
Single-Cell Transcriptional Analysis of Neuronal Progenitors

RESEARCH ARTICLE
Inhibitors of CXCR4 Affect the Migration andFate of CXCR4� Progenitors in the DevelopingLimb of Chick EmbryosFaisal Yusuf,1 Rizwan Rehimi,1 Gabriela Morosan-Puopolo,1 Fangping Dai,1 Xiaobing Zhang,2,3 andBeate Brand-Saberi1*
Chemokines and their receptors play major roles in numerous physiological and pathological processesduring development and disease. CXCR4 is the most abundantly expressed chemokine receptor duringdevelopment. In contrast to other chemokine receptors, CXCR4 binds and is activated exclusively by itsligand stromal derived factor-1 (SDF-1) or CXCL12. SDF-1 signaling has a wide range of effects on CXCR4-expressing cells depending on the cell type ranging from cell growth to adhesion, chemotaxis, andmigration. CXCR4 also serves as a co-receptor for HIV-1 entry into T-cells and has been implicated in thepathogenesis of rheumatoid arthritis and cancer growth and invasion. Numerous inhibitors andantagonists of CXCR4 have been produced and are being tested for their efficiency to target its role inpathogenesis. Our initial expression analysis revealed that CXCR4 is expressed by the migrating myogenicand angiogenic precursors in the developing chick limb. In this study, we used the most specific peptidicinhibitors of CXCR4, T140 and its analog TN14003, to analyse the effect of blocking CXCR4/SDF-1 signalingon the undetermined bioptent migratory progenitors in the developing chick limb. Our results point todefects in migration and an altered differentiation program of these CXCR4-expressing progenitor pool inthe limb. Developmental Dynamics 235:3007–3015, 2006. © 2006 Wiley-Liss, Inc.
Key words: chick; CXCR4; cell migration; myogenesis; angiogenesis; differentiation
Accepted 2 August 2006
INTRODUCTION
The epithelial somites undergo mor-phological changes leading to the for-mation of compartments (Brand-Sa-beri et al., 1996). The ventral halves ofthe somites form the sclerotome andthe dorsal halves the dermomyotome;only the latter retains its epithelialstate. The sclerotome gives rise to thebone, connective tissue, and cartilagethat surround the neural structures,whereas the dermomyotome contains
progenitors of the myogenic, dermo-genic, and angiogenic lineage (Brand-Saberi and Christ, 2000). At all axiallevels, the dermomyotome fuels themyotome with myoblasts that willeventually give rise to the deep mus-cles of the back or epaxial muscles andangioblasts that form the vascularnetwork around the newly formedstructures. However, at the cervicaland limb level, the dermomyotomealso gives rise to the migrating myo-
blasts, angioblasts, as well as unde-termined progenitors that undertakelong-range migrations to reach theirfinal sites where they settle down anddifferentiate to form muscle and endo-thelia (Christ et al., 1977; Ordahl andLe Douarin 1992). In the limb bud, themigrating myoblasts enter the desig-nated premuscle anlage, proliferate,and differentiate. A number of mark-ers have been identified that allow usto trace these migrating myogenic
1Institute of Anatomy and Cell Biology, Department of Molecular Embryology, University of Freiburg, Freiburg, Germany2School of Life Science, Lanzhou University, Lanzhou, China3Department of ENT Head-Neck, First Hospital of Lanzhou University, Lanzhou, ChinaGrant sponsor: Baden-Wurttemberg; Grant sponsor: EC; Grant number: 511978.*Correspondence to: Prof. Dr. Beate Brand-Saberi, Institute of Anatomy and Cell Biology, Department of Molecular Embry-ology, Freiburg University, Albertstrasse 17, 79104 Freiburg, Germany. E-mail: [email protected]
DOI 10.1002/dvdy.20951Published online 6 September 2006 in Wiley InterScience (www.interscience.wiley.com).
DEVELOPMENTAL DYNAMICS 235:3007–3015, 2006
© 2006 Wiley-Liss, Inc.

myoblasts from their initial static po-sition in the ventrolateral lip of thedermomyotome to their final sites ofdifferentiation in the premusclemasses (Birchmeier and Brohmann,2000). Pax3, Lbx1, and a tyrosine ki-nase receptor, cMet, are considered tomark the myogenic precursor cells inthe lateral lip of the dermomyotome.Pax3 is necessary for the correct spec-ification and establishment of themyogenic precursor pool in the ventraldermomyotome as in Pax3 mutants(Splotch) hypaxial muscle develop-ment is impaired (Franz et al., 1993;Bober et al., 1994 Tajbakhsh et al.,1997). Initially described as a myo-genic cell marker, work by Kardonand colleagues has pointed towards abipotential Pax3-expressing popula-tion in the limb contributing to boththe myogenic and endothelial com-partment (Kardon et al., 2002).
cMet interacts with its secretedligand, Scatter Factor/HepatocyteGrowth Factor, to cause delaminatonof the myogenic precursors that are toundertake long-range migrations. IncMet or SF/HGF mutant mice, all themuscles that derive from migratingmyogenic precursors are missing(Bladt et al., 1995). The only gene ex-pressed exclusively by the migratinghypaxial myogenic precursors is Lbx1.It is a homeodomain transcription fac-tor, and its impairment results in mi-gration defects affecting the myogenicprecursors that populate the limb,whereas other migrating myoblastsfind their targets (Schafer and Braun,1999; Gross et al., 2000). After delami-nation from the lateral lip of the der-momyotome, the myogenic precursorscontinue to express Pax3, cMet, andLbx1 along their migratory routes totheir final sites of differentiation.These myogenic progenitors prolifer-ate and later begin to express myo-genic regulatory factors, MyoD, Myf5,and Mrf4, which determine their ter-minal differentiation into myotubesand muscle fibres.
Very recently, CXCR4, a new markerfor the bipotent migratory progenitorshas been described (Vasyutina et al.,2005). CXCR4 is a chemokine receptorwith SDF-1� (CXCL12) as its sole li-gand. Its role has been well documentedin HIV-1 pathogenesis as a co-factor forenv-mediated HIV-1 and HIV-2 fusionand entry into CD4� expressing T-cells
(Deng et al., 1996; Doranz et al., 1996;Feng et al., 1996).
The CXCR4/SDF-1� axis has beenimplicated in numerous physiologicaland pathological processes rangingfrom cell growth, adhesion, chemotaxis,and migration to cancer metastasis andHIV pathogenesis (Murdoch, 2000).
