
Acylhydrazones Contribute to Serum Glucose Homeostasis Through Dual Physiological Targets
10
Send Orders of Reprints at [email protected] Current Topics in Medicinal Chemistry, 2012, 12, 2049-2058 2049 Acylhydrazones Contribute to Serum Glucose Homeostasis Through Dual Physiological Targets Marisa Jádna Silva Frederico 1 , Allisson Jhonatan Gomes Castro 1 , Alessandra Mascarello 2 , Camila Pires Mendes 1 , Virgínia Dermachi Kappel 1 , Taisa Regina Stumpf 2 , Paulo Cesar Leal 2 , Ricardo José Nunes 2 , Rosendo Augusto Yunes 2 and Fátima Regina Mena Barreto Silva 1,* 1 Universidade Federal de Santa Catarina, Departamento de Bioquímica, Centro de Ciências Biológicas, Campus Uni- versitário, Bairro Trindade, Cx. Postal 5069, CEP: 88040-970, Florianópolis, SC, Brazil; 2 Universidade Federal de Santa Catarina, Departamento de Química, Centro de Ciências Físicas e Matemáticas, Campus Universitário, Bairro Trindade, CEP: 88040-900, Florianópolis, SC, Brazil Abstract: In this study the in vivo and in vitro antidiabetic effects of four acylhydrazone derivatives were investigated in rats. The secretagogue action, oral glucose tolerance, insulinogenic index and mechanism of action of these acylhydra- zones in relation to calcium uptake in pancreatic islets were studied. Also, the insulinomimetic effect on glycemia in dia- betic rats was verified. Of the acylhydrazones studied, 1 and 4 were able to increase glucose tolerance in an acute time- course. A powerful secretagogue effect was exhibited by 1 and glibenclamide with an insulinogenic index around 3.9 and 1.3-fold higher than that of the hyperglycemic group, respectively. Moreover, an acute and dose-dependent effect of glibenclamide and 1 on calcium uptake in pancreatic islets was observed. The rapid stimulatory effect of 1 on calcium up- take seems to be mediated, at least in part, by ATP-dependent K + channels (K + -ATP) since the stimulatory effect of 1 was similar to that observed for glibenclamide but was not potentiated by sulphonylurea. Furthermore, both extracellular and calcium from stocks mediate the signal transduction of stimulatory effect of 1 on calcium uptake which may contribute to insulin secretion. In addition, the insulinomimetic effect of 1 was evidenced through the level of serum glucose lowering in alloxan-induced diabetic rats. Also, 1 induced a significant increase in glycogen content in vivo and glucose uptake in soleus muscle in vitro. The results of this study indicate dual physiological targets for the acylhydrazone 1, i.e., pancreatic islets and skeletal muscle, as a result of insulin secretagogue and insulinomimetic action. Keywords: Acylhydrazones, calcium, insulinomimetic, glucose, glycemia, pancreas, secretagogue, skeletal muscle. INTRODUCTION Diabetes mellitus (DM) is a global health problem and one of the major causes of morbidity and mortality [1]. The incidence of the disease is high worldwide and varies be- tween populations due to genetic and susceptibility differ- ences as well as other modifiable risk factors [2, 3]. While the prevalence of DM has reached epidemic proportions, most available treatments still focus on the symptoms of the disease, rather than the underlying pathology itself. There are basically two types of diabetes, type 1(T1DM) and type 2 (T2DM). T2DM accounts for the vast majority of cases of diabetes, and is a heterogeneous disease marked by -cell dysfunction [4, 5] combined with insulin resistance in target tissues [4, 6, 7]. The pathogenesis of the disease involves both genetic [8] and environmental factors, and is frequently associated with obesity [6, 9, 10, 11]. T1DM is characterized by the destruction of pancreatic -cells due to T-cell- mediated auto-immune attack [12], which not yet fully un- derstood. *Address correspondence to this author at the Departamento de Bioquímica, Centro de Ciências Biológicas, UFSC. Campus Universitário, Bairro Trin- dade, Cx Postal 5069, CEP: 88040-970 - Florianópolis, Santa Catarina, Brazil; Tel: +55 48 3721.6912; Fax: +55 48 3721.9672; E-mail: [email protected] Types 1 and 2 diabetes have in common a deficit in - cell mass. While the etiologies associated with T1DM and T2DM differ, both forms are characterized by a relative defi- cit in functional -cell mass such that the insulin require- ments of the body are not met and glycemia is uncontrolled [13]. Thus, studies that aim to investigate the mechanisms of action of new compounds in insulin targets in order to high- lights the diabetes treatments as well as improve glucose tolerance, either through improving the insulin secretory output of the remaining -cells (insulin secretagogues), as well as improving the response to insulin in target tissues (insulin sensitizers), or replacing the missing insulin (exoge- nous insulinomimetic compounds), are of great importance in relation to this pathogenesis. Acylhydrazones structural skeleton is similar to the an- tidiabetic compounds, chalcones, previously studied [14,15,16]. Acylhydrazones compounds presents a wide spectrum of biological properties including analgesic, anti- inflammatory, antithrombotic, antinociceptive and antiplate- let activity [17,18]. N-acylhydrazones have been shown to produce a cardio inotropic effect and vasodilation of vascular smooth muscle in rats [19]. Moreover, recently, the struc- ture–activity relationships of 3,4,5-trimethoxy-hydrazones has been described, these compounds showing potent anti- cancer activity and the ability to provoke tubulin depolym- 1873-5294/12 $58.00+.00 © 2012 Bentham Science Publishers
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Acylhydrazones Contribute to Serum Glucose Homeostasis Through Dual Physiological Targets

Send Orders of Reprints at [email protected]
Current Topics in Medicinal Chemistry, 2012, 12, 2049-2058 2049
Acylhydrazones Contribute to Serum Glucose Homeostasis Through Dual Physiological Targets
Marisa Jádna Silva Frederico1, Allisson Jhonatan Gomes Castro
1, Alessandra Mascarello
2,
Camila Pires Mendes1, Virgínia Dermachi Kappel
1, Taisa Regina Stumpf
2, Paulo Cesar Leal
2,
Ricardo José Nunes2, Rosendo Augusto Yunes
2 and
Fátima Regina Mena Barreto Silva
1,*
1Universidade Federal de Santa Catarina, Departamento de Bioquímica, Centro de Ciências Biológicas, Campus Uni-versitário, Bairro Trindade, Cx. Postal 5069, CEP: 88040-970, Florianópolis, SC, Brazil; 2Universidade Federal de Santa Catarina, Departamento de Química, Centro de Ciências Físicas e Matemáticas, Campus Universitário, Bairro Trindade, CEP: 88040-900, Florianópolis, SC, Brazil
Abstract: In this study the in vivo and in vitro antidiabetic effects of four acylhydrazone derivatives were investigated in
rats. The secretagogue action, oral glucose tolerance, insulinogenic index and mechanism of action of these acylhydra-
zones in relation to calcium uptake in pancreatic islets were studied. Also, the insulinomimetic effect on glycemia in dia-
betic rats was verified. Of the acylhydrazones studied, 1 and 4 were able to increase glucose tolerance in an acute time-
course. A powerful secretagogue effect was exhibited by 1 and glibenclamide with an insulinogenic index around 3.9 and
1.3-fold higher than that of the hyperglycemic group, respectively. Moreover, an acute and dose-dependent effect of
glibenclamide and 1 on calcium uptake in pancreatic islets was observed. The rapid stimulatory effect of 1 on calcium up-
take seems to be mediated, at least in part, by ATP-dependent K+ channels (K
+-ATP) since the stimulatory effect of 1 was
similar to that observed for glibenclamide but was not potentiated by sulphonylurea. Furthermore, both extracellular and
calcium from stocks mediate the signal transduction of stimulatory effect of 1 on calcium uptake which may contribute to
insulin secretion. In addition, the insulinomimetic effect of 1 was evidenced through the level of serum glucose lowering
in alloxan-induced diabetic rats. Also, 1 induced a significant increase in glycogen content in vivo and glucose uptake in
soleus muscle in vitro. The results of this study indicate dual physiological targets for the acylhydrazone 1, i.e., pancreatic
islets and skeletal muscle, as a result of insulin secretagogue and insulinomimetic action.
Keywords: Acylhydrazones, calcium, insulinomimetic, glucose, glycemia, pancreas, secretagogue, skeletal muscle.
INTRODUCTION
Diabetes mellitus (DM) is a global health problem and
one of the major causes of morbidity and mortality [1]. The
incidence of the disease is high worldwide and varies be-
tween populations due to genetic and susceptibility differ-
ences as well as other modifiable risk factors [2, 3]. While
the prevalence of DM has reached epidemic proportions,
most available treatments still focus on the symptoms of the
disease, rather than the underlying pathology itself. There are
basically two types of diabetes, type 1(T1DM) and type 2
(T2DM). T2DM accounts for the vast majority of cases of
diabetes, and is a heterogeneous disease marked by -cell
dysfunction [4, 5] combined with insulin resistance in target
tissues [4, 6, 7]. The pathogenesis of the disease involves
both genetic [8] and environmental factors, and is frequently
associated with obesity [6, 9, 10, 11]. T1DM is characterized
by the destruction of pancreatic -cells due to T-cell-
mediated auto-immune attack [12], which not yet fully un-
derstood.
