
Insulin-like growth factor binding protein-6 delays replicative senescence of human fibroblasts
28
Insulin-like growth factor binding protein-6 delays replicative senescence of human fibroblasts Lucia Micutkova a , Thomas Diener a , Chen Li b , Adelina Rogowska-Wrzesinska c , Christoph Mueck a , Eveline Huetter a , Birgit Weinberger a , Beatrix Grubeck-Loebenstein a , Peter Roepstorff c , Rong Zeng b , and Pidder Jansen-Duerr a,⁎ a Institute for Biomedical Aging Research, Austrian Academy of Sciences, Rennweg 10, A-6020 Innsbruck, Austria b Key Laboratory of Systems Biology, Institute of Biochemistry and Cell Biology, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences, Shanghai 200031, China c Protein Research Group, Department of Biochemistry and Molecular Biology, University of Southern Denmark, Campusvej 55, DK 5230 Odense M, Denmark Highlights ► Proteomic analysis of senescent secretome reveals upregulation of IGFBP-6 in fibroblasts. ► IGFBP-6 knockdown induces premature senescence in young fibroblasts. ► IGFBP-6 lentiviral overexpression delays replicative senescence in fibroblasts. Abstract Cellular senescence can be induced by a variety of mechanisms, and recent data suggest a key role for cytokine networks to maintain the senescent state. Here, we have used a proteomic LC-MS/MS approach to identify new extracellular regulators of senescence in human fibroblasts. We identified 26 extracellular proteins with significantly different abundance in conditioned media from young and senescent fibroblasts. Among these was insulin-like growth factor binding protein-6 (IGFBP-6), which was chosen for further analysis. When IGFBP-6 gene expression was downregulated, cell proliferation was inhibited and apoptotic cell death was increased. Furthermore, downregulation of IGFBP-6 led to premature entry into cellular senescence. Since IGFBP-6 overexpression increased cellular lifespan, the data suggest that IGFBP-6, in contrast to other IGF binding proteins, is a negative regulator of cellular senescence in human fibroblasts. Abbreviations IGF, insulin-like growth factor; IGFBP-6, insulin-like growth factor binding protein-6; LC-MS/ MS, liquid chromatography tandem mass spectrometry; SASP, senescence-associated secretory phenotype © 2011 Elsevier Ireland Ltd. ⁎ Corresponding author. Tel.: +43 512 583919 44; fax: +43 512 583919 8. [email protected]. This document was posted here by permission of the publisher. At the time of deposit, it included all changes made during peer review, copyediting, and publishing. The U.S. National Library of Medicine is responsible for all links within the document and for incorporating any publisher-supplied amendments or retractions issued subsequently. The published journal article, guaranteed to be such by Elsevier, is available for free, on ScienceDirect. Sponsored document from Mechanisms of Ageing and Development Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479. Sponsored Document Sponsored Document Sponsored Document
-
Upload
southerndenmark -
Category
Documents
-
view
1 -
download
0
Transcript of Insulin-like growth factor binding protein-6 delays replicative senescence of human fibroblasts

Insulin-like growth factor binding protein-6 delays replicativesenescence of human fibroblasts
Lucia Micutkovaa, Thomas Dienera, Chen Lib, Adelina Rogowska-Wrzesinskac, ChristophMuecka, Eveline Huettera, Birgit Weinbergera, Beatrix Grubeck-Loebensteina, PeterRoepstorffc, Rong Zengb, and Pidder Jansen-Duerra,⁎
aInstitute for Biomedical Aging Research, Austrian Academy of Sciences, Rennweg 10, A-6020Innsbruck, AustriabKey Laboratory of Systems Biology, Institute of Biochemistry and Cell Biology, ShanghaiInstitutes for Biological Sciences, Chinese Academy of Sciences, Shanghai 200031, ChinacProtein Research Group, Department of Biochemistry and Molecular Biology, University ofSouthern Denmark, Campusvej 55, DK 5230 Odense M, Denmark
Highlights► Proteomic analysis of senescent secretome reveals upregulation of IGFBP-6 in fibroblasts. ►IGFBP-6 knockdown induces premature senescence in young fibroblasts. ► IGFBP-6 lentiviraloverexpression delays replicative senescence in fibroblasts.
AbstractCellular senescence can be induced by a variety of mechanisms, and recent data suggest a key rolefor cytokine networks to maintain the senescent state. Here, we have used a proteomic LC-MS/MSapproach to identify new extracellular regulators of senescence in human fibroblasts. Weidentified 26 extracellular proteins with significantly different abundance in conditioned mediafrom young and senescent fibroblasts. Among these was insulin-like growth factor bindingprotein-6 (IGFBP-6), which was chosen for further analysis. When IGFBP-6 gene expression wasdownregulated, cell proliferation was inhibited and apoptotic cell death was increased.Furthermore, downregulation of IGFBP-6 led to premature entry into cellular senescence. SinceIGFBP-6 overexpression increased cellular lifespan, the data suggest that IGFBP-6, in contrast toother IGF binding proteins, is a negative regulator of cellular senescence in human fibroblasts.
AbbreviationsIGF, insulin-like growth factor; IGFBP-6, insulin-like growth factor binding protein-6; LC-MS/MS, liquid chromatography tandem mass spectrometry; SASP, senescence-associated secretoryphenotype
© 2011 Elsevier Ireland Ltd.⁎Corresponding author. Tel.: +43 512 583919 44; fax: +43 512 583919 8. [email protected] document was posted here by permission of the publisher. At the time of deposit, it included all changes made during peerreview, copyediting, and publishing. The U.S. National Library of Medicine is responsible for all links within the document and forincorporating any publisher-supplied amendments or retractions issued subsequently. The published journal article, guaranteed to besuch by Elsevier, is available for free, on ScienceDirect.
Sponsored document fromMechanisms of Ageing andDevelopment
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

KeywordsIGFBP-6; Senescence; Secretome; Proteomics; Fibroblast
1 IntroductionDuring the process of human aging, cellular degeneration and a partial loss of physiologicalfunction occurs in various tissues. There is increasing evidence, that age-associated changesin tissue functions can be triggered by the appearance of senescent cells in various tissues,e.g. vascular tissue of elderly donors (Vasile et al., 2001). Cellular senescence is believed tobe an important mechanism for preventing the cells from becoming malignant. However,senescent cells can also have detrimental effects on their microenvironment and neighboringcells due to the deregulation of their metabolism and changed composition of secretedproteins (for review see Campisi and d’Adda di Fagagna, 2007; Chen and Goligorsky,2006). The altered secretome of senescent cells has been previously referred to as thesenescence-associated secretory phenotype (SASP; Coppe et al., 2008) and the senescence-messaging secretome (SMS; Kuilman and Peeper, 2009), respectively. The secretome ofsenescent cells is complex and includes factors that are involved in senescence-associatedproliferation arrest (IGF-I; Ferber et al., 1993 and IGFBPs; Wajapeyee et al., 2008), immuneregulation (cytokines and chemokines; Maier et al., 1990), and extracellular matrixremodeling (MMP-1 and -3; Krizhanovsky et al., 2008). Fibroblasts play a central role intissue processes, especially in the skin, not only because of their involvement in thereorganization of the extracellular matrix but also due to their role in the communicationbetween different cell types, mediated mainly through the secretion of proteins withextracellular function, such as cytokines. Senescent fibroblasts have been shown to produceless collagen and to secrete increasing amounts of matrix-degrading enzymes during agingin vitro (Campisi, 1998; Toussaint et al., 2002). This is in agreement with in vivo findings,showing that the skin of aged individuals has a decreased content of collagen, leading tochanges in skin morphology. In studies with skin biopsies from humans and non-humanprimates, an age-related increase of senescent cells was observed in vivo (Dimri et al., 1995;Herbig et al., 2006; Ressler et al., 2006), and this may account for the age-related changes inskin morphology. Therefore, identification of the nature of proteins secreted by senescentfibroblasts becomes important for a better understanding of aging processes and thedevelopment of age-related diseases.
In this communication, we used liquid chromatography tandem mass spectrometry (LC-MS/MS) proteomic technique to analyze the secretome of young and senescent human diploidfibroblasts (HDFs). We identified 26 extracellular proteins with differential abundance insupernatants of young and senescent cells. Among these, insulin-like growth factor bindingprotein-6 (IGFBP-6) was chosen for further analysis.
IGFBP-6 belongs to the family of six high affinity IGF binding proteins (IGFBP-1 to -6),which play an important role in the distribution and modulation of biological effects of IGFs(Firth and Baxter, 2002; Jones and Clemmons, 1995). IGFBP-6 is a relatively new memberof the IGFBP family, with a unique about 50-fold preferential binding affinity for IGF-IIrelative to IGF-I (Bach, 1999; Roghani et al., 1989, 1991). This feature makes it a verypotent inhibitor of IGF-II actions, which is of particular interest especially for growthinhibition of IGF-II-dependent tumors, such as neuroblastomas (Grellier et al., 1998),rhabdomyosarcomas (Gallicchio et al., 2001) and colon cancer (Kim et al., 2002). IGFBP-6is expressed predominantly in quiescent, non-dividing cells (Ewton and Florini, 1995) andduring differentiation of diverse cell types, such as neuroblastoma-derived cells or prostatecells (Chambery et al., 1998; Lipinski et al., 2005). IGF-independent effects of IGFBP-6
Micutkova et al. Page 2
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

