Protective Effect of Topical Application of α-Tocopherol and/or N-Acetyl Cysteine on Argemone...
11
This article was downloaded by: [Mukul Das] On: 17 May 2013, At: 09:58 Publisher: Routledge Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Nutrition and Cancer Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/hnuc20 Protective Effect of Topical Application of α-Tocopherol and/or N-Acetyl Cysteine on Argemone Oil/Alkaloid- Induced Skin Tumorigenesis in Mice Anu Pal a , Shamshad Alam a , Jaya Singhal a , Rahul Kumar a , Kausar M. Ansari a & Mukul Das a a Food, Drug and Chemical Toxicology Group , CSIR-Indian Institute of Toxicology Research , Uttar Pradesh , India To cite this article: Anu Pal , Shamshad Alam , Jaya Singhal , Rahul Kumar , Kausar M. Ansari & Mukul Das (2013): Protective Effect of Topical Application of α-Tocopherol and/or N-Acetyl Cysteine on Argemone Oil/Alkaloid-Induced Skin Tumorigenesis in Mice, Nutrition and Cancer, 65:sup1, 78-87 To link to this article: http://dx.doi.org/10.1080/01635581.2013.785005 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Protective Effect of Topical Application of α-Tocopherol and/or N-Acetyl Cysteine on Argemone...

This article was downloaded by: [Mukul Das]On: 17 May 2013, At: 09:58Publisher: RoutledgeInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Nutrition and CancerPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/hnuc20
Protective Effect of Topical Application of α-Tocopheroland/or N-Acetyl Cysteine on Argemone Oil/Alkaloid-Induced Skin Tumorigenesis in MiceAnu Pal a , Shamshad Alam a , Jaya Singhal a , Rahul Kumar a , Kausar M. Ansari a & Mukul Dasa
a Food, Drug and Chemical Toxicology Group , CSIR-Indian Institute of Toxicology Research ,Uttar Pradesh , India
To cite this article: Anu Pal , Shamshad Alam , Jaya Singhal , Rahul Kumar , Kausar M. Ansari & Mukul Das (2013): ProtectiveEffect of Topical Application of α-Tocopherol and/or N-Acetyl Cysteine on Argemone Oil/Alkaloid-Induced Skin Tumorigenesisin Mice, Nutrition and Cancer, 65:sup1, 78-87
To link to this article: http://dx.doi.org/10.1080/01635581.2013.785005
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form toanyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses shouldbe independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims,proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly inconnection with or arising out of the use of this material.

Nutrition and Cancer, 65(S1), 78–87Copyright C© 2013, Taylor & Francis Group, LLCISSN: 0163-5581 print / 1532-7914 onlineDOI: 10.1080/01635581.2013.785005
Protective Effect of Topical Application of �-Tocopheroland/or N-Acetyl Cysteine on ArgemoneOil/Alkaloid-Induced Skin Tumorigenesis in Mice
Anu Pal, Shamshad Alam, Jaya Singhal, Rahul Kumar, Kausar M. Ansari,and Mukul DasFood, Drug and Chemical Toxicology Group, CSIR-Indian Institute of Toxicology Research,Uttar Pradesh, India
Since bioantioxidants in plasma of Epidemic Dropsy patients[a condition caused by consumption of adulterated mustard oilwith argemone oil (AO)] were found to be significantly decreased,the beneficial effect of N-acetyl cysteine (NAC) and α-tocopherol(TOCO) against AO- or sanguinarine (SANG)-induced tumori-genicity was undertaken in mice. Topical application of TOCOand NAC either alone or in combination showed significant protec-tion against AO/TPA- and SANG/TPA-induced skin tumorigenic-ity. Histopathological findings suggest that papillomatous growthin AO/TPA- and SANG/TPA-treated animals were substantiallyprotected following topical application of TOCO or NAC. Further,treatment of TOCO and NAC either alone or in combination toAO/TPA- or SANG/TPA-induced mice significantly decreased lipidperoxidation, along with significant revival in glutathione (GSH)content and activities of tyrosinase, histidase, catalase, SOD, GSHperoxidase, and GSH reductase in skin. In vitro studies showed thatTOCO and/or NAC significantly decreased the AO and SANG in-duced cell proliferation and activation of ERK, p38, JNK MAPKsand NF-κB signaling in HaCaT cells. In summary, TOCO andNAC may be useful in preventing the tumorigenic response of AOand SANG probably by acting as scavenger of free radicals andinhibiting MAPKs and NF-κB signaling.
INTRODUCTIONAccidental or willful adulteration of edible oil with arge-
mone oil (AO) and consumption of such contaminated oil,give rise to a number of clinical symptoms, collectively re-ferred as Epidemic Dropsy (1). A number of epidemics havebeen reported in different cities of India and other countries in-cluding Bangladesh, Fiji Island, Madagascar, Mauritius, Nepal,
Submitted 16 October 2012; accepted in final form 18 November2012.
Address correspondence to Mukul Das, Food, Drug and Chemi-cal Toxicology Group, CSIR-Indian Institute of Toxicology Research,Mahatma Gandhi Marg, P.O. Box 80, Lucknow-226001, Uttar Pradesh,India. Fax: +91-522-2628227. E-mail: [email protected]
and South Africa (2). Systemic manifestations of dropsy syn-drome include normocytic anemia, hyper pigmentation of bodyparts, burning sensation of eyes, bilateral pitting edema oflower limbs, erythema, breathlessness, and hepatomegaly. Inextreme cases, glaucoma and even death due to cardiac andrespiratory failure have been observed (1,2). Our recent studieshave shown that AO and sanguinarine (SANG) alkaloid pos-sess genotoxic and carcinogenic potential and this has beenimplicated to DNA damage in animals as well as in dropsypatients, which might be the reason for the incidence of hepato-biliary/gall bladder cancers in the North and Eastern regions ofIndia (3–6). The toxicity of AO has been attributed to its phys-iologically active benzophenanthridine alkaloids, SANG anddihydrosanguinarine (7).
Our prior studies in experimental animals and clinical sub-jects suggest that reactive oxygen species (ROS) are involvedin the toxicity of AO (8,9). Our recent clinical studies in dropsypatients suggest that ROS are involved in the toxicity of AO bydiminishing the plasma total antioxidant capacity, α-tocopherol(TOCO) and glutathione (GSH) levels which in turn cause ox-idative damage to plasma proteins and lipids (9). In addition, itwas also observed that depleted plasma GSH may be due to di-rect utilization of GSH peroxidase to scavenge SANG-inducedfree radicals production in dropsy patients (10). In a preliminarystudy, we observed that some of the bioantioxidants includingriboflavin and TOCO offered protection to Epidemic Dropsypatients in a limited trial (11,12). Thus, it is quite likely thatbioantioxidants may have beneficial effects against AO- andSANG-induced tumorigenicity.
Based on these premise, we designed a study to evaluate theefficacy of two antioxidants viz N-acetyl cysteine (NAC) andTOCO on AO/TPA and SANG/TPA-induced skin tumorigenic-ity. NAC works extracellulary of itself and intracellularly as aprecursor of L-cysteine and reduced GSH through variety ofmechanisms including scavenging of ROS (13,14). Tocopheroland related compounds are considered to be potent cellular
78
Dow
nloa
ded
by [
Muk
ul D
as]
at 0
9:58
17
May
201
3

PROTECTION OF AO INDUCED TUMORIGENESIS BY ANTIOXIDANTS 79
antioxidants capable of trapping the ROS and terminating thefree radical chain reactions (15,16). Further, for understandingof molecular mechanism of protection by NAC and TOCO, sig-naling pathways were investigated in AO- and SANG-inducedHaCaT cells.
