
Expression of granulocyte colony stimulating factor and its receptor by retinal pigment epithelial...
9
Blood Cells, Molecules, and Diseases (1995) 21(20) Oct 31: 192-200 S.S. Kothari et al. The Hope Heart Institute, 528 18th Avenue, Seattle, WA, USA; 1 Department of Biochemistry, the University of Washington School of Medicine, Seattle, WA, USA. Present address: Department of Veterinary 2 PathoBiology, University of Minnesota,St. Paul, MN 55108; Department of Medical Education, Providence Medical Center, and Department of Biochemistry, University of Washington School of Medicine, Seattle, 3 WA, USA; Department of Medicine, University of Washington School of Medicine, Seattle, Washington USA; 4 Reprint requests to: William P. Hammond, M.D., Department of Medical Education, Providence Medical Center, 500 17th Ave, C-34008, Seattle, WA 5 98124 USA, phone (206) 320-2552, fax (206) 320-3589, e-mail: [email protected]. ©The Blood Cells Foundation, La Jolla, California USA ISSN No. 1079-9796 Published by the Blood Cells Foundation Established by Springer-Verlag, Inc. in 1975 192 Expression of Granulocyte Colony Stimulating Factor (G-CSF) and Granulocyte/Macrophage Colony Stimulating Factor (GM-CSF) mRNA upon Stimulation with Phorbol Ester Submitted on 06/25/95; revised on 10/19/95 (communicated by Seymour J. Klebanoff, M.D., Ph.D., 10/26/95) Sudesh S. Kothari , Mitchell S. Abrahamsen , Toby Cole , and William P. Hammond 1 2 3 1,4,5 ABSTRACT: Stimulated human peripheral blood mononuclear cells (MNC) have been shown to express both G-CSF and GM-CSF. Furthermore, G-CSF is expressed by monocytes but not lymphocytes, whereas GM-CSF is expressed largely by T lymphocytes and at low levels in monocytes/ macrophages. Here we present the effect of TPA (12-O-tetradecanoyl phorbol-13-acetate) on G-CSF and GM-CSF expression in stimulated human MNCs and T lymphocytes. We observed that TPA (30nM) decreased G-CSF mRNA levels in MNCs, while ionomycin increased G-CSF in a dose-dependent manner. TPA and ionomycin individually increased GM-CSF mRNA levels in T-lymphocytes and MNCs. Further, GM-CSF was induced synergistically by TPA plus ionomycin, whereas this combination markedly decreased G-CSF mRNA levels in MNCs. These data suggest at least two signaling pathways by which G-CSF and GM-CSF mRNA levels are modulated in a mixed population of monocytes and T lymphocytes, namely protein kinase C (PKC) and calcium. These signals seem to act synergistically in lymphocytes to increase GM-CSF, and not G-CSF mRNA levels specifically. It would also appear these signals act on MNCs in an opposing manner to decrease G-CSF mRNA levels, indicating that activation of PKC and the calcium signaling pathway lead to a cell-type specific modulation of individual cytokines and precise regulation of granulocyte production. Keywords: granulocyte colony stimulating factor, granulocyte/macrophage colony stimulating factor, peripheral blood mononuclear cells, phorbol esters, mRNA expression INTRODUCTION hematopoietic progenitors into various cell types. Human peripheral blood mononuclear cells stimulating factor (G-CSF) and granulocyte/ (MNC) are known to express several different macrophage colony stimulating factor (GM- CSF), cytokines in response to specific stimuli, which can act upon granulocyte/ macrophage lead to the proliferation and differentiation of multipotential progenitors (CFU-GM) that have Two such cytokines, granulocyte colony
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Expression of granulocyte colony stimulating factor and its receptor by retinal pigment epithelial...

Blood Cells, Molecules, and Diseases (1995) 21(20) Oct 31: 192-200 S.S. Kothari et al.