were also reported, including modulation of cell migration (Fu et al., 2007, 2010) andinduction of apoptosis (Iosef et al., 2008). In addition to IGFBP-3 and -5, which bothpossess functional nuclear localization signals (NLS; Schedlich et al., 2000), NLS was alsoidentified in the C-terminal domain of IGFBP-6. It was shown, that IGFBP-6 can be activelyimported into the nuclei of rhabdomyosarcoma and HEK-293 cells, and nucleartranslocation seems to be required for its IGF-independent apoptotic effects (Iosef et al.,2008). Many studies show regulation of IGFBP-6 predominantly in conditions consistentwith its growth inhibitory and proapoptotic effects (for review see Bach, 2005). However,there are studies indicating that in some situations, IGFBP-6 is associated withproproliferative (Schmid et al., 1999) and/or protective effects (Beilharz et al., 1998);however, these IGFBP-6 actions are not clearly understood.
Insulin/IGF signaling plays a major role in determining the rate of aging in many species(Bartke, 2005; Rincon et al., 2005), although the precise role of IGFBPs in this processremains to be elucidated. In our previous work, we studied the role of IGFBP-3 in thesenescence response of endothelial cells (Hampel et al., 2006; Muck et al., 2008), indicatingthat IGFBP-3 acts as an antiproliferative and premature senescence-inducing protein.Accumulation of IGFBP-3 in conditioned medium of senescent human fibroblasts was alsoreported (Goldstein et al., 1991; Moerman et al., 1993), where it contributes to the growtharrest phenotype of these cells (Grigoriev et al., 1995; Hampel et al., 2004, 2005). The roleof IGFBP-6 in cellular senescence was not studied yet. However, increased levels ofIGFBP-6 protein were detected in conditioned medium of human fibroblasts madeprematurely senescent by exposure to H2O2 (Xie et al., 2005). In the same work,upregulation of IGFBP-6 protein was also observed in the plasma of aging mice and ofyoung mice treated with doxorubicin, which leads to a premature senescence phenotype. Inthis communication, we focused on the role of IGFBP-6 in cellular senescence in HDFs. Wedepleted and/or overexpressed IGFBP-6 in primary human fibroblasts using a lentiviralapproach. Our data suggest that IGFBP-6 depletion has detrimental effects on HDFs andleads to decreased proliferation, increased cell death and early occurrence of senescence. Incontrast, IGFBP-6 overexpression had protective effects on HDFs and markedly delayed theonset of senescence.
2 Materials and methods2.1 Chemicals
Urea, thiourea, tris, SDS, glycine, acrylamide, bis-acrylamide, TEMED, bromophenole blue,CHAPs, dithiothreitol (DTT) and iodoacetamide (IAA) were purchased from Bio-Rad.Sequencing grade trypsin was purchased from Promega. All other chemicals were purchasedfrom Sigma unless indicated otherwise.
2.2 Cell cultureTwo different pools of human primary diploid fibroblasts (HDFs) were obtained fromATCC (American Type Culture Collection). Each pool consisted of HDFs isolated fromseveral different newborn donors. Cells were cultivated in Dulbecco's modified Eagle'smedium (DMEM; Sigma), supplemented with 10% heat-inactivated fetal calf serum (FCS;Biochrom), 4 mM L-glutamine and 100 U/ml penicillin with 0.1 mg/ml streptomycin (GibcoInvitrogen). HDFs were serially passaged until they reached replicative senescence.Population doublings (PDL) were estimated using the following equation:n = (log10 F − log10 I)/0.301 (with n = population doublings, F = number of cells at the endof one passage, I = number of cells that were seeded at the beginning of one passage). Afterroughly 45 population doublings, the cells reached growth arrest. The maximal number ofpopulation doublings achieved by untreated HDF was set 100%, and used to calculate the
Micutkova et al. Page 3
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

percentage of lifespan completed for each culture. In particular, this procedure was used todetermine the percentage of elapsed lifespan at the time when cells were transduced withvectors for IGFBP-6 overexpression and knockdown, respectively (see text). The senescentstatus was verified by in situ staining for senescence associated-β-galactosidase as described(Dimri et al., 1995). For production of lentiviral particles, HEK-293FT cells (humanembryonic kidney 293 cells expressing the large T-antigen of SV40; Invitrogen) weremaintained in DMEM (10% not-heat-inactivated FCS, 4 mM L-glutamine, 100 U/mlpenicillin, 0.1 mg/ml streptomycin). U-2OS human osteosarcoma cells used for estimationof virus particle titer and the knockdown efficiency of individual IGFBP-6 shRNAs, wereobtained from ATCC (American Type Culture Collection) and maintained in DMEM (10%heat-inactivated FCS, 4 mM L-glutamine, 100 U/ml penicillin, 0.1 mg/ml streptomycin).Cells were grown in an atmosphere of 5% CO2 at 37 °C and were subcultured bytrypsinization with 0.05% trypsin–EDTA (Gibco Invitrogen) every 3–4 days.
2.3 Production of cellular supernatantsYoung (18% of lifespan completed) and replicatively senescent (93% of lifespan completed)HDFs from pool 1 were seeded on 145 cm2 cell culture dishes in 10% FCS DMEM at thedensity of 2.5 × 106. After 24 h cells were washed twice with PBS and incubated withserum-free DMEM for 5 h to get rid of the residual FCS. Then the medium was exchangedand fresh serum-free DMEM was added to the cells. After 72 h the supernatants wereharvested and filtered with a 0.45 μm Steritop Filter (Millipore). Supernatants were furtherconcentrated in a Stericup filtration column with cut-off of 5 kDa (Millipore) and proteinswere precipitated by a methanol/chloroform precipitation (Wessel and Flugge, 1984).Protein pellets were freeze-dried and stored at −80 °C. The whole procedure was repeated 3times, in order to produce 3 independent technical replicates for both young and senescentHDFs.
2.4 Protein sample preparationThe freeze-dried samples were solubilized in cold lysis buffer (8 M urea, 4% (v/v) CHAPS,40 mM Tris–base), then centrifuged under 4 °C, 15,000 × g for 60 min to remove insolublefraction and transferred to a new tube. The protein concentration of each sample wasquantified using Bradford kit (Bio-Rad). For each sample, about 100 μg of protein wassubjected to in-solution digestion. The samples were reduced for 2.5 h at 37 °C by additionof 1 M dithiothreitol (DTT) to a final concentration of 2 mM DTT, and then alkylated for45 min at room temperature by addition of 1 M iodoacetamide (IAA) to a final concentrationof 10 mM IAA. Samples were precipitated using 1 ml of 50% acetone, 49.9% ethanol, 0.1%acetic acid and incubated at −20 °C for 16 h. The samples were then centrifuged at15,000 × g, 4 °C for 45 min and the pellets were collected. The pellets were further washedtwice with pure acetone and pure ethanol separately and repeated centrifugation. Finally, thepellets were resuspended in 100 mM NH4HCO3 and digested using 2 μg of modified trypsin(Promega) at 37 °C for 20 h. Before mass spectrometry analysis, the peptide mixtures wereultra-filtered using 10 kDa cut-off centrifugal filter units (Millipore) to remove trypsin andother undigested proteins.
2.5 Liquid chromatography tandem mass spectrometry (LC-MS/MS) analysisThe peptide mixtures from each sample were separated by reverse phase high performanceliquid chromatography RP-HPLC followed by tandem mass spectrometry analysis. RP-HPLC was performed on a surveyor LC system (Thermo Finnigan). The C18 column (RP,180 μm × 150 mm) was obtained from Column Technology Inc. The pump flow was split1:120 to achieve a column flow rate of 1.5 nL/min. Mobile phase A was 0.1% formic acid inwater, and mobile phase B was 0.1% formic acid in acetonitrile. The tryptic peptidemixtures were eluted using a gradient of 2–98% B over 180 min. The LC system was
Micutkova et al. Page 4
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

interfaced to a LTQ linear ion trap mass spectrometer (Thermo Finnigan) using electro-spray ionization operated in positive mode. The mass spectrometer was operated in a data-dependent mode to automatically switch between MS and MS/MS acquisition. Following afull ion scan (from m/z 400–2000) the 10 most intense precursor ions with charge stateslarger than +1 were selected for fragmentation. Former target ions selected for fragmentationwere dynamically excluded with the following settings: repeat count: 2; repeat duration:0.5 min; exclusion duration: 1.5 min. The normalized collision energy was 35.0.
2.6 Data processingAll acquired MS/MS spectra were processed using Bioworks 3.1 (Thermo Finnigan) andsearched against the Human International Protein Index protein sequence database (version3.39, www.ebi.ac.uk/IPI), using the TurboSEQUEST program. All cysteine residues weresearched as carboxamidomethycystein (+57.02 Da). Homemade Buildsummary softwarewas used to delete the redundant data as previously reported (Tang et al., 2007).Additionally, to estimate the rate of incorrect identifications (false positives), all the filteredspectra were subjected to database searching against a composite database containing humanprotein sequences in both the forward (correct) and reverse (incorrect) orientation.Thresholds for Xcorr according to peptide false-discovery rate (FDR) lower than 1% wereapplied, with a fixed DeltaCn of more than 0.1. Further more proteins with less than 2spectral counts were excluded and the final protein false discovery rate was lower than 5%.
2.7 Protein quantitationA newly developed proteomic strategy of label-free quantification by spectral counts wasutilized (Aebersold and Mann, 2003; Gramolini et al., 2008). Briefly, the raw peptide countsof each protein identified in each sample were used as a semi quantitative measure for theprotein relative abundance. To reduce the systematic bias from the proteomics data, thisabundance was then normalized using the “Global” normalization method (Callister et al.,2006). In this normalization method, the total raw spectral counts (T) in each LC-MS/MSrun were set to an equal number (Tconstant). For protein (i), the normalized abundance( ) in the LC-MS/MS run (j) should be computed as the following formula:
, where Xij is the original raw spectral counts for the protein. Theprotein abundance based on normalized spectral counts was then used to perform statisticalanalysis. Three independent technical replicates for each group were analyzed. Student t-testwas used to compare the differences between young (18% of lifespan completed) andsenescent (93% of lifespan completed) HDFs. p < 0.05 and fold changes >1.5 wereconsidered statistically significant.
2.8 Lentiviral IGFBP-6 knockdown and overexpressionProduction of lentiviral particles was carried out according to the manufacturer's protocol(Addgene). For lentiviral knockdown, lentiviral vector pLKO.1, containing 5 differentIGFBP-6-specific shRNAs (small-hairpin RNA) and control shRNA respectively werepurchased from Open Biosystems. Two IGFBP-6-shRNAs with the best level of knockdownwere chosen for further experiments (shRNA1: 5′-GAGAATCCTAAGGAGAGTAAA-3′;shRNA2: 5′-GCCTGCTGTTGCAGAGGAGAA-3′). For lentiviral overexpression, IGFBP-6cDNA was cloned into the pLenti6/CMV/V5-DEST Gateway system (Invitrogen). As acontrol empty vector and/or vector carrying green fluorescence protein (GFP) was used. Forpackaging of the lentivirus, HEK293FT cells were cultivated in 75 cm2 flasks to 90%confluence and transfected with a mixture of two packaging plasmids: 7.5 μg psPAX2 and2.5 μg pMD2.G (Addgene) together with 3 μg pLenti6/CMV and/or specific shRNAspLKO.1 by Lipofectamine 2000 (Invitrogen). Supernatants were harvested 48 h posttransfection. Lentiviral particles were tittered on U-2OS cells. Lentiviral infection was
Micutkova et al. Page 5
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

