
Endogenous natriuretic factors. 7. Biospecificity of a natriuretic gamma-tocopherol metabolite...
7
0022-3565/97/2822-0657S03.00/0 THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICS Copyright © 1997 by The American Society for Pharmacology and Expérimental Thorapeutics JPET 282:657-662, 1997 Vol. 282, No. 2 Printed m U.S.A. Endogenous Natriuretic Factors 7: Biospecificity of a Natriuretic y-Tocopherol Metabolite LLU-a1 E. DAVID MURRAY, JR., WILLIAM J. WECHTER, DARKO KANTOCI, WEN-HUI WANG, TIM PHAM, DAVID D. QUIGGLE, KARINA M. GIBSON, DOUGLAS LEIPOLD and BEATRICE M. ANNER Laboratory of Chemical Endocrinology, Loma Linda University School of Medicine, Loma Linda, California (E.D.M. Jr., W.J.W., O.K., T.P., D.D.Q., K.M.G., D.L), Department of Pharmacology, New York Médical Collège, Valhalta, New York (W.-H.W.) and Department of Pharmacology, Geneva University School of Medicine, Geneva, Switzerland (B.M.A.) Accepted for publication April 8, 1997 ABSTRACT The structural elucidation and mechanism of action of a poten- tial component, LLU-a, of what is possibly a multifactorial complex known as "natriuretic hormone" was recently reported [Wechter, W.J. et al. (1996a) Proc. Natl. Acad. Sci. U.S.A. 93: 6002-6007]. "Natriuretic hormone," a long-sought factor, is believed to regulate extracellular fluid volume and consequently be pathomimetic for hypertension, cirrhosis, congestive heart failure and other volume expanded states. The studies reported herein further characterize LLU-a. The precursor of thé endog- enous LLU-a was demonstrated to be y-tocopherol by radio- labeling studies. The pharmacokinetics of infused rac-LLU-a proved to be biphasic (half-lives: 12 min and 6 h). Specificity of thé inhibition of thé 70 pS potassium channel of thé thick ascending limb of thé loop of Henle was examined with thé natural S-enantiomer being thé most potent known inhibitor whereas thé analogous a-tocopherol metabolite, rac-5-Me- LLU-ct, showed no inhibition. fîac-LLU-a does not inhibit two isozymesof theNa+/K+-ATPase. LLU-a is natriuretic acting via inhibition of thé 70 pS potassium channel and not Na+/K+- ATPase, thé assumed mechanism of action of thé "natriuretic hormone." LLU-a, a metabolite of a vitamin, if it were found to play a rôle in thé régulation of extracellular fluid volume, would be thé second example of a vitamin acting as a precursor for a hormone. Of considérable interest is thé fact that this manu- script reports thé first biological activity of y-tocopherol, a member of thé vitamin E complex. "Natriuretic hormone," thé putative controller of extracel- lular fluid volume (de Wardener et al., 1961) has been sought for more than 30 years. This compound was believed to be responsible for thé three effects observed during saline-in- duced volume expansion: prolongea natriuresis, inhibition of sodium transport and increased vasoreactivity. Most search paradigms hâve focused on thé inhibition of sodium trans- port, specifically inhibition of thé Na+/K+-ATPase (sodium pump) or immunocross-reactivity to antibodies developed to thé pump inhibitors ouabain or digoxin, as an assay to detect this compound during isolation (Wechter and Benaksas, 1990; Benaksas et al., 1995). This has led to thé isolation of digoxin (Goto et al., 1990) and "iso-ouabain" (Ludens et al., 1991; Mathews et al., 1991; Tymiak et al., 1993; Zhao et al., 1995). Digoxin and ouabain, however, are kaliuretic and not natriuretic (Crabos et al., 1987; Murray et al., 1995; Pamnani et al, 1991; Smyth et al, 1992; Sekihara and Yuzaki, 1993). Received for publication December 6, 1996. ! This work was supported by Adventist Health Systems/Loma Linda (E.D.M., D.K., W.J.W.) and thé National Institûtes of Health (W.-H.W; grants DK 47402 and HL 34300) and Swiss National Science Foundation (B.M.A.; grant 31-37-552.93). Therefore, this search tool is inappropriate except for bufa- lin-like compounds which are natriuretic (Pamnani et al., 1991). Our search has concentrated on thé natriuresis attrib- uted to this putative hormone, resulting in détection of sev- eral natriuretic components in human urémie urine (Benak- sas et al., 1993; Murray et al., 1995). The isolation, purification and structure détermination of thé most interesting of thèse compounds to date, LLU-a, has been described (Wechter et al., 1996a). A probable metabolite of y-tocopherol, LLU-a, was shown to be natriuretic, an in- hibitor of thé 70 pS ATP-sensitive K+ (KATP) channel in thé TAL and without an effect on hemodynamics as measured by thé glomerular filtration rate (Wechter et al., 1996a). It was also demonstrated not to be an inhibitor of thé Na +/K+- ATPase in MDBK cells. Unlike atrial natriuretic peptide, which is also not an inhibitor of thé sodium pump (Chiou and Vesely, 1995; Charlton and Baylis, 1990), LLU-a exhibits a prolonged natriuresis like that described for "natriuretic hor- mone" (de Wardener et al., 1961). In thé présent study LLU-a is further characterized. Its origin from y-tocopherol is established. The pharmacokinet- ics of intravenously infused LLU-a is reported and compared ABBREVIATIONS: TAL, thick ascending limb; MDBK, Madin-Darby bovine kidney; HPLC, high-performance liquid chromatography; 5-Me-LLU-«, 5-methyl-LLU-a; PBS, phosphate-buffered saline; Hepes, 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid. 657
Transcript of Endogenous natriuretic factors. 7. Biospecificity of a natriuretic gamma-tocopherol metabolite...

0022-3565/97/2822-0657S03.00/0THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICSCopyright © 1997 by The American Society for Pharmacology and Expérimental ThorapeuticsJPET 282:657-662, 1997
Vol. 282, No. 2Printed m U.S.A.
Endogenous Natriuretic Factors 7: Biospecificity of aNatriuretic y-Tocopherol Metabolite LLU-a1
E. DAVID MURRAY, JR., WILLIAM J. WECHTER, DARKO KANTOCI, WEN-HUI WANG, TIM PHAM, DAVID D. QUIGGLE,KARINA M. GIBSON, DOUGLAS LEIPOLD and BEATRICE M. ANNER
Laboratory of Chemical Endocrinology, Loma Linda University School of Medicine, Loma Linda, California (E.D.M. Jr., W.J.W., O.K., T.P.,D.D.Q., K.M.G., D.L), Department of Pharmacology, New York Médical Collège, Valhalta, New York (W.-H.W.) and Department ofPharmacology, Geneva University School of Medicine, Geneva, Switzerland (B.M.A.)