Due to its various implications inmany pathological processes like can-cer metastasis, HIV pathogenesis, andrheumatoid arthritis, CXCR4 has be-come an effective therapeutic target.Efforts have been made by severalgroups to design CXCR4 antagoniststhat could be of therapeutic impor-tance in the future. Today there aremany known CXCR4 antagonists thathave been described (Tamamura etal., 2005; Princen and Schols, 2005),with different levels of efficiency ininhibiting HIV entry into target cellsor inhibiting CXCL12-induced migra-tion of breast cancer cells, leukaemiaT cells, pancreatic cancer cells, smallcell lung cancer cells, chronic lympho-cytic leukaemia B cells, and pre-Bacute lymphoblastic leukaemia cells(Tamamura et al., 2003a,b; Tama-mura and Fujii 2005; Hartmann et al.,2005). Tamamura and collegues claimto have developed highly selectiveCXCR4 antagonists, T22 and T140(18-mer and 14-mer peptides, respec-tively), which strongly suppress T-cellline-tropic HIV-1 (X4-HIV-1) entrythrough their specific binding toCXCR4 (Tamamura and Fujii, 2004)The most efficient and specific inhibi-tors of CXCR4 described to date arethe short peptide inhibitor T140 andits analogs. The inhibitors were origi-nally designed as HIV entry inhibi-tors. A peptidic CXCR4 antagonistT140 efficiently blocks the entry of Tcell line-tropic strains of HIV-1 (X4-HIV-1) into target cells (Tamamura etal., 2003b). Recently, Meike Burgerand collegues have studied the effectof T140 analogs on the CXCR4-SDF-1axis in vitro. TN14003 antagonizedthe antiapoptotic effect of syntheticCXCL12 and stromal cell-mediatedprotection of CLL cells from spontane-ous apoptosis (Burger et al., 2005).CXCR4-blocking agents effectively an-tagonize CXCL12-induced migratoryand signaling responses and stromalprotection of CLL cells from spontane-ous or fludarabine-induced apoptosis.Similar effects were observed by
Juarez and collegues for acute lym-phoblastic leukemia (ALL) (Juarez etal., 2003). They also report that T140works by blocking the CXCR4/SDF-1axis acting as a competitive inhibitorthat hinders SDF-1 mediated cell pro-liferation and cell migration of ALLcells. Moreover, T140 analogs have alsobeen shown to be of use as anti-meta-static agents in breast cancer and forthe treatment of rheumatoid arthritis(Tamamura et al., 2003a, 2004).
Earlier we have shown thatCXCR4� cells have been found in thepremuscle masses and in the develop-ing vessels of the limb bud in chickembryo (Yusuf et al., 2005). To targetthe CXCR4-expressing cells in the de-veloping avian limb, we used highlyspecific CXCR4 inhibitors, T140 andits analog TN14003, to block the SDF-1-driven responses mediated via theCXCR4 receptor. This is the first re-port of the use of these inhibitory pep-tides in the developing embryo. Weexamine the effect of CXCR4 inhibi-tors, T140 (14er peptide) andTN14003 (14er peptide), in avian em-bryos. The CXCR4� cells were effec-tively inhibited in their migratorypath resulting in a decreased numberof precursors reaching their respectivepremuscle anlage in the limb. More-over, we observed that the CXCR4�
cells that do not reach their destinedlocation in the premuscle anlage wereforced into an angiogenic fate at thesite of T140/TN14003 contact. We con-clude that SDF-1 signaling is neces-sary for the appropriate migration,and blocking this by using specificpeptidic inhibitors of CXCR4 effi-ciently interferes with the migrationof the CXCR4-expressing progenitorpool in the limb. In doing so, thesecells are denied access to sites of dif-ferentiation where they would nor-mally encounter differentiation sig-nals. The inability to access thesedifferentiation sites, forces these cellsto adopt a default angiogenic fate.
RESULTS
Effect of T140/TN14003 onCXCR4� Cells in theDeveloping Limb Bud ofChick Embryo
We implanted acrylic beads or affigelbeads soaked in T140 and TN14003 at
3008 YUSUF ET AL.

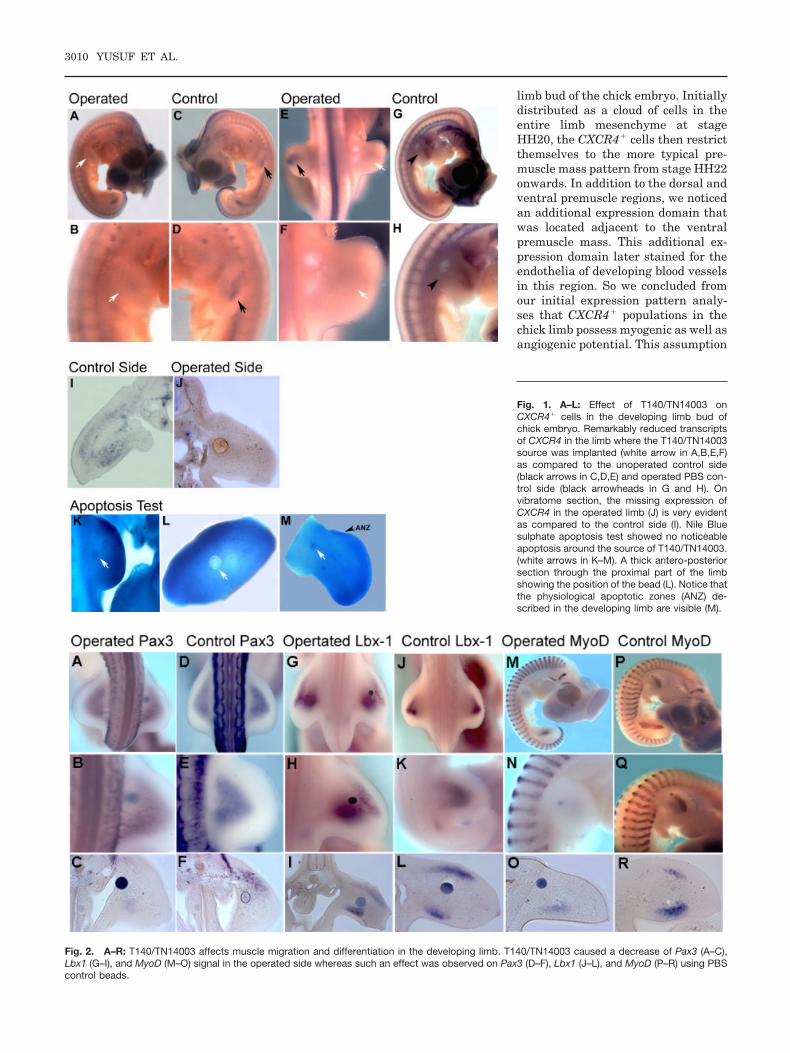
stage HH17–19 in the proximal regionof the limb bud. We were firstly inter-ested to observe the effect of the inhib-itors on their specific target, the che-mokine receptor CXCR4. The CXCR4expression in the limb where the beadwas implanted was remarkably re-duced (n � 11) (Fig. 1A,B,E,F,J). Thenumber of CXCR4� cells in the limbwere considerably less in the limbwith the T140/TN14003 bead as com-pared to the control side (Fig. 1C,D,I).PBS beads showed no effect on theCXCR4� population in the limb bud(n � 4) (Fig. 1G,H). Similar effectswere observed using acrylic and affi-gel beads. Using concentrations lessthan 5–10 mg/ml of T140/TN14003produced little if any effect.