carried out in young (21% of lifespan completed) HDFs pool 1 upon reaching 70–80%confluence, using lentiviral particles at multiplicity of infection of 4 in presence of 8 μg/mlhexadimethrine bromide (Sigma) as a transduction enhancer. 72 h post infection, transducedcells were selected by either puromycin (500 ng/ml) in the case of knockdown or blasticidin(10 μg/ml) in the case of overexpression.
2.9 Real time-PCRTotal RNA from HDFs was isolated with RNeasy Mini kit (Qiagen). IGFBP-6 geneexpression levels in transduced cells were analyzed at 32% of lifespan completed. Inaddition, IGFBP-6 overexpressing cells were checked for IGFBP-6 gene expression also atlatter passages (90% of lifespan completed, data not shown). 1 μg of RNA was reversetranscribed using Transcriptor First Strand cDNA Synthesis Kit (Roche Applied Science)and oligo(dT) primer. Primers for amplification of IGFBP-6 (Acc. No.: NM_002178) andhousekeeper beta-2-microglobulin (B2M; Acc. No.: NM_004048) were designed usingPrimer3 software (IGFBP-6 forward: 5′-AAGGAGAGTAAACCCCAAGCA-3′, IGFBP-6reverse: 5′-TTTGAGCCCCTCGGTAGAC-3′; B2M forward: 5′-GTGCTCGCGCTACTCTCTCT-3′; B2M reverse: 5′-TCAATGTCGGATGGATGAAA-3′).The cDNA equivalent of 5 ng of RNA was applied to PCR amplification in combinationwith 15 μl of LightCycler® 480 SYBR Green I Master (Roche Applied Science), a reactionmixture including FastStart Taq DNA Polymerase and SYBR Green I dye for productvisualization. Real-time PCR was performed in triplicates for two independentmeasurements using the LightCycler® 480 Instrument (Roche Applied Science). Cyclingconditions were as follows: 95 °C for 8 min (initial denaturation step) followed by 40 cyclesof target amplification (95 °C for 15 s, 60 °C for 8 s and 72 °C for 15 s) and final melting(95 °C for 1 min, 60 °C for 30 s, 95 °C continuous with five acquisitions/°C). Crossingpoints for IGFBP-6 and housekeeper B2M were used for calculation of IGFBP-6 relativegene expression.
2.10 Western blot and ELISACells and cell supernatants from young (18% of lifespan completed) and senescent (93% oflifespan completed) HDFs were harvested. Cells were lysed in lysis buffer containing50 mM Tris–HCl, 150 mM NaCl, 1% NP-40, 0.1% SDS, 0.5% Na-deoxycholate, 0.2 mMphenylmethylsulfonyl-fluoride, 1 mM NaF, 10 μg/ml aprotinin, 10 μg/ml leupeptin (pH 7.5).Cell supernatants were centrifuged at 300 × g and IGFBP-6 levels and assayed by standardWestern blot protocol using primary mouse monoclonal anti-human IGFBP-6 antibody(R&D Systems); goat anti-human Serpin E2 antibody (R&D Systems) and rabbit polyclonalanti-human GAPDH (glyceraldehyde-3-phosphate dehydrogenase) antibody (Santa GruzBiotechnology). IGFBP-6 levels in supernatants were quantified by ELISA (HumanIGFBP-6 DuoSet; R&D Systems) according to the manufacturer's protocol. Levels ofp21Waf1/Cip1 in IGFBP-6 knockdown (32–42% of lifespan completed) and overexpressingcells (90–95% of lifespan completed) were assessed by Western blot using mouse anti-human p21Waf1/Cip1 antibody (BD Pharmingen). As a loading control, mouse monoclonalantibody for α-tubulin was used (Sigma).
2.11 Cell proliferation assayCell proliferation was assayed by the 5-bromo-2-deoxyuridine Labeling and Detection Kit I(Roche Applied Science) as described by the manufacturer. Cells were incubated with BrdUlabeling reagent for approximately 60 min. After the staining procedure, the incorporationwas visualized by fluorescence microscopy and in addition analyzed by FACS. The numberof BrdU-positive cells was expressed as the percentage of total cell number. Themeasurements in IGFBP-6 knockdown cells were performed at 32–42% of lifespancompleted, in IGFBP-6 overexpressing cells at 90–95% of lifespan completed.
Micutkova et al. Page 6
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

2.12 Assessment of apoptotic cell deathFor detection of apoptosis, Annexin V staining and propidium iodide (PI) staining, wereused. For Annexin V staining, HDFs were detached and incubated with 5 μl of Annexin V-FITC (Pharmingen BD Biosciences) in Annexin V-buffer, containing 10 mM Hepes,140 mM sodium chloride and 2.5 mM CaCl2 (pH 7.4) for 15 min. Cells were washed withPBS and Annexin V-FITC positive cells were measured using Flow cytometry (FACS). Inthe case of PI staining, cells were washed in PBS and resuspended in 0.1% propidium iodide(50 mg/ml) with 0.1% Triton X-100. The PI fluorescence of individual nuclei was measuredby FACS. The measurements in IGFBP-6 knockdown cells were performed at 32–42% oflifespan completed, in IGFBP-6 overexpressing cells at 90–95% of lifespan completed.
2.13 Staining for senescence associated-β-galactosidase (SA-β-gal)The senescent status was verified by in situ staining for SA-β-gal as described (Dimri et al.,1995). Briefly, cells were grown on 6-well cell culture dishes, washed three times with PBSand fixed with 2% formaldehyde, 0.2% glutaraldehyde in PBS for 5 min. After anotherwashing step with PBS, the cells were incubated with SA-β-gal staining solution (150 mMNaCl, 2 mM MgCl2, 5 mM potassium ferricyanide, 5 mM potassium ferrocyanide, 40 mMcitric acid, 12 mM sodium phosphate, pH 6.0, containing 1 mg/ml 5-bromo-4-chloro-3-indolyl-b-D-galactoside (X-gal)) for 24 h at 37 °C. The reaction was stopped by washing thecells with PBS. The number of SA-β-gal positive cells was calculated as the percentage oftotal cell number. The measurements in IGFBP-6 knockdown cells were performed at 32–42% of lifespan completed, in IGFBP-6 overexpressing cells at 90–95% of lifespancompleted.
2.14 Starvation-mediated cell cycle arrestYoung (21% of lifespan completed) HDFs were seeded on 6-wells in 10% FCS DMEM atthe density of 5 × 105. After 24 h cells were washed twice with PBS and incubated withserum-free DMEM for additional 24 h. Afterwards, the medium was exchanged and freshserum-free DMEM was added to the cells. After 48 h the supernatants were harvested andstored at −80 °C. IGFBP-6 levels in supernatants were quantified by ELISA (HumanIGFBP-6 DuoSet; R&D Systems) according to the manufacturer's protocol. As a control forcell cycle arrest, cell proliferation was assayed by the 5-bromo-2-deoxyuridine Labeling andDetection Kit I (Roche Applied Science) as described above.
2.15 ELISA measurements of human serum samplesIGFBP-6 levels in human serum were analyzed by ELISA (Human IGFBP-6 DuoSet; R&DSystems). The measurements were performed in 140 healthy volunteers, divided into 2groups: the old age group (age: 66–92, n = 93) and the young/middle-age group (age: 25–41,n = 47). Blood draw was approved by the local ethics committee and all participants gavetheir written informed consent.
2.16 Ingenuity™ pathways analysesYoung (21% of lifespan completed) HDF were infected with IGFBP-6 shRNA1 andshRNA2 and/or SCR as described above. 20 days after lentiviral transduction (32% oflifespan completed) total mRNA was isolated by Trizol™ reagent (Invitrogen). Affymetrixmicroarray analysis using Human Genome U133 Plus 2.0 arrays was performed by theMicroarray Facility Tübingen (www.microarray-facility.com). Results were further analyzedfor relevant signaling pathways, using Ingenuity™ Pathways Analysis software(https://analysis.ingenuity.com; Ingenuity Systems, Inc.).
Micutkova et al. Page 7
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