*Address correspondence to this author at the Departamento de Bioquímica,
Centro de Ciências Biológicas, UFSC. Campus Universitário, Bairro Trin-
dade, Cx Postal 5069, CEP: 88040-970 - Florianópolis, Santa Catarina,
Brazil; Tel: +55 48 3721.6912; Fax: +55 48 3721.9672;
E-mail: [email protected]
Types 1 and 2 diabetes have in common a deficit in -
cell mass. While the etiologies associated with T1DM and
T2DM differ, both forms are characterized by a relative defi-
cit in functional -cell mass such that the insulin require-
ments of the body are not met and glycemia is uncontrolled
[13]. Thus, studies that aim to investigate the mechanisms of
action of new compounds in insulin targets in order to high-
lights the diabetes treatments as well as improve glucose
tolerance, either through improving the insulin secretory
output of the remaining -cells (insulin secretagogues), as
well as improving the response to insulin in target tissues
(insulin sensitizers), or replacing the missing insulin (exoge-
nous insulinomimetic compounds), are of great importance
in relation to this pathogenesis.
Acylhydrazones structural skeleton is similar to the an-
tidiabetic compounds, chalcones, previously studied
[14,15,16]. Acylhydrazones compounds presents a wide
spectrum of biological properties including analgesic, anti-
inflammatory, antithrombotic, antinociceptive and antiplate-
let activity [17,18]. N-acylhydrazones have been shown to
produce a cardio inotropic effect and vasodilation of vascular
smooth muscle in rats [19]. Moreover, recently, the struc-
ture–activity relationships of 3,4,5-trimethoxy-hydrazones
has been described, these compounds showing potent anti-
cancer activity and the ability to provoke tubulin depolym-
1873-5294/12 $58.00+.00 © 2012 Bentham Science Publishers

2050 Current Topics in Medicinal Chemistry, 2012, Vol. 12, No. 19 Frederico et al.
erization [20,21]. On the other hand, to the best of our
knowledge, the effect of acylhydrazones on diabetes was not
yet been reported. In this context, the aim of this study was
to determine the role of acylhydrazones on serum glucose
lowering and on insulin secretion as well as and the mecha-
nism of action associated with calcium uptake in pancreatic
islets. Insulomimetic effect of the acylhydrazone 1 in soleus
muscle was determined through glycogen content and glu-
cose uptake measurements. In addition, the acute toxicity of
acylhydrazones was investigated.
EXPERIMENTAL PROTOCOLS
Materials
Alloxan monohydrate [14], glibenclamide, diazoxide
[22], nifedipine [23], (1-[6-[((17 )-3-methoxyestra-
1,3,5[10]-trien-17-yl)amino]hexyl]-1H-pyrrole-2,5-dione) U-
73122 [21] and oyster glycogen type II were purchased from
Sigma Chemical Company (St. Louis, MO, USA). The io-
dine reagents (CaCl2 + I2 + KI) were purchased from
VETEC, Rio de Janeiro, Brazil. The commercial kit used to
determine the glycemia was produced by Analisa (Belo
Horizonte, MG, Brazil). The enzyme-linked immunosorbent
assay (ELISA) for quantitative determination of rat insulin
(catalogue no. EZRMI-13K) was purchased from Millipore
(St Charles, MO, USA). [45
Ca]CaCl2 (sp. act. 321 KBq/mg
Ca2+
), [U-14
C]-2-deoxy-D-glucose ([14
C]-DG), specific activ-
ity 10.6 GBq/mmol and biodegradable scintillation liquid
were obtained from Perkin Elmer Life and Analytical Sci-
ences (Boston, MA, USA). Salts and solvents were pur-
chased from Merck AG (Darmstadt. Germany).
For the in vivo experiments acylhydrazones were dis-
solved in a solution of 0.3% Tween 80. In all experiments,
the vehicle (solution of 0.3% Tween 80) alone was not able
to modify the results under any of the experimental condi-
tions, for the basal and treated groups (data not shown). For
the in vitro experiments 1 and 4 acylhydrazones were dis-
solved in dimethyl sulfoxide and added to both the control
and test media at final concentrations not exceeding 0.1%
(v/v). At this concentration, dimethyl sulfoxide failed to af-
fect islet function [24].
Experimental Animals
Male albino Wistar rats (180–210 g; 49 day-old) were
bred in an animal facility and housed in an air conditioned
room (approximately 21 ± 2 °C) with controlled lighting
(lights on from 06:00 to 18:00 h). Rats were maintained with
pelleted food (Nuvital, Nuvilab CR1, Curitiba, PR, Brazil),
and tap water was available ad libitum. For the in vivo and in vitro treatments rats were deprived of food for at least 16 h
but allowed free access to water. All the animals were moni-
tored and maintained in accordance with the ethical recom-
mendations of the Brazilian Veterinary-Medicine Council
and the Brazilian College of Animal Experimentation
(CEUA protocol PP00414/UFSC).
Studies on Effect of Acylhydrazones on Oral Glucose Tolerance
Hyperglycemic rats were divided into different groups of
four rats (see Tables and Figures), for the control and treated
animals. Control groups were randomly divided as follows:
rats which received vehicle (solution of 0.3% Tween 80);
and hyperglycemic control group of rats which received glu-
cose (4 g/kg; 8.9 M), p.o. gavage. Treated rats received the
acylhydrazones 1 (1, 10 and 100 mg/kg), 2, 3 and 4 (10
mg/kg) or glibenclamide (10 mg/kg) as a positive control
group, p.o. gavage. The serum glucose levels were measured
immediately prior to, and at 15, 30, 60 and 180 min after
treatment. After centrifugation, serum samples were used to
determine glucose and insulin levels or lactate dehydro-
genase activity. Blood samples taken from the tail vein were
collected and centrifuged and the serum was used to deter-
mine the glycemia by the glucose oxidase method [25].
Insulin Serum Measurements
The insulin was measured applying the enzyme-linked
immunosorbent assay (ELISA) according to the manufac-
turer’s instructions. The range of values detected by this as-
say was 0.63 ng/mL to 4.3 ng/mL. The intra- and inter-assay
coefficients of variation (CV) for insulin were 3.22 and 6.95,
respectively, with a sensitivity of 0.2 ng/ mL. All insulin
levels were estimated by means of colorimetric measure-
ments at 450 nm with an ELISA plate reader (Organon Tek-
nika, Roseland, NJ, USA) by interpolation from a standard
curve. Samples were analyzed in duplicate and results were
expressed as ng of insulin serum mL-1
. The incremental areas
under the response curves (AUCs) were calculated. The insu-
linogenic index (II) was calculated as the ratio between the
AUCinsulin and AUCglucose (from zero to 60 min) [14].
Pancreatic Islet Isolation
The rat pancreas was visualized by way of a central ab-
dominal incision. The bile duct was clamped at the tip of the
duodenum and cannulated at a point sufficiently proximal to
the liver. Krebs Ringer bicarbonate buffer with a composi-
tion of 122 mM NaCl, 3 mM KCl, 1.2 mM MgSO4, 1.3 mM
CaCl2, 0.4 mM KH2PO4, and 25 mM NaHCO3 supplemented
with 8 mM HEPES (KRb-HEPES) medium was introduced
slowly into the bile duct by syringe until the pancreas was
clearly distended. The pancreas was then gently removed and
then kept in Petri dishes with KRb-HEPES medium. The
pancreatic tissue was cut into small pieces (2 x 2 mm) and
incubated in plastic tubes in 1 mL of KRb-HEPES medium
supplemented with collagenase (3 mg). After incubation the
mixture was transferred to a conical tube (110 x 15 mm),
resuspended in 10 mL with collagenase free medium and
centrifuged at room temperature for 3 min at 45 x g in an
Excelsa Baby centrifuge (model 206), FANEM, São Paulo,
SP, Brazil. The supernatant was discarded and the sediment
resuspended in fresh KRb-HEPES medium. This washing
procedure was repeated five times; in the last two washings
the islets were allowed to settle without centrifugation. Ali-
quots (100 L) of the final sediment with the isolated islet
were transferred to eppendorff tubes with the incubation me-
dium KRb-HEPES [26,27].