Accepted for publication April 8, 1997
ABSTRACTThe structural elucidation and mechanism of action of a poten-tial component, LLU-a, of what is possibly a multifactorialcomplex known as "natriuretic hormone" was recently reported[Wechter, W.J. et al. (1996a) Proc. Natl. Acad. Sci. U.S.A. 93:6002-6007]. "Natriuretic hormone," a long-sought factor, isbelieved to regulate extracellular fluid volume and consequentlybe pathomimetic for hypertension, cirrhosis, congestive heartfailure and other volume expanded states. The studies reportedherein further characterize LLU-a. The precursor of thé endog-enous LLU-a was demonstrated to be y-tocopherol by radio-labeling studies. The pharmacokinetics of infused rac-LLU-aproved to be biphasic (half-lives: 12 min and 6 h). Specificity ofthé inhibition of thé 70 pS potassium channel of thé thick
ascending limb of thé loop of Henle was examined with thénatural S-enantiomer being thé most potent known inhibitorwhereas thé analogous a-tocopherol metabolite, rac-5-Me-LLU-ct, showed no inhibition. fîac-LLU-a does not inhibit twoisozymesof theNa+/K+-ATPase. LLU-a is natriuretic acting viainhibition of thé 70 pS potassium channel and not Na+/K+-ATPase, thé assumed mechanism of action of thé "natriuretichormone." LLU-a, a metabolite of a vitamin, if it were found toplay a rôle in thé régulation of extracellular fluid volume, wouldbe thé second example of a vitamin acting as a precursor for ahormone. Of considérable interest is thé fact that this manu-script reports thé first biological activity of y-tocopherol, amember of thé vitamin E complex.
"Natriuretic hormone," thé putative controller of extracel-lular fluid volume (de Wardener et al., 1961) has been soughtfor more than 30 years. This compound was believed to beresponsible for thé three effects observed during saline-in-duced volume expansion: prolongea natriuresis, inhibition ofsodium transport and increased vasoreactivity. Most searchparadigms hâve focused on thé inhibition of sodium trans-port, specifically inhibition of thé Na+/K+-ATPase (sodiumpump) or immunocross-reactivity to antibodies developed tothé pump inhibitors ouabain or digoxin, as an assay to detectthis compound during isolation (Wechter and Benaksas,1990; Benaksas et al., 1995). This has led to thé isolation ofdigoxin (Goto et al., 1990) and "iso-ouabain" (Ludens et al.,1991; Mathews et al., 1991; Tymiak et al., 1993; Zhao et al.,1995). Digoxin and ouabain, however, are kaliuretic and notnatriuretic (Crabos et al., 1987; Murray et al., 1995; Pamnaniet al, 1991; Smyth et al, 1992; Sekihara and Yuzaki, 1993).
Received for publication December 6, 1996.! This work was supported by Adventist Health Systems/Loma Linda
(E.D.M., D.K., W.J.W.) and thé National Institûtes of Health (W.-H.W; grantsDK 47402 and HL 34300) and Swiss National Science Foundation (B.M.A.;grant 31-37-552.93).
Therefore, this search tool is inappropriate except for bufa-lin-like compounds which are natriuretic (Pamnani et al.,1991). Our search has concentrated on thé natriuresis attrib-uted to this putative hormone, resulting in détection of sev-eral natriuretic components in human urémie urine (Benak-sas et al., 1993; Murray et al., 1995).
The isolation, purification and structure détermination ofthé most interesting of thèse compounds to date, LLU-a, hasbeen described (Wechter et al., 1996a). A probable metaboliteof y-tocopherol, LLU-a, was shown to be natriuretic, an in-hibitor of thé 70 pS ATP-sensitive K+ (KATP) channel in théTAL and without an effect on hemodynamics as measured bythé glomerular filtration rate (Wechter et al., 1996a). It wasalso demonstrated not to be an inhibitor of thé Na+/K+-ATPase in MDBK cells. Unlike atrial natriuretic peptide,which is also not an inhibitor of thé sodium pump (Chiou andVesely, 1995; Charlton and Baylis, 1990), LLU-a exhibits aprolonged natriuresis like that described for "natriuretic hor-mone" (de Wardener et al., 1961).
In thé présent study LLU-a is further characterized. Itsorigin from y-tocopherol is established. The pharmacokinet-ics of intravenously infused LLU-a is reported and compared
ABBREVIATIONS: TAL, thick ascending limb; MDBK, Madin-Darby bovine kidney; HPLC, high-performance liquid chromatography; 5-Me-LLU-«,5-methyl-LLU-a; PBS, phosphate-buffered saline; Hepes, 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid.
657

658 Murray et al. Vol. 282
with thé time course of natriuresis. The specifïcity of théinhibition of thé 70 pS K+ channel by thé enantiomers ofLLU-Œ is determined. The analogous a-tocopherol metaboliteis also investigated for ils ability to inhibit this potassiumchannel. The inhibition of thé Na+/K+-ATPase from différentsources is tested.
MethodsLLU-tt and derivatives. The compounds used in thèse studies
(fig. 1 ) were synthesized as described by Kantoci et al. (accompanyingpaper, 1997). The enantiomers of LLU-a were purified by prepara-tive chiral HPLC as described by Kantoci et al. (accompanying paper,1997).
Metabolism of y-tocopherol in rats. Two rats (Sprague-Daw-ley, Harlan) were each injected intraperitoneally with 500 /.(.Ci/kg ofl/'HJ-y-tocopherol (9.6 Ci/mmol, Amersham custom synthesis) in 2.5ml/kg of 10% ethanol in corn oil (Chiku et al., 1984; Simon et al.,1956a, b). The rats were then placed into metabolic cages for urinecollection for 4 days. The urine (220 ml) was pooled, solids removedby filtration. The solution was extracted with 2 g Amberlite XAD-2(Aldrich, Milwaukee, WI) with stirrîng for 18 h. The XAD was recov-ered by filtration and extracted with 4 volumes of methanol totaling60 ml. The methanol was removed under a stream of 5.0 fini filteredair. The dried residue was resuspended in 1.8 ml of 40:60 methanol/0.2 M acetic acid for chromatography. Beckman System Gold (126pump, 168 diode array détecter) controlled by System Gold softwarev 5.1 on PC was used for ail chromatography. A C]8 RP-HPLC(Beckman Ultrasphere ODS column; 5 (i.m; 10 X 250 mml was elutedat 2 rnl/rnin with a gradient of 0.2 M acetic acid (A), methanol (B) and70% toluène/30% methanol (C) (60% A:40% B for 5 min, a lineargradient to 50% A:50% B over 5 min, a linear gradient to 30% A:70%B over 55 min, a linear gradient to 100% B over 2 min, 100% B for 3min, 100% C for 8 min, 100% B for 7 min). The eluant was monitored
for absorbance at 265 and 290 nm and ultraviolet spectra werecollected (202-390 nm}. Seventy-eight fractions of 1-min durationwere collected. An aliquot (50 fû) of each fraction was counted in 5 mlof liquid scintillation cocktail (ScintiVerse™ Bio-HP, Fisher, Tustin,ÇA) in a LS 3801 Liquid Scintillation Counter (Beckman, SanRamon, ÇA). Fractions (39-42) eluting in thé région known to con-tain LLU-o were pooled, 25 /j,g of synthetic, racemic LLU-« wasadded and thé pooled material was dried under reduced pressure ina centrifugal vacuum concentrator (Jouan, Winchester, VA). Theresidue was dissolved in 1.8 ml of 15:85 45 mM acetic acid in aceto-nitrile/50 mM acetic acid (aq.) and chromatographed on C1H RP-HPLC (Beckman Ultrasphere ODS column; 5 ^m; 4.6 x 250 mm)eluted at 1 ml/min with a gradient of 50 mM acetic acid (A) and 45mM acetic acid in acetonitrile (B) (85% A:15% B for 3 min, a lineargradient to 100% B over 42 min, 100% B for 5 min). The eluant wasmonitored for absorbance at 265 and 295 nm by thé diode arraydétecter. Fifty 1-min fractions were collected with aliquots (100 fd)counted as described abovc. This experiment was performed induplicate.