MATERIALS AND METHODS
Chemicals and AntibodiesDulbecco’s modified Eagle’s medium (DMEM), strepto-
mycin, penicillin, fetal bovine serum (FBS), trypsin/ethylene-diamine tetra acetic acid (EDTA) solution, proteaseinhibitor cocktail set-I, EDTA disodium salt, 2-amino-2-(hydr-oxymethyl)propane-1,3 diol (Tris), triton X-100, Tween-20,TOCO, bovine serum albumin (BSA), 5,5′-dithio-bis-nitrobenzoic acid (DTNB), dimethylbenzanthracene (DMBA),L-DOPA, epinephrine, flavin adenine dinucleotide (FAD),glutathione (GSH), glutathione disulfide (GSSH), glutaralde-hyde, L-histidine, nicotinamide adenine dinucleotide phosphate(NADP), nicotinamide adenine dinucleotide phosphate reduced(NADPH), and 12-O-tetradecanoyl phorbol myristate acetate(TPA) were purchased from Sigma Chemicals Co. (St. Louis,MO). Antibodies specific for p-ERK1/2, p-p38, p-JNK, p-NF-κB, ERK1/2, p38, JNK, α-tubulin, and β-Actin-HRP were pur-chased from Santa Cruz Biotechnology (Santa Cruz, CA). [3H]-Thymidine (specific activity: 79.2 Ci/mmol) was purchasedfrom Amersham (Arlington Heights, IL). All other chemicalsand reagents used were of highest purity commercially available.
Collection of Argemone Seeds, Extraction of Oil,and Preparation of Sanguinarine Alkaloid from AO
Argemone mexicana seeds were procured from outskirts ofLucknow city, Uttar Pradesh, India. The oil extraction, SANGalkaloid isolation, and their purification was done as describedpreviously (17).
Animal Treatment to Study the Preventive Effectof �-Tocopherol and/or N-Acetyl Cysteine on AOand Sanguinarine-Induced Tumorigenicity
Six- to 7-wk-old female Swiss albino mice (20± 2g) derivedfrom the animal breeding colony of CSIR-Indian Institute ofToxicology Research, Lucknow, India, were acclimatized understandard laboratory conditions and given a commercial pelletdiet (Provimi Animal Nutrition India Pvt. Ltd. Bangalore, In-dia) and water ad libitum. Animals were housed in plastic cageshaving rice husk as bedding and maintained in controlled atmo-sphere of 12-h dark/light, 22 ± 2◦C temperature and 50–60%humidity as per rules laid down by Animal Welfare Commit-tee of CSIR-Indian Institute of Toxicology Research. The micewere shaved with an electric clipper (Oster, Racine, WI) and hairwas removed with hair depilatory cream (Anne French, WeythLimited, Mumbai, India) 1 wk prior to the beginning of exper-iment. Mice were divided into 12 groups of 15 animals each.TOCO was dissolved in mustard oil and topically applied twice
weekly at the dose of 1 μmol, which was 40 times less than thatselected for single topical application (18). The dose of NACwas decided on the basis of the study conducted by D’Agostiniet al. (19) where NAC was given in drinking water to yield a cal-culated daily intake of 1.0 g/kg body wt to study the inhibitionof UV radiation-induced skin tumorigenicity. Since in our studytwice weekly topical application of NAC against AO/TPA orSANG/TPA induced skin tumorigenesis was studied, a 24 timeslower dose was chosen for the present investigation
The following treatment schedule was adopted:
• Group 1: Acetone (100 μl) topically applied twiceweekly.• Group 2: TOCO (1.0 μmol) topically applied twice
weekly.• Group 3: NAC (5.0 μmol) topically applied twice
weekly.• Group 4: TOCO (1.0 μmol) and NAC (5.0 μmol) was
topically applied twice weekly at a difference of 1 h.• Group 5: Single topical application of AO (0.3 ml)
followed by twice weekly application of TPA (4.0nmol/100 μl) after a week.• Group 6: Single topical application of SANG
(18.0 μmol/100 μl) followed by twice weekly appli-cation of TPA (4.0 nmol/100 μl) after a week.• Group 7: Single topical application AO of (0.3 ml)
followed by twice weekly application of TPA (4.0nmol/100 μl) and TOCO (1.0 μmol/100 μl) at an in-terval of 1 h.• Group 8: Single topical application AO of (0.3 ml)
followed by twice weekly application of TPA (4.0nmol/100 μl) and NAC (5.0 μmol/100 μl) at an in-terval of 1 h.• Group 9: Single topical application of SANG
(18.0 μmol/100 μl) followed by twice weekly ap-plication of TPA (4.0 nmol/100 μl) and TOCO(1.0 μmol/100 μl) at an interval of 1h.• Group 10: Single topical application of SANG
(18.0 μmol/100 μl) followed by twice weekly ap-plication of TPA (4.0 nmol/100 μl) and NAC(5.0 μmol/100 μl) at an interval of 1h.• Group 11: Single topical application AO of (0.3 ml)
followed by twice weekly application of TPA (4.0nmol/100 μl) and TOCO (1.0 μmol/100 μl) and NAC(5.0 μmol/100 μl) at an interval of 1 h.• Group 12: Single topical application of SANG
(18.0 μmol/100 μl) followed by twice weekly ap-plication of TPA (4.0 nmol/100 μl) and TOCO(1.0 μmol/100 μl) and NAC (5.0 μmol/100 μl) at aninterval of 1 h.
Skin tumor formation and body weight were recorded twiceweekly for 25 wk and tumors greater than 1 mm in diameterwere included in the cumulative total, only if they persistedfor 2 wk or more. Cumulative tumors were defined as the total
Dow
nloa
ded
by [
Muk
ul D
as]
at 0
9:58
17
May
201
3

80 A. PAL ET AL.
number of tumors observed on each mouse during its lifetime.The latent period was computed by the method of Shimkin andAndervont (20) as described earlier (21).
Histopathological ProcessingThe animals were sacrificed by cervical dislocation at the
termination of experiment. Skin was removed, washed in coldnormal saline, soak dried over filter paper and weighed. A por-tion of skin was fixed in 10% buffered formalin and embeddedin paraffin after processing. Sections of 5.0-μm thickness werecut and stained with hematoxylin and eosin for microscopicexamination.
Biochemical ParametersFat from the dermal side of skin was removed by scrapping
and 10% w/v skin homogenate was prepared using Ultra TurraxPolytron (Janke & Kunkel, Staufen, Germany). A portion ofhomogenate was centrifuged at 10,000 g for 20 min at 4◦C toisolate the postmitochondrial supernatant and stored at −80◦Cfor the biochemical assays. Glutathione (GSH) content was as-sayed in homogenate according to the method of Ellman (22).The rate of lipid peroxidation (LPO) was determined accordingto the method of Utley et al. (23) by estimating malondialdehyde(MDA) formed with 2-thiobarbituric acid (TBA). Catalase ac-tivity was determined by the method of Sinha (24). The methodis based on the reduction of dichromate to chromic acetate whenheated in the presence of hydrogen peroxide, with the forma-tion of perchoromic acid as an unstable intermediate, whichwas read at 570 nm. Catalase activity was expressed as μmolesH2O2 consumed/min/mg protein. GSH reductase was assayedaccording to the method of Carlberg and Mannervik (25). Theoxidation of NADPH was recorded spectrophotometrically at340 nm after 30 second intervals for 3 minutes. The activity wasexpressed as nmoles of NADPH oxidized per minute per mgprotein. GSH peroxidase was assayed according to the methodof Rotruck et al (26) and was expressed as nmoles GSH con-sumed per min per mg protein. Superoxide dismutase (SOD)estimation was based on the method of the ability of enzymeto inhibit the auto-oxidation of epinephrine at the pH 10.2 (27).Histidase activity was assayed according to the method of Taborand Mehler (28). Enzyme activity was calculated using a molarextinction coefficient of 18,800 M−1 cm−1 for urocanic acid andexpressed as nanomoles of urocanic acid formed per min permg protein. Tyrosinase activity was measured according to themethod of Horowitz et al. (29) and expressed as nanomoles ofsubstrate transformed per min per mg protein.