2.17 Statistical analysisStudent t-test was used to compare the differences between individual samples. p valuesbelow 0.05 were considered statistically significant. The graphs represent meanvalues ± standard error of mean (SEM).
3 Results3.1 Upregulation of IGFBP-6 gene expression in fibroblast senescence
To address senescence-associated changes in the secretome of human diploid fibroblasts(HDFs), supernatants from young (18% of lifespan completed) and senescent (93% oflifespan completed) human foreskin fibroblasts were harvested and subjected to LC-MS/MSanalysis. A total of 407 proteins were identified by 6 LC-MS/MS runs, and 37 differentiallyexpressed proteins were selected, for which the fold change was >1.5 and the p-value waslower than 0.05. While the majority of the proteins identified in this analysis are wellestablished as extracellular proteins (26 proteins in total, see Table 1), we also identified 10differentially regulated proteins, which according to the literature are intracellular (data notshown) and one so far undescribed protein lacking annotations. The most likely explanationfor the occurrence of intracellular proteins in cellular supernatants is leakage subsequent tocell death. An alternative possibility is that the proteins, which are primarily annotated asintracellular, may reach the supernatant by processes different from cellular leakage (e.g.ordered secretion), however, this possibility remains to be addressed. One of the proteinsidentified by differential secretome analysis was IGFBP-6, which was found upregulated inthe supernatant of senescent cells (Fig. 1). Upregulation of IGFBP-6 in the supernatant ofsenescent cells was confirmed by Western blot (Fig. 2A) and ELISA (Fig. 2B), for bothHDF pools we tested. IGFBP-6 was readily detected in supernatants, but only traces ofIGFBP-6 were found in cellular lysates (Fig. 2A), suggesting that after synthesis IGFBP-6 israpidly secreted. In order to confirm, that the upregulation of IGFBP-6 is related to cellularsenescence, and does not simply reflect cell cycle-related changes, IGFBP-6 levels wereanalyzed in cells exposed to serum starvation for 72 h, which quantitatively arrested cells inthe G(0) phase of the cell cycle but did not show any impact on IGFBP-6 expression relativeto control cells (Fig. S1).
3.2 Depletion of IGFBP-6 reduces cell proliferation and induces apoptosis andsenescence
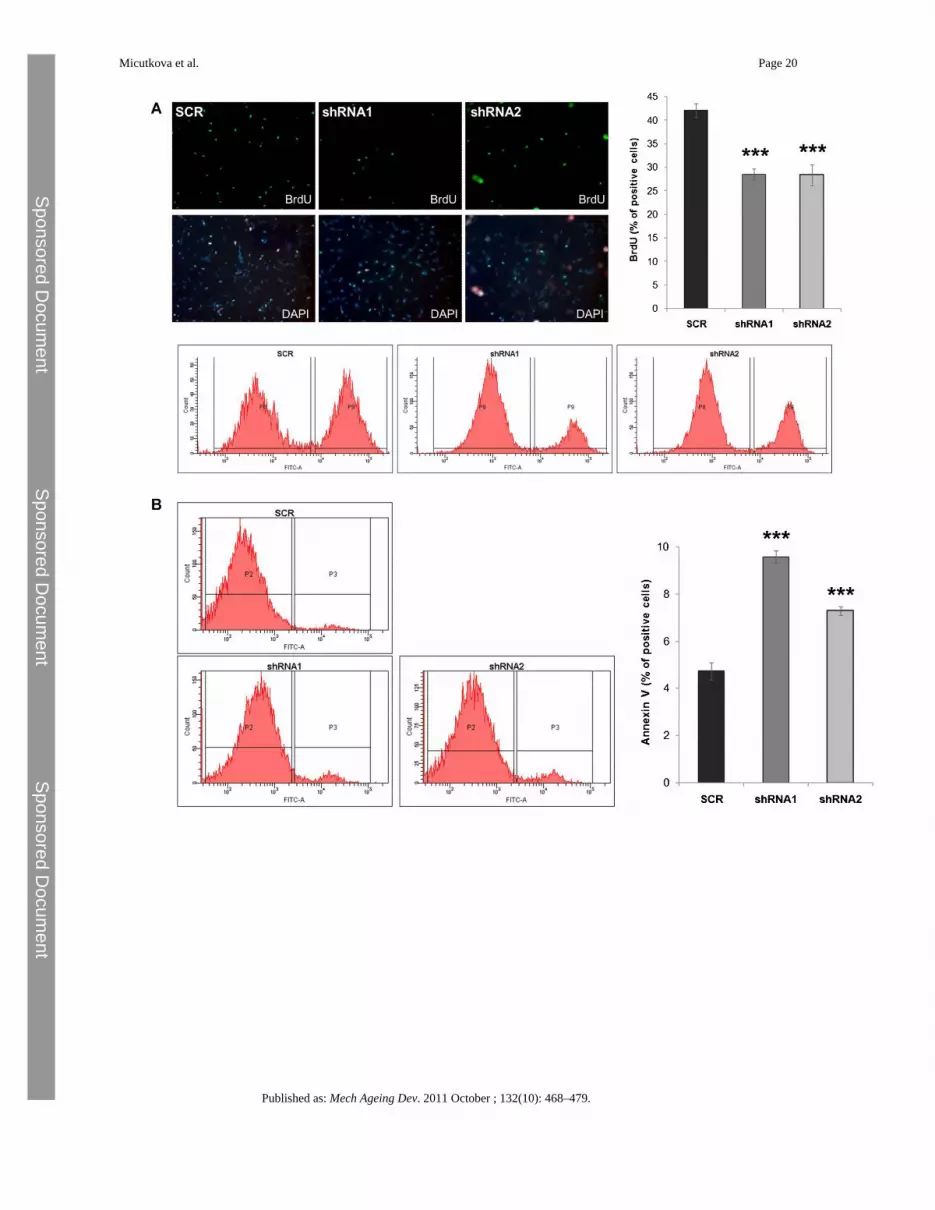
To address the functional significance of IGFBP-6 upregulation in HDF senescence, 5different commercially available shRNAs against IGFBP-6 were tested by transienttransfection in IGFBP-6 overexpressing U-2OS cells (data not shown). The two most potentshRNAs (referred to as shRNA1 and 2 below) were selected and lentiviral particles wereproduced, suitable for stable downregulation of IGFBP-6 expression in primary cells. Youngfibroblasts (21% of lifespan completed) were infected with lentiviruses expressing shRNAs1 and 2, and the effect on IGFBP-6 mRNA was determined 20 days after infection (32% oflifespan completed). We found that cells infected with both IGFBP-6 targeting shRNAsdisplayed a significant downregulation of IGFBP-6 mRNA (Fig. 3A) and protein (Fig. 3B),whereas infection with control shRNA had no effect. The analysis of transduced fibroblasts(32–42% of lifespan completed) revealed significant downregulation of cell numbers incultures infected with viruses carrying shRNA 1 and 2, whereas control shRNA had noeffect (Fig. 3C). The observed reduction in cell number could be due to a decreased rate ofcell proliferation or an increased rate of cell death, or a combination of both. When cellproliferation was assessed by bromo-deoxyuridine (BrdU) incorporation assays, knockdownof IGFBP-6 led to a significant reduction in the percentage of BrdU-positive cells (Fig. 4A),suggesting that IGFBP-6 is required for the full proliferation capacity of these cells. Whencell death was analyzed by Annexin V-staining and propidium iodide staining, we found a
Micutkova et al. Page 8
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

significant upregulation in the rate of apoptotic cells upon knockdown of IGFBP-6 (Fig. 4Band C), suggesting that IGFBP-6 might also protect fibroblasts from apoptosis. Together thedata suggest that the reduced cell numbers obtained with fibroblasts depleted for IGFBP-6 isdue to both a reduction of cell proliferation rate and an increase in cell death. To address apotential impact of IGFBP-6 knockdown on cellular senescence, cells were stained forsenescence associated-β-galactosidase (SA-β-gal). This experiment revealed a significantincrease in the percentage of SA-β-gal positive cells when IGFBP-6 was depleted by eithershRNA1 or shRNA2, whereas control shRNA had no effect (Fig. 4D). The increase in theabundance of SA-β-gal positive cells was accompanied by a strong upregulation ofp21Waf1/Cip1 protein (Fig. 4D) in IGFBP-6 depleted cells. These observations suggest thatIGFBP-6 acts to prevent early onset of cellular senescence.
3.3 IGFBP-6 overexpression prevents apoptosis and delays senescence of humanfibroblasts
The data obtained so far clearly document that IGFBP-6 is required to limit apoptotic celldeath and to ensure the full proliferation potential of diploid human fibroblasts, suggestingthat IGFBP-6 functions to delay cellular senescence in this cell type. To address thispossibility further, we constructed lentiviral vectors for IGFBP-6 overexpression andinfected young (21% of lifespan completed) HDFs with lentiviruses carrying this vector. Asa control, lentiviruses with empty vector and GFP expressing vector were used. A few daysafter infection (32% of lifespan completed), a significant upregulation of IGFBP-6 mRNA(Fig. 5A) and protein (Fig. 5B) was observed. When infected cells were further passaged,the control cells started to reach senescence-associated growth arrest about 90 days afterinfection, whereas the IGFBP-6 expressing cells continued to proliferate. At the end of thelifespan, overexpression of IGFBP-6 enabled the cells to perform 4–5 additional populationdoublings (Fig. 5C). This was accompanied by a significant increase in the cell proliferationrate as measured by BrdU incorporation (Fig. 5D) and a significant decrease in apoptoticcell death determined by Annexin V-staining (Fig. 5E) and propidium iodide staining (Fig.5F) at 90–95% of lifespan completed. 120 days after infection (ca. 95% of lifespancompleted), nearly all the cells in the controls stained SA-β-gal positive, whereas roughly50% of the IGFBP-6 expressing cells stained still SA-β-gal negative (Fig. 5G) and displayedreduced p21Waf1/Cip1 protein expression relative to controls (Fig. 5G). Together these datafully support the conclusion from the previous experiments, namely that IGFBP-6 can delayentry into the senescent phenotype in human diploid fibroblasts.
3.4 Gene expression changes induced by IGFBP-6 depletionNext we addressed potential mechanisms, underlying the actions of IGFBP-6. RNA wasprepared from IGFBP-6 knockdown cells and analyzed for changes in the gene expressionprofile, using Affymetrix™ genome-wide mRNA profiling technology. The gene expressionpattern was further analyzed for relevant signaling pathways, using Ingenuity™ PathwaysAnalysis software and led to identification of NFkB pathway as potential functional targetfor IGFBP-6 (Fig. S2). Thus, we observed significant downregulation of NFkB-regulatedgenes coding for cytokines such as CCL2, CXCL1, CXCL5, and IL-8 in IGFBP-6 depletedcells (Fig. S2). These findings imply potential role for IGFBP-6 in the regulation of NFkBactivity.
3.5 Upregulation of IGFBP-6 levels in serum from elderly donorsTo link our observations made in cell culture to aging phenotypes, IGFBP-6 levels weredetermined in serum of healthy probands in different age groups. We found that IGFBP-6levels were significantly increased in serum from elderly donors (Fig. 6), suggesting thatIGFBP-6 upregulation may be relevant for human aging.
Micutkova et al. Page 9
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

