
Functional specialization of transcription elongation factors

Succinate dehydrogenase assembly factor 2 is needed forassembly and activity of mitochondrial complex II and fornormal root elongation in Arabidopsis
Shaobai Huang, Nicolas L. Taylor, Elke Stroher, Ricarda Fenske and A. Harvey Millar*
Australian Research Council Centre of Excellence in Plant Energy Biology and Centre for Comparative Analysis of Biomolecu-
lar Networks, The University of Western Australia, Bayliss Building M316,35 Stirling Highway, Crawley, WA 6009, Australia
Received 11 June 2012; revised 22 September 2012; accepted 27 September 2012; published online 29 November 2012.
*For correspondence (e-mail [email protected]).
SUMMARY
Mitochondria complex II (succinate dehydrogenase, SDH) plays a central role in respiratory metabolism as a
component of both the electron transport chain and the tricarboxylic acid cycle. We report the identification
of an SDH assembly factor by analysis of T-DNA insertions in At5g51040, a protein with unknown function
that was identified by mass spectrometry analysis as a low abundance mitochondrial protein. This gene is
co-expressed with a number of genes encoding mitochondrial proteins, including SDH1-1, and has low partial
sequence similarity to human SDHAF2, a protein required for flavin-adenine dinucleotide (FAD) insertion into
SDH. In contrast to observations of other SDH deficient lines in Arabidopsis, the sdhaf2 line did not affect pho-
tosynthetic rate or stomatal conductance, but instead showed inhibition of primary root elongation with early
lateral root emergence, presumably due to the low SDH activity caused by the reduced abundance of SDHAF2.
Both roots and leaves showed succinate accumulation but different responses in the abundance of other
organic acids and amino acids assayed. Isolated mitochondria showed lowered SDH1 protein abundance, low-
ered maximal SDH activity and less protein-bound flavin-adenine dinucleotide (FAD) at the molecular mass of
SDH1 in the gel separation. The short root phenotype and SDH function of sdhaf2 was fully complemented by
transformation with SDHAF2. Application of the SDH inhibitor, malonate, phenocopied the sdhaf2 root archi-
tecture in WT. Whole root respiratory assays showed no difference between WT and sdhaf2, but micro-respi-
rometry of the tips of roots clearly showed low oxygen consumption in sdhaf2 which could explain a
metabolic deficit responsible for root tip growth.
Keywords: succinate dehydrogenase, plant mitochondria, electron transport chain.
INTRODUCTION
The mitochondrion is the powerhouse of the eukaryote cell
within which the tricarboxylic acid (TCA) cycle drives
reduction of the electron transport chain and the subse-
quent oxidative phosphorylation and synthesis of ATP.
Succinate dehydrogenase (SDH EC 1.3.99.1 or complex II,
EC 1.3.5.1) catalyses the oxidation of succinate to form
fumarate. It has a central role in mitochondrial metabolism
as it is a component of both the TCA cycle and the electron
transport chain. The classical SDH enzyme in eukaryotes is
comprised of four subunits, a flavoprotein (SDH1) that con-
tains a bound flavin-adenine dinucleotide (FAD) co-factor,
an iron-sulfur (Fe-S) protein (SDH2) containing three Fe-S
clusters, and two small integral membrane proteins (SDH3
and SDH4) that bind heme to form the b- type cytochrome
(Lemire and Oyedotun, 2002; Sun et al., 2005).
Plants, however, have a divergent SDH composition
when compared to mammalian and yeast forms of the
enzyme. The sequences of SDH1 and SDH2 are relatively
conserved between plants and animals (60–90% identity),
while there is very significant divergence in the sequences
for the membrane anchor proteins, SDH3 and SDH4 (20–
40% identity) (Burger et al., 1996). In addition, the isolated
plant SDH enzyme contains a further four plant-specific
subunits with unknown function (Eubel et al., 2003; Millar
et al., 2004; Huang et al., 2010). A recent study has revealed
SDH (complex II) is more abundant in non-green tissues of
Arabidopsis, such as root, which contrasts with mitochon-
drial complex I which is more abundant in photosynthetic
tissues (Peters et al., 2012). This indicates a potential differ-
ence in the metabolic requirements in shoots and roots in
© 2012 The AuthorsThe Plant Journal © 2012 Blackwell Publishing Ltd
429
The Plant Journal (2013) 73, 429–441 doi: 10.1111/tpj.12041

Arabidopsis for these two major protein complexes of
mitochondrial respiration.
The functional importance of the flavoprotein subunit
(SDH1) in plants has been studied with unexpected and
contrasting results. In Arabidopsis, SDH1 is encoded by two
genes, SDH1-1 (At5g66760) and SDH1-2 (At2g18450), the
former is the main protein found in SDH while the latter is
expressed at a very low level (Leon et al., 2007). Disruption
of SDH1-2 had no effect on the growth and development of
Arabidopsis mutants (Leon et al., 2007). SDH1-1 appears to
be essential for gametophyte development in Arabidopsis.
Down-regulation of SDH1-1 by RNA interference results in
pollen abortion and a reduced seed set, and homozygous
SDH1-1 T-DNA insertion lines failed to complete gameo-
phyte development (Leon et al., 2007). Heterozygous SDH1-
1/sdh1-1 plants are reported to have low SDH activity but
improved photosynthesis and growth in nitrogen-limiting
conditions due to altered stomatal conductance and nitro-
gen assimilation (Fuentes et al., 2011). We have recently
shown that a point mutation in SDH1-1, at a conserved
region in the substrate binding pocket, significantly
reduced SDH activity and the production of mitochondrial
reactive oxygen species in Arabidopsis (Gleason et al.,
2011). This mutation altered salicylic acid-mediated expres-
sion of specific stress and defence genes in Arabidopsis,
and mutant plants exhibited increased susceptibility to spe-
cific fungal and bacterial pathogens as a result (Gleason
et al., 2011).
The functional importance of the iron-sulfur subunit
(SDH2) in plants has also been studied, showing some
similarities and differences to the results for SDH1. The
iron-sulfur subunit (SDH2) of SDH in Arabidopsis is
encoded by three genes, SDH2-1 (At3g27380), SDH2-2
(At5g40650) and SDH2-3 (At5g65165). SDH2-1 and SDH2-2
in Arabidopsis have distinct cell specific expression pat-
terns and promoter activities, and only SDH2-2 appears to
be expressed at a significant level in root tips (Elorza et al.,
2004). The knockout of SDH2-1 did not have any pheno-
type, indicating functional redundancy between SDH2-1
and SDH2-2 (Elorza et al., 2004). SDH2-3 is specifically
expressed in the embryo during seed development (Elorza
et al., 2006). The disruption of SDH2-3 delayed seed germi-
nation, indicating its role at an early step of germination
(Roschzttardtz et al., 2009). In tomato, RNAi of SDH2
increased the rate of net photosynthesis and growth,
mainly due to greater stomatal aperture in these lines (Ara-
ujo et al., 2011). The repression of the SDH2-2 expression
using a guard cell–specific MYB60 promoter had no effect
on changes in stomatal aperture or rates of photosynthesis
in the mutant, suggesting a possible role of mesophyll
cells instead of guard cells in stomatal function (Araujo
et al., 2011). Those plants showed similarities to the phe-
notype of SDH1-1/sdh1-1 plants studied in Arabidopsis
(Fuentes et al., 2011). Based on analysis of metabolites, a
model of SDH regulation of stomatal aperture by mito-
chondrial metabolites such as malate and fumarate has
been proposed (Araujo et al., 2011).
In humans, SDH deficiency has been closely linked to
several different diseases, notably paraganglioma (PGL)
and infantile leukoencephalopathy. In total 70% of familial
cases of head and neck PGL are due to germline mutations
in SDH2, SDH3, or SDH4 (Baysal et al., 2002), and 10% of
sporadic PGL cases are associated with mutations in SDH2
or SDH4 (Schiavi et al., 2005; Timmers et al., 2009). In
comparison, mutations in SDH1 in humans are more rare,
some lead to PGL and some lead to a Leigh-like syndrome
(Burnichon et al., 2010). Two uncharacterized proteins
linked to infantile leukoencephalopathy and PGL have been
identified recently as SDH assembly factors and named
SDHAF1 (Ghezzi et al., 2009) and SDHAF2 (Hao et al.,
2009), respectively. Functional analysis indicated that
SDHAF1 is likely to be involved in Fe-S insertion into SDH2
based on its similarity to proteins with an analogous role
in complex I (Ghezzi et al., 2009), while SDHAF2 has been
show to interact with SDH1 and to be required for insertion
of the flavin into SDH1 (Hao et al., 2009).
We have developed a large list of Arabidopsis mitochon-
dria proteins using proteome analysis (Heazlewood et al.,
2004; Taylor et al., 2011). As a result of our in-depth analy-
sis of mitochondrial peptides for presequence identification
(Huang et al., 2009a) we identified a protein with unknown
function (At5g51040). The transcript for this protein was
co-expressed with SDH1 and SDH2 in plants, and the
protein sequence showed a low level of sequence similarity
to the human SDHAF2. This raised the hypothesis that a
SDHAF2-like protein could exist in plants and might provide
an independent means of altering SDH function in plants
and assessing the differing phenotypes of SDH mutants in
plants. Through characterizing the phenotype of T-DNA
insertion lines for At5g51040 we provided evidence that this
protein is required for normal SDH abundance and activity,
and that the lowered SDH activity did not lead to a stomatal
or photosynthetic phenotype but instead lowered primary
root elongation and root function in Arabidopsis.
