
A novel role of CXCR4 and SDF-1 during migration of cloacal muscle precursors
10
RESEARCH ARTICLE A Novel Role of CXCR4 and SDF-1 During Migration of Cloacal Muscle Precursors Rizwan Rehimi, 1,2 Nargis Khalida, 1,3 Faisal Yusuf, 1,3 Gabriela Morosan-Puopolo, 1,2 and Beate Brand-Saberi 1,3 * The cloaca acts as a common chamber into which gastrointestinal and urogenital tracts converge in lower vertebrates. The distal end of the cloaca is guarded by a ring of cloacal muscles or sphincters, the equiva- lent of perineal muscles in mammals. It has recently been shown that the development of the cloacal mus- culature depends on hindlimb muscle formation. The signaling molecules responsible for the outward migration of hindlimb myogenic precursors are not known. Based on the expression studies for CXCR4 and SDF-1, we hypothesized a role of this signaling pair during cloacal muscle precursor migration. The aim of our study was to investigate the role of SDF-1/CXCR4 during cloacal muscle precursor migration in the chicken embryos. We show that SDF-1 is expressed in the cloacal region, and by experimentally manipulating the SDF-1/CXCR4 signaling, we can show that SDF-1 guides the migration of CXCR4- expressing cloacal muscle precursors. Developmental Dynamics 239:1622–1631, 2010. V C 2010 Wiley-Liss, Inc. Key words: chicken embryo; SDF-1; CXCR4; cell migration; cloacal muscle Accepted 2 March 2010 INTRODUCTION During early embryonic development of mammals, the cloaca acts as a col- lecting chamber into which the intes- tinal, urinary, and genital tracts con- verge (Martinez-Frias et al., 2000; McBride et al., 2003; Dravis et al., 2004; Valasek et al., 2005). Later in development, the cloaca is partitioned into separate urogenital and alimen- tary tracts (Kimmel et al., 2000; de Santa Barbara et al., 2002; Penington and Hutson, 2003; Hynes and Fraher, 2004). In contrast to mammals, in adult birds, amphibians, reptiles, monotreme mammals, and chon- drichthyan fish, the cloaca persists throughout their lives as a single opening for urinary, digestive, and genital ducts (King and McLelland, 1981). In mammals, the equivalent of cloacal muscle are the perineal sphincters guarding the lower uro- genital and gastrointestinal tract. The origin of cloacal muscles has only recently been shown to lie in the hypaxial muscle precursors in the somites which are known to populate the limbs and the ventral body wall (Valasek et al., 2005). Following hypaxial myogenic pre- cursors epithelio-mesenchymal tran- sition from the ventrolateral dermo- myotome, they either migrate individually into the limb mesen- chyme or form cell sheets, myotomal extensions, that eventually give rise to ventral body wall muscles (Galis, 2001). The single individually migrat- ing myogenic progenitor cells gener- ate hypaxial muscles of the extrem- ities, tongue and diaphragm (Cheval- lier et al., 1977; Christ et al., 1977; Christ and Ordahl, 1995). The forma- tion of the hypaxial compartment in the dermomyotome, the delamination, the migration, and the eventual dif- ferentiation of these myogenic precur- sors are tightly regulated by several genes. Pax3 and Lbx1 are required for the specification of these myogenic precursors in the dermomyotome, cMet and scatter factor (SF/HGF) are involved in the delamination process, Lbx-1 enables the navigation of the myogenic precursors along migratory routes, while MyoD and Myogenin play a role in the myogenic differen- tiation program (Bober et al., 1994; Developmental Dynamics 1 Institute of Anatomy and Cell Biology, Department of Molecular Embryology, University of Freiburg, Freiburg, Germany 2 Department Faculty of Biology, University of Freiburg, Freiburg, Germany 3 Institute of Anatomy, Department of Anatomy and Molecular Embryology, Ruhr University, Bochum, Germany Grant sponsor: EU’s Sixth Framework Programme; Grant number: 511978; Grant sponsor: GKR 1104. *Correspondence to: Beate Brand-Saberi, Institute of Anatomy, Department of Anatomy and Molecular Embryology, Ruhr University Bochum, 44780 Bochum, Germany. E-mail: [email protected] DOI 10.1002/dvdy.22288 Published online 9 April 2010 in Wiley InterScience (www.interscience.wiley.com). DEVELOPMENTAL DYNAMICS 239:1622–1631, 2010 V C 2010 Wiley-Liss, Inc.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of A novel role of CXCR4 and SDF-1 during migration of cloacal muscle precursors

a RESEARCH ARTICLE
A Novel Role of CXCR4 and SDF-1 DuringMigration of Cloacal Muscle PrecursorsRizwan Rehimi,1,2 Nargis Khalida,1,3 Faisal Yusuf,1,3 Gabriela Morosan-Puopolo,1,2
and Beate Brand-Saberi1,3*
The cloaca acts as a common chamber into which gastrointestinal and urogenital tracts converge in lowervertebrates. The distal end of the cloaca is guarded by a ring of cloacal muscles or sphincters, the equiva-lent of perineal muscles in mammals. It has recently been shown that the development of the cloacal mus-culature depends on hindlimb muscle formation. The signaling molecules responsible for the outwardmigration of hindlimb myogenic precursors are not known. Based on the expression studies for CXCR4and SDF-1, we hypothesized a role of this signaling pair during cloacal muscle precursor migration. Theaim of our study was to investigate the role of SDF-1/CXCR4 during cloacal muscle precursor migration inthe chicken embryos. We show that SDF-1 is expressed in the cloacal region, and by experimentallymanipulating the SDF-1/CXCR4 signaling, we can show that SDF-1 guides the migration of CXCR4-expressing cloacal muscle precursors. Developmental Dynamics 239:1622–1631, 2010. VC 2010 Wiley-Liss, Inc.
Key words: chicken embryo; SDF-1; CXCR4; cell migration; cloacal muscle
Accepted 2 March 2010
INTRODUCTION
During early embryonic developmentof mammals, the cloaca acts as a col-lecting chamber into which the intes-tinal, urinary, and genital tracts con-verge (Martinez-Frias et al., 2000;McBride et al., 2003; Dravis et al.,2004; Valasek et al., 2005). Later indevelopment, the cloaca is partitionedinto separate urogenital and alimen-tary tracts (Kimmel et al., 2000; deSanta Barbara et al., 2002; Peningtonand Hutson, 2003; Hynes and Fraher,2004). In contrast to mammals, inadult birds, amphibians, reptiles,monotreme mammals, and chon-drichthyan fish, the cloaca persiststhroughout their lives as a singleopening for urinary, digestive, and
genital ducts (King and McLelland,1981). In mammals, the equivalent ofcloacal muscle are the perinealsphincters guarding the lower uro-genital and gastrointestinal tract.The origin of cloacal muscles has onlyrecently been shown to lie in thehypaxial muscle precursors in thesomites which are known to populatethe limbs and the ventral body wall(Valasek et al., 2005).
