
Upregulation of REG Iα accelerates tumor progression in pancreatic cancer with diabetes

Inhibition of Paraquat-Induced Autophagy Accelerates the ApoptoticCell Death in Neuroblastoma SH-SY5Y Cells
Rosa A. Gonzalez-Polo,*,1 Mireia Niso-Santano,* Miguel A. Ortız-Ortız,* Ana Gomez-Martın,* Jose M. Moran,*Lourdes Garcıa-Rubio,† Javier Francisco-Morcillo,‡ Concepcion Zaragoza,§ German Soler,* and Jose M. Fuentes*
*CIBER de Enfermedades Neurodegenerativas, Departamento de Bioquımica y Biologıa Molecular y Genetica, E.U. Enfermerıa y T.O., Universidad de
Extremadura, Avda. Universidad s/n 10071 Caceres, Spain; †Departamento de Quımica Organica, ‡Departamento de Biologıa Celular, §Departamento
de Medicina y Sanidad Animal, Facultad de Veterinaria, Universidad de Extremadura, Caceres, Spain
Received January 12, 2007; accepted February 27, 2007
Autophagy is a degradative mechanism involved in the recy-
cling and turnover of cytoplasmic constituents from eukaryotic
cells. This phenomenon of autophagy has been observed in
neurons from patients with Parkinson’s disease (PD), suggesting
a functional role for autophagy in neuronal cell death. On the
other hand, it has been demonstrated that exposure to pesticides
can be a risk factor in the incidence of PD. In this sense, paraquat
(PQ) (1,1#-dimethyl-4,4#-bipyridinium dichloride), a widely used
herbicide that is structurally similar to the known dopaminergic
neurotoxicant MPP+ (1-methyl-4-phenyl-pyridine), has been sug-
gested as a potential etiologic factor for the development of PD.
The current study shows, for the first time, that low concen-
trations of PQ induce several characteristics of autophagy in
human neuroblastoma SH-SY5Y cells. In this way, PQ induced
the accumulation of autophagic vacuoles (AVs) in the cytoplasm
and the recruitment of a LC3-GFP fusion protein to AVs.
Furthermore, the cells treated with PQ showed an increase of
the long-lived protein degradation which is blocked in the
presence of the autophagy inhibitor 3-methyladenine and regu-
lated by the mammalian target of rapamycin (mTOR) signaling.
Finally, the cells succumbed to cell death with hallmarks of
apoptosis such as phosphatidylserine exposure, caspase activation,
and chromatin condensation. While caspase inhibition retarded
cell death, autophagy inhibition accelerated the apoptotic cell
death induced by PQ. Altogether, these findings show the re-
lationship between autophagy and apoptotic cell death in human
neuroblastoma cells treated with PQ.
Key Words: paraquat; vacuoles; apoptosis; Parkinson.
Autophagy is an intracellular lysosome-mediated catabolicmechanism that is responsible for the bulk degradation andrecycling of damaged or dysfunctional cytoplasmic compo-nents and intracellular organelles (Klionsky et al., 2000).Autophagy is a cellular response to both extracellular (nutrientdeprivation or hypoxia) and intracellular (accumulation of
damaged organelles and cytoplasmic components) stress con-ditions. Autophagy, being an evolutionarily ancient cellularresponse to intra- and extracellular noxious stimuli, mayprecede or coexist with apoptosis, and it may be induced byapoptotic stimuli (Xue et al., 1999). Thus, autophagy andapoptosis may be interconnected (Bursch et al., 2000). Cellularautophagy is a physiological degradative process involved, likeapoptosis, in embryonic growth and development, cellularremodeling, and the biogenesis of some subcellular organelles(Filonova et al., 2000; Hariri et al., 2000; Sattler and Mayer,2000). Autophagic cell death involves accumulation of auto-phagic vacuoles (AVs) in the cytoplasm of dying cells as wellas mitochondria dilation and enlargement of the endoplasmicreticulum and the Golgi apparatus. Autophagic cell death hasbeen described during the normal nervous system development(Schweichel et al., 1973) and could be a consequence ofa pathological process such as those associated with neurode-generative diseases (Petersen et al., 2001). Of note, autophagyis highly enhanced in brain amyloidoses (Graeber et al., 2002),Alzheimer disease (Stadelmann et al., 1999), Huntington’sdisease (Landles et al., 2004), and Parkinson’s disease (PD)(Anglade et al., 1997).
PD is a chronic progressive neurodegenerative disease,affecting at least 1% of the population over the age of 55.Genetic forms of the disease represent less than 5% of currentcases, but the causes of the vast majority of sporadic cases ofPD are still unknown. Accumulating evidence strongly pointsto environmental toxins as feasible triggers of neurodegenera-tion of nigrostriatal dopaminergic neurons, and the commonuse of pesticides in rural life has been correlated to parkinson-ism in humans (Di Monte et al., 2000). In this sense, theherbicide 1,1#-dimethyl-4,4#-bipyridinium dichloride (paraquat[PQ]), widely used as a cationic nonselective bipyridylherbicide to control weeds and grasses in many agriculturalareas, has emerged as a putative risk factor on the basis of itsstructural homology to 1-methyl-4-phenyl-pyridine (MPPþ),the active metabolite of 1-methyl-4-phenyl-1,2,3,6 tetrahydro-pyridine, a neurotoxicant that induces Parkinson’s-like features
1 To whom correspondence should be addressed. Fax: þ34-927-257-451.
E-mail: [email protected].
� The Author 2007. Published by Oxford University Press on behalf of the Society of Toxicology. All rights reserved.For Permissions, please email: [email protected]
TOXICOLOGICAL SCIENCES 97(2), 448–458 (2007)
doi:10.1093/toxsci/kfm040
Advance Access publication March 6, 2007
by guest on October 28, 2014
http://toxsci.oxfordjournals.org/D
ownloaded from

in rodents, nonhuman primates, and humans (Langston andBallard, 1984; Tanner and Ben Shlomo, 1999).
Occupational PQ exposures have been associated withparkinsonism (Hertzman et al., 1990; Liou et al., 1997), althoughthe mechanism of PQ toxicity is yet poorly understood. Severalstudies have suggested the involvement of reactive oxygenspecies (ROS) in its effect (Gonzalez-Polo et al., 2004; Mollaceet al., 2003). Our group has demonstrated that free radicals playan active role in the PQ-induced apoptotic events that culminatewith cell death (Gonzalez-Polo et al., 2004).
Here, we show for the first time that low concentrations ofPQ induce several characteristics of autophagy in humanneuroblastoma SH-SY5Y cells, which are commonly selectedas a cellular PD model (Lai et al., 1997; Lee et al., 2000;Shavali et al., 2004). Finally, the cells succumbed to cell deathwith hallmarks of apoptosis. Thus, we investigated the re-lationship between autophagy and apoptosis in neurons treatedwith PQ to propose a new and exciting strategy to theknowledge of the PD mechanism and its possible treatment.
