
Insights from event-related potentials into the temporal and hierarchical organization of the...
16
Insights from event-related potentials into the temporal and hierarchical organization of the ventral and dorsal streams of the visual system in selective attention MANUEL MARTÍN-LOECHES, JOSÉ A. HINOJOSA, and FRANCISCO J. RUBIA Brain Mapping Unit, Pluridisciplinary Institute, Complutense University, Madrid, Spain Abstract The temporal and hierarchical relationships between the dorsal and the ventral streams in selective attention are known only in relation to the use of spatial location as the attentional cue mediated by the dorsal stream. To improve this state of affairs, event-related brain potentials were recorded while subjects attended simultaneously to motion direction ~mediated by the dorsal stream! and to a property mediated by the ventral stream ~color or shape!. At about the same time, a selection positivity ~SP! started for attention mediated by both streams. However, the SP for color and shape peaked about 60 ms later than motion SP. Subsequently, a selection negativity ~SN! followed by a late positive component ~ LPC! were found simultaneously for attention mediated by both streams. A hierarchical relationship between the two streams was not observed, but neither SN nor LPC for one property was completely insensitive to the values of the other property. Descriptors: Selective attention, Motion, Color, Shape, Visual system, Event-related potentials The event-related potential ~ ERP! is a useful tool in psychophys- iological research. ERP studies have made notable contributions to our knowledge about both the cognitive and the neural levels of selective attention. Specifically, ERPs can be particularly useful in determining the timing and hierarchical organization of perceptual processes ~for recent reviews, see Heslenfeld, Kenemans, Kok, & Molenaar, 1997; Hillyard et al., 1995!. In this regard, ERPs have a large advantage over other brain imaging techniques such as positron emission tomography ~ PET! or functional magnetic res- onance imaging ~fMRI !. Moreover, ERPs alone or in combination with other techniques have also shed some light on the anatomical localization of attentional processes ~e.g., Clark & Hillyard, 1996; Gomez-Gonzalez, Clark, Fan, Luck, & Hillyard, 1994; Heinze et al., 1994!. Regarding the temporal organization of selective attention, ERPs studies have usually determined that a very early attention effect is produced by spatial location of the stimuli ~ Mangun, 1995!, usu- ally followed in time by attention to nonspatial properties such as color or shape ~e.g., Hillyard & Münte, 1984; Mangun & Hillyard, 1995; Paz-Caballero & García Austt, 1992!. Spatial attention mod- ulates the early P1 and N1 components of the ERPs, whereas attention to nonspatial features is usually reflected by later mod- ulations, such as a selection positivity ~SP! and a selection nega- tivity ~SN!. However, it has also been found recently that attentional modulations produced by nonspatial features such as spatial fre- quency, color, stimulus size, or orientation may appear as early as the attention effects produced by spatial location ~ Karayanidis & Michie, 1997; Mangun & Hillyard, 1995; Zani & Proverbio, 1995, 1997!. Nonspatial properties of the stimuli, on the other hand, also seem to show attention effects at different timings. For instance, early selection based on shape has been described to be more effective than selection based on color, the reverse being true in later stages ~Smid, Jakob, & Heinze, 1997!. Closely related to timing is the question of the hierarchical organization of selective attention. In this regard, the terms hier- archical ~or contingent ! and independent processing have been applied. A contingent or hierarchical relationship between stimulus properties in selective attention would take place when the value of one stimulus property influences the effects of attention to other stimulus properties. This relationship has been observed in several studies: size being contingent upon color ~ Wijers, Mulder, Okita, & Mulder, 1989!, orientation upon spatial frequency ~ Kenemans, Kok, & Smulders, 1993!, and color and motion upon spatial loca- tion ~Anllo-Vento & Hillyard, 1996!. Usually, contingency or hi- erarchy have been synonymous with cancellation, that is, attention effects for one property may be completely absent under certain values of the other property. On the other hand, processing of stimulus properties may also be independent, that is, produced by several independent feature-specific pathways that work in an ex- haustive fashion ~ Hansen & Hillyard, 1983!. Of course, indepen- dent and contingent processing may coexist, as several degrees of This study was supported in part by grant CICYT-PM96-0052096. The authors thank Berenice Valdés for her help in data collection. We also thank LourdesAnllo-Vento, Werner Sommer, Cyma Van Petten, and five anonymous reviewers for highly valuable comments and help on a preliminary version of the manuscript. Address reprint requests to: M. Martín-Loeches, Brain Mapping Unit, Pluridisciplinary Institute, Complutense University, Po. Juan XXIII, 1, 28040- Madrid, Spain. E-mail: [email protected]. Psychophysiology, 36 ~1999!, 721–736. Cambridge University Press. Printed in the USA. Copyright © 1999 Society for Psychophysiological Research 721
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Insights from event-related potentials into the temporal and hierarchical organization of the...

Insights from event-related potentials into the temporaland hierarchical organization of the ventral and dorsalstreams of the visual system in selective attention
MANUEL MARTÍN-LOECHES, JOSÉ A. HINOJOSA,and FRANCISCO J. RUBIABrain Mapping Unit, Pluridisciplinary Institute, Complutense University, Madrid, Spain
Abstract
The temporal and hierarchical relationships between the dorsal and the ventral streams in selective attention are knownonly in relation to the use of spatial location as the attentional cue mediated by the dorsal stream. To improve this stateof affairs, event-related brain potentials were recorded while subjects attended simultaneously to motion direction~mediated by the dorsal stream! and to a property mediated by the ventral stream~color or shape!. At about the sametime, a selection positivity~SP! started for attention mediated by both streams. However, the SP for color and shapepeaked about 60 ms later than motion SP. Subsequently, a selection negativity~SN! followed by a late positivecomponent~LPC! were found simultaneously for attention mediated by both streams. A hierarchical relationshipbetween the two streams was not observed, but neither SN nor LPC for one property was completely insensitive to thevalues of the other property.
Descriptors: Selective attention, Motion, Color, Shape, Visual system, Event-related potentials
The event-related potential~ERP! is a useful tool in psychophys-iological research. ERP studies have made notable contributions toour knowledge about both the cognitive and the neural levels ofselective attention. Specifically, ERPs can be particularly useful indetermining the timing and hierarchical organization of perceptualprocesses~for recent reviews, see Heslenfeld, Kenemans, Kok, &Molenaar, 1997; Hillyard et al., 1995!. In this regard, ERPs havea large advantage over other brain imaging techniques such aspositron emission tomography~PET! or functional magnetic res-onance imaging~fMRI !. Moreover, ERPs alone or in combinationwith other techniques have also shed some light on the anatomicallocalization of attentional processes~e.g., Clark & Hillyard, 1996;Gomez-Gonzalez, Clark, Fan, Luck, & Hillyard, 1994; Heinzeet al., 1994!.
Regarding the temporal organization of selective attention, ERPsstudies have usually determined that a very early attention effect isproduced by spatial location of the stimuli~Mangun, 1995!, usu-ally followed in time by attention to nonspatial properties such ascolor or shape~e.g., Hillyard & Münte, 1984; Mangun & Hillyard,1995; Paz-Caballero & García Austt, 1992!. Spatial attention mod-ulates the early P1 and N1 components of the ERPs, whereas
attention to nonspatial features is usually reflected by later mod-ulations, such as a selection positivity~SP! and a selection nega-tivity ~SN!. However, it has also been found recently that attentionalmodulations produced by nonspatial features such as spatial fre-quency, color, stimulus size, or orientation may appear as early asthe attention effects produced by spatial location~Karayanidis &Michie, 1997; Mangun & Hillyard, 1995; Zani & Proverbio, 1995,1997!. Nonspatial properties of the stimuli, on the other hand, alsoseem to show attention effects at different timings. For instance,early selection based on shape has been described to be moreeffective than selection based on color, the reverse being true inlater stages~Smid, Jakob, & Heinze, 1997!.
Closely related to timing is the question of the hierarchicalorganization of selective attention. In this regard, the termshier-archical ~or contingent! and independent processing have beenapplied. A contingent or hierarchical relationship between stimulusproperties in selective attention would take place when the value ofone stimulus property influences the effects of attention to otherstimulus properties. This relationship has been observed in severalstudies: size being contingent upon color~Wijers, Mulder, Okita,& Mulder, 1989!, orientation upon spatial frequency~Kenemans,Kok, & Smulders, 1993!, and color and motion upon spatial loca-tion ~Anllo-Vento & Hillyard, 1996!. Usually, contingency or hi-erarchy have been synonymous with cancellation, that is, attentioneffects for one property may be completely absent under certainvalues of the other property. On the other hand, processing ofstimulus properties may also be independent, that is, produced byseveral independent feature-specific pathways that work in an ex-haustive fashion~Hansen & Hillyard, 1983!. Of course, indepen-dent and contingent processing may coexist, as several degrees of
This study was supported in part by grant CICYT-PM96-0052096.The authors thank Berenice Valdés for her help in data collection. We
also thank Lourdes Anllo-Vento, Werner Sommer, Cyma Van Petten, andfive anonymous reviewers for highly valuable comments and help on apreliminary version of the manuscript.
Address reprint requests to: M. Martín-Loeches, Brain Mapping Unit,Pluridisciplinary Institute, Complutense University, Po. Juan XXIII, 1, 28040-Madrid, Spain. E-mail: [email protected].
Psychophysiology, 36~1999!, 721–736. Cambridge University Press. Printed in the USA.Copyright © 1999 Society for Psychophysiological Research
721

independence and contingent relationship appear to be plausibleover time~Smid et al., 1997!.
As mentioned above, the anatomy of attentional processes canalso be studied by means of ERPs. Nevertheless, anatomical con-clusions drawn from ERPs studies are weaker than those based onother techniques such as PET or fMRI. However, although a pre-cise anatomical location based on ERPs may not be free of trade-offs, it is at least possible to infer the involvement of differentgenerating brain systems from different ERPs topographies~Mc-Carthy & Wood, 1985; Rugg & Coles, 1995!.
With regard to visual selective attention, there appear to be twomain systems of interest. We can dissociate a “ventral stream” ofoccipitotemporal visual areas for processing stimulus propertiessuch as color, or shape, and an occipitoparietal “dorsal stream” forspatial and movement information. This division was originallyfound and well documented in the macaque monkey, but recentstudies also support its presence in the human brain~McIntoshet al., 1994; Tootell, Dale, Sereno, & Malach, 1996; Ungerleider &Haxby, 1994!. The ventral stream has also been called the “tem-poral” or “what” pathway. The chief reason for calling it the “what”stream is its function in identifying objects, which is to a largeextent based on the main features processed by this pathway, thatis, color and form. On the other hand, the dorsal stream has alsobeen called the “parietal” or “where” pathway. This “where” path-way is not merely devoted to identifying object positions, butconstitutes a more general system involved in visual motion pro-cessing, eye movements, and spatial organization.
“What versus where” is the most common division of the visualsystem, although further subdivisions can be made, as there seemto be dissociable brain subsystems for processing particular prop-erties within the ventral stream such as color and shape~Corbetta,Miezin, Dobmeyer, Shulman, & Petersen, 1990!. Recently, it hasalso been reported that there are particular situations in which onestream may also process properties of the stimuli theoreticallycorresponding to the domain of the other stream. For instance,although dorsal areas such as the middle temporal~MT ! specializein motion information~Kaneoke, Bundoe, Koyama, Suzuki, &Kakigi, 1997; O’Craven, Rosen, Kwong, Treisman, & Savoy, 1997!,several types of motion processing may rely on ventral areas~Maun-sell & Ferrera, 1994!. In a like manner, shape identification maydepend on the dorsal stream under certain circumstances~Britten,Newsome, & Saunders, 1992!. However, the division of the visualsystem into a dorsal and a ventral stream, the dorsal stream spe-cialized in the processing of spatial and motion information andthe ventral stream in the processing of object features such as coloror shape, is currently the predominantly accepted model~e.g.,Ungerleider, Courtney, & Haxby, 1998!.