Following hypaxial myogenic pre-cursors epithelio-mesenchymal tran-sition from the ventrolateral dermo-myotome, they either migrateindividually into the limb mesen-chyme or form cell sheets, myotomalextensions, that eventually give riseto ventral body wall muscles (Galis,2001). The single individually migrat-
ing myogenic progenitor cells gener-ate hypaxial muscles of the extrem-ities, tongue and diaphragm (Cheval-lier et al., 1977; Christ et al., 1977;Christ and Ordahl, 1995). The forma-tion of the hypaxial compartment inthe dermomyotome, the delamination,the migration, and the eventual dif-ferentiation of these myogenic precur-sors are tightly regulated by severalgenes. Pax3 and Lbx1 are required forthe specification of these myogenicprecursors in the dermomyotome,cMet and scatter factor (SF/HGF) areinvolved in the delamination process,Lbx-1 enables the navigation of themyogenic precursors along migratoryroutes, while MyoD and Myogeninplay a role in the myogenic differen-tiation program (Bober et al., 1994;
Dev
elop
men
tal D
ynam
ics
1Institute of Anatomy and Cell Biology, Department of Molecular Embryology, University of Freiburg, Freiburg, Germany2Department Faculty of Biology, University of Freiburg, Freiburg, Germany3Institute of Anatomy, Department of Anatomy and Molecular Embryology, Ruhr University, Bochum, GermanyGrant sponsor: EU’s Sixth Framework Programme; Grant number: 511978; Grant sponsor: GKR 1104.*Correspondence to: Beate Brand-Saberi, Institute of Anatomy, Department of Anatomy and Molecular Embryology, RuhrUniversity Bochum, 44780 Bochum, Germany. E-mail: [email protected]
DOI 10.1002/dvdy.22288Published online 9 April 2010 in Wiley InterScience (www.interscience.wiley.com).
DEVELOPMENTAL DYNAMICS 239:1622–1631, 2010
VC 2010 Wiley-Liss, Inc.

Tajbakhsh et al., 1997; Dietrich et al.,1999; Schafer and Braun, 1999; Broh-mann et al., 2000; Gross et al., 2000).The experimental evidence that cloa-cal muscles originate from thesomites that provide myogenic pre-cursors for the hindlimb has comefrom the work of Valasek and col-leagues. They report that the cloacalmuscles in the developing chickenembryo derive their origin from thesame somites that also give rise to theleg muscles (Valasek et al., 2005). Infact, cells that participate in cloacalmuscle formation initially migratefrom the somites into the developinghindlimb bud (Christ et al., 1974,1977; Chevallier et al., 1977; Jacobet al., 1979; Rees et al., 2003; Bona-fede et al., 2006) and then translocatefrom the ventral muscle mass in thehindlimb toward the cloacal tubercle(Valasek et al., 2005; Evans et al.,2006). These cloacal precursors ex-press Pax7 and MyoD as they extendout of the proximal limb toward thecloacal tubercle.
In placental animals, the hindgutinitially ends blindly in the cloacawhere the cloacal membrane sepa-rates it from proctodeum. Mesenchy-mal condensations inside the cloacalater form the urorectal septum divid-ing the cloaca into a ventral urogeni-tal sinus and a dorsal anorectal canal.Later during development, the uror-ectal septum thickens and forms thedeep perinium where the anal andurogenital sphincter muscles arelocated. Only after the rupture of thecloacal membrane a connection of theendodermal and ectodermal part ofthe anal canal is established. An epi-thelial plug ensures a closed analcanal till late into the fetal period.Defects in the formation of the urorec-tal septum and failure in the ruptureof the urorectal membrane can giverise to severe forms of anorectal mal-formations. Persistent cloaca is nor-mal in birds, reptiles, and some fish,but in human infants appears as acomplex anorectal and genitourinarymalformation whereby the rectum,vagina, and urethra are fused. A com-mon opening larger than 3cm in di-ameter at birth is associated with apoor prognosis as the sphincter func-tion is compromised and is not ame-nable to an effective surgical correc-tion (Keith et al., 2005). Although
work over the years using the mouseand chicken model has revealed theinvolvement of Shh and Wnt signal-ing in anorectal malformations (Moet al., 2001; Tai et al., 2009), the etiol-ogy and pathogenesis of anorectalmalformations remains poorly under-stood and controversial.
Research using avian and mamma-lian model systems are leading to aconsensus on the evolutionary historyof muscle formation, which enables usto use the avian model to depict theorigin and formation of distinctmuscles in mammals (Brand-Saberiand Christ, 2000; Valasek et al.,2005). The external urogenital andanorectal sphincter muscles in mam-mals are functionally homologues tothe cloacal sphincter muscles in birds.The underlying genetic and molecularmechanisms governing the origin andthe development of the early commoncloacal muscles are not understood.Moreover, the mechanism active dur-ing remigration of these myogenicprecursors from the ventral musclemass to the cloaca has not yet beendescribed. In our present work, weattempted to decipher probable molec-ular players that are active duringthese migratory events. Chemokinereceptor CXCR4 and its ligand SDF-1have been implicated in numerous de-velopmental processes and precursormigration events during embryonicdevelopment (Murdoch, 2000). Pri-mordial germ cells express function-ally active CXCR4 on their surfacesand respond to SDF-1 cues leadingthem toward the gonads (Stebler atal., 2004). CXCR4 and SDF-1 mutantmice show defective B-cell lymphopoi-esis (Nagasawa et al., 1996; Zou et al.,1998) and abnormal vasculogenesis,developmental defects that occur dueto defective cell migration. We andothers have previously shown thatthe chemokine SDF-1 and its receptorCXCR4 are required for the migrationof myogenic precursors in the limbbuds of developing chicken embryos(Vasyutina et al., 2005; Yusuf et al.,2006). We hypothesized a role of che-mokine SDF-1 and its receptorCXCR4 during cloacal myogenic pre-cursor migration. Our hypothesis wasbased on the observation of SDF-1expression in the cloacal region fromstage HH27 onward (Rehimi et al.,2008) and CXCR4 expression in mi-
gratory cells (David et al., 2002; Liet al., 2004; Yusuf et al., 2005; Vasyu-tina et al., 2005; Haas and Gilmour,2006). To test our hypothesis, we tar-getted the SDF-1/CXCR4 signallingat the level of the proximal hindlimband analysed effects on the migrationof cloacal muscle precursors and even-tual cloacal muscle formation. Weshow here that SDF-1 and CXCR4play an important role in the remigra-tion of myogenic precursor cells fromthe ventral part of the hindlimb intothe cloaca. Our experimental manipu-lation targeting the SDF-1/CXCR4signaling at the level of the cloacalmuscle precursors in the proximalhindlimb affected their ability tomigrate out of the hindlimb towardthe cloaca and eventually leads todefects in cloacal muscle formation.We provide evidence based on manip-ulation experiments in chickenembryos that CXCR4/SDF-1 signalingis active during cloacal muscle forma-tion and SDF-1 serves as a guidingcue to migratory cloacal muscle cellsthat express its receptor CXCR4 ontheir path from hindlimb toward themidline to form cloacal/perinealmuscles.