MATERIALS AND METHODS
Cell line and culture condition. Human neuroblastoma SH-SY5Y cells
were grown in Dulbecco’s modified Eagle medium (Gibco BRL, Paisley, U.K.)
containing 100 U/ml penicillin, 100 lg/ml streptomycin, and 10% fetal bovine
serum (Hyclone, Brevieres, France). Cells were seeded at 5 3 105 in a 75-cm2
tissue culture flask (TPP, Trasadingen, Switzerland) and incubated at 37�Cunder a saturating humidity atmosphere of 5% CO2/95% air.
Reagents and cell death induction. Confluent cells (~80%) in 75-cm2
tissue culture flasks were trypsinized and seeded in tissue culture dishes at
a concentration of 5 3 104 cells/cm2. Twenty-four hours later, the medium was
aspirated and replaced with fresh medium alone or containing the indicated
concentrations of PQ (Niso-Santano et al., 2006). Serum and amino acid
starvation of cells were performed using serum-free Earle’s balanced salt
solution medium (Sigma, St Louis, MO) (Boya et al., 2005). For caspase
inhibiton, a sublethal dose of N-benzyloxycarbonyl-Val-Ala-Asp fluorome-
thylketone (Z-VAD-fmk, 50lM, Bachem AG, Bubendorf, Switzerland) was
added 1 h before PQ. The antioxidants N-acetylcysteine (NAC 1mM), vitamin E
(100lM), glutathione (1mM), and the inihibitor of autophagy, 3-methyladenine
(3-MA, 10mM), were added before the addition of PQ.
Flow cytometry. To determine apoptosis-associated changes by cytofluor-
ometry, we used 3,3#-dihexyloxacarbocyanine iodide (DiOC6(3), 40nM) for the
mitochondrial transmembrane potential quantification, propidium iodide (PI,
1 lg/ml) for the determination of cell viability (Boya et al., 2003; Gonzalez-Polo
et al., 2005a), and Annexin-V labeled with fluorescein isothiocyanate (all of
them from Molecular Probes, Eugene, OR) for the assessment of phosphati-
dylserine exposure. For the determination of superoxide anion generation,
hydroethidine (HE, 10lM) was used. After different experimental conditions,
cells were trypsinized and labeled with the fluorochromes at 37�C, followed by
cytoflurometric analysis with a FACS Scan (Becton Dickinson, Frankin Lakes,
NJ) (Gonzalez-Polo et al., 2005a,b).
Immunofluorescence and light microscopy. Cells were cultured on
coverslips. After the experimental conditions, the cells were stained with
CMFDA (5-chloromethylfluorescein diacetate, 1lM) (Molecular Probes) for
30 min at 37�C to visualize the vacuoles. Cells were fixed with para-
formaldehyde (4% wt:vol) for CMFDA staining and immunofluorescence
assays (Gonzalez-Polo et al., 2005a,2005b). Cells were stained for the detection
of activated caspase-3 with a polyclonal antibody from Cell Signaling
Technology, Inc. (Danvers, MA) developed with an anti-rabbit immunoglobulin
Alexa fluor conjugate (Molecular Probes) and counter-stained with Hoechst
33342, 2lM (Ho) (Sigma) before mounting. Fluorescence microscopy was
analyzed with an Olympus IX51 equipped with a DC300F camera.
Western blot analysis. Following experimental treatments, the cells
(cultured in 60 mm dishes) were rinsed twice with cold phosphate-buffered
saline (PBS) and removed by scrapping and then centrifuged at 9003 g for 5 min
at 4�C. Cells were lysed in a buffer containing 50mM N-2-hydroxyethyl-
piperazine-N#-2-ethanesulfonic acid pH 7.5, 100mM NaCl, 1% Triton X-100,
10% glycerol, 1mM ethyleneglycol-bis(aminoethylether)-tetraacetic acid, 5mM
MgCl2, 25mM NaF, and Protease Inhibitor Cocktail (Sigma). Cells were
centrifuged at 13,000 3 g 5 min at 4�C. Supernatants were stored at � 80�Cuntil analysis by Western blot. Protein concentration was measured according to
Bradford (1976) using bovine serum albumin (BSA) as standard. Equal amounts
of proteins (20 lg/condition) were resolved in 12–15% sodium dodecyl sulfate
gel electrophoresis and transferred to polyvinylidene difluoride (PVDF)
membranes according to conventional partially modified methods (Fuentes
et al., 2000). Briefly, proteins were transferred (250 mA for 60 min) to PVDF
membranes using Mini Trans-Blot Cell apparatus (Bio-Rad, Hercules, CA). The
procedure for immunodetection includes the transfer and blocking of the
membrane (60 min at room temperature) with Tween–Tris-buffered saline
(TTBS) (10mM Tris/HCl pH 7.5, 150mM NaCl, and 0.2% Tween-20) containing
10% nonfat-dried milk. Membranes were then incubated overnight at 4�C with
primary antibodies, p-mTORser2448, and cleaved caspase-3 from Cell Signal-
ling (Beverly, MA) and anti-LC3 from MBL Medical and Biological Labora-
tories Co., Ltd (Nagoya, Japan), all of them diluted 1:1000 in TTBS þ 10%
nonfat-dried milk or 5% BSA. After washing (for two 5-min periods with TTBS),
membranes were incubated (60 min at room temperature) with peroxidase-
conjugated secondary antibodies (1:5000 in TTBS with 10% nonfat-dried milk).
After washing (for two 5-min periods and one 10-min period), the detection of
bound antibodies was visualized by chemiluminiscence using the ECL-plus
reagent (GE Healthcare, Buckinghamshire, UK). Actin content was analyzed as
a control by means of a rabbit polyclonal antibody (from Sigma).
Analysis of protein degradation. Human neuroblastoma SH-SY5Y cells
were incubated with 0.2 mCi/ml. L-[14C]valine (GE Healthcare) in complete
medium (CM) for 24 h at 37�C. Unincorporated radioisotope was removed by
washing the cells three times with PBS (pH 7.4). Cells were then incubated in
Earle’s balanced salt solution buffer plus 0.1% BSA (nutrient-free [NF]
medium) in the presence of 10mM cold valine, for 1 h (prechase period).
After this time, the medium was replaced by the appropriate fresh medium (NF
or CM) plus cold valine 10mM in the presence or absence of 10mM 3-MA and
10lM PQ for 4 h (chase period). Cells and radiolabeled proteins from the
medium were precipitated in trichloroacetic acid at the final concentration of
10% (vol/vol). The precipitated proteins were separated from the soluble
radioactivity by centrifugation at 600 3 g for 20 min and then dissolved in 1 ml
of 0.2 N NaOH. The rate of protein degradation was calculated by determining
the ratio of acid-soluble radioactivity recovered from both cells and medium to
the ratio of radioactivity in trichloroacetic acid-precipitated proteins obtained
from both cells and medium (Pattingre et al., 2003).
