
Did a submarine landslide contribute to the 2011 Tohoku tsunami?

Ž .Journal of Neuroimmunology 103 2000 146–152www.elsevier.comrlocaterjneuroim
b7 integrins contribute to demyelinating disease of the central nervoussystem
Jagat R. Kanwar a, Jane E.B. Harrison a, Dongmao Wang a, Euphemia Leung a,Werner Mueller b, Norbert Wagner b, Geoffrey W. Krissansen a,)
a Department of Molecular Medicine, School of Medicine and Health Science, UniÕersity of Auckland, Auckland, New Zealandb Institute for Genetics, UniÕersity of Cologne, 50937 Cologne, Germany
Received 24 May 1999; received in revised form 18 August 1999; accepted 29 October 1999
Abstract
Ž .A role for a4 integrins in different forms of the multiple sclerosis-like disease experimental autoimmune encephalomyelitis EAE hasbeen demonstrated, but the individual contributions of a4b1, a4b7, and the related aEb7 integrin have not been determined. The b7integrins a4b7 and aEb7 play a central role in chronic inflammation, mediating the trafficking, entry, andror adhesion of lymphocytesin the inflamed pancreas and gut, and their ligands MAdCAM-1, VCAM-1 and E-cadherin are expressed on brain endothelial cellsandror on microvessels in the inflamed central nervous system. Here, we show that an antibody directed against the b7 subunit greatly
Ž .attenuates a non-remitting form of EAE, induced by adoptive transfer of myelin oligodendrocyte peptide MOG -stimulated T cells.35– 55
Combinational treatment with both anti-b7 and a4 integrin subunit antibodies led to more rapid and complete remission than thatobtained with anti-a4 antibody alone, potentially implicating a role for aEb7 in disease progression. Remission correlated with thedown-regulation of the vascular addressins VCAM-1, MAdCAM-1, and ICAM-1 on cerebral blood vessels. Attenuated forms of diseasewere induced by adoptive transfer of either wild-type encephalitogenic T cells to b7-deficient gene knockout mice, or of b7yry
encephalitogenic T cells to wild-type recipients. The former finding indicates that b7qve recruited cells contribute to disease progression.Thus a4b1, a4b7, and aEb7 integrins may all play a contributory role in the progression of chronic forms of demyelinating disease,and together with their ligands could represent potential targets for improved treatment of some forms of multiple sclerosis. q 2000Elsevier Science B.V. All rights reserved.
Keywords: Experimental autoimmune encephalomyelitis; b7 integrins; Cell adhesion molecules; Lymphocyte traffic; Central nervous system; Multiplesclerosis
1. Introduction
Ž .Experimental autoimmune encephalomyelitis EAE is aŽ .disease of the central nervous system CNS similar to
multiple sclerosis, in which encephalitogenic leukocytesŽ .penetrate the blood–brain barrier BBB and damage the
Žmyelin sheath of nerve fibres Raine, 1991, 1994; Cannellaand Raine, 1995; Martin and McFarland, 1995; Brosnan
.and Raine, 1996 . Both diseases lead to impaired nerveconduction and paralysis, which varies from relapsingrre-mitting to chronic progressive. The pathways by whichencephalitogenic T cells access the CNS have not beenfully delineated, but clearly involve integrin cell adhesion
) Corresponding author. Tel.: q64-9-373-7599 ext. 6280; fax: q64-9-373-7674; e-mail: [email protected]
molecules that mediate lymphocyte trafficking and recircu-lation.
The b7 integrin subfamily of cell adhesion moleculesŽconsists of two members, a4b7 Holzmann and Weiss-
. Žman, 1989; Yuan et al., 1992 and aEb7 Krissansen et.al., 1992; Parker et al., 1992 whose expression is re-
stricted to leukocytes. a4b7 mediates the adherence ofŽ .lymphocytes to high endothelial venules HEV via its
Žpreferred ligand MAdCAM-1 Berlin et al., 1993; Briskin.et al., 1993; Yang et al., 1995 , whereas aEb7 mediates
the adhesion of intraepithelial lymphocytes of the intestineto the intestinal epithelium via an interaction with E-
Ž .cadherin Cepek et al., 1994; Karecla et al., 1995 . In geneknockout mice deficient in the b7 integrin subfamily, the
Ž .gut-associated lymphoid tissue GALT is severely im-paired due to a failure of b7yry lymphocytes to adhere toblood vessel walls at the site of transmigration into the
0165-5728r00r$ - see front matter q 2000 Elsevier Science B.V. All rights reserved.Ž .PII: S0165-5728 99 00245-3

( )J.R. Kanwar et al.rJournal of Neuroimmunology 103 2000 146–152 147
Ž .GALT Wagner et al., 1996 . Further, in non-obese dia-Ž .betic NOD mice, treatment with anti-b7 or anti-
MAdCAM-1 mAbs provides protection against the sponta-neous development of diabetes and insulitis, presumablyby blocking lymphocyte migration into the inflamed pan-
Ž .creas Yang et al., 1997 . Hence a4b7-MAdCAM-1 inter-actions play critical roles in allowing the entry of leuko-cytes into chronically inflamed tissues.