The Hope Heart Institute, 528 18th Avenue, Seattle, WA, USA;1
Department of Biochemistry, the University of Washington School of Medicine, Seattle, WA, USA. Present address: Department of Veterinary2
PathoBiology, University of Minnesota,St. Paul, MN 55108;
Department of Medical Education, Providence Medical Center, and Department of Biochemistry, University of Washington School of Medicine, Seattle,3
WA, USA;
Department of Medicine, University of Washington School of Medicine, Seattle, Washington USA;4
Reprint requests to: William P. Hammond, M.D., Department of Medical Education, Providence Medical Center, 500 17th Ave, C-34008, Seattle, WA 5
98124 USA, phone (206) 320-2552, fax (206) 320-3589, e-mail: [email protected].
©The Blood Cells Foundation, La Jolla, California USAISSN No. 1079-9796
Published by the Blood Cells FoundationEstablished by Springer-Verlag, Inc. in 1975
192
Expression of Granulocyte Colony Stimulating Factor (G-CSF)and Granulocyte/Macrophage Colony Stimulating Factor(GM-CSF) mRNA upon Stimulation with Phorbol EsterSubmitted on 06/25/95; revised on 10/19/95(communicated by Seymour J. Klebanoff, M.D., Ph.D., 10/26/95)
Sudesh S. Kothari , Mitchell S. Abrahamsen , Toby Cole , and William P. Hammond1 2 3 1,4,5
ABSTRACT: Stimulated human peripheral blood mononuclear cells (MNC) have been shown to express bothG-CSF and GM-CSF. Furthermore, G-CSF is expressed by monocytes but not lymphocytes, whereas GM-CSFis expressed largely by T lymphocytes and at low levels in monocytes/ macrophages. Here we present the effectof TPA (12-O-tetradecanoyl phorbol-13-acetate) on G-CSF and GM-CSF expression in stimulated human MNCsand T lymphocytes. We observed that TPA (30nM) decreased G-CSF mRNA levels in MNCs, while ionomycinincreased G-CSF in a dose-dependent manner. TPA and ionomycin individually increased GM-CSF mRNA levelsin T-lymphocytes and MNCs. Further, GM-CSF was induced synergistically by TPA plus ionomycin, whereasthis combination markedly decreased G-CSF mRNA levels in MNCs. These data suggest at least two signaling pathways by which G-CSF and GM-CSF mRNA levels are modulatedin a mixed population of monocytes and T lymphocytes, namely protein kinase C (PKC) and calcium. Thesesignals seem to act synergistically in lymphocytes to increase GM-CSF, and not G-CSF mRNA levels specifically.It would also appear these signals act on MNCs in an opposing manner to decrease G-CSF mRNA levels,indicating that activation of PKC and the calcium signaling pathway lead to a cell-type specific modulation ofindividual cytokines and precise regulation of granulocyte production.Keywords: granulocyte colony stimulating factor, granulocyte/macrophage colony stimulating factor, peripheral blood mononuclear cells, phorbol esters, mRNAexpression
INTRODUCTION hematopoietic progenitors into various cell types.
Human peripheral blood mononuclear cells stimulating factor (G-CSF) and granulocyte/(MNC) are known to express several different macrophage colony stimulating factor (GM- CSF),cytokines in response to specific stimuli, which can act upon granulocyte/ macrophagelead to the proliferation and differentiation of multipotential progenitors (CFU-GM) that have
Two such cytokines, granulocyte colony

S.S. Kothari et al. Blood Cells, Molecules, and Diseases (1995) 21(20) Oct 31: 192-200
193
the capacity to proliferate and differentiate to form simultaneously down modulates G-CSF mRNAcolonies of granulocytes, monocytes or both. levels within a mixed lymphocyte /monocyte cellG-CSF specifically induces colonies of neutro- population. Thus it is suggested that TPAphilic granulocytes, whereas GM-CSF can induce mediated signal transduction regulatesformation of colonies containing both granulopoiesis through a combination of specificgranulocytes and macrophages [see reviews 1-3]. factor stimulation and inhibition.