Electron microscopy. Scraped cells were spinned and immersed in
a mixture of 2% glutaraldehyde and 2% paraformaldehyde in phosphate buffer
(pH 7.4) for 1 h at 4�C. Pellets were then rinsed in phosphate buffer, postfixed in
1% osmium tetroxide for 90 min, rinsed, dehydrated in a graded series of
acetone, and embedded in Araldite resin. Ultrathin 90-nm–thick sections were
obtained using a Leica Reichert Ultracut ultramicrotome equipped with a glass
knife. Sections were mounted on copper grids, stained with uranyl acetate and
lead citrate, examined, and photographed with a Jeol JEM-100 CX II
transmission electron microscope.
Transient transfections. SH-SY5Y cells were transiently transfected with
the Fugene procedure, with 1–2 lg DNA per 35-mm plate, according to the
manufacturer’s protocol (Roche Diagnostics, GmbH, Mannheim, Germany) to
label AVs with LC3-GFP plasmid (Kabeya et al., 2000).
PARAQUAT INDUCES AUTOPHAGY 449
by guest on October 28, 2014
http://toxsci.oxfordjournals.org/D
ownloaded from

Statistical analysis. Data are representative of at least three independent
experiments each in triplicate determination. Statistical analysis of the data was
performed using one-way ANOVA. Significance (*) was defined at p < 0.05.
RESULTS
PQ Induces Apoptotic Death in Neuroblastoma Cells
As previously described (Chun et al., 2001; Gonzalez-Poloet al., 2004; Niso-Santano et al., 2006), several cell lines,
including human neuroblastoma SH-SY5Y cells, are sensitiveto the PQ induced toxicity. We investigated, for the first time,the cell death type in neuroblastoma cells exposed to thebipiridinic pesticide PQ. As we show in Figure 1, the PQtreatment caused phosphatidylserine exposure on the outerleaflet of the plasma membrane, which can manifest beforeplasma membrane permeabilization. This phenomenon wasincreased in combination with PQ and starvation conditions.The cells treated with 10–25lM of PQ exhibited a range of25–50% loss of cell viability in CM and NF medium. In parallel
FIG. 1. PQ-mediated sensitization of SH-SY5Y cells to starvation-induced cell death. The human neuroblastoma SH-SY5Y cells were cultured in the
indicated conditions (CM, NF), in the absence (Co) or in the presence of the indicated doses of PQ for 24 h, followed by simultaneous staining of the
phosphatidylserine exposure with Annexin-V fluorescein isothiocyanate (FITC) and viability with PI. Quantitative data (error bars are SE n¼ 3) are shown in (A),
and representative FACS diagrams are shown in (B).
450 GONZALEZ-POLO ET AL.
by guest on October 28, 2014
http://toxsci.oxfordjournals.org/D
ownloaded from
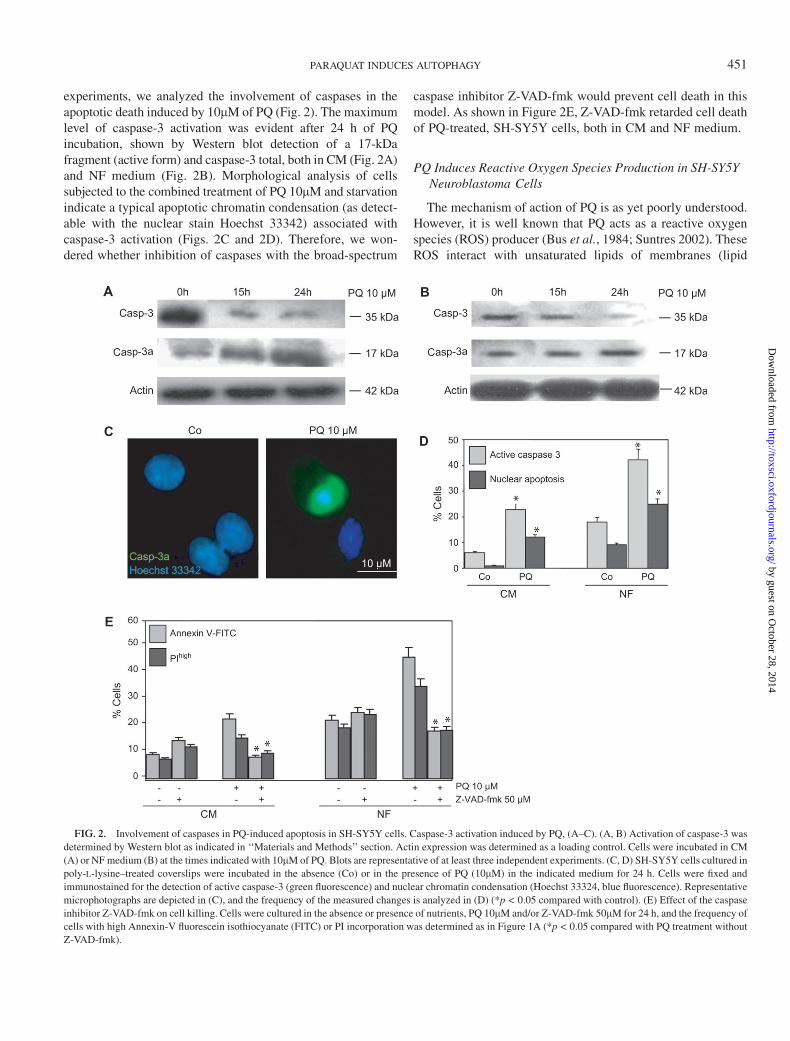
experiments, we analyzed the involvement of caspases in theapoptotic death induced by 10lM of PQ (Fig. 2). The maximumlevel of caspase-3 activation was evident after 24 h of PQincubation, shown by Western blot detection of a 17-kDafragment (active form) and caspase-3 total, both in CM (Fig. 2A)and NF medium (Fig. 2B). Morphological analysis of cellssubjected to the combined treatment of PQ 10lM and starvationindicate a typical apoptotic chromatin condensation (as detect-able with the nuclear stain Hoechst 33342) associated withcaspase-3 activation (Figs. 2C and 2D). Therefore, we won-dered whether inhibition of caspases with the broad-spectrum
caspase inhibitor Z-VAD-fmk would prevent cell death in thismodel. As shown in Figure 2E, Z-VAD-fmk retarded cell deathof PQ-treated, SH-SY5Y cells, both in CM and NF medium.
PQ Induces Reactive Oxygen Species Production in SH-SY5YNeuroblastoma Cells
The mechanism of action of PQ is as yet poorly understood.However, it is well known that PQ acts as a reactive oxygenspecies (ROS) producer (Bus et al., 1984; Suntres 2002). TheseROS interact with unsaturated lipids of membranes (lipid
FIG. 2. Involvement of caspases in PQ-induced apoptosis in SH-SY5Y cells. Caspase-3 activation induced by PQ, (A–C). (A, B) Activation of caspase-3 was
determined by Western blot as indicated in ‘‘Materials and Methods’’ section. Actin expression was determined as a loading control. Cells were incubated in CM
(A) or NF medium (B) at the times indicated with 10lM of PQ. Blots are representative of at least three independent experiments. (C, D) SH-SY5Y cells cultured in
poly-L-lysine–treated coverslips were incubated in the absence (Co) or in the presence of PQ (10lM) in the indicated medium for 24 h. Cells were fixed and
immunostained for the detection of active caspase-3 (green fluorescence) and nuclear chromatin condensation (Hoechst 33324, blue fluorescence). Representative
microphotographs are depicted in (C), and the frequency of the measured changes is analyzed in (D) (*p < 0.05 compared with control). (E) Effect of the caspase
inhibitor Z-VAD-fmk on cell killing. Cells were cultured in the absence or presence of nutrients, PQ 10lM and/or Z-VAD-fmk 50lM for 24 h, and the frequency of
cells with high Annexin-V fluorescein isothiocyanate (FITC) or PI incorporation was determined as in Figure 1A (*p < 0.05 compared with PQ treatment without
Z-VAD-fmk).