RESULTS
SDF-1, CXCR4, Pax7, and
MyoD Expression in the
Cloaca
We have previously defined theexpression pattern of SDF-1 in thedeveloping chicken embryo. Weobserved that SDF-1 has a wideexpression in different tissues duringdevelopment. At stage HH26–29,SDF-1 transcripts are expressed inthe cloacal cleft (Fig. 1A,B). Further-more, at stage HH29, SDF-1 isstrongly expressed in the proximalregion of the hindlimb (Fig. 1B). Atsimilar stages, CXCR4 transcripts arevisible around the developing cloacalregion (Fig. 1C), while a few CXCR4-positive cells were observed near thecloaca at stage HH29 (Fig. 1D). Aband of Pax7 expression extendingfrom the proximal side of hindlimb to-ward the cloaca is visible at stageHH26–HH29 (Fig. 1E,F). At similarstages, in MyoD expression domain isobservable in the proximal hindlimb
Dev
elop
men
tal D
ynam
ics
CXCR4/SDF-1 AXIS DURING CLOACAL MUSCLE FORMATION 1623
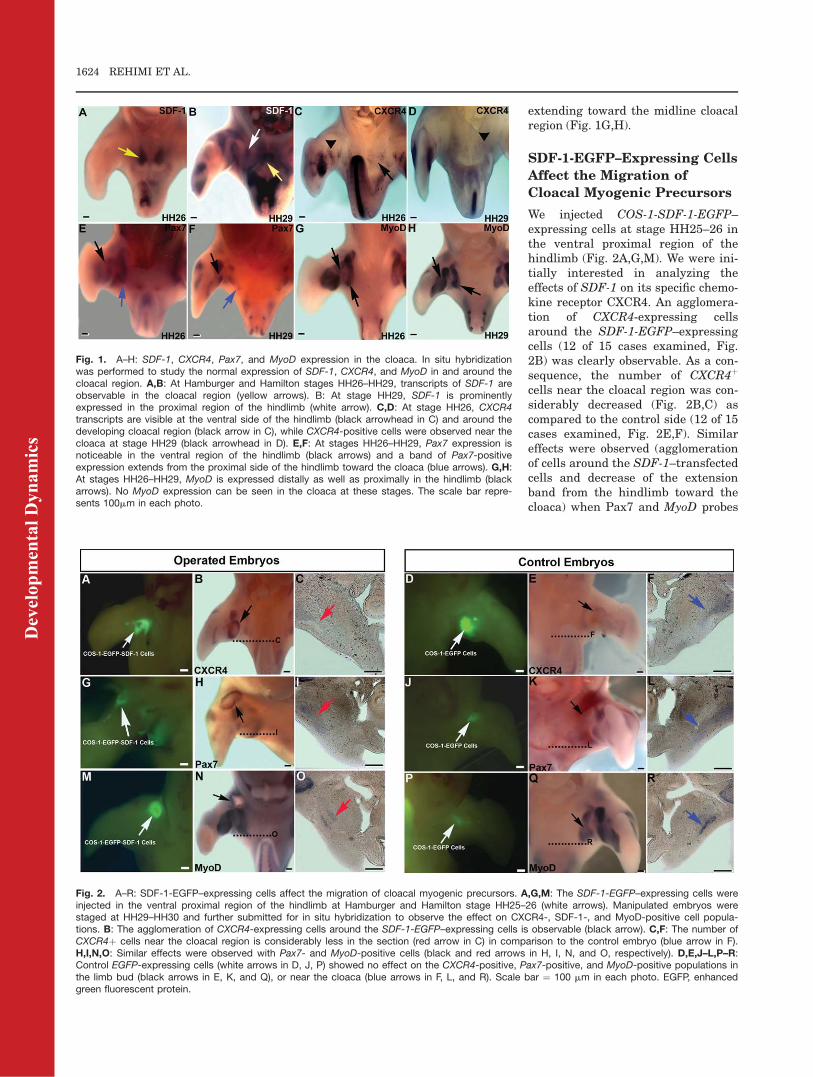
extending toward the midline cloacalregion (Fig. 1G,H).
SDF-1-EGFP–Expressing Cells
Affect the Migration of
Cloacal Myogenic Precursors
We injected COS-1-SDF-1-EGFP–expressing cells at stage HH25–26 inthe ventral proximal region of thehindlimb (Fig. 2A,G,M). We were ini-tially interested in analyzing theeffects of SDF-1 on its specific chemo-kine receptor CXCR4. An agglomera-tion of CXCR4-expressing cellsaround the SDF-1-EGFP–expressingcells (12 of 15 cases examined, Fig.2B) was clearly observable. As a con-sequence, the number of CXCR4þ
cells near the cloacal region was con-siderably decreased (Fig. 2B,C) ascompared to the control side (12 of 15cases examined, Fig. 2E,F). Similareffects were observed (agglomerationof cells around the SDF-1–transfectedcells and decrease of the extensionband from the hindlimb toward thecloaca) when Pax7 and MyoD probes
Fig. 1. A–H: SDF-1, CXCR4, Pax7, and MyoD expression in the cloaca. In situ hybridizationwas performed to study the normal expression of SDF-1, CXCR4, and MyoD in and around thecloacal region. A,B: At Hamburger and Hamilton stages HH26–HH29, transcripts of SDF-1 areobservable in the cloacal region (yellow arrows). B: At stage HH29, SDF-1 is prominentlyexpressed in the proximal region of the hindlimb (white arrow). C,D: At stage HH26, CXCR4transcripts are visible at the ventral side of the hindlimb (black arrowhead in C) and around thedeveloping cloacal region (black arrow in C), while CXCR4-positive cells were observed near thecloaca at stage HH29 (black arrowhead in D). E,F: At stages HH26–HH29, Pax7 expression isnoticeable in the ventral region of the hindlimb (black arrows) and a band of Pax7-positiveexpression extends from the proximal side of the hindlimb toward the cloaca (blue arrows). G,H:At stages HH26–HH29, MyoD is expressed distally as well as proximally in the hindlimb (blackarrows). No MyoD expression can be seen in the cloaca at these stages. The scale bar repre-sents 100mm in each photo.
Fig. 2. A–R: SDF-1-EGFP–expressing cells affect the migration of cloacal myogenic precursors. A,G,M: The SDF-1-EGFP–expressing cells wereinjected in the ventral proximal region of the hindlimb at Hamburger and Hamilton stage HH25–26 (white arrows). Manipulated embryos werestaged at HH29–HH30 and further submitted for in situ hybridization to observe the effect on CXCR4-, SDF-1-, and MyoD-positive cell popula-tions. B: The agglomeration of CXCR4-expressing cells around the SDF-1-EGFP–expressing cells is observable (black arrow). C,F: The number ofCXCR4þ cells near the cloacal region is considerably less in the section (red arrow in C) in comparison to the control embryo (blue arrow in F).H,I,N,O: Similar effects were observed with Pax7- and MyoD-positive cells (black and red arrows in H, I, N, and O, respectively). D,E,J–L,P–R:Control EGFP-expressing cells (white arrows in D, J, P) showed no effect on the CXCR4-positive, Pax7-positive, and MyoD-positive populations inthe limb bud (black arrows in E, K, and Q), or near the cloaca (blue arrows in F, L, and R). Scale bar ¼ 100 mm in each photo. EGFP, enhancedgreen fluorescent protein.
Dev
elop
men
tal D
ynam
ics
1624 REHIMI ET AL.

were tested (9 of 10 cases examined,Fig. 2H,I,N,O). Implantation of con-trol EGFP-expressing cells showed noeffect on the CXCR4, Pax7, and MyoDpopulation in the limb bud and nearthe cloaca (15 of 15 cases examined,Fig. 2K,L,Q,R). Therefore, implanta-
tion of SDF-1–expressing cells in theproximal hindlimb led to a decreasein myogenic cells moving out towardthe developing cloaca, whereas nosuch effect was observed after implan-tation of control EGFP-expressingcells.