The literature suggests that the dorsal stream plays an earlierattention role than the ventral stream. This is supported by theabove-mentioned findings that a very early attention effect is pro-duced by spatial location of the stimuli~processed by the dorsalstream!, usually being followed in time by effects of attention toproperties processed by the ventral stream~Harter & Aine, 1984;Mangun, 1995; Mangun & Hillyard, 1995!. But recent evidenceindicates that early effects may not be exclusive to spatial location~Karayanidis & Michie, 1997; Mangun & Hillyard, 1995; Zani &Proverbio, 1995, 1997!. However, with the exception of the studyby Zani and Proverbio~1997!, in all these studies attention to thenonspatial feature was never combined with simultaneous atten-tion to spatial location, as has been the usual case when spatialattention effects precede nonspatial effects. On the other hand, thetemporal prevalence of attention to spatial location is also accom-
panied by a contingent relationship: effects of attention to nonspa-tial features are greatly reduced or absent when the location of thestimuli is not the attended one~e.g., Mangun, 1995; Mangun &Hillyard, 1995!.
One might thus conclude that the “where” or spatial-motiondetection system is dominant in the visual information processingsystem. This dominance might tentatively be explained by adap-tive mechanisms. Giving priority to the subsystem devoted to lo-calizing stimuli and their motion might allow more adaptive benefitsthan the subsystem identifying objects, as the former could initiatepreparation to escape when necessary. This interpretation would bein consonance with the close link between the system for voluntaryorienting of visuospatial attention and the neural system for re-sponse preparation~Yamaguchi & Kobayashi, 1998!.
However, these assertions regarding the relationships betweenthe dorsal and ventral streams might not extrapolate to all situa-tions. Apart from very few exceptions~Hillyard & Münte, 1984;Zani & Proverbio, 1997!, they can be perfectly applicable only inrelation to spatial location as a property analyzed by the dorsalstream. However, it is a fact that no other property of the dorsalstream has been studied in combination with properties of theventral stream.
With the exception of two recent studies~Anllo-Vento & Hill-yard, 1996; Girelli & Luck, 1997!, nonspatial properties of thedorsal stream have not been used as attentional cues in ERP stud-ies. Anllo-Vento and Hillyard combined direction of motion withspatial location, both properties of the dorsal system. This studythus addressed contingent relationships and timing of processespertaining to the dorsal stream itself, rather than relationships be-tween the dorsal and ventral systems. The results showed a stronghierarchical relationship between the two properties, motion atten-tion effects being completely absent at unattended locations. Atattended locations, selective attention to motion showed ERP ef-fects consisting mainly of a frontal SP and a posterior SN, whereaslocation selection showed earlier effects, the usual P10N1 enhance-ment. Similar results were obtained for selective attention to colorin the same study. The motion-related SN, nevertheless, was foundto have a different topography from color-related SN, as wouldbe expected if two different underlying streams processed theseproperties.
In the other study using motion as an attentional cue, Girelliand Luck~1997! required subjects to detect a single, salient fea-ture. Hence, and again, conclusions regarding the contingent rela-tionships between the dorsal and ventral streams when other thanspatial location is used as attentional cue of the dorsal subsystemcannot be stated. Also, Girelli and Luck focused their efforts on aparticular time segment~containing the N2pc component, thoughtto be functionally similar to SN!, thus disregarding the timing ofprocesses of selective attention to motion.
Therefore, it seems that further research is required to elucidatethe temporal and hierarchical relationships between the dorsal andventral streams in visual selective attention. Particularly, it is nec-essary to use a dorsal stream feature other than spatial location, incombination with attentional cues processed by the ventral stream.With this aim, the present study examines two unique combina-tions of to-be-attended stimulus properties: motion-plus-color andmotion-plus-shape. Within each combination the dorsal stream an-alyzes one of the relevant properties~motion!, while the ventralstream analyzes the other~color, shape!. We can thus further ex-plore the timing and hierarchical relationships between the twostreams when properties other than spatial location are used as theattentional cue for the dorsal stream. If it is true that the dorsal or
722 M. Martín-Loeches, J.A. Hinojosa, and F.J. Rubia

“where” stream prevails within the visual information processingsystem, the ERP modulations due to motion attention effects wouldtake place earlier than those pertaining to attention effects medi-ated by the ventral or “what” stream, that is, color or shape atten-tion effects. In addition to or instead of this temporal prevalence,a hierarchical prevalence of the dorsal stream might be observedfor if color or shape attention effects are contingent upon motionattention effects, that is, if color or shape attention effects are slightor absent for unattended values of motion.
Methods
SubjectsTwenty-four subjects~12 women; age range5 18–26 years,M522 years! participated in the study. Subjects had normal or corrected-to-normal vision, and reported no neurological complaints. Allsubjects were right handed, with average handedness scores~Old-field, 1971! of 1 .82, with a range of1 .38 to1 .100. Subjectswere paid for participating.
StimuliEach stimulus consisted of a pair of sequentially flashed squares orcircles. Squares were 0.93 0.98 in size and circles were 1.18 indiameter~equivalent area for both squares and circles!. The secondflash of the pair was centered 0.58 left or right of the first, therebyproducing the perception of a horizontally moving square or circle,which could be either red or blue. Screen background was alwayswhite. Accordingly, there were four types of stimulus pairs, de-fined by orthogonal combinations of color and direction of motion—red leftward, red rightward, blue leftward, blue rightward.
The center of the first flash could appear randomly located atany point ~1 pixel resolution! within a 0.83 2.68 ~vertical andhorizontal dimensions, respectively! rectangle located 0.88 abovethe central fixation point~Figure 1!. The limits of this rectangle
were never visible to subjects, though they were advised of thisprocedure. Because squares and circles differed slightly in size~toobtain equivalent areas!, this procedure yielded different total ar-eas into which either squares or circles could appear. Thus, thetotal entire area for squares was 1.253 3.058, and for circles1.353 3.158. The stimuli appeared randomly within these areas,and could cross the vertical midline. In this way, movement di-rection was designed to be largely independent of spatial location.The total stimulus field was small in order to avoid eye movementsthat could contaminate recordings. The total area of stimulationcan be considered a “central” space, given its small surface.
Each flash in the pair had a duration of 33 ms, and the stimulusonset asynchrony~SOA! could be of 50 ms~standard trials,p 5.85! or 150 ms~target trials,p 5 .15!.
There were 1,000 stimuli~pairs of flashes! per block. Eachstimulus was sampled randomly from 16 stimulus classes definedby direction of motion~leftward0rightward, p 5 .5 each!, color~red0blue, p 5 .5 each!, shape~square0circle, p 5 .5 each!, andtype of stimulus~standard0target!. Interstimulus intervals~ISIs!varied at random between 250 and 750 ms~rectangular distribution!.
ProcedureSubjects were seated in a comfortable chair and were trained tomaintain fixation on the central cross while discriminating theinfrequent targets from the common standards. Each subject wastested in two different blocks but in a single session. In one block,motion direction and color of the stimuli were task relevant~motion-color block!. Here, subjects were told to attend simultaneously toa specific motion direction~leftward or rightward! and a specificcolor ~red or blue!. In the other block the relevant properties weremotion direction and shape~motion-shape block!, in which sub-jects had to attend to a specific motion direction~leftward orrightward! and a specific shape~square or circle!. The order ofpresentation of the type of block, the attended motion direction, theattended color in the motion-color block, and the attended shape inthe motion-shape block were all counterbalanced across subjects.Within each session, the attended motion direction in the secondblock was the unattended one in the first block.
Subjects were instructed to press a button every time they de-tected a target, that is, a pair of flashes separated by a SOA of150 ms, and having the attended values of the stimulus properties.Given the probabilities outlined above, the number of targets withthe attended values of the stimulus properties was about 62–63 per1,000 stimuli in a block. The number of standards with the at-tended values of the stimulus properties was about 212–213 per1,000 stimuli in a block. Each block included a rest period in themiddle. Response hand was switched midway through each blockafter the rest period; initial response hand was counterbalancedacross subjects. Before the recordings, subjects were trained bypracticing the task for 2–3 min.
Electrophysiological RecordingElectroencephalographic data were amplified with SYN-AMPSamplifiers ~NeuroScan, Inc., Sterling, VA!. The electrode siteswere Fp1, Fp2, F7, F3, Fz, F4, F8, T7, C3, Cz, C4, T8, P7, P5, P3,P1, Pz, P2, P4, P6, P8, PO7, PO1, PO2, PO8, O1, Oz, and O2 ofthe revised 10020 International System~American Electroenceph-alographic Society, 1991!. The electrooculogram~EOG! was ob-tained from below versus above the right eye~vertical EOG! andthe left versus right lateral orbital rim~horizontal EOG!. Electrodeimpedances were kept below 5 kV. Amplification bandpass was0.01–50 Hz, and sampling rate was 250 Hz. All electrodes were
Figure 1. Each stimulus consisted of a pair of squares or circles flashedbriefly separated by a short interval~50 or 150 ms! to produce the per-ception of moving stimuli. Stimuli always moved along the horizontalmeridian, in a leftward or rightward direction. Also, stimuli could be eitherred or blue. Here, a rightward-moving circle is shown. The center of eachfirst flash could appear located at any point within the dotted-line rectangle~not visible to subjects!.
Ventral and dorsal streams in selective attention 723

referred to linked mastoids, and a ground electrode was located onthe forehead~Fpz position!.
Offline averaging of the ERPs was performed by extractingoverlapping epochs of the EEG beginning 200 ms before the firststimulus of each pair~triggering stimulus! and continuing for800 ms poststimulus. Artifact rejection was performed before av-eraging. Trials in which EOG activity exceeded 40mV were au-tomatically rejected. On average, 18% of the trials were rejectedby this method, there being no notable differences across condi-tions in this respect~a range of 15–22% of rejected trials!. Offlinecorrection of smaller eye movement artifacts was also made, usingthe method described by Semlitsch, Anderer, Schuster, and Preelich~1986!. Trials with incorrect responses were excluded from analysis.
Due to the short ISIs used in this study, the ERPs responses tosuccessive stimuli overlapped. However, the averaged ERPs to thetemporally adjacent stimuli were “smeared” out in the averagesbecause of the ISI randomization~Hillyard & Münte, 1984!. None-theless, even in the case that IS randomization was not fully ef-fective, the overlap should not differ across conditions.
Data AnalysisBehavioral performance was measured in terms of reaction times~RTs! and percentage of correct responses. Responses were scoredas correct if they occurred within 200–1,000 ms of targets with theattended values of motion and color~or shape!. All other responseswere classified as false alarms~FAs!.
A separate average was computed for each of the 16 types ofstimulus defined by the direction of motion~attended0unattended!,color~attended0unattended!, shape~attended0unattended!, and typeof stimulus~standard0target!, and for each electrode location. OnlyERPs responses elicited in behaviorally correct trials were in-cluded in the averages. All amplitudes were calculated with respectto the mean voltage over the 200 ms preceding stimulus onset.
Mean voltages of the ERPs to standard stimuli were obtainedfor 24 ms time windows starting at 48 ms after stimulus onset andcontinuing until 360 ms. From 360 to 760 ms, mean voltages werecalculated for 100-ms windows. These fine-grained analyses werenecessary to achieve our principal aim of analyzing the temporalrelationships between streams. However, given that P1 peak la-tency could be precisely located midway between two time win-dows, and given the special relevance of this component for selectiveattention studies, it was decided to include an additional timewindow in the analyses to specifically explore this component.Based on the latency of P1 peak amplitude, this window comprisedthe time period from 108 to 132 ms.