PARAQUAT INDUCES AUTOPHAGY 451
by guest on October 28, 2014
http://toxsci.oxfordjournals.org/D
ownloaded from

peroxidation) and destroy organelles subsequently leading tocell death (Dodge, 1971). In this sense, several studies havesuggested the involvement of ROS in the toxicity induced byPQ, including studies in vitro in several cell lines as in humanlung epithelial cells (Cappelletti et al., 1998) or in cerebellargranule cells (Bus et al., 1984; Gonzalez-Polo et al., 2004),as well as studies in vivo (Mollace et al., 2003). In our study, weshowed that exposure to low concentrations of PQ (10lM) for24 h caused an increase of intracellular levels of ROS in humanneuroblastoma SH-SY5Y cells (as measured by the oxidationof the nonfluorescent dye HE into its fluorescent productethidium [Eth]) (Fig. 3). This production was enhanced whenthe cells were treated with a combination of nutrient depletionconditions and PQ. The ROS generation was abolished inthe presence of several antioxidants as N-acetylcysteine as wellas the ROS scavenger vitamin E (a-tocopherol) (data notshown).
The Treatment of SH-SY5Y Neuroblastoma Cells withLow Concentrations of PQ Produces Characteristicsof Autophagy
It has been described that autophagy is a cellular response toextracellular (nutrient deprivation, hypoxia) and intracellularstress conditions (accumulation of damaged organelles andcytoplasmic components). This phenomenon is characterizedmorphologically by the accumulation of AVs in the cytoplasm.
We have tested in our model that, in NF medium, that is, in theabsence of serum and amino acids, as well as in CM, controlcells did not develop any discernible vacuoles, while cellstreated with PQ 10lM exhibited an increase in cytoplasmicvacuolization visible as ‘‘holes’’ in CMFDA-stained cells ofa time-dependent way (Figs. 4A and 4B). Moreover, thesevacuoles induced in the presence of PQ showed, clearly, thecharacteristic double membranes of AVs as visible by trans-mission electron microscopy (Fig. 4C). Figures 4A and 4Cshow images in CM conditions. Microphotographs in NFconditions are equivalents (data not shown).
Similarly, after treatment of serum and amino acid starva-tion, 10lM of PQ induced the recruitment of a LC3-GFP fusionprotein to AV in discrete foci (Kabeya et al., 2000; Mizushima2004) (Figs. 5A and 5B). In agreement with the microscopicobservations, we detected by Western blotting the accumula-tion of the autophagosome marker LC3-II (Boya et al., 2005;Kabeya et al., 2000) both in CM (Fig. 5C) and under theconditions of starvation (data not shown) in SH-SY5Y treatedwith PQ 10lM. Furthermore, as compared to the prototypicautophagy inhibitor 3-MA, PQ (10lM) induced the autophagy(as determined by quantifying the turnover of proteins instarved cells) as shown in Figure 6A. In this sense, exposure ofhuman neuroblastoma cells with 10lM of PQ produceda marked decrease in the phosphorylation (Ser2448) status ofmTOR (Figs. 6B and 6C), protein involved in the autophagysignaling, with a maximal effect after 30 min of herbicideexposure in CM (Fig. 6B). In starved cells incubated with PQ,the activation of phosphorylated mTOR was invaluable to allthe tested times (Fig. 6C).
The Inhibition of PQ-Induced Autophagy Acceleratesthe Apoptotic Cell Death in SH-SY5Y Cells
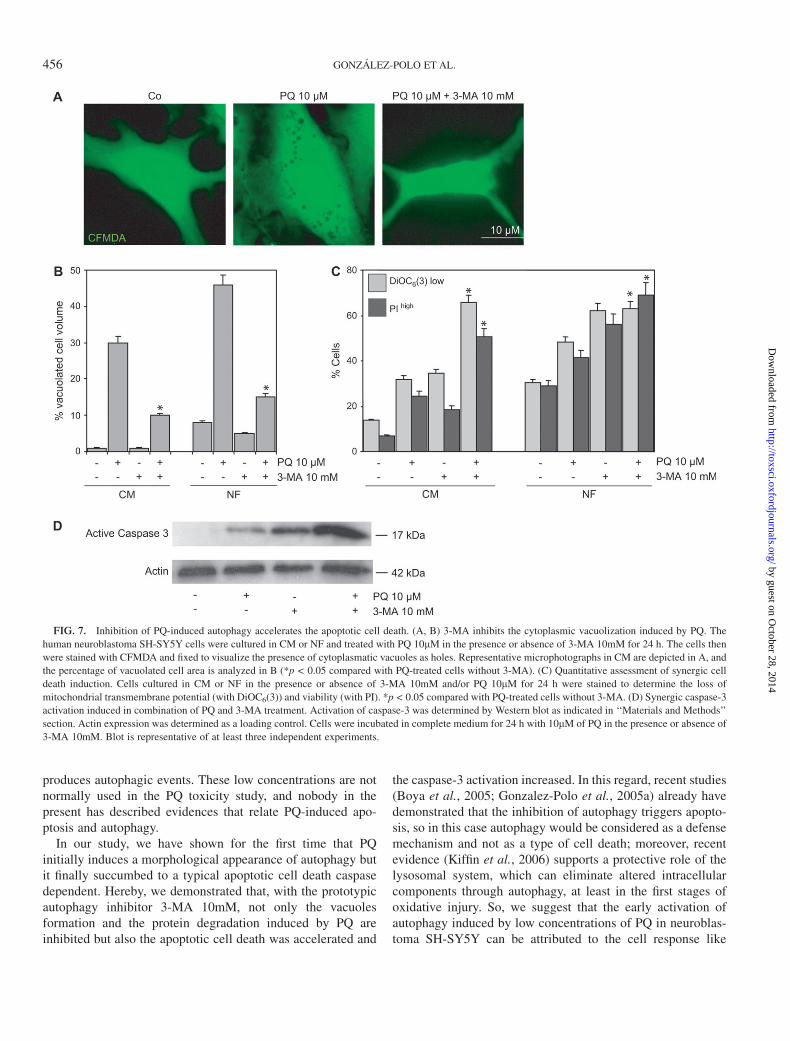
We investigated the possible connection between the accu-mulation of autophagic phenomenon and the apoptotic celldeath induced by PQ in SH-SY5Y cells. The morphologicalanalysis (Figure 7A) showed that the prototypic autophagyinhibitor 3-MA inhibited the PQ-induced autophagic vacuoli-zation as well as protein degradation, already previouslymentioned in Figure 6A. Moreover, cytofluorometric quantifi-cation revealed that a significant fraction of cells treated with3-MA and/or PQ lost the capacity to retain the dye DiOC6(3)and hence dissipated the mitochondrial transmembrane poten-tial (Dwm). Among this Dwm low population, a fraction of cellsincorporated the vital dye PI and thus lost the barrier functionof their plasma membrane (Fig. 7B). This event was moreevident in combination of 3-MA and the herbicide PQ. In thesame way, the caspase-3 activation, as measured by Westernblot detection of 17-kDa fragment (Fig. 7C), was moreincreased with PQ and 3-MA together that each one separately.Therefore, the combination of these results clearly suggeststhat the inhibition of PQ-induced autophagy accelerates theapoptotic cell death in SH-SY5Y cells.