G-CSF has been reported to be expressed bymonocytes upon stimulation with various factors MATERIALS AND METHODSsuch as LPS, phorbol ester (TPA), ionophores,IL-1b, and TNFa (3-6). At the promoter level, the Reagentsspecificity of gene expression has been shown tobe mediated via the CK-1 DNA element, which Escherichia coli lipopolysaccharide (LPS),binds specific factor(s) in response to IL-1b and 12-O-Tetradecanoyl phorbol 13-acetate (TPA),TNFa stimulation (7). Also upregulation of G-CSF and L-Asparagine were purchased from Sigmagene expression does not effect GM-CSF gene Chemical Co. (St. Louis, MO.). Ionomycin wasexpression and visa versa (8). Lymphocytes purchased from Calbiochem-Novobiochem Co.appear not to express G-CSF (9,10,11). (La Jolla, CA.), penicillin-streptomycin solution
GM-CSF is predominantly expressed by from, Gibco BRL (Gaithersburg, MD), andT-lymphocytes and to a lesser extent by NCTC109 medium, from Whittaker Bioproductsmonocytes (5,10,12,13). GM-CSF gene (Walkersville, MD)expression is greatly induced by phorbol estersand ionophores, as well as IL-1b and TNFa. TheCell Preparation and CultureGM-CSF promoter sequence is shown to have atleast two DNA elements that mediate specific Human blood was obtained from subjectsgene expression, namely the TPA responsive ranging in age from 30 to 53 years, underelement GM2 and the TPA plus ionomycin protocols approved by the Human Subjectsresponsive element CLEO (conserved lymphokine Review Committee of the University ofelement 0) (14). In fibroblasts GM-CSF is shown Washington and the Clinical Research Center atto be induced by at least 3 signal transduction the University of Washington Medical Center.mechanisms, involving i) [Ca ] , ii) PKC, and iii) Human peripheral blood mononuclear cells2+
I
a G protein-dependent activation pathway (15). (MNCs) were separated by Ficoll-Hypaque(FH)Our hypothesis was to test whether stimulation density centrifugation (sp. gravity 1.077).
with phorbol ester elevated mRNA levels of both Lymphocytes were enriched by passage of MNCsG- and GM-CSFs in mixed monocyte and through nylon wool (NW) (16). Cells were restedlymphocyte cell population, and how this might overnight at 4 C prior to aliquoting andeffect the modulation of granulopoiesis. In theseexperiments, we investigated the effects ofvarying concentrations of TPA and/or ionomycinstimulation of GM-CSF and G-CSF geneexpression, to ascertain whether stimulation ofspecific signal pathways induced either both genessimultaneously or differently in a mixedpopulation of monocytes and lymphocytes(MNCs), and in a lymphocyte-enriched popula-tion. Here we show that TPA indeed stimulatesGM-CSF mRNA levels in lymphocytes and
o
stimulating. Cells were cultured in NCTC109medium (500ml was supplemented with 10% fetalcalf serum, 2.5ml penicillin (100U/ml)-streptomycin (100U/ml) solution, 5ml 4mg/mlL-asparagine and 5ml 1mM Sodium pyruvate).Approximately 1x10 cells were stimulated in7
plastic flasks (Falcon, T25) with various agents atdifferent concentrations as well as for variousdurations.

Blood Cells, Molecules, and Diseases (1995) 21(20) Oct 31: 192-200 S.S. Kothari et al.
194
RNA Extraction, Northern Blotting and of signal strength of stimulated versusHybridization unstimulated control, after normalization using
Total cellular RNA was isolated using loaded (data not shown). Inferences about peakguanidinium isothiocyanate (17), size-fractionated message levels were made from the densitometricon 1.2% agarose gels containing formaldehyde analysis.and blotted onto nitrocellulose membranes. Ineach experiment the entire RNA sample wasDNA Probesloaded for each data point and represented a fixednumber of cells within each experiment. A 600bp human cDNA GM-CSF insertHybridization and washing conditions were (corresponding to GM-CSF cDNA sequenceperformed as described previously (11). Blots 0-600bp) and 1.455kb human cDNA G-CSFwere restripped and rehybridized to other probes. insert (corresponding to G-CSF cDNA sequenceDensitometric image analysis was performed on a 30-1485bp) were purified from their respectivePower Macintosh using public domain NIH Image plasmid vectors and labeled with P-dCTP byprogram, version 1.55 (written by Wayne Rasband nick translation (18). Human glyceraldehydeat the U.S. National Institutes of Health and 3-phosphate dehydrogenase (G3PDH) cDNAavailable from the internet by anonymous ftp from probe (corresponding to cDNA sequencezippy.nimh.nih.gov or on floppy disk from 71-1053bp) was obtained by using PCR (poly-NTIS,5285 Port Royal Rd., Springfield VA merase chain reaction) amplimer set obtained22161, part no. PB93-504868). Increase or from Clontech Laboratories, Inc., (Palo Alto,decrease in mRNA levels were based on the ratio CA.).
