
Integrins and EGFR coordinately regulate the pro-apoptotic protein Bim to prevent anoikis
9
LETTERS NATURE CELL BIOLOGY VOLUME 5 | NUMBER 8 | AUGUST 2003 733 Integrins and EGFR coordinately regulate the pro-apoptotic protein Bim to prevent anoikis Mauricio J. Reginato 1 , Kenna R. Mills 1 , Jessica K. Paulus 1 , Danielle K. Lynch 1 , Dennis C. Sgroi 2 , Jayanta Debnath 1 , Senthil K. Muthuswamy 1,3 and Joan S. Brugge 1,4 Epithelial cells must adhere to the extracellular matrix (ECM) for survival, as detachment from matrix triggers apoptosis or anoikis 1 . Integrins are major mediators of adhesion between cells and ECM proteins, and transduce signals required for cell survival 2 . Recent evidence suggests that integrin receptors are coupled to growth factor receptors in the regulation of multiple biological functions 2 ; however, mechanisms involved in coordinate regulation of cell survival are poorly understood and mediators responsible for anoikis have not been well characterized. Here, we identify the pro-apoptotic protein Bim as a critical mediator of anoikis in epithelial cells. Bim is strongly induced after cell detachment and downregulation of Bim expression by RNA interference (RNAi) inhibits anoikis. Detachment-induced expression of Bim requires a lack of β 1 -integrin engagement, downregulation of EGF receptor (EGFR) expression and inhibition of Erk signalling. Overexpressed EGFR was uncoupled from integrin regulation, resulting in the maintenance of Erk activation in suspension, and a block in Bim expression and anoikis. Thus, Bim functions as a key sensor of integrin and growth factor signals to the Erk pathway, and loss of such coordinate regulation may contribute to tumour progression. Adhesion of cells to ECM proteins is important for the structural organization of tissues of multicellular organisms. Integrin-mediated regulation of cell survival is thought to contribute to maintenance of tissue homeostasis by ensuring that cells remain in their proper tissue environment. Tumour cells display significant resistance to anoikis, as most tumour-derived epithelial cells survive in the absence of adhe- sion 3 . Many oncogenes mimic integrin-dependent signals, thus allow- ing oncogene-expressing cells to survive under conditions in which normal cells undergo apoptosis 1 . Elucidation of the mechanisms underlying anoikis will provide important insights into adhesion- mediated regulation of cell death and tumour progression. The Bcl-2 family of proteins are major regulators of apoptosis 4 . Anti-apoptotic members, such as Bcl-x L and Bcl-2, contain four Bcl-2- homology (BH) domains. The pro-apoptotic members of this family fall into two categories: Bax-like proteins, such as Bax/Bak, that con- tain multiple BH domains, or BH3-only members, such as Bad, Bid and Bim 5 . The Bax-like members and the BH3-only members can both initiate apoptosis, but only the Bax-like members contain innate cytotoxic function 6 . The BH3-only members seem to function as sen- sors of cell stress and can promote apoptosis either by heterodimerizing and antagonizing the function of anti-apoptotic Bcl-2/Bcl-x L mem- bers and/or activating the function of Bax-like members 7 . As cells that overexpress the pro-survival proteins Bcl-2 and Bcl-x L are highly resistant to anoikis 8 , we investigated the mechanism through which overexpression of Bcl-x L blocks anoikis in MCF-10A immortalized mammary epithelial cells 9 .We detected high levels of apoptosis at both 48 and 72 h after detachment from matrix, as measured by a DNA fragmentation cell death ELISA (see Supplementary Information, Fig. S1a) or by live/dead staining (40% dead at 48 h, 50% at 72 h; see Supplementary Information, Fig. S1b). However, stable expression of Bcl-x L in MCF-10A cells induced a fourfold reduction of both DNA frag- mentation and ethidium-bromide-positive cells (see Supplementary Information, Fig. S1b and S1c, respectively) in cells detached from the matrix for 48 h. Furthermore, expression of a Bcl-x L mutant (Bcl-x L F131V/D133A ) that binds to BH3-only proteins, but not to Bax-like proteins 7 , rescued MCF-10A cells from anoikis (see Supplementary Information, Fig. S1c). In contrast, expression of Mt8 (Bcl-x L G138E/R139L/I140N ), which lacks functional BH3-domain and is unable to bind BH3-only proteins 7 , did not rescue cells from anoikis (see Supplementary Information, Fig. S1c). These results indicate that bind- ing of Bcl-x L to BH3-domain-only proteins is critical for the anti-apop- totic function of Bcl-x L during anoikis and that BH3-only proteins are involved in mediating anoikis. To determine whether expression of BH3-only proteins is regulated during anoikis, we analysed expression of several BH3-only proteins (Bad, Bim, Bmf and Bid) after detachment of MCF-10A cells. Of these proteins, only levels of Bim changed after loss of attachment (Fig. 1a). It has been reported that Bim RNA undergoes alternative splicing to generate three variants: BimEL, BimL and BimS 10 . Attached MCF-10A cells contain low levels of BimEL and undetectable levels of BimL and BimS. All three isoforms were highly upregulated by 24 h after detach- ment from matrix and reached maximal levels at or around 48 h in suspension (Fig. 1a). In normal cells, BimEL and BimL are thought to be sequestered in a cytoskeleton-associated motor complex and apoptotic stimuli induce 1 Department of Cell Biology, Harvard Medical School, Boston, MA 02115, USA. 2 Department of Pathology, Harvard Medical School, Massachusetts General Hospital, Boston, MA 02129, USA. 3 Present address: Cold Spring Harbor Laboratory, 1 Bungtown Road, Cold Spring Harbor, NY 11274, USA. 4 Correspondence should be addressed to J.S.B. (e-mail: [email protected]). ©2003 Nature Publishing Group
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Integrins and EGFR coordinately regulate the pro-apoptotic protein Bim to prevent anoikis

L E T T E R S
NATURE CELL BIOLOGY VOLUME 5 | NUMBER 8 | AUGUST 2003 733
Integrins and EGFR coordinately regulate the pro-apoptotic protein Bim to prevent anoikisMauricio J. Reginato1, Kenna R. Mills1, Jessica K. Paulus1, Danielle K. Lynch1, Dennis C. Sgroi2, Jayanta Debnath1,Senthil K. Muthuswamy1,3 and Joan S. Brugge1,4
Epithelial cells must adhere to the extracellular matrix (ECM)for survival, as detachment from matrix triggers apoptosis oranoikis1. Integrins are major mediators of adhesion betweencells and ECM proteins, and transduce signals required for cellsurvival2. Recent evidence suggests that integrin receptors arecoupled to growth factor receptors in the regulation of multiplebiological functions2; however, mechanisms involved incoordinate regulation of cell survival are poorly understood andmediators responsible for anoikis have not been wellcharacterized. Here, we identify the pro-apoptotic protein Bimas a critical mediator of anoikis in epithelial cells. Bim isstrongly induced after cell detachment and downregulation ofBim expression by RNA interference (RNAi) inhibits anoikis.Detachment-induced expression of Bim requires a lack of β1-integrin engagement, downregulation of EGF receptor(EGFR) expression and inhibition of Erk signalling.Overexpressed EGFR was uncoupled from integrin regulation,resulting in the maintenance of Erk activation in suspension,and a block in Bim expression and anoikis. Thus, Bimfunctions as a key sensor of integrin and growth factor signalsto the Erk pathway, and loss of such coordinate regulation maycontribute to tumour progression.