RESULTS
Identification, expression and linkage of unknown
function protein At5g51040 to roles associated with
succinate dehydrogenase
Analysis of the mitochondrial proteome from cell culture
using peptide mass spectrometry approaches for prese-
quence analysis (Huang et al., 2009a) revealed a range of
proteins that had not previously been reported in plant
mitochondria. One of these was a single high quality pep-
tide match to At5g51040, a hydrophilic protein with
unknown function containing the DUF339 domain (Taylor
et al., 2011). Our further analyses of Arabidopsis cell
© 2012 The AuthorsThe Plant Journal © 2012 Blackwell Publishing Ltd, The Plant Journal, (2013), 73, 429–441
430 Shaobai Huang et al.

culture mitochondrial matrix extracts have identified three
additional peptides matched to the At5g51040 sequence
(Figure 1a). The four peptides cover 48% of the amino acid
sequence of At5g51040 with a high confidence total protein
score. At5g51040 is predicted to be targeted to mitochon-
dria by many common bioinformatics prediction algo-
rithms (http://suba.plantenergy.uwa.edu.au) (Heazlewood
et al., 2007). Based on genvestigator database, At5g51040
is highly expressed in root and flowering tissues (http//
www.genevestigator.com). Co-expression analysis in Ara-
bidopsis (http://atted.jp) (Obayashi et al., 2009) revealed
that expression of At5g51040 clusters with the expression
of several mitochondrial components, most notably
SDH1 and SDH2 (Figure 1b). The AtProteome database of
plant tissues (http://fgcz-atproteome.unizh.ch/) (Baerenfal-
ler et al., 2008) also showed that peptides for At5g51040
were rare and only found in cell culture and root tissue
samples (Figure 1c). BLASTP (Stephen et al., 2007) of the
At5g51040 protein sequence showed it shared 30% identity
to human SDHAF2 (NP_060311) in a specific 96 AA region
(position 69–165 in At5g51040) and 32% identity to yeast
SDHAF2 (B3LIY9) in the similar region. This succinate
dehydrogenase assembly factor (SDHAF2) is required for
SDH1 flavination in human and yeast (Hao et al., 2009).
However, the expectation score for the match between the
Arabidopsis and human (approximately 1 9 e�5) sequences
was significantly higher than that for the sequences of
SDH1/SDH2 between the two species (Figure 1d) and above
normal limits for claims of functional assignments
(<1 9 e�11) or biological process/pathway assignments
(<1 9 e�23) (Thomas et al., 2003, 2006).
SDHAF2 disruption affects root but not shoot growth and
leads to succinate accumulation
To experimentally investigate the role of At5g51040 in
Arabidopsis we obtained two independent T-DNA insertion
lines, Sail_622_A09 and GABI_879B11 from the Salk Insti-
tute Genomic Analysis Laboratory (http://signal.salk.edu/
cgi-bin/tdnaexpress). Sail_622_A09 had an insert in an in-
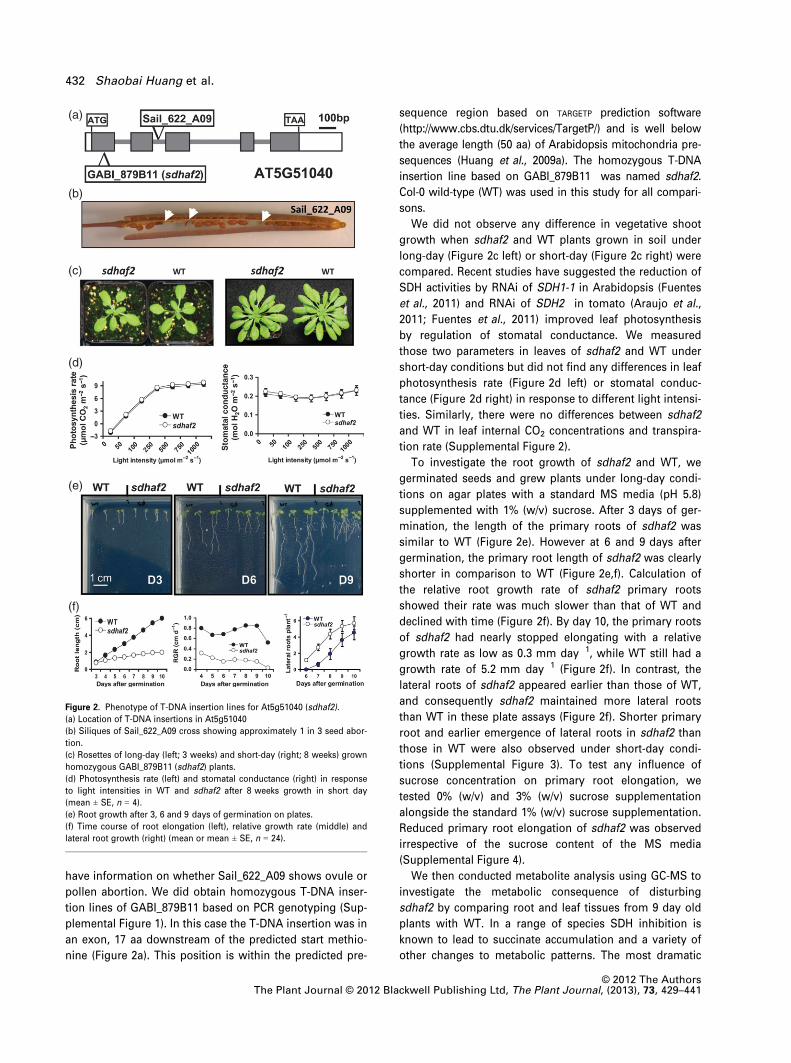
tronic region of the gene (Figure 2a). We could not obtain a
Sail_622_A09 T-DNA homozygous knockout line based on
PCR genotyping (Supplemental Figure 1). Inspection of sili-
ques showed a 1:3 ratio of seed abortion in Sail_622_A09,
suggesting the homozygous genotype is lethal at an early
stage (Figure 2b). This seed abortion phenotype maybe
similar to that observed for SDH1-1 homozygous T-DNA
knockout lines that were shown to fail to complete gameo-
phyte development (Leon et al., 2007), however we do not
(a)
(b)
(d)
(c)
Figure 1. Identification, expression and sequence
similarity of At5g51040.
(a) Mass spectrometry data for identification of
At5g51040 peptides in isolated Arabidopsis mito-
chondrial matrix samples, showing experimental
and theoretical peptide mass, Mascot Ion score
and expectation value.
(b) Co-expression network of At5g51040 in Arabid-
opsis from AttedII analysis (version 6; http://atted.
jp/) with Pearson’s correlation coefficients between
genes pairs inset, with the gene of interest in black
and SDH subunits in grey.
(c) Presence of At5g51040 peptides in the
AtProteome study of different tissue types
(http://fgcz-atproteome.unizh.ch/).
(d) Sequence similarity analysis of At5g51040,
SDH1-1 and SDH2-1 from Arabidopsis with the
nearest homolog in the human proteome,
SDHAF2 (NP_060311), SDHA (NP_004159), SDHB
(NP_002991), showing bit score, expectation
value and maximum sequence identity.
© 2012 The AuthorsThe Plant Journal © 2012 Blackwell Publishing Ltd, The Plant Journal, (2013), 73, 429–441
SDHAF2 in complex II function in Arabidopsis 431
have information on whether Sail_622_A09 shows ovule or
pollen abortion. We did obtain homozygous T-DNA inser-
tion lines of GABI_879B11 based on PCR genotyping (Sup-
plemental Figure 1). In this case the T-DNA insertion was in
an exon, 17 aa downstream of the predicted start methio-
nine (Figure 2a). This position is within the predicted pre-
sequence region based on TARGETP prediction software
(http://www.cbs.dtu.dk/services/TargetP/) and is well below
the average length (50 aa) of Arabidopsis mitochondria pre-
sequences (Huang et al., 2009a). The homozygous T-DNA
insertion line based on GABI_879B11 was named sdhaf2.
Col-0 wild-type (WT) was used in this study for all compari-
sons.
We did not observe any difference in vegetative shoot
growth when sdhaf2 and WT plants grown in soil under
long-day (Figure 2c left) or short-day (Figure 2c right) were
compared. Recent studies have suggested the reduction of
SDH activities by RNAi of SDH1-1 in Arabidopsis (Fuentes
et al., 2011) and RNAi of SDH2 in tomato (Araujo et al.,
2011; Fuentes et al., 2011) improved leaf photosynthesis
by regulation of stomatal conductance. We measured
those two parameters in leaves of sdhaf2 and WT under
short-day conditions but did not find any differences in leaf
photosynthesis rate (Figure 2d left) or stomatal conduc-
tance (Figure 2d right) in response to different light intensi-
ties. Similarly, there were no differences between sdhaf2
and WT in leaf internal CO2 concentrations and transpira-
tion rate (Supplemental Figure 2).