Cell CultureHuman keratinocytes (HaCaT) were a generous gift from
Prof. Susan M. Fischer, MD Anderson Cancer Centre,Smithville, TX, USA. These cells were derived from a spon-taneously immortalized human keratinocytes and are non-tumorigenic epidermal cell line that exhibits many of the mor-
phological and functional properties of normal human ker-atinocytes. HaCaT cells were cultured in DMEM supplementedwith 10% FBS, and 1% penicillin/streptomycin solution. Thecells were maintained under standard cell culture conditions at37◦C and 5% CO2 in a humid environment.
Cell ProliferationTo show the effect of TOCO and NAC individually or in
combination on cell proliferation, [3H]-thymidine incorporationwas performed in AO- and SANG-induced HaCaT cells. HaCaTcells were grown to about 80–85% confluence in 6-well platesand treated with 10 μM TOCO and 5 mM NAC alone or incombination in the presence or absence of AO (0.1 μl/ml) andSANG (8.0 μg/ml) for 20 h. Two hours prior to harvest, cellswere pulsed with 1 μCi per ml media of [3H]-thymidine. Thecultures were processed as previously described (30) and [3H]-thymidine incorporation in DNA was measured.
Preparation of Cell Extracts and Western Blot AnalysisThe confluent HaCaT cells were serum starved for 24 h to
establish quiescence, followed by 24 h of exposure with 10 μMTOCO and 5 mM NAC, individually or in combination. Sub-sequently, cells were stimulated with 0.25% DMSO, AO (0.1μl/ml) or SANG (8.0 μg/ml) for 1 h. After treatment schedule,cells washed with cold PBS and lysed with ice-cold RIPA buffersupplemented with protease inhibitors. Forty μg proteins weresubjected to 10% SDS-PAGE and transferred to a polyvinyli-dene difluoride (PVDF) membrane (Millipore, Billerica, MA)and incubated with specific primary antibodies at 4◦C overnightfollowed by incubation with HRP-conjugated secondary an-tibody (Sigma Chemical Co, St. Louis, MO). Bound antibodywas detected by enhanced chemiluminescence using ECL West-ern blotting detection kit following the instructions provided bymanufacturer (Amersham, Fairfield, CT). For the detection ofphosphorylated protein, separate SDS/PAGE run was performedand their respective total and actin/tubulin was performed bystripping and re-probing to ensure equal loading of protein.
Statistical AnalysisThe results were expressed as the mean± SE. The statistical
significance of difference between the values of control andtreatment groups was determined using Student’s t test. A Pvalue of <0.05 was considered statistically significant.
RESULTSThe effect of topical application of TOCO and/or NAC
on AO- and SANG-induced skin tumorigenesis is depicted inFig. 1. The first incidence of tumorgenesis in groups treatedwith AO/TPA and SANG/TPA occurred at 10th and 18th wk,respectively (Fig. 1a). The cumulative number of tumors inAO/TPA and SANG/TPA groups was 43 and 19, respectively,after 25 wk of exposure (Fig. 1a). Twice weekly topical appli-cation of TOCO after 1 h of TPA application to AO/TPA- and
Dow
nloa
ded
by [
Muk
ul D
as]
at 0
9:58
17
May
201
3

PROTECTION OF AO INDUCED TUMORIGENESIS BY ANTIOXIDANTS 81
0
5
10
15
20
25
30
35
40
45
1 4 7 10 13 16 19 22 25
Cu
mu
lati
ve n
um
ber
of
tum
ors
Weeks on Test
AO/TPA
SANG/TPA
AO/TPA+TOCO
AO/TPA+NAC
AO/TPA+TOCO+NAC
0
20
40
60
80
100
120
1 4 7 10 13 16 19 22 25
% o
f m
ice
wit
h t
um
ors
Weeks on Test
AO/TPA
SANG/TPA
AO/TPA+TOCO
AO/TPA+NAC
AO/TPA+TOCO+NAC
0
0.5
1
1.5
2
2.5
3
3.5
1 4 7 10 13 16 19 22 25
Tum
ors
per
tu
mo
r b
eari
ng
m
ou
se
Weeks on Test
AO/TPA
SANG/TPA
AO/TPA+TOCO
AO/TPA+NAC
AO/TPA+TOCO+NAC
0
0.5
1
1.5
2
2.5
3
3.5
1 4 7 10 13 16 19 22 25
Tum
ors
/mo
use
Weeks on Test
AO/TPA
SANG/TPA
AO/TPA+TOCO
AO/TPA+NAC
AO/TPA+TOCO+NAC
A B
D C
FIG. 1. Effect of α-tocopherol (TOCO) and N-acetyl cysteine (NAC) on argemone oil (AO)/12-O-tetradecanoyl phorbol myristate acetate (TPA) and sanguinarine(SANG)/TPA induced skin tumorigenesis in terms of (A) cumulative number of tumors; (B) percentage of mice with tumors; (C) tumors/tumor bearing mouse;and (D) tumor/mouse. Twice weekly topical application of acetone, TOCO (40 μmol) or NAC (5 μmol) to mice showed no signs of tumor formation. Similarly,no tumors were observed in SANG/TPA+TOCO, SANG/TPA+NAC, and SANG/TPA+TOCO+NAC treated groups.
SANG/TPA-treated animals resulted in significant decrease of93% and 100%, respectively, in cumulative number of tumorsafter 25 wk. Similarly twice weekly topical application of NACafter 1 h of TPA application to AO/TPA- and SANG/TPA-treatedanimals resulted in decrease of 63% and 100%, respectively, andcombination of TOCO and NAC showed pronounced decreaseof 98% and 100%, respectively. The percentage of mice withtumors during TOCO treatment to AO/TPA- and SANG/TPA-induced animals is shown in Fig. 1b. One hundred percent micetreated with AO/TPA showed tumors after 25 wk, whereas 47%of mice developed tumors following SANG/TPA treatment af-ter 25 wk. Furthermore, topical application of TOCO and NACindividually or in combination to AO/TPA-treated animals re-sulted in significant decrease (80%, 73%, and 94%, respectively)in percentage of mice with tumors (Fig. 1b).
The effect of TOCO and/or NAC on the number of tumorsper tumor bearing mouse in AO/TPA- and SANG/TPA-treatedgroups is shown in Fig. 1c. A total of 2.9 and 2.7 tumors per tu-mor bearing mouse were observed in AO/TPA- and SANG/TPA-treated groups after 25 wk. Treatment with TOCO to AO/TPA-and SANG/TPA-induced groups caused a significant decreaseof 66% and 100% in tumors per tumor bearing mouse, re-spectively (Fig. 1c). Similarly topical application of NAC alsocaused significant decrease of 49% and 100%, and combinationof TOCO and NAC showed 68% and 100% decrease in AO/TPAand SANG/TPA groups, respectively. The data when expressedas tumors per mouse showed 2.9 and 1.2 values for AO/TPAand SANG/TPA groups after 25 wk (Fig. 1d). Treatment with
TOCO to AO/TPA and SANG/TPA groups resulted in signifi-cant decrease of 93% and 100% in the number of tumors permouse after 25 wk. Similarly topical application of NAC alsocaused significant decrease of 86% and 100%, and combinationof TOCO and NAC showed 98% and 100% decrease in AO/TPAand SANG/TPA groups, respectively.
The effect of TOCO on histopathological changes in skin ofAO/TPA and SANG/TPA induced animals is elicited in Fig. 2.The skin of mice treated with twice weekly application of ace-tone and TOCO, NAC, or combination of TOCO and NAC re-vealed normal epidermis, dermis, and muscular layers (data notshown). The skin of AO/TPA- and SANG/TPA-treated animalsdemonstrated papillomatous growth comprising of proliferationof squamous cells, hyperkeratinization, and keratin pearl forma-tion, whereas dermis showed significant oedematous reactionalong with marked infiltration of mononuclear and fibroblas-tic activity (Fig. 2A and 2E). Treatment of TOCO and/or NACindividually or in combination to AO/TPA- and SANG/TPA-induced animals showed normal skin structure similar to that ofcontrol with mild edema in dermis (Figure 2B–2D and 2F–2H).