FIG. 3. PQ-induced formation of superoxide. SH-SY5Y cells were
incubated in the indicated medium, CM or NF, in the absence (Co) or in the
presence of PQ (10lM) for 24 h followed by quantitation of the frequency of
cells that exhibit oxidation of HE into the fluorescent Eth. All results are
representative of at least three independent determinations. Numbers in FACS
diagrams refer to the percentage of cells that exhibit oxidation of HE into the
fluorescent Eth.
452 GONZALEZ-POLO ET AL.
by guest on October 28, 2014
http://toxsci.oxfordjournals.org/D
ownloaded from

DISCUSSION
Several studies have shown the possibility that environmen-tal neurotoxicants such as pesticides may be related to the
development of nongenetic PD (Cory-Slechta et al., 2005;Landrigan et al., 2005; Norris et al., 2004; Ritz et al., 2000;Sherer et al., 2001). PQ is one of the possible pesticidesinvolved in PD genesis because of the similar chemical
FIG. 4. PQ induces accumulation of AVs. (A, B) SH-SY5Y cells were cultured in CM or NF in the presence of PQ 10lM the times indicated. The presence of
cytoplasmic vacuoles was determined by staining with CFMDA as indicated in ‘‘Materials and Methods’’ section. Cytoplasmic vacuoles are visualized as holes.
Representative microphotographs in CM are depicted in (A), and the percentage of vacuolated cell area is analyzed in (B) (error bars are SE n ¼ 3).
(C) Ultrastructure of PQ-treated SH-SY5Y cells. The cells were treated as in (A). Electron microscopy of representative cells are shown.
PARAQUAT INDUCES AUTOPHAGY 453
by guest on October 28, 2014
http://toxsci.oxfordjournals.org/D
ownloaded from

structure to the dopaminergic neurotoxicant MPPþ and thestrong correlation between the incidence of the disease and theamount of PQ used (Ritz et al., 2000). In the present study wereport, for the first time, that low concentrations of PQ inducemolecular events compatible with autophagy in neuroblastomaSH-SY5Y cells. We also show that the inhibition of autophagyexacerbates PQ-induced apoptosis.
The human dopaminergic cell line SH-SY5Y, which hasmany qualities of nervous system neurons, was used as targetneuronal cells. First, we confirmed that PQ induced neurotox-icity in SH-SY5Y cells in a dose-dependent manner both inCM and in starvation conditions (Fig. 1). This result wassimilar to the results reported by Gonzalez-Polo et al. (2004)and Yang et al. (2005). Our group found that PQ induced dose-dependent cell death and apoptosis in SH-SY5Y cells withina period of 24 h. PQ is a well-known oxidative-stress inducer(Gonzalez-Polo et al., 2004; Mollace et al., 2003), so most ofthe reports about it are focused on ROS production. In thissense, we show in our study that low concentrations of PQ(10lM) induce a robust ROS generation (Fig. 3), which is moreevident in NF conditions. However, the major finding of thisstudy is that low PQ concentrations produce characteristics ofautophagy.
Autophagy was described for the first time by Schweichelet al. (1973). It is characterized by the appearance of numerouscytoplasmic AVs of lysosomal origin, followed by mitochon-drial dilation and enlargement of the endoplasmic reticulum
and the Golgi apparatus. Dysfunction of cellular degradationhas been implicated in PD pathogenesis. Macroautophagy isthe primary mechanism of degrading long-lived proteins anddamaged organelles. The autophagic process begins with theremodeling of subcellular membranes into double-membranevesicles that sequester cytoplasm and organelles in AVs. Thesevacuoles are fused with the lysosome to form the autophago-lysosome. The content of the autophagolysosome are ulti-mately degraded by lysosomal degradative enzymes (Klionskyet al., 2000). The electron microscopy images in Figure 4 showthat PQ induces the accumulation of double-membrane AVs inthe cytoplasm. This event is accompanied by other autophagiccharacteristics as accumulation of the autophagosome markerLC3-II (Fig. 5) or protein degradation regulated negatively bythe mTOR kinase signaling (Fig. 6). In this sense, it has beendemonstrated that the inactivation of mTOR in yeast ormammalian cells leads to induction of autophagy, even innutrient-rich medium, indicating that mTOR negatively con-trols starvation-induced autophagy (Blommaart et al., 1995).So, under unfavorable conditions, mTOR is inactive, leading toa reduction in protein synthesis and an upregulation of proteindegradation (Dennis et al., 1999). Thus, we report here that PQis involved in the control of autophagy in neuroblastomaSH-SY5Y cells. So, one possibility is that the cell treated withPQ triggers autophagy as a defense mechanism to degrade theoxidized proteins by ROS and damaged or dysfunctionalcytoplasmic components.
FIG. 5. Effect of PQ on the subcellular localization and status of the AV marker LC3. (A, B) Redistribution of LC3-GFP. The cells were transfected with an
LC3-GFP fusion construct as indicated in ‘‘Materials and Methods’’ section. Twenty-four hours after transient transfection, cells were incubated in the indicated
medium, CM or NF, treated in the absence (Co) or presence of PQ 10lM for 15 h and fixed. Representative fluorescence microphotographs are shown in (A), and
the frequency of cells exhibiting the accumulation of LC3-GFP in vacuoles was quantified (*p < 0.05 compared with control) in (B). (C) Immunoblot analyses of
accumulating LC3-II protein in cells treated with PQ 10lM the indicated times. Actin expression was determined as a loading control in the same conditions. Blots
are representative of at least three independent experiments.