A role for a4 integrins in the establishment of EAE wasdemonstrated by in vivo administration of anti-a4 integrinantibodies which diminished the paralysis associated with
Ž .EAE Yednock et al., 1992; Baron et al., 1993 . Further,the suppressive effects of TNFbp on the development ofEAE appears to correlate with down-regulation of VCAM-
Ž .1rVLA-4 Selmaj et al., 1998 . Hence, a4 integrins play acrucial role in allowing encephalitogenic T cells to enterthe brain, but the individual contributions of the tworelated a4 integrins a4b1 and a4b7 to the various stagesof the different forms of EAE has not been determined. Arecent study of a relapsing–remitting form of EAE inducedby proteolipid protein in the SJL mice, revealed that b7
Žintegrins play no role in the early stages of EAE En-.gelhardt et al., 1998 . However, this finding does not rule
out a role for b7 integrins in the chronic phase of non-re-mitting forms of the disease, as it is established thatMAdCAM-1, the preferential ligand for a4b7, is predomi-nantly expressed on specialized HEV formed at chroni-
Ž .cally inflamed sites Streeter et al., 1988 . Further, E-cadherin has also been found expressed on brain microves-
Ž .sel endothelial cells Pal et al., 1997 . Since MAdCAM-1expression is particularly noticeable on HEV-like endothe-
Žlia in relapsing brain lesions of mice with EAE Cannella.et al., 1991 , we sought to show a role for b7 integrins in
the chronic phase of a non-remitting form EAE.
2. Materials and methods
2.1. Mice and antibodies
C57BLr6 wild-type and b7yry gene knockout micewere bred and housed by the Animal Resource Unit,School of Medicine and Health Science, University ofAuckland, Auckland, New Zealand. They were randomlybred on a C57BLr6 background. Rat hybridomas secreting
ŽmAbs against mouse integrin a4 subunit PSr2 rat IgG2b. Ž . ŽmAb Miyake et al., 1991 , b7 subunit FIB 504.64 rat
. Ž . ŽIgG2a mAb Andrew et al., 1994 , VCAM-1 MK1.9 rat. Ž . ŽIgG1 mAb Miyake et al., 1991 , and ICAM-1 BE29G1
. Ž .rat IgG2a mAb Kuhlman et al., 1991 were purchasedfrom the American Type Culture Collection, Rockville,
Ž . ŽMD. The rat hybridoma MECA-367 rat IgG2a Berlin et.al., 1993 , which secretes a mAb against mouse MAd-
CAM-1, was kindly provided by Dr. Eugene Butcher,Stanford University, Stanford, CA. Rat IgG obtained from
Sigma, USA, served as a control for the latter antibodies inin vivo blocking and immunohistological studies.
2.2. Encephalitogenic lymphocytes
Encephalitogenic lymphocytes were obtained by subcu-taneous immunization of 8-10 week old wild-type andb7yry C57BLr6 mice with 300 mg of MOG peptide35 – 55Ž . ŽMEVGWYRSPFSRVVHLYRNGK Mendel et al.,
.1995 ; synthesized by Chiron Technologies, Clayton, Aus-tralia and emulsified in CFA containing 500 mg of My-
Žcobacterium tuberculosis H37RA DIFCO Laboratories,.Detroit, USA . Administration of pertussis toxin was not
necessary, and was omitted. Two injections were given 1week apart in opposite flanks, and 14 days later spleen anddraining lymph nodes were removed, and a single cellsuspension obtained by teasing the tissue through a stain-less steel wire mesh. Cells were then washed three times in
ŽHanks’ balanced salt solution HBSS, GIBCO, Grand Is-.land, NY, USA and resuspended to a concentration of
6=106 cellsrml in complete medium consisting of RPMIŽ .1640 GIBCO BRL, Gaithersburg, MD, USA supple-
mented with 1 mM L-glutamine, pencillin–streptomycin,10% FCS, and 2-mercaptoethanol. Cells were fed with 50
Ž .Urml of IL-2 R&D Systems, Minneapolis, MN, USAand 30 mgrml MOG peptide antigen, and incubated in ahumidified 378C, 10% CO incubator. After 4 d of activa-2
tion lymphoblasts were isolated by centrifugation on Fi-Žcoll-paque Pharmacia LKB Biotechnology AB, Uppsala,
.Sweden gradients. Blasts from the interface were washedthree times, and resuspended in complete medium. Cellviability as determined by trypan blue exclusion was )
97%. EAE was transferred to both wild-type and b7yry
C57BLr6 mice by injecting 6=108 MOG peptide-stimu-Ž .lated lymphocytes into the tail vein 70% and intraperi-
Ž .toneally 30% .
2.3. ProliferatiÕe response of lymphocytes to MOG peptide
The cell proliferation enzyme-linked immunosorbentŽ .assay ELISA was done with a non-radioactive and colori-
Žmetric immunoassay kit Boehringer Mannheim,.Mannheim, Germany to measure the incorporation of
Ž .5-bromo-2-deoxyuridine BrdU . In brief, lymphocytesfrom the spleens of wild-type and b7yry C57BLr6 mice
Žwere purified on Ficoll-paque Pharmacia LKB Biotech-.nology AB, Uppsala, Sweden . Equal numbers of wild-type
yry Ž 4.and b7 cells 1=10 were incubated in the presenceor absence of 30 mg of MOG peptide in 96-well35 – 55
Žflat-bottomed plates Becton Dickinson Labware, Frankin.Lakes, NJ, USA in triplicate for 4 days. The cells were
labelled with BrdU for 25 h, and fixed for 30 min at roomtemperature. The plates were washed and incubated withperoxidase-conjugated anti-BrdU Fab fragment. After
( )J.R. Kanwar et al.rJournal of Neuroimmunology 103 2000 146–152148
washing, substrate was added, the reaction stopped, andabsorbance measured in an ELISA plate reader.
2.4. EAE induction, and inhibition with antibodies
MOG peptide-stimulated lymphoblasts were injectedŽ . Žinto the tail vein 70% of cells and intraperitoneally 30%
.of cells into mice, and animals were observed daily forclinical signs of EAE. The mice were scored according tothe following scale: 0, no clinical signs of EAE; 1, limptail; 2, partial hind limb paralysis; 3, complete hind limbparalysis; 4, complete hind limb and partial fore limbparalysis; 5, paralysis extending to diaphragm; 6, hind andfore limb paralysis; 7, death due to EAE. Animals werecared for according to the guidelines of the University ofAuckland Animal Ethics Committee. They had to be main-tained in a state of hind-limb paralysis, hence care wasshown in the provision of drinking water, food, and com-fort. The daily mean clinical score for each group is themean disease score of at least five mice.