Adhesion of cells to ECM proteins is important for the structuralorganization of tissues of multicellular organisms. Integrin-mediatedregulation of cell survival is thought to contribute to maintenance oftissue homeostasis by ensuring that cells remain in their proper tissueenvironment. Tumour cells display significant resistance to anoikis, asmost tumour-derived epithelial cells survive in the absence of adhe-sion3. Many oncogenes mimic integrin-dependent signals, thus allow-ing oncogene-expressing cells to survive under conditions in whichnormal cells undergo apoptosis1. Elucidation of the mechanismsunderlying anoikis will provide important insights into adhesion-mediated regulation of cell death and tumour progression.
The Bcl-2 family of proteins are major regulators of apoptosis4.Anti-apoptotic members, such as Bcl-xL and Bcl-2, contain four Bcl-2-homology (BH) domains. The pro-apoptotic members of this familyfall into two categories: Bax-like proteins, such as Bax/Bak, that con-tain multiple BH domains, or BH3-only members, such as Bad, Bid
and Bim5. The Bax-like members and the BH3-only members canboth initiate apoptosis, but only the Bax-like members contain innatecytotoxic function6. The BH3-only members seem to function as sen-sors of cell stress and can promote apoptosis either by heterodimerizingand antagonizing the function of anti-apoptotic Bcl-2/Bcl-xL mem-bers and/or activating the function of Bax-like members7. As cells thatoverexpress the pro-survival proteins Bcl-2 and Bcl-xL are highlyresistant to anoikis8, we investigated the mechanism through whichoverexpression of Bcl-xL blocks anoikis in MCF-10A immortalizedmammary epithelial cells9. We detected high levels of apoptosis at both48 and 72 h after detachment from matrix, as measured by a DNAfragmentation cell death ELISA (see Supplementary Information, Fig.S1a) or by live/dead staining (40% dead at 48 h, 50% at 72 h; seeSupplementary Information, Fig. S1b). However, stable expression ofBcl-xL in MCF-10A cells induced a fourfold reduction of both DNA frag-mentation and ethidium-bromide-positive cells (see SupplementaryInformation, Fig. S1b and S1c, respectively) in cells detached from thematrix for 48 h. Furthermore, expression of a Bcl-xL mutant (Bcl-xL
F131V/D133A) that binds to BH3-only proteins, but not to Bax-likeproteins7, rescued MCF-10A cells from anoikis (see SupplementaryInformation, Fig. S1c). In contrast, expression of Mt8 (Bcl-xL
G138E/R139L/I140N), which lacks functional BH3-domain and isunable to bind BH3-only proteins7, did not rescue cells from anoikis (seeSupplementary Information, Fig. S1c). These results indicate that bind-ing of Bcl-xL to BH3-domain-only proteins is critical for the anti-apop-totic function of Bcl-xL during anoikis and that BH3-only proteins areinvolved in mediating anoikis.
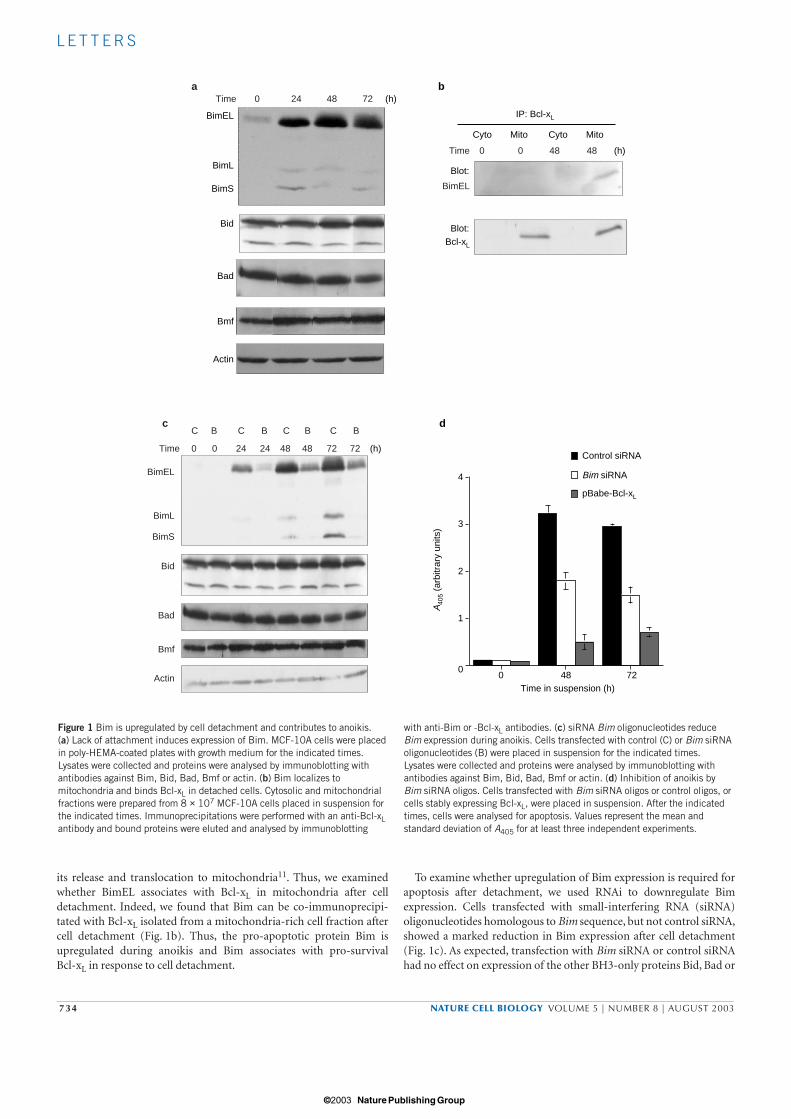
To determine whether expression of BH3-only proteins is regulatedduring anoikis, we analysed expression of several BH3-only proteins(Bad, Bim, Bmf and Bid) after detachment of MCF-10A cells. Of theseproteins, only levels of Bim changed after loss of attachment (Fig. 1a).It has been reported that Bim RNA undergoes alternative splicing togenerate three variants: BimEL, BimL and BimS10. Attached MCF-10Acells contain low levels of BimEL and undetectable levels of BimL andBimS. All three isoforms were highly upregulated by 24 h after detach-ment from matrix and reached maximal levels at or around 48 h insuspension (Fig. 1a).
In normal cells, BimEL and BimL are thought to be sequestered in acytoskeleton-associated motor complex and apoptotic stimuli induce
1Department of Cell Biology, Harvard Medical School, Boston, MA 02115, USA. 2Department of Pathology, Harvard Medical School, Massachusetts General Hospital,Boston, MA 02129, USA. 3Present address: Cold Spring Harbor Laboratory, 1 Bungtown Road, Cold Spring Harbor, NY 11274, USA. 4Correspondence should beaddressed to J.S.B. (e-mail: [email protected]).
© 2003 Nature Publishing Group
L E T T E R S
734 NATURE CELL BIOLOGY VOLUME 5 | NUMBER 8 | AUGUST 2003
its release and translocation to mitochondria11. Thus, we examinedwhether BimEL associates with Bcl-xL in mitochondria after celldetachment. Indeed, we found that Bim can be co-immunoprecipi-tated with Bcl-xL isolated from a mitochondria-rich cell fraction aftercell detachment (Fig. 1b). Thus, the pro-apoptotic protein Bim isupregulated during anoikis and Bim associates with pro-survival Bcl-xL in response to cell detachment.
