
Embryo production and quality of Holstein heifers and cows supplemented with β-carotene and...
14
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/6346597 Embryo production and quality of Holstein heifers and cows supplemented with beta-carotene and tocopherol Article in Animal Reproduction Science · July 2008 DOI: 10.1016/j.anireprosci.2007.04.001 · Source: PubMed CITATIONS 28 READS 142 5 authors, including: Lilian Dias University of São Paulo 8 PUBLICATIONS 29 CITATIONS SEE PROFILE Ana Viveiros Universidade Federal de Lavr… 50 PUBLICATIONS 819 CITATIONS SEE PROFILE All content following this page was uploaded by Ana Viveiros on 02 December 2016. The user has requested enhancement of the downloaded file.
Transcript of Embryo production and quality of Holstein heifers and cows supplemented with β-carotene and...
Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/6346597
EmbryoproductionandqualityofHolsteinheifersandcowssupplementedwithbeta-caroteneandtocopherol
ArticleinAnimalReproductionScience·July2008
DOI:10.1016/j.anireprosci.2007.04.001·Source:PubMed
CITATIONS
28
READS
142
5authors,including:
LilianDias
UniversityofSãoPaulo
8PUBLICATIONS29CITATIONS
SEEPROFILE
AnaViveiros
UniversidadeFederaldeLavr…
50PUBLICATIONS819CITATIONS
SEEPROFILE
AllcontentfollowingthispagewasuploadedbyAnaViveiroson02December2016.
Theuserhasrequestedenhancementofthedownloadedfile.

Please cite this article in press as: Sales, J.N.S. et al., Embryo production and quality of Hol-stein heifers and cows supplemented with �-carotene and tocopherol, Anim. Reprod. Sci. (2007),doi:10.1016/j.anireprosci.2007.04.001
ARTICLE IN PRESS+ModelANIREP-3346; No. of Pages 13
Animal Reproduction Science xxx (2007) xxx–xxx
Embryo production and quality of Holsteinheifers and cows supplemented with
�-carotene and tocopherol
J.N.S. Sales ∗, L.M.K. Dias, A.T.M. Viveiros,M.N. Pereira, J.C. Souza
Department of Animal Science, Universidade Federal de Lavras, Lavras-MG,Postal Code: 37200-000, Post-Box: 37, Brazil
Received 10 December 2006; accepted 3 April 2007
Abstract
The hypothesis was that the intramuscular injection (i.m.) of �-carotene associated to tocopherol improvescow (n = 86) and heifer (n = 91) embryo production and quality. Time of estrus was synchronized in animalswith an ear implant with 3 mg of norgestomet associated with an i.m. injection of 6 mg of norgestomet and10 mg of estradiol valerate (CRESTAR®, Intervert International B.V., Boxmeer, Holland) and superovulatedby 8 i.m. FSH/LHp injections (400 IU-heifers and 500 IU-cows) in decreasing concentrations at 12 h inter-vals. Animals were inseminated 12 and 24 h after observed onset of estrus and embryos recovered 7 dayslater. Animals were randomly allocated to one of three treatments: (1) vegetable oil vehicle (control), (2)800 mg of �-carotene and 500 mg of tocopherol (T800) and (3) 1200 mg of �-carotene and 750 mg of toco-pherol (T1200). Supplemental injections were given at the day norgestomet implants were inserted and atfirst superovulatory injection. An index (Embryo Quality Index or EQI) was proposed to more precisely eval-uate embryo quality (excellent*1 + good*2 + regular*3 + poor*4 + degenerate*5 + unfertilized ova*5)/total.There was an interaction between physiological stage (heifer or cow) and treatment on EQI (P = 0.01) andon the proportion of viable embryos (P = 0.03), where both variables were improved in T1200 cows, but notin heifers. The average EQI for heifers and cows in control, T800 and T1200 were 2.6 ± 0.3 and 3.6 ± 0.3;2.5 ± 0.3 and 3.6 ± 0.3; 2.9 ± 0.3 and 2.7 ± 0.3, respectively. The average total number of viable embryoswas greater (P = 0.01) in supplemented cows (3.5 ± 1.1; 5.4 ± 1.4 and 7.5 ± 1.2 in control, T800 and T1200,respectively), but less (P = 0.01) in heifers (7.5 ± 1.2; 5.6 ± 1.2 and 4.0 ± 1.1 in control, T800 and T1200,respectively). Supplementation injections of �-carotene associated to tocopherol improved embryo qualityin superovulated Holstein cows, in the present experimental conditions and may be advantageous in similar
∗ Corresponding author. Tel.: +55 35 3829 1686; fax: +55 35 3829 1231.E-mail addresses: [email protected], [email protected] (J.N.S. Sales).
0378-4320/$ – see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.anireprosci.2007.04.001

Please cite this article in press as: Sales, J.N.S. et al., Embryo production and quality of Hol-stein heifers and cows supplemented with �-carotene and tocopherol, Anim. Reprod. Sci. (2007),doi:10.1016/j.anireprosci.2007.04.001
ARTICLE IN PRESS+ModelANIREP-3346; No. of Pages 13
2 J.N.S. Sales et al. / Animal Reproduction Science xxx (2007) xxx–xxx
embryo production systems. However, at dosages applied in the present experiment, this treatment shouldnot be recommended for nuliparous heifers.© 2007 Elsevier B.V. All rights reserved.
Keywords: Cattle; �-carotene; Tocopherol; Embryo transfer; Superovulation
1. Introduction
The compound, �-carotene, is a lipid soluble isoprenoid pigment naturally produced by plantsand photosynthetic organisms. This carotenoid is the natural vitamin A precursor in feed and isconverted to retinol (its storage form) in the liver and used by tissues as retinal (Bendich andOlson, 1989). Tocopherol is an organic lipid soluble and biologically active alcohol responsiblefor most of feed vitamin E activity (Bregelius and Traber, 1999; Metre and Callan, 2001). Bothvitamins are potent anti-oxidative agents, so that, any substance, present in lesser concentrationscompared to the oxidizable substrate delays or inhibits oxidation of this substrate in an efficientmanner (Sies and Stahl, 1995).
Beneficial effects of �-carotene in the reproduction has been reported for various species. Par-enteral administration of �-carotene in mice (Chew and Archer, 1983) and pigs (Brief and Chew,1985; Kostoglou et al., 2000) decreased embryonic mortality and increased litter size. In dairycows, supplemental dietary �-carotene decreased retained placenta and endometritis incidencecompared to no supplementation or only vitamin A supplementation (Michal et al., 1994).