4 DiscussionThe data obtained in this study suggest that IGFBP-6 is upregulated, when HDFs undergoreplicative senescence. This has been discovered by an unbiased proteomic analysis ofcellular supernatants and confirmed by Western blot and ELISA. When IGFBP-6 geneexpression was downregulated by shRNA-mediated gene silencing, cell proliferation wasinhibited and apoptotic cell death was increased. Furthermore, downregulation of IGFBP-6led to premature entry into cellular senescence. Since IGFBP-6 overexpression led to aslight but significant increase in cellular lifespan, the data suggest that IGFBP-6, in contrastto other IGF-binding proteins, is a negative regulator of cellular senescence in humanfibroblasts.
4.1 Role of the IGF/IGFBP axis in cellular senescenceSignaling through the insulin/IGF signal transduction pathway is known to contribute to themodulation of lifespan in all metazoan organisms, where this has been studied (Bartke,2005; Rincon et al., 2005). IGF-binding proteins are essential modulators of IGF signaling.They do so by regulating the availability of IGF for its receptors, and IGFBP-6 might play aparticular role because its affinity for IGF-II is approximately 50-fold higher than for IGF-I.Studies of cellular senescence have revealed an important contribution of IGF-bindingproteins to the regulation of cellular lifespan. Thus, it was shown that IGFBP-3 isupregulated in senescent fibroblasts, endothelial cells and other cell types (Goldstein et al.,1991; Moerman et al., 1993) and IGFBP-3 prevents mitogenic signaling throughextracellular IGF in senescent fibroblasts (Grigoriev et al., 1995) and thereby inducescellular senescence (Muck et al., 2008; Kim et al., 2007a). IGFBP-5 is also upregulated incellular models of replicative senescence (Hampel et al., 2006) and was shown to inducesenescence in HUVEC (Kim et al., 2007b). IGFBP-5 was also shown to contribute tofibrosis in vivo (Yasuoka et al., 2006). IGFBP-6 was found upregulated in stress-inducedcellular senescence (Xie et al., 2005), but its regulation in replicative senescence of humanfibroblasts has not been studied so far.
Among the members of the IGFBP gene family, IGFBP-3, -5 and -6 seem to shareadditional functions besides IGF binding, since these three proteins contain nuclearlocalization sequences. Accordingly, a nuclear function was suggested for IGFBP-3, whichseems to be independent of its capability to sequester IGF. Thus, it was shown that IGFBP-3can interact physically and functionally with retinoid X receptor (Liu et al., 2000; Schedlichet al., 2004; for recent review, see Yamada and Lee, 2009), leading to changes in geneexpression. Recent studies also suggested that IGFBP-3 can inhibit transcriptional activationby NFkB (Williams et al., 2007; Han et al., 2011), a family of structurally relatedtranscription factors that regulate immune and inflammatory responses, development, cellgrowth, and apoptosis. There is increasing evidence that NFkB belongs to the key regulatorsof the senescence-associated secretome, and Acosta et al. (2008) have shown NFkB-dependent activation of several cytokines in oncogene-induced senescence. Hence, NFkBmay play an active role in at least some forms of cellular senescence.
4.2 Role of IGFBP-6 in regulation of cell survival, proliferation, and senescenceConsistent with the here reported antiapoptotic activity of IGFBP-6 in human fibroblasts,several studies support a specific role for IGFBP-6 for cell survival in the nervous system, asIGF-II is the most abundantly expressed IGF in the adult CNS (Naeve et al., 2000). Robustupregulation of IGFBP-6 mRNA in lesioned motoneurons suggests that this binding proteinmay be of special relevance for severed neuronal cells (Hammarberg et al., 1998). Beilharzet al. (1998) have shown that IGFBP-6 was significantly upregulated in injured hemispherefollowing a severe hypoxic-ischemic (HI) injury. Since treatment of an HI brain injury with
Micutkova et al. Page 10
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

IGF-II was shown to worsen outcome and block the neuroprotective effects of IGF-I (Guanet al., 1996), it can be hypothesized that IGFBP-6 may potentiate the neuroprotective actionsof endogenous IGF-I (Beilharz et al., 1998) by a so far unknown mechanism.
Several previous studies showed that IGFBP-6 is accompanied mainly with antiproliferativephenotype due to its binding of IGF-II and blocking the IGF/insulin signaling (Bach, 2005),and IGFBP-6 was reported as tumor suppressor in nasopharyngeal carcinoma (Kuo et al.,2010). Moreover, IGFBP-6 gene expression is upregulated by agents which lead to thedevelopment of prematurely senescent phenotype, such as 5AZA (Kulaeva et al., 2003),doxorubicin (Chang et al., 2002; Xie et al., 2005) and H2O2 (Xie et al., 2005). However,some studies show upregulation of IGFBP-6 in situations accompanied by increased cellproliferation and survival. Thus, a Gli1 binding element was found in the promoter ofIGFBP-6 (Yoon et al., 2002) and recently, it was shown that, transcription factor Gli1maintained cell survival by binding the promoter regions and facilitating transcription ofBcl-2 genes along with IGFBP-6 in a parallel manner in pancreatic cancer cells (Xu et al.,2009). IGFBP-6 expression was also shown to be increased in invasive but not non-invasivemammary carcinoma cell lines (Evtimova et al., 2003). Finally, direct mitogenic andantiapoptotic effects of IGFBP-6 were described in the human osteoblastic osteosarcomacell line Saos-2/B-10 (Schmid et al., 1999). The studies reported here clearly suggest thatIGFBP-6, unlike other members of the IGFBP family, delays cellular senescence in diploidhuman fibroblasts, and this activity seems to involve regulation of the cdk inhibitorp21Waf1/Cip1. A significant upregulation of p21Waf1/Cip1 protein levels was observed inIGFBP-6 depleted cells, which contrasts with IGFBP-6 overexpressing cells, where theexpression of p21Waf1/Cip1 was strongly downregulated. It has been shown, thatoverexpression of p21Waf1/Cip1 is able to induce a senescence-like growth arrest in somecells (McConnell et al., 1998; Chang et al., 2000), while deletion of p21Waf1/Cip1 canpostpone senescent arrest (Medcalf et al., 1996; Brown et al., 1997). It may be interesting tonote that in both cases the levels of p21Waf1/Cip1 mRNA were not regulated (Micutkova etal., unpublished results), indicating that regulation of p21Waf1/Cip1 gene expression byIGFBP-6 occurs at the posttranscriptional level, consistent with several previouspublications reporting posttranscriptional regulation of p21Waf1/Cip1 under variousconditions (Li et al., 1996; Gong et al., 2003).
Circumstantial evidence presented here may also support the NFkB pathway as target forIGFBP-6 action. Using Affymetrix™ genome-wide mRNA profiling technology incombination with RT-qPCR (Laschober et al., 2010), we found a significant downregulationof NFkB-driven genes coding for cytokines such as CCL2, CXCL1, CXCL5, and IL-8 inIGFBP-6 depleted cells (see Fig. S2), suggesting that IGFBP-6 can influence NFkB activity;however, more work will be required to substantiate these findings.
We think that the increased production of IGFBP-6 by senescent cells may reflect anadaptive mechanism by which the cells avoid/delay senescence-associated growth arrest.Along these lines, we found that IGFBP-6 expression level is increased in serum fromelderly donors, suggesting that IGFBP-6 upregulation may be relevant for human aging.
ReferencesAcosta J.C. O’Loghlen A. Banito A. Guijarro M.V. Augert A. Raguz S. Fumagalli M. Da Costa M.
Brown C. Popov N. Takatsu Y. Melamed J. d’Adda di Fagagna F. Bernard D. Hernando E. Gil J.Chemokine signaling via the CXCR2 receptor reinforces senescence. Cell. 2008; 133:1006–1018.[PubMed: 18555777]
Aebersold R. Mann M. Mass spectrometry-based proteomics. Nature. 2003; 422:198–207. [PubMed:12634793]
Micutkova et al. Page 11
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