CXCR4 Inhibitors (T140/
TN14003) Affect the Migration
of Cloacal Myogenic
Precursors
We implanted acrylic beads soaked inT140 and TN14003 at stage HH25–26in the ventral proximal region of thehindlimb. The expression of CXCR4was prominently reduced in the ven-tral proximal side of hindlimb wherethe beads were placed as well as inthe extension band toward the cloaca(16 of 18 cases examined, Fig. 3A).Similar effects were observed withPax7 (Fig. 3E) and MyoD (12 of 15cases examined, Fig. 3I). The reducedexpression of CXCR4 (Fig. 3B), Pax7(Fig. 3F), and MyoD (Fig. 3J) could belikewise appreciated in histologicalsections of these animals. Controlphosphate buffered saline (PBS)beads showed no effect on theCXCR4, Pax7, and MyoD populationin the limb bud (10 of 10 cases exam-ined, Fig. 3C,D,G,H,K,L).
SDF-1-EGFP Cells and CXCR4
Inhibitors Affect the
Formation of Cloacal Muscle
Mass
In our next experiments, we wereinterested in determining if applica-tion of SDF-1–expressing cells andCXCR4 inhibitors to the proximalhindlimb would have an impact oncloacal muscle formation. After im-plantation of SDF-1 cells or T140/TN14003 beads into the proximalhindlimb (Figs. 4A, 5A,B), we reincu-bated manipulated embryos forextended time periods and analyzed
Fig. 3. A–L: CXCR4 inhibitor (T140/TN14003) affects the migration of cloacal myogenic precur-sors. Acrylic beads soaked in T140/TN14003 were implanted in the ventral proximal side of thehindlimb. Operated embryos were reincubated until Hamburger and Hamilton stage HH29–HH30. These embryos were submitted for in situ hybridization to observe the effect on genes ofinterest. A: The expression of CXCR4 was remarkably reduced in the proximal ventral region ofthe hindlimb, where the T140/TN14003 source was implanted (green arrow). E,I: Similar effectswere observed with Pax7 and MyoD, respectively (green arrows). B,F,J: In vibratome sections,the decreased expression of CXCR4, Pax7, and MyoD is evident (black arrows). C,G,K: Phos-phate buffered saline (PBS) beads showed no effect on CXCR4, Pax7, and MyoD expression inthe limb bud (white arrows). D,H,L: Sections showed no decrease in the migratory myoblasts to-ward the cloaca either (black arrows). The scale bar represents 100mm in each photo.
Fig. 4. A–F: COS-1-SDF-1-EGFP–expressingcells affect the formation of cloacal muscle. A:To observe the alterations in the cloacal mor-phology at Hamburger and Hamilton stageHH33, after injecting the SDF-1-EGFP–express-ing cells (white arrow) into the ventral proximalside of the hindlimb at HH26, we hybridizedmanipulated embryos with a probe for MyoD.B,C: Decreased in MyoD expression showedthat the cloacal muscle is decreased due to theinhibition of the muscle precursor migrationfrom the ventral side of the hindlimb toward thecloaca (red arrows). D–F: Control embryos thatwere injected with control EGFP-expressingcells (white arrow in D) showed no effect oncloacal musculature (red arrowheads in E,F).Scale bar ¼ 100 mm in each photo. EGFP,enhanced green fluorescent protein.
Dev
elop
men
tal D
ynam
ics
CXCR4/SDF-1 AXIS DURING CLOACAL MUSCLE FORMATION 1625

these embryos at stage HH33 to lookfor any possible alteration in cloacalmuscle formation. We observed thatthe cloacal muscle is decreased due todecreased muscle precursor cellmigration from the ventral side of thehindlimb toward the cloaca (7 of 8cases examined, 8 of 10 cases exam-ined after SDF-1 cells injection andT140/TN14003 bead implantation,respectively, Fig. 4B,C, and Fig. 5A–
C). Control embryos that wereinjected with control EGFP-express-ing cells or PBS beads showed noeffect on cloacal musculature (10 of 10cases examined, Fig. 4D–F and Fig.5D–F). The beads do not cause a me-chanical hindrance or shielding effectto perturb the migration of myogenicprecursors as no decrease of musclemass is observed in the embryos inwhich PBS beads were inserted.
T140/TN4003 Do Not Cause
Cell Death
To ensure that the reduction inexpression of CXCR4, Pax7, andMyoD was not a result of cell deathfollowing T140/TN14003 bead implan-tation, we used the Nile blue sulfatecell death detection method (van denEijnde et al., 1997; Zuzarte-Luis andHurle, 2002) after T140/TN14003 andPBS bead implantation at intervals of24 hr. No increased uptake of Nileblue sulfate was observed indicatingabsence of cell death (5 of 5 casesexamined, Fig. 6A,B). We have alsopreviously documented that T140/TN14003 at concentrations of 5–10mg/ml does not induce cell death(Yusuf et al., 2006).
Cloacal Muscle Precursors Are
CXCR4- and Pax7-Positive
We were interested to know whetherCXCR4þ cloacal muscle precursorcells also express Pax7. For this rea-son, we have performed double immu-nohistostaining on paraffin sections ofmouse embryos at stage embryonicday (E) 13.5. We observed that the clo-acal muscle precursor cells coexpressCXCR4 (Fig. 7A) and Pax7 (Fig. 7B; 4of 5 cases examined, Fig. 7C).
Ectopic Application of COS-1-
SDF-1-EGFP Cells in the
Chicken and Mice Hindlimb
Attract Migrating Cloacal
Myoblast
To determine if CXCR4þ muscle pre-cursors respond to ectopic SDF-1source, we applied COS-1-SDF-1-EGFPcells ectopically within the hindlimb ofchicken and mouse embryos at stageHH26 and E12.5, respectively, in limbcultures. For this, COS-1 cells weretransiently transfected with an SDF-1-EGFP expression construct andimplanted into the hindlimb bud ofchicken (HH26) and mouse (E12.5)embryos and thereafter the operatedlimbs were removed and cultured incollagen culture gel system. An accu-mulation of CXCR4þ muscle precursorsectopically around the SDF-1-EGFP–expressing cells was observable in themurine hindlimbs (16 of 18 casesexamined, Fig. 8A,B). Furthermore, asimilar agglomeration of Pax7þ cells
Fig. 5. A–F: CXCR4 inhibitors (T140/TN14003) affect the formation of cloacal muscle mass.A,B: To analyze the changes in the cloacal morphology at the later stage (Hamburger and Hamil-ton stage HH33) following implantion of beads soaked in T140/TN14003 (black arrows in A andB) in the ventral proximal side of the hindlimb at HH26. Similarly, in situ hybridization have beendone along with MyoD probe. A–C: The expression of MyoD is remarkably reduced in the cloa-cal region, showing that the cloacal muscle decreased due to inhibition of muscle precursormigration from the ventral side of the hindlimb toward the cloaca (green arrows). D–F: Controlembryos that were implanted with control phosphate buffered saline (PBS) beads (black arrowsin D and E) showed no effect on cloacal musculature (red arrowheads in D, E, and F). Scale bar¼ 100 mm in each photo.
Fig. 6. A,B: CXCR4 inhibitor (T140/TN14003) does not cause cell death. The Nile blue sulfatetest for cell death showed no noticeable changes around the source of T140/TN14003 andaround the control phosphate buffered saline (PBS) beads (black arrows). Scale bar ¼ 100 mmin each photo.