Ž .For antibody blocking, anti-b7 Fib 504 and anti-a4Ž .PSr2 subunit mAbs were administered into the tail veinŽ . Ž .70% of mAb and intraperitoneally 30% of mAb at 10mgrkg separately or in combination three times on alter-nate days when the clinical score reached 3 or 4.5. Control
Žgroups of mice received either PBS or rat IgG Sigma,.USA . Experiments were repeated to confirm reproducibil-
ity.
2.5. Immunohistology
Brains were frozen in isopentane at y708C, and trans-verse 10 mm sections were made through the cerebrum,cerebellum, and mid-brain. Sections were mounted on polyL-lysine-coated slides, and endogenous peroxidase block-ing was done by incubating slides for 30 min in 0.3%H O in methanol. The sections were stained with anti-2 2
CAM mAbs using VECTASTAIN ABC Elite PeroxidaseŽ . ŽRat IgG Kit Vector Laboratories, Burlingame, CA,
.USA . In brief, sections were washed with PBS and blockedwith rabbit serum, and incubated with different anti-CAMmAbs. After three washings with PBS, slides were incu-bated with biotinylated anti-rat secondary antibody for 10min. They were then washed, incubated with ABC peroxi-dase reagent, and developed with SIGMA FAST DABŽ X .3,3 -diaminobenzidine tetrahydrochloride with metal en-hancer CoCl tablets. Slides were counterstained with2
haematoxylin and eosin. Controls for the staining protocolconsisted of omitting the primary anti-CAM mAb, or
Ž .substituting it with rat IgG Sigma . Representative sec-tions are reported in the case of analysis of leukocyteinfiltration, whereas for visualisation of CAMs we deliber-ately chose blood vessels lacking extensive perivascularaccumulation of leukocytes for clarity.
3. Results
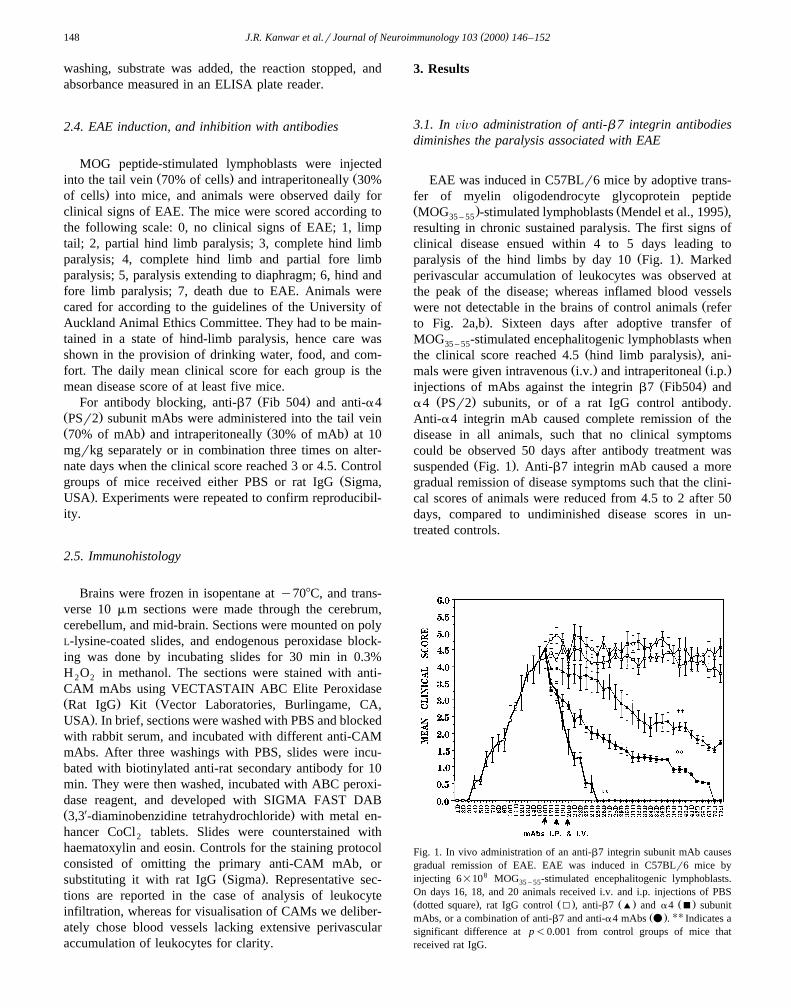
3.1. In ÕiÕo administration of anti-b 7 integrin antibodiesdiminishes the paralysis associated with EAE
EAE was induced in C57BLr6 mice by adoptive trans-fer of myelin oligodendrocyte glycoprotein peptideŽ . Ž .MOG -stimulated lymphoblasts Mendel et al., 1995 ,35 – 55
resulting in chronic sustained paralysis. The first signs ofclinical disease ensued within 4 to 5 days leading to
Ž .paralysis of the hind limbs by day 10 Fig. 1 . Markedperivascular accumulation of leukocytes was observed atthe peak of the disease; whereas inflamed blood vessels
Žwere not detectable in the brains of control animals refer.to Fig. 2a,b . Sixteen days after adoptive transfer of
MOG -stimulated encephalitogenic lymphoblasts when35 – 55Ž .the clinical score reached 4.5 hind limb paralysis , ani-
Ž . Ž .mals were given intravenous i.v. and intraperitoneal i.p.Ž .injections of mAbs against the integrin b7 Fib504 and
Ž .a4 PSr2 subunits, or of a rat IgG control antibody.Anti-a4 integrin mAb caused complete remission of thedisease in all animals, such that no clinical symptomscould be observed 50 days after antibody treatment was
Ž .suspended Fig. 1 . Anti-b7 integrin mAb caused a moregradual remission of disease symptoms such that the clini-cal scores of animals were reduced from 4.5 to 2 after 50days, compared to undiminished disease scores in un-treated controls.