In vivo bioassay. The assay for natriuresis in conscious rats haspreviously been described in détail (Benaksas et al., 1993), but willbe briefly reiterated hère. The bladder of ether-anesthetized femaleSprague-Dawley (Harlan) rats (200-250 g) was catheterized for col-lection of urine in 10-min periods. The fémoral artery and vein werecannulated for monitoring of mean arterial pressure and infusion ofsaline (0.49 ml/h) and samples, respectively. After recovery fromanesthesia, furosemide (100 /^g, approximately 0.4 mg/kg b.wt; 1mg/ml in 0.17% saline) was infused as a positive control at thébeginning of thé sixth 10-min urine collection period. The samplewas infused at thé beginning of thé 17th 10-min period. Urine wascollected for another 150 min. The volume of thé urine was deter-mined gravimetrically. The urine Na~ and K"1" concentrations weredetermined with a Beckman E2A electrolyte analyzer. From thèse
H,C COOH
Synthetic, racemic LLU-cc
COOH
Synthetic, racemic 5-Me-LLU-ct
H,C COOH H3C COOHFig. 1. Compounds used in thèse stud-ies. S-LLU-a is thé naturel enantiomer(Kantoci et al., accompanying paper,1997).
Lactone of oxidation product of LLU-a

1997 Natriuretic y-Tocopherol Metabolite 659
data thé sodium excrétion values (UNaV; LUNaV = urine sodiumconcentration X urine volume per time]) were calculated.
The net sodium excrétion for thé infusion of furosemide or samplewas calculated as follows. The médian (or middlc) sodium excrétionvalue Immoles Na VIO-min period) for thé five periods before infusionof furosemide or sample was used to establish a baseline value for thécalculation of AUNaV (= ^moles Na+ in a 10-min period - base-linejumelés Na+) for thé administration of either furosemide or sample,respectively. The sum of AUNaV for thé four periods after infusion offurosemide was thé net sodium excreted for furosemide, defined asfurosemide response (FR). The sum of AUNaV for thé 15 periods (150rnin) after infusion of thé sample was thé net sodium excreted for thésample, defined as sample response (SR). The natriuretic response ofa sample was then normalized to thé natriuretic response of thé doseof furosemide infused. This natriuretic ratio (RN) (or normalizednatriuretic response) of a sample was calculated by dividing SR byFR (RN = SR/FR). A sample is considered natriuretically active if RN
^ .55 (greater than 99% confidence limite).Pharmacokinetics of LLU-cc. The in vivo bioassay for natriure-
sis (described above) was adapted for thèse experiments. At thé timeof sample infusion, 10 /ng of [!'H|LLU-« (4.2 ^.Ci/nmol, AmershamCustom Synthesis, 0.25 mCi/ml ethanol; 640 jil dried under a streamof argon and resuspended in 100 /.il of 47.5% ethanol) was infused,and thé saline infusion rate was changed to 1 ml/min. The experi-ment was performed on two animais. Blood (approximately 0.5 ml)was drawn from each animal into heparinized microfuge tubes atvarious time points. Blood was drawn from animal 1 at 1, 5, 30 and120 rnin and from animal 2 at 2, 15, 60 and 240 min. The blood wascentrifuged for 5 min. An aliquot of plasma (100 jLtl) was withdrawnand placed into a 15-ml centrifuge tube. HC1 (2 N, 20 /J) was addedto each centrifuge tube and vortexed for 30 sec. Hexane/ether (4:1, 2ml) was added to each tube and thé tube was vortexed for 3 min. Thesamples were sonicated and centrifuged as necessary and 1.5 ml ofthé hexane/ether sample was removed and placed in a glass testtube. Unlabeled synthetic, racemic LLU-a (50 /i.g) was added to eachsample. The solvent was removed under a stream of filtered air. Thesample was resuspended in 1.8 ml of 15:85 45 mM acetic acid inmethanoL/50 mM acetic acid (aq.) and chromatographed on aceticacid/acetonitrile RP-HPLC as described in "Metabolism of y-Tocoph-erol in Rats." Fifty 1-min fractions were collected, with aliquots (100(Ltl) counted as above. A pilot study was conducted using one animalinfused with 10 jig of [3HlLLU-« and blood drawn at 5,15, 30, 60 and160 min.
Na+/K+-ATPase inhibition assays. Inhibition of thé Na+/K+-ATPase by thé réduction of 86Rb+ uptake into human lymphocyteswas examined. Lymphocytes were isolated from heparinized bloodwith a Histopaque-1077 (Sigma, Buchs, Switzerland) density gradi-ent and then washed with buffer by centrifugation similar to previ-ously described methods (Armer et al., 1994). Cell viability wasexamined by thé trypan bluc dye exclusion test and was found to begreater than 99%. Lymphocytes (5 X 106 cells) were incubated in théprésence or absence of synthetic, racemic LLU-o: (125 fjM, finalconcentration) in 50 /A of PBS (154 mM NaCl, 10 mM sodiumphosphate, pH 7.2-7.4) with 50 nCi 86Rb+ for 30 min at 36°C withshaking. Cells were washed three times with PBS (once) or D-PBS(twice; 137 mM NaCl, 2.68 mM KC1, 0.90 mM CaCl2, 0.49 mMMgCla, 6.46 mM Na2HPO4, 1.47 mM KH^PO,,) by centrifugation at0°C at 500 X g The recovered cells were then counted. Cell viabilitywas checkeci by trypan blue dye exclusion after incubation under thésame conditions but without 8t'Rb(. Na h/K+-ATPase was isolatedfrom lamb rénal outer medulla as described previously (Anner et al.,1992). Na+/K * -ÂTPase activity was measured by thé linked enzymeassay method described (Anner et al., 1992). Synthetic, racemicLLU-a in 50 mM sodium phosphate buffer, pH 7.2, was added to afinal concentration of 125 [tM either during thé assay or 30 rnin.before thé assay for preincubation at 37°C. Only buffer was added forcontrol assays.