G3PDH signals to estimate the amount of RNA
32
Figure 1. Expression of G-CSF and GM-CSF mRNA in MNCs stimulated for 24 hr with varying concentration of TPA, ionomycin, LPS and TPA plusionomycin. Control lanes 1 and 2 represent RNA extracted from unstimulated MNCs after 24 hr incubation, whereas in Control lane 3, RNA was extracted fromunstimulated MNCs after 6 hr incubation in plastic flask.

S.S. Kothari et al. Blood Cells, Molecules, and Diseases (1995) 21(20) Oct 31: 192-200
195
RESULTS concentrations above 3nM and when
MNCs consisted of approximately 30% with various combinations of different TPAmonocytes and 70% lymphocytes. A Northern blot concentrations (ranging from 3nM to 30nM) andof the total cellular RNA extracted from cells ionomycin concentrations (ranging from 0.2µM tostimulated for 24 hours (Fig. 1) with varying 1µM) did not show any G-CSF mRNA expressionconcentrations of TPA, ionomycin, LPS, and TPA (Fig. 1 and Fig. 2, and data not shown). plus ionomycin, showed there was a TPA However, GM-CSF mRNA levels showed adose-dependent increase in GM-CSF mRNA mean fold increase of 4.85 and 7.16, whenlevels, which was maximal at 30nM (as stimulated with 3nM and 30nM TPAdetermined by densitometry). Likewise, induction concentrations, respectively (Fig. 2).with ionomycin was maximal at a concentration Co-stimulation with 0.2µM ionomycin plus thebelow 1µM. There was a dose-dependent above concentrations of TPA showed marginalsynergistic effect upon co-stimulation with TPA increases to 5.05 and 8.8 fold, respectively. Theseplus ionomycin, when the TPA concentration was levels were elevated to as much as 20-fold, aftermaintained at 3nM and ionomycin concentrations stimulation with 30nM TPA plus 1µM ionomycinwere varied between 0.1µM and 1µM. (data not shown). Stimulation with 0.2µM
G-CSF mRNA expression levels (Fig. 1), in ionomycin alone, resulted in a 4.8-fold meanboth the TPA and ionomycin-stimulated MNCs increase in GM-CSF mRNA levels as compared todecreased in a dose-dependent manner. However, 1.2-fold increase upon stimulation with LPS.at low doses of TPA (peak at 1.5nM) and of Since T-lymphocytes have previously beenionomycin (peaked at 0.2µM), there was a small shown to express GM-CSF but not G-CSFincrease in G-CSF mRNA levels. No