To examine whether upregulation of Bim expression is required forapoptosis after detachment, we used RNAi to downregulate Bimexpression. Cells transfected with small-interfering RNA (siRNA)oligonucleotides homologous to Bim sequence, but not control siRNA,showed a marked reduction in Bim expression after cell detachment(Fig. 1c). As expected, transfection with Bim siRNA or control siRNAhad no effect on expression of the other BH3-only proteins Bid, Bad or
(h)
(h)
Actin
BimS
BimEL
BimL
Bad
Bid
Bmf
a
c
Actin
BimS
BimEL
BimL
Bad
Bid
Bmf
C B C B C B C B
0
1
2
3
4
0 48 72
pBabe-Bcl-xL
Bim siRNA
Control siRNA
dA
405
(arb
itrar
y un
its)
b0
0 0
24
24 24
48
48 48
72
72 72
Time in suspension (h)
(h) Cyto Mito Cyto Mito
Blot:
Blot:
0 0 48 48
IP: Bcl-xL
Bcl-xL
BimEL
Time
Time
Time
Figure 1 Bim is upregulated by cell detachment and contributes to anoikis. (a) Lack of attachment induces expression of Bim. MCF-10A cells were placedin poly-HEMA-coated plates with growth medium for the indicated times.Lysates were collected and proteins were analysed by immunoblotting withantibodies against Bim, Bid, Bad, Bmf or actin. (b) Bim localizes tomitochondria and binds Bcl-xL in detached cells. Cytosolic and mitochondrialfractions were prepared from 8 × 107 MCF-10A cells placed in suspension forthe indicated times. Immunoprecipitations were performed with an anti-Bcl-xLantibody and bound proteins were eluted and analysed by immunoblotting
with anti-Bim or -Bcl-xL antibodies. (c) siRNA Bim oligonucleotides reduceBim expression during anoikis. Cells transfected with control (C) or Bim siRNAoligonucleotides (B) were placed in suspension for the indicated times.Lysates were collected and proteins were analysed by immunoblotting withantibodies against Bim, Bid, Bad, Bmf or actin. (d) Inhibition of anoikis byBim siRNA oligos. Cells transfected with Bim siRNA oligos or control oligos, orcells stably expressing Bcl-xL, were placed in suspension. After the indicatedtimes, cells were analysed for apoptosis. Values represent the mean andstandard deviation of A405 for at least three independent experiments.
© 2003 Nature Publishing Group

L E T T E R S
NATURE CELL BIOLOGY VOLUME 5 | NUMBER 8 | AUGUST 2003 735
Bmf (Fig. 1c). Inhibition of Bim expression resulted in twofold protec-tion from anoikis both at 48 and 72 h after cell detachment (Fig. 1d).However, siRNA-mediated inhibition of Bim expression did not inhibitanoikis as effectively as overexpressing Bcl-xL (4–5-fold protection; Fig.1d). The inability of Bim siRNA to inhibit apoptosis to the same extentas Bcl-xL could be caused by the residual level of Bim protein in the BimsiRNA-transduced cells or involvement of other BH3-only proteins inanoikis. Among BH3-only proteins, the recently identified protein Bmfis implicated in anoikis, as it localizes to mitochondria and binds Bcl-2
after cell detachment in the mammary cancer cell line MCF-7 (ref. 12);however, its contribution to anoikis has not been examined.Nevertheless, inhibition of Bim expression during cell detachment ren-ders cells more resistant to anoikis, suggesting that upregulation of Bimexpression during detachment contributes to cell death during anoikis.
To investigate directly whether Bim is regulated by ECM signals, weincubated single cells in suspension in the presence of exogenous ECM(Matrigel) and monitored Bim expression. Under these conditions,Bim upregulation was blocked in suspension (Fig. 2a). Moreover,
Actin
+ IgG
BimEL
– Ins
– Ins
– Ins
a
b
Actin
BimEL
c
EGFR
ErbB2
pErk
BimEL
Erk2
pAkt
Akt
Matrix
BimEL
EGFR
Actin
MCF-10A MMEC MDCK IEC-18
d
EGFR
0 48 48 48 48 (h) (h)
– – + + +
+ β1
Con – EGF – EGF
– EGF
– Serum
0 24 48 72
A S A S A S A S
Time Time
Figure 2 Regulation of Bim by integrins and EGFR signalling. (a) Regulation of Bim by β1 integrins. Cells were placed in suspension forthe indicated times in the presence or absence of 5% Matrigel (Matrix),anti-integrin-β1 blocking antibody13 (100 µg ml−1) or IgG control antibody(100 µg ml−1). Lysates were collected and proteins were analysed byimmunoblotting with anti-Bim, -EGFR and -actin antibodies. (b)Withdrawal of EGF in attached cells upregulates expression of Bim.Attached cells were incubated for 24 h in growth medium in the absenceof EGF, insulin or serum, as indicated. Lysates were collected and proteinswere analysed by immunoblotting with antibodies against Bim or actin.
(c) Downregulation of EGFR levels during anoikis. Cells were placed insuspension for the indicated times. Lysates were collected and proteinswere analysed by immunoblotting with anti-EGFR, -ErbB2, -Bim, -phospho-Erk, -Erk2, -phospho-Akt or -Akt antibodies. (d) Loss of EGFRexpression and induction of Bim in multiple epithelial cells. Cell lysateswere collected from MCF-10A, primary mouse mammary epithelial cells(MMEC), and MDCK cells either attached (A) or in suspension for 24 h(S). IEC-18 cells were placed in suspension for 12 h, as they undergorapid apoptosis after detachment17. Proteins were analysed byimmunoblotting with anti-EGFR, -Bim and -actin antibodies.
© 2003 Nature Publishing Group
L E T T E R S
736 NATURE CELL BIOLOGY VOLUME 5 | NUMBER 8 | AUGUST 2003
addition of a blocking antibody to β1 integrin13 restored upregulationof Bim expression in cells cultured in suspension with exogenous ECM(Fig. 2a). Thus, Bim is regulated by ECM signals through β1 integrinreceptors.
Integrins and growth factors activate multiple common downstreamsignalling pathways that can regulate cell survival, including the phos-phatidylinositol 3-OH kinase (PI(3)K) pathway and its target proteinkinase B (PKB)/Akt14,15, as well as the Erk/mitogen-activated proteinkinase (MAPK) pathway16. Therefore, we also examined if growth fac-tors present in MCF-10A medium could regulate Bim expression.Withdrawal of EGF, but not insulin, from attached MCF-10A cells
increased expression of Bim (Fig. 2b), suggesting that engagement ofEGFRs, as well as β1 integrins, can inhibit expression of Bim.