T140/TN4003 Do Not CauseApoptosis in the DevelopingLimb
To ensure that the observed effects ofCXCR4 inhibitors on the CXCR4 pop-ulations in the limb was not a result ofapoptosis, we performed an apoptosistest following the T140/TN14003 beadimplantation at intervals of 8, 12, and24 hr. No apoptosis was observedaround the bead (n � 15) (Fig.1K–M).The physiological anterior necroticzone (ANZ) described in the develop-ing limb (Zuzarte-Luis and Hurle,2002) can be observed (Fig. 1M).
T140/TN14003 Affects theMigration andDifferentiation of theMigratory Progenitors in theDeveloping Limb
During their course of migration, themyogenic precursors express Pax3. Af-ter having completed their long-rangemigration beginning at the ventrolat-eral lip of the dermomyotome and end-ing in their designated regions in limbmesenchyme, the myogenic precur-sors begin to terminally differentiateand express myogenic regulatory fac-tors like MyoD. It was of interest forus to gain insight into the fate of mus-cle development in the limb bud. Insitu hybridization done using a Pax3-specific probe at stage HH21–23showed also a decrease of Pax3� cellsmigrating into the limb bud (n � 7)(Fig. 2A–C) as compared to the control
side (Fig. 2D–F). However, the effectwas not as pronounced as that ob-served using the CXCR4 RNA probe.Similar effects were also observed onthe expression domains of anothermyogenic precursor marker, Lbx1(n � 6) (Fig. 2G–I). Control beadssoaked in PBS did not produce a sim-ilar decrease of the Pax3 and Lbx1(Fig. 2J–L) signal in the operated limb(n � 4 and n � 5, respectively). Atlater stages of development, at stageHH25 and above, we looked into thedifferentiation events of the myogenicprecursors in the limb using theMyoD-specific RNA probe. A decreaseof the MyoD signal was observed inthe operated limb (n � 9) (Fig. 2M–O)as compared to the unoperated controlside. Control operations with PBSbeads exhibited no decrease of theMyoD signal on the operated side (n �5) (Fig. 2P–R).
T140/TN14003 Can EnhanceCommitment to AngiogenicFate
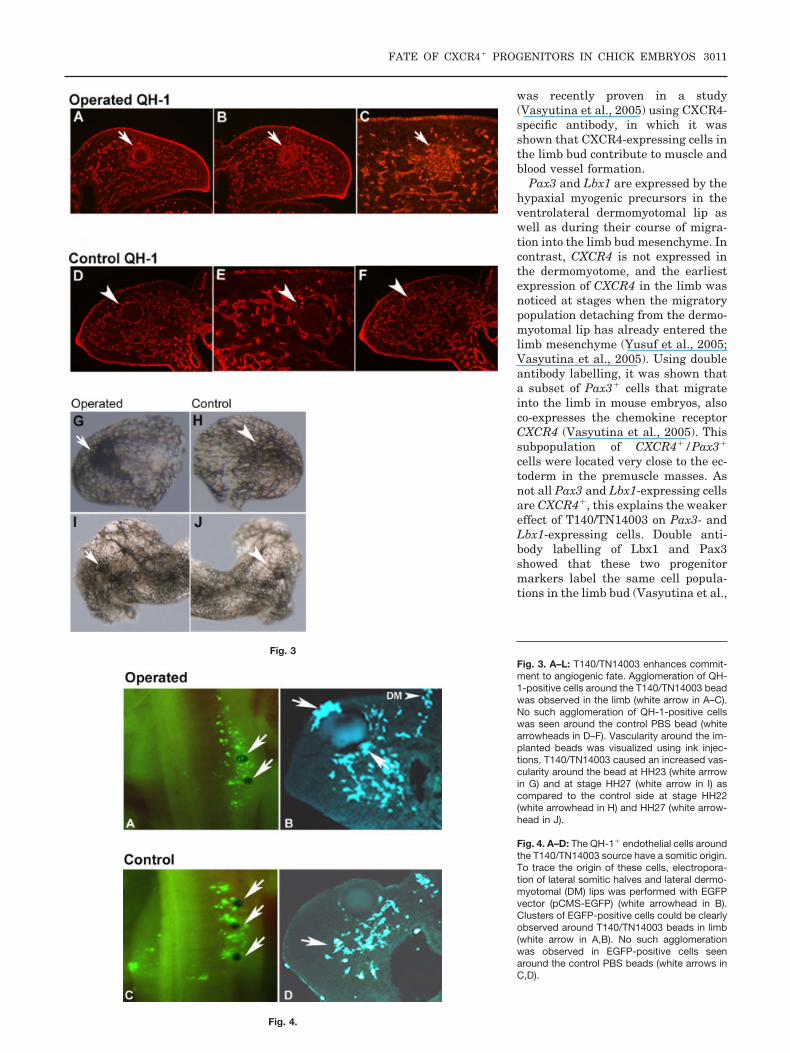
As there were fewer CXCR4� cellsfound in the limb bud mesenchyme,and there was a decrease in the mus-cle mass in the limb as observed byMyoD staining, we were interested toknow the fate of the CXCR4� cellsthat fail to reach the muscle anlageafter exposure to T140/TN14003. Weknow that a subpopulation ofCXCR4� cells entering the limb alsocontributes to vessel formation, andwe used a vascular marker, QH-1 an-tibody to observe the effect of CXCR4inhibitors on vessel formation in thelimb. Cryosections revealed an in-creased numbers of QH-1� cells ag-glomerating around the bead (n � 3)(Fig. 3A–C). The patterns of QH-1marking in the operated limb distantto the T140/TN14003 source were notaltered as compared to the non-oper-ated side (Fig. 3A,B). No such agglom-eration of QH-1� cells was observedaround PBS beads (n � 4) (Fig. 3D–F).