Patch-clamp experiments. Pathogen-free Sprague-Dawley rats(rnale or female, 80-120 g) were maintained on normal rat chowbefore being sacrificed as described previously (Wang, 1994). Kid-neys were then removed and thin (1 mm) coronal sections were eut.The cortical and medullary TAL tubules were dissected at roomtempérature. The TAL tubules were immobilized onto cover glasscoated with Cell-Tak (Collaborative Research, Bedford, MA). Thecover glass was then placed in a chamber mounted on an invertedmicroscope and thé tubules superfused at 37°C with Hepes-bufferedRinger's solution (135 mM NaCl, 5 mM KC1, 1.5 mM MgCl2, 1.8 mMCaCl2, 5 mM glucose, 10 mM Hepes, pH 7.4). The apical membranewas exposed by cutting thé tubule with a sharpened micropipette.Single-channel currents were recorded in cell-attached, inside-outconfiguration with 140 mM KC1 in thé pipette with a résistance ofapproximately 4 to 6 megohm. The recordings were collected with anAxon 200A patch-clamp amplifier. Single-channel currents were low-pass filtered at 1 kHz with an eight-pole Bessel filter (FrequencyDevices 902LPF) and converted to digitized signais at a samplingrate of 44 kHz by a Sony PCM-501ES puise code modulator and avideotape recorder. Analysis was done with use of thé pCLAMPsoftware (v. 6.0, Axon Instruments, Burlingame, ÇA) on an IBM-compatible 486 computer. The potency of thé inhibition of thé K 'channel is expressed as IC50, which was calculated by linear régres-sion analysis.
Results
70 pS K+ channel inhibition. Racemic, synthetic LLU-ahas been previously shown to inhibit thé 70 pS K+ channel inthé TAL of thé loop of Henle with an IC50 of 6 nM (Wechteret al., 1996a and fig. 2). R- and S-LLU-a as well as racemic,synthetic 5-methyl-LLU-a (thé analogous metabolite of a-to-copherol) were tested in this system. 5-Methyl-LLU-a dem-onstrated no inhibition of thé 70 pS K ' channel in thé con-centration range of 100 to 500 nM (fig. 2), whereas S-LLU-t*(IC50 = 3 nM) was a more potent inhibitor than thé racemateand R-IAÀJ-a was about 20-fold less effective, IC50 = 51 nM(fig. 2).
Isolation of [3H]LLU-a from [3H]y-tocopherol-treat-ed rats. To test thé hypothesis that LLU-a originales as aresuit of side-chain dégradation of y-tocopherol, rats were
100
80
g 60
40
20
I'
10 100Concentration (nM)
1000
Fig. 2. Inhibition of thé 70 pS K+ channels from TAL S-enantiomer ofrac-LLU-a {closed squares), R-enantiomer of rac-LLU-u (closed trian-gles), rac-LLU-u (closed circles) and rac-5-methyl-LLU-a (open circles)were examined for their ability to inhibit thé potassium channel asdescribed under "Methods." Data are présentée! as mean ± S.E.M.
660 Murray et al.
« Q innn
E1-5cin
**mg 1-0o
m•fi0
•.Q< 0.5
0.0
x
>
XX 1
//
/»
['0 10 20
i/
//
X/
X/•
,
.
.
h. [rmr^-r- , , 1
2500
co
2000 po
ii
1500 ~EQ.o -
1000 £OH
-
500
-
100
80
0>
60 'C«cOo
40 <
S?
20
0
30 40 50
Time (Minutes)
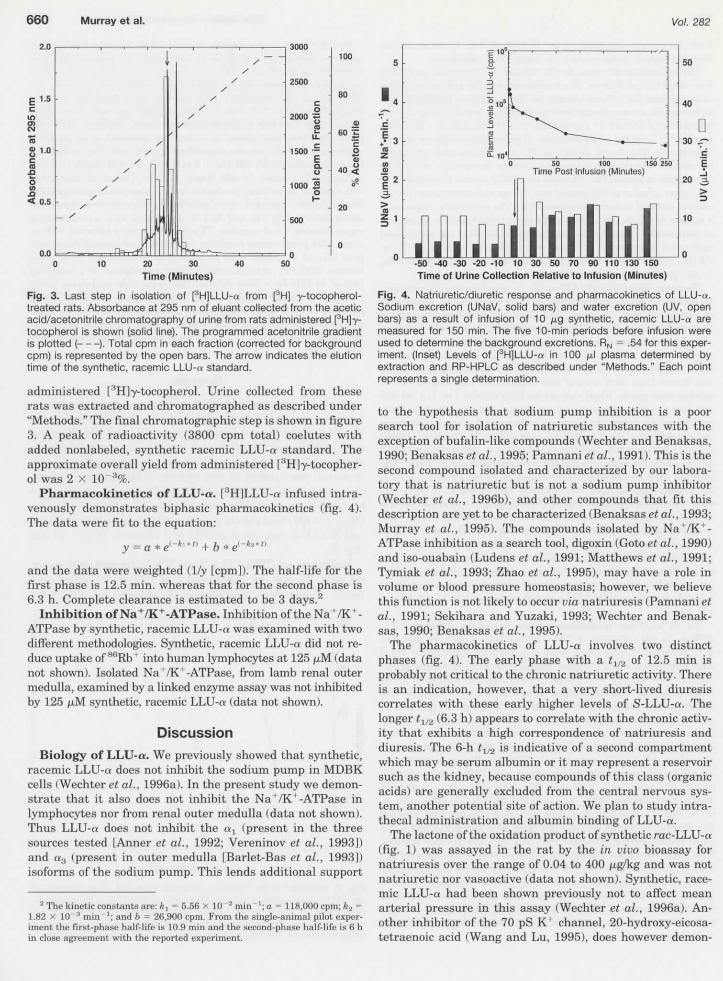
Fig. 3. Last step in isolation of fHJLLU-a from [3H] y-tocopherol-treated rats. Absorbance at 295 nrn of eluant collected from thé aceticacid/acetonitrile chromatography of urine from rats administered [3H]y-tocopherol is shown (solid line). The programmed acetonitrile gradientis plotted ( }. Total cpm in each fraction (corrected for backgroundcpm) is represented by thé open bars. The arrow indicates thé elutiontime of thé synthetic, racemic LLU-a standard.
administered [3H]y-tocopherol. Urine collected from thèserats was extracted and chromatographed as described under"Methods." The final chromatographic step is shown in figure3. A peak of radioactivity (3800 cpm total) coelutes withadded nonlabeled, synthetic racemic LLU-a standard. Theapproximate overall yield from administered [''H^-tocopher-ol was 2 X 10 3%.