Calcium Uptake Experiments
The isolated islets were pre-incubated for 60 min in a
Dubnoff metabolic incubator to equilibrate in KRb-HEPES
buffer containing 5 mM glucose and 0.1 Ci/mL45
Ca2+
at

Antihyperglycemic and Hypoglycemic Effects of Acylhydrazones Current Topics in Medicinal Chemistry, 2012, Vol. 12, No. 19 2051
37°C, pH 7.4 and gassed with O2:CO2(95:5; v/v). The islets
were incubated for 10 min in KRb-HEPES without (control)
or with the acylhydrazone 1 (0.2, 2 and 20 M). In some
experiments a channel antagonist, agonist or phospholipase
C (PLC) inhibitor were added during the last 15 min of incu-
bation before the treatment and maintained during the entire
incubation period (see figure legends). The following drugs
were used: nifedipine (0.1, 1 and 10 M), diazoxide (250
M) [28,29], glibenclamide (0.2, 2 and 20 M), U-73122 (1
M) [23]. The incubation was halted using the technique
reported by Batra and Sjogren [30] with modifications [28].
Cold buffer (1 mL with lanthanum chloride (10 mM) at 2°C
was added to the samples to stop calcium fluxes. The tubes
were centrifuged for 1 min at 45 x g. The supernatant was
preserved and the islets were washed twice in cold lantha-
num chloride solution. The islets were added to screw-cap
tubes containing 300 L of 0.5 M NaOH solution and boiled
at 100°C for 5 min. Aliquots of 50 L were taken from each
sample for the measurement of radioactivity in scintillation
liquid Optiphase Hisafe III (Wallac Oy, Turku, Finland) in
an LKB rack beta liquid scintillation spectrometer (model LS
6500; Multi-Purpose Scintillation Counter-Beckman Coulter,
Boston, USA) and 5 L aliquots were used for protein quan-
tification by the Lowry method [31].
Studies on Effect of Acylhydrazones on Serum Glucose Levels in Diabetic Rats
Diabetes was induced by a single intravenous injection of
5% (w/v) alloxan monohydrate in a saline solution at a dose
of 50 mg/kg body weight. Blood samples were collected 3
days later and glucose levels were determined to confirm the
development of diabetes [14]. Fasted diabetic rats received
the acylhydrazone 1 (10 mg/kg) by oral gavage. The serum
glucose was measured immediately prior to and after 1, 2
and 3 h of treatment.
Skeletal Muscle and Liver Glycogen Measurements
Soleus muscle and liver were harvested from hypergly-
cemic rats, untreated and treated with the acylhydrazone 1
(10 mg/kg), and then used for the assay of glycogen content
immediately after 3 h of treatment. Glycogen was isolated
from these tissues as described by Krisman [32], with minor
modifications [33]. The tissues were weighed, homogenized
in 33% KOH and boiled at 100°C for 20 min, with occa-
sional stirring. After cooling, 96% ethanol was added to the
samples which were heated to boiling followed by cooling in
an ice bath to aid the precipitation of glycogen. The ho-
mogenates were centrifuged at 1300 x g for 15 min, the su-
pernatant was discarded and the pellets were neutralized with
saturated NH4Cl before being heated to 100°C for 5 min,
washed and resolubilized in water. Glycogen content was
determined by treatment with iodine reagent and the absor-
bance was measured at 460 nm. The results are expressed as
mg of glycogen/g of tissue.
Studies on Glucose Uptake in Rat Soleus Muscle
For the [U-14
C]-2-deoxy-D-glucose uptake experiments,
soleus muscles from normal fasted rats were used. Soleus
muscles were distributed (alternately left and right) between
basal and treated groups. The muscles were dissected and
then preincubated (30 min) and incubated (60 min) at 37°C
in KRb buffer with 3 mM glucose bubbled with O2/CO2
(95%:5%, v/v) until pH 7.4. The acylhydrazone 1 (20 M)
was added to the incubation medium (60 min) in the pres-
ence of 14
C-DG (0.1 Ci/mL). After incubation the muscles
were placed in screw cap tubes containing 500 L of a 0.5 M
NaOH solution and boiled at 100°C for 5 min. Aliquots of 25
L were taken from each sample for radioactivity measure-
ments in scintillation liquid Optiphase Hisafe III (Wallac Oy,
Turku, Finland) on an LKB rack beta liquid scintillation
spectrometer (model LS 6500; Multi-Purpose Scintillation
Counter-Beckman Coulter, Boston, USA) and 5 L aliquots
were used for protein quantification by the Lowry method
[31]. The results were expressed as the nmol glycosyl units/
mg protein [34], with modifications.
Lactate Dehydrogenase Assay (LDH Assay)
The in vivo toxicity was assessed by measuring the re-
lease of the cytosolic enzyme lactate dehydrogenase into
serum after 180 min of treatment with the acylhydrazone 1.
The LDH activity was measured applying a spectropho-
tometric method and the level of activity was calculated by
measuring the oxidation of NADH with the results being
expressed as % of control [35].
Data and Statistical Analysis
Data were expressed as mean ± S.E.M. One-way analysis
of variance (ANOVA) followed by the Bonferroni post-hoc test or unpaired Student’s t-test was used to determine the
significance of differences between groups. Differences were
considered to be significant at p 0.05.
Chemistry
The acylhydrazones were prepared by condensation of
3,4,5-trimethoxybenzohydrazide and the appropriate alde-
hyde, under reflux [36]. The 3,4,5-
trimethoxybenzohydrazide was prepared as previously de-
scribed [37]. Details on the synthesis of the compounds have
been previously published [15,16,21].
RESULTS AND DISCUSSION
Studies on Effect of Acylhydrazones on Oral Glucose
Tolerance
The results related to the effect of acylhydrazones and
glibenclamide on serum glucose levels in hyperglycemic rats
can be seen in Fig. (1) and (Table 1). Initially, a dose-
response curve on oral glucose tolerance for the acylhydra-
zone 1 (1, 10 and 100 mg/kg), was obtained. As expected, in
the hyperglycemic control group after starting the oral glu-
cose tolerance test, the serum glucose level was typically
increased when compared with zero time for the same group.
However, at a dose of 10 mg/kg the acylhydrazone 1
produced remarkable serum glucose lowering at 15, 30 and
60 min (28%, 22% and 19%, respectively) compared to the
hyperglycemic control group. As it can be seen at Fig. 1, the
oral hypoglycemic compound, glibenclamide, produced a
serum glucose lowering after 15, 30 and 60 min at dose of 10
mg/kg (23%, 32% and 32%) compared to the hyperglycemic
control group, respectively. This dose of glibenclamide is in
line with that reported by Sakamoto et al. [38] and León-
Reyes et al. [39], concerning the significant therapeutic
effect on glycemic control. On the other hand, at doses of 1

2052 Current Topics in Medicinal Chemistry, 2012, Vol. 12, No. 19 Frederico et al.
and 100 mg/kg of the acylhydrazone 1 slightly reduced
glycemia (by around 14% and 12%) only after 30 min,
respectively [Fig. (1)]. Based on these results, the dose of 10
mg/kg was used for further studies with other
acylhydrazones.
Our previous studies identified the substituents at differ-
ent positions on the core structure of chalcones that deter-
mine their antihyperglycemic activity as well as the specific-
ity of action [14,40,41]. Taking in mind that the structural
skeleton of acylhydrazones is similar to that of chalcones,
some selected modifications to the core structure of acylhy-
drazones were made to assay them in vivo and in vitro
[Scheme 1].
Scheme 1. Structure of the acylhydrazones.
Based on the preliminary results for 1 (substituent 1-
naphthyl), we tested the structural analogue of 2 (substituent
2-naphthyl). As can be observed in (Table 1), the compound
2 produced serum glucose lowering of around 20% and 11%
at 15 and 30 min, respectively, when compared with
hyperglycemic control group. These data indicate that 2 was
less potent than compound 1, namely due to a change in the
conformation of the naphthyl group. Also, we analyzed the
influence of an electron donor group in the core of the
acylhydrazone. The oral administration of 3 (substituent p-
N(CH3)2) slightly reduced the serum glucose by 18% and
23% at 15 and 30 min, respectively [Table 1]. On the other
hand, the oral administration of an acylhydrazone with the
electron acceptor group 4 (substituent p-Br) showed signifi-
cant, acute and lasting serum glucose lowering, of around
25%, 1.8% and 16% at 15, 30 and 60 min, respectively.
Based on the results outlined herein the development of a
series of novel molecules is underway in order to improve
the biological activity.
This first goal was to demonstrate that the acylhydra-
zones 1 and 4 markedly reduce serum glucose levels and
then to determine the insulinogenic index, mechanism of
action and insulinomimetic effect for these compounds.
Effect of Acylhydrazone and Glibenclamide on Serum Insulin Measurements
(Table 2) shows a typical pattern of insulin secretion after
glucose overloading (4 g/kg) in hyperglycemic control
groups with an insulinogenic index (II) of 0.66. The acylhy-
drazones 1 and 4 exhibited high II values of around 2.5.