G-CSF mRNA, it was interesting to see whether wemRNA was detected in cells treated with TPA would obtain similar results in a cell populationconcentrations above 3nM, whereas GM-CSF consisting largely of lymphocytes uponmRNA expression, as shown in Fig. 1, was stimulation with the various agents. In a 24 hourgreatly induced by TPA at 3nM and above. When time course experiment, nylon wool (NW)TPA (3nM) was combined with ionomycin enriched lymphocytes were stimulated with TPA(0.1µM), G-CSF mRNA levels were detected but (30nM), ionomycin (1µM), TPA plus ionomycindid not show an increase over TPA alone. Rather, (30nM and 1µM, respectively), or LPSG-CSF mRNA levels fell below basal expression(200ng/ml), over 4, 6, 8 or 24 hours (Fig. 3). Inwhen stimulated with higher concentrations of this experiment the NW separated cells consistedTPA alone or co-stimulation with TPA plus of 2% monocytes and 98% lymphocytes and theionomycin. Stimulation with 2µM and higher MNCs consisted of 28% monocytes and 72%concentration of ionomycin abolished G-CSF lymphocytes. MNCs were unstimulated and themRNA expression. G-CSF mRNA levels were time course was done in duplicate (Fig. 3).induced significantly with LPS stimulation. Unstimulated MNCs expressed both GM-CSF and
No difference in GCSF gene expression was G-CSF mRNA in a similar fashion, peaking at 8seen between MNCs stimulated with TPA for 6 hours. However, no GM-CSF mRNA wasand 24 hours (data not shown). detected at the zero time point, whereas G-CSF
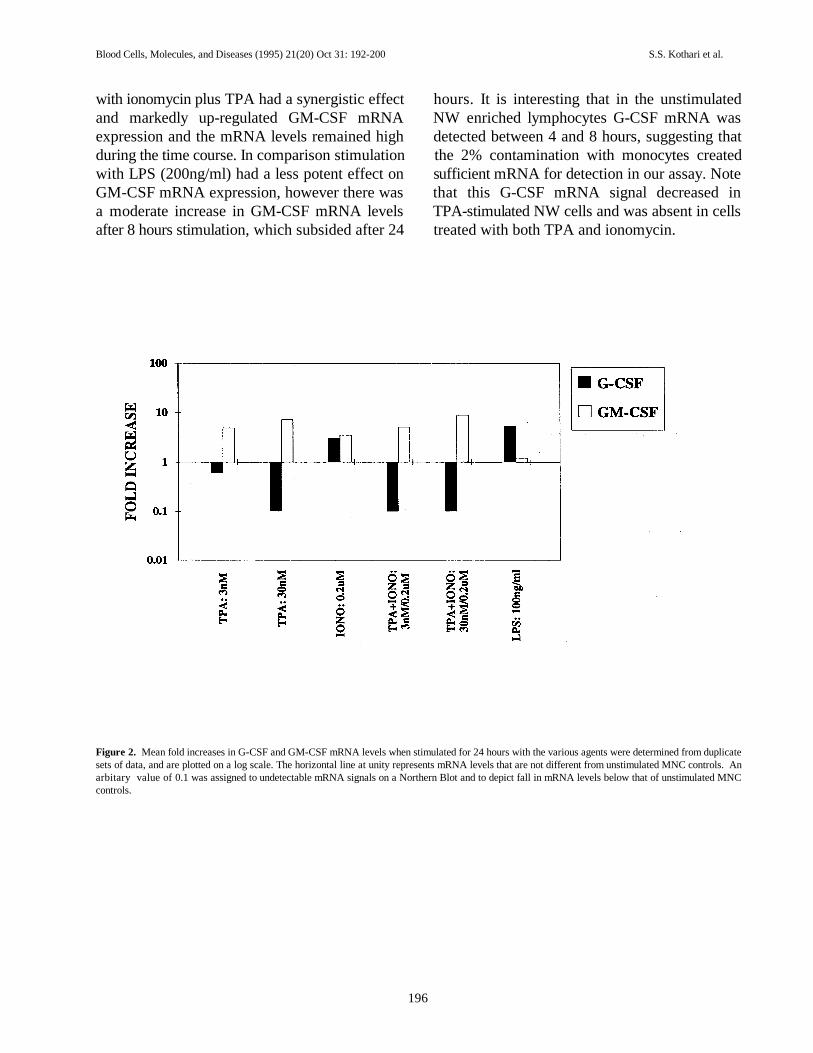
Fold increases in G-CSF and GM-CSF mRNA mRNA was present.levels were calculated based upon standardization NW enriched lymphocytes stimulated withof controls to equal 1 and undetectable signals on TPA alone caused an increase in the GM-CSFthe Northern blot to equal 0.1. The mean fold mRNA levels over the time course (Fig. 3).increases from duplicate experiments were plotted Ionomycin alone also showed an increase in theon a log scale and summarized in Fig. 2. G-CSF GM-CSF mRNA levels at between 6 and 8 hoursmRNA levels were undetectable at TPA and then declined by 24 hours. Co-stimulation
co-stimulated with ionomycin. Co-stimulation
Blood Cells, Molecules, and Diseases (1995) 21(20) Oct 31: 192-200 S.S. Kothari et al.