Pharmacokinetics of LLU-«. [JH]LLU-a infused intra-venously demonstrates biphasic pharmacokinetics (fïg. 4).The data were fit to thé équation:
y — a * e
and thé data were weighted (1/y [cpm]). The half-life for théfirst phase is 12.5 min. whereas that for thé second phase is6.3 h. Complète clearance is estimated to be 3 days.2
Inhibition of Na+/K+-ATPase. Inhibition of thé Na*/K4 -ATPase by synthetic, racemic LLU-a was examined with twodifférent méthodologies. Synthetic, racemic LLU-a did not re-duce uptake of 8GRb ' into human lymphocytes at 125 /xM (datanot shown). Isolated Na+/K+-ATPase, from lamb rénal outermedulla, examined by a linked enzyme assay was not inhibitedby 125 /xM synthetic, racemic LLU-a (data not shown).
Discussion
Biology of LLU-a. We previously showed that synthetic,racemic LLU-a does not inhibit thé sodium pump in MDBKcells (Wechter et al., 1996a). In thé présent study we démon-strate that it also does not inhibit thé Na h/Kf-ATPase inlymphocytes nor from rénal outer medulla (data not shown).Thus LLU-a does not inhibit thé a^ (présent in thé threesources tested [Anner et al., 1992; Vereninov et al., 1993])and a,j (présent in outer medulla IBarlet-Bas et al., 19931)isoforms of thé sodium pump. This lends additional support
|2
1
Vol. 282
50
40
50 100 150 250Time Posl Infusion (Minutes)
20
10
2 The kinetic constants are: k1 = 5.56 x 10 2min ';a = 118,000 iîpm;A2 =1.82 X 10~:1 min"1; and b = 26,900 cpm. From thé single-animal pilot exper-iment thé first-phase half-life is 10.9 min and thé second-phase half-life is 6 hin close agreement with thé reported experiment.
-50 -40 -30 -20 -10 10 30 50 70 90 110 130 150
Time of Urine Collection Relative to Infusion (Minutes)
Fig. 4. Natriuretic/diuretic response and pharmacokinetics of LLU-a.Sodium excrétion (UNaV, solid bars) and water excrétion (UV, openbars) as a resuit of infusion of 10 fïg synthetic, racemic LLU-tt aremeasured for 150 min. The five 10-min periods before infusion wereused to détermine thé background excrétions. RN = .54 for this exper-irnent. (Inset) Levels of [3H]LLU-u in 100 /il plasma determined byextraction and RP-HPLC as described under "Methods." Each pointrepresents a single détermination.
to thé hypothesis that sodium pump inhibition is a poorsearch tool for isolation of natriuretic substances with théexception of bufalin-like compounds (Wechter and Benaksas,1990; Benaksas et al., 1995; Pamnani et al., 1991). This is thésecond compound isolated and characterized by our labora-tory that is natriuretic but is not a sodium pump inhibitor(Wechter et al., 1996b), and other compounds that fit thisdescription are yet to be characterized (Benaksas et al., 1993;Murray et al., 1995). The compounds isolated by Na^/K"1"-ATPase inhibition as a search tool, digoxin (Goto et al., 1990)and iso-ouabain (Ludens et al., 1991; Matthews et al., 1991;Tymiak et al., 1993; Zhao et al., 1995), may hâve a rôle involume or blood pressure homeostasis; however, we believethis function is not likely to occur via natriuresis (Pamnani etal., 1991; Sekihara and Yuzaki, 1993; Wechter and Benak-sas, 1990; Benaksas et al., 1995).
The pharmacokinetics of LLU-a involves two distinctphases (fïg. 4). The early phase with a t1/2 of 12.5 min isprobably not critical to thé chronic natriuretic activity. Thereis an indication, however, that a very short-lived diuresiscorrelates with thèse early higher levels of S-LLU-a. Thelonger tl/2 (6.3 h) appears to correlate with thé chronic activ-ity that exhibits a high correspondence of natriuresis anddiuresis. The 6-h ti/2 is indicative of a second compartmentwhich may be sérum albumin or it may represent a réservoirsuch as thé kidney, because compounds of this class (organicacids) are generally excluded from thé central nervous sys-tem, another potential site of action. We plan to study intra-thecal administration and albumin binding of LLU-a.
The lactone of thé oxidation product of synthetic rac-LLU-a(fïg. 1) was assayed in thé rat by thé in vivo bioassay fornatriuresis over thé range of 0.04 to 400 /-(.g/kg and was notnatriuretic nor vasoactive (data not shown). Synthetic, race-mic LLU-a had been shown previously not to affect meanarterial pressure in this assay (Wechter et al., 1996a). An-other inhibitor of thé 70 pS K+ channel, 20-hydroxy-eicosa-tetraenoic acid (Wang and Lu, 1995), does however démon-

1997 Natriuretic y-Tocopherol Metabolite 659
data thé sodium excrétion values (UNaV; |UNaV = urine sodiumconcentration X urine volume per time]) were calculated.
The net sodium excrétion for thé infusion of furosemide or samplewas calculated as follows. The médian (or middle) sodium excrétionvalue (jamoles Na+/10-rnin period) for thé five periods before infusionof furosemide or sample was used to establish a baseline value for thécalculation of AUNaV (= jtmoles Na+ in a 10-min period - base-line/iinoles Na+) for thé administration of either furosemide or sample,respectively. The sum of AUNaV for thé four periods after infusion offurosemide was thé net sodium excreted for furosemide, defined asfurosemide response (FR). The sum of AUNaV for thé 15 periods (150min) after infusion of thé sample was thé net sodium excreted for thésample, defined as sample response (SR). The natriuretic response ofa sample was then normalized to thé natriuretic response of thé doseof furosemide infused. This natriuretic ratio (RN) (or normalizednatriuretic response) of a sample was calculated by dividing SR byFR (RN = SR/FR). A sample is considered natriuretically active if RN
^ .55 (greater than 99% confidence limits).Pharmacokinetics of LLU-«. The in vivo bioassay for natriure-
sis (described above) was adapted for thèse experiments. At thé timeof sample infusion, 10 jiig of |''H|LLU-« (4.2 /iCi/nmol, AmershamCustom Synthesis, 0.25 mCi/ml ethanol; 640 (û dried under a streamof argon and resuspended in 100 /xl of 47.5% ethanol) was infused,and thé saline infusion rate was changed to 1 ml/min. The experi-ment was performed on two animais. Blood (approximately 0.5 ml)was drawn from each animal into heparinized microfuge tubes atvarious time points. Blood was drawn from animal 1 at 1, 5, 30 and120 min and from animal 2 at 2, 15, 60 and 240 min. The blood wascentrifugea for 5 min. An aliquot of plasma (100 /il) was withdrawnand placed into a 15-ml centrifuge tube. HC1 (2 N, 20 /xl) was addedto each centrifuge tube and vortexed for 30 sec. Hexane/ether (4:1, 2ml) was added to each tube and thé tube was vortexed for 3 min. Thesamples were sonicated and centrifuged as necessary and 1.5 ml ofthé hexane/ether sample was removed and placed in a glass testtube. Unlabeled synthetic, racemic LLU-n (50 /_tg) was added to eachsample. The solvent was removed under a stream of filtered air. Thesample was resuspended in 1.8 ml of 15:85 45 mM acetic acid inmethanol/50 mM acetic acid (aq.) and chromatographed on aceticacid/acetonitrile RP-HPLC as described in "Metabolism of -y-Tocoph-erol in Rats." Fifty 1-min fractions were collected, with aliquots (100^1) counted as above. A pilot study was conducted using one animalinfused with 10 fig of [3H]LLU-a and blood drawn at 5,15, 30, 60 and160 min.