Bach L.A. Insulin-like growth factor binding protein-6: the “forgotten” binding protein? Horm. Metab.Res.. 1999; 31:226–234. [PubMed: 10226806]
Bach L.A. IGFBP-6 five years on; not so ‘forgotten’? Growth Horm. IGF Res.. 2005; 15:185–192.[PubMed: 15914054]
Bartke A. Minireview: role of the growth hormone/insulin-like growth factor system in mammalianaging. Endocrinology. 2005; 146:3718–3723. [PubMed: 15919742]
Beilharz E.J. Russo V.C. Butler G. Baker N.L. Connor B. Sirimanne E.S. Dragunow M. Werther G.A.Gluckman P.D. Williams C.E. Scheepens A. Co-ordinated and cellular specific induction of thecomponents of the IGF/IGFBP axis in the rat brain following hypoxic-ischemic injury. Brain Res.Mol. Brain Res.. 1998; 59:119–134. [PubMed: 9729323]
Brown J.P. Wei W. Sedivy J.M. Bypass of senescence after disruption of p21CIP1/WAF1 gene innormal diploid human fibroblasts. Science. 1997; 277:831–834. [PubMed: 9242615]
Callister S.J. Barry R.C. Adkins J.N. Johnson E.T. Qian W.J. Webb-Robertson B.J. Smith R.D. LiptonM.S. Normalization approaches for removing systematic biases associated with mass spectrometryand label-free proteomics. J. Proteome Res.. 2006; 5:277–286. [PubMed: 16457593]
Campisi J. The role of cellular senescence in skin aging. J. Investig. Dermatol. Symp. Proc.. 1998;3:1–5.
Campisi J. d’Adda di Fagagna F. Cellular senescence: when bad things happen to good cells. Nat. Rev.Mol. Cell Biol.. 2007; 8:729–740. [PubMed: 17667954]
Chambery D. de Galle B. Babajko S. Retinoic acid stimulates IGF binding protein (IGFBP)-6 anddepresses IGFBP-2 and IGFBP-4 in SK-N-SH human neuroblastoma cells. J. Endocrinol.. 1998;159:227–232. [PubMed: 9795362]
Chang B.D. Watanabe K. Broude E.V. Fang J. Poole J.C. Kalinichenko T.V. Roninson I.B. Effects ofp21Waf1/Cip1/Sdi1 on cellular gene expression: implications for carcinogenesis, senescence, andage-related diseases. Proc. Natl. Acad. Sci. U. S. A.. 2000; 97:4291–4296. [PubMed: 10760295]
Chang B.D. Swift M.E. Shen M. Fang J. Broude E.V. Roninson I.B. Molecular determinants ofterminal growth arrest induced in tumor cells by a chemotherapeutic agent. Proc. Natl. Acad. Sci.U. S. A.. 2002; 99:389–394. [PubMed: 11752408]
Chen J. Goligorsky M.S. Premature senescence of endothelial cells: Methusaleh's dilemma. Am. J.Physiol.: Heart Circ. Physiol.. 2006; 290:H1729–H1739. [PubMed: 16603702]
Coppe J.P. Patil C.K. Rodier F. Sun Y. Munoz D.P. Goldstein J. Nelson P.S. Desprez P.Y. Campisi J.Senescence-associated secretory phenotypes reveal cell-nonautonomous functions of oncogenicRAS and the p53 tumor suppressor. PLoS Biol.. 2008; 6:2853–2868. [PubMed: 19053174]
Dimri G.P. Lee X. Basile G. Acosta M. Scott G. Roskelley C. Medrano E.E. Linskens M. Rubelj I.Pereira-Smith O. Smith J. A biomarker that identifies senescent human cells in culture and inaging skin in vivo. Proc. Natl. Acad. Sci. U. S. A.. 1995; 92:9363–9367. [PubMed: 7568133]
Evtimova V. Zeillinger R. Weidle U.H. Identification of genes associated with the invasive status ofhuman mammary carcinoma cell lines by transcriptional profiling. Tumour Biol.. 2003; 24:189–198. [PubMed: 14654713]
Ewton D.Z. Florini J.R. IGF binding proteins-4, -5 and -6 may play specialized roles during L6myoblast proliferation and differentiation. J. Endocrinol.. 1995; 144:539–553. [PubMed: 7537792]
Ferber A. Chang C. Sell C. Ptasznik A. Cristofalo V.J. Hubbard K. Ozer H.L. Adamo M. Roberts C.T.Jr. LeRoith D. Dumenil G. Baserg R. Failure of senescent human fibroblasts to express the insulin-like growth factor-1 gene. J. Biol. Chem.. 1993; 268:17883–17888. [PubMed: 7688732]
Firth S.M. Baxter R.C. Cellular actions of the insulin-like growth factor binding proteins. Endocr.Rev.. 2002; 23:824–854. [PubMed: 12466191]
Fu P. Thompson J.A. Bach L.A. Promotion of cancer cell migration: an insulin-like growth factor(IGF)-independent action of IGF-binding protein-6. J. Biol. Chem.. 2007; 282:22298–22306.[PubMed: 17519236]
Fu P. Liang G.J. Khot S.S. Phan R. Bach L.A. Cross-talk between MAP kinase pathways is involvedin IGF-independent, IGFBP-6-induced Rh30 rhabdomyosarcoma cell migration. J. Cell. Physiol..2010; 224:636–643. [PubMed: 20432455]
Micutkova et al. Page 12
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

Gallicchio M.A. Kneen M. Hall C. Scott A.M. Bach L.A. Overexpression of insulin-like growth factorbinding protein-6 inhibits rhabdomyosarcoma growth in vivo. Int. J. Cancer. 2001; 94:645–651.[PubMed: 11745458]
Goldstein S. Moerman E.J. Jones R.A. Baxter R.C. Insulin-like growth factor binding protein 3accumulates to high levels in culture medium of senescent and quiescent human fibroblasts. Proc.Natl. Acad. Sci. U. S. A.. 1991; 88:9680–9684. [PubMed: 1719537]
Gong J. Ammanamanchi S. Ko T.C. Brattain M.G. Transforming growth factor beta 1 increases thestability of p21/WAF1/CIP1 protein and inhibits CDK2 kinase activity in human colon carcinomaFET cells. Cancer Res.. 2003; 63:3340–3346. [PubMed: 12810668]
Gramolini A.O. Kislinger T. Alikhani-Koopaei R. Fong V. Thompson N.J. Isserlin R. Sharma P. OuditG.Y. Trivieri M.G. Fagan A. Kannan A. Higgins D.G. Huedig H. Hess G. Arab S. Seidman J.G.Seidman C.E. Frey B. Perry M. Backx P.H. Liu P.P. MacLennan D.H. Emili A. Comparativeproteomics profiling of a phospholamban mutant mouse model of dilated cardiomyopathy revealsprogressive intracellular stress responses. Mol. Cell. Proteomics. 2008; 7:519–533. [PubMed:18056057]
Grellier P. De Galle B. Babajko S. Expression of insulin-like growth factor-binding protein 6complementary DNA alters neuroblastoma cell growth. Cancer Res.. 1998; 58:1670–1676.[PubMed: 9563481]
Grigoriev V.G. Moerman E.J. Goldstein S. Overexpression of insulin-like growth factor bindingprotein-3 by senescent human fibroblasts: attenuation of the mitogenic response to IGF-I. Exp.Cell Res.. 1995; 219:315–321. [PubMed: 7543848]
Guan J. Williams C.E. Skinner S.J. Mallard E.C. Gluckman P.D. The effects of insulin-like growthfactor (IGF)-1, IGF-2, and des-IGF-1 on neuronal loss after hypoxic-ischemic brain injury in adultrats: evidence for a role for IGF binding proteins. Endocrinology. 1996; 137:893–898. [PubMed:8603600]
Hammarberg H. Risling M. Hokfelt T. Cullheim S. Piehl F. Expression of insulin-like growth factorsand corresponding binding proteins (IGFBP 1–6) in rat spinal cord and peripheral nerve afteraxonal injuries. J. Comp. Neurol.. 1998; 400:57–72. [PubMed: 9762866]
Hampel B. Malisan F. Niederegger H. Testi R. Jansen-Durr P. Differential regulation of apoptotic celldeath in senescent human cells. Exp. Gerontol.. 2004; 39:1713–1721. [PubMed: 15582287]
Hampel B. Wagner M. Teis D. Zwerschke W. Huber L. Jansen-Durr P. Apoptosis resistance ofsenescent human fibroblasts is correlated with the absence of nuclear IGFBP-3. Aging Cell. 2005;4:325–330. [PubMed: 16300485]
Hampel B. Fortschegger K. Ressler S. Chang M.W. Unterluggauer H. Breitwieser A. SommergruberW. Fitzky B. Lepperdinger G. Jansen-Durr P. Voglauer R. Grillari J. Increased expression ofextracellular proteins as a hallmark of human endothelial cell in vitro senescence. Exp. Gerontol..2006; 41:474–481. [PubMed: 16626901]
Han J. Jogie-Brahim S. Harada A. Oh Y. Insulin-like growth factor-binding protein-3 suppressestumor growth via activation of caspase-dependent apoptosis and cross-talk with NF-κB signaling.Cancer Lett.. 2011 [Epub ahead of print].
Herbig U. Ferreira M. Condel L. Carey D. Sedivy J.M. Cellular senescence in aging primates. Science.2006; 311:1257. [PubMed: 16456035]
Iosef C. Gkourasas T. Jia C.Y. Li S.S. Han V.K. A functional nuclear localization signal in insulin-likegrowth factor binding protein-6 mediates its nuclear import. Endocrinology. 2008; 149:1214–1226. [PubMed: 18039785]
Jones J.I. Clemmons D.R. Insulin-like growth factors and their binding proteins: biological actions.Endocr. Rev.. 1995; 16:3–34. [PubMed: 7758431]
Kim E.J. Schaffer B.S. Kang Y.H. Macdonald R.G. Park J.H. Decreased production of insulin-likegrowth factor-binding protein (IGFBP)-6 by transfection of colon cancer cells with an antisenseIGFBP-6 cDNA construct leads to stimulation of cell proliferation. J. Gastroenterol. Hepatol..2002; 17:563–570. [PubMed: 12084030]
Kim K.S. Kim M.S. Seu Y.B. Chung H.Y. Kim J.H. Kim J.R. Regulation of replicative senescence byinsulin-like growth factor-binding protein 3 in human umbilical vein endothelial cells. Aging Cell.2007; 6:535–545. [PubMed: 17635417]
Micutkova et al. Page 13
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