Also, the acylhydrazone 1 showed a potent insulin
secretagogue effect within 15 min after oral treatment. As it
was expected, the glibenclamide was effective to induce in-
sulin secretion exhibiting an II around 2. It is reported that
the most important pathway for quantitative insulin release
from islet interstitial is regulated by Ca2 +
ion [42-47]. In this
context, the mechanism of action of this efficient insulin
secretagogue and antihyperglycemic compound, acylhydra-
zone 1, was studied in calcium uptake in pancreatic islets.
Effect of Acylhydrazone on Ca2+
Uptake in Pancreatic Islets
After the equilibrium of calcium in the pancreatic islets
following 60 min of incubation with 45
Ca2+
(0.1 Ci/mL),
calcium measurements were carried out after 10 min in the
presence of glibenclamide or the acylhydrazone 1 (0.2, 2 and
20 M). (Table 3) shows that glibenclamide stimulated sig-
nificantly the calcium uptake at 20 M. Percentile, the dose
of 20 M of the acylhydrazone 1 showed a stimulatory effect
on calcium uptake around 3.0 fold (p < 0.001) greater than
that observed for the control group after 10 min. The rapid
calcium uptake is associated with the first phase of insulin
secretion which is sustained due to two functional pools de-
fined as the readily releasable pool and the immediately re-
leasable pool [48,49]. This is consistent with the literature
where it is demonstrated that an intracellular calcium in-
crease in –cells is essential for exocytose of insulin gran-
ules [50-54]. Therefore, based on the dose-response curve, in
all subsequent experiments 20 M of the acylhydrazone 1
and a 10 min incubation period were used, since 10 min is
related to the first phase of insulin release in –cells.
In the –cell, the closing of potassium channels leads to
the depolarization of the plasma membrane with subsequent
opening of voltage-dependent Ca2+
channels and acceleration
of Ca2+
uptake. This abrupt increase in the concentration of
cytosolic-free Ca2+
([Ca2+
]i) is mandatory for the triggering
of insulin secretion [53-57]. Thus, the mechanism of action
of the acylhydrazone 1 associated with the stimulatory effect
on 45
Ca2+
uptake was investigated in pancreatic islets. To
assess the involvement of potassium channels in the
stimulatory effect of the acylhydrazone 1 on calcium uptake
one antagonist and one agonist of potassium channels were
used, that is, glibenclamide (20 μM) and diazoxide (250 μM)
[22], respectively.
Compound 1 was able to increase significantly the
calcium uptake, by around 77% [Fig. (2A)]. Glibenclamide,
a blocker of K+-ATP channels, stimulated calcium uptake by
a factor of 1.5 when compared with the control. On the other
hand, when glibenclamide was preincubated before 1, a
significant decrease in the stimulatory effect of 1 on calcium
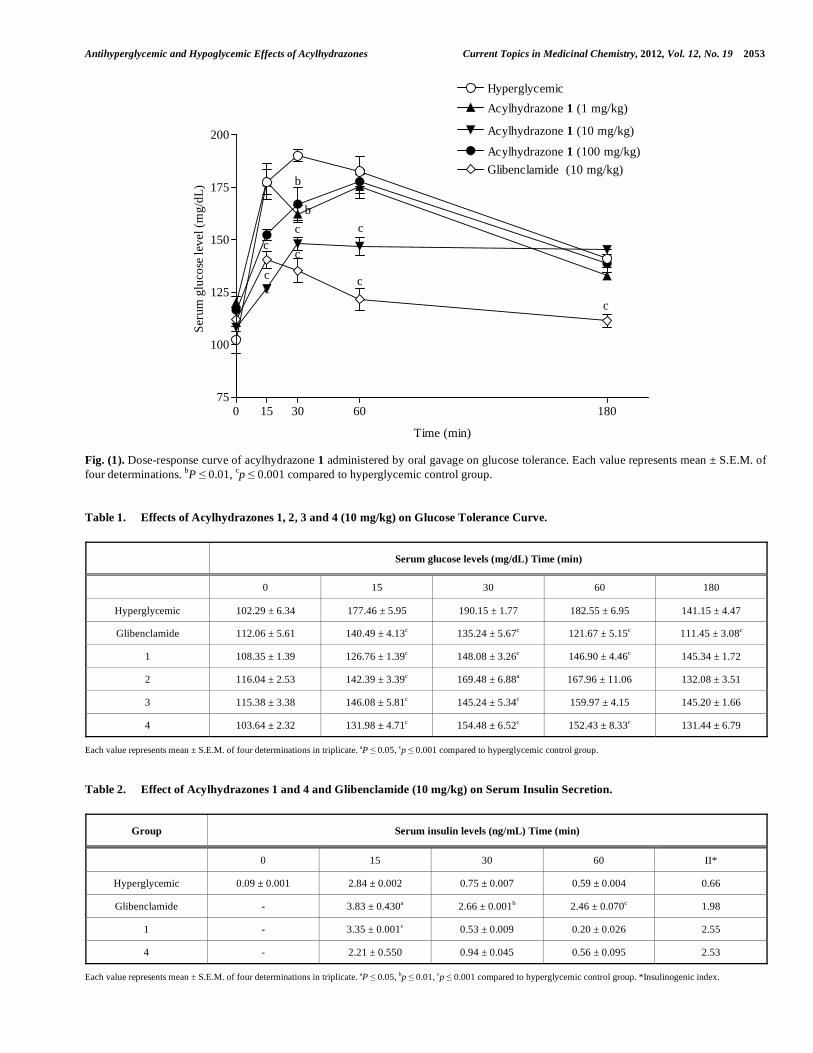
Antihyperglycemic and Hypoglycemic Effects of Acylhydrazones Current Topics in Medicinal Chemistry, 2012, Vol. 12, No. 19 2053
Fig. (1). Dose-response curve of acylhydrazone 1 administered by oral gavage on glucose tolerance. Each value represents mean ± S.E.M. of
four determinations. bP 0.01,
cp 0.001 compared to hyperglycemic control group.
Table 1. Effects of Acylhydrazones 1, 2, 3 and 4 (10 mg/kg) on Glucose Tolerance Curve.
Serum glucose levels (mg/dL) Time (min)
0 15 30 60 180
Hyperglycemic 102.29 ± 6.34 177.46 ± 5.95 190.15 ± 1.77 182.55 ± 6.95 141.15 ± 4.47
Glibenclamide 112.06 ± 5.61 140.49 ± 4.13c 135.24 ± 5.67
c 121.67 ± 5.15
c 111.45 ± 3.08
c
1 108.35 ± 1.39 126.76 ± 1.39c 148.08 ± 3.26
c 146.90 ± 4.46
c 145.34 ± 1.72
2 116.04 ± 2.53 142.39 ± 3.39c 169.48 ± 6.88
a 167.96 ± 11.06 132.08 ± 3.51
3 115.38 ± 3.38 146.08 ± 5.81c 145.24 ± 5.34
c 159.97 ± 4.15 145.20 ± 1.66
4 103.64 ± 2.32 131.98 ± 4.71c 154.48 ± 6.52
c 152.43 ± 8.33
c 131.44 ± 6.79
Each value represents mean ± S.E.M. of four determinations in triplicate. aP 0.05,
cp 0.001 compared to hyperglycemic control group.
Table 2. Effect of Acylhydrazones 1 and 4 and Glibenclamide (10 mg/kg) on Serum Insulin Secretion.
Group Serum insulin levels (ng/mL) Time (min)
0 15 30 60 II*
Hyperglycemic 0.09 ± 0.001 2.84 ± 0.002 0.75 ± 0.007 0.59 ± 0.004 0.66
Glibenclamide - 3.83 ± 0.430a 2.66 ± 0.001
b 2.46 ± 0.070
c 1.98
1 - 3.35 ± 0.001c 0.53 ± 0.009 0.20 ± 0.026 2.55
4 - 2.21 ± 0.550 0.94 ± 0.045 0.56 ± 0.095 2.53
Each value represents mean ± S.E.M. of four determinations in triplicate. aP 0.05,
bp 0.01, cp 0.001 compared to hyperglycemic control group. *Insulinogenic index.