196
with ionomycin plus TPA had a synergistic effect hours. It is interesting that in the unstimulatedand markedly up-regulated GM-CSF mRNA NW enriched lymphocytes G-CSF mRNA wasexpression and the mRNA levels remained high detected between 4 and 8 hours, suggesting thatduring the time course. In comparison stimulation the 2% contamination with monocytes createdwith LPS (200ng/ml) had a less potent effect on sufficient mRNA for detection in our assay. NoteGM-CSF mRNA expression, however there was that this G-CSF mRNA signal decreased ina moderate increase in GM-CSF mRNA levels TPA-stimulated NW cells and was absent in cellsafter 8 hours stimulation, which subsided after 24 treated with both TPA and ionomycin.
Figure 2. Mean fold increases in G-CSF and GM-CSF mRNA levels when stimulated for 24 hours with the various agents were determined from duplicatesets of data, and are plotted on a log scale. The horizontal line at unity represents mRNA levels that are not different from unstimulated MNC controls. Anarbitary value of 0.1 was assigned to undetectable mRNA signals on a Northern Blot and to depict fall in mRNA levels below that of unstimulated MNCcontrols.

S.S. Kothari et al. Blood Cells, Molecules, and Diseases (1995) 21(20) Oct 31: 192-200
197

Blood Cells, Molecules, and Diseases (1995) 21(20) Oct 31: 192-200 S.S. Kothari et al.
198
DISCUSSION plus ionomycin have been shown to release
Unstimulated MNCs, consisting of a mixture can subsequently stimulate monocytes to expressof monocytes and lymphocytes, showed G-CSFand GM-CSF gene expression that peaked ataround 6-8 hours and then subsequently declinedover 24 hours. The presence of G-CSF mRNA,unlike GM-CSF mRNA, was probably due toactivation of monocytes upon adhering to plastic,as reported by others (10)
MNCs stimulated with LPS showed a greaterincrease in G-CSF mRNA levels then GM-CSFmRNA levels, in agreement with current literature(19). Ionomycin stimulated MNCs showedmaximal expression of G-CSF and GM-CSFmRNA levels at low concentration. Stimulation ofmonocytes with TPA at around 1nM concentrationhas been shown to increase G-CSF mRNA levels(5,7,20). Our results are in agreement at similardoses, but here we show that at 3nM and above,G-CSF mRNA levels appeared to bedown-regulated. In contrast, GM-CSF mRNA wasup regulated at similar doses, suggesting that instimulated MNCs, variation in TPA concentrationleads to a switch in expression of specific colonystimulating factor. Interestingly, our resultsconfirm the findings of upregulated GM-CSFmRNA level with TPA plus ionomycin (14,15)and extend these findings by showing that TPAabolished the increase in G-CSF mRNA levelinduced by ionomycin alone. Importantly, asimilar pattern of response was observed in MNCsstimulated for only 6 hours. This suggests thatthere is not a qualitative difference at earlier andlater time points. It would be interesting to seewhether TPA stimulation could abolish G-CSFexpression induced by other stimulators, such asIL-1 or TNFa (21). Furthermore, although proteinassays were not performed, we would expectabrogation of G-CSF mRNA expression tocorrelate with fall in protein levels.
These data suggest that TPA at high dosesacted upon MNCs to decrease G-CSF mRNAproduction. This effect was either directly onmonocytes or indirectly via lymphocytes (10) bya hitherto undefined mechanism, present in amixed cell population. T cells activated by TPA
several lymphokines, such as TNFa (22) which
G-CSF (21). We have not directly looked at theeffect of TPA on cytokine expression in purifiedmonocytes. However, others have observed inmonocytes/macrophages stimulated with TPA,either no induction of G-CSF gene expression (23)or post-transcriptional stabilization of G-CSFmRNA levels rather than an increase intranscriptional rate (7,18,19). Our data do notpermit resolution of the mechanism(s) by whichalteration in the mRNA levels resulted, such asfrom increased transcriptional rates orpost-transcriptional mRNA stability. However,since purified monocytes stimulated with TPAeither do not express GM-CSF (24) or expressGM-CSF at very low levels (5), it is suggestedthat stimulation with TPA or TPA plus ionomycinleads to GM-CSF mRNA expression bylymphocytes and down modulation of G-CSF inmonocytes within a mixed population.