The best described role of tocopherol in mammalian species refers to its antioxidative function,by cellular control of lipid peroxidation explained by effects on inflammatory responses, immunesystem, striated musculature and reproduction (Metre and Callan, 2001). Vitamin E treatmentaccelerates uterine involution in cows with metritis (Harrison et al., 1984). Vitamin E protects theearly embryo from the effects of ambient heat stress (Arechiga et al., 1994), a cytotoxic event thatis probably mediated by free radicals (Loven, 1988).
Embryo transfer is a biotechnique that allows embryo collection from a donor female andsubsequent transfer to recipients to carry a pregnancy to term (Reichenbach et al., 2002). Embryoquality is very important in embryo transfer programs, because it reflects directly upon pregnancyoutcome after embryo transfer (Abe et al., 2002). Various factors may affect embryo quality.Nutrition is very important but it is still not well studied or understood. Among nutritional factorsthat could affect embryo quality, antioxidants, such as, �-carotene and tocopherol show greatpotential. The injectable form used in the present study was used to ensure delivery and avoid theusual potency loss that occurs in feed due to humidity, light, heat and management errors, thus,targeted plasma concentrations would be reached and maintained (Herdt and Stowe, 1991).
In the present study �-carotene was studied along with tocopherol with the objective of assess-ing synergistic effects of the two substances, for the latter is also recognized as an excellent naturalantioxidative agent that protects carotenoids in the animal body (Weiss et al., 1994; Lal Thankurand Srivastava, 1996). This association improves reproductive efficiency in many animal species.However, any effect on embryo quality in superovulated cattle has yet to be assessed. These twoantioxidants were given to superovulated donor females in an attempt to decrease the oxidativeeffects in animals that are likely to be undergoing hormonal stress and are also producing a largenumber of corpora lutea, which, according to Haliloglu et al. (2002), have significant �-caroteneconcentrations which may be necessary for steroidogenesis. So, the rationale was that the extracorpora lutea may concentrate �-carotene and possibly tocopherol.

Please cite this article in press as: Sales, J.N.S. et al., Embryo production and quality of Hol-stein heifers and cows supplemented with �-carotene and tocopherol, Anim. Reprod. Sci. (2007),doi:10.1016/j.anireprosci.2007.04.001
ARTICLE IN PRESS+ModelANIREP-3346; No. of Pages 13
J.N.S. Sales et al. / Animal Reproduction Science xxx (2007) xxx–xxx 3
The objective of the present study was to evaluate the effect of an intramuscular injection of �-carotene combined with tocopherol on embryo production and quality of superovulated Holsteinheifers and cows. Additionally, a novel embryo grading system is proposed, offering an option tothe current IETS embryo classification system.
2. Material and methods
2.1. Donors, animal facilities and diets
The present experiment was conducted from October 2003 through September 2004, at theFazenda Vista Alegre III, in the county of Curvelo in the central-north region of the state ofMinas Gerais, at 633 meters of altitude, south latitude 18◦45′56′′ and west Greenwich longitude44◦27′27′′. Treatments for the different physiological stage (heifer or cow) were applied simultane-ously at various time periods, and a balanced proportion of physiological stage and treatment wasmaintained throughout the trial. Heifers (n = 91) and cows (n = 86) averaged (means ± S.D.) 21 ± 2and 71 ± 29 months of age, 363 ± 27 and 687 ± 128 kg, 3.5 ± 0.3 and 3.4 ± 0.4 of body condition(on a 1–5 scale; Wildman et al., 1982), respectively. Lactating cows averaged (means ± S.D.)113.1 ± 45.1 days in milk and 23.2 ± 2.7 kg daily milk production across all treatments (no statis-tical differences were found between treatments). Pastures were mainly Brachiaria brizantha andCynodon dactylon (tifton). Heifers were maintained in tifton paddocks and supplemented dailywith tifton hay and balanced concentrate (corn bran, soybean meal, dried citrus, lime, dicalciumphosphate, sodium chloride and mineral mix). As for cows, heifer diet was designed to meet100% of NRC requirements (NRC, 1989). Cows were kept in a free-stall system. Brachiariabrizantha small paddocks served as rest areas and cows received elephant grass silage (Pennise-tum purpureum), tifton hay and concentrate (same ingredients as the heifer concentrate added ofcottonseed).
2.2. Estrous synchronization, superovulation and artificial insemination
On Day 0 (day of implant administration), when donors were evaluated for reproductivesoundness by rectal palpation, subcutaneous ear implant inserted with 3 mg of norgestomet(CRESTAR®, Intervert International B.V., Boxmeer, Holland) was placed, plus an i.m. injec-tion of 6 mg of norgestomet, and 10 mg of estradiol valerate. On Day 5, superovulation wasinitiated. In the superovulation protocol 500 IU (cows) and 400 IU (heifers) of FSH (PLUSET®
I.F. Serono, Rome, Italy), were distributed in decreasing dosages of 100, 100, 75, 75, 50, 50, 25, 25IU (cows) and 80, 80, 60, 60, 40, 40, 20, 20 IU (heifers) during 4 days, at 12 h intervals. Luteolysiswas induced by a single i.m. injection of 500 �g of DL cloprostenol (SINCROSIN®, Vallee S.A.,Montes Claros, Brazil). The ear implants were removed on the seventh FSH injection (Day 8)and estrus was observed on Day 9. Animals in estrus were inseminated by a single experiencedtechnician 12 and 24 h after the onset of estrus. Animals were flushed on Day 16 (7 days afterestrus). Only uterine flushes yielding at least two ova/embryos were included in the statisticalanalysis to ensure that animals were actually superovulated.
2.3. Treatments
Donors (n = 177) were allocated to treatments randomly as they entered the chute to be evaluatedfor reproduction soundness on Day 0. The first i.m. supplementation was given on Day 0 and the

Please cite this article in press as: Sales, J.N.S. et al., Embryo production and quality of Hol-stein heifers and cows supplemented with �-carotene and tocopherol, Anim. Reprod. Sci. (2007),doi:10.1016/j.anireprosci.2007.04.001
ARTICLE IN PRESS+ModelANIREP-3346; No. of Pages 13
4 J.N.S. Sales et al. / Animal Reproduction Science xxx (2007) xxx–xxx
second on the first day of superovulation or Day 5. Treatments were as follows: (1) donors receiveda 10 mL i.m. injection of vegetable oil (control; n = 59, 29 cows and 30 heifers), (2) donors received800 mg of �-carotene and 500 mg of tocopherol (T800; n = 59, 28 cows and 31 heifers) and (3)donors received 1200 mg of �-carotene and 750 mg of tocopherol (T1200; n = 59, 29 cows and30 heifers).