Dev
elop
men
tal D
ynam
ics
1626 REHIMI ET AL.

around the SDF-1-EGFP–expressingcells occurred in cultured chicken hind-limb. These results show that CXCR4þ/Pax7þ cells are able to respond andmigrate to ectopic SDF-1 source (11 of15 cases examined, Fig. 8C,D). Similareffects were absent in control limbsthat received transplants of EGFP-transfected COS-1 cells (data notshown).
DISCUSSION
Cell migration is a central process inthe normal physiology, development,and maintenance of multicellular
organisms. During development, thechemokine, SDF-1 and its receptor,CXCR4 play an important role in dif-ferent processes such as vasculariza-tion and embryogenesis. Mutations ofCXCR4 or SDF-1 affect the migrationof cerebellar granule cells and hippo-campal and cortical neuronal progeni-tors (Lazarini et al., 2003; Pujol et al.,2005). CXCR4/SDF-1 signaling is alsonecessary for the migration of primor-dial germ cells in fish, avians, andmammals (Knaut et al., 2003; Stebleret al., 2004). We have previously ana-lyzed the expression pattern ofCXCR4 (Yusuf et al., 2005) and SDF-1
(Rehimi et al., 2008) in the developingchicken embryo and found that it isexpressed in a multitude of develop-ing embryonic structures undergoingactive morphogenesis. In this presentwork, we report the expression ofCXCR4 and SDF-1 during cloacalmuscle myogenic precursor migrationin chicken embryos and unravel theirrole during the migration of these clo-acal muscle precursors from the prox-imal hindlimb toward the developingcloaca.In an elaborate work, Valasek et al.
(2005) could show that the cloacalmusculature is of hypaxial origin andthat cloacal muscle precursors tempo-rarily reside within the ventral mus-cle mass of the hindlimbs in chickenand mouse embryos. Surgical experi-ments and detailed analyses ofgenetic mutants allowed us to con-clude that the formation of a normalproximal hindlimb field and thedelamination of the myogenic precur-sors from the dermomyotome are anessential prerequisite for the correctdevelopment of cloacal muscles (Vala-sek et al., 2005). Limbless (Prahladet al., 1979) and met mutants (Die-trich et al., 1999), from which the clo-acal muscles are completely absent,strengthen this hypothesis. Interest-ingly however, analysis of Lbx1mutants showed a normal cloacal de-velopment (Valasek et al., 2005), prob-ably because progenitors of the ven-tral premuscle masses developnormally in this mutant.Cloacal muscle precursors originate
predominantly from the same axialsomitic levels that give rise to hind-limb muscles, namely somite 30 to 34in chicken embryos. Only somite 34provides muscle precursors exclu-sively for the cloaca, whereas somites30–33 (together with somite 26–29)additionally provide limb muscle pre-cursors. Stepwise analysis of the mi-gratory pattern of cloacal muscle pre-cursors shows that two separablemigratory events are involved duringcloacal muscle formation. In thechicken embryo, the myogenic precur-sors first delaminate from the hypax-ial dermomyotome at appropriateaxial levels and migrate into thehindlimb field as individual mig-rating precursors. This migrationevent requires an effective delamina-tion of myogenic precursors from the
Fig. 7. A–C: Cloacal muscle precursors coexpress CXCR4 and Pax7. A,B: Antibody stainingon paraffin sections of embryonic day (E) 13.5 mouse embryos for CXCR4 (white arrowheads inA) and Pax7 (green arrowheads in B). C: We analyzed that a majority of cloacal muscle precur-sors coexpress CXCR4 and Pax7 (white arrows). Scale bar ¼ 100 mm in each photo.
Fig. 8. A–D: Ectopic application of COS-1-SDF-1-EGFP cells in the chicken and mouse hind-limb in collagen culture system attracts migrating cloacal myoblast. To determine if CXCR4þ
muscle progenitor cells respond to SDF-1, SDF-1 cells were ectopically implanted into the hind-limb of chicken (HH26) and mouse (E12.5) embryos. These limbs were then stained with CXCR4and Pax7 antibody. An agglomeration of CXCR4- and Pax7-expressing cells around the COS-1-SDF-1-EGFP–expressing cells was clearly observable in chicken and mouse, respectively (bluearrows). Scale bar ¼ 100 mm in each photo.
Dev
elop
men
tal D
ynam
ics
CXCR4/SDF-1 AXIS DURING CLOACAL MUSCLE FORMATION 1627

hypaxial dermomyotome and is de-pendent on Met and SF/HGF signal-ing, while migration of these hindlimbmuscle precursors deeper into thelimb mesenchyme additionally usesthe CXCR4/SDF-1 signaling (Vasyu-tina et al., 2005) and Lbx1 (Birchme-ier and Brohmann, 2000). The secondmigration event occurs once themigration of muscle precursors fromthe somites toward the hindlimb mes-enchyme has ceased around stageHH25–26. Cloacal precursors nowextend similar to myotomal exten-sions for the ventral body wallmuscles from the proximal ventralmuscle mass of the hindlimb towardthe future cloacal anlage in the mid-line (Valasek et al., 2005). During theinitial migratory event, the cells thatdelaminate from the hypaxial dermo-myotome express met, Pax3, Lbx-1,and only after they have entered thelimb mesenchyme, CXCR4 expressionis detectable. Data from mouse andchicken support the notion thatCXCR4-positive cells form a subpopu-lation of a larger Lbx1-expressingpool of the muscle precursors invad-ing the limb (Vasyutina et al., 2005).Eventually, as the initial migrationevent ceases, genes necessary for theinitiation of the myogenic differentia-tion such as MyoD, Myogenin, andMrf4 are turned on. It has beenshown that, as differentiation pro-gresses in the ventral and dorsal pre-muscle masses in the limb, there is agradual decrease of the Pax3þ/Lbx1þ–expressing precursor pool coupledwith a gradual rise of MyoDþ precur-sors. Analysis at the cellular resolu-tion has shown that apart from theexclusively Lbx1þ- and MyoDþ-expressing precursors, a mixed popu-lation of the MyoDþ/Pax3þ/Lbx1þ
pool is frequently encountered; how-ever, none or very few CXCR4þ/MyoDþ precursors are seen (Vasyu-tina et al., 2005). Therefore, CXCR4expression in limb muscle precursorsis lost as the differentiation begins.The scenario is different during thesecond migratory event when cloacalmuscle precursors move from theproximal ventral hindlimb toward thecloacal tubercle. These precursorshave already initiated the myogenicprogram and express MyoD. This issimilar to the hypaxial myotomalextensions that partially express
MyoD and form the ventral body wallmusculature.
We report here for the first timethat CXCR4 and its ligand SDF-1 areexpressed at appropriate stages dur-ing the second migratory event andthat they play a role during thismigration. After experimentally per-turbing the CXCR4/SDF-1 signalingpathway in the proximal hindlimb,the migration of cloacal muscle pre-cursors was affected. The secondmigration event during cloacal muscleformation is reminiscent of the forma-tion of myotomal/dermomyotomalextensions giving rise to ventral bodywall muscles and may possibly sharecommon molecular effectors. We haveobserved the expression of CXCR4 inthe hypaxial myotomes/dermomyo-tome that extend ventrally to formthe ventral body wall muscles (datanot shown).