454 GONZALEZ-POLO ET AL.
by guest on October 28, 2014
http://toxsci.oxfordjournals.org/D
ownloaded from

The pesticide induced, in the first place, the morphologicalappearance of autophagy (Figs. 4–6). But finally the cellssuccumbed in morphological and biochemical changes that aretypical of apoptotic cell death, as phosphatidylserine exposure(Fig. 1), Dwm dissipation (Fig. 7), caspase-3 activation (Fig. 2),and chromatin condensation (Fig. 2). Interestingly, inhibitionof caspases retarded cell death (Fig. 2), yet had no influence onautophagic vacuolization (data not shown). In this sense, thereis some controversy in the bibliography regarding the preciserole that autophagy might play in programed cell death. Thereare many reports describing situations in which autophagyactivation accompanies apoptosis, but there are also cases inwhich autophagy enhancement leads to cell death in theabsence of caspase activation (Bursch et al., 1996; Butleret al., 2000; Chi et al., 1999; Xiang et al., 1996). Moreover,autophagic cell death has been described in association withseveral neurodegenerative diseases, including PD (Angladeet al., 1997; Marino et al., 2004; Petersen et al., 2001).Increased levels of autophagy were observed in neuronal cell
lines expressing mutant proteins associated with such diseases.Expression of PD-associated A53T, but not wild-type,a-synuclein in PC12 cells induced massive AV formation(Stefanis et al., 2001). In addition, dopaminergic neurons withAVs have been observed in the substantia nigra of PD patients,along with neurons displaying apoptotic features (Angladeet al., 1997). However, the functional consequence of increasedautophagic activity for neurodegeneration is not clear but hasbeen suggested that excessive autophagy may directly lead toneuronal cell death (Yuan et al., 2003).
In the literature, concentrations of high micromolar ormillimolar range of PQ are normally used to study itsneurotoxicity and apoptosis induction (Gonzalez-Polo et al.,2004; Kim et al., 2004; Peng et al., 2004). The evidence thatPQ triggers apoptotic cell death has been demonstrated in othercellular models, as in human lung epithelial cells (Cappellettiet al., 1998) as well as in PC12 cells (Li et al., 1999). Ourexperiments show that PQ induces apoptosis in SH-SY5Y cellsat low concentrations (10lM) at the same concentrations that
FIG. 6. Study of the protein degradation and changes of mTOR phosphorylation in the PQ-mediated neurotoxicity. (A) The protein half-life was determined by
pulse chasing with radioactive valine as indicated in ‘‘Materials and Methods’’ section. The cells were incubated in CM or NF and treated with PQ 10lM in the
presence or absence of 3-MA 10mM for 6 h. *p < 0.05 compared with control. (B, C) mTOR dephosphorylation induced by PQ. The SH-SY5Y cells were
incubated with 10lM of PQ the indicated times in CM (B) or in NF medium (C). Cell lysates were analyzed for mTOR activation by Western blotting using
a phospho-specific mTOR (ser2448) antibody as indicated in ‘‘Materials and Methods’’ section. Actin expression was determined as a loading control in the same
conditions. Blots are representative of at least three independent experiments.
PARAQUAT INDUCES AUTOPHAGY 455
by guest on October 28, 2014
http://toxsci.oxfordjournals.org/D
ownloaded from
produces autophagic events. These low concentrations are notnormally used in the PQ toxicity study, and nobody in thepresent has described evidences that relate PQ-induced apo-ptosis and autophagy.
In our study, we have shown for the first time that PQinitially induces a morphological appearance of autophagy butit finally succumbed to a typical apoptotic cell death caspasedependent. Hereby, we demonstrated that, with the prototypicautophagy inhibitor 3-MA 10mM, not only the vacuolesformation and the protein degradation induced by PQ areinhibited but also the apoptotic cell death was accelerated and
the caspase-3 activation increased. In this regard, recent studies(Boya et al., 2005; Gonzalez-Polo et al., 2005a) already havedemonstrated that the inhibition of autophagy triggers apopto-sis, so in this case autophagy would be considered as a defensemechanism and not as a type of cell death; moreover, recentevidence (Kiffin et al., 2006) supports a protective role of thelysosomal system, which can eliminate altered intracellularcomponents through autophagy, at least in the first stages ofoxidative injury. So, we suggest that the early activation ofautophagy induced by low concentrations of PQ in neuroblas-toma SH-SY5Y can be attributed to the cell response like
FIG. 7. Inhibition of PQ-induced autophagy accelerates the apoptotic cell death. (A, B) 3-MA inhibits the cytoplasmic vacuolization induced by PQ. The
human neuroblastoma SH-SY5Y cells were cultured in CM or NF and treated with PQ 10lM in the presence or absence of 3-MA 10mM for 24 h. The cells then
were stained with CFMDA and fixed to visualize the presence of cytoplasmatic vacuoles as holes. Representative microphotographs in CM are depicted in A, and
the percentage of vacuolated cell area is analyzed in B (*p < 0.05 compared with PQ-treated cells without 3-MA). (C) Quantitative assessment of synergic cell
death induction. Cells cultured in CM or NF in the presence or absence of 3-MA 10mM and/or PQ 10lM for 24 h were stained to determine the loss of
mitochondrial transmembrane potential (with DiOC6(3)) and viability (with PI). *p < 0.05 compared with PQ-treated cells without 3-MA. (D) Synergic caspase-3
activation induced in combination of PQ and 3-MA treatment. Activation of caspase-3 was determined by Western blot as indicated in ‘‘Materials and Methods’’
section. Actin expression was determined as a loading control. Cells were incubated in complete medium for 24 h with 10lM of PQ in the presence or absence of
3-MA 10mM. Blot is representative of at least three independent experiments.
456 GONZALEZ-POLO ET AL.
by guest on October 28, 2014
http://toxsci.oxfordjournals.org/D
ownloaded from

a defense mechanism against the rapid oxidative stress inducedafter PQ exposure. Our results indicated that the systems ofdefense of the cell seemed not to be sufficient because the cellfinally succumbed to the apoptotic cell death. In addition, wehave seen for the first time in our study that when we inhibitedPQ-induced autophagy in presence of 3-MA, we acceleratedthe apoptotic death process.
In conclusion, our results help to suggest PQ neurotoxicity asan emerging PD model system. This study also provides newmolecular bases for the knowledge of the relationship betweenautophagy and the cell response to stress and apoptosis celldeath. However, further studies are required to elucidate theexact mechanism involved in PQ-induced autophagy and itsrelation with neuronal cell death.
ACKNOWLEDGMENTS
Supported by grants 2PR04B002, 2PR06B124, and 3PR05C015 (Junta de
Extremadura, Spain) and PI040828 (Fondo de Investigacion Sanitaria, Ministerio
de Sanidad y Consumo, Spain). R.A.G.P. was supported by the Junta de
Extremadura re-incorporation fellowship. M.N.S. was supported by the Valhon-
do-Calaff Foundation fellowship. M.A.O. was supported by the Junta of
Extremadura predoctoral fellowship. The authors would like to thank P. Delgado,
R. Tarazona, and V. Llorente Vera for their invaluable technical assistance. We
also thank Dr Guido Kroemer (Institute Gustave Roussy, Villejuif, France) and
Tamotsu Yoshimori (National Institute of Genetics, Mishima, Japan) for regeants.
REFERENCES
Anglade, P., Vyas, S., Javoy-Agid, F., Herrero, M. T., Michel, P. P., Marquez, J.,
Mouatt-Prigent, A., Ruberg, M., Hirsch, E. C., and Agid, Y. (1997).
Apoptosis and autophagy in nigral neurons of patients with Parkinson’s
disease. Histol. Histopathol. 12, 25–31.