2.4. Embryo transfer
Embryos were collected from donors seven days after the first AI by a non-surgical procedure(Wright, 1981) with 1000 mL of Dulbecco’s modified phosphate buffered saline (DMPBS-Nutricell®, Campinas, Brazil). Embryos were evaluated by consensus of two experiencedveterinarians according to Lindner and Wright (1983). The embryo grading system recommendedby IETS (International Embryo Transfer Society, 1999) may be influenced by a variable degreeof subjectiveness. However, a very comprehensive study, done by Hasler (2001), demonstratedclearly that the traditional grading system (Lindner and Wright, 1983) is efficient and is relateddirectly to embryo survival after transfer, more so than any other variable considered in theirstudy.
An index (Embryo Quality Index or EQI) was proposed to more precisely evaluate embryoquality (excellent embryos *1 + good*2 + regular*3 + poor*4 + degenerate*5 + unfertilizedova*5)/total embryos recovered. For example: if a uterine flush yielded four excellent, three goodand one degenerate the index would be (4 × 1 + 3 × 2 + 0 × 3 + 0 × 4 + 1 × 5)/8 = 1.87. Valuescloser to one indicate greater quality. The proportion of viable embryos (VIAVP), based on thesame classification, was defined as the sum of viable embryos (excellent + good + regular + poor),divided by the total number of ova/embryos.
2.5. Blood sampling and serum β-carotene analysis
Blood samples were collected by coccygeal vessel venipuncture in 12 donors (four fromeach treatment), for �-carotene serum concentration determination. Samples were collected fromeach animal on Days 0, 5 and 16. Samples were centrifuged (2000 × g) for 10 min and freezerstored in cryogenic tubes (Cryogenic vial-Corning®) at −21 ◦C. Concentration of �-carotene wasdetermined by high pressure liquid chromatography described elsewhere (Njeru et al., 1992).
2.6. Statistical analysis
The experimental design was a 2 by 3 factorial, where two physiological stages (cows orheifers) were compared across three treatments (control, T800 and T1200). The variables EQI,VIAVP, total ova/embryos and viable embryos, were analyzed by the GENMOD procedure ofSAS (SAS Technical Report -243, 1993) and, as determined by the UNIVARIATE procedure ofSAS (SAS, 1995), characterized as a Poisson distribution. Because the Poisson curve implies inover dispersion, the GENMOD procedure was applied with a scale parameter (pscale option) toaccount for it and adjust the model adequately. Least square means (generated by the GENMOD)were compared by orthogonal contrasts. Fertilization rate data were analyzed by chi-square withthe PROCFREG procedure of SAS. The following model was used:
Yij = μ + Pi + Tj + P × Tij + eij

Please cite this article in press as: Sales, J.N.S. et al., Embryo production and quality of Hol-stein heifers and cows supplemented with �-carotene and tocopherol, Anim. Reprod. Sci. (2007),doi:10.1016/j.anireprosci.2007.04.001
ARTICLE IN PRESS+ModelANIREP-3346; No. of Pages 13
J.N.S. Sales et al. / Animal Reproduction Science xxx (2007) xxx–xxx 5
where μ = constant associated to all observations; Pi = effect of physiological stage i (i = heiferand cow); Tj = effect of treatment j (j = control, T800 and T1200); P × Tij = interaction betweenphysiological stage and treatment; eij = experimental error associated to Yij, which, by hypothesis,has a Poisson distribution where the mean is equal the variance.
3. Results
Supplementary injections (800 mg or 1200 mg) on Days 0 and 5 increased �-carotene con-centrations from 3.4 ± 1.1 and 5.1 ± 1.1 �g/mL before injection (Day 0) to 9.6 ± 0.9 and10.9 ± 3.4 �g/mL (Day 16), respectively (Fig. 1). Similar findings were observed by Arechigaet al. (1998a,b), where �-carotene concentrations after three injections remained elevated for 19days after the first injection.
Of the 177 donors only 132 (65 cows and 67 heifers) were considered for analysis, based onthe production of at least two ova/embryos. Because of lack of response to the superovulationprotocol, 34 animals (5 heifers and 6 cows in control, 6 heifers and 8 cows in T800 and 4 heifersand 5 cows in T1200) were removed from the trial. An additional group (n = 11) were not usedbecause they were unsuitable for insemination, as they presented abnormal mucous discharge orbecause of problems such as, implant loss or death. None of these events were due to treatmenteffects as they were included in an initial broader model and withdrawn for the final analysis.
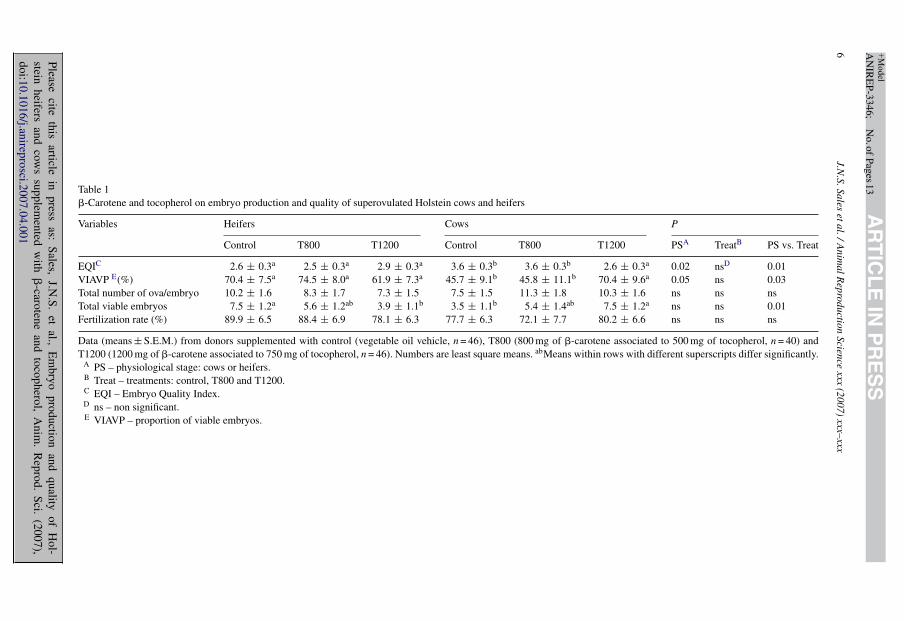