The mean amplitudes of each time window were submitted toanalysis of variance~ANOVA !, using the Greenhouse–Geisser ep-silon correction for nonsphericity of the variance-covariance ma-trix ~Jennings & Wood, 1976!. A four-way repeated-measuresANOVA was applied to each time window of ERPs to standards,with the following factors: attended ventral stream feature~colorvs. shape, or C vs. S!; attended versus unattended value of dorsalstream feature, that is, motion direction~M1 vs. M2!; attendedversus unattended value of ventral stream feature~C1 vs. C2, orS1 vs. S2!; and electrode~28 levels!. For the sake of brevity theywill be referred to as ventral feature, value of dorsal feature, valueof ventral feature, and electrode, respectively. As midline elec-trodes~Fz, Cz, Pz, and Oz! were recorded and analyzed, it wasconsidered more appropriate not to include hemisphere as an ad-ditional factor in the ANOVA. However, when a detailed analy-sis of waves or maps revealed a possible asymmetry of theeffects, a direct comparison between left and right hemispheres
was performed by means of ANOVA using only those leads wherethe effects appeared larger. Finally, when more than two consec-utive windows are found to be significant in relation to the sameeffect, the smallest and largestF, p, andE values in that range arereported.
ERPs to target stimuli were analyzed following similar proce-dures, but considering other time windows based on their maindifferences with respect to standard stimuli, as will be more fullydescribed in the Results section corresponding to targets. Seem-ingly, however, target and standard stimuli should not be comparedas they differ physically~there were different SOAs between thefirst and the second flashes!. Nevertheless, we decided to makethis comparison, in the same way as Anllo-Vento and Hillyard~1996!, finding some results of interest.
Results
Behavioral DataRTs to targets when color was combined with motion~638 ms!were shorter than when shape was the combined feature~656 ms!,but this difference only approached significance,F~1,23! 5 4.2,p , .1. The average percentage of correct target detections wasrelatively low, 56.5%. When color was combined with motion, thepercentage of correct responses was slightly but significantlyhigher than when shape was the combined feature~59% vs. 54%!,F~1,23! 5 5.1, p , .05. These low values of correct percentageswere striking, as a preliminary study with eight subjects yielded anaverage of 70% correct responses. The low values in the presentstudy may well be due to a relative difficulty for discriminatingbetween targets and standards, reported by most subjects. It ap-pears that the difference in SOA between targets and standards wasrelatively difficult to distinguish in our study, and this problemmay be because the center of the stimuli varied randomly from trialto trial within a stimulation area that covered both the vertical andhorizontal dimensions. Actually, this was one of the major differ-ences from Anllo-Vento and Hillyard’s~1996! study, in which higherlevels of correct responses were obtained. Another difference fromthat study was the use in the present study of rather unusually longstimulus series~of about 10 min!, which may have led to problemsin sustaining attention at the end of the runs. However, this secondpossibility can be discarded, as an inspection of the data did notreveal any particular accumulation of errors over the series.
In any case, this high level of difficulty may be advantageousfor our purposes, as it would have motivated subjects to maintainhigher levels of attention. The percentage of FAs was very low:1.5% on average, indicating that the subjects were conservative intheir decision criteria. No significant difference in percentage ofFAs was observed between blocks combining color with motion~1.7%! and blocks combining shape with motion~1.4%!.
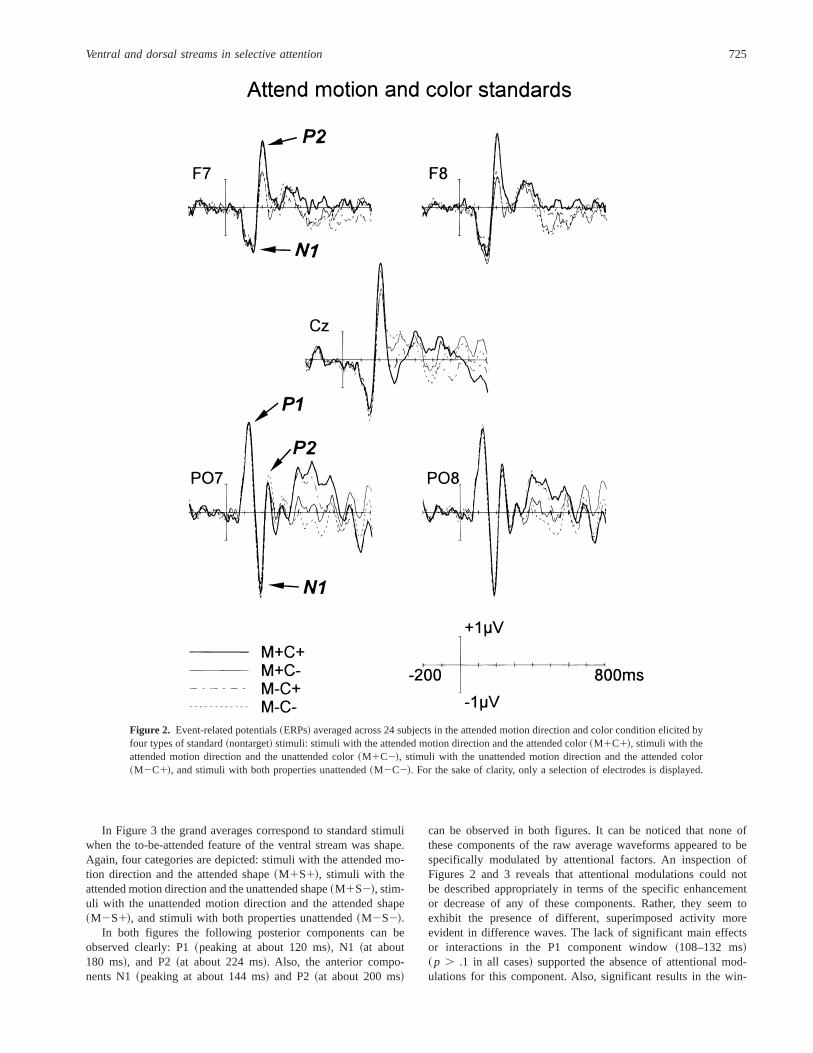
ERP DataERPs to Standard StimuliFigure 2 shows the grand-average ERP waveforms to standard
stimuli when color was the to-be-attended feature of the ventralstream. For the sake of clarity, only a selection of electrodes isdisplayed. ERPs to standards were averaged separately in fourcategories: stimuli with the attended motion direction and the at-tended color~M1C1!, stimuli with the attended motion directionand the unattended color~M1C2!, stimuli with the unattendedmotion direction and the attended color~M2C1!, and stimuli withboth properties unattended~M2C2!.
724 M. Martín-Loeches, J.A. Hinojosa, and F.J. Rubia
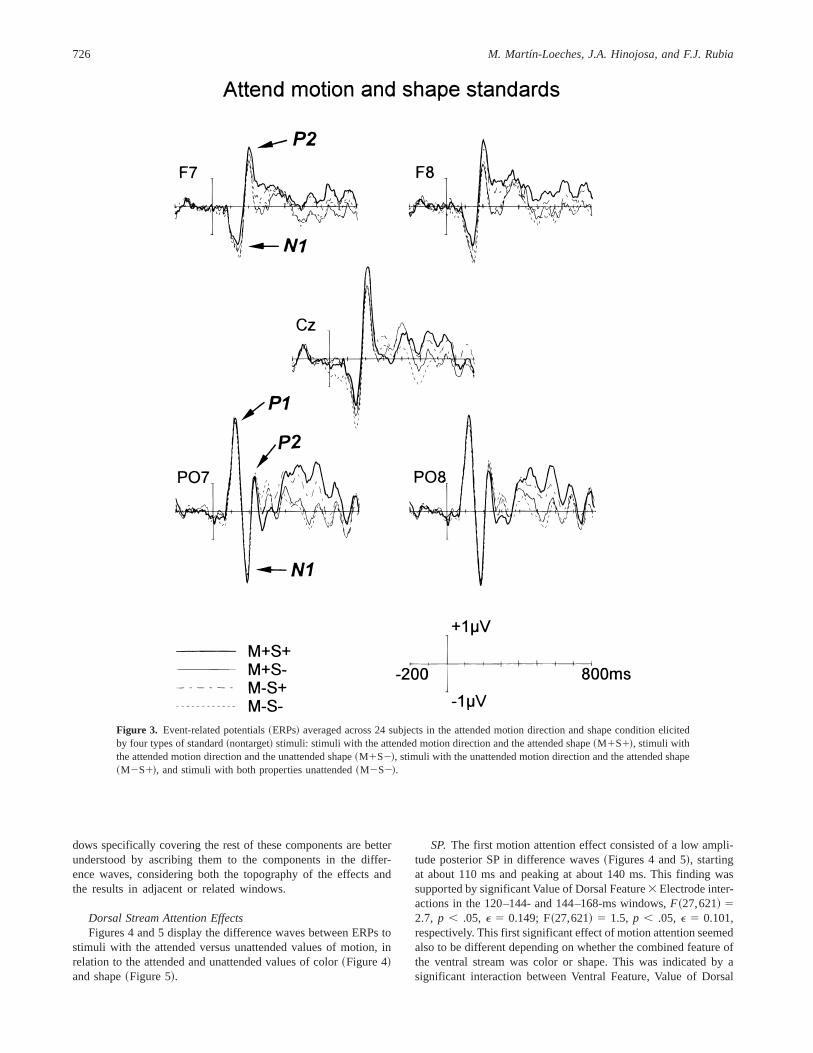
In Figure 3 the grand averages correspond to standard stimuliwhen the to-be-attended feature of the ventral stream was shape.Again, four categories are depicted: stimuli with the attended mo-tion direction and the attended shape~M1S1!, stimuli with theattended motion direction and the unattended shape~M1S2!, stim-uli with the unattended motion direction and the attended shape~M2S1!, and stimuli with both properties unattended~M2S2!.
In both figures the following posterior components can beobserved clearly: P1~peaking at about 120 ms!, N1 ~at about180 ms!, and P2~at about 224 ms!. Also, the anterior compo-nents N1~peaking at about 144 ms! and P2~at about 200 ms!
can be observed in both figures. It can be noticed that none ofthese components of the raw average waveforms appeared to bespecifically modulated by attentional factors. An inspection ofFigures 2 and 3 reveals that attentional modulations could notbe described appropriately in terms of the specific enhancementor decrease of any of these components. Rather, they seem toexhibit the presence of different, superimposed activity moreevident in difference waves. The lack of significant main effectsor interactions in the P1 component window~108–132 ms!~ p . .1 in all cases! supported the absence of attentional mod-ulations for this component. Also, significant results in the win-
Figure 2. Event-related potentials~ERPs! averaged across 24 subjects in the attended motion direction and color condition elicited byfour types of standard~nontarget! stimuli: stimuli with the attended motion direction and the attended color~M1C1!, stimuli with theattended motion direction and the unattended color~M1C2!, stimuli with the unattended motion direction and the attended color~M2C1!, and stimuli with both properties unattended~M2C2!. For the sake of clarity, only a selection of electrodes is displayed.
Ventral and dorsal streams in selective attention 725
dows specifically covering the rest of these components are betterunderstood by ascribing them to the components in the differ-ence waves, considering both the topography of the effects andthe results in adjacent or related windows.