To investigate the root growth of sdhaf2 and WT, we
germinated seeds and grew plants under long-day condi-
tions on agar plates with a standard MS media (pH 5.8)
supplemented with 1% (w/v) sucrose. After 3 days of ger-
mination, the length of the primary roots of sdhaf2 was
similar to WT (Figure 2e). However at 6 and 9 days after
germination, the primary root length of sdhaf2 was clearly
shorter in comparison to WT (Figure 2e,f). Calculation of
the relative root growth rate of sdhaf2 primary roots
showed their rate was much slower than that of WT and
declined with time (Figure 2f). By day 10, the primary roots
of sdhaf2 had nearly stopped elongating with a relative
growth rate as low as 0.3 mm day�1, while WT still had a
growth rate of 5.2 mm day�1 (Figure 2f). In contrast, the
lateral roots of sdhaf2 appeared earlier than those of WT,
and consequently sdhaf2 maintained more lateral roots
than WT in these plate assays (Figure 2f). Shorter primary
root and earlier emergence of lateral roots in sdhaf2 than
those in WT were also observed under short-day condi-
tions (Supplemental Figure 3). To test any influence of
sucrose concentration on primary root elongation, we
tested 0% (w/v) and 3% (w/v) sucrose supplementation
alongside the standard 1% (w/v) sucrose supplementation.
Reduced primary root elongation of sdhaf2 was observed
irrespective of the sucrose content of the MS media
(Supplemental Figure 4).
We then conducted metabolite analysis using GC-MS to
investigate the metabolic consequence of disturbing
sdhaf2 by comparing root and leaf tissues from 9 day old
plants with WT. In a range of species SDH inhibition is
known to lead to succinate accumulation and a variety of
other changes to metabolic patterns. The most dramatic
(a)
(b)
(c)
(d)
(e)
(f)
Figure 2. Phenotype of T-DNA insertion lines for At5g51040 (sdhaf2).
(a) Location of T-DNA insertions in At5g51040
(b) Siliques of Sail_622_A09 cross showing approximately 1 in 3 seed abor-
tion.
(c) Rosettes of long-day (left; 3 weeks) and short-day (right; 8 weeks) grown
homozygous GABI_879B11 (sdhaf2) plants.
(d) Photosynthesis rate (left) and stomatal conductance (right) in response
to light intensities in WT and sdhaf2 after 8 weeks growth in short day
(mean ± SE, n = 4).
(e) Root growth after 3, 6 and 9 days of germination on plates.
(f) Time course of root elongation (left), relative growth rate (middle) and
lateral root growth (right) (mean or mean ± SE, n = 24).
© 2012 The AuthorsThe Plant Journal © 2012 Blackwell Publishing Ltd, The Plant Journal, (2013), 73, 429–441
432 Shaobai Huang et al.

response was accumulation of succinate to 7.5 and 6.4-fold
higher in the leaves and roots of sdhaf2 when compared
with WT (Figure 3). Changes in organic acids of the TCA
cycle and amino acids with significant differences in abun-
dance are presented in Figure 3 and other detected amino
acids and metabolites are listed in full in Supplemental
Table 1. In leaf tissues a range of organic acids (malate,
fumarate, citrate, aconitate) and amino acids (Ser, Gly, Glu,
Pro, homoserine) accumulated in sdhaf2 (Figure 3). In root
tissues, a low abundance of malate and a-ketoglutaratewere observed in sdhaf2, potentially due to a slowing of
the TCA cycle via SDH disruption. The significant differ-
ence in accumulation of these metabolites between roots
and leaves also demonstrates the restricted transportation
of those metabolites between the two tissues. The high
accumulation of glycine, homoserine and glutamate in
leaves was not observed in sdhaf2 roots (Figure 3). Low
homoserine and glutamate in the roots of sdhaf2 could be
related to the low level of their TCA cycle precursors,
malate and a-ketoglutarate, respectively (Figure 3).
Low SDH assembly, activity and less flavinated SDH1
in sdhaf2
To assess protein abundance differences in sdhaf2, mito-
chondria were isolated from hydroponic shoot material
and compared to WT mitochondria using Differential in
Gel Electrophoresis (DIGE). No dramatic differences in
mitochondrial protein abundances between WT and sdhaf2
were observed (Figure 4a). However, through image analy-
sis we found two protein spots with molecular mass of
approximately 70 kDa which were significantly decreased
in abundance in sdhaf2, by 1.9- and 1.6- fold, respectively
(Figure 4b). Both protein spots were analysed by mass
spectrometry and the protein identified in both cases was
SDH1-1 (At5g66760) (Figure 4c). To more precisely
determine the relative abundance of SDHAF2 and SDH1-1
in sdhaf2, we optimised selected reaction monitoring
assays for SDHAF2 and SDH1-1 (At5g66760), and used suc-
cinyl-CoA ligase abundance (At5g23250) as a control.
Quantification of these peptides in trypsin digestions of
whole mitochondria showed that SDHAF2 and SDH1-1 are
both present at significantly reduced levels in sdhaf2,
about 50 and 30% respectively. No difference in the abun-
dance of the mitochondrial control, succinyl-CoA ligase,
was observed between sdhaf2 and WT (Figure 4d). There-
fore the phenotype of sdhaf2 may be explained by a
knockdown of SDHAF2 and the concomitant reduction of
SDH1-1 abundance in mitochondria. To confirm the impact
of this we then investigated mitochondrial SDH function by
directly measuring succinate-dependent DCPIP reduction
and succinate-dependent O2 consumption rate in mito-
chondrial samples isolated from sdhaf2 and WT. Mitochon-
dria from sdhaf2 had only 35% of SDH enzymatic activity
and 40% of the succinate-dependent O2 consumption
rate of mitochondria isolated from WT plants (Figure 5a,b).
We also confirmed that there was no major effect of sdhaf2
on the activity of the remainder of the electron transport
chain by assaying NADH-dependent O2 consumption rate
(Figure 5c). SDH blue-native in-gel enzymatic activity stain-
ing also revealed a significant reduction of SDH activity in
Figure 3. Changes in organic acids and amino
acids in root and shoot material from sdhaf2
compared to WT plants. Significant fold differ-
ences (n = 5, P < 0.05) in organic acids and
amino acids in the roots and leaves of 9-day
old plants. Black arrow indicates reaction direc-
tion. The inhibition site of SDH by sdhaf2 is
indicated.
© 2012 The AuthorsThe Plant Journal © 2012 Blackwell Publishing Ltd, The Plant Journal, (2013), 73, 429–441
SDHAF2 in complex II function in Arabidopsis 433
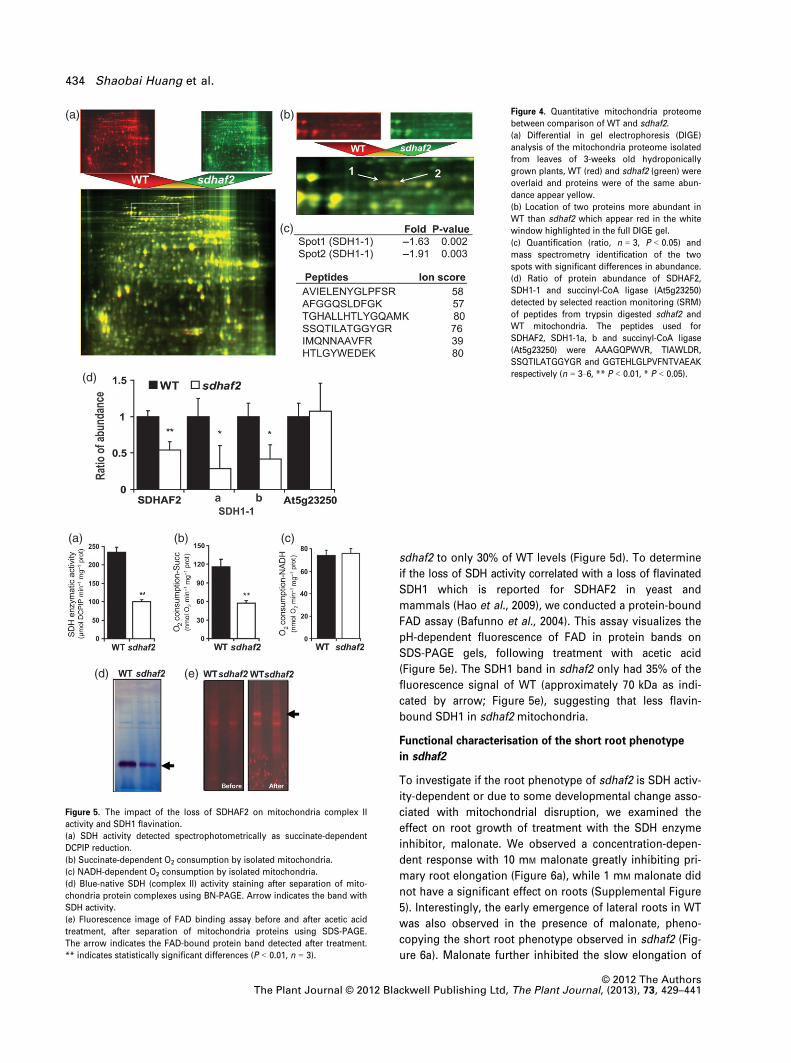
sdhaf2 to only 30% of WT levels (Figure 5d). To determine
if the loss of SDH activity correlated with a loss of flavinated
SDH1 which is reported for SDHAF2 in yeast and
mammals (Hao et al., 2009), we conducted a protein-bound
FAD assay (Bafunno et al., 2004). This assay visualizes the
pH-dependent fluorescence of FAD in protein bands on
SDS-PAGE gels, following treatment with acetic acid
(Figure 5e). The SDH1 band in sdhaf2 only had 35% of the
fluorescence signal of WT (approximately 70 kDa as indi-
cated by arrow; Figure 5e), suggesting that less flavin-
bound SDH1 in sdhaf2 mitochondria.