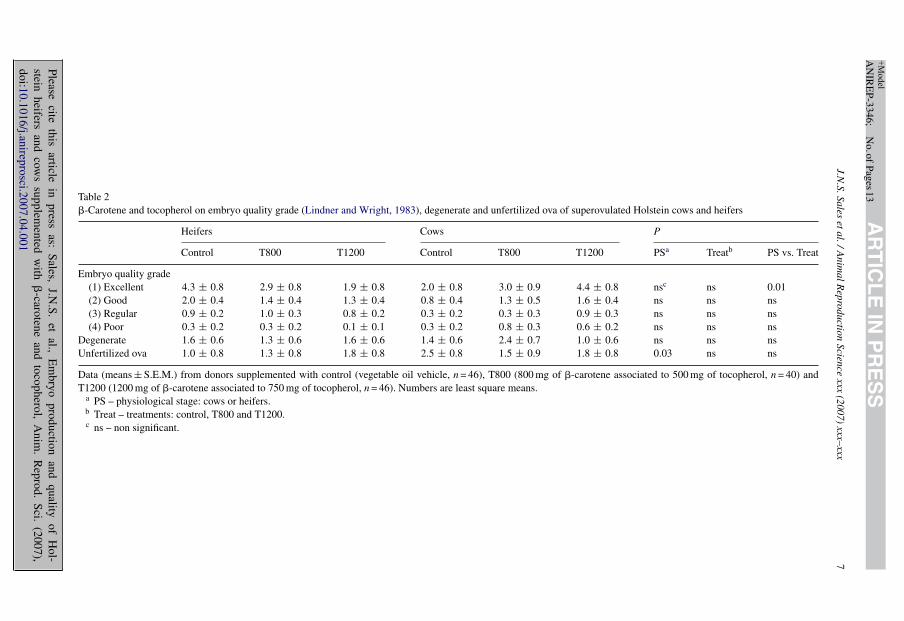
Treatments did not affect embryo quality index (Table 1). However, there was a physiologicalstage *treatment interaction effect on embryo quality (Table 1). Embryo quality improved in cowsthat received the greatest vitamin dose (T1200). In contrast, the T1200 dose decreased embryoquality in heifers, compared to control. Physiological stage affected embryo quality (Table 1)which was better in heifers than in cows. The embryo quality index scores may be comparedto the average numbers of embryo as graded in the traditional method (Table 2). Note that themain effect of physiological stage, which affected the EQI, was not detectable with the traditionalgrading system, except for unfertilized ova. The proportion of viable embryos (VIAVP) also wasnot affected by treatment but there were effects of physiological stage and of the interaction
Fig. 1. Mean �-carotene serum concentrations (determined by HPLC) in animals receiving i.m. injections of vegetableoil (�), 800 mg of �-carotene (�) and 1200 mg of �-carotene (�) on Days 0 and 5 of the experimental. Arrows representsupplementary injection days. The concentrations of �-carotene were different (P < 0.05) between all treatments on Day5 (*) and between control and the other treatments on Day 16 (**). Values were averaged between physiological stages(because values did not differ significantly, P > 0.05 between physiological stages). Only samples from four heifers andfive cows were used in this analysis.
Pleasecite
thisarticle
inpress
as:Sales,
J.N.S.
etal.,
Em
bryoproduction
andquality
ofH
ol-stein
heifersand
cows
supplemented
with
�-carotene
andtocopherol,
Anim
.R
eprod.Sci.
(2007),doi:10.1016/j.anireprosci.2007.04.001
AR
TIC
LE
IN P
RE
SS
+Model
AN
IRE
P-3346;N
o.ofPages13
6J.N
.S.Salesetal./A
nimalR
eproductionScience
xxx(2007)
xxx–xxx
Table 1�-Carotene and tocopherol on embryo production and quality of superovulated Holstein cows and heifers
Variables Heifers Cows P
Control T800 T1200 Control T800 T1200 PSA TreatB PS vs. Treat
EQIC 2.6 ± 0.3a 2.5 ± 0.3a 2.9 ± 0.3a 3.6 ± 0.3b 3.6 ± 0.3b 2.6 ± 0.3a 0.02 nsD 0.01VIAVP E(%) 70.4 ± 7.5a 74.5 ± 8.0a 61.9 ± 7.3a 45.7 ± 9.1b 45.8 ± 11.1b 70.4 ± 9.6a 0.05 ns 0.03Total number of ova/embryo 10.2 ± 1.6 8.3 ± 1.7 7.3 ± 1.5 7.5 ± 1.5 11.3 ± 1.8 10.3 ± 1.6 ns ns nsTotal viable embryos 7.5 ± 1.2a 5.6 ± 1.2ab 3.9 ± 1.1b 3.5 ± 1.1b 5.4 ± 1.4ab 7.5 ± 1.2a ns ns 0.01Fertilization rate (%) 89.9 ± 6.5 88.4 ± 6.9 78.1 ± 6.3 77.7 ± 6.3 72.1 ± 7.7 80.2 ± 6.6 ns ns ns
Data (means ± S.E.M.) from donors supplemented with control (vegetable oil vehicle, n = 46), T800 (800 mg of �-carotene associated to 500 mg of tocopherol, n = 40) andT1200 (1200 mg of �-carotene associated to 750 mg of tocopherol, n = 46). Numbers are least square means. abMeans within rows with different superscripts differ significantly.
A PS – physiological stage: cows or heifers.B Treat – treatments: control, T800 and T1200.C EQI – Embryo Quality Index.D ns – non significant.E VIAVP – proportion of viable embryos.
Pleasecite
thisarticle
inpress
as:Sales,
J.N.S.
etal.,
Em
bryoproduction
andquality
ofH
ol-stein
heifersand
cows
supplemented
with
�-carotene
andtocopherol,
Anim
.R
eprod.Sci.
(2007),doi:10.1016/j.anireprosci.2007.04.001
AR
TIC
LE
IN P
RE
SS
+Model
AN
IRE
P-3346;N
o.ofPages13
J.N.S.Sales
etal./Anim
alReproduction
Sciencexxx
(2007)xxx–xxx
7
Table 2�-Carotene and tocopherol on embryo quality grade (Lindner and Wright, 1983), degenerate and unfertilized ova of superovulated Holstein cows and heifers
Heifers Cows P
Control T800 T1200 Control T800 T1200 PSa Treatb PS vs. Treat
Embryo quality grade(1) Excellent 4.3 ± 0.8 2.9 ± 0.8 1.9 ± 0.8 2.0 ± 0.8 3.0 ± 0.9 4.4 ± 0.8 nsc ns 0.01(2) Good 2.0 ± 0.4 1.4 ± 0.4 1.3 ± 0.4 0.8 ± 0.4 1.3 ± 0.5 1.6 ± 0.4 ns ns ns(3) Regular 0.9 ± 0.2 1.0 ± 0.3 0.8 ± 0.2 0.3 ± 0.2 0.3 ± 0.3 0.9 ± 0.3 ns ns ns(4) Poor 0.3 ± 0.2 0.3 ± 0.2 0.1 ± 0.1 0.3 ± 0.2 0.8 ± 0.3 0.6 ± 0.2 ns ns ns
Degenerate 1.6 ± 0.6 1.3 ± 0.6 1.6 ± 0.6 1.4 ± 0.6 2.4 ± 0.7 1.0 ± 0.6 ns ns nsUnfertilized ova 1.0 ± 0.8 1.3 ± 0.8 1.8 ± 0.8 2.5 ± 0.8 1.5 ± 0.9 1.8 ± 0.8 0.03 ns ns
Data (means ± S.E.M.) from donors supplemented with control (vegetable oil vehicle, n = 46), T800 (800 mg of �-carotene associated to 500 mg of tocopherol, n = 40) andT1200 (1200 mg of �-carotene associated to 750 mg of tocopherol, n = 46). Numbers are least square means.
a PS – physiological stage: cows or heifers.b Treat – treatments: control, T800 and T1200.c ns – non significant.