The effect of topical application of TOCO and/or NAC onhistidase and tyrosinase enzymes in AO/TPA and SANG/TPAtreated mice is tabulated in Table 1. AO/TPA and SANG/TPAtreatment showed significant reduction of dermal tyrosinase(29–35%) and histidase (29–30%) activities. Treatment ofTOCO to AO/TPA- and SANG/TPA-induced animals ledto significant increase in tyrosinase (32–44%) and histi-dase (26–27%) activities. Topical application of NAC to
Dow
nloa
ded
by [
Muk
ul D
as]
at 0
9:58
17
May
201
3

82 A. PAL ET AL.
FIG. 2. Photomicrograph of skin showing (A) Squamous cell carcinoma in argemone oil (AO)/12-O-tetradecanoyl phorbol myristate acetate (TPA) treated mice;(B) AO/TPA+α-tocopherol (TOCO) treated group; (C) AO/TPA+N-acetyl cysteine (NAC) treated group; (D) AO/TPA+TOCO+NAC treated group; (E) papillo-matous growth with proliferation of epidermal layers in sanguinarine (SANG)/TPA treated mice; (F) SANG/TPA+TOCO treated animal; (G) SANG/TPA+NACtreated group; and (H) SANG/TPA+TOCO+NAC treated group (H&E, magnification = ×50) (Color figure available online).
AO/TPA- and SANG/TPA-induced animals caused significantincrease in tyrosinase (27–39%) and histidase (24%) activi-ties. Combined treatment of TOCO and NAC to AO/TPA-and SANG/TPA-induced animals showed increase in tyrosinase(31–32%) and histidase (23–24%) activities (Table 1).
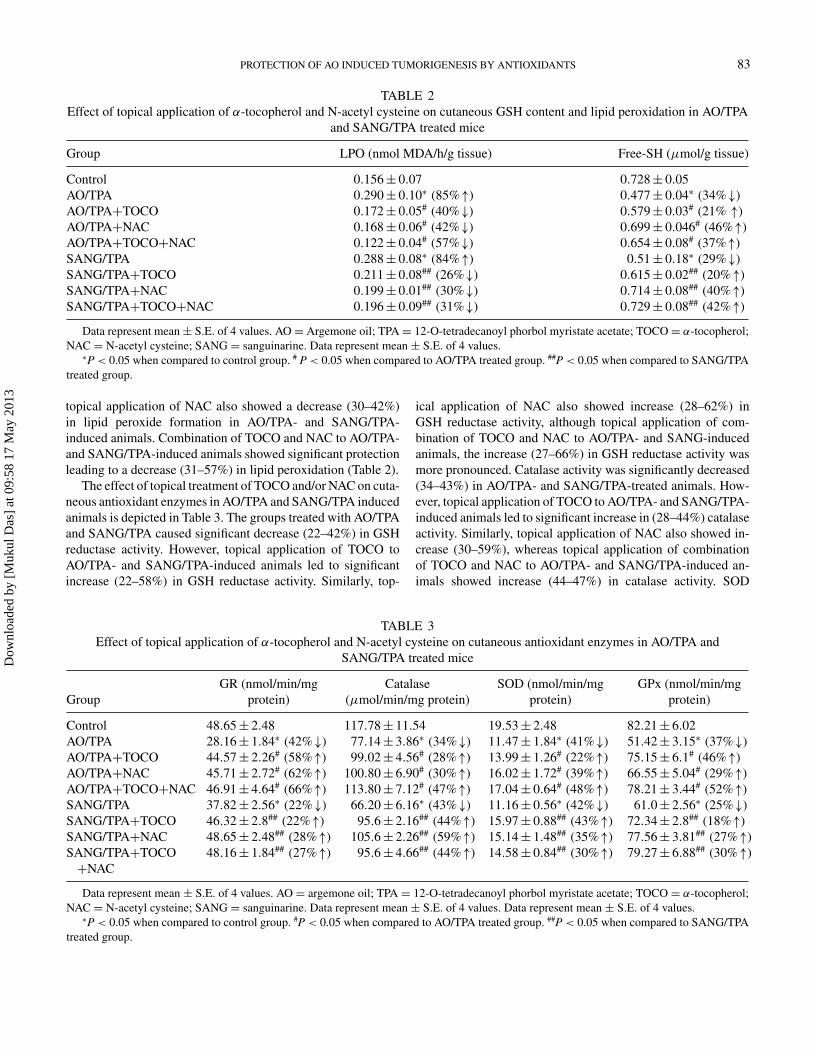
Effect of treatment of TOCO and/or NAC on cutaneous GSHcontent and LPO in AO/TPA and SANG/TPA induced ani-mals is shown in Table 2. The groups treated with AO/TPAand SANG/TPA caused significant decrease (29–34%) in GSHcontent. However, topical application of TOCO to AO/TPA-
and SANG/TPA-induced animals led to significant increase(20–21%) in GSH content. Similarly, topical application ofNAC also showed an increase (40–46%) in GSH content;however, topical application of combination of NAC andTOCO to AO/TPA- and SANG/TPA-induced animals causedincrease (37%–42%) in GSH content. On the contrary, LPOwas found to be substantially enhanced (84–85%) in AO/TPA-and SANG/TPA-treated groups. Topical application of TOCOto AO/TPA- and SANG/TPA-treated groups resulted in signifi-cant decrease (26–40%) in lipid peroxide formation. Similarly
TABLE 1Effect of topical application of α-tocopherol and N-acetyl cysteine on skin marker enzymes in AO/TPA and SANG/TPA treated
mice
Group Tyrosinase (nmol/min/mg protein) Histidase (nmol/min/mg protein)
Control 6.60± 0.70 194.00± 8.05AO/TPA 4.29± 0.20∗ (35%↓) 136.70± 4.04∗ (29%↓)AO/TPA+TOCO 6.18± 0.50# (44%↑) 172.90± 6.03# (26%↑)AO/TPA+NAC 6.00± 0.46# (39%↑) 169.90± 8.46# (24%↑)AO/TPA+TOCO+NAC 5.62± 0.44# (31%↑) 170.65± 9.08# (24%↑)SANG/TPA 4.68± 0.40∗ (29%↓) 134.30± 6.18∗ (30%↓)SANG/TPA+TOCO 6.21± 0.58## (32%↑) 171.50± 9.02## (27%↑)SANG/TPA+NAC 5.99± 0.51## (27%↑) 167.14± 8.08## (24%↑)SANG/TPA+TOCO+NAC 6.19± 0.47## (32%↑) 165.90± 9.08## (23%↑)
Data represent mean ± S.E. of 4 values. AO = Argemone oil; TPA = 12-O-tetradecanoyl phorbol myristate acetate; TOCO = α-tocopherol;NAC = N-acetyl cysteine; SANG = sanguinarine.∗P < 0.05 when compared to control group. #P < 0.05 when compared to AO/TPA treated group. ## P < 0.05 when compared to SANG/TPA
treated group.