Functional characterisation of the short root phenotype
in sdhaf2
To investigate if the root phenotype of sdhaf2 is SDH activ-
ity-dependent or due to some developmental change asso-
ciated with mitochondrial disruption, we examined the
effect on root growth of treatment with the SDH enzyme
inhibitor, malonate. We observed a concentration-depen-
dent response with 10 mM malonate greatly inhibiting pri-
mary root elongation (Figure 6a), while 1 mM malonate did
not have a significant effect on roots (Supplemental Figure
5). Interestingly, the early emergence of lateral roots in WT
was also observed in the presence of malonate, pheno-
copying the short root phenotype observed in sdhaf2 (Fig-
ure 6a). Malonate further inhibited the slow elongation of
(a)
(d)
(b)
(c)
Figure 4. Quantitative mitochondria proteome
between comparison of WT and sdhaf2.
(a) Differential in gel electrophoresis (DIGE)
analysis of the mitochondria proteome isolated
from leaves of 3-weeks old hydroponically
grown plants, WT (red) and sdhaf2 (green) were
overlaid and proteins were of the same abun-
dance appear yellow.
(b) Location of two proteins more abundant in
WT than sdhaf2 which appear red in the white
window highlighted in the full DIGE gel.
(c) Quantification (ratio, n = 3, P < 0.05) and
mass spectrometry identification of the two
spots with significant differences in abundance.
(d) Ratio of protein abundance of SDHAF2,
SDH1-1 and succinyl-CoA ligase (At5g23250)
detected by selected reaction monitoring (SRM)
of peptides from trypsin digested sdhaf2 and
WT mitochondria. The peptides used for
SDHAF2, SDH1-1a, b and succinyl-CoA ligase
(At5g23250) were AAAGQPWVR, TIAWLDR,
SSQTILATGGYGR and GGTEHLGLPVFNTVAEAK
respectively (n = 3–6, ** P < 0.01, * P < 0.05).
(a)
(d) (e)
(b) (c)
Figure 5. The impact of the loss of SDHAF2 on mitochondria complex II
activity and SDH1 flavination.
(a) SDH activity detected spectrophotometrically as succinate-dependent
DCPIP reduction.
(b) Succinate-dependent O2 consumption by isolated mitochondria.
(c) NADH-dependent O2 consumption by isolated mitochondria.
(d) Blue-native SDH (complex II) activity staining after separation of mito-
chondria protein complexes using BN-PAGE. Arrow indicates the band with
SDH activity.
(e) Fluorescence image of FAD binding assay before and after acetic acid
treatment, after separation of mitochondria proteins using SDS-PAGE.
The arrow indicates the FAD-bound protein band detected after treatment.
** indicates statistically significant differences (P < 0.01, n = 3).
© 2012 The AuthorsThe Plant Journal © 2012 Blackwell Publishing Ltd, The Plant Journal, (2013), 73, 429–441
434 Shaobai Huang et al.

primary roots of sdhaf2 (Figure 6a), presumably due to the
even more limited SDH activity in the mutant. To check
whether the respiration rate of root mitochondria was
contributing to the short root phenotype of sdhaf2, we
measured respiration rate of whole root systems using a
Clark-type O2 electrode. We found slightly higher root res-
piration in sdhaf2 than in WT, potentially due to the larger
number of lateral roots in sdhaf2 contributing to a higher
O2 consumption rate (Figure 6b). To remove the impact of
lateral roots and other expanded regions of primary root,
we applied a micro-respirometry approach based on fluo-
rescence analysis of O2 consumption to measure the respi-
ration of just the primary root tips (5 mm) of WT and
sdhaf2 in a small liquid volume (2.5 ll). As shown in Fig-
ure 6c, we found the O2 consumption rate of sdhaf2 root
tips over 2 h was only about 46% of that measured for WT,
indicating a limitation of respiration and thus of energy
production in the primary root tips of sdhaf2.
Complemented of sdhaf2 by a full length cDNA restores
the root phenotype and SDH function
As the T-DNA insertion in the presequence of the protein
only decreased SDHAF2 protein abundance (Figure 4d),
rather than eliminating SDHAF2, we sought to genetically
confirm that the short root phenotype in sdhaf2 was
caused by this partial reduction. We cloned an At5g51040
full length cDNA (Supplemental Figure 1 for primer infor-
mation) and transformed it under the control of the 35S
CMV promoter in the pGreen pMDC43 vector (Curtis and
Grossniklaus, 2003) into sdhaf2. We obtained seven inde-
pendent lines that restored primary root elongation of
sdhaf2 to WT levels and images for two lines are shown
here as examples (Figure 7a). Assays on purified shoot
mitochondria from these lines showed full recovery of the
level of FAD-SDH1, the activity of the intact native complex
and succinate-dependent respiration after the complemen-
tation (Figure 7b–e). To ensure that similar changes were
occurring in roots and correlated with the root phenotype,
we isolated root mitochondria and measured low levels of
FAD-SDH1, succinate-dependent oxygen consumption, and
SDH activity in sdhaf2. All three parameters were fully
recovered in the complemented line (Figure 7f–h). There-
fore, we concluded that knockdown of the product of
At5g51040 is responsible for the short root phenotype and
dysfunction of SDH in the sdhaf2 line.
DISCUSSION
As a component of both mitochondrial electron transport
chain and the TCA cycle, succinate dehydrogenase (SDH)
has attracted significant attention in research of mammalian,
yeast and plant systems. The similarity and diversity of the
sequences of the four classical subunits of SDH in mammals
and plants have been highlighted (Burger et al., 1996;
Adams et al., 2001; Huang et al., 2010). However, the factors
responsible for the assembly of SDH in any eukaryote have
been unknown until the recent discovery of two assembly
factors for SDH associated with different diseases in humans
(Ghezzi et al., 2009; Hao et al., 2009). In this study, we pres-
ent evidence that a low similarity homolog of SDHAF2 exists
in plants and had already been identified as the unknown
function protein At5g51040 in the mitochondrial proteome of
Arabidopsis at the time that SDHAF1 and SDHAF2 were first
reported in humans. Functional analysis has now revealed
this protein has a role as a SDH assembly factor in plants
and also shows that loss of SDH function can impact pri-
mary root elongation in Arabidopsis.
Assembly of SDH in plants and a role for sdhaf2
Comparisons at a number of levels suggest the product
of At5g51040 is not a SDH subunit in plants, but that it
has a similar function to SDHAF2 in mammals and yeast
in SDH assembly. The product of At5g51040 is a low
abundance component in plant mitochondria that has
not been identified by gel-based proteomics but has
been observed in mitochondrial samples analysed by
LC-MS/MS (Taylor et al., 2011). It is sub-stoichiometric
to SDH subunits and has not been identified in purified
SDH complexes (Eubel et al., 2003; Millar et al., 2004;
Klodmann et al., 2011). Its proposed role as an assembly
factor comes from its low level sequence similarity to
(a)
(b) (c)
Figure 6. Respiration rate dependence of root phenotype in sdhaf2.
(a) Root growth after 8 days of germination treated with or without 10 mM
malonate.
(b) Oxygen consumption rate of whole roots from 9-day old plants.
(c) Micro-respirometry traces of five 9-day old plant primary root tip oxygen
consumption rates from sdhfa2 and WT with two biological replicates and
11 technical measurements over 120 mins of assay.
© 2012 The AuthorsThe Plant Journal © 2012 Blackwell Publishing Ltd, The Plant Journal, (2013), 73, 429–441
SDHAF2 in complex II function in Arabidopsis 435

human SDHAF2 (Figure 1) and knockdown of sdhaf2
leads to suppressed SDH activity, less FAD-SDH1 and a
loss of SDH1 protein abundance (Figures 4 and 5). In
yeast, SDHAF2 co-immunoprecipitates a proportion of
SDH1 and while deletion of SDHAF2 leads to a complete
loss of FAD-SDH1, some unbound SDH1 remains in yeast
mitochondria (Hao et al., 2009). In Arabidopsis the
decrease in SDH activity and function in sdhaf2 is not as
dramatic due to the presence of significant residual lev-
els of SDHAF2 protein in sdhaf2 (Figure 4d). We would
expect the complete deletion of SDHAF2 function in
plants to be lethal, as was observed for Sail_622_A09,
and as is reported for the knockout of SDH1 (Leon et al.,
2007). In yeast and human SDHAF2, a conserved glycine
residue is required for SDH1:SDHAF2 physical interac-
tion, this residue is also conserved in the Arabidopsis
SDHAF2 sequence (Supplemental Figure 6), suggesting
conservation of this SDH1 interaction site despite the
divergence of these sequences. BN-PAGE activity stain-
ing did not reveal any impact on the molecular mass of
SDH in sdhaf2 and thus there is no evidence for the
accumulation of any assembly intermediates in the
mutant. There is a close correlation between the amount
of SDH1-1 protein (Figure 4) and the loss of SDH activity
(Figure 5) which suggests that, unlike in yeast (Hao et al.,
2009), non-flavinated SDH1 does not exists in sdhaf2.