As EGF can suppress expression of Bim, we investigated why Bimprotein was still upregulated during cell detachment, despite the pres-ence of EGF. To determine if cell detachment disrupts signalling initi-ated by EGF, we examined expression of EGFR in suspended cells.Indeed, EGFR expression was markedly reduced by 24 h after celldetachment and co-incident with upregulation of BimEL expression(Fig. 2c). Loss of EGFR expression was specific, as expression of ErbB2(Fig. 2c) and the c-Met receptors (Fig. 3a) were unaffected. In addi-tion, activation of downstream signalling proteins regulated by EGF
(h)
(h)
pLPCX EGFR
BimEL
c-Met
EGFR
Actin
0
0.5
1
1.5
2
2.5
b
d
BimEL
Actin
AP1510
BimEL
Actin
0
10
20
30
40
50
– Matrix + Matrix
a c
pErk
0
0 0
8 24 48 0 8 24 48
48 48 48
ErbB
2
pBab
e
pBab
e
ErbB
2
ErbB
2
– + + – +
A40
5 (a
rbitr
ary
units
)
Cel
l dea
th(p
erce
ntag
e)
Time
Time
Time in suspensionTime in suspension
0 48 0 480
484848 48 48 48
(h)(h)
pLPC
XpL
PCX
EGFR
EGFR
ErbB
2 –
APEr
bB2
+ A
P
Con
trol
Con
trol
Con
trol
– EG
F
+ AG
1478
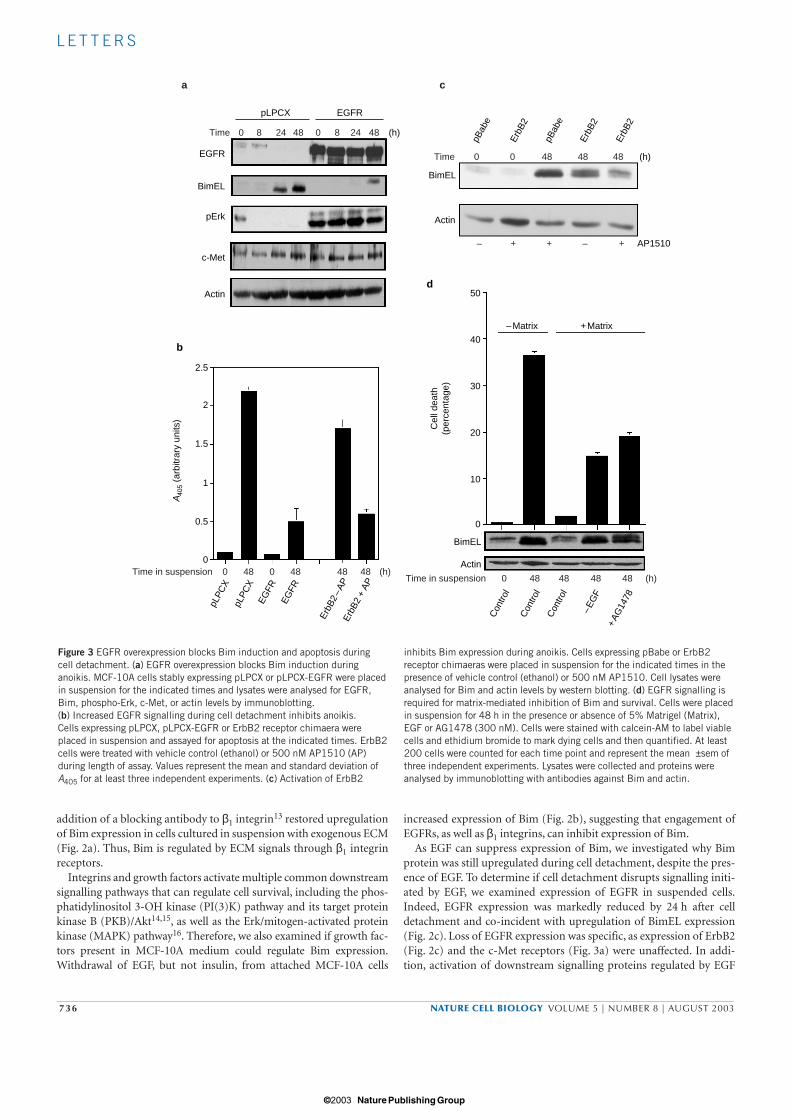
Figure 3 EGFR overexpression blocks Bim induction and apoptosis duringcell detachment. (a) EGFR overexpression blocks Bim induction duringanoikis. MCF-10A cells stably expressing pLPCX or pLPCX-EGFR were placedin suspension for the indicated times and lysates were analysed for EGFR,Bim, phospho-Erk, c-Met, or actin levels by immunoblotting.(b) Increased EGFR signalling during cell detachment inhibits anoikis. Cells expressing pLPCX, pLPCX-EGFR or ErbB2 receptor chimaera wereplaced in suspension and assayed for apoptosis at the indicated times. ErbB2cells were treated with vehicle control (ethanol) or 500 nM AP1510 (AP)during length of assay. Values represent the mean and standard deviation ofA405 for at least three independent experiments. (c) Activation of ErbB2
inhibits Bim expression during anoikis. Cells expressing pBabe or ErbB2receptor chimaeras were placed in suspension for the indicated times in thepresence of vehicle control (ethanol) or 500 nM AP1510. Cell lysates wereanalysed for Bim and actin levels by western blotting. (d) EGFR signalling isrequired for matrix-mediated inhibition of Bim and survival. Cells were placedin suspension for 48 h in the presence or absence of 5% Matrigel (Matrix),EGF or AG1478 (300 nM). Cells were stained with calcein-AM to label viablecells and ethidium bromide to mark dying cells and then quantified. At least200 cells were counted for each time point and represent the mean ±sem ofthree independent experiments. Lysates were collected and proteins wereanalysed by immunoblotting with antibodies against Bim and actin.
© 2003 Nature Publishing Group

L E T T E R S
NATURE CELL BIOLOGY VOLUME 5 | NUMBER 8 | AUGUST 2003 737
Actin
BimEL
Control PD LY PP2
pErk
a
pBabe
BimEL
Actin
pErk
c
Control PD LYControl Control PD LYPP2 PP2
pBab
e
pBab
e
Ras
G12
V
MEK
2-D
D
Raf
-CAA
X
BimEL
Actin
pErk
Erk2
b
0
1
2
3
d e
0
2
4
6
8
0 48 48 48 48 (h)
(h)
RasG12V
0 24 24 24 24 24 24 24 24
A40
5 (a
rbitr
ary
units
)
Rel
ativ
e B
im m
RN
A e
xpre
ssio
n(a
rbitr
ary
units
)
Time
pBab
epB
abe
Ras
G12
VR
af-C
AAX
Mek
2-D
D
MER
-Akt
– T
MER
-Akt
+ T
Time in suspension Time in suspension0 0 24 0 2448 48 48 48 48 48
pBab
e
pBab
e
MEK
2-D
D
MEK
2-D
D
Time
(h) (h)
Figure 4 Erk signalling regulates Bim expression and anoikis. (a) Bimexpression is induced in attached cells treated with Mek inhibitor. Attachedcells were incubated for 24 h in the presence of vehicle control (DMSO),PD98069 (40 µM), LY294002 (50 µM) or PP2 (10 µM), as indicated.Lysates were collected and proteins were analysed by immunoblotting withanti-Bim, -phospho-Erk or -actin antibodies. (b) Activation of the MAPKpathway blocks Bim induction during anoikis. Cells were transfected asindicated and placed in suspension for the indicated times. Lysates werecollected and proteins were analysed by immunoblotting with anti-Bim, -phospho-Erk, -Erk2 or -actin antibodies. (c) Ras-mediated downregulation ofBim during detachment is dependent on MAPK activity. Cells expressingpBabe or pBabe-H-RasV12 were placed in suspension in the presence ofvehicle control (DMSO), PD98069 (40 µM), LY294002 (50 µM) or PP2
(10 µM), as indicated. Lysates were collected and proteins were analysed byimmunoblotting with anti-Bim, -phospho-Erk or –actin antibodies. (d) ActiveRas, Raf, Mek and Akt inhibit anoikis. Cells were transfected as indicatedand placed in suspension and assayed for apoptosis at the indicated times.Myr-Akt-ER cells were treated with vehicle control (ethanol; −T) or tamoxifen(1 µM; +T) during the length of assay. Values represent the mean andstandard deviation of A405 for at least three independent experiments. (e) The MAPK pathway regulates Bim mRNA levels in MCF-10A cells. TotalRNA was collected from cells transfected with pBabe or pBabe-MEK2-DDafter 0 or 24 h in suspension and used to assay Bim transcripts using real-time RT–PCR. Data are expressed as relative Bim expression normalized totransfection of pBabe at time zero from an average of three independentexperiments.