Dow
nloa
ded
by [
Muk
ul D
as]
at 0
9:58
17
May
201
3
PROTECTION OF AO INDUCED TUMORIGENESIS BY ANTIOXIDANTS 83
TABLE 2Effect of topical application of α-tocopherol and N-acetyl cysteine on cutaneous GSH content and lipid peroxidation in AO/TPA
and SANG/TPA treated mice
Group LPO (nmol MDA/h/g tissue) Free-SH (μmol/g tissue)
Control 0.156± 0.07 0.728± 0.05AO/TPA 0.290± 0.10∗ (85%↑) 0.477± 0.04∗ (34%↓)AO/TPA+TOCO 0.172± 0.05# (40%↓) 0.579± 0.03# (21% ↑)AO/TPA+NAC 0.168± 0.06# (42%↓) 0.699± 0.046# (46%↑)AO/TPA+TOCO+NAC 0.122± 0.04# (57%↓) 0.654± 0.08# (37%↑)SANG/TPA 0.288± 0.08∗ (84%↑) 0.51± 0.18∗ (29%↓)SANG/TPA+TOCO 0.211± 0.08## (26%↓) 0.615± 0.02## (20%↑)SANG/TPA+NAC 0.199± 0.01## (30%↓) 0.714± 0.08## (40%↑)SANG/TPA+TOCO+NAC 0.196± 0.09## (31%↓) 0.729± 0.08## (42%↑)
Data represent mean ± S.E. of 4 values. AO = Argemone oil; TPA = 12-O-tetradecanoyl phorbol myristate acetate; TOCO= α-tocopherol;NAC = N-acetyl cysteine; SANG = sanguinarine. Data represent mean ± S.E. of 4 values.∗P < 0.05 when compared to control group. # P < 0.05 when compared to AO/TPA treated group. ##P < 0.05 when compared to SANG/TPA
treated group.
topical application of NAC also showed a decrease (30–42%)in lipid peroxide formation in AO/TPA- and SANG/TPA-induced animals. Combination of TOCO and NAC to AO/TPA-and SANG/TPA-induced animals showed significant protectionleading to a decrease (31–57%) in lipid peroxidation (Table 2).
The effect of topical treatment of TOCO and/or NAC on cuta-neous antioxidant enzymes in AO/TPA and SANG/TPA inducedanimals is depicted in Table 3. The groups treated with AO/TPAand SANG/TPA caused significant decrease (22–42%) in GSHreductase activity. However, topical application of TOCO toAO/TPA- and SANG/TPA-induced animals led to significantincrease (22–58%) in GSH reductase activity. Similarly, top-
ical application of NAC also showed increase (28–62%) inGSH reductase activity, although topical application of com-bination of TOCO and NAC to AO/TPA- and SANG-inducedanimals, the increase (27–66%) in GSH reductase activity wasmore pronounced. Catalase activity was significantly decreased(34–43%) in AO/TPA- and SANG/TPA-treated animals. How-ever, topical application of TOCO to AO/TPA- and SANG/TPA-induced animals led to significant increase in (28–44%) catalaseactivity. Similarly, topical application of NAC also showed in-crease (30–59%), whereas topical application of combinationof TOCO and NAC to AO/TPA- and SANG/TPA-induced an-imals showed increase (44–47%) in catalase activity. SOD
TABLE 3Effect of topical application of α-tocopherol and N-acetyl cysteine on cutaneous antioxidant enzymes in AO/TPA and
SANG/TPA treated mice
GroupGR (nmol/min/mg
protein)Catalase
(μmol/min/mg protein)SOD (nmol/min/mg
protein)GPx (nmol/min/mg
protein)
Control 48.65± 2.48 117.78± 11.54 19.53± 2.48 82.21± 6.02AO/TPA 28.16± 1.84∗ (42%↓) 77.14± 3.86∗ (34%↓) 11.47± 1.84∗ (41%↓) 51.42± 3.15∗ (37%↓)AO/TPA+TOCO 44.57± 2.26# (58%↑) 99.02± 4.56# (28%↑) 13.99± 1.26# (22%↑) 75.15± 6.1# (46%↑)AO/TPA+NAC 45.71± 2.72# (62%↑) 100.80± 6.90# (30%↑) 16.02± 1.72# (39%↑) 66.55± 5.04# (29%↑)AO/TPA+TOCO+NAC 46.91± 4.64# (66%↑) 113.80± 7.12# (47%↑) 17.04± 0.64# (48%↑) 78.21± 3.44# (52%↑)SANG/TPA 37.82± 2.56∗ (22%↓) 66.20± 6.16∗ (43%↓) 11.16± 0.56∗ (42%↓) 61.0± 2.56∗ (25%↓)SANG/TPA+TOCO 46.32± 2.8## (22%↑) 95.6± 2.16## (44%↑) 15.97± 0.88## (43%↑) 72.34± 2.8## (18%↑)SANG/TPA+NAC 48.65± 2.48## (28%↑) 105.6± 2.26## (59%↑) 15.14± 1.48## (35%↑) 77.56± 3.81## (27%↑)SANG/TPA+TOCO+NAC
48.16± 1.84## (27%↑) 95.6± 4.66## (44%↑) 14.58± 0.84## (30%↑) 79.27± 6.88## (30%↑)
Data represent mean ± S.E. of 4 values. AO = argemone oil; TPA = 12-O-tetradecanoyl phorbol myristate acetate; TOCO = α-tocopherol;NAC = N-acetyl cysteine; SANG = sanguinarine. Data represent mean ± S.E. of 4 values. Data represent mean ± S.E. of 4 values.∗P < 0.05 when compared to control group. #P < 0.05 when compared to AO/TPA treated group. ##P < 0.05 when compared to SANG/TPA
treated group.
Dow
nloa
ded
by [
Muk
ul D
as]
at 0
9:58
17
May
201
3
84 A. PAL ET AL.
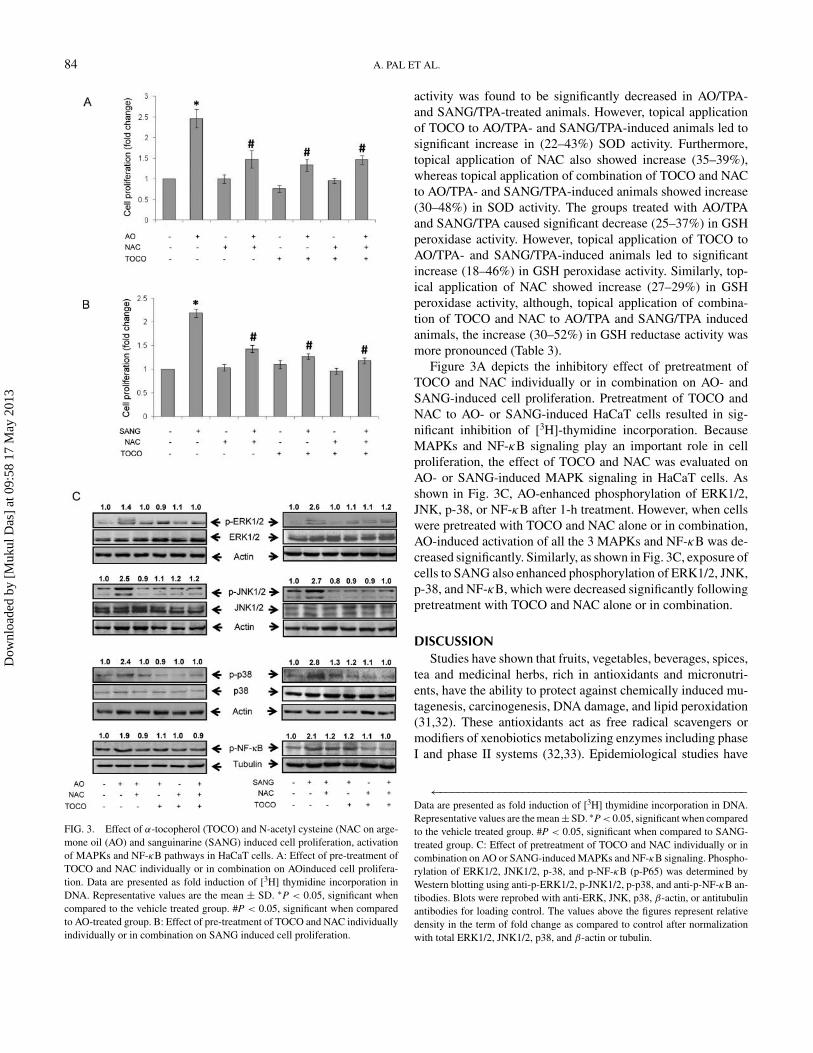
FIG. 3. Effect of α-tocopherol (TOCO) and N-acetyl cysteine (NAC on arge-mone oil (AO) and sanguinarine (SANG) induced cell proliferation, activationof MAPKs and NF-κB pathways in HaCaT cells. A: Effect of pre-treatment ofTOCO and NAC individually or in combination on AOinduced cell prolifera-tion. Data are presented as fold induction of [3H] thymidine incorporation inDNA. Representative values are the mean ± SD. ∗P < 0.05, significant whencompared to the vehicle treated group. #P < 0.05, significant when comparedto AO-treated group. B: Effect of pre-treatment of TOCO and NAC individuallyindividually or in combination on SANG induced cell proliferation.