Although it is possible that, the reduced SDH1 is the con-
sequence of a reduction in the capacity to have FAD
incorporated in SDH1 (leading to SDH1 instability or deg-
radation) we are relying on yeast and mammalian evi-
dence to make this claim. In rice, the TPR protein
Os11g32480 shares 69% identity to At5g51040 at the
amino acid sequence level. This rice protein contains the
same conserved glycine residue described in mammals
and is likely to function as the rice SDHAF2. However,
we have not identified this protein to date in our rice
mitochondria proteome studies (Huang et al., 2009b).
Similarity searches using the sequence of the SDHAF1
protein from humans (Ghezzi et al., 2009) also reveals a
low similarity match (45% identity in a 40 aa window,
E = 2.3 9 e�2) in Arabidopsis (At2g39725). This protein is
also predicted to be a mitochondrial protein, but has not
been identified to date in plant mitochondrial proteomic
(a)
(b) (c)
(f) (g)
(h)
(d) (e)
Figure 7. Complementation of sdhaf2 with full-length cDNA of SDHAF2 dri-
ven by the 35S promoter.
(a) Root growth of WT, sdhaf2 and complemented lines 1 and 2 after 9 days
of germination.
(b) FAD binding assay following acetic acid treatment after separation of
shoot mitochondria protein using SDS-PAGE. The arrow indicates the FAD
band induced by acetic acid treatment.
(c) Succinate-dependent O2 consumption by isolated shoot mitochondria.
(d) Succinate dehydrogenase activity from shoot mitochondria detected
spectrophotometrically using succinate-dependent DCPIP reduction.
(e) Blue-native SDH (complex II) activity staining after separation of shoot
mitochondrial protein complexes using BN-PAGE. Arrow indicates band with
SDH activity. ** indicates statistically significant difference (P < 0.01, n = 3).
(f) FAD binding assay from root mitochondria.
(g) Succinate-dependent O2 consumption by root mitochondria. (h) Succi-
nate dehydrogenase activity from root mitochondria using DCPIP reduction.
© 2012 The AuthorsThe Plant Journal © 2012 Blackwell Publishing Ltd, The Plant Journal, (2013), 73, 429–441
436 Shaobai Huang et al.

studies. Therefore is seems plausible that functional ho-
mologs of both SDHAF2 and SDHAF1 are present in
plants, albeit with very diverse sequence.
Different metabolic consequences of SDH disruption
across organisms
Reduction in SDH activity by mutations of SDH1, SDH2,
and SDH3 inhibit yeast growth when supplied with
non-fermentable carbon sources and lead to organic acid
accumulation and acetate secretion (Lombardo et al., 1990;
Daignan-Fornier et al., 1994). In humans, mutations in SDH
subunits decreases SDH activity and are linked to a series
of neurological and blood disorders (Baysal et al., 2002).
SDH mutations in human patients lead to accumulation of
succinate and fumarate in the cytosol of cells that inhibit
propyl hydroxylase and lead to tumourigenesis (Rutter
et al., 2010). In plants, however, inhibition of SDH by par-
tial reduction of SDH subunits has been reported to
improved leaf photosynthesis by increasing stomatal con-
ductance in two different species (Araujo et al., 2011; Fuen-
tes et al., 2011). One possible explanation of regulation of
stomatal aperture in these SDH depleted lines has been
low organic acid abundances (such as malate and fuma-
rate) in the mesophyll cells of leaves of the mutants that
limit their subsequent transportation to guard cells for
osmotic regulation (Araujo et al., 2011). In this study, no
differences in photosynthetic rate or stomatal conductance
were found in comparisons of sdhaf2 and WT, despite
decreases in SDH activity similar to the levels in these
recent reports (Araujo et al., 2011; Fuentes et al., 2011).
Rather, leaves of sdhaf2 accumulated larger amounts of
TCA cycle-related metabolites (such as malate, fumarate)
than those of WT (Figure 3). While this is starkly different
from the metabolic observations in tomato leaves with
SDH2 dysfunction (Araujo et al., 2011), it is similar to what
is known from studies on yeast and human SDH muta-
tions. This finding may expalin why no difference in sto-
matal conductance occurred in sdhaf2, based on the idea
that TCA-cycle related organic acids rather than succinate
accumulation played the key role in this stomatal regula-
tion (Araujo et al., 2011). It is unclear how mutants in
which SDH activity is lowered in various ways show these
very different outcomes in plant leaves. However, we did
observe decreases of malate and a-ketoglutarate in roots
of sdhaf2 (Figure 3) which are tissues in which the main
phenotype of sdhaf2 plants is observed.
A role for SDHAF2 in root elongation
We have found sdhaf2 had a short root phenotype (Figure 2),
which has not been reported in other SDH1 or SDH2 deficient
mutants in plants to our knowledge. It can be envisaged that
mitochondria play an essential role during root growth and
active cell division in the root meristem and that these cells
may require more cellular energy than other root tissue
types. Reduction of enzymatic activities of several TCA cycle
enzymes using RNAi in tomato have resulted in low rates of
root respiration and reduction in root dry mass in 5-weeks-
old plants (van der Merwe et al., 2010). SDH RNA anti-sense
plants have been the exception as they were reported to have
no change in root dry mass (Araujo et al., 2011). In Arabidop-
sis, a significant number of mutants inmitochondrial compo-
nents display short roots on plant assays but normally this is
also accompanied with gross losses of vegetative shoot
growth. For example, mutants of mitochondrial protease
Lon1 (Rigas et al., 2009) and prohibitin (Van Aken et al.,
2007) have short roots and very reduced shoot growth.
Mutants affecting mitochondrial complex I are often small
plants with reduced root and shoot growth (de Longevialle
et al., 2007; Kuhn et al., 2009; Meyer et al., 2009; Koprivova
et al., 2010), indicating that these genes are essential for
whole plant growth characteristics. In contrast, sdhaf2 shows
a specific inhibition of primary root but not shoot growth,
and an acceleration of lateral root growth. The role of
SDHAF2 in root elongation could be linked to the rela-
tively higher abundance of SDH compared to other electron
transport chain complexes in non-green tissues (Peters
et al., 2012) and the presence of more SDHAF2 peptides in
roots than leaves in the AtProteome dataset (Figure 1). Stud-
ies of 13C labelling and flux analysis in Xanthium strumarium
leaves suggest the TCA cycle operates as two split sets of
reactions yielding glutamate and fumarate in the light (Tcher-
kez et al., 2009). This would decrease steady-state SDH flux
to very low levels in photosynthetic tissues in the light and
would argue that SDH would play a maximal role in the dark
and in non-photosynthetic tissues. Therefore, root-specific
phenotypes in SDH mutants like sdhaf2may be explained by
a higher control coefficient in mitochondrial energy produc-
tion by SDH in root tips than in leaf tissues. The possibility of
the involvement of SDHAF2 in mitochondrial dysfunctions in
addition to the one in SDH function cannot be ruled out by
our data. Multiple targets might explain different phenotypes
in SDHmutants in plants. However, the largemajority of elec-
tron transport chain assembly factors known to date have
single sites of action (Kim et al., 2012; Mimaki et al., 2012),
the role of SDHAF2 in yeast is specifically in SDH1 binding
that then allows FAD entry, and no other target in mitochon-
dria for SDHAF2 has been found in yeast and mammals (Hao
et al., 2009). Discovery and study of other mitochondrial
mutants causing short root phenotypes without shoot
growth effects deserve further investigation to resolve this
question.
EXPERIMENTAL PROCEDURES
Plant lines
Seeds of Arabidopsis (Arabidopsis thaliana) ecotype Columbia-0(WT) and T-DNA insertion lines (Sail_622_A09 and GABI_879B11)were obtained from ABRC. Plants were screened for homozygous
© 2012 The AuthorsThe Plant Journal © 2012 Blackwell Publishing Ltd, The Plant Journal, (2013), 73, 429–441
SDHAF2 in complex II function in Arabidopsis 437

insertion using standard PCR based methods using the primerslisted in Supplemental Figure 1.
Plant growth conditions
Seeds were sown on a 1:3:1 of Perlite (Chillagoe Perlite Pty Ltd.,http://chillagoeperlite.com): shamrock compost (http://www.bum.ie): vermiculite soil mix (Peter Bacon Enterprises, Australia) cov-ered with transparent acrylic hoods. After 3 days at 4°C in dark,plants were grown under controlled conditions as follows: ashort-day photoperiod (8 h of light/16 h of dark), a light intensityof 250 lM m�2 s�1, a relative humidity of 70%, and a temperaturecycle of 22°C day/17°C night; a long-day photoperiod (16 h oflight/8 h of dark), a light intensity of 200 lM m�2 s�1, a relativehumidity of 70%, and a temperature cycle of 22°C day/17°C night.The hoods were removed after 1 week. For plate growth, seedswere sown on ½ MS Gamborg B5 (SIGMA G1019; http://www.sigmaaldrich.com) plates containing 0.75% agar, 1% sucrose,1.8 mM MES with pH at 5.8 adjusted by KOH. For sucrose treat-ment, 0%, 1 and 3% (w/v) of sucrose were added to the medium.For specific SDH inhibitor treatment, malonate were added to thefinal concentration of 0, 1, and 10 mM in the media. After 2 days at4°C in the dark, plates were transferred to long-day controlledconditions as described above and set in a vertical position tofacilitate analysis of root length.