Na+/K+-ATPase inhibition assays. Inhibition of thé Na'/K4"-ATPase by thé réduction of 86Rb+ uptake into human lymphocyteswas examined. Lymphocytes were isolated from heparinized bloodwith a Histopaque-1077 (Sigma, Buchs, Switzerland) density gradi-ent and then washed with buffer by centrifugation similar to previ-ously described methods (Anner et al., 1994). Cell viability wasexamined by thé trypan blue dye exclusion test and was found to begreater than 99%. Lymphocytes (5 X 106 cells) were incubated in théprésence or absence of synthetic, racemic LLU-a (125 /i,M, finalconcentration) in 50 jtl of PBS (154 mM NaCl, 10 mM sodiumphosphate, pH 7.2-7.4) with 50 nCi 86Rb+ for 30 min at 36°C withshaking. Cells were washed three times with PBS (once) or D-PBS(twice; 137 mM NaCl, 2.68 mM KC1, 0.90 mM CaCl2, 0.49 mMMgCla, 6.46 mM Na2HPO4, 1.47 mM KH2PO4) by centrifugation at0°C at 500 X g The recovered cells were then counted. Cell viabilitywas checked by trypan blue dye exclusion after incubation under thésame conditions but without 86Rb h . Na'/K+-ATPase was isolatedfrom lamb rénal outer medulla as described previously (Anner et al.,1992). Na^/K^ATPase activity was measured by thé linked enzymeassay method described (Anner et al., 1992). Synthetic, racemicLLU-o in 50 mM sodium phosphate buffer, pH 7.2, was added to afinal concentration of 125 fiM either during thé assay or 30 min.before thé assay for preincubation at 37°C. Only buffer was added forcontrol assays.
Patch-clamp experiments. Pathogen-free Sprague-Dawley rats(mâle or female, 80-120 g) were maintained on normal rat chowbefore being sacrificed as described previously (Wang, 1994). Kid-neys were then removed and thin (1 mm) coronal sections were eut.The cortical and medullary TAL tubules were dissected at roomtempérature. The TAL tubules were immobilized onto cover glasscoated with Cell-Tak (Collaborative Research, Bedford, MA). Thecover glass was then placed in a chamber mounted on an invertedmicroscope and thé tubules superfused at 37°C with Hepes-bufferedRinger's solution (135 mM NaCl, 5 mM KC1,1.5 mM MgCl2, 1.8 mMCaCl2, 5 mM glucose, 10 mM Hepes, pH 7.4). The apical membranewas exposed by cutting thé tubule with a sharpened micropipette.Single-channel currents were recorded in cell-attached, inside-outconfiguration with 140 mM KC1 in thé pipette with a résistance ofapproximately 4 to 6 megohm. The recordings were collected with anAxon 200A patch-clamp amplifier. Single-channel currents were low-pass filtered at 1 kHz with an eight-pole Bessel filter {FrequencyDevices 902LPF) and converted to digitized signais at a samplingrate of 44 kHz by a Sony PCM-501ES puise code modulator and avideotape recorder. Analysis was done with use of thé pCLAMPsoftware (v. 6.0, Axon Instruments, Burlingame, ÇA) on an IBM-compatible 486 computer. The potency of thé inhibition of thé K 'channel is expressed as ICril), which was calculated by linear régres-sion analysis.
Results
70 pS K+ channel inhibition. Racemic, synthetic LLU-ahas been previously shown to inhibit thé 70 pS K+ channel inthé TAL of thé loop of Henle with an IC50 of 6 nM (Wechteret al., 1996a and fig. 2). R- and S-LLU-a as well as racemic,synthetic 5-methyl-LLU-a (thé analogous metabolite of a-to-copherol) were tested in this system. 5-Methyl-LLU-a dem-onstrated no inhibition of thé 70 pS K ' channel in thé con-centration range of 100 to 500 nM (fig. 2), whereas S-LLU-a(ICSO — 3 nM) was a more potent inhibitor than thé racemateand R-LLU-u was about 20-fold less effective, IC50 = 51 nM(fig. 2).
Isolation of [3H]LLU-a from [3H]y-tocopherol-treat-ed rats. To test thé hypothesis that LLU-a originales as aresuit of side-chain dégradation of y-tocopherol, rats were
100
80
60
40
20
0
10 100Concentration (nM)
1000
Fig. 2. Inhibition of thé 70 pS K+ channels from TAL. S-enantiomer ofrac-LLU-a (closed squares), fî-enantiomer of rac-LLU-u {closed trian-gles), rac-LLU-tï (closed circles) and rac-5-methyl-LLU-« (open circles)were examined for their ability to inhibit thé potassium channel asdescribed under "Methods." Data are présentée! as mean ± S.E.M.

1997 Natriuretic y-Tocopherol Metabolîte 661
strate vasoactivity in thé spontaneously hypertensive rat(Schwartzman and McGiff, 1995). The vasoactivity of LLU-tttherefore needs to be examined in other Systems. Once syn-thesized, thé hydroquinone of thé lactone of thé oxidationproduct of LLU-a will be examined for natriuresis and vaso-activity.
In previous studies, thé effect of synthetic, racemic LLU-aon thé 70 pS KATF channel in thé TAL cells was investigatedas a potential mechanism of natriuresis (Wechter et al.,1996a). Inhibition of this channel in thé TAL leads to natri-uresis by preventing potassium recycling for thé Na+/K+/2C1 cotransporter (Greger, 1985; Hébert and Andreoli,1984; Greger and Schlatter, 1981). In thé current study théindividual enantiomers of LLU-tt were examined in patch-clamp expérimente. The S-enantiomer of LLU-a (thé natu-rally occurring enantiomer; Kantoci et al., accompanying pa-per, 1997) proved to be an even more potent inhibitor of thé70 pS K' channel than thé racemate (fig. 2). The R-enanti-omer, however, although not devoid of activity was about20-fold less potent (fig. 2) than thé S-enantiomer. Signifï-cantly, thé analogous compound derived from a-tocopherol(5-Me-LLU-a; Schônfeld et al., 1993; Schultz et al., 1995) wascompletely inactive as a potassium channel inhibitor at théconcentrations tested (100-500 nM; fig. 2). Thèse resultsindicate gréât structural specificity to thé biology of LLU-aand its binding to thé 70 pS K+ channel. The addition of asingle methyl group being biologically significant is not un-usual (e.g., methylated steroids).
