to investigate the effect ofG-CSF antibody on signal transduction pathway in irradiatedanimals.
Acknowledgements
We gratefully acknowledge Dr. Cara Olsen for her statisticalhelp, and AFRRI’s Veterinary Sciences Department and CobaltRadiation Facility for their support. The authors also acknowledgeSSGT Asline Khnachar for performing histopathology of jejunumcells. This study was supported by an AFRRI intramural researchgrant (RAB2CZ) to VKS. The authors are thankful to Dr. ChristopherR. Lissner, Scientific Director, and Dr. John R. Gilstad, DepartmentHead, for helpful discussions, and to Dr. Mark H. Whitnall, Counter-measures Program Advisor, for critical review of this manuscript.
References
[1] Benjamin GC, McGeary M, McCutchen SR. Assessing medical preparedness torespond to a terrorist nuclear event: workshop report. Washington, D.C.: TheNational Academies Press; 2009.
[2] Singh VK, Seed TM. Radiation effects. In: Roy MJ, editor. Physician’s guide toterrorist attack. Totowa: Humana Press; 2003. p. 339–62.
[3] Waselenko JK, MacVittie TJ, Blakely WF, Pesik N, Wiley AL, Dickerson WE, et al.Medical management of the acute radiation syndrome: recommendations ofthe strategic national stockpile radiation working group. Ann Intern Med2004;140:1037–51.
[4] Pellmar TC, Rockwell S. Priority list of research areas for radiological nuclearthreat countermeasures. Radiat Res 2005;163:115–23.
[5] Hosseinimehr SJ. Trends in the development of radioprotective agents. DrugDiscov Today 2007;12:794–805.
[6] Nair CK, Devi PU, Shimanskaya R, Kunugita N, Murase H, Gu YH, et al. Watersoluble vitamin E (TMG) as a radioprotector. Indian J Exp Biol2003;41:1365–71.
[7] Devi PU. Withania somnifera Dunal (Ashwagandha): potential plant source of apromising drug for cancer chemotherapy and radiosensitization. Indian J ExpBiol 1996;34:927–32.
[8] Yoshida Y, Niki E, Noguchi N. Comparative study on the action of tocopherolsand tocotrienols as antioxidant: chemical and physical effects. Chem PhysLipids 2003;123:63–75.
[9] Brigelius-Flohe R, Traber MG. Vitamin E: function and metabolism. FASEBJournal 1999;13:1145–55.
[10] Traber MG. Vitamin E regulatory mechanisms. Ann Rev Nutr 2007;27:347–62.[11] Singh VK, Shafran RL, Jackson III WE, Seed TM, Kumar KS. Induction of
cytokines by radioprotective tocopherol analogs. Exp Mol Pathol2006;81:55–61.
[12] Sen CK, Khanna S, Roy S. Tocotrienols: Vitamin E beyond tocopherols. Life Sci2006;78:2088–98.
[13] Sen CK, Khanna S, Rink C, Roy S. Tocotrienols: the emerging face of naturalvitamin E. Vitam Horm 2007;76:203–61.
[14] Atkinson J, Epand RF, Epand RM. Tocopherols and tocotrienols in membranes:a critical review. Free Rad Biol Med 2008;44:739–64.
[15] Ghosh SP, Kulkarni S, Hieber K, Toles R, Romanyukha L, Kao TC, et al. Gamma-tocotrienol, a tocol antioxidant as a potent radioprotector. Int J Radiat Biol2009;85:598–606.
[16] Beekman R, Touw IP. G-CSF and its receptor in myeloid malignancy. Blood2010;115:5131–6.
[17] Souza LM, Boone TC, Gabrilove J, Lai PH, Zsebo KM, Murdock DC, et al.Recombinant human granulocyte colony-stimulating factor: effects on normaland leukemic myeloid cells. Science 1986;232:61–5.
[18] Dainiak N. Rationale and recommendations for treatment of radiation injurywith cytokines. Health Phys 2010;98:838–42.
[19] Dainiak N, Waselenko JK, Armitage JO, MacVittie TJ, Farese AM. Thehematologist and radiation casualties. Hematology Am Soc Hematol EducProgram 2003:473–96.
[20] Singh VK, Brown DS, Kao TC. Tocopherol succinate: a promising radiationcountermeasure. Int Immunopharmacol 2009;9:1423–30.
[21] Singh VK, Parekh VI, Brown DS, Kao TC, Mog SR. Tocopherol succinate:modulation of antioxidant enzymes and oncogene expression, andhematopoietic recovery. Int J Radiat Oncol Biol Phys 2011;79:571–8.
[22] Singh VK, Brown DS, Kao TC. Alpha-tocopherol succinate protects mice fromgamma-radiation by induction of granulocyte-colony stimulating factor. Int JRadiat Biol 2010;86:12–21.