0 15 30 60 18075
100
125
150
175
200
Acylhydrazone 1 (1 mg/kg)
Acylhydrazone 1 (10 mg/kg)
Hyperglycemic
c
Acylhydrazone 1 (100 mg/kg)
c c
c
b
b
Glibenclamide (10 mg/kg)
c
c
cc
Time (min)
Seru
m g
luco
se le
vel (
mg/
dL)
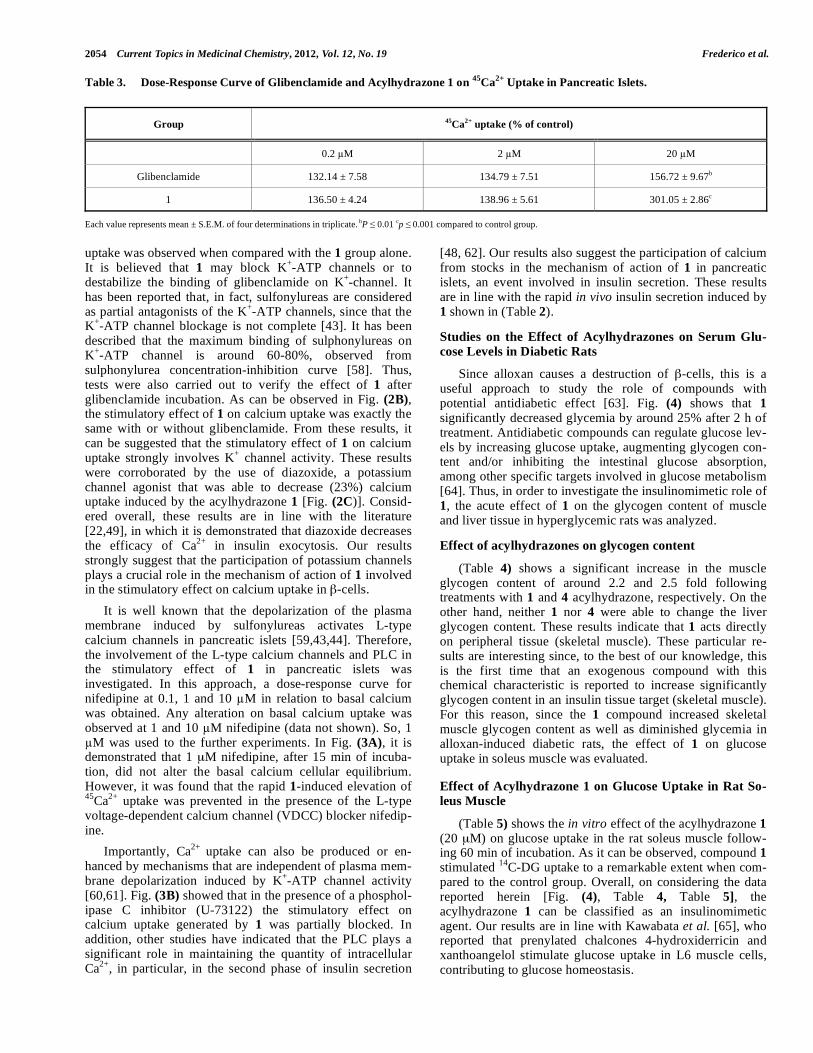
2054 Current Topics in Medicinal Chemistry, 2012, Vol. 12, No. 19 Frederico et al.
Table 3. Dose-Response Curve of Glibenclamide and Acylhydrazone 1 on 45
Ca2+
Uptake in Pancreatic Islets.
Group 45
Ca2+
uptake (% of control)
0.2 M 2 M 20 M
Glibenclamide 132.14 ± 7.58 134.79 ± 7.51 156.72 ± 9.67b
1 136.50 ± 4.24 138.96 ± 5.61 301.05 ± 2.86c
Each value represents mean ± S.E.M. of four determinations in triplicate. bP 0.01
cp 0.001 compared to control group.
uptake was observed when compared with the 1 group alone.
It is believed that 1 may block K+-ATP channels or to
destabilize the binding of glibenclamide on K+-channel. It
has been reported that, in fact, sulfonylureas are considered
as partial antagonists of the K+-ATP channels, since that the
K+-ATP channel blockage is not complete [43]. It has been
described that the maximum binding of sulphonylureas on
K+-ATP channel is around 60-80%, observed from
sulphonylurea concentration-inhibition curve [58]. Thus,
tests were also carried out to verify the effect of 1 after
glibenclamide incubation. As can be observed in Fig. (2B), the stimulatory effect of 1 on calcium uptake was exactly the
same with or without glibenclamide. From these results, it
can be suggested that the stimulatory effect of 1 on calcium
uptake strongly involves K+ channel activity. These results
were corroborated by the use of diazoxide, a potassium
channel agonist that was able to decrease (23%) calcium
uptake induced by the acylhydrazone 1 [Fig. (2C)]. Consid-
ered overall, these results are in line with the literature
[22,49], in which it is demonstrated that diazoxide decreases
the efficacy of Ca2+
in insulin exocytosis. Our results
strongly suggest that the participation of potassium channels
plays a crucial role in the mechanism of action of 1 involved
in the stimulatory effect on calcium uptake in -cells.
It is well known that the depolarization of the plasma
membrane induced by sulfonylureas activates L-type
calcium channels in pancreatic islets [59,43,44]. Therefore,
the involvement of the L-type calcium channels and PLC in
the stimulatory effect of 1 in pancreatic islets was
investigated. In this approach, a dose-response curve for
nifedipine at 0.1, 1 and 10 M in relation to basal calcium
was obtained. Any alteration on basal calcium uptake was
observed at 1 and 10 M nifedipine (data not shown). So, 1
M was used to the further experiments. In Fig. (3A), it is
demonstrated that 1 μM nifedipine, after 15 min of incuba-
tion, did not alter the basal calcium cellular equilibrium.
However, it was found that the rapid 1-induced elevation of 45
Ca2+
uptake was prevented in the presence of the L-type
voltage-dependent calcium channel (VDCC) blocker nifedip-
ine.
Importantly, Ca2+
uptake can also be produced or en-
hanced by mechanisms that are independent of plasma mem-
brane depolarization induced by K+-ATP channel activity
[60,61]. Fig. (3B) showed that in the presence of a phosphol-
ipase C inhibitor (U-73122) the stimulatory effect on
calcium uptake generated by 1 was partially blocked. In
addition, other studies have indicated that the PLC plays a
significant role in maintaining the quantity of intracellular
Ca2+
, in particular, in the second phase of insulin secretion
[48, 62]. Our results also suggest the participation of calcium
from stocks in the mechanism of action of 1 in pancreatic
islets, an event involved in insulin secretion. These results
are in line with the rapid in vivo insulin secretion induced by
1 shown in (Table 2).
Studies on the Effect of Acylhydrazones on Serum Glu-
cose Levels in Diabetic Rats
Since alloxan causes a destruction of -cells, this is a
useful approach to study the role of compounds with
potential antidiabetic effect [63]. Fig. (4) shows that 1
significantly decreased glycemia by around 25% after 2 h of
treatment. Antidiabetic compounds can regulate glucose lev-
els by increasing glucose uptake, augmenting glycogen con-
tent and/or inhibiting the intestinal glucose absorption,
among other specific targets involved in glucose metabolism
[64]. Thus, in order to investigate the insulinomimetic role of
1, the acute effect of 1 on the glycogen content of muscle
and liver tissue in hyperglycemic rats was analyzed.
Effect of acylhydrazones on glycogen content
(Table 4) shows a significant increase in the muscle
glycogen content of around 2.2 and 2.5 fold following
treatments with 1 and 4 acylhydrazone, respectively. On the
other hand, neither 1 nor 4 were able to change the liver
glycogen content. These results indicate that 1 acts directly
on peripheral tissue (skeletal muscle). These particular re-
sults are interesting since, to the best of our knowledge, this
is the first time that an exogenous compound with this
chemical characteristic is reported to increase significantly
glycogen content in an insulin tissue target (skeletal muscle).
For this reason, since the 1 compound increased skeletal
muscle glycogen content as well as diminished glycemia in
alloxan-induced diabetic rats, the effect of 1 on glucose
uptake in soleus muscle was evaluated.
Effect of Acylhydrazone 1 on Glucose Uptake in Rat So-
leus Muscle
(Table 5) shows the in vitro effect of the acylhydrazone 1
(20 μM) on glucose uptake in the rat soleus muscle follow-
ing 60 min of incubation. As it can be observed, compound 1
stimulated 14
C-DG uptake to a remarkable extent when com-
pared to the control group. Overall, on considering the data
reported herein [Fig. (4), Table 4, Table 5], the
acylhydrazone 1 can be classified as an insulinomimetic
agent. Our results are in line with Kawabata et al. [65], who
reported that prenylated chalcones 4-hydroxiderricin and
xanthoangelol stimulate glucose uptake in L6 muscle cells,
contributing to glucose homeostasis.