Analysis of Arabidopsis mitochondria proteome and
identification of At5g51040
Previously, we had conducted in-depth analysis of the mitochon-drial proteome to define mitochondrial protein pre-sequencesusing mitochondrial matrix proteins isolated from Arabidopsis cellculture. We did not detect a pre-sequence of At5g51040, but inthese analyses of separated matrix proteins we found high qualityMS/MS spectra matching to At5g51040 as listed in Figure 1a. Datacollection was according to the methods outlined in Huang et al.(2009a) and Taylor et al. (2011).
Isolation of mitochondria from hydroponic plants using
gradient centrifugation
Shoot mitochondria were isolated from 3-week-old hydroponicallygrown Arabidopsis seedlings using the method previouslydescribed (Lee et al., 2008). The final mitochondrial samples wascollected and washed three times by dilution with washing buffer(0.3 M Sucrose; 10 mM TES, pH 7.4) and centrifugation at 20000g for15 min at 4�C. The crude root mitochondria were isolated from roottissue of 3-week-old hydroponic grown Arabidopsis seedling usingthe differential centrifugation steps outlined in (Lee et al., 2008) andsamples were collected and washed with an buffered solution (0.3 M
Sucrose, 10 mM TES, pH 7.4,) by centrifugation at 20,000 g for15 min at 4�C. Aliquots were used directly for assays or frozen forfurther analysis.
Protein bound-flavin assay
A protein bound-flavin assay was adapted from a methoddescribed by (Bafunno et al., 2004). After SDS-PAGE separation ofmitochondrial proteins, the protein gel was incubated for 30 minin 10% acetic acid to adjust pH. The gel was scanned before andafter acetic acid treatment. Protein bands with bound-flavin werevisualized by acetic acid treatment revealing increased fluores-cence. Excitation at 488 nm and emission at 526 nm was moni-tored as evidence of protein-bound flavin (FAD) using a TyphoonTrio laser imager (GE Healthcare, http://www.gelifesciences.com).
Succinate-dependent and NADH-dependent oxygen
consumption using an O2 electrode
Oxygen consumption by purified mitochondria was measured by acomputer-controlled Clark-type O2 electrode (Hansatech-Instruments,http://www.hansatech-instruments.com). All reactions were carriedout at 25°C using 1 ml of mitochondrial reaction medium (0.3 M
sucrose, 10 mM TES, 10 mM NaCl, 4 mM MgSO4, 0.1% (w/v) BSA, pH7.2) and 100 lg of mitochondrial protein. To investigate the succi-nate dependent O2 consumption rate, succinate (5 mM) was addedto the reaction solution. To investigate the NADH-dependent O2 con-sumption, NADH (1 mM) instead of succinate was added to the reac-tion solution.
SDH activity assay by spectrophotometric monitoring of
DCPIP reduction
The SDH activity was determined based on the method describedby Huang et al. (2010). The mitochondrial fraction (approximately20 lg protein) was assayed for activity spectrophotometrically bymonitoring absorbance at 600 nm, at 25°C, in 1 ml of a reactionmedium (50 mM potassium phosphate pH 7.4, 10 mM sodium suc-cinate, 0.1 mM EDTA, 0.1% (w/v) BSA, 10 mM potassium cyanide,0.12 mM dichlorophenolindophenol (DCPIP), 1.6 mM phenazinemethosulfate). An extinction coefficient of 21 mM
�1 cm�1 at600 nm for reduced DCPIP was used for calculations.
Blue-native gel SDH activity staining
Blue-native (BN) gel electrophoresis was carried using the methodof (Eubel et al., 2005). For SDH activity staining, the BN-native gelwere soaked in a staining medium (84 mM succinate Na, 0.2 mM
phenazine methosulfate, 2 mg ml�1 nitro tetrazolium blue, 4.5 mM
EDTA, 10 mM potassium cyanide, 50 mM potassium phosphatebuffer, pH 7.5). After 2–3 h of staining, the gels were scanned.
Differential In-Gel Electrophoresis (DIGE) analysis
Mitochondria proteins (50 lg) of WT and sdhaf2, as well as 50 lgof 1:1 mixture of both samples, were acetone-precipated andre-solubilized in lysis buffer [7 M urea, 2 M thiourea, 4% (w/v)CHAPS, 40 mM Tris base, pH 8.5]. The individual samples labelledwith 400 pmol of fluorescent dyes Cy2, Cy3 and Cy5 (GE Health-care) and then combined and separated on IEF strips (pH 3–10non-linear, 24 cm; GE Healthcare) according to the manufacturer’sinstructions. After a brief wash in 19 gel buffer, the strips weretransferred on top of a 12% (v/v) polyacrylamide gel where theywere covered with 1.2% (w/v) agarose in 19 gel buffer. Two-dimensional gels were run at 50 mA current per gel for 6 h. Thefluorescence-labelled proteins were visualized on a Typhoon Triolaser scanner (GE Healthcare) and image comparison was con-ducted using the DECYDER
TM software package (version 6.5; GEHealthcare) with statistical analysis of three independent experi-ments. Gel pictures were overlaid using the IMAGE QUANT TL
TM soft-ware (GE Healthcare). Mass spectrometry identification of proteinspots was conducted as outlined in Lee et al. (2008).
Quantification of peptide abundance using selected
reaction monitoring
Protein extracts from whole mitochondrial trypsin digests wereanalysed on an Agilent 6430 QqQ mass spectrometer with an HPLCChip Cube source (Agilent Technologies, http://www.home.agilent.com). The chip consisted of a 160-nl enrichment column (Zorbax300SB-C18, 5-lm pore size) and a 150-mm separation column
© 2012 The AuthorsThe Plant Journal © 2012 Blackwell Publishing Ltd, The Plant Journal, (2013), 73, 429–441
438 Shaobai Huang et al.

(Zorbax 300SB-C18, 5-lm pore size) driven by Agilent Technologies1200 series nano/capillary LC system. Both systems were con-trolled by MassHunter Workstation Data Acquisition for QqQ (ver-sion B.03.01, build 2600; Agilent Technologies). Peptides wereloaded onto the trapping column at 3 ll min�1 in 5% (v/v) acetoni-trile and 0.1% (v/v) formic acid with the chip switched to enrich-ment and using the capillary pump. The chip was then switched toseparation, and peptides were eluted during a 30-min gradient (5%[v/v] acetonitrile to 100% [v/v] acetonitrile) directly into the massspectrometer. Parent ions and transitions (three transitions per par-ent ion, one quantifier and two qualifiers) were selected based onprevious Q-TOF experiments and were 478.3/742.4 (478.3/557.3,478.3/813.4) for AAAGQPWVR (SDHAF2); 437.7/660.3 (437.7/589.3and 437.7/403.2) for TIAWLDR (SDH1-1 a); 655.8/794.4 (655.8/681.3and 655.8/610.3) for SSQTILATGGYGR (SDH1-1 b) and 614/879.5(614/618.3 and 614/418.2) for GGTEHLGLPVFNTVAEAK (At5g23250).The mass spectrometer was run in positive ion mode, and for eachparent/transition the fragmentor was set to 130, dwell time was5 ms and collision energy was 12.8 V (AAAGQPWVR), 6.8 V (TIA-WLDR), 17.9 V (SSQTILATGGYGR) and 12.9 V (GGTEHLGLPVFNT-VAEAK). SRM chromatograms were analysed in MassHunterWorkstation Quantitative Analysis (version B.04.00, build 4.0.225.0;Agilent Technologies) and quantitative results were obtained byintegrating the area under the peak of each quantifier transition.Qualifiers and retention time information were used to confirm theright peak was integrated. The data was expressed as relative peakarea abundance compared with WT.
Metabolite extraction and GC-MS analysis
Metabolites were extracted from frozen 9-day-old Arabidopsisroots and leaves according to the method described by (Shingaki-Wells et al., 2011) and derivatized metabolite samples were analy-sed on an Agilent GC/MSD system (Agilent Technologies). RawGC-MS data pre-processing and statistical analysis were per-formed using METABOLOME-EXPRESS software (version 1.0, http://www.metabolome-express.org). Detailed methods using this analysisapproach have already been documented (Carroll et al., 2010).
Leaf gas exchange measurements
Gas-exchange parameters were measured on leaves of 8-week-oldplants grown in short day conditions using a LI-6400 XT infraredgas analyzer (Li-Cor, http://www.licor.com). All measurementswere performed after at least 2 h of illumination. One fully devel-oped leaf per plant was enclosed in a 60-mm2 leaf chamber. Theleaf was acclimated in the leaf chamber to 22°C, relative humidity70% and a CO2 concentration of 400 ppm with light intensity of250 lM m�2 s�1. Then, a series of light intensities (0, 50, 100, 250,500, 700, 1000 lM m�2 s�1) were applied. Gas-exchange parame-ters, including CO2 assimilation rate, stomatal conductance, tran-spiration rate and internal CO2 concentration, were recorded infour independent measurements.