TN140/TN14003 Cause anIncrease in Vessel Formation
The presence of QH-1 cells around thebead prompted us to check for changesin the vascular pattern that may occurin the vicinity of the implanted sourceof the T140/TN14003. To this effect,
we injected Indian ink into thevitelline vessels of stage HH22–26embryos to visualize the vascular net-work. We observed an increased vas-cularization at stage HH23 (n � 6,Fig. 3G) and at later stages, HH27, anincrease of vessel density around thesource of T140/TN14003 (n � 4, Fig.3I) as compared to the control sides(Fig. 3H and J).
The QH-1� Endothelial CellsAround the T140/TN14003Source Have a SomiticOrigin
In connection with the reduction ofmuscle precursor cells after T140/TN14003 application, an alteration ofthe differentiation program of the bi-potent migratory precursors from thesomites could be suspected. Thus, wewere further interested to trace theorigin of the of QH-1� cells that ag-glomerate around the T140/TN14003source. To find this out, we electropo-rated lateral somitic halves and lat-eral dermomyotomal lips from wherethe myogenic and endothelial precur-sors originate and migrate into thelimb mesenchyme with a reporterEGFP vector (pCMS-EGFP). Wefound the agglomeration of EGFP-ex-pressing cells around the source ofT140/TN14003 beads (n � 5, Fig.4A,B). As the origin of these EGFP liesin the somites that were initially elec-troporated, we can conclude that theQH-1-expressing cells that jamaround the bead have a somitic origin.Control beads soaked in PBS showedno such agglomeration around thebeads (n � 5, Fig. 4C,D). Hence, wecan conclude that differentiation isalso perturbed by interfering withCXCR4 signaling.
DISCUSSION
In this study, we examined the effectof a specific peptidic CXCR4 inhibitorT140 and its analog TN14003 on themigratory behaviour and differentia-tion of CXCR4� populations in the de-veloping limb of the chick embryo,which later contribute to muscle andblood vessel formation. We had re-ported earlier the expression patternof CXCR4 in the chick embryo (Yusufet al., 2005). Starting from stage 19,CXCR4� cells can be traced in the
FATE OF CXCR4� PROGENITORS IN CHICK EMBRYOS 3009
limb bud of the chick embryo. Initiallydistributed as a cloud of cells in theentire limb mesenchyme at stageHH20, the CXCR4� cells then restrictthemselves to the more typical pre-muscle mass pattern from stage HH22onwards. In addition to the dorsal andventral premuscle regions, we noticedan additional expression domain thatwas located adjacent to the ventralpremuscle mass. This additional ex-pression domain later stained for theendothelia of developing blood vesselsin this region. So we concluded fromour initial expression pattern analy-ses that CXCR4� populations in thechick limb possess myogenic as well asangiogenic potential. This assumption
Fig. 2. A–R: T140/TN14003 affects muscle migration and differentiation in the developing limb. T140/TN14003 caused a decrease of Pax3 (A–C),Lbx1 (G–I), and MyoD (M–O) signal in the operated side whereas such an effect was observed on Pax3 (D–F), Lbx1 (J–L), and MyoD (P–R) using PBScontrol beads.
Fig. 1. A–L: Effect of T140/TN14003 onCXCR4� cells in the developing limb bud ofchick embryo. Remarkably reduced transcriptsof CXCR4 in the limb where the T140/TN14003source was implanted (white arrow in A,B,E,F)as compared to the unoperated control side(black arrows in C,D,E) and operated PBS con-trol side (black arrowheads in G and H). Onvibratome section, the missing expression ofCXCR4 in the operated limb (J) is very evidentas compared to the control side (I). Nile Bluesulphate apoptosis test showed no noticeableapoptosis around the source of T140/TN14003.(white arrows in K–M). A thick antero-posteriorsection through the proximal part of the limbshowing the position of the bead (L). Notice thatthe physiological apoptotic zones (ANZ) de-scribed in the developing limb are visible (M).
3010 YUSUF ET AL.
was recently proven in a study(Vasyutina et al., 2005) using CXCR4-specific antibody, in which it wasshown that CXCR4-expressing cells inthe limb bud contribute to muscle andblood vessel formation.
Pax3 and Lbx1 are expressed by thehypaxial myogenic precursors in theventrolateral dermomyotomal lip aswell as during their course of migra-tion into the limb bud mesenchyme. Incontrast, CXCR4 is not expressed inthe dermomyotome, and the earliestexpression of CXCR4 in the limb wasnoticed at stages when the migratorypopulation detaching from the dermo-myotomal lip has already entered thelimb mesenchyme (Yusuf et al., 2005;Vasyutina et al., 2005). Using doubleantibody labelling, it was shown thata subset of Pax3� cells that migrateinto the limb in mouse embryos, alsoco-expresses the chemokine receptorCXCR4 (Vasyutina et al., 2005). Thissubpopulation of CXCR4�/Pax3�
cells were located very close to the ec-toderm in the premuscle masses. Asnot all Pax3 and Lbx1-expressing cellsare CXCR4�, this explains the weakereffect of T140/TN14003 on Pax3- andLbx1-expressing cells. Double anti-body labelling of Lbx1 and Pax3showed that these two progenitormarkers label the same cell popula-tions in the limb bud (Vasyutina et al.,
Fig. 3
Fig. 4.
Fig. 3. A–L: T140/TN14003 enhances commit-ment to angiogenic fate. Agglomeration of QH-1-positive cells around the T140/TN14003 beadwas observed in the limb (white arrow in A–C).No such agglomeration of QH-1-positive cellswas seen around the control PBS bead (whitearrowheads in D–F). Vascularity around the im-planted beads was visualized using ink injec-tions. T140/TN14003 caused an increased vas-cularity around the bead at HH23 (white arrrowin G) and at stage HH27 (white arrow in I) ascompared to the control side at stage HH22(white arrowhead in H) and HH27 (white arrow-head in J).
Fig. 4. A–D: The QH-1� endothelial cells aroundthe T140/TN14003 source have a somitic origin.To trace the origin of these cells, electropora-tion of lateral somitic halves and lateral dermo-myotomal (DM) lips was performed with EGFPvector (pCMS-EGFP) (white arrowhead in B).Clusters of EGFP-positive cells could be clearlyobserved around T140/TN14003 beads in limb(white arrow in A,B). No such agglomerationwas observed in EGFP-positive cells seenaround the control PBS beads (white arrows inC,D).