Please cite this article in press as: Sales, J.N.S. et al., Embryo production and quality of Hol-stein heifers and cows supplemented with �-carotene and tocopherol, Anim. Reprod. Sci. (2007),doi:10.1016/j.anireprosci.2007.04.001
ARTICLE IN PRESS+ModelANIREP-3346; No. of Pages 13
8 J.N.S. Sales et al. / Animal Reproduction Science xxx (2007) xxx–xxx
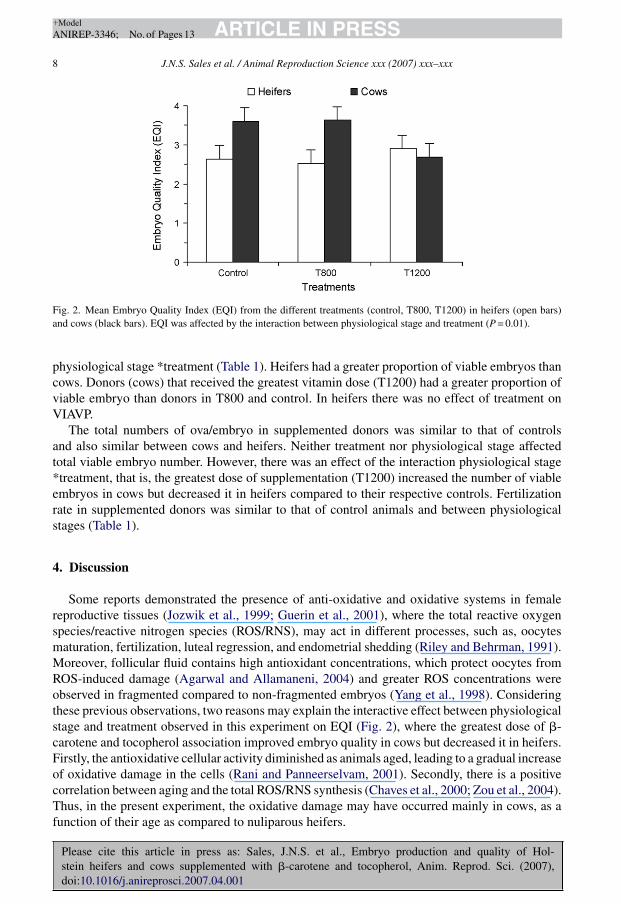
Fig. 2. Mean Embryo Quality Index (EQI) from the different treatments (control, T800, T1200) in heifers (open bars)and cows (black bars). EQI was affected by the interaction between physiological stage and treatment (P = 0.01).
physiological stage *treatment (Table 1). Heifers had a greater proportion of viable embryos thancows. Donors (cows) that received the greatest vitamin dose (T1200) had a greater proportion ofviable embryo than donors in T800 and control. In heifers there was no effect of treatment onVIAVP.
The total numbers of ova/embryo in supplemented donors was similar to that of controlsand also similar between cows and heifers. Neither treatment nor physiological stage affectedtotal viable embryo number. However, there was an effect of the interaction physiological stage*treatment, that is, the greatest dose of supplementation (T1200) increased the number of viableembryos in cows but decreased it in heifers compared to their respective controls. Fertilizationrate in supplemented donors was similar to that of control animals and between physiologicalstages (Table 1).
4. Discussion
Some reports demonstrated the presence of anti-oxidative and oxidative systems in femalereproductive tissues (Jozwik et al., 1999; Guerin et al., 2001), where the total reactive oxygenspecies/reactive nitrogen species (ROS/RNS), may act in different processes, such as, oocytesmaturation, fertilization, luteal regression, and endometrial shedding (Riley and Behrman, 1991).Moreover, follicular fluid contains high antioxidant concentrations, which protect oocytes fromROS-induced damage (Agarwal and Allamaneni, 2004) and greater ROS concentrations wereobserved in fragmented compared to non-fragmented embryos (Yang et al., 1998). Consideringthese previous observations, two reasons may explain the interactive effect between physiologicalstage and treatment observed in this experiment on EQI (Fig. 2), where the greatest dose of �-carotene and tocopherol association improved embryo quality in cows but decreased it in heifers.Firstly, the antioxidative cellular activity diminished as animals aged, leading to a gradual increaseof oxidative damage in the cells (Rani and Panneerselvam, 2001). Secondly, there is a positivecorrelation between aging and the total ROS/RNS synthesis (Chaves et al., 2000; Zou et al., 2004).Thus, in the present experiment, the oxidative damage may have occurred mainly in cows, as afunction of their age as compared to nuliparous heifers.

Please cite this article in press as: Sales, J.N.S. et al., Embryo production and quality of Hol-stein heifers and cows supplemented with �-carotene and tocopherol, Anim. Reprod. Sci. (2007),doi:10.1016/j.anireprosci.2007.04.001
ARTICLE IN PRESS+ModelANIREP-3346; No. of Pages 13
J.N.S. Sales et al. / Animal Reproduction Science xxx (2007) xxx–xxx 9
The main ROS/RNS targets may be the oocyte during ovarian follicular development, especiallyat the final stages of growth, and the embryo during its early development. These antioxidativeagents not only alter the majority of cell molecular types, but also are induced at an early stagethe blockage and retardation of embryonic development (Guerin et al., 2001). Elevated ROS/RNSconcentrations are toxic to the embryo and fetus apparently due to the fact that the free radicalsgenerated under such conditions are in excess compared to the developing embryo defense capacityagainst oxidation. This embryo vulnerability may lead to the production of highly reactive oxygenspecies, inducing oxidative stress and cell damage (Miki et al., 1988). These findings are consistentwith the notion that, as it may have occurred in the present study, supplementation of �-caroteneand tocopherol promoted equilibrium between oxidative agents and the antioxidative systemwhich may have improved embryo quality.