activity was found to be significantly decreased in AO/TPA-and SANG/TPA-treated animals. However, topical applicationof TOCO to AO/TPA- and SANG/TPA-induced animals led tosignificant increase in (22–43%) SOD activity. Furthermore,topical application of NAC also showed increase (35–39%),whereas topical application of combination of TOCO and NACto AO/TPA- and SANG/TPA-induced animals showed increase(30–48%) in SOD activity. The groups treated with AO/TPAand SANG/TPA caused significant decrease (25–37%) in GSHperoxidase activity. However, topical application of TOCO toAO/TPA- and SANG/TPA-induced animals led to significantincrease (18–46%) in GSH peroxidase activity. Similarly, top-ical application of NAC showed increase (27–29%) in GSHperoxidase activity, although, topical application of combina-tion of TOCO and NAC to AO/TPA and SANG/TPA inducedanimals, the increase (30–52%) in GSH reductase activity wasmore pronounced (Table 3).
Figure 3A depicts the inhibitory effect of pretreatment ofTOCO and NAC individually or in combination on AO- andSANG-induced cell proliferation. Pretreatment of TOCO andNAC to AO- or SANG-induced HaCaT cells resulted in sig-nificant inhibition of [3H]-thymidine incorporation. BecauseMAPKs and NF-κB signaling play an important role in cellproliferation, the effect of TOCO and NAC was evaluated onAO- or SANG-induced MAPK signaling in HaCaT cells. Asshown in Fig. 3C, AO-enhanced phosphorylation of ERK1/2,JNK, p-38, or NF-κB after 1-h treatment. However, when cellswere pretreated with TOCO and NAC alone or in combination,AO-induced activation of all the 3 MAPKs and NF-κB was de-creased significantly. Similarly, as shown in Fig. 3C, exposure ofcells to SANG also enhanced phosphorylation of ERK1/2, JNK,p-38, and NF-κB, which were decreased significantly followingpretreatment with TOCO and NAC alone or in combination.
DISCUSSIONStudies have shown that fruits, vegetables, beverages, spices,
tea and medicinal herbs, rich in antioxidants and micronutri-ents, have the ability to protect against chemically induced mu-tagenesis, carcinogenesis, DNA damage, and lipid peroxidation(31,32). These antioxidants act as free radical scavengers ormodifiers of xenobiotics metabolizing enzymes including phaseI and phase II systems (32,33). Epidemiological studies have
←−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−Data are presented as fold induction of [3H] thymidine incorporation in DNA.Representative values are the mean± SD. ∗P < 0.05, significant when comparedto the vehicle treated group. #P < 0.05, significant when compared to SANG-treated group. C: Effect of pretreatment of TOCO and NAC individually or incombination on AO or SANG-induced MAPKs and NF-κB signaling. Phospho-rylation of ERK1/2, JNK1/2, p-38, and p-NF-κB (p-P65) was determined byWestern blotting using anti-p-ERK1/2, p-JNK1/2, p-p38, and anti-p-NF-κB an-tibodies. Blots were reprobed with anti-ERK, JNK, p38, β-actin, or antitubulinantibodies for loading control. The values above the figures represent relativedensity in the term of fold change as compared to control after normalizationwith total ERK1/2, JNK1/2, p38, and β-actin or tubulin.
Dow
nloa
ded
by [
Muk
ul D
as]
at 0
9:58
17
May
201
3

PROTECTION OF AO INDUCED TUMORIGENESIS BY ANTIOXIDANTS 85
provided evidence for protective role of vitamin A, its precur-sor β-carotene, TOCO, and vitamin C on the development ofvarious type of cancers, including skin cancer (34,35).
Present study indicates that topical application of TOCOand NAC individually or in combination, efficiently decreasedthe tumor incidence and multiplicity induced by AO/TPA orSANG/TPA in mice. In vitro results showed that TOCO and/orNAC significantly decreased the AO- and SANG-induced cellproliferation and activation of ERK, p38, JNK MAPKs, andNF-κB signaling. The protection of AO/TPA- or SANG/TPA-induced tumorigenesis by TOCO may be due to its antioxidantactivity. In this regard it has been observed that ROS are in-volved in the toxicity of AO, thereby leading to decrease thetotal antioxidant capacity and bioantioxidant pool includingTOCO (10,11,36,37). The lipophillic TOCO is considered asone of the most effective endogenous antioxidants to protectmembrane structures being peroxidized (38) and this antioxida-tive effect may be responsible for prevention of carcinogenesis(39). It has been demonstrated that TOCO protects skin fromoxidative stress, acute and chronic photodamage, and tumorformation induced by UVB and chemicals (40,41). Similarly,NAC, a precursor of GSH biosynthesis, has been shown to in-hibit the formation of UVB and benzo(a)pyrene induced skinsquamous cell carcinoma (19). The ability of NAC to inhibitthese skin tumors is likely to be in part due to NAC per se or inpart due to the generation of L-cysteine and GSH in skin cells,which may scavenge the free radical formation or conjugatewith electrophilic metabolite(s) of chemicals (42,43). Topicalapplication of TOCO and NAC to AO/TPA- and SANG/TPA-induced animals enhances the decreased activity of tyrosinase.Since loss of tyrosinase activity has been related to skin tumorsusceptibility in mice due to the deficiency of melanin pigmentproduction, thereby modulating oxidative DNA damage (44).Thus the present results of TOCO and NAC may indicate theprotective role for DNA molecule from oxidative attack causedby AO and SANG, which may be one of the possible mechanismof anticarcinogenic activity. The enzyme histidase found in nor-mal epidermis enhances cellular concentration of histidine thatcan be used for stimulated synthesis of histidine-rich proteinsinvolved in the keratin formation during tumorigenesis (45).Histidase activity in skin carcinoma bearing mouse has beenfound to be absent and lowered in TPA-treated mice (45,46).Animals showing squamous cell carcinoma following AO/TPAor SANG/TPA administration caused inhibition of histidase ac-tivity, which was found to be enhanced following topical appli-cation of TOCO and NAC, implicating the decreased synthesisof keratin as observed in histopathological studies.
The collective action of antioxidants and nonenzymatic smallmolecular antioxidants such as the sulphydryl compound, GSH,is to afford protection against the adverse effect of oxidant or re-active metabolites of carcinogens (47,48). In addition, the levelof antioxidant defense enzymes are lowered in transformedcells and/ or tumor tissue (47,48). The result of the presentstudy showed that GSH content, lipid peroxidation, and antiox-
idant enzymes were found to be modulated in AO/TPA- andSANG/TPA-induced tumor formation in mice. Earlier studieshave also shown that antioxidants enzymes were inhibited inDMBA/TPA-induced skin tumors (49). Furthermore, depletionof these enzymes following exposure to carcinogens and/or tu-mor promoters is well documented (47,48). Interestingly, topicalapplication of TOCO and NAC individually or in combination,to AO/TPA- and SANG/TPA-treated animals showed signifi-cant decrease in LPO along with revival of GSH content andactivities of catalase, SOD, GSH peroxidase, and GSH re-ductase. In this regard, earlier studies have shown thatTOCO significantly enhanced the activity of antioxidant en-zymes in complete and multistage carcinogenesis in mouseskin (15,49). Our data also indicates that NAC is involvedin the revival of antioxidants enzymes in AO/TPA- andSANG/TPA-treated animals and this may lead to scavengeROS generated by AO or SANG, thereby causing inhibition incarcinogenesis.