Kim K.S. Seu Y.B. Baek S.H. Kim M.J. Kim K.J. Kim J.H. Kim J.R. Induction of cellular senescenceby insulin-like growth factor binding protein-5 through a p53-dependent mechanism. Mol. Biol.Cell. 2007; 18:4543–4552. [PubMed: 17804819]
Krizhanovsky V. Yon M. Dickins R.A. Hearn S. Simon J. Miething C. Yee H. Zender L. Lowe S.W.Senescence of activated stellate cells limits liver fibrosis. Cell. 2008; 134:657–667. [PubMed:18724938]
Kuilman T. Peeper D.S. Senescence-messaging secretome: SMS-ing cellular stress. Nat. Rev. Cancer.2009; 9:81–94. [PubMed: 19132009]
Kulaeva O.I. Draghici S. Tang L. Kraniak J.M. Land S.J. Tainsky M.A. Epigenetic silencing ofmultiple interferon pathway genes after cellular immortalization. Oncogene. 2003; 22:4118–4127.[PubMed: 12821946]
Kuo Y.S. Tang Y.B. Lu T.Y. Wu H.C. Lin C.T. IGFBP-6 plays a role as an oncosuppressor gene inNPC pathogenesis through regulating EGR-1 expression. J. Pathol.. 2010; 222:299–309. [PubMed:20635349]
Laschober G.T. Ruli D. Hofer E. Muck C. Carmona-Gutierrez D. Ring J. Hutter E. Ruckenstuhl C.Micutkova L. Brunauer R. Jamnig A. Trimmel D. Herndler-Brandstetter D. Brunner S. ZenzmaierC. Sampson N. Breitenbach M. Frohlich K.U. Grubeck-Loebenstein B. Berger P. Wieser M.Grillari-Voglauer R. Thallinger G.G. Grillari J. Trajanoski Z. Madeo F. Lepperdinger G. Jansen-Durr P. Identification of evolutionarily conserved genetic regulators of cellular aging. Aging Cell.2010; 9:1084–1097. [PubMed: 20883526]
Li X.S. Rishi A.K. Shao Z.M. Dawson M.I. Jong L. Shroot B. Reichert U. Ordonez J. Fontana J.A.Posttranscriptional regulation of p21WAF1/CIP1 expression in human breast carcinoma cells.Cancer Res.. 1996; 56:5055–5062. [PubMed: 8895764]
Lipinski R.J. Cook C.H. Barnett D.H. Gipp J.J. Peterson R.E. Bushman W. Sonic hedgehog signalingregulates the expression of insulin-like growth factor binding protein-6 during fetal prostatedevelopment. Dev. Dyn.. 2005; 233:829–836. [PubMed: 15906375]
Liu B. Lee H.Y. Weinzimer S.A. Powell D.R. Clifford J.L. Kurie J.M. Cohen P. Direct functionalinteractions between insulin-like growth factor-binding protein-3 and retinoid X receptor-alpharegulate transcriptional signaling and apoptosis. J. Biol. Chem.. 2000; 275:33607–33613.[PubMed: 10874028]
Maier J.A. Voulalas P. Roeder D. Maciag T. Extension of the life-span of human endothelial cells byan interleukin-1 alpha antisense oligomer. Science. 1990; 249:1570–1574. [PubMed: 2218499]
McConnell B.B. Starborg M. Brookes S. Peters G. Inhibitors of cyclin-dependent kinases inducefeatures of replicative senescence in early passage human diploid fibroblasts. Curr. Biol.. 1998;8:351–354. [PubMed: 9512419]
Medcalf A.S. Klein-Szanto A.J. Cristofalo V.J. Expression of p21 is not required for senescence ofhuman fibroblasts. Cancer Res.. 1996; 56:4582–4585. [PubMed: 8840965]
Moerman E.J. Thweatt R. Moerman A.M. Jones R.A. Goldstein S. Insulin-like growth factor bindingprotein-3 is overexpressed in senescent and quiescent human fibroblasts. Exp. Gerontol.. 1993;28:361–370. [PubMed: 7693501]
Muck C. Micutkova L. Zwerschke W. Jansen-Durr P. Role of insulin-like growth factor bindingprotein-3 in human umbilical vein endothelial cell senescence. Rejuvenation Res.. 2008; 11:449–453. [PubMed: 18171112]
Naeve G.S. Vana A.M. Eggold J.R. Verge G. Ling N. Foster A.C. Expression of rat insulin-likegrowth factor binding protein-6 in the brain, spinal cord, and sensory ganglia. Brain Res. Mol.Brain Res.. 2000; 75:185–197. [PubMed: 10686339]
Ressler S. Bartkova J. Niederegger H. Bartek J. Scharffetter-Kochanek K. Jansen-Durr P. WlaschekM. p16INK4A is a robust in vivo biomarker of cellular aging in human skin. Aging Cell. 2006;5:379–389. [PubMed: 16911562]
Rincon M. Rudin E. Barzilai N. The insulin/IGF-1 signaling in mammals and its relevance to humanlongevity. Exp. Gerontol.. 2005; 40:873–877. [PubMed: 16168602]
Roghani M. Hossenlopp P. Lepage P. Balland A. Binoux M. Isolation from human cerebrospinal fluidof a new insulin-like growth factor-binding protein with a selective affinity for IGF-II. FEBS Lett..1989; 255:253–258. [PubMed: 2551732]
Micutkova et al. Page 14
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

Roghani M. Segovia B. Whitechurch O. Binoux M. Purification from human cerebrospinal fluid ofinsulin-like growth factor binding proteins (IGFBPs). Isolation of IGFBP-2, an altered form ofIGFBP-3 and a new IGFBP species. Growth Regul.. 1991; 1:125–130. [PubMed: 1726837]
Schedlich L.J. Le Page S.L. Firth S.M. Briggs L.J. Jans D.A. Baxter R.C. Nuclear import of insulin-like growth factor-binding protein-3 and -5 is mediated by the importin beta subunit. J. Biol.Chem.. 2000; 275:23462–23470. [PubMed: 10811646]
Schedlich L.J. O’Han M.K. Leong G.M. Baxter R.C. Insulin-like growth factor binding protein-3prevents retinoid receptor heterodimerization: implications for retinoic acid-sensitivity in humanbreast cancer cells. Biochem. Biophys. Res. Commun.. 2004; 314:83–88. [PubMed: 14715249]
Schmid C. Keller C. Gosteli-Peter M. Zapf J. Mitogenic and antiapoptotic effects of insulin-likegrowth factor binding protein-6 in the human osteoblastic osteosarcoma cell line Saos-2/B-10.Biochem. Biophys. Res. Commun.. 1999; 263:786–789. [PubMed: 10512758]
Tang L.Y. Deng N. Wang L.S. Dai J. Wang Z.L. Jiang X.S. Li S.J. Li L. Sheng Q.H. Wu D.Q. Zeng R.Quantitative phosphoproteome profiling of Wnt3a-mediated signaling network: indicating theinvolvement of ribonucleoside-diphosphate reductase M2 subunit phosphorylation at residueserine 20 in canonical Wnt signal transduction. Mol. Cell. Proteomics. 2007; 6:1952–1967.[PubMed: 17693683]
Toussaint O. Royer V. Salmon M. Remacle J. Stress-induced premature senescence and tissue ageing.Biochem. Pharmacol.. 2002; 64:1007–1009. [PubMed: 12213599]
Vasile E. Tomita Y. Brown L.F. Kocher O. Dvorak H.F. Differential expression of thymosin beta-10by early passage and senescent vascular endothelium is modulated by VPF/VEGF: evidence forsenescent endothelial cells in vivo at sites of atherosclerosis. FASEB J.. 2001; 15:458–466.[PubMed: 11156961]
Wajapeyee N. Serra R.W. Zhu X. Mahalingam M. Green M.R. Oncogenic BRAF induces senescenceand apoptosis through pathways mediated by the secreted protein IGFBP7. Cell. 2008; 132:363–374. [PubMed: 18267069]
Wessel D. Flugge U.I. A method for the quantitative recovery of protein in dilute solution in thepresence of detergents and lipids. Anal. Biochem.. 1984; 138:141–143. [PubMed: 6731838]
Williams A.C. Smartt H. AM H.Z. Macfarlane M. Paraskeva C. Collard T.J. Insulin-like growth factorbinding protein 3 (IGFBP-3) potentiates TRAIL-induced apoptosis of human colorectal carcinomacells through inhibition of NF-kappaB. Cell Death Differ.. 2007; 14:137–145. [PubMed:16645643]
Xie L. Tsaprailis G. Chen Q.M. Proteomic identification of insulin-like growth factor-bindingprotein-6 induced by sublethal H2O2 stress from human diploid fibroblasts. Mol. Cell. Proteomics.2005; 4:1273–1283. [PubMed: 15958393]
Xu X.F. Guo C.Y. Liu J. Yang W.J. Xia Y.J. Xu L. Yu Y.C. Wang X.P. Gli1 maintains cell survivalby up-regulating IGFBP6 and Bcl-2 through promoter regions in parallel manner in pancreaticcancer cells. J. Carcinog.. 2009; 8:13. [PubMed: 19736394]
Yamada P.M. Lee K.W. Perspectives in Mammalian IGFBP-3 biology: local vs. systemic action. Am.J. Physiol.: Cell Physiol.. 2009; 296:C954–C976. [PubMed: 19279229]
Yasuoka H. Jukic D.M. Zhou Z. Choi A.M. Feghali-Bostwick C.A. Insulin-like growth factor bindingprotein 5 induces skin fibrosis: a novel murine model for dermal fibrosis. Arthritis Rheum.. 2006;54:3001–3010. [PubMed: 16947625]
Yoon J.W. Kita Y. Frank D.J. Majewski R.R. Konicek B.A. Nobrega M.A. Jacob H. Walterhouse D.Iannaccone P. Gene expression profiling leads to identification of GLI1-binding elements in targetgenes and a role for multiple downstream pathways in GLI1-induced cell transformation. J. Biol.Chem.. 2002; 277:5548–5555. [PubMed: 11719506]
Appendix A Supplementary dataRefer to Web version on PubMed Central for supplementary material.
Micutkova et al. Page 15
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