Blommaart, E. F., Luiken, J. J., Blommaart, P. J., van Woerkom, G. M., and
Meijer, A. J. (1995). Phosphorylation of ribosomal protein S6 is inhibitory
for autophagy in isolated rat hepatocytes. J. Biol. Chem. 270, 2320–2326.
Boya, P., Gonzalez-Polo, R. A., Casares, N., Perfettini, J. L., Dessen, P.,
Larochette, N., Metivier, D., Meley, D., Souquere, S., Yoshimori, T., et al.
(2005). Inhibition of macroautophagy triggers apoptosis. Mol. Cell Biol. 25,
1025–1040.
Boya, P., Gonzalez-Polo, R. A., Poncet, D., Andreau, K., Vieira, H. L.,
Roumier, T., Perfettini, J. L., and Kroemer, G. (2003). Mitochondrial
membrane permeabilization is a critical step of lysosome-initiated apoptosis
induced by hydroxychloroquine. Oncogene 22, 3927–3936.
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein-dye
binding. Anal. Biochem. 72, 248–254.
Bursch, W., Ellinger, A., Kienzl, H., Torok, L., Pandey, S., Sikorska, M.,
Walker, R., and Hermann, R. S. (1996). Active cell death induced by the anti-
estrogens tamoxifen and ICI 164 384 in human mammary carcinoma cells
(MCF-7) in culture: The role of autophagy. Carcinogenesis 17, 1595–1607.
Bursch, W., Hochegger, K., Torok, L., Marian, B., Ellinger, A., and Hermann,
R. S. (2000). Autophagic and apoptotic types of programmed cell death
exhibit different fates of cytoskeletal filaments. J. Cell Sci. 113(Pt 7),
1189–1198.
Bus, J. S. and Gibson, J. E. (1984). Paraquat: Model for oxidant-initiated
toxicity. Environ. Health Perspect. 55, 37–46.
Butler, R., Mitchell, S. H., Tindall, D. J., and Young, C. Y. (2000).
Nonapoptotic cell death associated with S-phase arrest of prostate
cancer cells via the peroxisome proliferator-activated receptor gamma
ligand, 15-deoxy-delta12,14-prostaglandin J2. Cell Growth Differ. 11,
49–61.
Cappelletti, G., Maggioni, M. G., and Maci, R. (1998). Apoptosis in human
lung epithelial cells: Triggering by paraquat and modulation by antioxidants.
Cell Biol. Int. 22, 671–678.
Chi, S., Kitanaka, C., Noguchi, K., Mochizuki, T., Nagashima, Y., Shirouzu, M.,
Fujita, H., Yoshida, M., Chen, W., Asai, A., et al. (1999). Oncogenic Ras
triggers cell suicide through the activation of a caspase-independent cell
death program in human cancer cells. Oncogene 18, 2281–2290.
Chun, H. S., Gibson, G. E., DeGiorgio, L. A., Zhang, H., Kidd, V. J., and Son,
J. H. (2001). Dopaminergic cell death induced by MPP(þ), oxidant
and specific neurotoxicants shares the common molecular mechanism.
J. Neurochem. 76, 1010–1021.
Cory-Slechta, D. A., Thiruchelvam, M., Barlow, B. K., and Richfield, E. K.
(2005). Developmental pesticide models of the Parkinson disease phenotype.
Environ. Health Perspect. 113, 1263–1270.
Dennis, P. B., Fumagalli, S., and Thomas, G. (1999). Target of rapamycin
(TOR): Balancing the opposing forces of protein synthesis and degradation.
Curr. Opin. Genet. Dev. 9, 49–54.
Di Monte, D. A., McCormack, A., Petzinger, G., Janson, A. M., Quik, M., and
Langston, W. J. (2000). Relationship among nigrostriatal denervation,
parkinsonism, and dyskinesias in the MPTP primate model. Mov. Disord.
15, 459–466.
Dodge, A. D. (1971). The mode of action of the bipyridylium herbicides,
paraquat and diquat. Endeavour 30, 130–135.
Filonova, L. H., Bozhkov, P. V., Brukhin, V. B., Daniel, G., Zhivotovsky, B., and
von Arnold, S. (2000). Two waves of programmed cell death occur during
formation and development of somatic embryos in the gymnosperm, Norway
spruce. J. Cell Sci. 113(Pt 24), 4399–4411.
Fuentes, J. M., Lompre, A. M., Moller, J. V., Falson, P., and le Maire, M. (2000).
Clean Western blots of membrane proteins after yeast heterologous
expression following a shortened version of the method of Perini et al.
Anal. Biochem. 285, 276–278.
Gonzalez-Polo, R. A., Boya, P., Pauleau, A. L., Jalil, A., Larochette, N.,
Souquere, S., Eskelinen, E. L., Pierron, G., Saftig, P., and Kroemer, G.
(2005a). The apoptosis/autophagy paradox: Autophagic vacuolization before
apoptotic death. J. Cell Sci. 118, 3091–3102.
Gonzalez-Polo, R. A., Carvalho, G., Braun, T., Decaudin, D., Fabre, C.,
Larochette, N., Perfettini, J. L., Djavaheri-Mergny, M., Youlyouz-Marfak, I.,
Codogno, P., et al. (2005b). PK11195 potently sensitizes to apoptosis
induction independently from the peripheral benzodiazepin receptor. Onco-
gene 24, 7503–7513.
Gonzalez-Polo, R. A., Rodriguez-Martin, A., Moran, J. M., Niso, M., Soler, G.,
and Fuentes, J. M. (2004). Paraquat-induced apoptotic cell death in
cerebellar granule cells. Brain Res. 1011, 170–176.
Graeber, M. B., and Moran, L. B. (2002). Mechanisms of cell death in
neurodegenerative diseases: Fashion, fiction, and facts. Brain Pathol. 12,
385–390.
Hariri, M., Millane, G., Guimond, M. P., Guay, G., Dennis, J. W., and Nabi, I. R.
(2000). Biogenesis of multilamellar bodies via autophagy. Mol. Biol. Cell 11,
255–268.
Hertzman, C., Wiens, M., Bowering, D., Snow, B., and Calne, D. (1990).
Parkinson’s disease: A case-control study of occupational and environmental
risk factors. Am. J. Ind. Med. 17, 349–355.
Kabeya, Y., Mizushima, N., Ueno, T., Yamamoto, A., Kirisako, T., Noda, T.,
Kominami, E., Ohsumi, Y., and Yoshimori, T. (2000). LC3, a mammalian
homologue of yeast Apg8p, is localized in autophagosome membranes after
processing. EMBO J. 19, 5720–5728.
PARAQUAT INDUCES AUTOPHAGY 457
by guest on October 28, 2014
http://toxsci.oxfordjournals.org/D
ownloaded from

Kiffin, R., Bandyopadhyay, U., and Cuervo, A. M. (2006). Oxidative stress and
autophagy. Antioxid. Redox Signal. 8, 152–162.
Kim, S. J., Kim, J. E., and Moon, I. S. (2004). Paraquat induces apoptosis of
cultured rat cortical cells. Mol. Cells 17, 102–107.