FATE OF CXCR4� PROGENITORS IN CHICK EMBRYOS 3011

2005). As T140/TN14003 have beendescribed to be highly specific CXCR4inhibitors (Tamamura et al., 2003a,b)that act by blocking SDF-1-mediatedresponses (Juazez et al., 2002) we no-ticed that it only seems to affect theCXCR4� cells, which are also proba-bly Pax3� and Lbx1�. Therefore, thePax3�/Lbx1�/CXCR4� cells can mi-grate into the limb bud and are notaffected by the T140/TN14003. Forthis reason, we observe only a de-crease in the muscle mass on the sidewhere the T140/TN140033 was im-planted and never an absence. Thisimplies that the CXCR4�-expressingcells in the limb bud, although havingthe potential to form blood vessels andmuscle, are only a subset of the bipo-tent Pax3� cells in the limb bud (Kar-don et al., 2002).
Studies of T140 and its analogTN14003 described in the literaturepoint towards a perturbation of theCXCR4/SDF-1 axis. Burger and colle-gues have shown that T140 andTN14003 can affect the migratoryabilities of the CXCR4-expressingchronic lymphoblastic leukemia (CLL)cells by inhibiting actin polymeriza-tion and chemotaxis, and, further-more, that the CXCL12 or SDF-1-in-duced phosphorylation of p44/42mitogen-activated protein kinase(MAPK) and signal transducer and ac-tivator of transcription 3 (STAT3) wasalso abolished (Burger et al., 2005). Inanother study, it was shown thatT140, T134, and AMD3100 could ef-fectively inhibit binding of monoclonalantibody against CXCR4. T140 andAMD3100 also blocked the chemo-taxis of acute lymphoblastic leukemia(ALL) cells to SDF-1 and attenuatedthe migration of ALL cells into bonemarrow stromal layers (Juarez et al.,2003). In another report, Juarez andcolleagues reported that T140 and itsanalogs inhibited the proliferation ofthe pre-B ALL cell line NALM6 re-markably. Proliferation of ALL cellscultured on bone marrow stromal lay-ers was also reduced in the presence ofT140 (Juarez et al., 2002).
In our experiments, we observedthat the migration of the CXCR4�
cells in the limb mesenchyme was hin-dered due to the application of T140/TN14003. Distal migration of theseprogenitors was blocked resulting infewer cells reaching the premuscle
masses and we could put this into ev-idence using probes for CXCR4, Pax3,and Lbx-1. As fewer migratory cellssucceed to migrate distally into thelimb, the muscle development and dif-ferentiation program is severely ham-pered as seen by a decrease of MyoDsignal.
SDF-1 has been shown to promotesurvival of CXCR4-expressing cells,and hematopoietic stem and progeni-tor cells (Joo et al., 2004). SyntheticSDF-1/CXCL12 also exhibited an an-tiapoptotic effect on chronic lympho-blastic leukemia cells in in vitro stud-ies reported by Meike Burger andcollegues (Burger et al., 2005). InCXCR4 mutants, significant apoptosiswas observed in the sensory neuronsof the dorsal root ganglia (DRG),which need an activated CXCR4 tosurvive the physiological apoptosis oc-curring during development. It is,however, of interest to note that thedecrease in the size of the dorsal rootganglia was not uniform along the en-tire anterio-posterior axis of the mu-tants and CXCR4 inactivation did notresult in a major loss of thoracic DRGneurons, although these cells also ex-press both CXCR4 and CXCL12 (Ode-mis et al., 2005). Odemis and col-leagues suggest that there may existcompensatory mechanisms that stillneed to be elucidated. This impliesthat there exist intrinsic differencesbetween the actions of CXCR4/SDF1signaling in the embryo. In our exper-iments, we did not observe any apo-ptotic activity occurring near the ec-topically implanted source of T140/TN14003 in the developing chick limb.It may be that in addition to SDF-1signaling in the limb, other factorslike Scatter Factor/Hepatocyte GrowthFactor (SF/HGF) may also play a role inthe survival and proliferation of thesomite-derived migratory progenitors.
The CXCR4� undetermined migra-tory progenitors that failed to reachtheir destined sites of differentiationin the premuscle anlage were forcedinto an angiogenic fate at the site toT140/TN14003 contact. This strength-ened the notion that the cells del-aminating from the ventrolateraldermomyotome at limb levels areheterogeneous in terms of their com-mitment to form muscle and endothe-lia, the majority being uncommitted(Kardon et al., 2002). The Pax3� cells
in the limb are bipotent, possessingthe ability to either develop into endo-thelia or myoblasts depending on thelocal signal they encounter in the limbenvironment. As all the CXCR4� cellsin the limb are Pax3� and Lbx1�, it isprobable that the CXCR4� cells rep-resent the bipotent Pax3� cell in thelimb whereas the Pax3�/CXCR4�
cells represent the committed myo-genic precursors. Therefore, when wetarget the CXCR4� in the limb, theuncommitted progenitors of the myo-genic and angiogenic lineages are pre-vented from migrating further intothe limb environment where theywould normally differentiate intomyoblasts under the influence of localsignals. The nature of these local sig-nals has not yet been determined.However, recent work in this directionhas pointed to the role of ectodermalWnt6 in myogenic determination ofthe progenitor populations migratingfrom the dermomyotome into the de-veloping limb (Geetha-Loganathan etal., 2005). Geetha-Loganathan andcolleagues showed that in the absenceof ectodermal Wnt6 signaling, migra-tory progenitors in the limb bud fail todifferentiate into muscle and insteadtake up an endothelial fate. Wntshave been also implicated in the mod-ulation of terminal differentiation ofmyogenic cells and the slow and intri-cate slow/fast limb muscle patterning(Anakwe et al., 2003). Another candi-date for the local signals is Tcf4, adownstream element of the canonicalWnt signaling pathway, which is ex-pressed in the premuscle masses limbmesenchyme (Kardon et al., 2003).Work in our lab has shown thatchanges in the patterning signals inthe limb like BMPs can bring aboutchanges in the expression pattern ofTcf4, which is also reflected by changesin muscle patterning (Bonafede et al.,2006).
Kardon and collegues postulate thatthere may be signals in an evenshorter range that determine myo-genic and endothelial fate as myoblastand endothelia differentiate in closeproximity to each other in the limbmesenchyme (Kardon et al., 2002).
Once these bipotent migratory pro-genitors are prevented from reachingthe premuscle anlage where theywould encounter the local signals thataid in their differentiation program,
3012 YUSUF ET AL.

they are forced into a default angio-genic fate as evidenced by the recruit-ment of somite derived QH-1� cellsand increased vascularity around theT140/TN14003 bead.