The beneficial effect of �-carotene on cow embryo quality observed in the present experiment,may also be explained by the: (1) increase in progesterone synthesis through the stimulatory roleof vitamin A on the cholesterol side chain cleavage enzyme (Ganguly et al., 1980; Weng et al.,2000); (2) increase in the intra-uterine secretion of important factors linked to embryonic initialdevelopment (Clawitter et al., 1990; Harney et al., 1990); (3) gap junction formation (Zhang etal., 1991, 1992; Frommel et al., 1994; Bertram and Bortkiewicz, 1995; Stahl et al., 1997) thatmay be important to coordinate luteal cell function (Redmer et al., 1991; Bilska et al., 1996).
Vitamin E in vitro protects embryos from ambient heat stress (Arechiga et al., 1994), a citotoxicevent that could be mediated by free radicals (Loven, 1988). This beneficial effect is explainedby the antioxidative properties of vitamin E which protects the spermatozoa and the oocytes fromattacks by oxidative agents that damage the early embryo. This also may have been a possiblereason for the improvement on embryo quality in cows in the present experiment.
There was an interactive effect between physiological stage and treatment for the total numberof viable embryos (Fig. 3), which was increased in cows by T1200, but decreased in heifers atthe same supplementation. Similarly, a greater number of viable embryos were found in donors(multiparous Nelore cows) that received vitamin A injection on the first day of the superovu-lation, compared to non-treated cows (Amaral, 2003). This finding concurs with the resultsobserved for cows in the present experiment may be justified by the effects of the establishmentof equilibrium between oxidative agents and the animal anti oxidative system, on steroidoge-
Fig. 3. Mean viable embryos within treatments (control, T800, T1200) in heifers (open bars) and cows (black bars). Meanembryo number was affected by the interaction between physiological stage and treatment (P = 0.01).

Please cite this article in press as: Sales, J.N.S. et al., Embryo production and quality of Hol-stein heifers and cows supplemented with �-carotene and tocopherol, Anim. Reprod. Sci. (2007),doi:10.1016/j.anireprosci.2007.04.001
ARTICLE IN PRESS+ModelANIREP-3346; No. of Pages 13
10 J.N.S. Sales et al. / Animal Reproduction Science xxx (2007) xxx–xxx
nesis, on gap junctions and on free radicals. In contrast, the reverse phenomenon observed inheifers in the present study was similar to the toxic effect was observed in vitro (Wang et al.,2002) when greater tocopherol dosages promoted embryo death in mice. The supplementaryinjections in heifers may have produced excessive vitamin concentrations causing an unbalancedratio between oxidative agents and the antioxidant defense system. According to Bedaiwy et al.(2002), embryonic development rate may be influenced by the ROS concentration in the femalegenital system. Additionally, in murine embryos the development rate was greater for embryoscultured in a lesser ROS concentration medium compared to embryos grown under a lesserROS conditioned medium. The lesser ROS concentration may actually represent physiologicalamounts and thus characterize a positive relationship between ROS concentrations and embryodevelopmental rate (Bedaiwy et al., 2002), where the ROS may alter gene expression duringdevelopment, affecting cytoskeleton formation, nuclear matrix proteins, chromatin configura-tion and the cellular ionic distribution (Allen, 1991). A substantial change on ROS productionmay significantly impact embryo development. Considering the described mechanisms it may beimplied that supplementation in heifers led to an excessive, non physiological, oxidative agentreduction in heifers which culminated in inadequate oocyte and embryo development. In practicethis deleterious effect by unknown mechanisms resulted in less viable embryos. It can be con-cluded that an appropriate balance must exist between the amount of oxidative agent producedand the capacity of the antioxidative defense mechanism to reduce it. This balance may also beobtained by exogenous antioxidants such as those from the diet or from parenteral supplementaryinjections.
The total numbers of ova/embryos were similar to those previously reported (Shaw et al., 1995;Sartori et al., 2004). The present results indicate that �-carotene and vitamin E did not act onthe number of recruited follicles from the ovulatory wave, neither on embryo yield, and weremost likely not involved in ovarian follicular development, at least in the way the experiment wasdesigned, which did not allow sufficient time for such effect to be evaluated.
Average fecundity rate (81.3%) was similar to that of one report (Dalton et al., 2000) butinferior (90.9%) to that of another (Sartori et al., 2004). Thus the artificial insemination processdid not interfere with fecundity in the present experiment. Additionally, it was observed that,the proposed treatments have most likely functioning to impact embryo viability and not eventsrelated to fertilization.
The novel embryo quality grading system or EQI provided consistent results as the traditionalgrading system. Unfortunately there were no similar data in the literature for comparisons, as EQIis unique at the present. Overall, the embryo grades assigned in the present experiment may be areliable indication, based on a large amount of evidence (Donaldson, 1985; Coleman et al., 1987;Hasler et al., 1987; Hasler, 2001; Spell et al., 2001), of ability to develop after implantation. Itremains to be tested whether this novel grading system reduces subjectivity or whether it mayexpress more precisely the donor status on its capacity to produce embryos compared to thetraditional methods.
5. Conclusion
The use of �-carotene combined with tocopherol may be an alternative to improve embryoquality in superovulated cows. However it is not recommended for heifers under similar conditionsof the present experiment. The use of the quality index or EQI will require more studies to beeffectively compared. Future studies must be directed to test the optimum dosage of vitamins aswell as their effect over time on embryo production.

Please cite this article in press as: Sales, J.N.S. et al., Embryo production and quality of Hol-stein heifers and cows supplemented with �-carotene and tocopherol, Anim. Reprod. Sci. (2007),doi:10.1016/j.anireprosci.2007.04.001
ARTICLE IN PRESS+ModelANIREP-3346; No. of Pages 13
J.N.S. Sales et al. / Animal Reproduction Science xxx (2007) xxx–xxx 11
Acknowledgements
We are thankful to Paulo Newton de Paiva Ferreira and Carlos Humberto Guimaraes for theirhelp which was essential for the completion of all the work. We are also grateful to Vallee S.A.,especially to Sergio Luiz Cirillo, who was able to finance the project, as the product develop-ment director of this genuinely Brazilian enterprise. We are especially thankful to CAPES whichprovided invaluable support in the form of a scholarship to the first author.
References
Abe, H., Matsuzaki, S., Hoshi, H., 2002. Ultrastructural differences in bovine morulae classified as high and low qualitiesby morphological evaluation. Theriogenology 57, 1273–1283.