Fig. 1. In vivo administration of an anti-b7 integrin subunit mAb causesgradual remission of EAE. EAE was induced in C57BLr6 mice byinjecting 6=108 MOG -stimulated encephalitogenic lymphoblasts.35 – 55
On days 16, 18, and 20 animals received i.v. and i.p. injections of PBSŽ . Ž . Ž . Ž .dotted square , rat IgG control I , anti-b7 ' and a4 B subunit
Ž . UUmAbs, or a combination of anti-b7 and anti-a4 mAbs v . Indicates asignificant difference at p-0.001 from control groups of mice thatreceived rat IgG.
( )J.R. Kanwar et al.rJournal of Neuroimmunology 103 2000 146–152 149
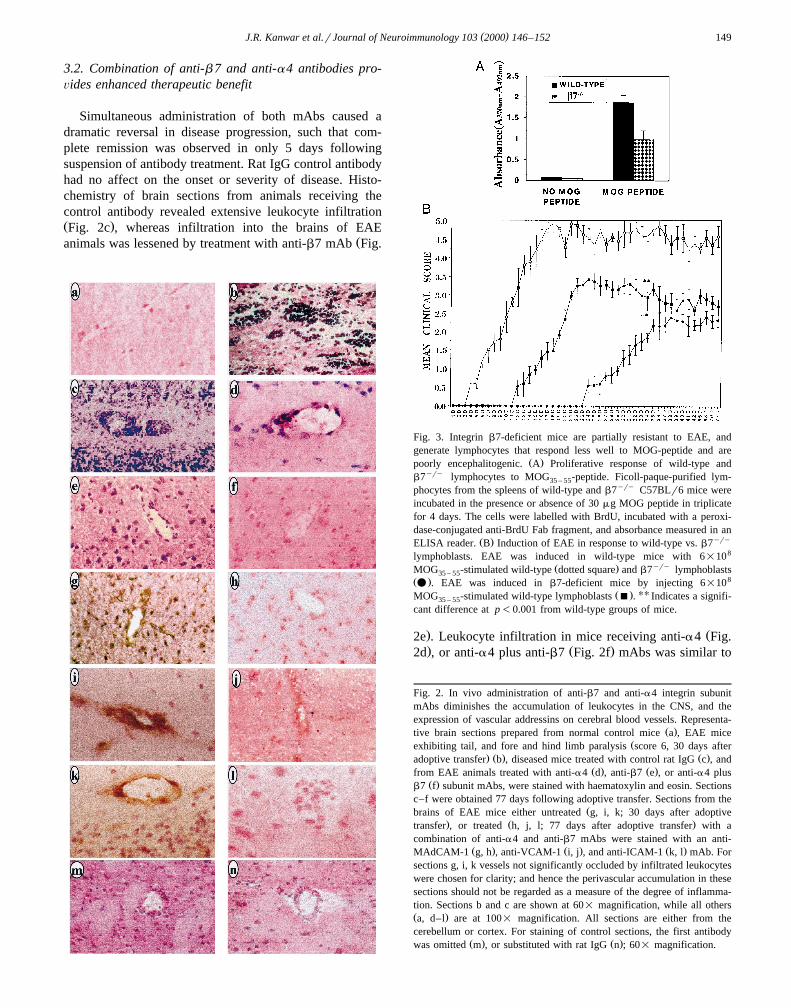
3.2. Combination of anti-b 7 and anti-a4 antibodies pro-Õides enhanced therapeutic benefit
Simultaneous administration of both mAbs caused adramatic reversal in disease progression, such that com-plete remission was observed in only 5 days followingsuspension of antibody treatment. Rat IgG control antibodyhad no affect on the onset or severity of disease. Histo-chemistry of brain sections from animals receiving thecontrol antibody revealed extensive leukocyte infiltrationŽ .Fig. 2c , whereas infiltration into the brains of EAE
Žanimals was lessened by treatment with anti-b7 mAb Fig.
Fig. 3. Integrin b7-deficient mice are partially resistant to EAE, andgenerate lymphocytes that respond less well to MOG-peptide and are
Ž .poorly encephalitogenic. A Proliferative response of wild-type andb7yry lymphocytes to MOG -peptide. Ficoll-paque-purified lym-35 – 55
phocytes from the spleens of wild-type and b7yry C57BLr6 mice wereincubated in the presence or absence of 30 mg MOG peptide in triplicatefor 4 days. The cells were labelled with BrdU, incubated with a peroxi-dase-conjugated anti-BrdU Fab fragment, and absorbance measured in an
Ž . yryELISA reader. B Induction of EAE in response to wild-type vs. b7lymphoblasts. EAE was induced in wild-type mice with 6=108
Ž . yryMOG -stimulated wild-type dotted square and b7 lymphoblasts35 – 55Ž . 8v . EAE was induced in b7-deficient mice by injecting 6=10
Ž . UUMOG -stimulated wild-type lymphoblasts B . Indicates a signifi-35 – 55
cant difference at p-0.001 from wild-type groups of mice.
. Ž2e . Leukocyte infiltration in mice receiving anti-a4 Fig.