The effect of the CXCR4 inhibitorthat has been observed is possibly dueto the lack of the ability of theCXCR4� cells in the limb to bind withSDF-1 in the presence of T140/TN14003. A chemotactic effect ofSDF-1 has been described for primor-dial germ cell migration in fish, birds,and mammals where an SDF-1 gradi-ent is established along determinedroutes of migration. Furthermore, theSDF-1/CXCR4 interaction is also nec-essary for promotion of survival ofthese progenitors (Doitsidou et al.,2002; Knaut et al., 2003; Molyneauxet al., 2003, Stebler et al., 2004). How-ever, it still needs to be determined ifthe SDF-1 in the limb mesenchymeacts as a chemotactic signal for themigrating myogenic and endothelialcells into the limb bud. Vasyutina andcoworkers have shown that the ectopicsource of SDF-1 can affect the normalmigratory pattern of myogenic precur-sors. Furthermore, the differentiationof the CXCR4� cells in the limb wasdelayed (Vasyutina et al., 2005). Inour experiments, the highly specificpeptidic inhibitors of CXCR4, T140,and TN14003, could affect the normalmigratory behaviour of CXCR4� cellsas does the ectopic source of SDF-1,but this was associated with an al-tered differentiation program. Themuscle mass in the developing limbbud was reduced on the side whereT140/TN14003 was implanted as evi-denced by a decrease of MyoD signalalong with an increase of vasculardensity in the vicinity of the T140/TN14003 source. This shows thatCXCR4/SDF-1 interaction may be nec-essary for proper myogenesis in theCXCR4� population in the limb mes-enchyme. Recent work has pointed to-wards the role of CXCR4/SDF-1 sig-naling in secondary myogenesis(Odemis et al., 2005).
We suppose that the SDF-1 andCXCR4 interaction may be requiredfor the realization of the myogenicprogram in the CXCR4� cell in thelimb bud or, alternatively, theCXCR4/SDF-1 axis maintains the un-determined state of the bipotent (myo-genic and angiogenic) progenitors un-
til they have reached their final sitesto encounter local cues for differentia-tion. The CXCR4/SDF-1 axis has beenimplicated in the preservation of aprogenitor or stem cell status (Mur-doch, 2000). Therefore, by blockingthe CXCR4 and SDF-1 interaction us-ing highly specific CXCR4 antagonistslike T140 and its analog TN14003, theSDF-1 driven responses are blunted.
Moreover, Ratajczak and colleagueshave shown that murine cell lines de-rived from muscle satellite cells ex-press functional CXCR4 on their sur-faces and respond to the SDF-1secreted by muscle tissue (Ratajczaket al., 2003). SDF-1 acts as chemoat-tractant for the CXCR4-expressingmyoblasts and hematopoietic progeni-tors and keeps them in an undifferen-tiated state. We hypothesize that theCXCR4� cells in the limb are bipotentprogenitors that use SDF-1 as a che-moattractant signal for migration andlater use the SDF-1 niches to preservetheir progenitor status upon receivingappropriate signals at their respectivesites of differentiation, and then areable to differentiate into myogenicand angiogenic lineages. Blocking thisCXCR4/SDF-1 axis by the use of spe-cific inhibitors of CXCR4 like T140/TN14003 perturbs not just the migra-tion but also the differentiationprogram of these progenitors in thelimb bud.
As a result of blocking CXCR4/SDFsignaling, CXCR4-expressing cells areunable to migrate along their appro-priate routes. In the absence of localsignals that support myogenesis,these bipotent uncommitted precur-sors adopt a default angiogenic fate.This suggests that SDF-1 signalingmay be of importance not just for themigration of the CXCR4� cells butalso for the preservation of their un-differentiated progenitor status. Fur-thermore, it is of clinical relevancewhether such a mechanism also playsa role in the treatment of tumors withT140/TN14003.
EXPERIMENTALPROCEDURES
Chicken Embryos
Fertilized chicken and quail eggs ob-tained from a local breeder were incu-bated at 38°C and 80% humidity. The
embryos were staged using the Ham-burger Hamilton staging system. Fol-lowing operative procedure, the eggswere reincubated at 38°C and 80% hu-midity to develop further to the re-quired stages. Staging was done ac-cording to Hamburger and Hamilton(1951).
Operations
T140 and TN14003 beads were pre-pared by soaking the acrylic beads oraffigel beads overnight or 6 hr in T140or TN14003 at concentrations rangingfrom 5–10 mg/ml. The eggs were win-dowed and after removal of the extra-embryonic membranes, the beadswere inserted into the developing limbbud of stage HH17–19 chick and quailembryos using a tungsten needle. PBSbeads were inserted as controls. Theembryos were then reincubated toreach stage HH22–27. Some werefixed with 4% PFA and further sub-mitted for in situ hybridisation andimmunohistochemistry. A few em-bryos were injected with Indian inkinto the viteline vessels to visualizethe vascular pattern of the limbs.These ink-injected embryos were thenfixed overnight in 100% ethanol. Thefollowing day, these embryos weretransferred to methyl salicylate for24 hr.
Apoptosis Test
An apoptosis test was performed afterthe implantation of T140/TN14003beads at 8, 12, and 24 hr of reincuba-tion, using Nile Blue sulphate (vanden Eijnde et al.,1997). After removalof the extra embryonic membranes,the embryos were stained in toto for30 min at 37°C in a Nile Blue sulfatesolution (1:20,000 w/v). After incuba-tion, the specimens were washed withPBS and put on ice and immediatelyphotographed.
In Situ Hybridization
Selected embryos were tested forCXCR4, Pax3, Lbx1, and MyoD usinghighly specific RNA probes. TheCXCR4 probe used was a 1,100-bpchicken probe. The Pax3 probe usedwas a 950-bp quail Pax3 probe. TheMyoD probe was a 1,518-bp specificchicken probe and an 826-bp Lbx1
FATE OF CXCR4� PROGENITORS IN CHICK EMBRYOS 3013

probe. Riboprobes were labeled with adigoxigenin RNA labeling kit fromBoehringer, Mannheim, Germany.Whole mount in situ hybridizationwas performed as described previ-ously (Nieto et al. 1996).
In Ovo Electroporation andFate Mapping
Fertilized eggs of Gallus gallus (WhiteLeghorn) obtained from a localbreeder were incubated at 38°C and80% humidity until the requiredstages of HH15–17 were reached. Af-ter lowering the embryos by extract-ing albumin from the side, the eggswere windowed and Indian ink wasinjected under the embryo to make iteasily visible. The extra membranewas partially removed in the region tobe electroporated. The pCMS-EGFPconstructs (2–3 �g/�l) were dissolvedin an adjunct solution (50 �l stock so-lution: 25 �l carboxymethlycellulose,7.5 �l fast green, 3.0 �l MgCl2, 7.5 �l20� PBS, and 7.0 �l H2O) in a ratio of2:1 as previously reported by us (Daiet al 2005). The DNA constructs weremicroinjected into the somites. Theelectrodes were placed at each side ofthe microinjected embryo, and fivesquare pulses of 45–60 V, 20-msecwidth were applied for each embryo.After 24- to 36-hr re-incubation, theEGFP expression in the electropo-rated embryos was visualized underfluorescence microscopy.