We therefore conclude that TPA acts in adose-dependent manner upon a mixed populationof monocytes and lymphocytes (MNCs) toregulate G-CSF and GM-CSF mRNA levels inopposite directions. Hence granulocyte productionappears to be regulated by these two cytokines'relative levels via intracellular signal transductionpathways which depend upon the magnitude ofthe stimulus. Whether similar mechanisms existsin other cell types which express both G-CSF andGM-CSF mRNAs, such as endothelial cells (25),fibroblasts (26) or bone marrow stromalfibroblasts (27) remains to be determined.
ACKNOWLEDGMENT
We would like to acknowledge the support of Dr.David R. Morris in whose laboratory these studieswere initiated and Eva Csiba for excellent technicalskills. We wish to thank Mary-Ann Nelson and DaveCriss at the Hope Heart Institute, for their help withthe figures. This work was supported in part by NIHgrant DK42083.

S.S. Kothari et al. Blood Cells, Molecules, and Diseases (1995) 21(20) Oct 31: 192-200
199
REFERENCES Uamaguchi-Iwai Y. Activation of lymphokine genes
1. Kaushansky K. The molecular biology of the colony respond to T cell activation signals. Pharmac Therstimulating factors. Blood Cell 13:3-15, 1987. 55:303-318, 1992.
2. Rapoport AP, Abboud CN, DiPersio JF. 15. Yamato K, El-Hajjaoui Z, Kuo JF, Koeffler P.Granulocyte-macrophage colony stimulating factor Granulocyte-macrophage colony-stimulating factor:(GM-CSF) and granulocyte colony stimulating factor signals for its mRNA accumulation. Blood(G-CSF): receptor biology, signal transduction, and 74:1314-1320, 1989.neutrophil activation. Blood Rev 6:43-57, 1992. 16. Chait A, Henze K, Mazzone T, Jensen M, Hammond
3. Metcalf D. The molecular control of cell division, W. Low density lipoprotein receptor activity in freshlydifferentiation commitment and maturation in isolated human blood monocytes and lymphocytes.haemopoietic cells. Nature 339:27-30,1989. Metabolism 31:721-727, 1982.
4. Herrmann F, Mertelsmann R. Polypeptides controlling 17. Chomcynski P, Sacchi N. Single step method of RNAhematopoietic cell development and activation. Blut isolation by guanidinium thiocyanate phenol-58:117-128, 1989. chloroform extraction. Anal Biochem 162:156-159,
5. Vellenga E, Rambaldi A, Ernst TJ, Ostapovicz D, 1987.Griffin JD. Independent regulation of M-CSF and 18. Maniatis T, Fritsch EF, Sambrook J. MolecularG-CSF gene expression in human monocytes. Blood cloning: A laboratory manual, 2nd ed. Cold Spring71:1529-1532, 1988. Harbor, NY: Cold Spring Harbor Laboratory Press,
6. Oster W, Lindemann A, Mertelsmann R, Herrmann F. 1989. Regulation of gene expression of M-, G-, GM-, and 19. Ernst TJ, Ritchie AR, Demetri GD, Griffin JD.multi-CSF in normal and malignant hematopoietic Regulation of G- and M-CSF mRNA levels in humancells. Blood Cells 14:443-462, 1988. blood monocytes is mediated primarily at a
7. Shannon MF, Coles LS, Goodall GJ, Fielke RK, post-transcriptional level. J Biol Chem 264:5700-Lagnado CA, Vadas MA. Three essential promoter 5703, 1989. elements mediate tumour necrosis factor and 20. Lee SM, Knoppel E, van de Van C, Cairo MS.interleukin-1 activation of the granulocyte colony Transcriptional rates of granulocyte-macrophagestimulating factor gene. Grow Fact 7:181-193, 1992. stimulating factor, granulocyte colony-stimulating
8. Segal GM, Smith TD, Heinrich MC, Ey FS, Bagby GC factor, interleukin-3, and macrophage colony stimulat-Jr. Specific repression of granulocyte-macrophage ing factor genes in activated cord versus adultand granulocyte colony stimulating factor gene mononuclear cells: alteration in cytokine expressionexpression in interleukin-1 stimulated endothelial cells may be secondary to post-transcriptional instability.with antisense oligodeoxynucleotides. Blood Pediatr Res 34:560-564, 1993.80:609-616, 1992. 21. Oster W, Lindemann A, Horn S, Mertalsmann R,
9. Clark SC, Kamen R. The human hematopoietic Herrmann F. Tumor necrosis factor (TNF)-alpha butcolony-stimulating factors. Science 236:1229-1237, not TNF-beta induces secretion of colony stimulating1987. factor for macrophages (CSF-1) by human monocytes.