[23] Singh VK, Brown DS, Kao TC, Seed TM. Preclinical development of a bridgingtherapy for radiation casualties. Exp Hematol 2010;38:61–70.
[24] Withers HR, Elkind MM. Microcolony survival assay for cells of mouseintestinal mucosa exposed to radiation. Int J Radiat Biol Relat Stud Phys ChemMed 1970;17:261–7.
[25] Paris F, Fuks Z, Kang A, Capodieci P, Juan G, Ehleiter D, et al. Endothelialapoptosis as the primary lesion initiating intestinal radiation damage in mice.Science 2001;293:293–7.
[26] Kee N, Sivalingam S, Boonstra R, Wojtowicz JM. The utility of Ki-67 and BrdUas proliferative markers of adult neurogenesis. J Neurosci Methods2002;115:97–105.
[27] Scholzen T, Gerdes J. The Ki-67 protein: from the known and the unknown. JCell Physiol 2000;182:311–22.
[28] Berbee M, Fu Q, Boerma M, Wang J, Kumar KS, Hauer-Jensen M. Gamma-Tocotrienol ameliorates intestinal radiation injury and reduces vascularoxidative stress after total-body irradiation by an HMG-CoA reductase-dependent mechanism. Radiat Res 2009;171:596–605.
[29] Singh VK, Grace MB, Jacobsen KO, Chang CM, Parekh VI, Inal CE, et al.Administration of 5-androstenediol to mice. pharmacokinetics and cytokinegene expression. Exp Mol Pathol 2008;84:178–88.
[30] Singh VK, Grace MB, Parekh VI, Whitnall MH, Landauer MR. Effects of genisteinadministration on cytokine induction in whole-body gamma irradiated mice.Int Immunopharmacol 2009;9:1401–10.
[31] Kulkarni S, Ghosh SP, Hauer-Jensen M, Kumar KS. Hematological targets ofradiation damage. Curr Drug Targets 2010;11:1375–85.
[32] Park E, Ahn G, Yun JS, Kim MJ, Bing SJ, Kim DS, et al. Dieckol rescues mice fromlethal irradiation by accelerating hemopoiesis and curtailingimmunosuppression. Int J Radiat Biol 2010;86:848–59.
[33] Singh VK, Yadav VS. Role of cytokines and growth factors in radioprotection.Exp Mol Pathol 2005;78:156–69.
[34] Wingard JR, Dainiak N. Treatment of radiation injury in the adult. Up-to-Date,Version 17.2. Available at: http://www.uptodate.com; 2010 [accessed March2010].
[35] Asano S. Multi-organ involvement: lessons from the experience of one victimof the Tokai-mura criticality accident. Br J Radiol Suppl 2005;27:9–12.
[36] Maekawa K. Overview of medical care for highly exposed victims in theTokaimura accident. In: Ricks RC, Berger ME, O’Hara FM, editors. The medicalbasis for radiation accident preparedness. The clinical care of victime. NewYork: Parthenon; 2002. p. 313–8.
[37] Ishii T, Futami S, Nishida M, Suzuki T, Sakamoto T, Suzuki N, et al. Brief noteand evaluation of acute-radiation syndrome and treatment of a Tokai-muracriticality accident patient. J Radiat Res (Tokyo) 2001;42(Suppl.):S167–82.
[38] Igaki H, Nakagawa K, Uozaki H, Akahane M, Hosoi Y, Fukayama M, et al.Pathological changes in the gastrointestinal tract of a heavily radiation-exposed worker at the Tokai-mura criticality accident. J Radiat Res (Tokyo)2008;49:55–62.
[39] Baranov AE, Guskova AK, Nadejina NM, Nugis VY. Chernobyl experiences:biological indications of exposure to ionizing radiation. Stem Cells1995;13(Suppl):69–77.
[40] Herodin F, Grenier N, Drouet M. Revisiting therapeutic strategies in radiationcasualties. Exp Hematol 2007;35:28–33.
[41] Herodin F, Roy L, Grenier N, Delaunay C, Bauge S, Vaurijoux A, et al.Antiapoptotic cytokines in combination with pegfilgrastim soon afterirradiation mitigates myelosuppression in nonhuman primates exposed tohigh irradiation dose. Exp Hematol 2007;35:1172–81.
[42] Mac Vittie TJ, Monroy RL, Vigneulle RM. The relative biological effectiveness ofmixed fission-neutron-gamma radiation on the hematopoietic syndrome inthe canine: effect of therapy on survival. Radiat Res 1991;128:S29.