Sectioning andImmunohistochemistry
Selected quail embryos with im-planted T140/TN14003 and PBSbeads were cryosectioned after embed-ding them in tissue freezing medium(Leica Instruments, Nussloch). Twen-ty-micrometer-thick sections werestained using quail-endothelial cell-specific monoclonal antibodies (QH-1).Antigen detection was done using acy3 fluorescent secondary antibodydetection kit.
To enhance the green flourescencesignal in the electroporated embryos,these embryos were cryosectioned asdescribed and then submitted for im-munohistochemistry using anti-GFPantibodies (Torrey Pines Biolabs) fol-lowed by cy2 fluorescent antibodies forantigen detection.
Microscopy and Imaging
Hybridized samples were observedand photographed using the LeicaMZFLIII microscope and LeicaDC300F digital camera. The sectionswere observed and photographed us-ing an Axioscope 20 (Zeiss) and aLeica DFC320 digital camera.
ACKNOWLEDGMENTSWe thank Hirokazu Tamamura andMeike Burger for providing theCXCR4 inhibitors, T140 andTN14003, and Tanja Hartmann forhelpful discussions. All authors wouldlike to extend their thanks to EllenGimbel, Susanna Konradi, and UlrikePein for their excellent technical as-sistances. We acknowledge the sup-port of “Molecular Mechanisms for Mi-gration, Invasion and Metastasis,” aBaden-Wurttemberg grant, and theMyores Project (511978) funded by theEC’s sixth framework Programme.
REFERENCES
Anakwe K, Robson L, Hadley J, Buxton P,Church V, Allen S, Hartmann C, HarfeB, Nohno T, Brown AM, Evans DJ, Fran-cis-West P. 2003. Wnt signaling regu-lates myogenic differentiation in the de-veloping avian wing. Development 130:3503–3514.
Birchmeier C, Brohmann H. 2000. Genesthat control the development of migrat-ing muscle precursor cells. Curr OpinCell Biol 12:725–730.
Bladt F, Riethmacher D, Isenmann S,Aguzzi A, Birchmeier C. 1995. Essentialrole for the c-met receptor in the migra-tion of myogenic precursor cells into thelimb bud. Nature 376:768–771.
Bober E, Franz T, Arnold HH, Gruss P,Tremblay P. 1994. Pax-3 is required forthe development of limb muscles: a pos-sible role for the migration of dermomyo-tomal muscle progenitor cells. Develop-ment 120:603–612.
Bonafede A, Kohler T, Rodriguez-Nieden-fuhr M, Brand-Saberi B. 2006. BMPs re-strict the position of premuscle massesin the limb buds by influencing Tcf4 ex-pression. Dev Biol (in press).
Brand-Saberi B, Christ B. 2000. Evolutionand development of distinct cell lineagesderived from somites. Curr Top Dev Biol48:1–42.
Brand-Saberi B, Wilting J, Ebensperger C,Christ B. 1996. The formation of somitecompartments in the avian embryo. Int JDev Biol 40:411–420.
Burger M, Hartmann T, Krome M, RawlukJ, Tamamura H, Fujii N, Kipps TJ,Burger JA. 2005. Small peptide inhibi-tors of the CXCR4 chemokine receptor(CD184) antagonize the activation, mi-
gration, and antiapoptotic responses ofCXCL12 in chronic lymphocytic leuke-mia B cells. Blood 106:1824–1830.
Christ B, Jacob HJ, Jacob M. 1977. Exper-imental analysis of the origin of the wingmusculature in avian embryos. AnatEmbryol (Berl) 150:171-186.
Dai F, Yusuf F, Farjah GH, Brand-SaberiB. 2005. RNAi-induced targeted silenc-ing of developmental control genes dur-ing chicken embryogenesis. Dev Biol285:80–90.
Deng H, Liu R, Ellmeier W, Choe S, Unut-maz D, Burkhart M, Di Marzio P, Mar-mon S, Sutton RE, Hill CM, Davis CB,Peiper SC, Schall TJ, Littman DR,Landau NR. 1996. Identification of a ma-jor co-receptor for primary isolates ofHIV-1. Nature 381:661–666
Doitsidou M, Reichman-Fried M, Stebler J,Koprunner M, Dorries J, Meyer D, Es-guerra CV, Leung T, Raz E. 2002. Guid-ance of primordial germ cell migrationby the chemokine SDF-1. Cell 111:647–659.
Doranz BJ, Rucker J, Yi Y, Smyth RJ,Samson M, Peiper SC, Parmentier M,Collman RG, Doms RW. 1996. A dual-tropic primary HIV-1 isolate that usesfusin and the beta-chemokine receptorsCKR-5, CKR-3, and CKR-2b as fusioncofactors. Cell 85:1149–1158
Feng Y, Broder CC, Kennedy PE, BergerEA. 1996. HIV-1 entry cofactor: func-tional cDNA cloning of a seven-trans-membrane, G protein-coupled receptor.Science 272:872–877.
Franz T, Kothary R, Surani MA, Halata Z,Grim M. 1993. The Splotch mutationinterferes with muscle development inthe limbs. Anat Embryol (Berl). 187:153–160.
Geetha-Loganathan P, Nimmagadda S,Huang R, Scaal M, Christ B. 2005. Roleof Wnt-6 in limb myogenesis. Anat Em-bryol (Berl) 21:1–6.
Gross MK, Moran-Rivard L, Velasquez T,Nakatsu MN, Jagla K, Goulding M.2000. Lbx1 is required for muscle precur-sor migration along a lateral pathwayinto the limb. Development 127:413–424.
Hamburger V, Hamilton HL. 1951. A se-ries of normal stages in the developmentof the chick embryo. J Morphol 88:49–92.
Hartmann TN, Burger JA, Glodek A, FujiiN, Burger M. 2005. CXCR4 chemokinereceptor and integrin signaling co-oper-ate in mediating adhesion and chemore-sistance in small cell lung cancer (SCLC)cells. Oncogene 4462–4471.
Joo EK, Broxmeyer HE, Kwon HJ, KangHB, Kim JS, Lim JS, Choe YK, Choe IS,Myung PK, Lee Y. 2004 Enhancement ofcell survival by stromal cell-derived fac-tor-1/CXCL12 involves activation ofCREB and induction of Mcl-1 and c-Fosin factor-dependent human cell lineMO7e. Stem Cells Dev 13:563–570.