10. Nioche S, Tazi A, Lecossier D, Hance AJ. Production Blood 70:1700-1703, 1987.of granulocyte colony-stimulating factor (GCSF) by 22. Thompson CB, Lindsten T, Ledbetter JA, et al. CD-28human cells: T lymphocyte-dependent and T activation pathway regulates the production oflymphocyte-independant release of GCSF by blood multiple T-cell derived lymphokines/cytokines. Procmonocytes. Eur J Immunol 18:1021-1026, 1988. Natl Acad Sci USA 86:1333-1337, 1989.
11. Kaushansky K, Miller EJ, Morris DR, Wilson CB, 23. Nishizawa M, Nagata S. Regulatory elementsHammond WP. The role of lymphocytes and responsible for inducible expression of themonocytes in hematopoietic growth factor production granulocyte colony-stimulating factor gene inby peripheral blood mononuclear cells. Cell Immunol macrophages. Mol Cell Biol 10:2002-2011, 1990.122:62-70, 1989. 24. Oster W, Lindemann A, Mertelsmann R, Herrmann F.
12. Thorens B, Mermod JJ, Vassalli P. Phagocytosis and Production of macrophage-, granulocyte-,inflammatory stimuli induce GM-CSF mRNA in granulocyte- macrophage- and multi-colony-macrophages through post-transcriptional control. Cell stimulating factor by peripheral blood cells. Eur J48:671-679, 1987. Immunol 19:543-547, 1988.
13. Chan JY, Slamon DJ, Nimer SD, Golde DW, Gasson 25. Broudy VC, Kaushansky K, Harlan JM, Adamson JW.JC. Regulation of expression of human granulocyte/ Interleukin 1 stimulates human endothelial cells tomacrophage colony-stimulating factor. Proc Natl Acad produce granulocyte-macrophage colony stimulatingSci USA 83:8669-8679, 1986. factor and granulocyte colony stimulating factor. J
14. Arai N, Naito Y, Watanabe M, Masuda ES, Immunol 139:464-468, 1987.
in T cells: Role of Cis-acting DNA elements that

Blood Cells, Molecules, and Diseases (1995) 21(20) Oct 31: 192-200 S.S. Kothari et al.
200
26. Koeffler HP, Gasson J, Tobler A. Transcriptional and fibroblasts secrete interleukin-6 and granulocyte-posttranscriptional modulation of myeloid colony- macrophage colony stimulating factor in the absencestimulating factor expression by tumor necrosis factor of inflammatory stimulation: demonstrating by serum-and other agents. Mol Cell Biol 8:3432-3438, 1988. free bioassay, enzyme-linked immunosorbent assay,
27. Guba SC, Sartor CI, Gottschalk LR, Ye-Hu J, and reverse transcriptase polymerase chain reaction.Mulligan T, Emerson SG. Bone marrow stromal Blood 80:1190-1198, 1992.




