Furthermore, attempts were made to study the effect ofTOCO and NAC on AO- or SANG-induced cell proliferationand signaling pathways in HaCaT cells. It was observed thatpretreatment of TOCO and NAC individually or in combina-tion significantly inhibits the AO- or SANG-induced cell pro-liferation. In agreement with these findings, TOCO has beenshown to inhibit the proliferation of vascular smooth musclecells, human osteosarcoma cells, and fibroblasts cells (50). Ithas been demonstrated that TOCO inhibits proliferation of sev-eral cancerous cell lines by reducing the levels of cyclin D1and cyclin E (51). Our data showed that AO- and SANG-induced MAPKs and NF-κB signaling was also significantlyinhibited by TOCO and NAC. MAPKs constitute a super fam-ily of proteins that include ERK1/2, JNK1/2, and p38 kinase(52). The involvement of MAPKs pathway in tumor prolifera-tion is well documented (53). Treatment of HaCaT cells withAO and SANG alkaloid showed marked increase in phospho-rylation of ERK1/2, JNK1/2, and p38, which may play a majorrole in cellular function, such as proliferation and transforma-tion (54) whereas ERK pathway suppresses apoptosis and en-hances cell survival or tumorigenesis (55). Studies have shownthat ERK1/2 and p38 are involved in the transcriptional ac-tivation of NF-κB and regulation of COX-2 expression (52).Earlier studies have shown that TOCO inhibits PKC and 5-lipooxygenase at the post-translational level under in vitro study(50,56). Thus our findings indicate that application of TOCOand NAC individually or in combination prevents the AO- andSANG-induced activation of ERK1/2, JNK1/2, and p38 asso-ciated with NF-κB signaling, which may have significance ininhibiting tumor formation in AO/TPA- and SANG/TPA-treatedanimals.
ACKNOWLEDGMENTSThe authors are grateful to Director, CSIR-Indian Institute of
Toxicology Research (IITR) for his keen interest in the present
Dow
nloa
ded
by [
Muk
ul D
as]
at 0
9:58
17
May
201
3

86 A. PAL ET AL.
study. Financial support of Supra-Institutional project-08 grate-fully acknowledged. The article is CSIR-IITR communication#3046.
REFERENCES1. Das M and Khanna SK: Clinicoepidemiological and safety evaluation stud-
ies on argemone oil. CRC Crit Rev Toxicol 27, 273–297, 1997.2. Das M and Khanna SK: Epidemic dropsy. Natl Med J India 11, 207–208,
1998.3. Ansari KM, Chauhan LKS, Dhawan A, Khanna SK, and Das M: Unequiv-
ocal evidence of genotoxic potential of argemone oil in mice. Intl J Cancer112, 890–895, 2004.
4. Ansari KM, Dhawan A, Khanna SK, and Das M: In vivo DNA damagingpotential of sanguinarine alkaloid isolated from argemone oil using alkalinecomet assay. Food Chem Toxicol 43, 147–153, 2005.
5. Das M, Ansari KM, Dhawan A, Shukla Y, and Khanna SK: Correlationof DNA damage in Epidemic Dropsy patients to carcinogenic potential ofargemone oil and isolated sanguinarine alkaloid in mice. Intl J Cancer 117,709–717, 2005.
6. Mishra V, Mishra M, Ansari KM, Chaudhari BP, Khanna R, et al.: Edibleoil adulterants, argemone oil and butter yellow, as aetiological factors forgall bladder cancer. Eur J Cancer 48, 2075–2085, 2012.
7. Sarkar SN: Isolation from argemone oil of dihydrosanguinarine and san-guinarine, Toxicity of sanguinarine. Nature 162, 265–268, 1948.
8. Das M, Upreti KK, and Khanna SK: Biochemical toxicology of Argemoneoil: Role of reactive oxygen species in iron catalyzed lipid peroxidation.Bull Environ Cotam Toxicol 46, 422–430, 1991.
9. Das M, Babu K, Reddy NP, and Srivastava LM: Oxidative damage of plasmaproteins and lipids in epidemic dropsy patients: alterations in antioxidantstatus. Biochim Biophys Acta 1722, 209–217, 2005.
10. Babu CK, Khanna SK, and Das M: Antioxidant status of erythrocytesand their response to oxidative challenge in humans with argemone oilpoisoning. Toxicol Appl Pharmacol 230, 304–311, 2008.
11. Kumar A, Hussain F, Das M, and Khanna SK: An outbreak of epidemicdropsy in the Barabanki district of Uttar Pradesh, India. A limited trial forthe scope of antioxidants in the management of symptoms. Biomed EnvironSci 5, 251–256, 1992.
12. Babu CK, Khanna SK, and Das M: Adulteration of mustard cooking oil withargemone oil: do Indian food regulatory policies and antioxidant therapyboth need revisitation? Antioxid Redox Signal 9, 515–525, 2007.
13. De Flora S, Izzotti A, D’Agostini F, and Balansky RM: Mechanisms of N-acetyl cysteine in the prevention of DNA damage and cancer, with specialreference to smoking-related end-points. Carcinogenesis 22, 999–1013,2001.
14. De Flora S, Izzotti A, Albini A, D’Agostini F, Bagnasco M, et al.: Antigeno-toxic and cancer preventive mechanisms of N-acetyl-L-cysteine. In: Can-cer Chemoprevention, Vol. 1, Promising Cancer Chemopreventive Agents,Kelloff GJ, Hawk ET, and Sigman CC (eds.). Totowa, NJ: Humana Press,2004, pp. 37–67.
15. Rahman S, Bhatia K, Khan AQ, Kaur M, Ahmad F, et al.: Topically appliedvitamin E prevents massive cutaneous inflammatory and oxidative stressresponses induced by double application of 12-O-tetradecanoylphorbol-13-acetate (TPA) in mice. Chem Biol Interact 15, 195–205, 2008.
16. Devaraj S, Li D, and Jialal I: The effects of alpha-tocopherol supplementa-tion on monocyte function. Decreased lipid oxidation, interleukin 1 beta se-cretion, and monocyte adhesion to endothelium. J Clin Invest 98, 756–763,1996.
17. Upreti KK, Das M, and Khanna SK: Biochemical toxicology of argemoneoil. I. Effect on hepatic cytochrome P-450 and xenobiotic metabolizingenzymes. J Appl Toxicol 11, 203–209, 1991.
18. Perchellet JP, Owen MD, Posey TD, Orten DK, and Schneider BA: In-hibitory effects of glutathione level-raising agents and D-α tocopherolon ornithine decarboxylase induction and mouse skin tumor promotion
by 12-O-tetradecanoyl-phorbol-13-acetate. Carcinogenesis, 6, 567–573,1985.
19. D’Agostini F, Balansky RM, Camoirano A, and Flora SD: Modulationof light-induced skin tumors by N-acetyl cysteine and/or ascorbic acid inhairless mice. Carcinogenesis, 26, 657–664, 2005.
20. Shimkin MB and Andervont HB: Comparative carcinogenicity of threecarcinogenic hydrocarbons. J Nat Cancer Inst 1, 57–62, 1940.
21. Das M, Bickers DR, and Mukhtar H: Protection against chemically inducedskin tumorigenesis in SENCAR mice by tannic acid. Intl J Cancer 43,468–470, 1989.