Agarwal, A., Allamaneni, S.S.R., 2004. Oxidants and antioxidants in human fertility. Middle East Fertil. Soc. J. 9, 187–197.Allen, R.G., 1991. Oxygen-reactive species and antioxidant response during development. The metabolic paradox of
cellular differentiation. Soc. Exp. Biol. Med. 196, 117–129.Amaral, B.C., 2003. Utilizacao de vitamina A injetavel em diferentes concentracoes na producao e qualidade de embrioes
bovinos, Dissertation (Master in Animal Science) – Universidade Federal de Lavras, Lavras, 48 pp.Arechiga, C.G., Ealy, A.D., Hansen, P.J., 1994. Efficacy of vitamin E and glutathione for thermoprotection of murine
morulae. Theriogenology 41, 1545–1553.Arechiga, C.G., Staples, C.R., McDowell, L.R., Hansen, P.J., 1998a. Effects of time insemination and suplemental �-
carotene on reproduction and milk yield of dairy cows under heat stress. J. Dairy Sci. 81, 390–402.Arechiga, C.G., Flores, S.V., Ortiz, O., Ceron, J.H., Porras, A., Mcdowell, L.R., Hansen, P.J., 1998b. Effect of injection
of �-carotene or vitamin E and selenium on fertility of lactating dairy cows. Theriogenology 50, 65–76.Bedaiwy, M.A., Goldberg, J.M., Falcone, T., Singh, M., Nelson, D., Azab, H., 2002. Relationship between oxidative stress
and embryotoxicity of hydrosalpingeal fluid. Hum. Reprod. 17, 601–604.Bendich, A., Olson, J.A., 1989. Biological actions of carotenoids. FASEB J. 3, 1927–1932.Bertram, J.S., Bortkiewicz, H., 1995. Dietary carotenoids inhibit neoplastic transformation and modulate gene expression
in mouse and human cells. Am. J. Clin. Nutr. 62, 1327–1336.Bilska, A.T.G., Reynolds, L.P., Kirsch, J.D., Redmer, D.A., 1996. Gap junctional intercellular communication of bovine
luteal cells from several stages of the estrous cycle: effects of cyclic adenosine 3,5-monophosphate. Biol. Reprod. 54,538–545.
Bregelius, F.R., Traber, M.G., 1999. Vitamin E: function and metabolism. FASEB J. 13, 1145–1155.Brief, S., Chew, B.P., 1985. Effects of vitamin A and �-carotene on reproductive performance in gilts. J. Anim. Sci. 60,
998–1004.Chaves, M.M., Vieira, E.R., Reis, A.P., Silva, R.L., Gerzstein, N.C., Machado, J.A.N., 2000. Increase of reactive oxygen
(ROS) and nitrogen (RNS) species generated by phagocyting granulocytes related to age. Mech. Age. Dev. 119, 1–8.Chew, B.P., Archer, R.G., 1983. Comparative role of vitamin A and �-carotene on reproduction and neonate survival in
rats. Theriogenology 20, 459–472.Clawitter, J., Trout, W.E., Burke, M.G., Araghi, S., Roberts, R.M., 1990. A novel family of progesterone-induced, retinol-
binding proteins from uterine secretions of the pig. J. Biol. Chem. 265, 3248–3255.Coleman, D.A., Dailey, R.A., Leffel, R.E., Baker, R.D., 1987. Estrous synchronization and establishment of pregnancy
in bovine embryo transfer recipients. J. Dairy Sci. 70, 858–866.Dalton, J.C., Nadir, S., Bame, J.H., Noftsinger, M., Saacke, R.G., 2000. The effect of time of artificial insemination on
fertilization status and embryo quality in superovulated cows. J. Anim. Sci. 78, 2081–2085.Donaldson, L.E., 1985. Matching of embryo stages and grades with recipient oestrous synchrony in bovine embryo
transfer. Vet. Rec. 117, 489–491.Frommel, T.O., Lietz, H., Mobarhan, S., 1994. Expression of mRNA for the gap-junctional protein connexin 43 in human
colonic tissue is variable in response to �-carotene supplementation. Nutr. Cancer 22, 257–265.Ganguly, J., Rao, M.R.S., Murthy, S.K., Sarada, K., 1980. Systemic mode of action of vitamin A. Vitam. Horm. 38, 1–54.Guerin, P., El Mouatassim, S., Menezo, Y., 2001. Oxidative stress and protection against reactive oxygen species in the
pre-implantation embryo and its surroundings. Hum. Reprod. Update 7, 175–189.Haliloglu, S., Baspinar, N., Serpek, B., Erdem, H., Bulut, Z., 2002. Vitamin A and betacarotene levels in plasma, corpus
luteum and follicular fluid of cyclic and pregnant cattle. Reprod. Domestic. Anim. 37, 96–99.

Please cite this article in press as: Sales, J.N.S. et al., Embryo production and quality of Hol-stein heifers and cows supplemented with �-carotene and tocopherol, Anim. Reprod. Sci. (2007),doi:10.1016/j.anireprosci.2007.04.001
ARTICLE IN PRESS+ModelANIREP-3346; No. of Pages 13
12 J.N.S. Sales et al. / Animal Reproduction Science xxx (2007) xxx–xxx
Harney, J.P., Mirando, M.A., Smith, L.C., Bazer, F.W., 1990. Retinol-binding protein: a major secretory product of thepig conceptus. Biol. Reprod. 42, 523–532.
Harrison, J.H., Hancock, D.D., Conrad, H.R., 1984. Vitamin E and selenium for reproduction of the dairy cow. J. DairySci. 67, 123–132.
Hasler, J.F., 2001. Factors affecting frozen and fresh embryo transfer pregnancy rates in cattle. Theriogenology 56,1401–1415.
Hasler, J.F., McCauley, A.D., Lathrop, W.F., Foote, R.H., 1987. Effect of donor-embryo-recipient interactions on pregnancyrate in a large-scale bovine embryo transfer program. Theriogenology 27, 139–168.
Herdt, T.H., Stowe, H.D., 1991. Fat-soluble vitamin nutrition for dairy cattle. Vet. Clin. North Am.: Food Anim. Pract. 7,391–415.
International Embryo Transfer Society. 1999. Manual da Sociedade Internacional de Transferencia de Embrioes. Savoy,Illinois. 180 pp.
Jozwik, M., Wolczynski, S., Szamatowicz, M., 1999. Oxidative stress markers in preovulatory follicular fluid in humans.Mol. Hum. Reprod. 5, 409–413.
Kostoglou, P., Kyriakis, S.C., Papasteriadis, A., Roumpies, N., Alexopoulos, C., Saoulidis, K., 2000. Effect of �-caroteneon health status and performance of sows and their litters. J. Anim. Physiol. Anim. Nutr. 83, 150–157.