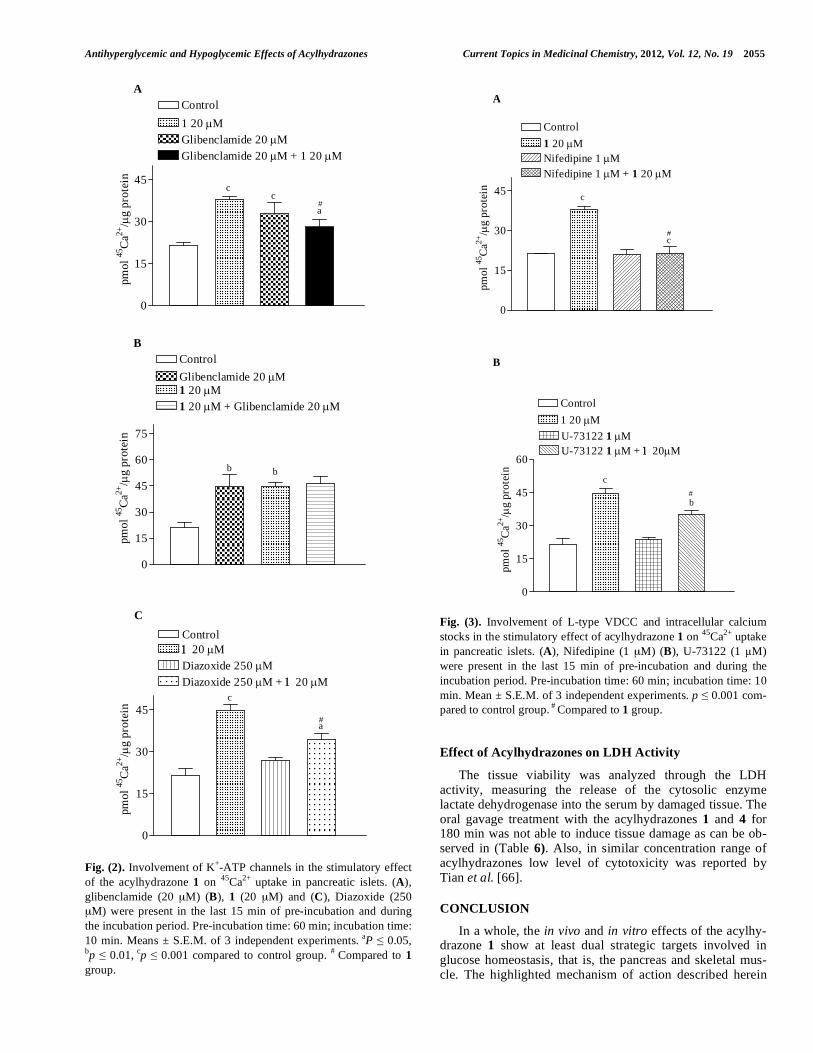
Antihyperglycemic and Hypoglycemic Effects of Acylhydrazones Current Topics in Medicinal Chemistry, 2012, Vol. 12, No. 19 2055
Fig. (2). Involvement of K+-ATP channels in the stimulatory effect
of the acylhydrazone 1 on 45
Ca2+
uptake in pancreatic islets. (A),
glibenclamide (20 μM) (B), 1 (20 μM) and (C), Diazoxide (250
μM) were present in the last 15 min of pre-incubation and during
the incubation period. Pre-incubation time: 60 min; incubation time:
10 min. Means ± S.E.M. of 3 independent experiments. aP 0.05,
bp 0.01, cp 0.001 compared to control group.
# Compared to 1
group.
Fig. (3). Involvement of L-type VDCC and intracellular calcium
stocks in the stimulatory effect of acylhydrazone 1 on 45
Ca2+
uptake
in pancreatic islets. (A), Nifedipine (1 μM) (B), U-73122 (1 M)
were present in the last 15 min of pre-incubation and during the
incubation period. Pre-incubation time: 60 min; incubation time: 10
min. Mean ± S.E.M. of 3 independent experiments. p 0.001 com-
pared to control group. #
Compared to 1 group.
Effect of Acylhydrazones on LDH Activity
The tissue viability was analyzed through the LDH
activity, measuring the release of the cytosolic enzyme
lactate dehydrogenase into the serum by damaged tissue. The
oral gavage treatment with the acylhydrazones 1 and 4 for
180 min was not able to induce tissue damage as can be ob-
served in (Table 6). Also, in similar concentration range of
acylhydrazones low level of cytotoxicity was reported by
Tian et al. [66].
CONCLUSION
In a whole, the in vivo and in vitro effects of the acylhy-
drazone 1 show at least dual strategic targets involved in
glucose homeostasis, that is, the pancreas and skeletal mus-
cle. The highlighted mechanism of action described herein
A
0
15
30
45
Control1 20 μMGlibenclamide 20 μMGlibenclamide 20 μM + 1 20 μM
c
ac
a
pmol
45C
a2+/μ
g pr
otei
n
#
B
0
15
30
45
60
75
ControlGlibenclamide 20 μM1 20 μM1 20 μM + Glibenclamide 20 μM
b b
a
pmol
45C
a2+/μ
g pr
otei
n
C
0
15
30
45
Control1 20 μMDiazoxide 250 μMDiazoxide 250 μM + 1 20 μM
c
a
pmol
45C
a2+/μ
g pr
otei
n
#
A
0
15
30
45
Control1 20 μMNifedipine 1 μMNifedipine 1 μM + 1 20 μM
c
a
pmol
45C
a2+/μ
g pr
otei
n
c#
B
0
15
30
45
60
Control1 20 μMU-73122 1 μMU-73122 1 μM + 1 20μM
c
pmol
45C
a2+/μ
g pr
otei
nb#

2056 Current Topics in Medicinal Chemistry, 2012, Vol. 12, No. 19 Frederico et al.
Fig. (4). Effect of acylhydrazone 1 on fasted serum glucose levels in diabetic Wistar rats, means ± SEM, n= 6, cp 0.001 compared to zero
time.
Table 4. Effect of Acylhydrazones 1 and 4 (10 mg/kg) on Glycogen Content in Rats.
Group
Hyperglycemic 1 4
Muscle 7.72 ± 0.40 16.81 ± 0.61c 18.76 ± 2.16
c
Liver 14.60 ± 1.27 11.71 ± 2.81 16.10 ± 1.23
Each value represents the mean ± S.E.M. of four determinations. cP 0.001 compared to hyperglycemic control group. Results expressed in mg/g of tissue.
strongly suggests both an insulin secretagogue and insuli-
nomimetic role for this compound. The effects of the acyl-
hydrazone 1 on serum glucose lowering and insulin secre-
tion, and its acute stimulatory action on calcium uptake in
pancreatic islets, glycogen accumulation and glucose uptake
in skeletal muscle on in vivo treatment, did not indicate cyto-
toxicity. Finally, the acylhydrazone 1 appears to represent a
potentially useful candidate for diabetes therapy in the near
future.
Table 5. Effect of 20 M Acylhydrazone 1 on Glucose Uptake
Glucose uptake (nmol glysosyl units/ mg protein)
Control 0.22 ± 0.02
1 0.77 ± 0.18b
Each value represents the mean ± S.E.M. of three determinations. bP 0.01, compared
to control group.
Table 6. Effect of Acylhydrazones 1 and 4 on Extracellular
LDH Activity in Serum
Group LDH % of Control Time (180 min)
1 115.2 ± 11.9
4 116.6 ± 5.8
1 and 4 (10 mg/kg). Each value represents the mean ± S.E.M. of four determinations.
CONFLICT OF INTEREST
The author(s) confirm that this article content has no con-
flicts of interest.
ACKNOWLEDGEMENTS
This study was supported by grants and fellowships from
the governmental agencies Conselho Nacional de Desenvol-
vimento Científico e Tecnológico-Brasil (CNPq), Coordena-
ção de Pessoal de Nível Superior (CAPES-PPG-Bioquímica)
and Fundação de Amparo à Pesquisa do Estado de Santa
Catarina (FAPESC). We also thank to Louise D. Chiaradia
for the support chemical. M. J. S. F is registered on the PPG-
Bioquímica. C. P. M. (CNPq fellowship).
ABBREVIATIONS
DM = Diabetes Mellitus
T1DM = Diabetes type1
T2DM = Diabetes type 2
U-73122 = 1-[6-[((17 )-3-methoxyestra-1,3,5[10]-
trien-17-yl)amino]hexyl]-1H-pyrrole-2,5-
dione
II = Insulinogenic index
AUCs = Areas under the response curves
KRb-HEPES = Krebs Ringer bicarbonate buffer with
HEPES
LDH = Lactate dehydrogenase
0 1 2 3
100
250
400
550Acylhydrazone 1 (10 mg/kg)
c
Time (h)
Seru
m g
luco
se le
vel (
mg/
dL)

Antihyperglycemic and Hypoglycemic Effects of Acylhydrazones Current Topics in Medicinal Chemistry, 2012, Vol. 12, No. 19 2057
PLC = Phospholipase C
VDCC = L-type voltage-dependent calcium channel
K+-ATP = ATP-dependent K
+ channel
REFERENCES
[1] World Health Organization. Diabetes programe: Country and
regional data on diabetes.
http://www.who.int/diabetes/facts/world_figures/en/ . [Accessed
on jun 2012].
[2] Quaseem, A.; Vijan, S.; Snow, V.; Cross, J.T.; Weiss, K.B.;
Owens, D.K. Clinical Efficacy. Assessment Subcommittee of the
American College of Physicians. Ann. Intern. Med., 2007, 18, 417-
422.