Klionsky, D. J., and Emr, S. D. (2000). Autophagy as a regulated pathway of
cellular degradation. Science 290, 1717–1721.
Lai, C. T., and Yu, P. H. (1997). Dopamine- and L-beta-3,4-dihydroxypheny-
lalanine hydrochloride (L-Dopa)-induced cytotoxicity towards catechol-
aminergic neuroblastoma SH-SY5Y cells. Effects of oxidative stress and
antioxidative factors. Biochem. Pharmacol. 53, 363–372.
Landles, C., and Bates, G. P. (2004). Huntingtin and the molecular pathogenesis
of Huntington’s disease. Fourth in molecular medicine review series. EMBO
Rep. 5, 958–963.
Landrigan, P. J., Sonawane, B., Butler, R. N., Trasande, L., Callan, R., and
Droller, D. (2005). Early environmental origins of neurodegenerative disease
in later life. Environ. Health Perspect. 113, 1230–1233.
Langston, J. W., and Ballard, P. (1984). Parkinsonism induced by 1-methyl-4-
phenyl-1,2,3,6-tetrahydropyridine (MPTP): Implications for treatment and
the pathogenesis of Parkinson’s disease. Can. J. Neurol. Sci. 11, 160–165.
Lee, H. S., Park, C. W., and Kim, Y. S. (2000). MPP(þ) increases the
vulnerability to oxidative stress rather than directly mediating oxidative
damage in human neuroblastoma cells. Exp. Neurol. 165, 164–171.
Li, X., and Sun, A. Y. (1999). Paraquat induced activation of transcription
factor AP-1 and apoptosis in PC12 cells. J. Neural Transm. 106, 1–21.
Liou, H. H., Tsai, M. C., Chen, C. J., Jeng, J. S., Chang, Y. C., Chen, S. Y., and
Chen, R. C. (1997). Environmental risk factors and Parkinson’s disease:
A case-control study in Taiwan. Neurology 48, 1583–1588.
Marino, G., and Lopez-Otin, C. (2004). Autophagy: Molecular mechanisms,
physiological functions and relevance in human pathology. Cell Mol. Life
Sci. 61, 1439–1454.
Mizushima, N. (2004). Methods for monitoring autophagy. Int. J. Biochem.
Cell Biol. 36, 2491–2502.
Mollace, V., Iannone, M., Muscoli, C., Palma, E., Granato, T., Rispoli, V.,
Nistico, R., Rotiroti, D., and Salvemini, D. (2003). The role of oxidative
stress in paraquat-induced neurotoxicity in rats: Protection by non peptidyl
superoxide dismutase mimetic. Neurosci. Lett. 335, 163–166.
Niso-Santano, M., Moran, J. M., Garcia-Rubio, L., Gomez-Martin, A.,
Gonzalez-Polo, R. A., Soler, G., and Fuentes, J. M. (2006). Low concen-
trations of paraquat induces early activation of extracellular signal-regulated
kinase 1/2, protein kinase B, and c-jun N-terminal kinase 1/2 pathways: Role
of c-jun N-terminal kinase in paraquat-induced cell death. Toxicol. Sci. 92,
507–515.
Norris, E. H., Giasson, B. I., and Lee, V. M. (2004). Alpha-synuclein: Normal
function and role in neurodegenerative diseases. Curr. Top. Dev. Biol. 60,
17–54.
Pattingre, S., Bauvy, C., and Codogno, P. (2003). Amino acids interfere with
the ERK1/2-dependent control of macroautophagy by controlling the activa-
tion of Raf-1 in human colon cancer HT-29 cells. J. Biol. Chem. 278,
16667–16674.
Peng, J., Mao, X. O., Stevenson, F. F., Hsu, M., and Andersen, J. K. (2004). The
herbicide paraquat induces dopaminergic nigral apoptosis through sustained
activation of the JNK pathway. J. Biol. Chem. 279, 32626–32632.
Petersen, A., Larsen, K. E., Behr, G. G., Romero, N., Przedborski, S., Brundin, P.,
and Sulzer, D. (2001). Expanded CAG repeats in exon 1 of the Huntington’s
disease gene stimulate dopamine-mediated striatal neuron autophagy and
degeneration. Hum. Mol. Genet. 10, 1243–1254.
Ritz, B., and Yu, F. (2000). Parkinson’s disease mortality and pesticide
exposure in California 1984–1994. Int. J. Epidemiol. 29, 323–329.
Sattler, T., and Mayer, A. (2000). Cell-free reconstitution of microautophagic
vacuole invagination and vesicle formation. J. Cell Biol. 151, 529–538.
Schweichel, J. U., and Merker, H. J. (1973). The morphology of various types
of cell death in prenatal tissues. Teratology 7, 253–266.
Shavali, S., Carlson, E. C., Swinscoe, J. C., and Ebadi, M. (2004). 1-Benzyl-
1,2,3,4-tetrahydroisoquinoline, a Parkinsonism-inducing endogenous toxin,
increases alpha-synuclein expression and causes nuclear damage in human
dopaminergic cells. J. Neurosci. Res. 76, 563–571.
Sherer, T. B., Betarbet, R., and Greenamyre, J. T. (2001). Pathogenesis of
Parkinson’s disease. Curr. Opin. Investig. Drugs 2, 657–662.
Stadelmann, C., Deckwerth, T. L., Srinivasan, A., Bancher, C., Bruck, W.,
Jellinger, K., and Lassmann, H. (1999). Activation of caspase-3 in single
neurons and autophagic granules of granulovacuolar degeneration in
Alzheimer’s disease. Evidence for apoptotic cell death. Am. J. Pathol. 155,
1459–1466.
Stefanis, L., Larsen, K. E., Rideout, H. J., Sulzer, D., and Greene, L. A. (2001).
Expression of A53T mutant but not wild-type alpha-synuclein in PC12
cells induces alterations of the ubiquitin-dependent degradation system,
loss of dopamine release, and autophagic cell death. J. Neurosci. 21,
9549–9560.
Suntres, Z. E. (2002). Role of antioxidants in paraquat toxicity. Toxicology 180,
65–77.
Tanner, C. M., and Ben Shlomo, Y. (1999). Epidemiology of Parkinson’s
disease. Adv. Neurol. 80, 153–159.
Xiang, J., Chao, D. T., and Korsmeyer, S. J. (1996). BAX-induced cell death
may not require interleukin 1 beta-converting enzyme-like proteases. Proc.
Natl. Acad. Sci. U.S.A 93, 14559–14563.
Xue, L., Fletcher, G. C., and Tolkovsky, A. M. (1999). Autophagy is activated
by apoptotic signalling in sympathetic neurons: An alternative mechanism of
death execution. Mol. Cell Neurosci. 14, 180–198.
Yuan, J., Lipinski, M., and Degterev, A. (2003). Diversity in the mechanisms of
neuronal cell death. Neuron 40, 401–413.
458 GONZALEZ-POLO ET AL.
by guest on October 28, 2014
http://toxsci.oxfordjournals.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