[43] Neelis KJ, Dubbelman YD, Qingliang L, Thomas GR, Eaton DL, Wagemaker G.Simultaneous administration of TPO and G-CSF after cytoreductive treatmentof rhesus monkeys prevents thrombocytopenia, accelerates platelet and redcell reconstitution, alleviates neutropenia, and promotes the recovery ofimmature bone marrow cells. Exp Hematol 1997;25:1084–93.
[44] Moon C, Kim SH, Kim JC, Hyun JW, Lee NH, Park JW, et al. Protective effect ofphlorotannin components phloroglucinol and eckol on radiation-inducedintestinal injury in mice. Phytother Res 2008;22:238–42.
[45] Oh H, Seong J, Kim W, Park S, Koom WS, Cho NH, et al. Recombinant humanepidermal growth factor (rhEGF) protects radiation-induced intestine injury inmurine system. J Radiat Res (Tokyo) 2010;51:535–41.
[46] Potten CS, Booth D, Haley JD. Pretreatment with transforming growth factorbeta-3 protects small intestinal stem cells against radiation damage in vivo. BrJ Cancer 1997;75:1454–9.
[47] Park E, Hwang I, Song JY, Jee Y. Acidic polysaccharide of Panax ginseng as adefense against small intestinal damage by whole-body gamma irradiation ofmice. Acta Histochem 2011;113:19–23.
[48] Fu Q, Berbee M, Boerma M, Wang J, Schmid HA, Hauer-Jensen M. Thesomatostatin analog SOM230 (pasireotide) ameliorates injury of the intestinalmucosa and increases survival after total-body irradiation by inhibitingexocrine pancreatic secretion. Radiat Res 2009;171:698–707.
[49] Yoon SC, Park JM, Jang HS, Shinn KS, Bahk YW. Radioprotective effect ofcaptopril on the mouse jejunal mucosa. Int J Radiat Oncol Biol Phys1994;30:873–8.
420 P.K. Singh et al. / Cytokine 56 (2011) 411–421

Author's personal copy
[50] Klocke R, Kuhlmann MT, Scobioala S, Schabitz WR, Nikol S. Granulocytecolony-stimulating factor (G-CSF) for cardio- and cerebrovascular regenerativeapplications. Curr Med Chem 2008;15:968–77.
[51] Harada M, Qin Y, Takano H, Minamino T, Zou Y, Toko H, et al. G-CSF preventscardiac remodeling after myocardial infarction by activating the Jak-Statpathway in cardiomyocytes. Nat Med 2005;11:305–11.
[52] Kuhlmann MT, Kirchhof P, Klocke R, Hasib L, Stypmann J, Fabritz L, et al. G-CSF/SCF reduces inducible arrhythmias in the infarcted heart potentially viaincreased connexin43 expression and arteriogenesis. J Exp Med2006;203:87–97.
[53] Schneider A, Kruger C, Steigleder T, Weber D, Pitzer C, Laage R, et al. Thehematopoietic factor G-CSF is a neuronal ligand that counteracts programmedcell death and drives neurogenesis. J Clin Invest 2005;115:2083–98.
[54] Wadhwa M, Thorpe R. Haematopoietic growth factors and their therapeuticuse. Thromb Haemost 2008;99:863–73.
[55] Singh VK, Singh PK, Wise SY, Seed TM. Mobilized progenitor cells as a bridgingtherapy for radiation casualties: A brief review of tocopherol succinate-basedapproaches. Int Immunopharmacol 2011;11:839–44.
[56] Luider J, Brown C, Selinger S, Quinlan D, Karlsson L, Ruether D, et al. Factorsinfluencing yields of progenitor cells for allogeneic transplantation:optimization of G-CSF dose, day of collection, and duration of leukapheresis.J Hematother 1997;6:575–80.
[57] Damon LE. Mobilization of hematopoietic stem cells into the peripheral blood.Expert Rev Hematol 2009;2:717–33.
[58] Jun HS, Lee YM, Song KD, Mansfield BC, Chou JY. G-CSF improves murineG6PC3-deficient neutrophil function by modulating apoptosis and energyhomeostasis. Blood 2011;117:3881–92.
[59] Kojima H, Otani A, Oishi A, Makiyama Y, Nakagawa S, Yoshimura N.Granulocyte colony-stimulating factor attenuates oxidative stress-inducedapoptosis in vascular endothelial cells and exhibits functional andmorphologic protective effect in oxygen-induced retinopathy. Blood2011;117:1091–100.
[60] Tsai RK, Chang CH, Sheu MM, Huang ZL. Anti-apoptotic effects of humangranulocyte colony-stimulating factor (G-CSF) on retinal ganglion cells afteroptic nerve crush are PI3K/AKT-dependent. Exp Eye Res 2011;90:537–45.
P.K. Singh et al. / Cytokine 56 (2011) 411–421 421




