[3] Hjelm, K.; Bard, K.; Nyberg, P.; Apelqvist, J. Religious and
cultural distance in beliefs about health and illness in women with
diabetes mellitus of different origin living in Sweden. Int. J. Nurs. Stud., 2003, 40, 627-643.
[4] Kahn, S.E. The relative contributions of insulin resistance and
beta-cell dysfunction to the pathophysiology of type 2 diabetes.
Diabetologia., 2003, 46, 3-19.
[5] Rhodes, C.J. Type 2 diabetes - a matter of beta-cell life and death?
Science., 2005, 21: 380-384.
[6] Yki-Jarvinen, H. Role of insulin resistance in the pathogenesis of
NIDDM. Diabetologia., 1995, 38, 1378-1388.
[7] Saltiel, A.R. New perspectives into the molecular pathogenesis and
treatment of type 2 diabetes. Cell.,2001, 23, 517-529.
[8] Sladek, R.; Rocheleau, G.; Rung, J.; Dina, C.; Shen, L.; Serre, D.;
Boutin, P.; Vincent, D.; Belisle, A.; Hadjadj, S.; Balkau, B.;
Heude, B.; Charpentier, G.; Hudson, T.J.; Montpetit, A.;
Pshezhetsky, A.V.; Prentki, M.; Posner, B.I.; Balding, D.J.; Meyre,
D.; Polychronakos, C.; Froguel, P. A genome-wide association
study identifies novel risk loci for type 2 diabetes. Nature., 2007,
22, 881-885.
[9] Karam, J.H. Type II diabetes and syndrome X. Pathogenesis and
glycemic management. Endocrinol. Metab.Clin. North. Am., 1992,
21, 329-350.
[10] Zimmet, P.Z. Diabetes epidemiology as a tool to trigger diabetes
research and care. Diabetologia., 1999, 42, 499-518.
[11] Baggio, L.L.; Drucker, D.J. Therapeutic approaches to preserve
islet mass in type 2 diabetes. Annu. Rev. Med., 2006, 57, 265-281.
[12] Atkinson, M.A.; Eisenbarth, G.S. Type 1 diabetes: new perspec-
tives on disease pathogenesis and treatment. Lancet., 2001, 21, 221-229.
[13] Hanley, S. Pancreatic -cell mass as a pharmacologic target in
diabetes. Mcgill. J. Med., 2009, 16, 51.
[14] Damazio, R.G.; Zanatta, A.P.; Cazarolli, L.H.; Mascarello, A.;
Chiaradia, L.D.; Nunes, R,J.; Yunes, R.A.; Silva,
F.R.Nitrochalcones: potential in vivo insulin secretagogues.
Biochimie., 2009, 91(11-12),1493-1498.
[15] Sehata, I.A.; Nasr, M.N.; El-Subbagh, H.I.; Gineinah, M.M.; Khei-
ra, S.M. Synthesis and biological testing of certain 1,3,4-
oxadiazole and 1,2,4-triazole derivatives as potential antimicrobial
agents. Sci Pharm., 1996, 64, 133.
[16] Borchhardt, D.M.; Mascarello, A.; Chiaradia, L.D.; Nunes, R.J.;
Oliva G.; Yunes R.A.; Adriano, D.; Andricopulo, A.D. Biochemi-
cal evaluation of a series of synthetic chalcone and hydrazide deri-
vatives as inhibitors of cruzain from trypanosoma cruzi. J. Braz. Chem. Soc., 2009, 21, 142-150.
[17] Fraga, A.G.; Rodrigues, C.R.; de Miranda, A.L.; Barreiro, E.J.;
Fraga, C.A. Synthesis and pharmacological evaluation of novel
heterotricyclic acylhydrazone derivatives, designed as PAF
antagonists. Eur. J. Pharm. Sci., 2000, 11, 285-290.
[18] Fraga, C.A.; Barreiro, E.J. Medicinal chemistry of N-
acylhydrazones: new lead-compounds of analgesic,
antiinflammatory and antithrombotic drugs. Curr. Med. Chem., 2006, 13, 167-198.
[19] Silva, A.G.; Zapata-Sudo, G.; Kummerle, A.E.; Fraga, C.A.; Bar-
reiro, E.J.; Sudo, R.T. Synthesis and vasodilatory activity of new
N-acylhydrazone derivatives, designed as LASSBio-294 analo-
gues. Bioorg. Med. Chem., 2005, 16, 3431-3437.
[20] Wang, Z.; Lu, Y.; Seibel, W.; Miller, D.D.; Li, W. Identifying
novel molecular structures for advanced melanoma by ligand-
based virtual screening. J. Chem. Inf. Model., 2009, 49(6): 1420-
1427.
[21] UFSC. Universidade Federal de Santa Catarina. Acil-hidrazonas
como protótipo de novos fármacos para leucemia linfóide aguda
(LLA). B. R. Patent 0000221109711624, November 25, 2011.
[22] Anello, M.; Gilon, P.; Henquin, J.C. Alterations of insulin secre-
tion from mouse islets treated with sulphonylureas: perturbations
of Ca2+
regulation prevail over changes in insulin content. Br. J. Pharmacol., 1999, 127,1883-1891.
[23] Zanatta, L.; Zamoner, A.; Gonçalves, R.; Zanatta, A.P.; Bouraïma-
Lelong, H.; Carreau, S.; Silva, FR. 1 ,25-Dihydroxyvitamin D(3)
signaling pathways on calcium uptake in 30-day-old rat Sertoli
cells. Biochemistry., 2011, 50, 10284-10292.
[24] Lebrun, P.; Antoine, M.H.; Ouedraogo, R.; Kane, C.; Dunne, M.;
Hermann, M.; Herchuelz, A.; Masereel, B.; Delarge, J.; de Tullio,
P.; Pirotte B. Activation of ATP-dependent K+ channels and inhi-
bition of insulin release: effect of BPDZ 62. J. Pharmacol. Exp. Ther., 1996, 277, 156–162.
[25] Varley, H.; Gowenlock, A.H.; Bell, M. Pratical Biochemistry; Eds. Heinemann IW: 5
rd ed.; London,Vol.1, 1976, pp. 389-391.
[26] Lacy, P.E.; Kostianovsky, M. Method for the isolation of intact
islets of Langerhans from the rat pancreas. Diabetes., 1967,16, 35-
39.
[27] Grillo, M.L.; Jacobus, A.P.; Scalco, R.; Grillo, M.L.; Jacobus,
A.P.; Scalco, R.; Amaral, F.; Rodrigues, D.O.; Loss, E.S.;
Wassermann, G.F. Testosterone rapidly stimulates insulin release
from isolated pancreatic islets through a non-genomic dependent
mechanism. Horm. Metab. Res., 2005, 37, 662-665.
[28] Zanatta, L.; Zamoner, A.; Gonçalves, R.; Zanatta, A.P.; Bouraïma-
Lelong, H.; Bois, C.; Carreau, S.; Silva, F.R. Effect of 1 ,25-
dihydroxyvitamin D3 in plasma membrane targets in immature rat
testis: ionic channels and gamma-glutamyl transpeptidase activity.
Arch. Biochem. Biophy., 2011, 515, 46–53.
[29] Rosso, A.; Pansera, M.; Zamoner, A.; Zanatta, L.; Bouraïma-
Lelong, H.; Carreau, S.; Silva, F.R. 1 ,25(OH)2-Vitamin D3 sti-
mulates rapid plasma membrane calcium influx via MAPK activa-
tion in immature rat Sertoli cells. Biochimie., 2012, 94, 146-154.
[30] Batra, S.; Sjögren, C. Effect of estrogen treatment on calcium
uptake by the rat uterine smooth muscle. Life. Sci., 1983, 32, 315-
319.
[31] Lowry, O.H.; Rosebrough, N.J.; Farr, A,L.; Randall, R.J. Protein
measurement with the Folin phenol reagent. J. Biol. Chem., 1951,
193, 265-275.
[32] Krisman CR. A method for the colorimetric estimation of glycogen
with iodine. Anal. Biochem., 1962, 4, 14-22.
[33] Cazarolli, L.H., Kappel, V.D., Pereira, D.F., Moresco, H.H.,
Brighente, I.M.C., Pizzolatti, M.G., Silva, F.R.M.B. Acute anti-
hyperglycemic role of crude extract, fractions and flavonoids from
Averrhoa carambola. Fitoterapia, 2012, 83, 1176 – 1183.
[34] Cazarolli, L.H., Folador, P., Moresco, H.H., Brighente, I.M.C.,
Pizzolatti, M.G., Silva, F.R.M.B. Stimulatory effect of apigenin-6-
-L-fucopyranoside on insulin secretion and glycogen synthesis.
Eur. J. Med.Chem., 2009, 44, 4668-4673.
[35] Hutchesson, A.; Preece, M.A.; Gray, G.; Green, A. Measurement
of lactate in cerebrospinal fluid in investigation of inherited meta-
bolic disease. Clin. Chem. 1997, 43, 158-161.