The interesting observation is that a metabolite of a minorcomponent of thé Vitamin E complex, obtained only throughdiet, is an effector of thé 70 pS K+ channel. In récent studiesin which 5-Me-LLU-a was isolated frorn human urine, théurine donors were given relatively large doses of a-tocopherol(650 mg/day and greater; Schultz et al., 1995; Schônfeld et al.,1993). 5-Me-LLU-a was not detected in any of thé pools ofurine from which LLU-a was isolated. It may be that 5-Me-LLU-a was présent as a conjugate. In those studies in which5-Me-LLU-a was isolated (Schultz et al, 1995; Schônfeld etal., 1993), thé investigators questioned thé rôle played byVitamin E (a-tocopherol) as an antioxidant, because thé ma-jor isolated metabolite was not thé oxidized Simon's metab-olites (Simon et al., 1956 a, b). The antioxidant potential of ana-tocopherol, however, is greater than that of y-tocopherol(reviewed by Kamal-Eldin and Appelqvist, 1996), which thusincreases thé likelihood of thé chroman ring of y-tocopherolremaining intact. At least for y-tocopherol thé proposed side-chain oxidation without thé chroman ring oxidation meta-bolic pathway may be necessary for thé production of animportant regulator of thé 70 pS KATP channel and possiblyin thé régulation of extracellular fluid volume.
Production of LLU-a from y-tocopherol. When LLU-awas first identifïed it was assumed that it originated frommetabolism of y-tocopherol, most likely by side-chain oxida-tion. In this study it is demonstrated that [ H3-LLU-a can beobtained from urine of rats administered [''H]y-tocopherol,thus providing évidence that LLU-a is a metabolite of y-to-copherol. This is in agreement with previous studies of a-to-copherol in humans and S-tocopherol in rats (Chiku et al.,1984; Schônfeld et al, 1993; Schultz et al., 1995). That LLU-aoriginales from side-chain dégradation without oxidation ofthé chroman ring is confirmed by thé study of Kantoci et al.(accompanying paper, 1997) demonstrating that thé stereo-
chemistry at C-2 is S or thé same configuration as that of théparent y-tocopherol (Burton and Ingold, 1989).
We currently hâve a study underway to détermine théconcentration of S-LLU-a in thé urine of pregnant women,congestive heart failure, cirrhosis, urémie and head traumapatients to détermine thé relationship between S-LLU-a lev-els and volume-expanded states. If there is a corrélationprésent, then thé multifactorial control of extracellular vol-ume including y-tocopherol might prove to be analogous tothat of vitamin D being a precursor to a hormone. The pos-sibility of y-tocopherol acting as a precursor to a hormonewould imply thé need for régulation of production of S-LLU-uvia storage, metabolism and other processes. This will un-doubtedly prove a very interesting area for future studies.The discovery of LLU-a is also thé first time that a biologicalactivity has been reported for y-tocopherol.
Ackno wledgm ent s
We acknowledge thé excellent technical assistance of M. Moos-mayer (Geneva) in performing thé Na+/K+-ATPase measurements.
Références
ANNER, B. M., MOIJSMAYER, M. AND IMESCH, E.: Mercury blocks Na-K-ATPase bya ligand-dependent and réversible mechanism. Am. J. Physiol. 262: F830-F836, 1992.
ANNER, B. M., LACOTTE, D., ANNËR, R. M. AND MOOSMAYER, M.: Interaction ofhypothalamic Na, K-ATPase inhibitor with isolated human peripheral bloodmononuclear cells. Biosci. Rep. 14: 231-242, 1994.
BARLET-BAS, C., ARYSTARKHOVA, E., CHEVAL, L., MARSY, S., SWEADNER, K., Moov-ANOV, N. AND DOUCET, A.: Are there several isoforrns of Na, K-ATPases alphasubunit in thé rabbit kîdney? J. Biol. Chem. 268: 11512-11515, 1993.
BENAKSAS, E. J., MURRAY, E. D., JK., RODGEKS, C. L., PHAM, T., BIGORNIA, A. E.,DEWIND, S. A., GIEBEL, R., BRUBACHER, E. S. AND WECHTER, W. J.: Endogenouanatriuretic factors 1: Sodium pump inhibition does not correlate with natri-uretic or pressor activities from urémie urine. Life Sci. 52:1045-1054, 1993.
BENAKSAS, E. J., MURRAY, E. D., JR. AND WECHTER, W. J,: Natriuretic hormonesII. Prog. Drug Res. 45: 245-288, 1995.
BURTON, G. W. AND INGULU, K. U.: Vitamin E as an in vitro and in vivoantioxidant. Ann. NY Acad. Sci. 570: 7-22, 1989.
CHARLTON, J. A. AND BAYLIS, P. H.: Lack of inhibition of vasopressin-stimulatedNa ' K ' ATPase by atrial natriuretic factor in thé rat rénal medullary thickascending limb of Henle's loop. Cell. Biochem. Funct. 8: 25-29, 1990.
CHIKU, S., HAMAMURA, K. AND NAKAMUKA, T.: Novel urinary metabolite of d-S-tocopherol in rats. J. Lipid Res. 25: 40-48, 1984.
CHIOU, S. ANDVESELY, D. L.: Dimethyl sullbxide inhibits rena! Na f-K"-ATPaseat a site différent from ouabain and atrial peptides. Life Sci. 57: 945-955,1995.
CRABOS, M., GRICHOIS, M. L., GUICHENEY, P., WAINER, I. W. AND CLOIX, J.: Furtherbiochemical characterization of an Na ' -pump inhibitor purified from humanurine. Eur. J. Biochem. 162: 129-135, 1987.
DE WARDENER, H. E., MILLS, I. H., CLAPHAM, W. F. AND HAYTER, C. J.: Studies onthé efferent mechanism of thé sodium diuresis which follows thé adminis-tration of intr ave nous saline in thé dog. Clin. Sci. 21: 249-258, 1961.
GOTO, A., ISHIGURO, T., YAMAHA, K., ISHII, M., YOSHIOKA, M., EUGCHI, C., SHIMORA,M. AND SUGIMOTO, T.: Isolation of a urinary digitalis-like factor indistinguish-able from digoxin. Biochem. Biophys. Res. Commun. 173: 1093-1101, 1990.