22. Ellman GL: Tissue sulfhydryl groups. Arch Biochem Biophys 82, 70–77,1959.
23. Utley HC, Burnheim E, and Hochestein P: Effect of sulfhydryl agents onperoxidation in microsomes. Arch Biochem Biophys 118, 29–32, 1967.
24. Sinha AK: Colorimetric assay of catalase. Anal Biochem 47, 389–394,1972.
25. Carlberg I and Mannervik B: Glutathione reductase. In: Methods in enzy-mology, Meister A (ed.). Orlando, FL: Academic Press, 1985, pp. 484–490.
26. Rotruck JT, Pope AL, Ganther HE, Swanson AB, Hafeman DG, et al.:Selenium: Biochemical role as a component of glutathione peroxidase.Science 179, 588–590, 1973.
27. Misra HP and Fridovich I: The role of superoxide anion in the auto oxidationof epinephrine and a simple assay for superoxide dismutase. J Biol Chem247, 3170–3175, 1972.
28. Tabor H and Mehler AH: Enzymes of protein metabolism, histidase anduroconase. In: Methods in Enzymology, Colowick SP and Kaplan HO (eds.).New York: Academic Press, 1955, pp. 615–632.
29. Horowitz MH, Fling M, and Horn E: Tyrosinase. In: Methods in Enzymol-ogy, Tabor H and Tabor CW (eds.). New York: Academic Press, 1970, pp.615–632.
30. Ansari KM, Rundhaug JE, and Fischer SM: Multiple signaling pathways areresponsible for prostaglandin E2-induced murine keratinocyte proliferation.Mol Cancer Res 6, 1003–1016, 2008.
31. Ames BN: Dietary carcinogens and anticarcinogens, oxygen radicals anddegenerative diseases. Science 221, 1256–1264, 1983.
32. Conney AH: Enzyme induction and dietary chemicals as approaches tocancer chemoprevention: The seventh DeWitt S. Goodman Lecture. CancerRes 63, 7005–7031, 1987.
33. Dgawa K, Futakuchi M, Hirose M, Boonuaphiphat P, Miziguchi Y, et al.:Stage and organ dependent effect of 1-O-hexyl-2, 3,5- trimethyl hydro-quinone and phenylethyl isothiocyanate in a rat multiorgan carcinogenesismodel. Intl J Cancer 76, 851–856, 1987.
34. Berger MR, Berger I, and Schmahl D: Vitamins and cancers. In: Nutrition,Toxicology and Cancer, Rowland IR (ed.). Boca Raton, FL: CRC Press,1991, pp. 517–548..
35. Burke KE, Clive J, Combs GF Jr, Commisso J, Keen CL et al. : Effectsof topical and oral vitamin E on pigmentation and skin cancer inducedby ultraviolet irradiation in SKH:2 hairless mice. Nutr Cancer 38, 87–97,2000.
36. Turveson RW, Carsson RA, Marley KA, Wang G, and Berenbaum MR:Sanguinarine, a phototoxic H2O2 producing alkaloid. Photochem Photobiol50, 733–738, 1989.
37. Upreti KK, Das M, and Khanna SK: Role of antioxidants and scavengerson argemone oil induced toxicity in rats. Arch Environ Cont Toxicol 20,531–537, 1991.
38. Witting LA: Vitamin E and lipid antioxidants in free-radicals-initiated reac-tions. In: Free Radical in Biological, Pryor WA (ed.). New York: AcademicPress, 1980, 295–319.
39. Cook MG and McNamara P: Effect of dietary vitamin E on dimethylhydrazine-induced colonic tumors in mice. Cancer Res 40, 1329–1331,1980.
40. Foote JA, Ranger-Moore JR, Einspahr JG, Saboda K, Kenyon J,et al.: Chemoprevention of human actinic keratosis by topical DL-alpha-tocopherol. Cancer Prev Res 4, 394–400, 2009.
Dow
nloa
ded
by [
Muk
ul D
as]
at 0
9:58
17
May
201
3

PROTECTION OF AO INDUCED TUMORIGENESIS BY ANTIOXIDANTS 87
41. Berton TR, Conti CJ, Mitchell DL, Aldaz CM, Lubet RA, et al.: The effectof vitamin E acetate on ultraviolet-induced mouse skin carcinogenesis. MolCarcinogenesis 23, 175–184, 1998.
42. Martin KR, Trempus C, Saulnier M, Kari FW, Barrett JC, et al.: Di-etary N-acetyl-L-cysteine modulates benzo[a]pyrene-induced skin tumorsin cancer-prone p53 haploin sufficient Tg.AC (v-Ha-ras) mice. Carcino-genesis 22, 1373–1378, 1998.
43. Steenvoorden DP, Hasselbaink DM, Beijersbergen van, and HenegouwenGM: Protection against UV-induced reactive intermediates in human cellsand mouse skin by glutathione precursors: a comparison of N-acetylcysteine and glutathione ethylester. Photochem Photobiol 67, 651–656,1998.
44. Suran A, Spinola M, Pazzaglia S, Peissel B, Tiverson C, et al.: Loss of tyrosi-nase activity confers increased skin tumor susceptibility in mice. Oncogene23, 4130–4135, 2004.
45. Colburn NH, Shigeko L, and Rebecca H: Decrease of epidermal histidaseactivity by tumor promoting phorbol esters. Cancer Res 35, 3154–3159,1975.
46. Barnhisel ML, Priest RE, and Priest JH: Histidase function in human epi-dermis cells. J Cell Physiol 76, 7–15, 1970.
47. Sun Y: Free radicals, antioxidants enzymes, and carcinogenesis. Free Rad-ical Biol Med 8, 583–599, 1990.
48. Perchellet JP and Perchellet EM: Antioxidants and multistage carcinogen-esis in mouse skin. Free Red Biol Med 7, 377–408, 1989.
49. Perchellet JP, Abney NL, Thomas RM, Guislain YL, and Perchellet EM:Effects of combined treatments with selenium, glutathione, and vitamin E
on glutathione peroxidase activity, ornithine decarboxylase induction, andcomplete and multistage carcinogenesis in mouse skin. Cancer Res 47,477–485, 1987.
50. Boscoboinik D, Szewczyk A, and Azzi A: α-tocopherol (vitamin E) regu-lates vascular smooth muscle cell proliferation and protein kinase C activity.Arch Biochem Biophys 286, 264–269, 1991.
51. Gysin R, Azzi A, and Visarius T: Gamma-tocopherol inhibits human cancercell cycle progression and cell proliferation by down-regulation of cyclins.FASEB J 16, 1952–1954, 2002.
52. Chun KS, Kim SH, Song YS, and Surh YJ: Celecoxib inhibits phorbol esterinduced expression of COX-2 and activation of AP-1 and p38 MAP kinasein mouse skin. Carcinogenesis 25, 13–22, 2004.
53. Yano T, Yajima S, Hagiwara K, Kumadaki I, and Yano Y: Vitamin Einhibits cell proliferation and the activation of extracellular signal-regulatedkinase during the promotion phase of lung tumourigenesis irrespective ofantioxidative effect. Carcinogenesis 21, 2129–2133, 2000.
54. Potapova O, Gorospe M, Bost F, Dean NM, Gaarde WA, et al.: c-Jun N-terminal kinase is essential for growth of human T 98G glioblastoma cells.J Biol Chem 275, 24767–24775, 2000.
55. Huang C, Ma WY, Young MR, Colburn N, and Dong Z: Shortage of mitogenactivated protein kinase is responsible for resistance to AP-1 transactiva-tion and transformation in mouse JB6 cells. Proc Natl Acad Sci USA 95,156–161, 1998.
56. Freedman JE, Farhat JH, Losalzo J, and Keaney JF: Alpha-tocopherol in-hibits aggregation of human platelets by a protein kinase C dependentmechanism. Circulation 94, 2434–2440, 1996.
Dow
nloa
ded
by [
Muk
ul D
as]
at 0
9:58
17
May
201
3