AcknowledgmentsWe acknowledge excellent technical support by Michael Neuhaus and Hans-Peter Viertler. The work wassupported by the European Union (Integrated Project PROTEOMAGE). The work in PJD's laboratory was alsosupported by a grant from the Austrian Science Funds (L342-B05).
Micutkova et al. Page 16
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

Fig. 1.Identification of IGFBP-6 protein in supernatants from human diploid fibroblasts. A) MS/MS spectrum of HLDSVLQQLQTEVYR peptide assigned to IGFBP-6 protein. B)Sequence of IGFBP-6 protein showing all 4 peptides identified in quantitative LC-MS/MSanalysis with total 31.25% sequence coverage and 115 spectral counts. Different peptidesare highlighted with different colors. (For interpretation of the references to color in thisfigure legend, the reader is referred to the web version of the article.)
Micutkova et al. Page 17
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

Fig. 2.Levels of endogenous IGFBP-6 protein in HDFs. A) Representative Western blot ofIGFBP-6 in cell lysates and supernatants of young (Y; 18% of lifespan completed) andsenescent (S; 93% of lifespan completed) fibroblasts is shown. As a loading control,SERPINE2 with unchanged abundance in young and senescent supernatants was used. B)Concentration of IGFBP-6 in supernatants was measured by IGFBP-6 specific ELISA intwo different HDF isolates (HDF1: p = 0.0011; HDF2: p = 0.0002; n = 3).
Micutkova et al. Page 18
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

Fig. 3.IGFBP-6 lentiviral knockdown in HDFs. A) Young HDFs (21% of lifespan completed) wereinfected with two different IGFBP-6 shRNAs (shRNA1 and shRNA2). IGFBP-6 knockdownlevels were assessed by real-time PCR at 32% of lifespan completed. Results from twoindependent infections are shown (I, II) (p < 0.001; n = 6). B) Immunoblots show theIGFBP-6 protein levels in cell lysates and supernatants of knockdown cells compared tocontrol shRNA (SCR). As a loading control, GAPDH and SERPINE2 were used. C) Growthcurve of knockdown HDFs is represented as number of population doublings (PDL) afterviral transduction.
Micutkova et al. Page 19
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent
Micutkova et al. Page 20
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

Fig. 4.Functional consequences of IGFBP-6 knockdown. A) Cell proliferation of infected HDFswas assessed by a BrdU incorporation assay and subsequent FACS analysis. Representativefigures show BrdU positive cells (green) with DAPI counterstained nuclei (blue) along withcorresponding FACS profiles. Summary BrdU graph represents the mean value ± SEM(shRNA1: p = 0.000004; shRNA2: p = 0.0005; n = 12). B) Cell death of HDFs withIGFBP-6 knockdown was determined by Annexin V-FACS. Representative FACS profilesare shown, along with summary graph (shRNA1: p = 0.00002; shRNA2: p = 0.0005; n = 9).C) The apoptosis frequency of cells treated as in panel B was independently assessed bypropidium iodide staining (shRNA1: p = 0.05; shRNA2: p = 0.04; n = 4). D) Changes in thesenescence status were monitored by senescence associated β-galactosidase staining (SA-β-gal: shRNA1: 0.000009; shRNA2: 0.000005; n = 8). Representative Western blot shows thelevels of p21Waf1/Cip1 protein. As a control α-tubulin (α-Tub) was used. All measurements
Micutkova et al. Page 21
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

were performed at 32-42% of lifespan completed. (For interpretation of the references tocolor in this figure legend, the reader is referred to the web version of the article.)
Micutkova et al. Page 22
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

Micutkova et al. Page 23
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

Fig. 5.IGFBP-6 lentiviral overexpression in HDFs. A) Young HDFs (21% of lifespan completed)were infected with IGFBP-6 overexpressing lentiviruses. Levels of IGFBP-6 mRNA werequantified by real-time PCR at 32% of lifespan completed. As a control, cells were infectedeither with lentiviruses containing empty vector (Mock) and/or overexpressing greenfluorescent protein (GFP). Results from two independent infections are shown (IGFBP6_1:p = 0.00004; IGFBP6_2: p = 0.0007; n = 6). B) Immunoblots show the IGFBP-6 proteinlevels in cell lysates and supernatants of HDFs with IGFBP-6 overexpression compared toMock and GFP control cells. As a loading control, GAPDH and SERPINE2 were used. C)Growth curve of HDF with overexpression of IGFBP-6, Mock and/or GFP is represented as
Micutkova et al. Page 24
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

a number of population doublings (PDL) upon viral transduction. Data are shown induplicates. D) Cell proliferation was estimated by BrdU incorporation assay. Representativefigures show BrdU positive cells (green) with DAPI counterstained nuclei (blue) (p = 0.02;n = 8). E) Cell apoptosis was measured by Annexin V-FACS (p = 0.002; n = 4). F) Thefrequency of apoptosis of cells treated as in panel E was independently assessed by PI-FACS (p = 0.00045; n = 4). G) Changes in the percentage of senescent cells were monitoredby SA-β-gal staining (p = 0.00005; n = 8). Representative Western blot shows the levels ofp21Waf1/Cip1 protein. As a control α-tubulin (α-Tub) was used. All functional assays wereperformed at 90–95% of lifespan completed. (For interpretation of the references to color inthis figure legend, the reader is referred to the web version of the article.)
Micutkova et al. Page 25
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent

Fig. 6.IGFBP-6 levels in human serum. IGFBP-6 protein levels were quantified by ELISA inserum samples derived from healthy volunteers in the old age group (age: 66–92, n = 93)and young/middle-age group (age: 25–41, n = 47; p = 0.000009).
Micutkova et al. Page 26
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent
Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent
Micutkova et al. Page 27
Table 1
Differentially expressed secreted proteins. Quantitative proteomic analysis using LC-MS/MS combined withlabel free spectral counting approach identified 26 extracellular proteins that are differentially expressed byyoung (Y1–Y3; 18% of lifespan completed) and senescent (S1–S3; 93% of lifespan completed) fibroblasts (t-test, p < 0.05, fold change >1.5, n = 3). IPI – international protein index; s.c. – spectral counts.
IPI Name Y1s.c. Y2s.c. Y3s.c. S1s.c. S2s.c. S3s.c. S/Y ratio t-testp value
IPI00296713 GRN Isoform 1 of Granulins precursor 0 1 0 2 3 5 12.489 0.0264
IPI00295741 CTSB Cathepsin B precursor 4 3 6 11 17 27 5.061 0.0324
IPI00008561 MMP1 Interstitial collagenase precursor 6 8 15 29 41 42 4.693 0.0043
IPI00011229 CTSD Cathepsin D precursor 2 5 3 9 6 8 2.784 0.0075
IPI00029235 IGFBP6 Insulin-like growth factor-binding protein 6 precursor
9 10 19 27 26 24 2.449 0.0049
IPI00296099 THBS1 Thrombospondin-1 precursor 49 45 48 61 102 115 2.345 0.0344
IPI00465439 ALDOA Fructose-bisphosphate aldolaseA
1 1 2 3 2 2 2.104 0.0379
IPI00844360 COL4A1 Collagen alpha-1(IV) chainprecursor
3 6 3 7 7 5 1.921 0.0452
IPI00302679 LTBP1 latent transforming growth factorbeta binding protein 1 isoform LTBP-1S
3 2 2 4 3 4 1.852 0.0193
IPI00013508 ACTN1 Alpha-actinin-1 14 15 15 9 8 7 0.654 0.0025
IPI00296537 FBLN1 Isoform C of Fibulin-1 precursor 79 67 83 48 27 30 0.545 0.0225
IPI00029658 EFEMP1 Isoform 1 of EGF-containingfibulin-like extracellular matrix protein 1precursor
6 11 10 2 4 3 0.407 0.0226
IPI00218725 LAMA2 laminin alpha 2 subunit isoformb precursor
19 17 17 10 5 2 0.382 0.0226
IPI00029568 PTX3 Pentraxin-related protein PTX3precursor
35 23 46 12 10 11 0.378 0.0320
IPI00296165 C1R Complement C1r subcomponentprecursor
115 81 111 26 26 29 0.314 0.0066
IPI00329573 COL12A1 Isoform 1 of Collagenalpha-1(XII) chain precursor
28 34 29 10 8 4 0.291 0.0010
IPI00792115 CLEC3B Putative uncharacterizedprotein DKFZp686H17246
5 10 10 1 2 1 0.197 0.0112
IPI00012119 DCN Isoform A of Decorin precursor 17 30 41 2 5 4 0.153 0.0176
IPI00783987 C3 Complement C3 precursor(Fragment)
6 10 10 0 2 0 0.098 0.0045
IPI00013179 PTGDS Prostaglandin-H2 D-isomeraseprecursor
5 4 4 1 0 0 0.089 0.0035
IPI00000860 FMOD Fibromodulin precursor 2 2 1 0 0 0 Only in Y 0.0097
IPI00003865 HSPA8 Isoform 1 of Heat shock cognate71 kDa protein
2 2 1 0 0 0 Only in Y 0.0097
IPI00005292 SPOCK1 Testican-1 precursor 2 2 4 0 0 0 Only in Y 0.0128
IPI00165972 CFD Complement factor Dpreproprotein
8 7 5 0 0 0 Only in Y 0.0035
IPI00444378 AEBP1 CDNA FLJ45634 fis, cloneCHONS2002829, moderately similar toadipocyte enhancer binding protein 1
2 3 2 0 0 0 Only in Y 0.0012
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.

Sponsored Docum
ent Sponsored D
ocument
Sponsored Docum
ent
Micutkova et al. Page 28
IPI Name Y1s.c. Y2s.c. Y3s.c. S1s.c. S2s.c. S3s.c. S/Y ratio t-testp value
IPI00643034 PLTP Isoform 1 of Phospholipid transferprotein precursor
1 1 2 0 0 0 Only in Y 0.0128
Published as: Mech Ageing Dev. 2011 October ; 132(10): 468–479.




