GREttER, R. AND SCHLATTER, E.: Présence ofluminal K ' , a prerequisite for activeNaCl transport in thé cortical thick ascending limb of Henle's loop of rabbitkidney. Pflugers Archiv. 392: 92-94, 1981.
GREGER, R.: Ion transport mechanisms in thick ascending limb of Henle's loopof mammalian nephron. Physiol. Rev. 65: 760-797, 1985.
HEBERT, S. C. AND ANDREOLI, T. E.: Control of NaCl transport in thick ascendinglimb. Am. J. Physiol. 246: F745-F756, 1984.
KAMAL-ELDIN, A. AND AFPELQVIST, L.-Â.: The chemistry and antioxidant proper-ties of tocopherols and tocotrienols. Lipids 31: 671-701, 1996.
KANTOCI, D., MUHRAY, E. D., JR., DEWlND, S. A., BORCIIARDT, D., KHAN, S. ANDWECHTER, W. J.: Endogenous natriuretic factors 6: The stereochemistry of anatriuretic y-tocopherol metabolite LLU-a. J. Pharmacol. Exp. Ther. 282:000-000, 1997
LUDENS, J. H., CLARK, M. A., DUCHARME, D. W., HARRIS, D. W., LUTZKE, B. S.,MANUEL, F., MATTHEWS, W. R., SUTLER, D. M. AND HAMLYN, J. M.: Purificationof an endogenous digitalis-like factor from human plasma for structuralanalysis. Hypertension 17: 923-929, 1991.
MATHEWS, W. R., DUCHARME, D. W., HAMLYN, J. M., HARRIS, D. W., MANDEL, F.,CLARK, M. A. AND LUDENS, J. H.: Mass spectral characterization of an endog-enous digitalislike factor from human plasma. Hypertension 17: 930-935,1991.

662 Murray et al. Vol. 282
MURRAY, E. D., JR., KAMTOCI, D., D^WiND, S. A., BICORNIA, A. E., D'Aïuicu, D. C.,KINC, J. G., JR., PHAM, T., LEVINE, B. H., JUNG, M. E. AND WECHTER, W. J.:Endogenous natriuretic factors 3: Isolation and characterization of humannatriuretic factors LLU-a, LLU-fr, and LLU-y. Life Sci. 57: 2145-2161,1995.
PAMNANI, M. B., CHGN, S., BRYANT, H. J., SCHOOLEY, J. F., JR., ELIADES, D. C.,YUAN, C. M. AND HADDY, F. J.: Effects of three sodium-potassium adenosinetriphosphatase inhibitors. Hypertension 18: 316-324, 1991.
SCHONFELD, A., SCHULTZ, M., PETKizKA, M. AND GASSMAN, B.: A novel metaboliteof flfîR-a-tocopherol in human urine. Die Nahrung 37: 498-500, 1993.
SCHULTZ, M., LEIST, M., PETRZIKA, M., GASSMANN, B. AND B RE ic EL rus-FLORE, R.:Novel urinary metabolite of a-tocopherol, 2,5,7,8-tetramethyl-2 (2'-carboxyethyl)-6-hydroxychroman, as an indicator of an adéquate vitamin Esupply? Am. J. Clin. Nutr. 62: suppl., 1527S-1534S, 1995.
SCHWARTZMAK, M. L. AND McGiFK, J. C.: Rénal cytochrome P450. J. Lipid Med.Cell Sig. 12: 229-242, 1995.
SEKIHARA, H. AND YUZAKI, Y.: Ouabain causes kaliuresis and works synergisti-cally with aldosterone in vivo. Life Sci. 53: 975-980, 1993.
SIMON, E. J., GROSS, C. S. AND MILHORAT, A. T.: The metabolism of vitaminE. I. J. Biol. Chem. 221: 707-805, 1956a.
SIMON, E. J., EISENGART, A., SUNDHEIM, L. AND MILHORAT, A. T.: The metabolismof vitamin E. II. Purification and characterization of urinary metabolites ofa-tocopherol. J. Biol. Chem. 221: 807-817, 1956b.
SMYTH, D. D., TKMPLETON, J. F., SASHI KUMAR, V. P., YAN, Y., WIDAJEWICZ, W. ANDLABELLA, F. S.: Digitaloid pregnanes promote potassium-sparing diuresis inthé guinea pig. Can. J. Physiol. Pharmacol. 70: 723-727, 1992.
TYMIAK, A. A., NORMAN, J. A., BOLT.AR, M., DiDoNATO, G. C., LEE, H., PARKER,W. L., Lo, L.-C-, BEROVA, N., NAKANISHI, K., HABER, E. AND HAUPERT, G. T., JR.:
Physiochemical characterization of a ouabain isomer isolated from bovinehypothalamus. Proc. Natl. Acad. Sci. U.S.A. 90: 8189-8193, 1993.
VERENINOV, A. A., MARAKHOVA, I. I., OSIPOV, V. V. AND TOROPOVA, F. V.: Blasttransformation of lymphocytes is accompanied by and increase in expressionof mRNA, coding for Na, K-ATPase. FEBS Lett. 316: 37-^0, 1993.
WANG, W. H.: Two types of K+ channel in thick ascending limb of rat kidney.Am. J. Physiol. 267: F599-F605, 1994.
WANG, W. H. AND Lu, M.: Effect of arachidonic acidon activity of thé apical K h
channel in thé thick ascending limb of thé rat kidney. J. Gen. Physiol. 106:727-743, 1995.
WECHTER, W. J. AND BENAKSAS, E. J.: Natriuretic hormones. Prog. Drug Res. 34:231-260, 1990.
WECHTER, W. J., KANTOCI, D., MURRAY, E. D. JR., D'AMICO, D. C., JUNG, M. E. ANDWANG, W.-H.: A new endogenous natriuretic factor: LLU-a. Proc. Natl. Acad.Sci. U.S.A. 93: 6002-6007, 1996a.
WECHTER, W. J., MURRAY, E. D. JR., KANTOCI, D. AND JUNG, M. E.: Character-ization oftwo new endogenous natriuretic factors. J. Am. Soc. Nephrol. 7:1819, 1996b.
ZHAO, N., Lo., L.-C., BEROVA, N., NAKANISHI, K., TYMIAK, A. A., LUDENS, J. H. ANDHAUPERT, G. T., JR.: Na, K-ATPase inhibitors from bovine hypothalamus andhuman plasma are différent from ouabain: Nanogram scalde CD structuralanalysis. Biochemistry 34: 9893-9896, 1995.
Send reprint requests to: William J. Wechter, Ph.D., FCP, Laboratory ofChemical Endocrinology, Room 1512, Department of Medicine, School of Med-icine, Loma Linda University, Loma Linda, ÇA 92350.




