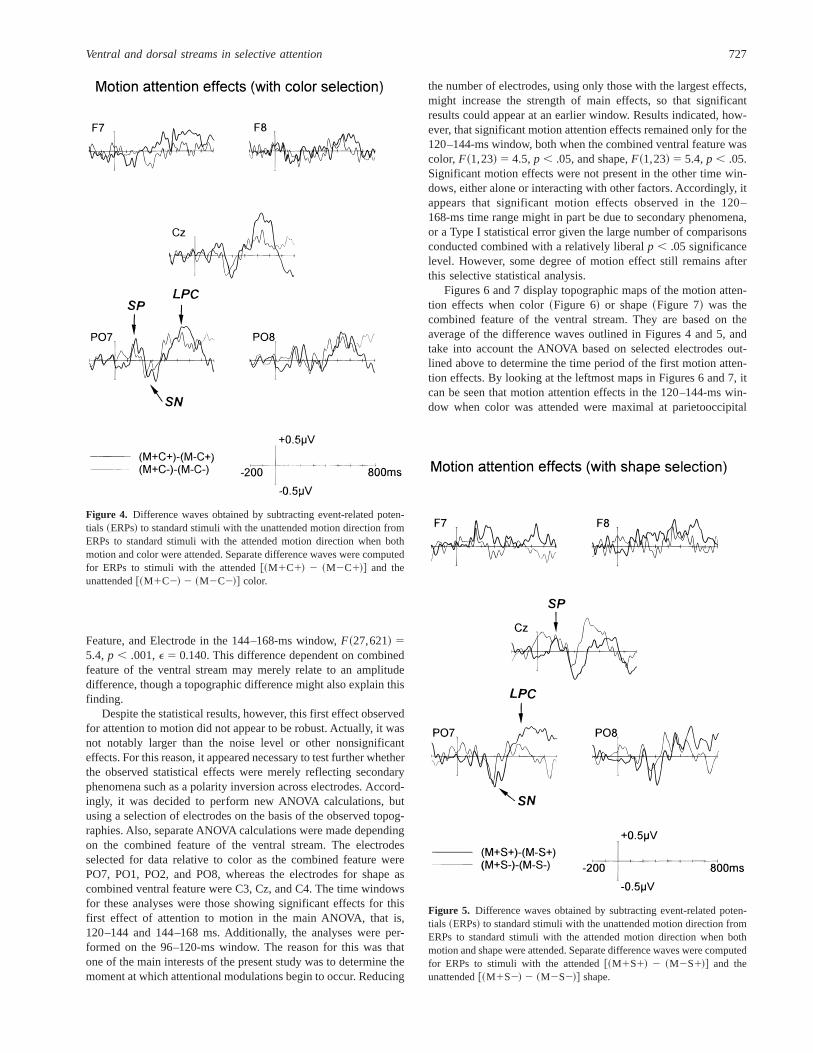
Dorsal Stream Attention EffectsFigures 4 and 5 display the difference waves between ERPs to
stimuli with the attended versus unattended values of motion, inrelation to the attended and unattended values of color~Figure 4!and shape~Figure 5!.
SP. The first motion attention effect consisted of a low ampli-tude posterior SP in difference waves~Figures 4 and 5!, startingat about 110 ms and peaking at about 140 ms. This finding wassupported by significant Value of Dorsal Feature3 Electrode inter-actions in the 120–144- and 144–168-ms windows,F~27,621! 52.7, p , .05, E 5 0.149; F~27,621! 5 1.5, p , .05, E 5 0.101,respectively. This first significant effect of motion attention seemedalso to be different depending on whether the combined feature ofthe ventral stream was color or shape. This was indicated by asignificant interaction between Ventral Feature, Value of Dorsal
Figure 3. Event-related potentials~ERPs! averaged across 24 subjects in the attended motion direction and shape condition elicitedby four types of standard~nontarget! stimuli: stimuli with the attended motion direction and the attended shape~M1S1!, stimuli withthe attended motion direction and the unattended shape~M1S2!, stimuli with the unattended motion direction and the attended shape~M2S1!, and stimuli with both properties unattended~M2S2!.
726 M. Martín-Loeches, J.A. Hinojosa, and F.J. Rubia
Feature, and Electrode in the 144–168-ms window,F~27,621! 55.4,p , .001,E5 0.140. This difference dependent on combinedfeature of the ventral stream may merely relate to an amplitudedifference, though a topographic difference might also explain thisfinding.
Despite the statistical results, however, this first effect observedfor attention to motion did not appear to be robust. Actually, it wasnot notably larger than the noise level or other nonsignificanteffects. For this reason, it appeared necessary to test further whetherthe observed statistical effects were merely reflecting secondaryphenomena such as a polarity inversion across electrodes. Accord-ingly, it was decided to perform new ANOVA calculations, butusing a selection of electrodes on the basis of the observed topog-raphies. Also, separate ANOVA calculations were made dependingon the combined feature of the ventral stream. The electrodesselected for data relative to color as the combined feature werePO7, PO1, PO2, and PO8, whereas the electrodes for shape ascombined ventral feature were C3, Cz, and C4. The time windowsfor these analyses were those showing significant effects for thisfirst effect of attention to motion in the main ANOVA, that is,120–144 and 144–168 ms. Additionally, the analyses were per-formed on the 96–120-ms window. The reason for this was thatone of the main interests of the present study was to determine themoment at which attentional modulations begin to occur. Reducing
the number of electrodes, using only those with the largest effects,might increase the strength of main effects, so that significantresults could appear at an earlier window. Results indicated, how-ever, that significant motion attention effects remained only for the120–144-ms window, both when the combined ventral feature wascolor,F~1,23! 5 4.5,p , .05, and shape,F~1,23! 5 5.4,p , .05.Significant motion effects were not present in the other time win-dows, either alone or interacting with other factors. Accordingly, itappears that significant motion effects observed in the 120–168-ms time range might in part be due to secondary phenomena,or a Type I statistical error given the large number of comparisonsconducted combined with a relatively liberalp , .05 significancelevel. However, some degree of motion effect still remains afterthis selective statistical analysis.
Figures 6 and 7 display topographic maps of the motion atten-tion effects when color~Figure 6! or shape~Figure 7! was thecombined feature of the ventral stream. They are based on theaverage of the difference waves outlined in Figures 4 and 5, andtake into account the ANOVA based on selected electrodes out-lined above to determine the time period of the first motion atten-tion effects. By looking at the leftmost maps in Figures 6 and 7, itcan be seen that motion attention effects in the 120–144-ms win-dow when color was attended were maximal at parietooccipital
Figure 4. Difference waves obtained by subtracting event-related poten-tials ~ERPs! to standard stimuli with the unattended motion direction fromERPs to standard stimuli with the attended motion direction when bothmotion and color were attended. Separate difference waves were computedfor ERPs to stimuli with the attended@~M1C1! 2 ~M2C1!# and theunattended@~M1C2! 2 ~M2C2!# color.
Figure 5. Difference waves obtained by subtracting event-related poten-tials ~ERPs! to standard stimuli with the unattended motion direction fromERPs to standard stimuli with the attended motion direction when bothmotion and shape were attended. Separate difference waves were computedfor ERPs to stimuli with the attended@~M1S1! 2 ~M2S1!# and theunattended@~M1S2! 2 ~M2S2!# shape.
Ventral and dorsal streams in selective attention 727
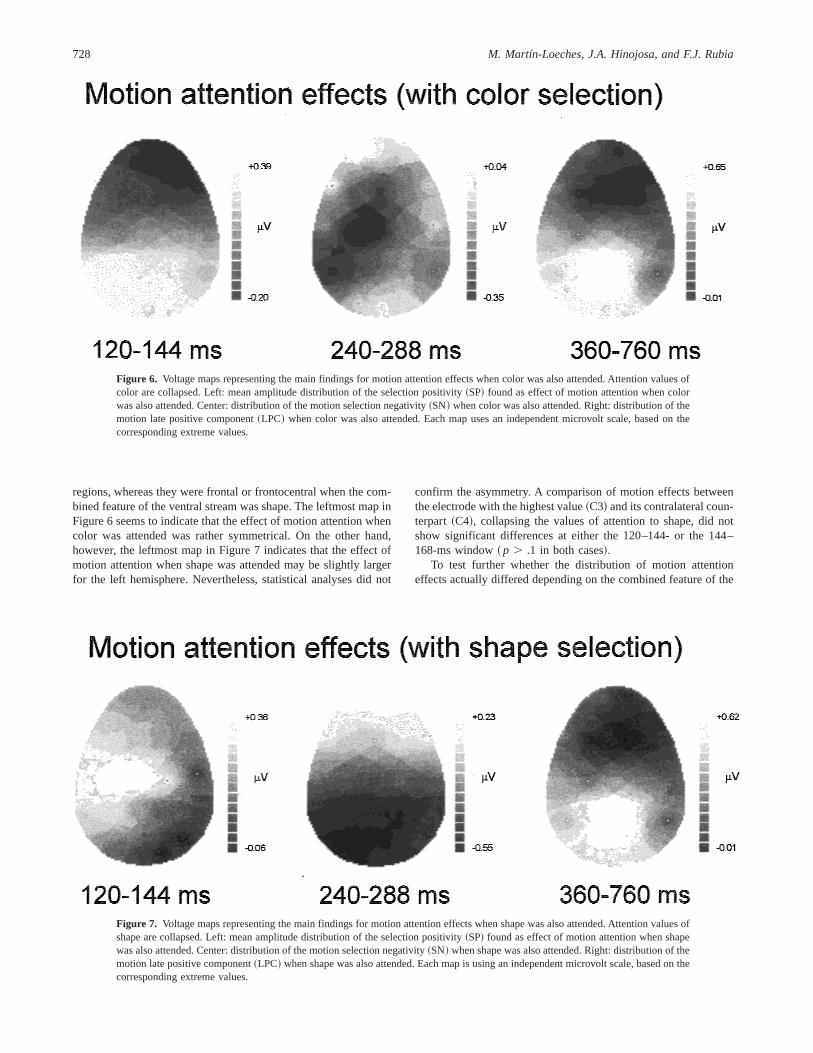
regions, whereas they were frontal or frontocentral when the com-bined feature of the ventral stream was shape. The leftmost map inFigure 6 seems to indicate that the effect of motion attention whencolor was attended was rather symmetrical. On the other hand,however, the leftmost map in Figure 7 indicates that the effect ofmotion attention when shape was attended may be slightly largerfor the left hemisphere. Nevertheless, statistical analyses did not
confirm the asymmetry. A comparison of motion effects betweenthe electrode with the highest value~C3! and its contralateral coun-terpart ~C4!, collapsing the values of attention to shape, did notshow significant differences at either the 120–144- or the 144–168-ms window~ p . .1 in both cases!.
To test further whether the distribution of motion attentioneffects actually differed depending on the combined feature of the
Figure 6. Voltage maps representing the main findings for motion attention effects when color was also attended. Attention values ofcolor are collapsed. Left: mean amplitude distribution of the selection positivity~SP! found as effect of motion attention when colorwas also attended. Center: distribution of the motion selection negativity~SN! when color was also attended. Right: distribution of themotion late positive component~LPC! when color was also attended. Each map uses an independent microvolt scale, based on thecorresponding extreme values.
Figure 7. Voltage maps representing the main findings for motion attention effects when shape was also attended. Attention values ofshape are collapsed. Left: mean amplitude distribution of the selection positivity~SP! found as effect of motion attention when shapewas also attended. Center: distribution of the motion selection negativity~SN! when shape was also attended. Right: distribution of themotion late positive component~LPC! when shape was also attended. Each map is using an independent microvolt scale, based on thecorresponding extreme values.
728 M. Martín-Loeches, J.A. Hinojosa, and F.J. Rubia
ventral stream, a profile analysis was performed~McCarthy &Wood, 1985!. Although the interaction between ventral feature,value of dorsal feature, and electrode was not significant in the120–144-ms window, it appeared appropriate to carry out a profileanalysis, given the evidence for different topographies from theleftmost maps in Figures 6 and 7. For this purpose, differencewaves for motion attention effects when color was attended weremade, collapsing the values of attention to color. The same pro-cedure was followed to obtain difference waves for motion atten-tion effects when shape was attended. Amplitude differences wereeliminated by scaling. For the time window of interest~120–144 ms! in the difference waves, mean amplitudes were scaled foreach subject across all electrodes, with the average distance fromthe mean, calculated from the grand-mean ERPs, as denominator.Significant differences in ANOVAs with these scaled data, wherepossible effects of source strength are eliminated, provide unambig-uous evidence for different scalp distributions. However, this to-pography comparison was not significant,F~27,621! 5 2.1, p ,.1, E 5 0.140.