© 2003 Nature Publishing Group

L E T T E R S
738 NATURE CELL BIOLOGY VOLUME 5 | NUMBER 8 | AUGUST 2003
and integrins, such as Erk (Figs 2c and 3a) and Akt (Fig. 2c), was alsodecreased during anoikis. Expression of EGFR was dependent onmatrix signals, in particular β1 integrin signals, as cells in suspensionincubated with exogenous ECM maintained EGFR levels (Fig. 2a) andEGFR was lost when cells suspended in Matrigel were incubated withβ1 integrin blocking antibody (Fig. 2a). Thus, EGFR expression is reg-ulated by matrix attachment through β1 integrin receptors and isreduced during anoikis in MCF-10A cells. We observed a similardecrease of EGFR levels and induction of Bim during anoikis in multi-ple types of epithelial cells, including primary mouse mammaryepithelial cells as well as two epithelial cell lines frequently used tostudy anoikis: MDCK (dog kidney epithelial)14 and IEC-18 (rat intes-tinal epithelial)17 cells (Fig. 2d). Thus, loss of EGFR and induction ofBim is a general consequence of cell detachment in a number ofepithelial cell lines. The mechanism responsible for loss of EGFR afterdetachment in epithelial cells is not understood. In mouse embryofibroblasts, detachment from matrix results in loss of platelet-derivedgrowth factor receptor β expression18 involving a ubiquitin-dependentdegradation pathway.
To investigate the effects of ectopic expression of elevated levels ofEGFR on Bim upregulation during anoikis, we stably overexpressedEGFR in MCF-10A cells. In contrast to endogenous EGFR, overex-pressed EGFR was not downregulated in suspension. Importantly,overexpression of EGFR blocked upregulation of Bim expression aftercell detachment (Fig. 3a) and prevented anoikis (Fig. 3b).Downstream signalling, specifically activation of Erk, was also main-tained in EGFR-overexpressing cells after detachment (Fig. 3a). Thus,overexpressed EGFR can escape downregulation caused by loss of inte-grin engagement and allow cell survival from anoikis. As many cancer-derived cell lines possess the ability to survive in suspension, weexamined whether a tumour-derived cell line could maintain EGFRexpression after detachment from matrix. The breast cancer cell lineMDA-MB-231 is highly resistant to anoikis (data not shown) and con-tains high levels of EGFR, compared with MCF-10A cells. EGFR wasnot downregulated and Bim was not upregulated when MDA-MB-231cells were detached from matrix (see Supplementary Information,Fig. S2), suggesting that this coordinate integrin/EGFR regulation canbe lost in transformed cells. EGFR-mediated inhibition of Bim expres-sion during anoikis may be critical for cell survival, as exogenousexpression of Bim in MCF-10A cells overexpressing EGFR (as well asMDA-MB-231 cells) induces high levels of apoptosis in attached cells(data not shown). Similar regulation of EGFR expression by integrinligation was observed in a transformed mammary epithelial cell line(T4 cells), in which blocking β1 integrin engagement reduced EGFRexpression19. Downregulation of EGFR by inhibition of β1 integrin lig-and binding was associated with reversion of the tumour phenotype,suggesting that this crosstalk has important functional consequences.These results indicate that the coordinate regulation of matrix attach-ment and EGFR expression may be lost during tumorigenesis andcould contribute to anchorage independence.
We further investigated whether signals from other EGFR familymembers could also inhibit Bim expression. MCF-10A cellsexpressing a chimaeric ErbB2–FKBP (FK-506-binding protein)receptor can be activated by homodimerizing with the FKBP-dimerizing small molecule AP1510 (ref. 20). Activation of ErbB2homodimers in MCF-10A cells inhibited Bim expression duringcell detachment (Fig. 3c). Consistent with inhibition of Bim induc-tion, activation of ErbB2 blocked anoikis (Fig. 3b). Thus, unregu-lated expression of EGFR or activation of ErbB2 signallingpathways is sufficient to inhibit Bim expression and confer resist-ance to anoikis.
The above gain-of-function experiments showed that maintenanceof high EGFR levels is sufficient to inhibit Bim expression and anoikis.To examine whether EGFR activation is required for matrix-mediatedinhibition of Bim and anoikis, suspended cells were incubated in thepresence of exogenous matrix and EGFR signalling was blocked by cul-turing cells in the absence of EGF or addition of a specific EGFR kinaseinhibitor AG1478. Inhibition of Bim expression by exogenous matrixwas reversed by withdrawal of EGF from the media or addition ofAG1478 (Fig. 3d). Consistent with Bim induction under these condi-tions, loss of EGFR signalling reduced cell survival by exogenousmatrix (Fig. 3d). Thus, adhesion to matrix is not sufficient to inhibitBim expression, as signals from EGFR also contribute to matrix-medi-ated cell survival in epithelial cells.
To characterize integrin- and EGFR-stimulated pathways that regu-late Bim expression, we examined the effects of inhibiting severalkinases that regulate cell survival on Bim expression in attached cells.Incubation of MCF-10A cells for 24 h with a MEK inhibitor(PD98059), but not a PI(3)K (LY294002) or Src inhibitor (PP2),induced upregulation of Bim (Fig. 4a), suggesting that signallingdownstream of Mek regulates Bim expression. Thus, in attached cells,it is probable that integrin and EGFR receptors cooperate to inhibitBim expression through the Mek/Erk pathway.
To determine whether activation of the Erk pathway is sufficient toregulate Bim expression in suspension, we expressed activated variantsof Ras, Raf and Mek, several protein components of the Erk signallingpathway. Expression of H-RasG12V, Raf-CAAX or MEK2-DD com-pletely inhibited expression of Bim in suspension (Fig. 4b). As onco-genic Ras can activate both Erk and PI(3)K pathways, we examinedinhibitors that would block either pathway. Indeed, the ability ofH-RasV12 to block Bim expression was lost when cells were incubatedwith PD98059, but not with LY294002 or PP2 (Fig. 4c). Thus, multipleactivators of the Erk pathway can inhibit Bim expression induced bycell detachment.
As Bim can be regulated through an Akt-Forkhead transcriptionfactor pathway by cytokine withdrawal in haematopoietic cells21, weexamined whether Akt could regulate Bim expression during anoikisin MCF-10A cells. Cells expressing a myristoylated variant of the Akt1kinase domain fused to the oestrogen receptor (Myr-Akt–ER)22 weretreated with 4-hydroxytamoxifen to activate Akt kinase activity.Although this treatment resulted in phosphorylation of Akt kinasesubstrates in suspension, upregulation of Bim expression was unal-tered (see Supplementary Information, Fig. S3).
We also examined whether suppression of Bim expression by theErk pathway correlated with inhibition of anoikis. H-RasG12V, Raf-CAAX and MEK2-DD were very effective in protecting cells fromanoikis (Fig. 4d). Activation of Akt in MCF-10A cells also inhibitedanoikis (Fig. 4d). As activated Akt had no effect on Bim expression,these data suggest that Akt-mediated protection from apoptosis isindependent of Bim regulation, possibly involving other BH3-onlyproteins (for example, Bad23) or events that block Bim-induced apop-tosis, such as induction of pro-survival members of the Bcl-2 family24
or caspase inactivation25. Nevertheless, expression of oncogenes suchas RasV12, Raf, Mek2 and ErbB2 that can activate the Erk pathway andprotect epithelial cells from anoikis also prevented upregulation ofBim expression. Taken together, our observations establish that Bim iscritical for regulation of apoptosis in detached epithelial cells and isalso a downstream target of many oncogenes that promote anchorage-independent survival/proliferation of epithelial cells.