Lal Thankur, M., Srivastava, U.S., 1996. Vitamin-E metabolism and its application. Nutr. Res. 16, 1767–1809.Lindner, G.M., Wright, R.W., 1983. Bovine embryo morphology and evaluation. Theriogenology 20, 407–416.Loven, D.P., 1988. A role for reduce oxygen species in heat-induce cell killing and the induction thermotolerance. Med.
Hypotheses 26, 39–50.Metre, D.C.V., Callan, R.J., 2001. Selenium and vitamin. Vet. Clin. North Am.: Food Anim. Pract. 17, 373–402.Michal, J.J., Heirman, L.R., Wong, T.W., Chew, B.P., Frigg, M., Volker, L., 1994. Modulatory effects of dietary
beta-carotene on blood and mammary leukocyte function in periparturient dairy cows. J. Anim. Sci. 77, 1408–1421.
Miki, A., Fujimoto, E., Obsaki, T., Mizoquti, H., 1988. Effects of oxygen concentrations on embryonic development inrats using whole embryo culture. Anat. Embryol. 178, 337–339.
National Research Council, 1989. Nutrient Requirements of Dairy Cattle. sixth ed. (rev.). Nat. Acad. Sci., Washington,DC.
Njeru, C.A., MsDowell, L.R., Wilkinson, N.S., Linda, S.B., Williams, S.N., Lentz, E.L., 1992. Serum �-tocopherolconcentration in sheep after intramuscular injection of DL-�-tocopherol. J. Anim. Sci. 70, 2562–2567.
Rani, P.J.A., Panneerselvam, C., 2001. Carnitine as a free radical scavenger in aging. Exp. Geront. 36, 1713–1726.Redmer, D.A., Bilska, A.T.G., Reynolds, L.P., 1991. Contact-dependent intercellular communication of bovine luteal cells
in culture. Endocrinology 129, 2757–2766.Reichenbach, H.D., Oliveira, M.A.L., Lima, P.F., Santos Filho, A.S., Andrade, J.C., 2002. Transferencia e criopreservacao
de embrioes bovinos. In: Goncalves, P.B.D., Figueiredo, J.R., Freitas, V.J. (Eds.), Biotecnicas aplicadas a reproducaoanimal, Varela, Rio de Janeiro, pp. 127–178.
Riley, J.C., Behrman, H.R., 1991. Oxygen radicals and reactive oxygen species in reproduction. Proc. Soc. Exp. Biol.Med. 198, 781–791.
Sartori, R., Souza, A.H., Guenther, J.N., Caraviello, D.Z., Geiger, L.N., Schenk, J.L., Wiltbank, M.C., 2004. Fertilizationrate and embryo quality in superovulated Holstein heifers artificially inseminated with X-sorted or unsorted sperm.Anim. Reprod. 1, 86–90.
SAS Technical Report P-243, SAS/STAT software: The GENMOD Procedure, 1993. Release 6.09, SAS Institute Inc.,Cary, NC, 88 pp.
SAS SAS/STAT User’s Guide, 1995. Version 6.12, SAS Institute Inc., Cary, NC, 2:1686 pp.Shaw, D.W., Farin, P.W., Washburn, S.P., Britt, J.H., 1995. Effect of retinal palmitate on superovulation rate and embryo
quality in superovulated cattle. Theriogenology 44, 51–58.Sies, H., Stahl, W., 1995. Vitamins E and C, �-carotene, and other carotenoids as antioxidants. Am. J. Clin. Nutr. 62,
1315S–1321S.Spell, A.R., Beal, W.E., Corah, L.R., Lamb, G.C., 2001. Evaluating recipient and embryo factors that affect pregnancy
rates of embryo transfer in beef cattle. Theriogenology 56, 287–297.Stahl, W., Nicolai, S., Briviba, K., Hanusch, M., Broszeit, G., Peters, M., Martin, H.D., Sies, H., 1997. Biological activities
of natural and synthetic carotenoids: induction of gap junctional communication and singlet oxygen quenching.Carcinogenesis 18, 89–92.
Wang, X., Falcone, T., Attaran, M., Goldberg, J.M., Agarwal, A., Sharma, R.K., 2002. Vitamin C and vitamin E supple-mentation reduce oxidative stress-induced embryo toxicity and improve the blastocyst development rate. Fertil. Steril.78, 1272–1277.

Please cite this article in press as: Sales, J.N.S. et al., Embryo production and quality of Hol-stein heifers and cows supplemented with �-carotene and tocopherol, Anim. Reprod. Sci. (2007),doi:10.1016/j.anireprosci.2007.04.001
ARTICLE IN PRESS+ModelANIREP-3346; No. of Pages 13
J.N.S. Sales et al. / Animal Reproduction Science xxx (2007) xxx–xxx 13
Weiss, W.P., Hogan, J.S., Smith, K.L., Williams, S.N., 1994. Effect of dietary fat and vitamin E on �-tocopherol and�-carotene in blood of peripartum cows. J. Dairy Sci. 77, 1422–1429.
Weng, B.C., Chew, B.P., Wong, T.S., Park, J.S., Kim, H.W., Lepine, A.J., 2000. �-Carotene uptake and changes in ovariansteroids and uterine proteins during the estrous cycle in the canine. J. Anim. Sci. 78, 1284–1290.
Wildman, E.E., Jones, G.M., Wagner, P.E., Boman, R.L., Troutt Junior, H.F., Lesch, T.N., 1982. A dairy cow bodycondition scoring system and its relationship to selected production characteristics. J. Dairy Sci. 65, 495–501.
Wright, J.M., 1981. Non-surgical embryo transfer in cattle: embryo-recipient interactions. Theriogenology 15, 43–51.Yang, H.W., Hwang, K.J., Kwon, H.C., Kim, H.S., Choi, K.W., Oh, K.S., 1998. Detection of reactive oxygen species
(ROS) and apoptosis in human fragmented embryos. Hum. Reprod. 13, 998–1002.Zhang, L., Cooney, R.V., Bertram, J.S., 1991. Carotenoids enhance gap junctional communication and inhibit lipid
peroxidation in C3H/lOCONTROLE/2 cells: relationship to their cancer chemopreventive action. Carcinogenesis, 12,2109–2114.
Zhang, L., Cooney, R.V., Bertram, J.S., 1992. Carotenoids up-regulate connexin-43 gene expression independent of theirpro-vitamin A or antioxidant properties. Cancer. Res. 52, 5707–5712.
Zou, Y., Jung, K.J., Kim, J.W., Yu, B.P., Chung, H.Y., 2004. Alteration of soluble adhesion molecules during aging andtheir modulation by calorie restriction. FASEB J. 18, 320–322.




