SN. The second phase of the motion attention effect was a SNpeaking at about 280 ms. A significant value of dorsal featureeffect was obtained in the 240–312-ms range windows,F~1,23! 58.5–11.8,p , .01 in all cases, and significant Value of DorsalFeature3 Electrode interactions in the 240–264- and 264–288-mswindows,F~27,621! 5 4.5,p , .01,E5 0.146;F~27,621! 5 2.7,p , .05,E5 0.137, respectively. Figures 6 and 7 show that this SNseems to present a slightly different topography depending on thecombined feature of the ventral stream. Motion SN when color wasalso attended showed the highest values at parietal and centralelectrodes, bilaterally, but tending to be predominantly left. Sim-ilarly, motion SN when shape was also attended showed thehighest values at parietal electrodes, bilaterally, but also largerover the left. Neither asymmetry was statistically significant incomparisons of bilateral pairs of electrodes showing the largesteffects ~ p . .1 both in the 240–264- and 264–288-ms win-dows!. Neither were there any significant interaction betweenventral feature and value of dorsal feature, nor between ventralfeature, value of dorsal feature, and electrode in the time win-dows corresponding to the SN~ p . .1 in all cases!. Thus, it canbe asserted that motion SN exhibits the same distribution regard-less of the combined feature of the ventral stream. Moreover,this finding was confirmed by profile analyses following theprocedures described above, which failed to show significantdifferences in either the 240–264- or 264–288-ms windows~ p . .1!. The motion SN can be summarized as showing aparietal central distribution. Sequential maps during the 240–288-ms interval~not shown! also revealed the stability over timeof this topographic distribution of motion SN.
Late positive component.The final significant finding for mo-tion attention effects corresponds to a late positive component~LPC! that covers the time range 360–760 ms. Within this timeperiod, significant value of dorsal feature effects were found in the460–560- and 560–660-ms windows,F~1,23! 5 16.6,p , .001;F~1,23! 5 7.6, p , .05, respectively. Also, significant Value ofDorsal Feature3 Electrode effects were found in the 360–760-msrange windows,F~27,621! 5 2.7–4.8;p , .05–.01;E 5 0.128–0.202. Rightmost maps in Figures 6 and 7 display these effects.This component was largest parietocentrally, both when the com-bined feature of the ventral stream was color and when it wasshape.
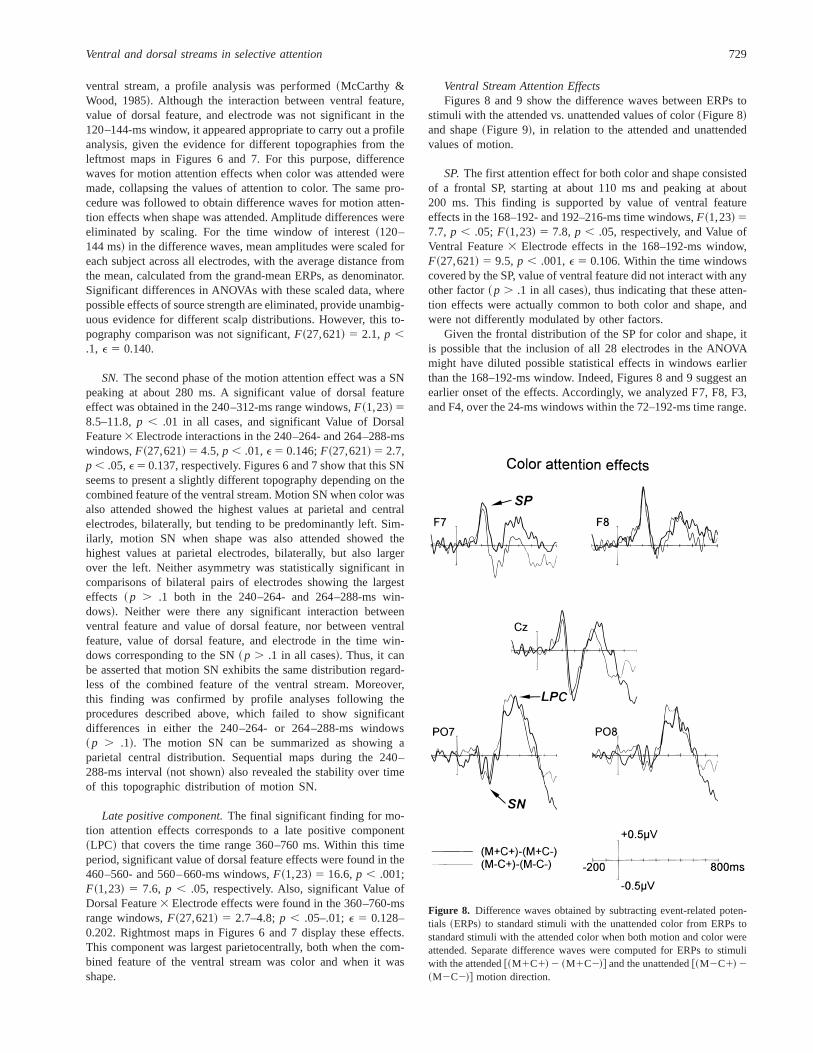
Ventral Stream Attention EffectsFigures 8 and 9 show the difference waves between ERPs to
stimuli with the attended vs. unattended values of color~Figure 8!and shape~Figure 9!, in relation to the attended and unattendedvalues of motion.
SP.The first attention effect for both color and shape consistedof a frontal SP, starting at about 110 ms and peaking at about200 ms. This finding is supported by value of ventral featureeffects in the 168–192- and 192–216-ms time windows,F~1,23! 57.7, p , .05; F~1,23! 5 7.8, p , .05, respectively, and Value ofVentral Feature3 Electrode effects in the 168–192-ms window,F~27,621! 5 9.5, p , .001,E 5 0.106. Within the time windowscovered by the SP, value of ventral feature did not interact with anyother factor~ p . .1 in all cases!, thus indicating that these atten-tion effects were actually common to both color and shape, andwere not differently modulated by other factors.
Given the frontal distribution of the SP for color and shape, itis possible that the inclusion of all 28 electrodes in the ANOVAmight have diluted possible statistical effects in windows earlierthan the 168–192-ms window. Indeed, Figures 8 and 9 suggest anearlier onset of the effects. Accordingly, we analyzed F7, F8, F3,and F4, over the 24-ms windows within the 72–192-ms time range.
Figure 8. Difference waves obtained by subtracting event-related poten-tials ~ERPs! to standard stimuli with the unattended color from ERPs tostandard stimuli with the attended color when both motion and color wereattended. Separate difference waves were computed for ERPs to stimuliwith the attended@~M1C1! 2 ~M1C2!# and the unattended@~M2C1! 2~M2C2!# motion direction.
Ventral and dorsal streams in selective attention 729
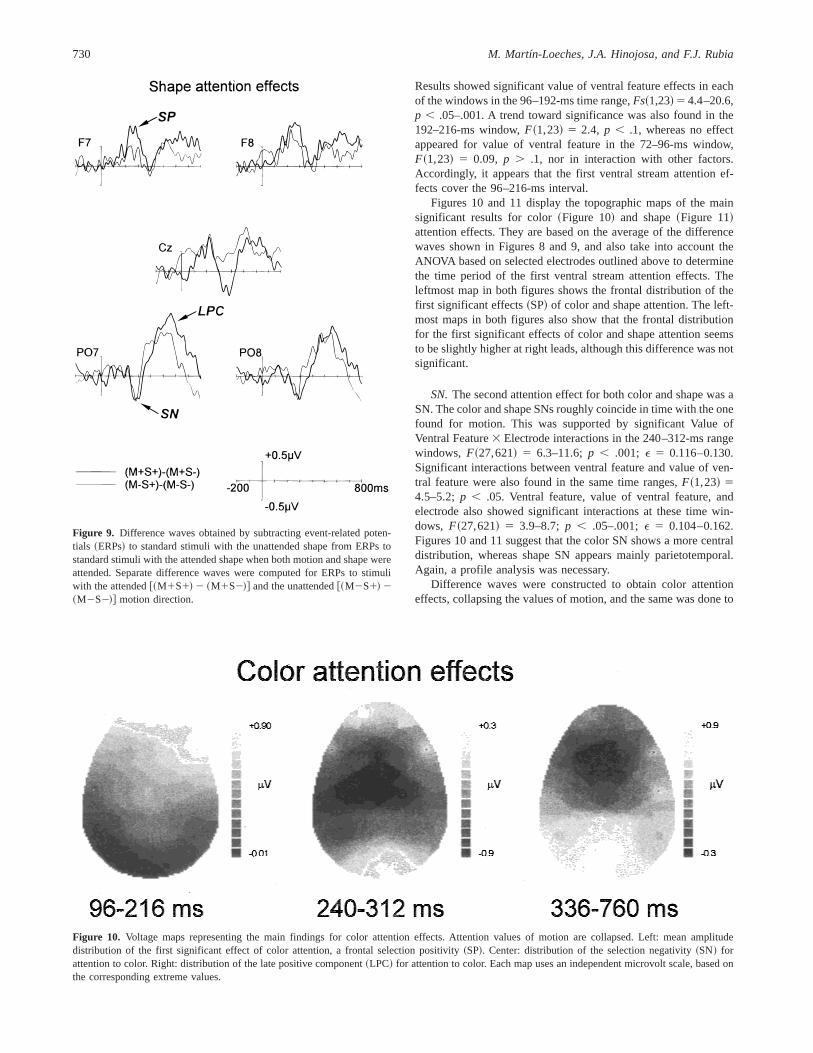
Results showed significant value of ventral feature effects in eachof the windows in the 96–192-ms time range,Fs~1,23! 5 4.4–20.6,p , .05–.001. A trend toward significance was also found in the192–216-ms window,F~1,23! 5 2.4, p , .1, whereas no effectappeared for value of ventral feature in the 72–96-ms window,F~1,23! 5 0.09, p . .1, nor in interaction with other factors.Accordingly, it appears that the first ventral stream attention ef-fects cover the 96–216-ms interval.
Figures 10 and 11 display the topographic maps of the mainsignificant results for color~Figure 10! and shape~Figure 11!attention effects. They are based on the average of the differencewaves shown in Figures 8 and 9, and also take into account theANOVA based on selected electrodes outlined above to determinethe time period of the first ventral stream attention effects. Theleftmost map in both figures shows the frontal distribution of thefirst significant effects~SP! of color and shape attention. The left-most maps in both figures also show that the frontal distributionfor the first significant effects of color and shape attention seemsto be slightly higher at right leads, although this difference was notsignificant.
SN.The second attention effect for both color and shape was aSN. The color and shape SNs roughly coincide in time with the onefound for motion. This was supported by significant Value ofVentral Feature3 Electrode interactions in the 240–312-ms rangewindows, F~27,621! 5 6.3–11.6;p , .001; E 5 0.116–0.130.Significant interactions between ventral feature and value of ven-tral feature were also found in the same time ranges,F~1,23! 54.5–5.2;p , .05. Ventral feature, value of ventral feature, andelectrode also showed significant interactions at these time win-dows, F~27,621! 5 3.9–8.7; p , .05–.001;E 5 0.104–0.162.Figures 10 and 11 suggest that the color SN shows a more centraldistribution, whereas shape SN appears mainly parietotemporal.Again, a profile analysis was necessary.
Difference waves were constructed to obtain color attentioneffects, collapsing the values of motion, and the same was done to
Figure 9. Difference waves obtained by subtracting event-related poten-tials ~ERPs! to standard stimuli with the unattended shape from ERPs tostandard stimuli with the attended shape when both motion and shape wereattended. Separate difference waves were computed for ERPs to stimuliwith the attended@~M1S1! 2 ~M1S2!# and the unattended@~M2S1! 2~M2S2!# motion direction.