Tissue oxygen consumption assays
O2 consumption by root tissues (approximately 80 mg) wasmeasured by a computer-controlled Clark-type O2 electrode unitas described above for O2 consumption by isolated mitochon-dria. The respiration medium for root tissues contained 10 mM
HEPES, 10 mM MES, 2 mM CaCl2 at pH 7.2. O2 consumption byroot tips (5 x 5 mm primary root tips) was recorded in thesame medium as root tissues using an XF96 extracellular fluxanalyser (Seahorse Biosciences, http://www.seahorsebio.com)over 120 min according to manufacturer’s instructions. Measure-ments were performed by assays in a 2.5 ll volume by
repeated movement of the XF96 analyser heads and mixing ina 100 ll of respiration solution in the wells between sets of5 min analyses.
Complementation of the sdhaf2 line
The complementation of the sdhaf2 mutant (GABI_879B11) linewas conducted using the full length At5g51040 cDNA. The result-ing sequence was recombined into the GATEWAY pDONRTM
vector (Invitrogen, http://www.invitrogen.com), verified by DNAsequencing, and recombined into a modified binary vectorpMDC43 with cauliflower mosaic virus 35S promoter (Curtis andGrossniklaus, 2003). This construct was introduced into Agrobac-terium tumefaciens which was then used to transform homozy-gous sdhaf2 mutants by a modified floral dip method(Pracharoenwattana et al., 2005). Transformed plants wereselected by germinating seedlings on agar plates containing25 mg L�1 hygromycin.
ACKNOWLEDGEMENTS
This research was funded by support from the Australian ResearchCouncil Centre of Excellence in Plant Energy Biology (CE0561495)and The University of Western Australia and Agilent TechnologiesAustralia support for the Centre for Comparative Analysis of Biomo-lecular Networks to AHM. AHM was funded as an ARC AustralianFuture Fellow (FT110100242), ES was funded as an ARC Post-doc-toral Fellow (DP110104865). We thank ABRC for provision of T-DNAinsertion lines.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the online ver-sion of this article.Table S1. GC-MS metabolomic analysis of roots and leaves of WTand sdhaf2 line of 9-old day plants.
Figure S1. PCR genotyping and primers for Sail_622_A09 andGABI_879B11 and primers for amplification of full lengthAt5g51040 cDNA for complementation.
Figure S2. Internal CO2 concentrations (left) and transpiration rate(right) in response to light intensities in WT and sdhaf2 after8 weeks growth in short day conditions.
Figure S3. Root growth of WT and sdhaf2 under short-day condi-tions after 3, 6, 10 and 12 days of germination on plates.Figure S4. Root growth on the plates supplemented with 0%, 1and 3% sucrose after 9 days of germination in long day condi-tions.Figure S5. Root growth under long-day conditions after 8 days ofgermination treated with or without 1 mM malonate.
Figure S6. Protein sequence alignment of the At5g51040 proteinsequence in Arabidopsis with the sequence of Human SDHAF2(NP_060311) and Yeast SDHAF2 (B3LIY9).
REFERENCES
Adams, K.L., Rosenblueth, M., Qiu, Y.-L. and Palmer, J.D. (2001) Multiple
losses and transfers to the nucleus of two mitochondrial succinate dehy-
drogenase genes during angiosperm evolution. Genetics, 158, 1289–1300.
Araujo, W.L., Nunes-Nesi, A., Osorio, S. et al. (2011) Antisense inhibition of
the iron-sulphur subunit of succinate dehydrogenase enhances photo-
synthesis and growth in tomato via an organic acid-mediated effect on
stomatal aperture. Plant Cell, 23, 600–627.Baerenfaller, K., Grossmann, J., Grobei, M.A., Hull, R., Hirsch-Hoffmann, M.,
Yalovsky, S., Zimmermann, P., Grossniklaus, U., Gruissem, W. and Ba-
ginsky, S. (2008) Genome-scale proteomics reveals Arabidopsis thaliana
gene models and proteome dynamics. Science, 320, 938–941.
© 2012 The AuthorsThe Plant Journal © 2012 Blackwell Publishing Ltd, The Plant Journal, (2013), 73, 429–441
SDHAF2 in complex II function in Arabidopsis 439

Bafunno, V., Giancaspero, T.A., Brizio, C., Bufano, D., Passarella, S., Boles,
E. and Barile, M. (2004) Riboflavin uptake and FAD synthesis in saccharo-
myces cerevisiae mitochondria. J. Biol. Chem. 279, 95–102.Baysal, B., Willett-Brozick, J., Lawrence, E. et al. (2002) Prevalence of SDHB,
SDHC, and SDHD germline mutations in clinic patients with head and
neck paragangliomas. J. Med. Genet., 39, 178–183.Burger, C., Lang, F., Reith, M. and Gray, W. (1996) Genes encoding the same
three subunits of respiratory complex II are present in the mitochondrial
DNA of two phylogenetically distinct eukarytoes. Proc. Natl Acad. Sci.
USA, 93, 2328–2332.Burnichon, N., Briere, J.J., Libe, R. et al. (2010) SDHA is a tumor suppressor
gene causing paraganglioma. Hum. Mol. Genet., 19, 3011–3020.Carroll, A.J., Badger, M.R. and Millar, A.H. (2010) The MetabolomeExpress
Project: enabling web-based processing, analysis and transparent
dissemination of GC/MS metabolomics datasets. BMC Bioinformatics,
11, 376.
Curtis, M.D. and Grossniklaus, U. (2003) A gateway cloning vector set for
high-throughput functional analysis of genes in planta. Plant Physiol.,
133, 462–469.Daignan-Fornier, B., Valens, M., Lemire, B. and Bolotin-Fukuhara., M. (1994)
Structure and regulation of SDH3, the yeast gene encoding the cyto-
chrome b560 subunit of respiratory complex II. J. Biol. Chem., 269,
15469–15472.Elorza, A., LeA³n, G., GA³mez, I., Mouras, A., Holuigue, L., Araya, A. and Jor-
dana, X. (2004) Nuclear SDH2-1 and SDH2-2 genes, encoding the iron-
sulfur subunit of mitochondrial complex II in Arabidopsis, have distinct
cell-specific expression patterns and promoter activities. Plant Physiol.
136, 4072–4087.Elorza, A., Roschzttardtz, H., Gomez, I., Mouras, A., Holuigue, L., Araya, A.
and Jordana, X. (2006) A nuclear gene for the iron-sulfur subunit of mito-
chondrial complex II is specially expressed during Arabidopsis seed
development and germination. Plant Cell Physiol., 47, 14–21.Eubel, H., Jansch, L. and Braun, H.-P. (2003) New insights into the respira-
tory chain of plant mitochondria Supercomplexes and a unique composi-
tion of Complex II. Plant Physiol., 133, 274–286.Eubel, H., Braun, H.P. and Millar, A.H. (2005) Blue-native PAGE in
plants: a tool in analysis of protein-protein interactions. Plant Meth-
ods, 1, 1–11.Fuentes, D., Meneses, M., Nunes-Nesi, A., Araujo, W.L., Tapia, R., Gomez, I.,
Holuigue, L., Gutierrez, R.A., Fernie, A.R. and Jordana, X. (2011) A defi-
ciency in the flavoprotein of Arabidopsis mitochondrial complex II
results in elevated photosynthesis and better growth in nitrogen-limiting
conditions. Plant Physiol., 157, 1114–1127.Ghezzi, D., Goffrini, P., Uziel, G. et al. (2009) SDHAF1, encoding a LYR com-
plex-II specific assembly factor, is mutated in SDH-defective infantile leu-
koencephalopathy. Nat. Genet., 41, 654–656.Gleason, C., Huang, S., Thatcher, L.F., Foley, R.C., Anderson, C.R., Carroll,
A.J., Millar, A.H. and Singh, K.B. (2011) Mitochondrial complex II has a
key role in mitochondrial-derived reactive oxygen species influence on
plant stress gene regulation and defense. Proc. Natl Acad. Sci. USA, 108,
10768–10773.Hao, H.-X., Khalimonchuk, O., Schraders, M. et al. (2009) SDH5, a gene
required for flavination of succinate dehydrogenase, is mutated in para-
ganglioma. Science, 325, 1139–1142.Heazlewood, J.L., Tonti-Filippini, J.S., Gout, A.M., Day, D.A., Whelan, J. and
Millar, A.H. (2004) Experimental analysis of the Arabidopsis mitochon-
drial proteome highlights signaling and regulatory components, pro-
vides assessment of targeting prediction programs, and Indicates plant-
specific mitochondrial proteins. Plant Cell, 16, 241–256.Heazlewood, J.L., Verboom, R.E., Tonti-Filippini, J., Small, I. and Millar, A.
H. (2007) SUBA: the Arabidopsis subcellular database. Nuc. Acids Res,
35, D213–D218.Huang, S., Taylor, N.L., Whelan, J. and Millar, A.H. (2009a) Refining the defi-
nition of plant mitochondrial presequences through analysis of sorting
signals, N-terminal modifications, and cleavage motifs. Plant Physiol.,
150, 1272–1285.Huang, S., Taylor, N.L., Narsai, R., Eubel, H., Whelan, J. and Millar, A.H.