Whether CXCR4þ cells represent asubpopulation of the cloacal muscleprecursors participating in the secondmigratory event needs to be furtherstudied. There are indications for thisas never during our experiments dowe observe the entire cloacal muscleformation can be impaired, pointingtoward the existence of a CXCR4�
subpopulation that is not affected byour experimental manipulations tar-geting the CXCR4þ cell pool. How-ever, this issue needs to be furtheraddressed at the cellular level to showif any such subpopulations exist. Asmentioned, similar CXCR4þ andCXCR4� myogenic presursor cellshave been described in the context oflimb muscle precursors (Vasyutinaet al., 2005).
The role of chemokine receptorCXCR4 and its ligand SDF-1 duringmyogenic cell migration for the forma-tion of cloacal musculature add up tothe list of several roles of this signal-ing pair during development. The for-mation of the cloaca is of utmost im-portance for the development andpatterning of the lower gastrointesti-nal and urogenital tract. Defect in thecloacal formation could lead to vary-ing degress of anorectal malforma-tions often accompanied with func-tional disability for the newborninfants needing immediate surgicalintervention. How far defective cloa-cal muscle formation resulting fromperturbed cell migration contribute to
cloacal defects and eventual anorectalmalformation has not been addressedas molecular mechanisms of cloacalmuscle formation have just started toemerge. Understanding developmen-tal cloacal myogenic migration andcloacal muscle formation, would helpin understanding the pathogenesis ofanorectal anomalies where defectivemyogenic cell migration is the pri-mary underlying cause. Several tissueinteractions have been described inthe literature that influence myogenicpatterning like epithelio-mesenchy-mal crosstalk (Anakwe et al., 2003),myogenic precursor-limb mesen-chyme precursor interactions (Bona-fede et al., 2006) and blood vessel for-mation and muscle splitting (Tozeret al., 2007). Therefore, further stud-ies investigating the role of signa-ling molecules such as Shh and Wntssignaling aimed at analyzing theinteraction of cloaca endodermal lin-ing with the surrounding mesen-chymal tissue and muscle tissuewould greatly help in under-standing the pathogenesis of anorec-tal malformations.
EXPERIMENTAL
PROCEDURES
Chicken Embryos
Fertilized chicken eggs obtained froma local breeder were incubated at37�C and 80% humidity. Following op-erative procedures, the eggs were re-incubated at 37�C and 80% humidityto develop further to the requiredstages. Staging was done according toHamburger and Hamilton (1992).
Construction of the SDF-1
Transgene Plasmid
For obtaining the chicken SDF-1gene, reverse transcriptase-polymer-ase chain reaction (RT-PCR) wasperformed with a pair of cSDF-1gene-specific primers (sense primer:50 GCCTGCACCGTCGCCAGAATG30; and antisense primer: 50 AGGCCAACTCCAAACCCATCTTCA 30). TheRT-PCR product (425 bp) was clonedinto pDrive vector and the cSDF-1insert was confirmed by sequencing.The full coding sequence of the cSDF-1 cDNA can be re-harvested from theconstructed pDrive- cSDF-1 plasmid
Dev
elop
men
tal D
ynam
ics
1628 REHIMI ET AL.

digested with both NheI and PstI. Forconstruction of the cSDF-1 overex-pression plasmid, the NheI–PstI frag-ment containing the full-lengthchicken SDF-1 coding sequence wasre-cloned into a prelinearizedpIRES2-EGFP vector (BD BiosciencesClontech) treated with the samerestriction enzymes.
Cell Culture, Transfection,
and Implantation In Ovo
COS-1 cells were a gift from Prof. Dr.Carmen Birchmeier (Max-Delbruck-Center for Molecular Medicine, Ber-lin). COS-1 cells were grown andtransfected with SDF-1-EGFP con-structs as described previously(Vasyutina et al., 2005). In brief,COS-1 cells were grown in DMEM(Dulbecco’s modified Eagle’s medium;Invitrogen, USA) supplemented with10% FCS (fetal calf serum; Invitro-gen) in a 37�C humidified atmosphereof 5% CO2 in air. Cells were passagedby treatment with 0.25% trypsin (w/v)/1 mM EDTA (ehtylenediaminete-traacetic acid; Invitrogen), harvested,centrifuged, and then resuspended inDMEM (DMEM; Invitrogen, USA)supplemented with 10% FCS (Invitro-gen) onto fresh flasks.
Cells were seeded 1 day beforetransfection at a density of 3 � 105
cells per T25 flask. Cells were approx-imately 70% confluent on the day ofgene transfection. SDF-1-EGFPtransfection was carried out with Lip-ofectamine (Invitrogen) according tothe manufacturer’s protocol. Trans-fection efficiency was estimated byenhanced green fluorescent protein(EGFP) expression. One or two daysafter gene transfection, cells weresubjected to drug selective medium,which was additionally supplementedwith 400 mg/ml G418 (Invitrogen).Individual colonies appeared after2–3 weeks and were picked by micro-pipette controlled by a three-dimen-sional controller. Resuspended colo-nies were transferred into new T25flasks. For controls, 10 mg pcDNA-EGFP was transfected into COS cells.
Eggs were windowed, and afterremoval of the extraembryonic mem-branes with a tungsten needle, con-centrated SDF-1-EGFP–transfectedCOS-1 cell suspensions were injectedproximally into the developing hind-
limb bud of stage HH25–26 chickenembryos using a capillary glass nee-dle. EGFP transfected COS-1 cellswere injected as controls.
CXCR4 Inhibitors (T140/
TN14003) and Beads
Implantation In Ovo
Highly specific peptidic inhibitors ofCXCR4 (T140/TN14003), obtained asa gift from the group of HirokazuTamamura (Institute of Biomaterialsand Bioengineering, Tokyo Medicaland Dental University, Chiyoda-ku,Tokyo, 101-0062, Japan), were dis-solved in PBS to a concentration 15mg/ml (Yusuf et al., 2006). For appli-cation of inhibitor, the acrylic beadsapproximately 80–120 mm in diame-ter (Bio-Rad Laboratories) wererinsed in PBS and incubated withCXCR4 inhibitor (T140/TN14003) so-lution overnight at 4 �C. Controlbeads were incubated in PBS. Forbead implantation, extraembryonicmembranes (stage HH25–26) wereremoved with a sharpened tungstenneedle, and a bead was inserted intothe ventral proximal side of the hind-limb mesenchyme using a blunt glassneedle. Embryos were reincubated toreach stage HH29–HH30. PBS-soaked beads were used as controls.All embryos were fixed in 4% parafor-maldehyde (PFA; VWR International)and further submitted for in situhybridization.
In Situ Hybridization
Selected embryos were tested forCXCR4, Pax7, and MyoD using highlyspecific RNA probes. The CXCR4probe used was a 693-bp chickenprobe. The Pax7 probe used was a582-bp chicken probe. The MyoDprobe was a 1,518-bp specific chickenprobe. Riboprobes were labeled with adigoxigenin RNA labeling kit fromBoehringer, Mannheim, Germany.Whole-mount in situ hybridizationwas performed as described previ-ously (Nieto et al., 1996).