Figure 10. Voltage maps representing the main findings for color attention effects. Attention values of motion are collapsed. Left: mean amplitudedistribution of the first significant effect of color attention, a frontal selection positivity~SP!. Center: distribution of the selection negativity~SN! forattention to color. Right: distribution of the late positive component~LPC! for attention to color. Each map uses an independent microvolt scale, based onthe corresponding extreme values.
730 M. Martín-Loeches, J.A. Hinojosa, and F.J. Rubia
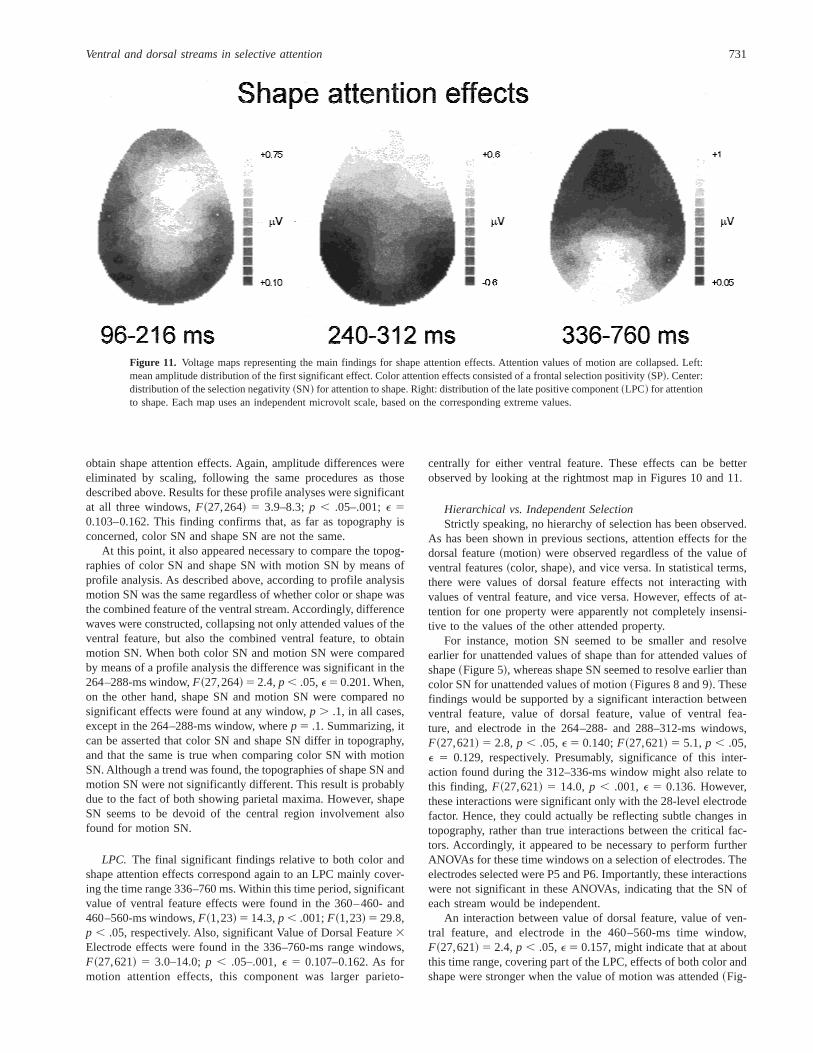
obtain shape attention effects. Again, amplitude differences wereeliminated by scaling, following the same procedures as thosedescribed above. Results for these profile analyses were significantat all three windows,F~27,264! 5 3.9–8.3;p , .05–.001;E 50.103–0.162. This finding confirms that, as far as topography isconcerned, color SN and shape SN are not the same.
At this point, it also appeared necessary to compare the topog-raphies of color SN and shape SN with motion SN by means ofprofile analysis. As described above, according to profile analysismotion SN was the same regardless of whether color or shape wasthe combined feature of the ventral stream. Accordingly, differencewaves were constructed, collapsing not only attended values of theventral feature, but also the combined ventral feature, to obtainmotion SN. When both color SN and motion SN were comparedby means of a profile analysis the difference was significant in the264–288-ms window,F~27,264! 5 2.4,p , .05,E5 0.201. When,on the other hand, shape SN and motion SN were compared nosignificant effects were found at any window,p . .1, in all cases,except in the 264–288-ms window, wherep 5 .1. Summarizing, itcan be asserted that color SN and shape SN differ in topography,and that the same is true when comparing color SN with motionSN. Although a trend was found, the topographies of shape SN andmotion SN were not significantly different. This result is probablydue to the fact of both showing parietal maxima. However, shapeSN seems to be devoid of the central region involvement alsofound for motion SN.
LPC. The final significant findings relative to both color andshape attention effects correspond again to an LPC mainly cover-ing the time range 336–760 ms. Within this time period, significantvalue of ventral feature effects were found in the 360–460- and460–560-ms windows,F~1,23! 5 14.3,p , .001;F~1,23! 5 29.8,p , .05, respectively. Also, significant Value of Dorsal Feature3Electrode effects were found in the 336–760-ms range windows,F~27,621! 5 3.0–14.0;p , .05–.001,E 5 0.107–0.162. As formotion attention effects, this component was larger parieto-
centrally for either ventral feature. These effects can be betterobserved by looking at the rightmost map in Figures 10 and 11.
Hierarchical vs. Independent SelectionStrictly speaking, no hierarchy of selection has been observed.
As has been shown in previous sections, attention effects for thedorsal feature~motion! were observed regardless of the value ofventral features~color, shape!, and vice versa. In statistical terms,there were values of dorsal feature effects not interacting withvalues of ventral feature, and vice versa. However, effects of at-tention for one property were apparently not completely insensi-tive to the values of the other attended property.
For instance, motion SN seemed to be smaller and resolveearlier for unattended values of shape than for attended values ofshape~Figure 5!, whereas shape SN seemed to resolve earlier thancolor SN for unattended values of motion~Figures 8 and 9!. Thesefindings would be supported by a significant interaction betweenventral feature, value of dorsal feature, value of ventral fea-ture, and electrode in the 264–288- and 288–312-ms windows,F~27,621! 5 2.8,p , .05, E5 0.140;F~27,621! 5 5.1,p , .05,E 5 0.129, respectively. Presumably, significance of this inter-action found during the 312–336-ms window might also relate tothis finding, F~27,621! 5 14.0, p , .001, E 5 0.136. However,these interactions were significant only with the 28-level electrodefactor. Hence, they could actually be reflecting subtle changes intopography, rather than true interactions between the critical fac-tors. Accordingly, it appeared to be necessary to perform furtherANOVAs for these time windows on a selection of electrodes. Theelectrodes selected were P5 and P6. Importantly, these interactionswere not significant in these ANOVAs, indicating that the SN ofeach stream would be independent.
An interaction between value of dorsal feature, value of ven-tral feature, and electrode in the 460–560-ms time window,F~27,621! 5 2.4,p , .05,E5 0.157, might indicate that at aboutthis time range, covering part of the LPC, effects of both color andshape were stronger when the value of motion was attended~Fig-
Figure 11. Voltage maps representing the main findings for shape attention effects. Attention values of motion are collapsed. Left:mean amplitude distribution of the first significant effect. Color attention effects consisted of a frontal selection positivity~SP!. Center:distribution of the selection negativity~SN! for attention to shape. Right: distribution of the late positive component~LPC! for attentionto shape. Each map uses an independent microvolt scale, based on the corresponding extreme values.
Ventral and dorsal streams in selective attention 731
ures 8 and 9!. However, it appeared necessary to perform an ad-ditional ANOVA on a selection of electrodes, as these interactionswere significant only with the electrode factor. Only one electrode,Pz, was used for these additional analyses in the 460–560-mswindow. Results indicated that the interaction between value ofdorsal feature and value of ventral feature remained significant,F~1,23! 5 5.8, p , .05, therefore confirming some degree ofmutual influence between streams with regard to LPC.
The motion LPC appears to be resolved earlier when color andshape present the unattended values~Figures 4 and 5!. Addition-ally, the color and shape LPCs seem to be resolved earlier whenmotion presents the unattended value~Figures 8 and 9!. The sig-nificant interaction between combined feature, value of motion,value of feature, and electrode in the 660–760-ms time window,F~27,621! 5 2.7, p , .05, E 5 0.154, would support these find-ings. Once again, however, these interactions were significant onlywith the electrode factor, so that it was necessary to perform anadditional ANOVA on a selection of electrodes. Again, Pz wasused for these additional analyses in the 660–760-ms window.
Results showed that the interaction between combined feature,value of motion, and value of feature remained significant,F~1,23! 5 4.0, p , .05.
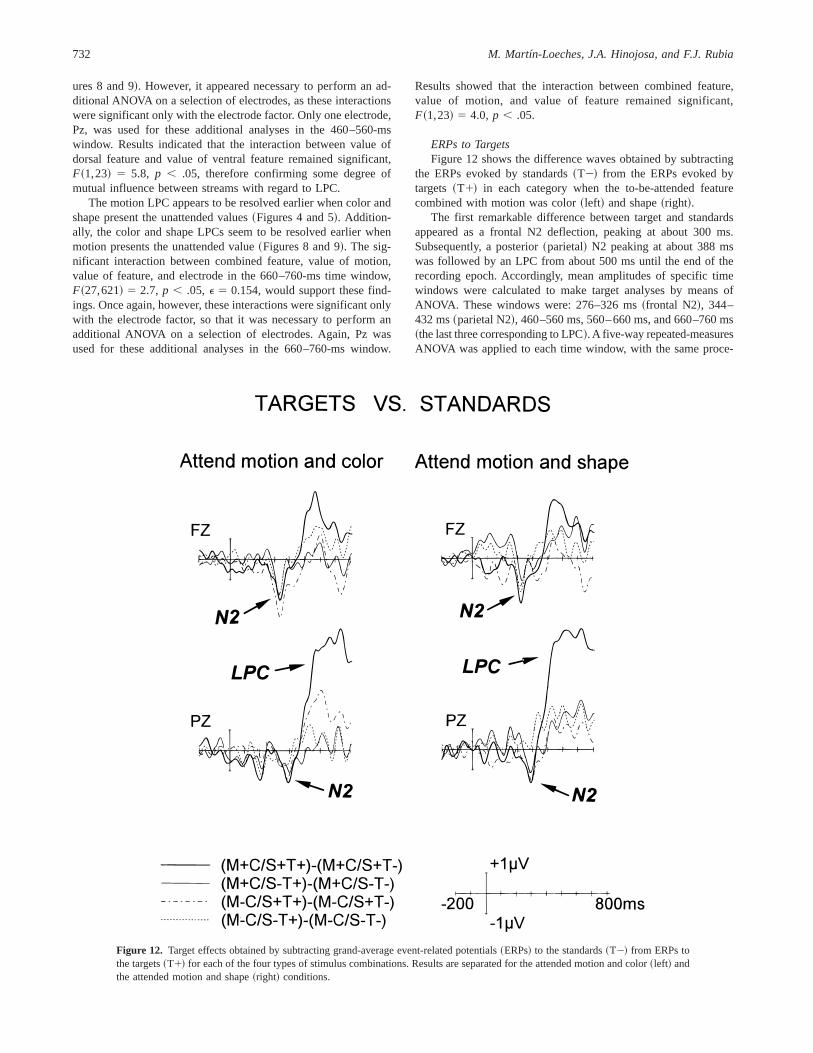
ERPs to TargetsFigure 12 shows the difference waves obtained by subtracting
the ERPs evoked by standards~T2! from the ERPs evoked bytargets ~T1! in each category when the to-be-attended featurecombined with motion was color~left! and shape~right!.