(2009b) Experimental analysis of the rice mitochondrial proteome, its
biogenesis and heterogeneity. Plant Physiol., 149, 719–734.Huang, S., Taylor, N.L., Narsai, R., Eubel, H., Whelan, J. and Millar, A.H.
(2010) Functional and composition differences between mitochondrial
complex II in Arabidopsis and rice are correlated with the complex genetic
history of the enzyme. Plant Mol. Biol., 72, 331–342.Kim, H.J., Khalimonchuk, O., Smith, P.M. and Winge, D.R. (2012) Structure,
function, and assembly of heme centers in mitochondrial respiratory
complexes. Biochim. Biophys. Acta, 1823, 1604–1616.Klodmann, J., Senkler, M., Rode, C. and Braun, H.-P. (2011) Defining the
protein complex proteome of plant mitochondria. Plant Physiol., 157,
587–598.Koprivova, A., des Francs-Small, C.C., Calder, G., Mugford, S.T., Tanz, S.,
Lee, B.-R., Zechmann, B., Small, I. and Kopriva, S. (2010) Identifica-
tion of a pentatricopeptide repeat protein implicated in splicing of
intron 1 of mitochondrial nad7 transcripts. J. Biol. Chem., 285, 32192–32199.
Kuhn, K., Richter, U., Meyer, E.H., Delannoy, E., Falcon de Longevialle,
A., O’Toole, N., Borner, T., Millar, A.H., Small, I.D. and Whelan, J.
(2009) Phage-type RNA polymerase RPOTmp performs gene-specific
transcription in mitochondria of Arabidopsis thaliana. Plant Cell, 21,
2762–2779.Lee, C.P., Eubel, H., O’Toole, N. and Millar, A.H. (2008) Heterogeneity of the
mitochondrial proteome for photosynthetic and non-photosynthetic Ara-
bidopsis metabolism. Mol. Cell. Proteomics, 7, 1297–1316.Lemire, B.L. and Oyedotun, K.S. (2002) The Saccharomyces cerevisiae mito-
chondrial succinate:ubiquinone oxidoreductase. Biochim. Biophys. Acta,
1553, 102–116.Leon, G., Holuigue, L. and Jordana, X. (2007) Mitochondrial complex II Is
essential for gametophyte development in Arabidopsis. Plant Physiol.,
143, 1534–1546.Lombardo, A., Carine, K. and Scheffler, I. (1990) Cloning and characteriza-
tion of the iron-sulfur subunit gene of succinate dehydrogenase from
Saccharomyces cerevisiae. J. Biol. Chem., 265, 10419–10423.de Longevialle, A.F., Meyer, E.H., Andres, C., Taylor, N.L., Lurin, C., Millar,
A.H. and Small, I.D. (2007) The pentatricopeptide repeat gene OTP43 Is
required for trans-splicing of the mitochondrial nad1 intron 1 in Arabid-
opsis thaliana. Plant Cell, 19, 3256–3265.van der Merwe, M.J., Osorio, S., Araujo, W.L., Balbo, I., Nunes-Nesi, A.,
Maximova, E., Carrari, F., Bunik, V.I., Persson, S. and Fernie, A.R. (2010)
Tricarboxylic acid cycle activity regulates tomato root growth via effects
on secondary cell wall production. Plant Physiol., 153, 611–621.Meyer, E.H., Tomaz, T., Carroll, A.J., Estavillo, G., Delannoy, E., Tanz, S.K.,
Small, I.D., Pogson, B.J. and Millar, A.H. (2009) Remodeled respiration in
ndufs4 with low phosphorylation efficiency suppresses Arabidopsis
germination and growth and alters control of metabolism at night. Plant
Physiol., 151, 603–619.Millar, H.A., Eubel, H., Jansch, L., Kruft, V., Heazlewood, J.L. and Braun, H.P.
(2004) Mitochondrial cytochrome c oxidase and succinate dehydrogenase
complexes contain plant specific subunits. Plant Mol. Biol., 56, 77–90.Mimaki, M., Wang, X., McKenzie, M., Thorburn, D.R. and Ryan, M.T. (2012)
Understanding mitochondrial complex I assembly in health and disease.
Biochim. Biophys. Acta, 1817, 851–862.Obayashi, T., Hayashi, S., Saeki, M., Ohta, H. and Kinoshita, K. (2009)
ATTED-II provides coexpressed gene networks for Arabidopsis. Nucleic
Acids Res., 37, D987–D991.Peters, K., Nießen, M., Peterhansel, C., Spath, B., Holzle, A., Binder, S.,
Marchfelder, A. and Braun, H.-P. (2012) Complex I–complex II ratio
strongly differs in various organs of Arabidopsis thaliana. Plant Mol.
Biol., 79, 273–284.Pracharoenwattana, I., Cornah, J.E. and Smith, S.M. (2005) Arabidopsis per-
oxisomal citrate synthase is required for fatty acid respiration and seed
germination. Plant Cell, 17, 2037–2048.Rigas, S., Daras, G., Laxa, M., Marathias, N., Fasseas, C., Sweetlove, L.J.
and Hatzopoulos, P. (2009) Role of Lon1 protease in post-germinative
growth and maintenance of mitochondrial function in Arabidopsis thali-
ana. New Phytol. 181, 588–600.Roschzttardtz, H., Fuentes, I., Vasquez, M., Corvalan, C., Leon, G., Gomez,
I., Araya, A., Holuigue, L., Vicente-Carbajosa, J. and Jordana, X. (2009) A
nuclear gene encoding the iron-sulfur subunit of mitochondrial complex
II is regulated by B3 domain transcription factors during seed develop-
ment in Arabidopsis. Plant Physiol., 150, 84–95.Rutter, J., Winge, D.R. and Schiffman, J.D. (2010) Succinate dehydroge-
nase–Assembly, regulation and role in human disease. Mitochondrion,
10, 393–401.
© 2012 The AuthorsThe Plant Journal © 2012 Blackwell Publishing Ltd, The Plant Journal, (2013), 73, 429–441
440 Shaobai Huang et al.

Schiavi, F., Boedeker, C.C., Bausch, B. et al. (2005) Predictors and preva-
lence of paraganglioma syndrome associated with mutations of the
SDHC gene. J. Amer. Med. Assoc., 294, 2057–2063.Shingaki-Wells, R.N., Huang, S., Taylor, N.L., Carroll, A.J., Zhou, W. and Mil-
lar, A.H. (2011) Differential molecular responses of rice and wheat co-
leoptiles to anoxia reveal novel metabolic adaptations in amino acid
metabolism for tissue tolerance. Plant Physiol., 156, 1706–1724.Stephen, A.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang, Z., Miller, W.
and Lipman, D.J. (1997) Gapped BLAST and PSI-BLAST: a new genera-
tion of protein database search programs. Nucleic Acids Res, 25, 3389–3402.
Sun, F., Huo, X., Zhai, Y., Wang, A., Xu, J., Su, D., Bartlam, M. and Rao, Z.
(2005) Crystal structure of mitochondrial respiratory membrane protein
Complex II. Cell, 121, 1043–1057.Taylor, N.L., Heazlewood, J.L. and Millar, A.H. (2011) The Arabidopsis thali-
ana 2-D gel mitochondrial proteome: refining the value of reference
maps for assessing protein abundance, contaminants and post-transla-
tional modifications. Proteomics, 11, 1720–1733.
Tcherkez, G., Mahe, A., Gauthier, P., Mauve, C., Gout, E., Bligny, R., Cornic, G.
and Hodges, M. (2009) In folio respiratory fluxomics revealed by 13C isotopic
labeling and H/D isotope effects highlight the noncyclic nature of the tricar-
boxylic acid “cycle” in illuminated leaves. Plant Physiol., 151, 620–630.Thomas, P.D., Campbell, M.J., Kejariwal, A., Mi, H., Karlak, B., Daverman,
R., Diemer, K., Muruganujan, A. and Narechania, A. (2003) PANTHER: a
library of protein families and subfamilies indexed by function. Genome
Res., 13, 2129–2141.Thomas, P.D., Kejariwal, A., Guo, N., Mi, H., Campbell, M.J., Muruganujan,
A. and Lazareva-Ulitsky, B. (2006) Applications for protein sequence-
function evolution data: mRNA/protein expression analysis and coding
SNP scoring tools. Nuc. Acids Res., 34, W645–W650.
Timmers, H.J.L.M., Gimenez-Roqueplo, A.-P., Mannelli, M. and Pacak, K.
(2009) Clinical aspects of SDHx-related pheochromocytoma and paragan-
glioma. Endocr. Relat. Cancer, 16, 391–400.Van Aken, O., Pecenkova, T., Van DeCotte, B. et al. (2007) Mitochondrial
type-I prohibitins of Arabidopsis thaliana are required for supporting pro-
ficient meristem development. Plant J., 52, 850–864.
© 2012 The AuthorsThe Plant Journal © 2012 Blackwell Publishing Ltd, The Plant Journal, (2013), 73, 429–441
SDHAF2 in complex II function in Arabidopsis 441
Copyright © 2022 FDOKUMEN




