Immunohistochemistry on
Paraffin Sections of Mice and
Chicken Embryos
For immunohistochemistry, histologi-cal sections of chicken and mouse
embryos were fixed overnight in 4%PFA (VWR International) in PBS.Nonspecific binding sites wereblocked with 1% bovin serum albumin(BSA, Sigma) in PBS for 30 min. Anti-CXCR4 antibody (ab2074, Abcam;1:200 in BSA) and anti-Pax7 antibody(Hybridoma bank; 1:50 in PBS) werediluted in 1% BSA (Sigma) in PBS.Sections were incubated with the pri-mary antibodies overnight in a humidchamber at 4�C. Primary antibodieswere detected by incubating sectionsfor 2 hr at room temperature with ei-ther fluorescein isothiocyanate-(FITC) or Cy3-labeled secondary anti-bodies (Cy3-conjugated goat anti-mouse IgG antibody; Jackson Immu-noResearch, 1:100 in PBS). Subse-quently, sections were washed inPBS, embedded in Aquatex (Mount-ing agent, Merk), and analyzed withan epifluorescence microscope (Axio-scope 20; Zeiss).
Immunohistochemistry on
Hindlimb in Collagen Gel
Cultures
Immunohistochemistry was per-formed on hindlimbs cultured in colla-gen gels following fixation in 4% PFA(8.18715, Merck) in PBS for 2 hr inwell plates. Collagen gels with oper-ated hindlimb were washed with PBSfor overnight. Nonspecific bindingsites were blocked with 5% lamb se-rum (LS, 16070096 Invitrogen), 20%dimethyl sulfoxide (DMSO; D8418,Sigma) in PBS for 1 hr. Anti-CXCR4antibody (ab2074, Abcam; 1:200 inBSA) and anti-Pax7 antibody (Hy-bridoma Bank; 1:50 in BSA) werediluted in 5% lamb serum (LS,Sigma), 20% DMSO (D8418, Sigma)in PBS. The gels containing the hin-dlimbs were then incubated with theprimary antibodies for 2 days in ahumid chamber at 4�C and washedagain with PBS overnight. Primaryantibodies were detected by incubat-ing gels overnight at room tempera-ture in dark with either FITC- orCy3-labeled secondary antibodies(Cy3-conjugated goat anti-mouse IgGantibody; Jackson ImmunoResearch,1:100 in PBS). Before mounting thecollagen gels on slides, we preparedspacers from plasticine to hold thecover slips. Coverslips with collagen
Dev
elop
men
tal D
ynam
ics
CXCR4/SDF-1 AXIS DURING CLOACAL MUSCLE FORMATION 1629

gels were carefully mounted in 90%glycerol (G7757, Sigma) in PBS.
Cell Death Test
Cell death test was performed afterthe implantation of T140/TN14003-soaked beads, using Nile Blue sulfate(van den Eijnde et al., 1997). After re-moval of the extra embryonic mem-branes, the embryos were stained intoto for 30 min at 37�C in a Nile Bluesulfate solution (1:20,000 w/v). Afterincubation, the specimens werewashed twice with PBS, put on ice,and immediately photographed undera microscope equipped with a digitalcamera connected to a computer(Leica DFG320).
Microscopy and Imaging
After in situ hybridization, sampleswere observed and photographedusing the Leica MZFLIII microscopeand Leica DC 300F digital camera.Hybridized sections were observedand photographed using an Axioscope20 (Zeiss) and a Leica DFC320 digitalcamera.
ACKNOWLEDGMENTSWe thank Prof. Dr. Carmen Birchme-ier (Max-Delbruck-Center for Molecu-lar Medicine, Berlin) for providing theCOS-1 cells, Prof. Dr. Erez Raz (CenterforMolecular Biology of Inflammation,Munster) for kindly providing theCXCR4 chicken probe and HirokazuTamamura (Institute of Biomaterialsand Bioengineering, Tokyo Medicaland Dental University) for sharing theCXCR4 inhibitor. We also thank Prof.Dr. Rujin Huang and Susanne Theis(Bonn) and Prof. Dr. B. Heimrich(Freiburg). All authors extend theirthanks to Ellen Gimbel, SusannaGlaser, and Ulrike Pein for their excel-lent technical assistances. We alsoacknowledge the support of ‘‘Molecu-lar mechanisms of migration, invasionand metastasis,’’ a Baden-Wurttem-berg grant, the Myores Project(511978) funded by the EU’s sixthframework Programme and GKR1104.
REFERENCES
Anakwe K, Robson L, Hadley J, Buxton P,Church V, Allen S, Hartmann C, HarfeB, Nohno T, Brown AM, Evans DJ,
Francis-West P. 2003. Wnt signallingregulates myogenic differentiation inthe developing avian wing. Develop-ment 130:3503–3514.
Birchmeier C, Brohmann H. 2000. Genesthat control the development of migrat-ing muscle precursor cells. Curr OpinCell Biol 12:725–730.
Bober E, Franz T, Arnold HH, Gruss P,Tremblay P. 1994. Pax-3 is required forthe development of limb muscles: a pos-sible role for the migration of dermo-myotomal muscle progenitor cells.Development 120:603–612.
Bonafede A, Kohler T, Rodriguez-Nieden-fuhr M, Brand-Saberi B. 2006. BMPsrestrict the position of premusclemasses in the limb buds by influencingTcf4 expression. Dev Biol 299:330–344.
Brand-Saberi B, Christ B. 2000. Evolutionand development of distinct cell line-ages derived from somites. Curr TopDev Biol 48:1–42.
Brohmann H, Jagla K, Birchmeier C.2000. The role of Lbx1 in migration ofmuscle precursor cells. Development127:437–445.
Chevallier A, Kieny M, Mauger A. 1977.Limb-somite relationship: origin of thelimb musculature. J Embryol Exp Mor-phol 41:245–258.
Christ B, Jacob HJ, Jacob M. 1974. [Ex-perimental studies on the developmentof the thoracic wall in chick embryos].Experientia 30:1449–1451.
Christ B, Jacob HJ, Jacob M. 1977. Ex-perimental analysis of the origin of thewing musculature in avian embryos.Anat Embryol (Berl) 150:171–186.
Christ B, Ordahl CP. 1995. Early stagesof chick somite development. AnatEmbryol (Berl) 191:381–396.
David NB, Sapede D, Saint-Etienne L,Thisse C, Thisse B, Dambly-ChaudiereC, Rosa FM, Ghysen A. 2002. Molecularbasis of cell migration in the fish lateralline: role of the chemokine receptorCXCR4 and of its ligand, SDF1. ProcNatl Acad Sci U S A 99:16297–16302.
de Santa Barbara P, van den Brink GR,Roberts DJ. 2002. Molecular etiology ofgut malformations and diseases. Am JMed Genet 115:221–230.
Dietrich S, Abou-Rebyeh F, Brohmann H,Bladt F, Sonnenberg-Riethmacher E,Yamaai T, Lumsden A, Brand-Saberi B,Birchmeier C. 1999. The role of SF/HGFand c-Met in the development of skeletalmuscle. Development 126:1621–1629.
Dravis C, Yokoyama N, Chumley MJ,Cowan CA, Silvany RE, Shay J, BakerLA, Henkemeyer M. 2004. Bidirectionalsignaling mediated by ephrin-B2 andEphB2 controls urorectal development.Dev Biol 271:272–290.
Evans DJ, Valasek P, Schmidt C, Patel K.2006. Skeletal muscle translocation invertebrates. Anat Embryol (Berl)211(suppl 1):43–50.
Galis F. 2001. Evolutionary history of ver-tebrate appendicular muscle. Bioessays23:383–387.