To determine how expression of Bim is inhibited by the MAPKpathway, we examined whether Bim mRNA is regulated during anoikisand whether the MAPK pathway is involved. Using real-time RT–PCR
© 2003 Nature Publishing Group

L E T T E R S
NATURE CELL BIOLOGY VOLUME 5 | NUMBER 8 | AUGUST 2003 739
to examine transcription of Bim mRNA during anoikis, we found thatBim transcripts were upregulated sevenfold 24 h after cell detachmentand that this upregulation was inhibited in cells expressing MEK2-DD(Fig. 4e). The Mek/Erk pathway has been implicated in regulating cellsurvival in a number of cell types. Previous studies using an oestrogen-inducible, activated form of Raf in MCF-10A (ref. 16) and MDCK26
cell lines also indicated that activation of Raf results in protection fromanoikis. In the MCF-10A study, the protective effect of activated Rafwas reported to be caused by production of EGF ligands16. However,our studies indicate that EGF itself provides only a short-term protec-tive effect and Raf induces a strong survival effect in the presence ofEGF in MCF-10A cells. In keratinocytes, activation of EGFR duringcell detachment results in Erk activation and protection fromanoikis27. The Mek/Erk pathway has been implicated in regulating Bimexpression in haematopoietic cells28, neurons29 and fibroblasts30. Inaddition, recent studies in neurons showed that BimEL can be phos-phorylated by an Mek/Erk-dependent pathway29. Furthermore, infibroblasts, phosphorylation may cause increased degradation ofBimEL31. We have also observed phosphorylation of BimEL by aMek/Erk pathway in MCF-10A cells (M.J.R. and J.S.B., unpublishedobservations), suggesting that the Mek/Erk pathway not only regulatesBim at the expression level, but also directly regulates its apoptoticfunction in epithelial cells. Taken together, our results suggest an intri-cate coupling between integrin and EGFRs in regulating cell survivalspecifically through modulation of Bim expression at the mRNA levelthrough the Mek/Erk pathway.
These studies provide a molecular mechanism of how ECM regu-lates cell survival through regulation of the BH3-only protein Bim. Inaddition, they provide a clear example of coordinate crosstalk betweenintegrin and EGFR signalling in regulating a specific apoptotic target.They highlight the intricate coupling of integrin and EGFR signallingin regulating both Bim expression and anoikis through the Mek/Erkpathway. Resistance to anoikis in tumour cells may be caused by loss ofnormal integrin and growth factor coupling, allowing inappropriatesurvival signals to be maintained in the absence of adhesion.Moreover, as pro-survival Bcl-2 members have been implicated intumorigenesis32 it is probable that loss of expression or function ofBH3-only proteins, such as Bim, could have a similar role in escapefrom apoptotic stress during tumour development.
METHODSCell culture and materials. MCF-10A, MDCK, IEC-18 and MDA-MB-231 cellswere obtained from American type culture collection (Manassas, VA). MCF-10A cells were maintained in DMEM/F12 (Invitrogen, Carlsbad, CA) supple-mented with 5% donor horse serum, 20 ng ml−1 EGF (Peprotech, Rocky Hill,NJ), 10 µg ml−1 insulin (Sigma, St Louis, MO), 1 ng ml−1 cholera toxin (Sigma),100 µg ml−1 hydrocortisone (Sigma), 50 U ml−1 penicillin, and 50 µg ml−1
streptomycin (Invitrogen). MDCK cells were maintained in DMEM(Invitrogen) supplemented with 10% foetal calf serum (FCS), 50 U ml−1
penicillin and 50 µg ml−1 streptomycin. IEC-18 cells were maintained inDMEM supplemented with 5% FCS, 4 µg ml−1 insulin, 50 U ml−1 penicillin,and 50 µg ml−1 streptomycin. MDA-MB-231 cells were maintained in DMEMsupplemented with 10% FCS, 50 U ml−1 penicillin, and 50 µg ml−1 strepto-mycin. Primary cultures of mouse mammary epithelium were prepared by lim-ited collagenase digestion as described33. Briefly, inguinal mammary glandsfrom adult female Balb/c mice were dissected, washed and subjected to multiplerounds of collagenase digestion in Hepes-buffered RPMI/2.5% FCS supple-mented with 1 mg ml−1 collagenase Blend L (Sigma) for 1 h at 37 °C. Pooledcell pellets containing epithelial organoids were filtred through a 400-mm poly-ester mesh (PE400; Clarcon, Warrington, UK), washed and allowed to adhere inprimary culture medium (Hams F12/DMEM supplemented with 10% FCS,100 ng ml−1 hydrocortisone, 5 µg ml−1 insulin, 10 ng ml−1 EGF and 5 ng ml−1
cholera toxin). Poly-HEMA, methylcellulose and 4-hydroxy-tamoxifen were
purchased from Sigma. PD98059, LY294002 and PP2 were purchased fromCalbiochem (San Diego, CA). AP1510 was obtained from AriadPharmaceuticals (Cambridge, MA). Growth-factor-reduced Matrigel (BDBiosciences, San Diego, CA) lots with protein concentrations between 9 and11 mg ml−1 were used. Antibodies: anti-Bim from Stressgen (Victoria, BritishColumbia, Canada); anti-actin, anti-c-Met, anti-Erk2 and anti-Bad from SantaCruz (Santa Cruz, CA); anti-Bcl-xL and anti-Bid from BD Biosciences, anti-Bmf from Alexis (San Diego, CA); anti-phospho Erk from BioSource(Camarillo, CA); anti-EGFR from Transduction Labs (Lexington, KY); anti-ErbB2 from Oncogene Research (Boston, MA); anti-phosphoAkt and anti-phosphoAkt substrate from Cell Signalling (Beverly, MA). M. Bissell (LawrenceBerkeley National Laboratory) and C. Damsky (UCSF) kindly provided AIIB2,the β1-integrin function-blocking antibody13.
Expression vectors and cell lines. pMig, pMig-Bcl-xL, pMig-Bcl-xL (Mt1) andpMig-Bcl-xL (Mt8) were provided by S. Korsmeyer (Dana-Farber CancerInstitute) and have been described7. O. Gozani and J. Yuan provided pBabe-Bcl-xL. pWZL-MER-Akt was a generous gift from R. Roth (Stanford University)22.The construct pBabe-H-RasV12 was created by cloning the H-RasV12 cDNAinsert from pCEFLAU5-H-RasV12 vector (provided by S. Gutkind, NationalInstitutes of Health) into the BamHI and EcoRI sites of pBabe-puro. The con-struct pLPCX-EGFR was created by cloning the EGFR cDNA insert frompMLVneo-EGFR (provided by A. Wells, University of Pittsburgh) into the XhoIand HindIII site of pLPCX. The construct pBabe-Raf-CAAX and pBabe-MEK2-DD were provided by R. Weinberg (Whitehead Institute) and S. Meloche(Institut Recherches Clinique de Montreal), respectively. All stable cell lines,except those expressing H-RasV12, were generated by retroviral infection ofMCF-10A cells followed with treatment with appropriate selectable marker.VSV-pseudotyped retroviruses were produced by transfection of the VSV-GPGproducer cell line34 (provided by R. Mulligan, Children’s Hospital, HarvardMedical School) with 15 µg of DNA using Lipofectamine 2000 (Invitrogen).Stable RasV12-expressing cells were made by transfecting MCF-10A cellswith10 µg pBabe-RasV12 cDNA using Lipofectamine 2000. Stable MCF-10Acells containing p75.B2, a chimeric ErbB2 receptor fused to the bivalent ligandFKBP, and its activation through dimerization with the bivalent ligand AP1510have been reported previously20.