. Ž .2d , or anti-a4 plus anti-b7 Fig. 2f mAbs was similar to
Fig. 2. In vivo administration of anti-b7 and anti-a4 integrin subunitmAbs diminishes the accumulation of leukocytes in the CNS, and theexpression of vascular addressins on cerebral blood vessels. Representa-
Ž .tive brain sections prepared from normal control mice a , EAE miceŽexhibiting tail, and fore and hind limb paralysis score 6, 30 days after
. Ž . Ž .adoptive transfer b , diseased mice treated with control rat IgG c , andŽ . Ž .from EAE animals treated with anti-a4 d , anti-b7 e , or anti-a4 plus
Ž .b7 f subunit mAbs, were stained with haematoxylin and eosin. Sectionsc–f were obtained 77 days following adoptive transfer. Sections from the
Žbrains of EAE mice either untreated g, i, k; 30 days after adoptive. Ž .transfer , or treated h, j, l; 77 days after adoptive transfer with a
combination of anti-a4 and anti-b7 mAbs were stained with an anti-Ž . Ž . Ž .MAdCAM-1 g, h , anti-VCAM-1 i, j , and anti-ICAM-1 k, l mAb. For
sections g, i, k vessels not significantly occluded by infiltrated leukocyteswere chosen for clarity; and hence the perivascular accumulation in thesesections should not be regarded as a measure of the degree of inflamma-tion. Sections b and c are shown at 60= magnification, while all othersŽ .a, d–l are at 100= magnification. All sections are either from thecerebellum or cortex. For staining of control sections, the first antibody
Ž . Ž .was omitted m , or substituted with rat IgG n ; 60= magnification.
( )J.R. Kanwar et al.rJournal of Neuroimmunology 103 2000 146–152150
that in healthy control mice. MAdCAM-1, VCAM-1, andICAM-1 were all highly expressed at the peak of disease
Ž .30 days after adoptive Fig. 2g,i,k , whereas the expressionof all three vascular addressins was dramatically downreg-ulated in anti-a4 and anti-b7 mAb-treated mice by day 77Ž .Fig. 2 h,j,l .
3.3. AdoptiÕe transfer of wild-type MOG -stimulated35- 55
lymphoblasts induces an attenuated form of disease inb 7-deficient mice
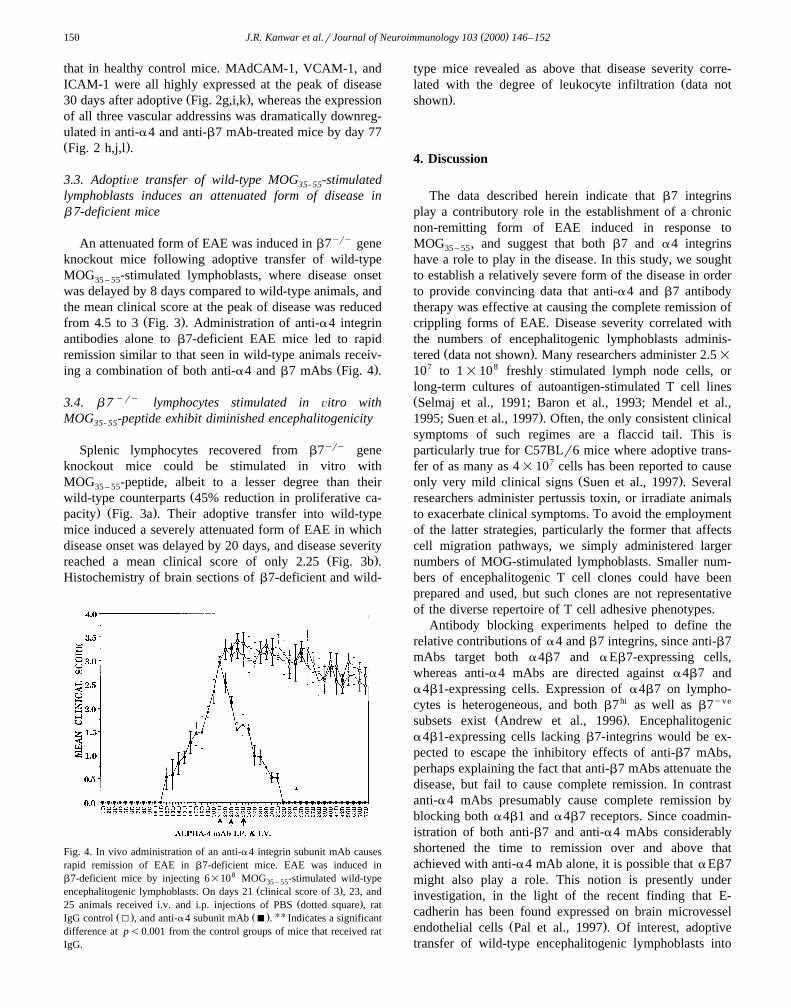
An attenuated form of EAE was induced in b7yry geneknockout mice following adoptive transfer of wild-typeMOG -stimulated lymphoblasts, where disease onset35 – 55
was delayed by 8 days compared to wild-type animals, andthe mean clinical score at the peak of disease was reduced
Ž .from 4.5 to 3 Fig. 3 . Administration of anti-a4 integrinantibodies alone to b7-deficient EAE mice led to rapidremission similar to that seen in wild-type animals receiv-
Ž .ing a combination of both anti-a4 and b7 mAbs Fig. 4 .
3.4. b 7 yry lymphocytes stimulated in Õitro withMOG -peptide exhibit diminished encephalitogenicity35- 55
Splenic lymphocytes recovered from b7yry geneknockout mice could be stimulated in vitro withMOG -peptide, albeit to a lesser degree than their35 – 55
Žwild-type counterparts 45% reduction in proliferative ca-. Ž .pacity Fig. 3a . Their adoptive transfer into wild-type
mice induced a severely attenuated form of EAE in whichdisease onset was delayed by 20 days, and disease severity
Ž .reached a mean clinical score of only 2.25 Fig. 3b .Histochemistry of brain sections of b7-deficient and wild-
Fig. 4. In vivo administration of an anti-a4 integrin subunit mAb causesrapid remission of EAE in b7-deficient mice. EAE was induced inb7-deficient mice by injecting 6=108 MOG -stimulated wild-type35 – 55
Ž .encephalitogenic lymphoblasts. On days 21 clinical score of 3 , 23, andŽ .25 animals received i.v. and i.p. injections of PBS dotted square , rat
Ž . Ž . UUIgG control I , and anti-a4 subunit mAb B . Indicates a significantdifference at p-0.001 from the control groups of mice that received ratIgG.
type mice revealed as above that disease severity corre-Žlated with the degree of leukocyte infiltration data not
.shown .