The first remarkable difference between target and standardsappeared as a frontal N2 deflection, peaking at about 300 ms.Subsequently, a posterior~parietal! N2 peaking at about 388 mswas followed by an LPC from about 500 ms until the end of therecording epoch. Accordingly, mean amplitudes of specific timewindows were calculated to make target analyses by means ofANOVA. These windows were: 276–326 ms~frontal N2!, 344–432 ms~parietal N2!, 460–560 ms, 560–660 ms, and 660–760 ms~the last three corresponding to LPC!. A five-way repeated-measuresANOVA was applied to each time window, with the same proce-
Figure 12. Target effects obtained by subtracting grand-average event-related potentials~ERPs! to the standards~T2! from ERPs tothe targets~T1! for each of the four types of stimulus combinations. Results are separated for the attended motion and color~left! andthe attended motion and shape~right! conditions.
732 M. Martín-Loeches, J.A. Hinojosa, and F.J. Rubia

dures followed as in the ANOVA for standards. The factors werealso the same as those used to analyze standard stimuli, but anadditional factor was included: target versus standard type of stim-ulus ~T1 vs. T2!, which will be referred to as type factor. Due tomainframe processor capacity, analyses were performed on a se-lection of anterior and posterior electrodes: F7, F3, Fz, F4, F8, C3,C4, P7, P3, Pz, P4, P8, PO7, PO1, PO2, and PO8.
In the 276–326-ms window, only a Type3 Value of Feature3Electrode interaction was found to be significant,F~15,345! 57.8, p , .0001,E 5 0.201. Accordingly, and by looking at Fig-ure 12, it seems that the frontal N2 showed significant results onlyunder certain circumstances, essentially for attended values of bothcolor and shape. Basically, exactly the same can be said regardingparietal N2 where, again, only the Type3 Value of Ventral Fea-ture3 Electrode interaction was significant,F~15,345! 5 3.2,p ,.05, E 5 0.244. Statistical results corresponding to LPC basicallyconfirm what was observed in Figure 12, although only for the660–760-ms time window: the conjunction of target and attendedvalues of both dorsal and ventral feature resulted in an LPC muchlarger than that for the other combinations~significant Type3Value of Dorsal Feature3 Value of Ventral Feature3 Electrodeinteraction,F @15,345# 5 3.1, p , .05, E 5 0.248!. Also at the660–760-ms window, there was a significant Type3 Value ofVentral Feature3 Electrode interaction,F~15,345! 5 3, p , .05,E 5 0.238. This result seems to be mainly related to the highervalues of LPC shown for attended values of color, combined withthe fact that target stimuli with attended values of both dorsal andventral features showed very large LPC.
Discussion
Observed attention effects for color and shape~ventral features!largely resemble those obtained in previous studies: an SP com-ponent peaking at about 200 ms, followed by an SN from about240 to 312 ms after stimulus onset, and finally an LPC around500 ms after stimulus onset. Similar results, both in terms of tim-ing and components involved, have been obtained for color orshape~Anllo-Vento & Hillyard, 1996; Hillyard & Münte, 1984;Karayanidis & Michie, 1996; Paz-Caballero and García-Austt, 1992;Smid et al., 1997; Wijers et al., 1989!. However, an early onset ofthe SP component mediated by the ventral stream~about 110 ms!has been reported rarely, but has been associated more typicallywith spatial location~e.g., Hillyard & Münte, 1984; Paz-Caballero& García-Austt, 1992; for a review, see Mangun, 1995!. It is ouropinion that the presence of these early findings for attention toventral features is mainly due to the fact that spatial location wasa completely irrelevant property in our study. This conclusionwould be in line with the few previous studies showing ventralstream attention effects starting at similar times. This has been thecase for color~Mangun & Hillyard, 1995!, orientation~Karayani-dis & Michie, 1997!, and size~Zani & Proverbio, 1995!. Only inthe study by Zani and Proverbio~1997! has a feature such asspatial frequency, processed by the ventral stream, been shown topresent such early attention effects combined with spatial location.
Some of our observations regarding attention to motion were inline with previous reports~Anllo-Vento & Hillyard, 1996; Girelli& Luck, 1997!, including the presence and timing of SN and LPCcomponents. The SN was found to show its main values in the240–312-ms period, therefore presenting a timing similar to that ofthe SNs for color and shape, although resolving slightly earlier.The LPC component for motion attention was also similar to theLPC found for color and shape.
A novel finding regarding attention to motion was the obser-vation of a significant effect previous to the SN. Even after aselection of critical electrodes to eliminate possible subtle changesin topography, in the 120–144-ms period we found significanteffects corresponding to an SP that peaked at about 140 ms. Thiswould be a notably earlier peaking for this component when com-pared with the only previous study considering the timing of se-lective attention to motion~Anllo-Vento & Hillyard, 1996!, whichfound it to be at about 230 ms. Again, the fact that spatial locationwas a completely irrelevant property in our study might be ad-duced. Accordingly, and at variance with the Anllo-Vento and Hill-yard ~1996! study, in the present study two properties of the samedorsal stream would not be “competing” in the allocation of se-lective attention. It seems, consequently, that in the presence ofattention to location, effects of attention to motion, although pro-cessed by the same stream, appear later. It is also important toremember that spatial location is indicated immediately at stimulusonset, whereas motion information can be extracted only after thestimulus “moves.” In the present study, apparent motion was in-duced by a 50-ms gap between two sequential stimuli. In this line,the previously reported hierarchical relationship between spatiallocation and motion selection is not, therefore, surprising, in thesense that no motion attention effects have been observed forunattended values of spatial location~Anllo-Vento & Hillyard,1996!.
Our data seem to support a temporal advantage of the dorsalstream relative to the ventral stream when peak amplitudes aloneare considered. Earlier peak amplitudes might indicate that partialanalyses related to selective attention were completed earlier in thedorsal than in the ventral stream. However, the processes under-taken by the two streams started at similar times, and significantresults appeared even earlier for ventral stream attention effects.Nevertheless, an additional but pertinent comment must be madein this regard: Ventral stream information was present at stimulusonset, whereas dorsal stream information required a 50-ms delayfor stimulus motion to occur. Hence, a temporal prevalence of thelatter over the former might have been more obvious and promi-nent if the two types of information were available simultaneously.Despite the temporal disadvantage of the dorsal stream, the twopathways showed their earliest attentional effects at similar times,and the dorsal effects resolved earlier than the ventral effects.These findings might be supporting some degree of temporal prev-alence of the “where” over the “what” pathway.
However, our earliest motion attention effects could be inter-preted in an alternative manner. Despite our efforts to make motiondirection independent of location, one could argue that our earliestmotion attention effects appeared at that time because they arebased, at least partially, on spatial location information. For ex-ample, the comparison between the location of the first member ofeach pair of stimuli and the location of the second might be ameans of detecting movement direction, something that is intrinsicto a simulated movement. Given the small area in which the stim-uli could appear, it might also be argued that attending to one-halfof the stimulus area was equivalent to attending the direction to-ward that half, it therefore being also possible to resolve the taskbased on spatial location information.1 Finally, motion SP was
1One reviewer pointed out the possibility that a certain proportion ofsecond flashes would fall on the left side of the screen in leftward motiontrials and on the right side in rightward motion trials. If this were the case,our stimulation procedure might be improved by a slight expansion of thestimulation area.
Ventral and dorsal streams in selective attention 733

found to be of small amplitude, even not particularly larger thannoise level or than other nonsignificant effects. For these reasons,this effect might be interpreted as a residue of a P1 effect based onlocation selection.
Nevertheless, other studies suggest that even simulated motionengages motion-related brain areas~e.g., MT-MST areas; Kaneokeet al., 1997!. It also seems unlikely that the subjects made such afine-grained spatial analysis as to attend to the position of thesecond flash, given both the short interstimulus interval and theshort interstimulus distance~50 ms and 0.58, respectively!. Finally,the well-known pattern for attention to spatial location is a P1enhancement followed by an N1 enhancement, which is differentfrom the pattern found here for motion. Our first motion effectsoccur mostly after the P1 falling slope, and an analysis of the P1latency window revealed no significant effects. No N1 enhance-ment could be found in our results. Moreover, the topography ofthe results for motion seems to differ from the parietal distributionof P10N1 enhancements, particularly in the case of motion atten-tion effects when the attended feature of the ventral stream wasshape, as they appeared as frontocentral. In any case, studies ofselective attention to motion are certainly scarce, so that furtherresearch would appear to be necessary to both confirm presentresults and improve the limitations imposed by the scarcity ofstudies on our knowledge about selective attention to motion.
Further research is also probably needed to determine whyearliest motion attention effects appeared to differ in topographydepending on the combined feature of the ventral stream. When thecombined ventral feature was color, early attention effects of mo-tion were predominantly parietooccipital. On the other hand, whenthe combined ventral feature was shape, early motion attentioneffects were mainly frontocentral. This finding is interesting, butdifficult to explain with the available data. An important fact to beconsidered is that the different topography was independent ofwhether the ventral features presented the attended or the un-attended values. Rather, the topography depended on which ventralfeature had to be attended. In this regard we consider the possi-bility that early motion effects may in fact be reflecting outputprocesses. In other words, the early motion effects may be reflect-ing not merely the motion information processing itself, but rathera partial outcome of this processing that takes different paths, ascolor and shape will also be subsequently processed by partiallydifferent subsystems~Corbetta et al., 1990!.
Regarding the other main objective of the present study, that is,to explore the hierarchical relationship between the ventral anddorsal streams, it appears that the relative temporal relationshipfound between the two subsystems did not entail a selection hier-archy in the common and strict sense when movement was theonly attentional cue for the dorsal stream. According to a hierar-chical view, the selection of an attribute does not take place forunattended values of an earlier selected attribute. Here, attentionalprocesses of the ventral stream~color, shape! did take place forunattended values of the property analyzed by the dorsal stream~motion!. Therefore, the two streams seem to be working in par-allel, in a relatively independent and exhaustive mode~using theterminology of Hansen & Hillyard, 1983!. This does not seem tobe the case, however, when the property of the dorsal stream usedas attentional cue was spatial location, as already mentioned, thoughseveral studies indicate that this assertion is not 100% true~Hill-yard & Münte, 1984; Zani & Proverbio, 1997!.
However, although a selection hierarchy or contingent relation-ship between the two streams has not, strictly speaking, been found,the selection of an attribute by one stream was not completely
independent of the selection of the attribute by the other stream.These mutual relationships mainly affected the LPC component.The motion LPC resolved earlier when color and shape presentedthe unattended value. Additionally, the color and shape LPCs re-solved earlier when motion presented the unattended value. Through-out part of the LPC component, effects of both color and shapewere stronger when the value of motion was the attended stimulus.Accordingly, the dorsal and the ventral streams seem to processtheir corresponding stimulus attributes in a not completely inde-pendent manner. What these findings indicate, therefore, is that thetwo streams are working in parallel, but have a mutual relationshipat some point during the processing. This mutual relationship mightbe described as having the form of a “cascade,” that is, results ofpartial analyses made by one stream are already transmitted to theother stream, resulting in a sort of continuous transmission ofinformation between the two subsystems~McClelland, 1979; Miller,1991!. Results of other ERPs studies also support the existence ofthis kind of information transmission between cognitive processes~e.g., Miller, 1991; Paz-Caballero & García-Austt, 1992; Smidet al., 1997!.
In sum, our results using dorsal stream properties other thanspatial location reveal that the temporal prevalence of this streamover the ventral stream can still be supported, though in relativeterms. Additionally, the hierarchical relationship between streamsusually found when spatial location is used as the relevant propertyof the dorsal stream, that is, the processing by the ventral stream iscontingent upon the dorsal stream, cannot be further supported. Inthis regard it might be more appropriate to state that the contingentrelationship is restricted to spatial location. Nevertheless, this hi-erarchy of spatial location has been called into question by recentreports~Zani & Proverbio, 1997!.