Detachment-induced apoptosis assay. MCF-10A cells were cultured inmedium containing multiple supplements that may provide survival signals toinfluence anoikis: for example, 5% horse serum, cholera toxin, insulin, EGF andhydrocortisone. Therefore, we performed all anoikis assays in growth mediumcontaining all supplements to ensure that the observed cell death was caused bylack of attachment alone and not deprivation of critical medium components.Primary MEC, MDCK, IEC-18 and MDA-MB-231 cells were placed in suspen-sion in their respective medium. Tissue culture plates were coated with Poly-HEMA (6 mg ml−1), incubated at 37 °C until dry and washed with PBS beforeuse. Cells were placed in growth medium in the presence of 0.5% methylcellu-lose (to avoid survival effects caused by clumping of cells) in suspension at adensity of 400,000 cells ml−1 and plated on poly-HEMA-coated 35-mm platesfor the indicated time. Cells were then washed with PBS and counted. 25,000cells were used to determine apoptosis with the cell death detection ELISA kit(Roche Diagnostics, Mannheim, Germany) according to the manufacturer’sinstructions. Each time point was performed in duplicate and each experimentwas carried out at least three times. To quantify percentage cell death, cells incu-bated in suspension in 0.5% methycellulose for the indicated times were stainedfor 15–30 min at 37 °C with a mixture of 1 µM ethidium bromide E and 5 µMcalcein-AM (Molecular Probes, Eugene, OR) in PBS. Indirect immunofluores-cence was performed on a Nikon TE300 microscope.
Western blotting and immunoprecipitation. Cell lysates from 2–5 × 106 MCF-10A cells either attached or in suspension for the indicated times were preparedin NP40 lysis buffer (50 mM Tris-HCl at pH 7.6, 150 mM NaCl, 10 mM NaF, 1%NP-40, 10% glycerol and 1mM Na3VO4) or RIPA buffer (150 mM NaCl, 1%NP40, 0.5% DOC, 50 mM TrisHCL at pH 8, 0.1% SDS, 10% glycerol, 5 mMEDTA, 20 mM NaF and 1 mM Na3VO4) supplemented with 1 µg ml−1 each ofpepstatin, leupeptin, aprotinin, and 200 µg ml−1 phenyl methylsulphonyl fluo-ride (PMSF). Lysates were cleared by centrifugation at 16,000g for 20 min at 4 °C
© 2003 Nature Publishing Group

L E T T E R S
740 NATURE CELL BIOLOGY VOLUME 5 | NUMBER 8 | AUGUST 2003
and analysed by SDS–PAGE and autoradiography. For immunoprecipitation,cytosolic and mitochondrial fractions were prepared using themitochondrial/cytosol fractionation kit (Alexis Biochemicals) according to themanufacturer’s instructions from 8 × 107 MCF-10A cells attached or after sus-pension for 48 h. Cytosol and mitochondrial lysates were incubated with 1 µg ofmouse anti-human Bcl-xL antibody (Chemicon, Temecula, CA) and ProteinA–Sepharose beads overnight at 4 °C. Beads were washed with NP-40 lysis bufferand boiled in SDS sample buffer. Proteins were analysed by western blottingusing primary antibodies to Bim and Bcl-xL and secondary Protein A–HRP anti-body (Zymed, San Francisco, CA).
RNAi. MCF-10A cells were plated onto 6-well plates at 300,000 cells per well.After 24 h, cells were transfected with double-stranded RNA–DNA hybrids at afinal concentration of 1 µg annealed oligo using Oligofectamine (Invitrogen)according to manufacturer’s instructions. After 24 h of transfection, cells wereplaced in anoikis assays or processed for SDS–PAGE analysis at the indicatedtimes. Oligonucleotides were obtained from Dharmacon Research (Lafayette,CO). Control sense 5′-(GGCUGUAACUUACGUGUACUU)d(TT)-3′; controlantisense, 5′-(AAGUACACGUAAGUUACAGCC)d(TT)-3′; Bim sense 5′-(GACCGAGAAGGUAGACAAUUG)d(TT)-3′; Bim antisense 5′(CAAU-UGUCUACCUUCUCGGUC)d(TT)-3′.
Real-time quantitative PCR. Total RNA was extracted from cells using RNASTAT-60 according to manufacturer’s instructions (Tel-Test). Equal amounts oftotal RNA (2 µg) was used as template for first-strand synthesis with oligo dTprimers (SuperScript first-strand synthesis system for RT–PCR; Invitrogen) ina 20-µl volume and the resulting first-strand cDNA was diluted and used astemplate in the RTQ-PCR analysis. Analysis of gene expression was performedwith an ABI 7900HT sequence detection system (Applied Biosystems, FosterCity, CA) using the relative standard curve method, as described35. Reactionsfor each sample were performed in triplicate using equal amounts of templatecDNA, and each triplicate performed in duplicate. The expression of the house-keeping gene, cyclophilin 33A, was used to normalize for variances in inputcDNA. The probe and primer pairs were designed using the ABI Primer Expresssoftware and the sequences of the PCR primer pairs and fluorogenic probe,respectively, are as follows: BIM: 5′-TGCAGACATTTTGCTTGTTCAA-3′,5′-GAACCGCTGGCTGCATAATAAT-3′, 5′-VIC-CCAACAAGACCCAGCACCGCG-TAMRA-3′; cyclophilin 33, 5′-GCTGCCTGTGCACTCATGAA-3′,5′-CAGTGCCATTG TGGTTTGTGA-3′, and 5′-6FAM-ATCACCGCCCTG-GCACATGAACTG-TAMRA-3′.
Note: Supplementary Information is available on the Nature Cell Biology website.
ACKNOWLEDGEMENTSWe would like to thank O. Gozani, R. Greenberg and members of the Brugge lab forstimulating discussions. We would like to thank Ariad Pharmaceuticals forproviding AP1510 (www.ariad.com/regulationkits). This work was supported bygrants from National Cancer Institute, Aventis Pharmaceuticals and the AmericanCancer Society (to J.S.B.); a Susan Komen Breast Cancer Postdoctoral Fellowship(to M.J.R.); an HHMI Physician Postdoctoral Fellowship (to J.D.); a MassachusettsBreast Cancer Research Award (to S.K.M.).
COMPETING FINANCIAL INTERESTSThe authors declare that they have no competing financial interests.
Received 16 January 2003; Accepted 22 JUNE 2003;Published online 6 June 2003; DOI: 10.1038/ncb1026.
1. Frisch, S. M. & Screaton, R. A. Anoikis mechanisms. Curr. Opin. Cell Biol. 13,555–562 (2001).
2. Miranti, C. K. & Brugge, J. S. Sensing the environment: a historical perspective onintegrin signal transduction. Nature Cell Biol. 4, E83–E90 (2002).
3. Schwartz, M. A. Integrins, oncogenes, and anchorage independence. J. Cell Biol.139, 575–578 (1997).
4. Cory, S. & Adams, J. M. The Bcl2 family: regulators of the cellular life-or-deathswitch. Nature Rev. Cancer 2, 647–656 (2002).
5. Puthalakath, H. & Strasser, A. Keeping killers on a tight leash: transcriptional andpost-translational control of the pro-apoptotic activity of BH3-only proteins. CellDeath Differ. 9, 505–512 (2002).
6. Zong, W. X., Lindsten, T., Ross, A. J., MacGregor, G. R. & Thompson, C. B. BH3-only
proteins that bind pro-survival Bcl-2 family members fail to induce apoptosis in theabsence of Bax and Bak. Genes Dev. 15, 1481–1486 (2001).
7. Cheng, E. H. et al. BCL-2, BCL-X(L) sequester BH3 domain-only molecules prevent-ing BAX- and BAK-mediated mitochondrial apoptosis. Mol. Cell 8, 705–711 (2001).
8. Rytomaa, M., Martins, L. M. & Downward, J. Involvement of FADD and caspase-8signalling in detachment-induced apoptosis. Curr. Biol. 9, 1043–1046 (1999).
9. Soule, H. D. et al. Isolation and characterization of a spontaneously immortalizedhuman breast epithelial cell line, MCF-10. Cancer Res. 50, 6075–6086 (1990).
10. O’Connor, L. et al. Bim: a novel member of the Bcl-2 family that promotes apoptosis.EMBO J. 17, 384–395 (1998).
11. Puthalakath, H., Huang, D. C., O’Reilly, L. A., King, S. M. & Strasser, A. Theproapoptotic activity of the Bcl-2 family member Bim is regulated by interaction withthe dynein motor complex. Mol. Cell 3, 287–296 (1999).