Juarez J, Bradstock KF, Gottlieb DJ, Ben-dall LJ. 2002. Antagonists of the Chemo-kine Receptor CXCR4 Block Chemotaxisand Inhibit Stromal Dependent Prolifer-ation of Acute Lymphoblastic Leukemia
3014 YUSUF ET AL.

Cells. American Society of HematologyMeeting. December, Philadelphia, PA.
Juarez J, Bradstock KF, Gottlieb DJ, Ben-dall LJ. 2003. Effects of inhibitors of thechemokine receptor CXCR4 on acutelymphoblastic leukemia cells in vitro.Leukemia 17:1294–1300.
Kardon G, Campbell JK, Tabin CJ. 2002Local extrinsic signals determine muscleand endothelial cell fate and patterningin the vertebrate limb. Dev Cell 3:533–545.
Kardon G, Harfe BD, Tabin CJ. 2003. ATcf4-positive mesodermal populationprovides a prepattern for vertebrate limbmuscle patterning. Dev Cell 5:937–944.
Knaut H, Werz C, Geisler R, Nusslein-Vol-hard C. Tubingen 2000 Screen Consor-tium. 2003. A zebrafish homologue of thechemokine receptor Cxcr4 is a germ-cellguidance receptor. Nature 421:279–282.
Molyneaux KA, Zinszner H, Kunwar PS,Schaible K, Stebler J, Sunshine MJ,O’Brien W, Raz E, Littman D, Wylie C,Lehmann R. 2003. The chemokine SDF1/CXCL12 and its receptor CXCR4 regu-late mouse germ cell migration and sur-vival. Development 130:4279–4286.
Murdoch C. 2000. CXCR4: chemokine re-ceptor extraordinaire. Immunol Rev 177:175–184.
Nieto MA, Patel K, Wilkinson DG. 1996. Insitu hybridization analysis of chick em-bryos in whole mount and tissue sec-tions. Methods Cell Biol 51:219–235.
Odemis V, Lamp E, Pezeshki G, Moepps B,Schilling K, Gierschik P, Littman DR,Engele J. 2005. Mice deficient in the che-mokine receptor CXCR4 exhibit im-paired limb innervation and myogenesis.Mol Cell Neurosci 30:494–505.
Ordahl CP, Le Douarin NM. 1992. Two myo-genic lineages within the developingsomite. 1992. Development 114:339–353.
Princen K, Schols D. 2005. HIV chemokinereceptor inhibitors as novel anti-HIVdrugs. Cytokine Growth Factor Rev 16:659–677.
Ratajczak MZ, Majka M, Kucia M, DrukalaJ, Pietrzkowski Z, Peiper S, Janowska-Wieczorek A. 2003. Expression of func-tional CXCR4 by muscle satellite cellsand secretion of SDF-1 by muscle-de-rived fibroblasts is associated with thepresence of both muscle progenitors inbone marrow and hematopoietic stem/progenitor cells in muscles. Stem Cells21:363–371.
Schafer K, Braun T. 1999. Early specifica-tion of limb muscle precursor cells by thehomeoboxgeneLbx1h.NatGenet23:213–216.
Stebler J, Spieler D, Slanchev K,Molyneaux KA, Richter U, Cojocaru V,Tarabykin V, Wylie C, Kessel M, Raz E.2004. Primordial germ cell migration inthe chick and mouse embryo: the role ofthe chemokine SDF-1/CXCL12. Dev Biol272:351–361.
Tajbakhsh S, Rocancourt D, Cossu G,Buckingham M. 1997. Redefining the ge-netic hierarchies controlling skeletalmyogenesis: Pax-3 and Myf-5 act up-stream of MyoD. Cell 89:127–138.
Tamamura H, Fujii N. 2004. Two orthogo-nal approaches to overcome multi-drugresistant HIV-1s: development of pro-tease inhibitors and entry inhibitorsbased on CXCR4 antagonists. Curr DrugTargets Infect Disord 4:103–110.
Tamamura H, Fujii N. 2005. The therapeu-tic potential of CXCR4 antagonists in thetreatment of HIV infection, cancer me-tastasis and rheumatoid arthritis. Ex-pert Opin Ther Targets 9:1267–1282.
Tamamura H, Hori A, Kanzaki N, Hira-matsu K, Mizumoto M, Nakashima H,Yamamoto N, Otaka A, Fujii N. 2003a.
T140 analogs as CXCR4 antagonistsidentified as anti-metastatic agents inthe treatment of breast cancer. FEBSLett 550:79–83.
Tamamura H, Hiramatsu K, Kusano S,Terakubo S, Yamamoto N, Trent JO,Wang Z, Peiper SC, Nakashima H,Otaka A, Fujii N. 2003b. Synthesis ofpotent CXCR4 inhibitors possessing lowcytotoxicity and improved biostabilitybased on T140 derivatives. Org BiomolChem 1:3656–3662.
Tamamura H, Fujisawa M, Hiramatsu K,Mizumoto M, Nakashima H, YamamotoN, Otaka A, Fujii N. 2004. Identificationof a CXCR4 antagonist, a T140 analog,as an anti-rheumatoid arthritis agent.FEBS Lett 569:99–104.
Tamamura H, Otaka A, Fujii N. 2005. De-velopment of anti-HIV agents targetingdynamic supramolecular mechanism:entry and fusion inhibitors based onCXCR4/CCR5 antagonists and gp41–C34-remodeling peptides. Curr HIV Res3:289–301.
van den Eijnde SM, Luijsterburg AJ,Boshart L, De Zeeuw CI, van Dieren-donck JH, Reutelingsperger CP, Ver-meij-Keers C. 1997. In situ detection ofapoptosis during embryogenesis with an-nexin V: from whole mount to ultrastruc-ture. Cytometry 29:313–320.
Vasyutina E, Stebler J, Brand-Saberi B,Schulz S, Raz E, Birchmeier C. 2005.CXCR4 and Gab1 cooperate to control thedevelopment of migrating muscle progen-itor cells. Genes Dev 19:2187–2198.
Yusuf F, Rehimi R, Dai F, Brand-Saberi B.2005. Expression of chemokine receptorCXCR4 during chick embryo develop-ment. Anat Embryol (Berl) 210:35–41.
Zuzarte-Luis V, Hurle JM. 2002. Pro-grammed cell death in the developinglimb. Int J Dev Biol 46:871–876.
FATE OF CXCR4� PROGENITORS IN CHICK EMBRYOS 3015
Copyright © 2022 FDOKUMEN




