4. Discussion
The data described herein indicate that b7 integrinsplay a contributory role in the establishment of a chronicnon-remitting form of EAE induced in response toMOG , and suggest that both b7 and a4 integrins35 – 55
have a role to play in the disease. In this study, we soughtto establish a relatively severe form of the disease in orderto provide convincing data that anti-a4 and b7 antibodytherapy was effective at causing the complete remission ofcrippling forms of EAE. Disease severity correlated withthe numbers of encephalitogenic lymphoblasts adminis-
Ž .tered data not shown . Many researchers administer 2.5=
107 to 1=108 freshly stimulated lymph node cells, orlong-term cultures of autoantigen-stimulated T cell linesŽSelmaj et al., 1991; Baron et al., 1993; Mendel et al.,
.1995; Suen et al., 1997 . Often, the only consistent clinicalsymptoms of such regimes are a flaccid tail. This isparticularly true for C57BLr6 mice where adoptive trans-fer of as many as 4=107 cells has been reported to cause
Ž .only very mild clinical signs Suen et al., 1997 . Severalresearchers administer pertussis toxin, or irradiate animalsto exacerbate clinical symptoms. To avoid the employmentof the latter strategies, particularly the former that affectscell migration pathways, we simply administered largernumbers of MOG-stimulated lymphoblasts. Smaller num-bers of encephalitogenic T cell clones could have beenprepared and used, but such clones are not representativeof the diverse repertoire of T cell adhesive phenotypes.
Antibody blocking experiments helped to define therelative contributions of a4 and b7 integrins, since anti-b7mAbs target both a4b7 and aEb7-expressing cells,whereas anti-a4 mAbs are directed against a4b7 anda4b1-expressing cells. Expression of a4b7 on lympho-cytes is heterogeneous, and both b7hi as well as b7yve
Ž .subsets exist Andrew et al., 1996 . Encephalitogenica4b1-expressing cells lacking b7-integrins would be ex-pected to escape the inhibitory effects of anti-b7 mAbs,perhaps explaining the fact that anti-b7 mAbs attenuate thedisease, but fail to cause complete remission. In contrastanti-a4 mAbs presumably cause complete remission byblocking both a4b1 and a4b7 receptors. Since coadmin-istration of both anti-b7 and anti-a4 mAbs considerablyshortened the time to remission over and above thatachieved with anti-a4 mAb alone, it is possible that aEb7might also play a role. This notion is presently underinvestigation, in the light of the recent finding that E-cadherin has been found expressed on brain microvessel
Ž .endothelial cells Pal et al., 1997 . Of interest, adoptivetransfer of wild-type encephalitogenic lymphoblasts into

( )J.R. Kanwar et al.rJournal of Neuroimmunology 103 2000 146–152 151
b7-deficient mice induced an attenuated form of the dis-ease, suggesting that b7 integrins may be important for theencephalitogenic functions of non-T leukocyte subsets such
Ž .as granulocytes, and myeloid Cua et al., 1995 cells thatfacilitate the disease, andror for the recruitment of addi-tional lymphocytes. We have previously shown that b7-in-tegrins are expressed de novo on differentiating mono-
Ž .cytesrmacrophages Yang et al., 1996 .All three leukocyte integrin ligands MAdCAM-1,
VCAM-1, and ICAM-1 were highly expressed on brainmicrovessels at the height of disease severity, and wereeither absent or expressed at low levels in antibody-treatedmice in remission. It was noticeable that MAdCAM-1expression was not restricted to endothelia, but was alsofound on neighbouring cells. Whilst these MAdCAM-1positive cells have not be identified, similar results have
Žbeen reported elsewhere Cannella et al., 1991; O’Neill et. Ž .al., 1991 . Cannella et al. 1991 noted that some astro-
cytes surrounding blood vessels stained with the anti-MAdCAM-1 mAb MECA-89. MAdCAM-1 has also been
Ž .detected on dendritic cells Szabo et al., 1997 , and choroidŽ .plexus epithelium Steffen et al., 1996 .
The b7 integrins could potentially act at various stagesof the immune response including antigen presentationduring epitope spreading, which occurs in MOG-induced
Ž .disease Bernard et al., 1997 . It should be rememberedŽ . Žthat a4b7 Lehnert et al., 1998 and aEb7 Berg et al.,
.1999 mediate T cell costimulation, and their ligandsŽ . ŽVCAM-1 Koopman et al., 1991 , MAdCAM-1 Szabo et. Ž .al., 1997 and E-cadherin Ebner et al., 1998 are all
expressed on dendritic cells. This notion seems unlikelygiven the demonstration that administration of anti-b7integrin antibodies prior to the development of clinicalsigns of disease has no inhibitory effect on disease pro-
Ž .gression Engelhardt et al., 1998 . However, it was notedthat b7yry lymphocytes did not proliferate to the sameextent as wild-type lymphocytes in response to MOGpeptide, indicating that b7 integrins may be involved in Tcell costimulation during antigen presentation. Alterna-tively, both a4 and b7 integrins may be required to allowtransmigration of different encephalitogenic leukocyte sub-
Ž .populations T and non-T across the BBB, as previouslyproposed for a4b1. Whatever the mechanism, effectivetherapies designed to treat chronic non-remitting forms ofinflammatory diseases of the CNS may need to targetmultiple adhesion pathways, including those involving theb7 integrins.