Another interesting set of findings of the present study re-lates to the topography of the SN component. We obtained dif-ferent SNs depending on the feature attended. Motion SN showeda parietocentral scalp distribution, but this distribution appearedto be slightly different depending on the combined ventral fea-ture ~more central when color was attended, more parietal whenshape was attended!, but a statistical procedure~profile analysis;see McCarthy & Wood, 1985! supported the unitary character ofmotion SN regardless of the attended ventral feature. On theother hand, color and shape, the ventral features, showed differ-ent SNs. Color SN appeared as largely central, while shape SNwas parietotemporal. Profile analyses confirmed this discrep-ancy, as well as the comparison between color and motion SN.Shape SN was not significantly different from motion SN, but atrend toward significance was found.
Anllo-Vento and Hillyard~1996! found that the SNs for motionand color displayed similar topography in an initial phase butdiffered significantly in a later phase. Smid et al.~1997! also foundthat SN topography for color and shape was not the same. Topog-raphies for color, shape, and~perhaps to a lesser extent! motionSNs in these studies resemble those found here. Considering presentand previous results, it seems that there are specialized brain areasinvolved in forming the SN. Moreover, it seems that these distinc-tive areas are not only specific for the ventral and dorsal streams,but also within the ventral stream. These data would neverthelessbe at variance with those of Girelli and Luck~1997!. These authorsfound a unitary distribution of ERPs effects of attention to color,orientation, and motion, supporting the existence of a commonlocus for selecting targets regardless of the features to be attended,and independent of the processing stream. However, in the studyby Girelli and Luck, subjects were required to detect a single,
734 M. Martín-Loeches, J.A. Hinojosa, and F.J. Rubia

salient feature, which is not usually regarded as an attention-demanding task~e.g., Braun & Sagi, 1990!. In this line, Girelli andLuck asserted in their conclusions that the unitary distribution theyfound would rather relate to operations specific to their tasks,namely the selection of a target object from an array of distractors.
Finally, we should briefly discuss the target results. In the presentstudy, target stimuli were defined by paired flashes with a longer-than-standard SOA. As is usually the case~e.g., Johnson, 1993!with the detection of infrequent, task-relevant target stimuli, it wasaccompanied by a late ERPs complex composed of an N2~orN200! with both frontal and parietal maxima~at about 300 and388 ms, respectively!, followed by an LPC~also known as P300!from about 500 to 800 ms. The N2 was nevertheless found to besignificant only for attended values of the ventral stream~color orshape!. This finding would indicate that at this stage the analysis ofthe target-defining feature~slower apparent motion! was contin-
gent upon color or shape selection. However, at this stage move-ment direction information seems to be irrelevant for N2 values,because they were equivalent regardless of the values of motiondirection. This result is noteworthy, because the analysis of motiondirection is probably already finished or at least at an advancedstage by the time of N2 appearance. Therefore, it appears that theconjunction of information relative to motion direction and thatrelative to speed of movement would take place after the conjunc-tion of information relative to ventral feature~color, shape! andthat relative to movement speed~presumably reflected by the N2processes!. Thereafter, only in the case that motion direction presentsthe attended value, is a large LPC obtained. At this last stage,however, color information also seems to have substantial weight,as attended values of color are able to produce some significantlylarge ~albeit small! values of LPC even for unattended values ofmotion direction.
REFERENCES
American Electroencephalographic Society.~1991!. Guidelines for stan-dard electrode position nomenclature.Journal of Clinical Neurophys-iology, 3, 38–42.
Anllo-Vento, L., & Hillyard, S. A. ~1996!. Selective attention to the colorand direction of moving stimuli: Electrophysiological correlates ofhierarchical feature selection.Perception and Psychophysics, 58, 191–206.
Braun, J., & Sagi, D.~1990!. Vision outside the focus of attention.Per-ception and Psychophysics, 48, 45–58.
Britten, K. H., Newsome, W. T., & Saunders, R. C.~1992!. Effects ofinferotemporal cortex lesions on form-form-motion discrimination inmonkeys.Experimental Brain Research, 88, 292–302.
Clark, V. P., & Hillyard, S. A.~1996!. Spatial selective attention affectsearly extrastriate but not striate components of the visual evoked po-tential.Journal of Cognitive Neuroscience, 8, 387–402.
Corbetta, M., Miezin, F. M., Dobmeyer, S., Shulman, G. L., & Petersen,S. E. ~1990!. Attentional modulation of neural processing of shape,color, and velocity in humans.Science, 248, 1556–1559.
Girelli, M., & Luck, S. J.~1997!. Are the same attentional mechanisms usedto detect visual search targets defined by color, orientation, and mo-tion? Journal of Cognitive Neuroscience, 9, 238–253.
Gomez-Gonzalez, C. M., Clark, V. P., Fan, S., Luck, S., & Hillyard, S. A.~1994!. Sources of attention-sensitive visual event-related potentials.Brain Topography, 7, 41–51.
Hansen, J. C., & Hillyard, S. A.~1983!. Selective attention to multidimen-sional auditory stimuli.Journal of Experimental Psychology: HumanPerception and Performance, 9, 1–19.
Harter, M. R., & Aine, C. J.~1984!. Brain mechanisms of visual selectiveattention. In R. Parasuraman & D. R. Davies~Eds.!, Varieties of at-tention ~pp. 293–321!. Orlando, FL: Academic Press.
Heinze, H. J., Mangun, G. R., Burchert, W., Hinrichs, H., Scholz, M.,Münte, T. F., Gos, A., Scherg, M., Johannes, S., Hundeshagen, H.,Gazzaniga, M. S., & Hillyard, S. A.~1994!. Combined spatial andtemporal imaging of brain activity during visual selective attention inhumans.Nature, 372, 543–546.
Heslenfeld, D. J., Kenemans, J. L., Kok, A., & Molenaar, P. C. M.~1997!.Feature processing and attention in the human visual system: An over-view. Biological Psychology, 45, 183–215.
Hillyard, S. A., Anllo-Vento, L., Clark, V. P., Heinze, H.-J., Luck, S. J., &Mangun, G. R.~1995!. In A. F. Kramer, M. G. H. Coles, & G. D. Logan~Eds.!, Converging operations in the study of visual selective attention~pp. 107–138!. Washington, DC: American Psychological Association.
Hillyard, S. A., & Münte, T. F.~1984!. Selective attention to color andlocation: An analysis with event-related brain potentials.Perceptionand Psychophysics, 36, 185–198.
Jennings, J. R., & Wood, C. C.~1976!.The E-adjustment procedure forrepeated-measures analyses of variance.Psychophysiology, 13, 277–278.
Johnson, R.~1993!. On the neural generators of the P300 component of theevent-related potential.Psychophysiology, 30, 90–97.
Kaneoke, Y., Bundoe, M., Koyama, S., Suzuki, H., & Kakigi, R.~1997!.Human cortical area responding to stimuli in apparent motion.Neu-roReport, 8, 677–682.
Karayanidis, F., & Michie, P. T.~1996!. Frontal processing negativity in avisual selective attention task.Electroencephalography and ClinicalNeurophysiology, 99, 38–56.
Karayanidis, F., & Michie, P. T.~1997!. Evidence of visual processingnegativity with attention to orientation and color in central space.Elec-troencephalography and Clinical Neurophysiology, 103, 282–297.
Kenemans, J. L., Kok, A., & Smulders, F. T. Y.~1993!. Event-relatedpotentials to conjunctions of spatial frequency and orientation as afunction of stimulus parameters and response requirements.Electro-encephalography and Clinical Neurophysiology, 88, 51–63.
Mangun, G. R.~1995!. Neural mechanisms of visual selective attention.Psychophysiology, 32, 4–18.
Mangun, G. R., & Hillyard, S. A.~1995!. Mechanisms and models ofselective attention. In M. D. Rugg & M. G. H. Coles~Eds.!, Electro-physiology of mind: Event-related brain potentials and cognition~pp. 40–85!. Oxford, UK: Oxford University Press.
Maunsell, J. H. R., & Ferrera, V. P.~1994!. Attentional mechanisms invisual cortex. In M.S. Gazzaniga~Ed.!, The cognitive neurosciences~pp. 451–461!. Cambridge, MA: MIT Press.
McCarthy, G., & Wood, C. C.~1985!. Scalp distributions of event-relatedpotentials: An ambiguity associated with analysis of variance models.Electroencephalography and Clinical Neurophysiology, 62, 203–208.
McClelland, J. L.~1979!. On the time relations of mental processes: Aframework for analyzing processes in cascade.Psychological Review,86, 287–330.
McIntosh, A. R., Grady, C. L., Ungerleider, L. G., Haxby, J. V., Rapoport,S. I., & Horwitz, B. ~1994!. Network analysis of cortical visual path-ways mapped with PET.Journal of Neuroscience, 14, 655–666.
Miller, J. ~1991!. Discrete versus continuous information processing: in-troduction and psychophysiology. In C. H. M. Brunia, G. Mulder, &M. N. Verbaten~Eds.!, Event-related brain potential research~EEGSupplement 42, pp. 244–259!. Amsterdam: Elsevier.
O’Craven, K. M., Rosen, B. R., Kwong, K. K., Treisman, A., & Savoy,R. L. ~1997!. Voluntary attention modulates fMRI activity in humanMT-MST. Neuron, 18, 591–598.
Oldfield, R. C. ~1971!. The assessment and analysis of handedness: TheEdinburgh Inventory.Neuropsychologia, 9, 97–113.
Paz-Caballero, M. D., & García-Austt, E.~1992!. ERP components relatedto stimulus selection processes.Electroencephalography and ClinicalNeurophysiology, 82, 369–376.
Rugg, M. D., & Coles, M. G. H.~1995!. The ERP and cognitive psychol-ogy: Conceptual issues. In M. D. Rugg & M. G. H. Coles~Eds.!,Electrophysiology of mind: Event-related brain potentials and cogni-tion ~pp. 27–39!. Oxford, UK: Oxford University Press.
Semlitsch, H. V., Anderer, P., Schuster, P., & Preelich, O.~1986!. A solutionfor reliable and valid reduction of ocular artifacts, applied to the P300ERP.Psychophysiology, 23, 695–703.
Smid, H. G. O. M., Jakob, A., & Heinze, H.-J.~1997!. The organization ofmulti-dimensional selection on the basis of color and shape: An event-related brain potential study.Perception and Psychophysics, 59, 693–713.
Tootell, R. B. H., Dale, A. M., Sereno, M. I., & Malach, R.~1996!. New
Ventral and dorsal streams in selective attention 735

images from human visual cortex.Trends in Neurosciences, 19, 481–489.
Ungerleider, L. G., Courtney, S. M., & Haxby, J. V.~1998!. A neural systemfor human visual working memory.Proceedings of the National Acad-emy of Sciences (USA), 95, 883–890.
Ungerleider, L. G., & Haxby, J. V.~1994!. “What” and “where” in thehuman brain.Current Opinion in Neurobiology, 4, 157–165.
Wijers, A. A., Mulder, G., Okita, T., & Mulder, L. J. M.~1989!. Event-related potentials during memory search and selective attention to lettersize and conjunctions of letter size and color.Psychophysiology, 26,529–547.
Yamaguchi S., & Kobayashi, S.~1998!. Contributions of the dopaminergicsystem to voluntary and automatic orienting of visuospatial attention.The Journal of Neuroscience, 18, 1869–1878.
Zani, A., & Proverbio, A. M.~1995!. ERP signs of early selective attentioneffects to check size.Electroencephalography and Clinical Neurophys-iology, 95, 277–292.
Zani, A., & Proverbio, A. M. ~1997!. Attentional modulation of shortlatency ERPs by selective attention to conjunction of spatial frequencyand location.Journal of Psychophysiology, 11, 21–32.
~Received May 18, 1998;Accepted January 13, 1999!
736 M. Martín-Loeches, J.A. Hinojosa, and F.J. Rubia




