12. Puthalakath, H. et al. Bmf: a proapoptotic BH3-only protein regulated by interactionwith the myosin V actin motor complex, activated by anoikis. Science 293,1829–1832 (2001).
13. Howlett, A. R., Bailey, N., Damsky, C., Petersen, O. W. & Bissell, M. J. Cellulargrowth and survival are mediated by β1 integrins in normal human breast epitheliumbut not in breast carcinoma. J. Cell Sci. 108, 1945–1957 (1995).
14. Khwaja, A., Rodriguez-Viciana, P., Wennstrom, S., Warne, P. H. & Downward, J.Matrix adhesion and Ras transformation both activate a phosphoinositide 3-OHkinase and protein kinase B/Akt cellular survival pathway. EMBO J. 16, 2783–2793(1997).
15. King, W. G., Mattaliano, M. D., Chan, T. O., Tsichlis, P. N. & Brugge, J. S.Phosphatidylinositol 3-kinase is required for integrin-stimulated AKT and Raf-1/mitogen-activated protein kinase pathway activation. Mol. Cell. Biol. 17,4406–4418 (1997).
16. Schulze, A., Lehmann, K., Jefferies, H. B., McMahon, M. & Downward, J. Analysis ofthe transcriptional program induced by Raf in epithelial cells. Genes Dev. 15,981–994 (2001).
17. Rosen, K. et al. Downregulation of the pro-apoptotic protein Bak is required for theras-induced transformation of intestinal epithelial cells. Curr. Biol. 8, 1331–1334(1998).
18. Baron, V. & Schwartz, M. Cell adhesion regulates ubiquitin-mediated degradation ofthe platelet-derived growth factor receptor β. J. Biol. Chem. 275, 39318–39323(2000).
19. Wang, F. et al. Reciprocal interactions between β1-integrin and epidermal growthfactor receptor in three-dimensional basement membrane breast cultures: a differentperspective in epithelial biology. Proc. Natl Acad. Sci. USA 95, 14821–14826(1998).
20. Muthuswamy, S. K., Li, D., Lelievre, S., Bissell, M. J. & Brugge, J. S. ErbB2, but notErbB1, reinitiates proliferation and induces luminal repopulation in epithelial acini.Nature Cell Biol. 3, 785–792 (2001).
21. Dijkers, P. F. et al. FKHR-L1 can act as a critical effector of cell death induced bycytokine withdrawal: protein kinase B-enhanced cell survival through maintenance ofmitochondrial integrity. J. Cell Biol. 156, 531–542 (2002).
22. Kohn, A. D. et al. Construction and characterization of a conditionally active versionof the serine/threonine kinase Akt. J. Biol. Chem. 273, 11937–11943 (1998).
23. Datta, S. R. et al. Akt phosphorylation of BAD couples survival signals to the cell-intrinsic death machinery. Cell 91, 231–241 (1997).
24. Leverrier, Y. et al. Role of PI3-kinase in Bcl-X induction and apoptosis inhibitionmediated by IL-3 or IGF-1 in Baf-3 cells. Cell Death Differ. 6, 290–296 (1999).
25. Cardone, M. H. et al. Regulation of cell death protease caspase-9 by phosphoryla-tion. Science 282, 1318–1321 (1998).
26. Le Gall, M. et al. The p42/p44 MAP kinase pathway prevents apoptosis induced byanchorage and serum removal. Mol. Biol. Cell 11, 1103–1112 (2000).
27. Jost, M., Huggett, T. M., Kari, C. & Rodeck, U. Matrix-independent survival of humankeratinocytes through an EGF receptor/MAPK-kinase-dependent pathway. Mol. Biol.Cell 12, 1519–1527 (2001).
28. Shinjyo, T. et al. Downregulation of Bim, a proapoptotic relative of Bcl-2, is a pivotalstep in cytokine-initiated survival signaling in murine hematopoietic progenitors.Mol. Cell. Biol. 21, 854–864 (2001).
29. Biswas, S. C. & Greene, L. A. NGF down-regulates the BH3-only protein Bim andsuppresses its pro-apoptotic activity by phosphorylation. J. Biol. Chem. 277,49511–49516 (2002).
30. Weston, C. R. et al. Activation of ERK1/2 by deltaRaf-1:ER* represses Bim expres-sion independently of the JNK or PI3K pathways. Oncogene 22, 1281–1293(2003).
31. Ley, R., Balmanno, K., Hadfield, K., Weston, C. R. & Cook, S. J. Activation of theERK1/2 signalling pathway promotes phosphorylation and proteasome-dependentdegradation of the BH3-only protein Bim. J. Biol. Chem. 278, 18811–18816(2003).
32. Evan, G. & Littlewood, T. A matter of life and cell death. Science 281, 1317–1322(1998).
33. Edwards, P. A. Tissue reconstitution, or transgenic mammary gland, technique formodeling breast cancer development. Cancer Treat Res. 83, 23–36 (1996).
34. Ory, D. S., Neugeboren, B. A. & Mulligan, R. C. A stable human-derived packagingcell line for production of high titer retrovirus/vesicular stomatitis virus G pseudo-types. Proc. Natl Acad. Sci. USA 93, 11400–11406 (1996).
35. Ma, X. J. et al. Gene expression profiles of human breast cancer progression. Proc.Natl Acad. Sci. USA 100, 5974–5979 (2003).
© 2003 Nature Publishing Group

supplementary information
1
Figure S1 Bcl-xL Inhibition of Anoikis in MCF-10A Cells Requires its Binding to BH3-only Proteins. a, Anoikis in MCF-10A cells. MCF-10A cells were place in suspensionin growth medium and 0.5% methycellulose in poly-HEMA coated plates. Cells wereanalyzed for apoptosis using the DNA Fragmentation Cell Death ELISA assay at indi-cated times. Values represent the mean and standard deviation of at least threeindependent experiments. b, Percent of Dying Cells During Anoikis. MCF-10A cellsexpressing pBabe, or pBabe-Bcl-xL were placed in suspension for indicated timesand stained with calcein AM to label viable cells and ethidium bromide (excluded byintact membranes of live cells but readily labels nucleic acid of dying cells withcompromised membranes) to mark dying cells. Percent of cells labeled positive forethidium bromide were quantified. At least 200 cells were counted for each timepoint and represents the mean ± SEM of three independent experiments. c,Regulation of anoikis by Bcl-xL Mutants. MCF-10A cells expressing pMig, pMig-Bcl-xL, pMig-Bcl-xL (Mt1) or pMig-Bcl-xL (Mt8) were placed in suspension and analyzedfor apoptosis using the DNA Fragmentation ELISA assay. Data shown represents atleast three independent experiments and standard deviation.
Figure S2 Breast tumor cell line MDA-MB-231 does not downregulate EGFR levelsduring cell detachment. MCF-10A and MDA-MB-231 cells were placed in suspensionfor indicated times and lysates were analyzed for EGFR, Bim, phospho-Erk, or Erk2levels by western analysis.
Figure S3 Activated Myr-Akt does not inhibit Bim expression during anoikis.Attached cells expressing pWZL-Myr-Akt-ER were incubated for 48 hrs with vehiclecontrol (ethanol) or 4-hydroxy-tamoxifen (Tam. 1 µM) or placed in suspension for 48hrs in the presence of vehicle control (ethanol) or (Tam. 1 µM). Lysates were col-lected and proteins were analyzed by immunoblotting with antibodies against Bim,phospho-Akt substrate, phospho-Akt or actin.
© 2003 Nature Publishing Group




