Acknowledgements
This work was supported in part by grants from theMultiple Sclerosis Society of New Zealand, the RoyalSociety of New Zealand, the Health Research Council ofNew Zealand, and The Lottery Grants Board of New
Ž .Zealand. G.W.K. was a recipient of a Wellcome UK
Senior Research Fellowship in Medical Science in NewZealand, and is now a James Cook Research Fellowfunded by the Royal Society of New Zealand. W.M. andN.W. are supported by the Deutsche Forschungsgemein-
Ž .schaft WA 1127r1-1 .
References
Andrew, D.P., Berlin, C., Honda, S., Yoshino, T., Hamann, A., Holz-mann, B., Kilshaw, P.J., Butcher, E.C., 1994. Distinct but overlappingepitopes are involved in alpha 4 beta 7-mediated adhesion to vascularcell adhesion molecule-1, mucosal addressin-1, fibronectin, and lym-phocyte aggregation. J. Immunol. 153, 3847–3861.
Andrew, D.P., Rott, L.S., Kilshaw, P.J., Butcher, E.C., 1996. Distributionof a4b7 and aEb7 integrins on thymocytes, intestinal epitheliallymphocytes and peripheral lymphocytes. Eur. J. Immunol. 26, 897–905.
Baron, J.L., Madri, J.A., Ruddle, N.H., Hashim, G., Janeway, C.A. Jr.,1993. Surface expression of a4 integrin by CD4 T cells is requiredfor their entry into brain parenchyma. J. Exp. Med. 177, 57–68.
Berg, R.W., Yang, Y., Lehnert, K., Krissansen, G.W., 1999. MouseM290 is the functional homologue of the human mucosal lymphocyteintegrin HML-1: antagonism between the integrin ligands E-Cadherinand RGD tripeptide. Immunol. Cell Biol. 77, 337–344.
Berlin, C., Berg, E.L., Briskin, M.J., Andrew, D.P., Kilshaw, P.J.,Holzmann, B., Weissman, I.L., Hamann, A., Butcher, E.C., 1993.a4b7 integrin mediates lymphocyte binding to the mucosal vascularaddressin MAdCAM-1. Cell 74, 185–195.
Bernard, C.C.A., Johns, T.G., Slavin, A., Ichikawa, M., Ewing, C., Liu,J., Bettadapura, J., 1997. Myelin oligodendrocyte glycoprotein: anovel candidate autoantigen in multiple sclerosis. J. Mol. Med. 75,77–88.
Briskin, M.J., McEvoy, L.M., Butcher, E.C., 1993. MAdCAM-1 hashomology to immunoglobulin and mucin-like adhesion receptors andto IgA1. Nature 363, 461–463.
Brosnan, C.F., Raine, C.S., 1996. Mechanisms of immune injury inmultiple sclerosis. Brain Pathol. 6, 243–257.
Cannella, B., Cross, A.H., Raine, C.S., 1991. Relapsing autoimmunedemyelination: a role for vascular addressins. J. Neuroimmunol. 35,295–300.
Cannella, B., Raine, C.S., 1995. The adhesion molecule and cytokineprofile of multiple sclerosis lesions. Ann. Neurol. 37, 424–435.
Cepek, K.L., Shaw, S.K., Parker, C.M., Russell, G.J., Morrow, J.S.,Rimm, D.L., Brenner, M.B., 1994. Adhesion between epithelial cellsand T lymphocytes mediated by E-cadherin and the aEb7 integrin.Nature 372, 190–193.
Cua, D.J., Hinton, D.R., Kirkman, L., Stohlman, S.A., 1995. Macrophagesregulate induction of delayed-type hypersensitivity and experimentalallergic encephalomyelitis in SJL mice. Eur. J. Immunol. 25, 2318–2324.
Ebner, S., Lenz, A., Reider, D., Fritsch, P., Schuler, G., Romani, N.,1998. Expression of maturation-rmigration-related molecules on hu-man dendritic cells from blood and skin. Immunobiology 198, 568–587.
Engelhardt, B., Laschinger, M., Schulz, M., Samulowitz, U., Vestweber,D., Hoch, G., 1998. The development of experimental autoimmuneencephalomyelitis in the mouse requires a4-integrin but not a4b7-in-tegrin. J. Clin. Invest. 102, 2096–2105.
Holzmann, B., Weissman, I.L., 1989. Peyer’s patch-specific lymphocytehoming receptors consist of a VLA-4-like a chain associated witheither of two integrin b chains, one of which is novel. EMBO J. 8,1735–1741.
Karecla, P.I., Bowden, S.J., Green, S.J., Kilshaw, P.J., 1995. Recognitionof E-cadherin on epithelial cells by the mucosal T cell integrin
Ž .aM290b7 aEb7 . Eur. J. Immunol. 25, 852–856.

( )J.R. Kanwar et al.rJournal of Neuroimmunology 103 2000 146–152152
Koopman, G., Parmentier, H.K., Schuurman, H.J., Newman, W., Meijer,C.J., Pals, S.T., 1991. Adhesion of human B cells to folliculardendritic cells involves both the lymphocyte function-associated anti-gen 1rintercellular adhesion molecule 1 and very late antigen4rvascular cell adhesion molecule 1 pathways. J. Exp. Med. 173,1297–1304.
Krissansen, G.W., Print, C.G., Prestidge, R.L., Hollander, D., Yuan, Q.,Jiang, W.-M., Jenkins, D.R., Leung, E., Mead, P., Yong, R., Amer-atunga, R.V., Cerf-Bensussan, N., Watson, J.D., 1992. Immunologicand structural relatedness of the integrin b7 complex and the humanintraepithelial lymphocyte antigen HML-1. FEBS Lett. 296, 25–28.
Kuhlman, P., Moy, V.T., Lollo, B.A., Brian, A.A., 1991. The accessoryfunction of murine intercellular adhesion molecule-1 in T lymphocyteactivation. J. Immunol. 146, 1773–1782.