[36] Troeberg, L.; Chen, X.; Flaherty, T.M.; Morty, R.E.; Cheng, M.;
Hua, H.; Springer, C.; McKerrow, J,H.; Kenyon, G,L.; Lonsdale-
Eccles, J,D.; Coetzer, T.H.; Cohen, F.E. Chalcone, acyl hydrazide,
and related amides kill cultured Trypanosoma brucei brucei. Mol. Med. 2000, 6, 660-669.
[37] Chida, A.S.; Vani, P.V.S.N.; Chandrasekharam, M.; Srinivasan,
R.; Singh, A.K. Synthesis of 2,3-dimethoxy-5-methyl-1,4-benzo
quinone: A key fragment in coenzyme-Q series. Synth. Commun., 2001, 31, 657.
[38] Sakamoto, K.; Yonoki, Y.; Fujioka, T.; Matsumura, M.; Mitsuta,
Y.; Sano, M.; Saito, M.; Nakahara, T.; Ishii, K. Disappearance of
glibenclamide-induced hypoglycemia in Wistar-Kyoto rats. Biol Pharm Bull 2006, 29,574-576.
[39] León-Reyes, M.R.; Castañeda-Hernández, G.; Ortiz, M.I.
Pharmacokinetic of diclofenac in the presence and absence of
glibenclamide in the rat. J Pharm Pharm Sci 2009,12, 280-287.
[40] Alberton, E.H.; Damazio, R.G.; Cazarolli, L.H.; Chiaradia, L.D.;
Leal, P.C.; Nunes, R.J.;, Yunes, R.A.; Silva, F.R. Influence of
chalcone analogues on serum glucose levels in hyperglycemic rats.
Chem. Biol. Interact. 2008, 171(3), 355-362.

2058 Current Topics in Medicinal Chemistry, 2012, Vol. 12, No. 19 Frederico et al.
[41] Damazio, R.G.; Zanatta, A.P.; Cazarolli, L.H.; Chiaradia, L.D.;
Mascarello, A.; Nunes, R.J.; Yunes, R.A.; Silva, F.R. Antihyper-
glycemic activity of naphthylchalcones. Eur. J. Med. Chem. 2010,
45, 1332-1337.
[42] Meyer, M.; Chudziak, F.; Schwanstecher, C.; Schwanstecher, M.;
Panten, U. Structural requirements of sulphonylureas and
analogues for interaction with sulphonylurea receptor subtypes. Br. J. Pharmacol., 1999, 128, 27–34.
[43] Proks, P.; Reimann, F.; Green, N.; Gribble, F.; Ashcroft, F.
Sulfonylurea stimulation of insulin secretion. Diabetes., 2002, 51, S368—S76.
[44] Gribble, F.M.; Reimann, F. Differential selectivity of insulin
secretagogues: mechanisms, clinical implications, and drug
interactions. J. Diabetes. Complications., 2003, 17, 11–15.
[45] Bryan, J.; Crane, A.; Vila-Carriles.; WH, Babenko AP, Aguilar-
Bryan L. Insulin secretagogues, sulfonylurea receptors and
K(ATP) channels. Curr. Pharm. Des., 2005, 11, 2699-26716.
[46] Barg, S.; Renstrom, E.; Berggren, P.O.; Barg, S.; Renström, E.;
Berggren, P.O.; Bertorello, A.; Bokvist, K.; Braun, M.; Eliasson,
L.; Holmes, W.E.; Köhler, M.; Rorsman, P.; Thévenod, F.The
stimulatory action of tolbutamide on Ca2+
-dependent exocytosis in
pancreatic cells is mediated by a 65-kDa mdr-like P-
glycoprotein. Proc. Natl. Acad. Sci. U S A., 1999, 96, 5539–5544. [47] Renstrom, E.; Barg, S.; Thevenod, F.; Rorsman, P. Sulfonylurea-
mediated stimulation of insulin exocytosis via an ATP-sensitive K+
channel-independent action. Diabetes. 2002, 51, 33–36.
[48] Yang, S.N.; Berggren, P.O. CaV2.3 channel and PKC: new players
in insulin secretion. J. Clin. Invest. 2005, 115, 16-20.
[49] Olofsson, C.S.; Salehi, A.; Holm, C.; Rorsman, P. Palmitate
increases L-type Ca2+
currents and the size of the readily releasable
granule pool in mouse pancreatic beta-cells. J. Physiol., 2004, 15, 935-948.
[50] Findlay, I.; Ashcroft, F.M.; Kelly, R.P.; Rorsman, P.; Petersen,
O.H.; Trube, G. Calcium currents in insulin-secreting -cells. Ann. NY. Acad. Sci., 1989, 560, 403-409.
[51] Rajan, A.S.; Aguilar-Bryan, L.; Nelson, D.A.; Yaney, G.C.; Hsu,
W.H.; Kunze, D.L.; Boyd, A.E 3rd. Ion channels and insulin
secretion. Diabetes. Care., 1990, 13, 340-363.
[52] Boyd III, A.E. The role of ion channels in insulin secretion. J. Cell. Biochem., 1992, 48, 235-241.
[53] Gilon, P.; Ravier, M.A.; Jonas, J.C.; Henquin, J.C. Control mecha-
nisms of the oscillations of insulin secretion in vitro and in vivo.
Diabetes, 2002, 51, 144-151.
[54] Matschinsky, FM. Regulation of pancreatic beta-cell glucokinase:
from basics to therapeutics. Diabetes, 2002, 51, 394-404.
[55] Aguilar-Bryan, L.; Bryan, J. Molecular biology of adenosine
triphosphate sensitive potassium channels. Endocr. Rev. 1999, 20, 101-135.
[56] Ashcroft, F.M.; Gribble, F.M. ATP-sensitive K+
channels and
insulin secretion: their role in health and disease. Diabetologia., 1999, 42, 903-919.
[57] Seino, S.; Iwanaga, T.; Nagashima, K.; Miki, T. Diverse roles of
K(ATP) channels learned from Kir6.2 genetically engineered mice.
Diabetes., 2000, 49, 311-318.
[58] Gribble, F.M.; Tucker, S.J.; Ashcroft, F.M. The interaction of
nucleotides with the tolbutamide block of K-ATP currents: a
reinterpretation. J. Physiol., 1997, 504, 35-45.
[59] Yang, S.N.; Berggren, P.O. The role of voltage-gated calcium
channels in pancreatic beta-cell physiology and pathophysiology.
Endocr. Rev., 2006, 27, 621-676.
[60] Henquin, J.C.; Ravier, M.A.; Nenquin, M.; Jonas, J.C.; Gilon, P.
Hierarchy of the beta-cell signals controlling insulin secretion.
Eur. J. Clin. Invest. 2003, 33, 742–50.
[61] MacDonald, P.E.; Wheeler, M.B. Voltage-dependent K+ channels
in pancreatic beta cells: role, regulation and potential as therapeu-
tic targets. Diabetologia., 2003, 46, 1046 -1062.
[62] Jing, X.; Li, D.Q.; Olofsson, C.S.; Salehi, A.; Surve, V.V.;
Caballero, J.; Ivarsson, R.; Lundquist, I.; Pereverzev, A.;
Schneider, T.; Rorsman, P.; Renström, E. CaV2.3 calcium channels
control second-phase insulin release. J. Clin. Invest. 2005, 115, 146-154.
[63] Federiuk, I.F.; Casey, H.M.; Quinn, M,J.; Wood, M.D.; Ward,
W.K. Induction of type-1 diabetes mellitus in laboratory rats by
use of alloxan: route of administration, pitfalls, and insulin
treatment. Comp. Med. 2004, 54, 252-257.
[64] Cazarolli, L.H.; Zanatta, L.; Alberton, E.H.; Figueiredo, M.S.;
Folador, P.; Damazio, R.G.; Pizzolatti, M.G.; Silva, F.R. Flavon-
oids: cellular and molecular mechanism of action in glucose ho-
meostasis. Mini. Rev. Med. Chem., 2008, 8, 1032-1038.
[65] Kawabata, K.; Sawada, K.; Ikeda, K.; Fukuda, I.; Kawasaki, K.;
Yamamoto, N.; Ashida, H. Prenylated chalcones 4-
hydroxyderricin and xanthoangelol stimulate glucose uptake in
skeletal muscle cells by inducing GLUT4 translocation. Mol. Nutr. Food Res., 2011, 55, 467-475.
[66] Tian, B.; He, M.; Tan, Z.; Tang, S.; Hewlett, I.; Chen, S.; Jin, Y.;
Yang, M. Synthesis and antiviral evaluation of new N-
acylhydrazones containing glycine residue. Chem. Biol. Drug Des., 2011, 77, 189-198.
Received: July 05, 2012 Accepted: October 11, 2012




