Gross MK, Moran-Rivard L, Velasquez T,Nakatsu MN, Jagla K, Goulding M.
2000. Lbx1 is required for muscle pre-cursor migration along a lateral path-way into the limb. Development 127:413–424.
Haas P, Gilmour D. 2006. Chemokine sig-naling mediates self-organizing tissuemigration in the zebrafish lateral line.Dev Cell 10:673–680.
Hamburger V, Hamilton HL. 1992. A se-ries of normal stages in the develop-ment of the chick embryo. 1951. DevDyn 195:231–272.
Hynes PJ, Fraher JP. 2004. The develop-ment of the male genitourinary system.I. The origin of the urorectal septumand the formation of the perineum. BrJ Plast Surg 57:27–36.
Jacob M, Christ B, Jacob HJ. 1979. Themigration of myogenic cells from thesomites into the leg region of avianembryos. An ultrastructural study.Anat Embryol (Berl) 157:291–309.
Keith A, George H, Patrick MJ. 2005.Pediaric surgery. ( 4th ed.), Philadel-phia: Elsevier Saunders. p 496–517.
Kimmel SG, Mo R, Hui CC, Kim PC.2000. New mouse models of congenitalanorectal malformations. J PediatrSurg 35:227–230; discussion 230–221.
King AS. McLelland. 1981. Form andfunction in birds. Vol 2. London: Aca-demic Press.
Knaut H, Werz C, Geisler R, Nusslein-Volhard C. 2003. A zebrafish homologueof the chemokine receptor Cxcr4 is agerm-cell guidance receptor. Nature421:279–282.
Lazarini F, Tham TN, Casanova P, Are-nzana-Seisdedos F, Dubois-Dalcq M.2003. Role of the alpha-chemokine stro-mal cell-derived factor (SDF-1) in thedeveloping and mature central nervoussystem. Glia 42:139–148.
Li Q, Shirabe K, Kuwada JY. 2004. Che-mokine signaling regulates sensory cellmigration in zebrafish. Dev Biol 269:123–136.
Martinez-Frias ML, Bermejo E, Rodri-guez-Pinilla E. 2000. Anal atresia, ver-tebral, genital, and urinary tractanomalies: a primary polytopic develop-mental field defect identified throughan epidemiological analysis of associa-tions. Am J Med Genet 95:169–173.
McBride HJ, Fatke B, Fraser SE. 2003.Wnt signaling components in thechicken intestinal tract. Dev Biol 256:18–33.
Mo R, Kim JH, Zhang J, Chiang C, HuiCC, Kim PC. 2001. Anorectal malforma-tions caused by defects in sonic hedgehogsignaling. Am J Pathol 159:765–774.
Murdoch C. 2000. CXCR4: chemokine re-ceptor extraordinaire. Immunol Rev 177:175–184.
Nagasawa T, Hirota S, Tachibana K,Takakura N, Nishikawa S, Kitamura Y,Yoshida N, Kikutani H, Kishimoto T.1996. Defects of B-cell lymphopoiesisand bone-marrow myelopoiesis in micelacking the CXC chemokine PBSF/SDF-1. Nature 382:635–638.
Nieto MA, Patel K, Wilkinson DG. 1996.In situ hybridization analysis of chick
Dev
elop
men
tal D
ynam
ics
1630 REHIMI ET AL.

embryos in whole mount and tissue sec-tions. Methods Cell Biol 51:219–235.
Penington EC, Hutson JM. 2003. The ab-sence of lateral fusion in cloacal parti-tion. J Pediatr Surg 38:1287–1295.
Prahlad KV, Skala G, Jones DG, BrilesWE. 1979. Limbless: a new genetic mu-tant in the chick. J Exp Zool 209:427–434.
Pujol F, Kitabgi P, Boudin H. 2005. Thechemokine SDF-1 differentially regu-lates axonal elongation and branchingin hippocampal neurons. J Cell Sci 118:1071–1080.
Rees E, Young RD, Evans DJ. 2003. Spa-tial and temporal contribution ofsomitic myoblasts to avian hind limbmuscles. Dev Biol 253:264–278.
Rehimi R, Khalida N, Yusuf F, Dai F,Morosan-Puopolo G, Brand-Saberi B.2008. Stromal-derived factor-1 (SDF-1)expression during early chick develop-ment. Int J Dev Biol 52:87–92.
Schafer K, Braun T. 1999. Early specifica-tion of limb muscle precursor cells bythe homeobox gene Lbx1h. Nat Genet23:213–216.
Stebler J, Spieler D, Slanchev K, Moly-neaux KA, Richter U, Cojocaru V, Tara-
bykin V, Wylie C, Kessel M, Raz E.2004. Primordial germ cell migration inthe chick and mouse embryo: the role ofthe chemokine SDF-1/CXCL12. DevBiol 272:351–361.
Tai CC, Sala FG, Ford HR, Wang KS, LiC, Minoo P, Grikscheit TC, Bellusci S.2009. Wnt5a knock-out mouse as a newmodel of anorectal malformation. JSurg Res 156:278–282.
Tajbakhsh S, Rocancourt D, Cossu G,Buckingham M. 1997. Redefining thegenetic hierarchies controlling skeletalmyogenesis: Pax-3 and Myf-5 actupstream of MyoD. Cell 89:127–138.
Tozer S, Bonnin MA, Relaix F, Di SavinoS, Garcia-Villalba P, Coumailleau P,Duprez D. 2007. Involvement of vesselsand PDGFB in muscle splitting duringchick limb development. Development134:2579–2591.
Valasek P, Evans DJ, Maina F, Grim M,Patel K. 2005. A dual fate of the hindlimb muscle mass: cloacal/perineal mus-culature develops from leg muscle cells.Development 132:447–458.
van den Eijnde SM, Luijsterburg AJ,Boshart L, De Zeeuw CI, van Dieren-donck JH, Reutelingsperger CP, Ver-
meij-Keers C. 1997. In situ detection ofapoptosis during embryogenesis withannexin V: from whole mount to ultra-structure. Cytometry 29:313–320.
Vasyutina E, Stebler J, Brand-Saberi B,Schulz S, Raz E, Birchmeier C. 2005.CXCR4 and Gab1 cooperate to controlthe development of migrating muscleprogenitor cells. Genes Dev 19:2187–2198.
Yusuf F, Rehimi R, Dai F, Brand-SaberiB. 2005. Expression of chemokine re-ceptor CXCR4 during chick embryo de-velopment. Anat Embryol (Berl) 210:35–41.
Yusuf F, Rehimi R, Morosan-Puopolo G,Dai F, Zhang X, Brand-Saberi B. 2006.Inhibitors of CXCR4 affect the migra-tion and fate of CXCR4þ progenitors inthe developing limb of chick embryos.Dev Dyn 235:3007–3015.
Zou YR, Kottmann AH, Kuroda M, Taniu-chi I, Littman DR. 1998. Function ofthe chemokine receptor CXCR4 in hae-matopoiesis and in cerebellar develop-ment. Nature 393:595–599.
Zuzarte-Luis V, Hurle JM. 2002. Pro-grammed cell death in the developinglimb. Int J Dev Biol 46:871–876.
Dev
elop
men
tal D
ynam
ics
CXCR4/SDF-1 AXIS DURING CLOACAL MUSCLE FORMATION 1631




