Lehnert, K., Print, C.G., Yang, Y., Krissansen, G.W., 1998. MucosalŽ .addressin cell adhesion molecule-1 MAdCAM-1 costimulates T cell
Ž .proliferation exclusively through integrin a4b7 LPAM-1 , whereasŽ .VCAM-1 and CS-1 peptide use a4b1 VLA-4 : Evidence for ‘‘re-
mote’’ costimulation and induction of hyperresponsiveness to B7molecules. Eur. J. Immunol. 28, 3605–3615.
Martin, R., McFarland, H.F., 1995. Immunological aspects of experimen-tal allergic encephalomyelitis and multiple sclerosis. Crit. Rev. Clin.Lab. Sci. 32, 121–182.
Mendel, I., Kerlero de Rosbo, N., Ben-Nun, A., 1995. A myelin oligo-dendrocyte glycoprotein peptide induces typical chronic experimentalautoimmune encephalomyelitis in H-2b mice: fine specificity and Tcell receptor V beta expression of encephalitogenic T cells. Eur. J.Immunol. 25, 1951–1959.
Miyake, K., Weissman, L.I., Greenberger, J.S., Kincade, P.W., 1991.Evidence for a role of the integrin VLA-4 in lympho-hemopoiesis. J.Exp. Med. 173, 599–607.
O’Neill, J.K., Butter, C., Baker, D., Gschmeissner, S.E., Kraal, G.,Butcher, E.C., Turk, J.L., 1991. Expression of vascular addressins andICAM-1 by endothelial cells in the spinal cord during chronic relaps-ing experimental allergic encephalomyelitis in the Biozzi ABrHmouse. Immunology 72, 520–525.
Pal, D., Audus, K.L., Siahaan, T.J., 1997. Modulation of cellular adhe-sion in bovine brain microvessel endothelial cells by a decapeptide.Brain Res. 747, 103–113.
Parker, C.M., Cepek, K.L., Russell, G.J., Shaw, S.K., Posnett, D.N.,Schwarting, R., Brenner, M.B., 1992. A family of b7 integrins onhuman mucosal lymphocytes. Proc. Natl. Acad. Sci. U.S.A. 89,1924–1928.
Raine, C.S., 1991. Multiple sclerosis: a pivotal role for the T cell inlesion development. Neuropathol. Appl. Neurobiol. 17, 265–274.
Raine, C.S., 1994. The Dale E. McFarlin Memorial Lecture: the im-munology of the multiple sclerosis lesion. Ann. Neurol. 36, S61–72.
Selmaj, K., Raine, C.S., Cross, A.H., 1991. Anti-tumor necrosis factortherapy abrogates autoimmune demyelination. Ann. Neurol. 30, 694–700.
Selmaj, K., Walczak, A., Mycko, M., Berkowicz, T., Kohno, T., Raine,C.S., 1998. Suppression of experimental autoimmune encephalomyeli-
Ž .tis with a TNF binding protein TNFbp correlates with down-regu-lation of VCAM-1rVLA-4. Eur. J. Immunol. 28, 2035–2044.
Steffen, B.J., Breier, G., Butcher, E.C., Schulz, M., Engelhardt, B., 1996.ICAM-1, VCAM-1, and MAdCAM-1 are expressed on choroid plexusepithelium but not endothelium and mediate binding of lymphocytesin vitro. Am. J. Pathol. 148, 1819–1838.
Streeter, P.R., Berg, E.L., Rouse, B.T., Bargatze, R.F., Butcher, E.C.,1988. A tissue-specific endothelial cell molecule involved in lympho-cyte homing. Nature 331, 41–46.
Suen, W.E., Bergman, C.M., Hjelmstrom, P., Ruddle, N.H., 1997. Acritical role for lymphotoxin in experimental allergic encephalomyeli-tis. J. Exp. Med. 186, 1233–1240.
Szabo, M.C., Butcher, E.C., McEvoy, L.M., 1997. Specialization ofmucosal follicular dendritic cells revealed by mucosal addressin-celladhesion molecule-1 display. J. Immunol. 158, 5584–5588.
Wagner, N., Lohler, J., Kunkel, E.J., Ley, K., Leung, E., Krissansen, G.,Rajewsky, K., Muller, W., 1996. Critical role for b7 integrins in theformation of the gut associated lymphoid tissue. Nature 382, 366–370.
Yang, X., Sytwn, H.-K., McDevitt, H.O., Michie, S.A., 1997. Involve-ment of b7 integrin and mucosal addressin cell adhesion molecule-1Ž .MAdCAM-1 in the development of diabetes in nonobese diabeticmice. Diabetes 46, 1542–1547.
Yang, Y., Sammar, M., Harrison, J.E.B., Lehnert, K., Print, C.G., Leung,E., Prestidge, R., Krissansen, G.W., 1995. Construction and adhesiveproperties of a soluble MAdCAM-1-Fc chimera expressed in a bac-ulovirus system: phylogenetic conservation of receptor-ligand interac-tion. Scand. J. Immunol. 42, 235–247.
Yang, Y., Harrison, J.E.B., Print, C.G., Lehnert, K., Sammar, M.,Lazarovits, A., Krissansen, G.W., 1996. Interaction of monocytoidcells with the mucosal addressin MAdCAM-1 via the integrins VLA-4and LPAM-1. Immunol. Cell Biol. 74, 383–393.
Yednock, T.A., Cannon, C., Fritz, L.C., Sanchez-Madrid, F., Steinman,L., Karin, N., 1992. Prevention of experimental autoimmune en-cephalomyelitis by antibodies against a4b1 integrin. Nature 356,63–66.
Yuan, Q., Jiang, W.-M., Leung, E., Hollander, D., Watson, J.D., Kris-sansen, G.W., 1992. Molecular cloning of the mouse integrin b7subunit. J. Biol. Chem. 267, 7352–7358.
Copyright © 2022 FDOKUMEN




















