
Bile and pancreatic juice replacement ameliorates early ligation-induced acute pancreatitis in rats
Upload
johnshopkinsCategory
view
4download
0
Dan Berkowitz and Mark RenehanSubroto Chatterjee, Djahida Bedja, Sumita Mishra, Christine Amuzie, Alberto Avolio, David Kass,
Apo E-/- Mice and Rabbits Fed a High Fat and Cholesterol DietInhibition of Glycosphingolipid Synthesis Ameliorates Atherosclerosis and Arterial Stiffness in
Print ISSN: 0009-7322. Online ISSN: 1524-4539 Copyright © 2014 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation published online April 7, 2014;Circulation.
http://circ.ahajournals.org/content/early/2014/04/07/CIRCULATIONAHA.113.007559World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circ.ahajournals.org/content/suppl/2014/04/07/CIRCULATIONAHA.113.007559.DC1.htmlData Supplement (unedited) at:
http://circ.ahajournals.org//subscriptions/
is online at: Circulation Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer available in the
Permissions in the middle column of the Web page under Services. Further information about this process isOnce the online version of the published article for which permission is being requested is located, click Request
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Editorial Office.Circulation Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
1
Inhibition of Glycosphingolipid Synthesis Ameliorates Atherosclerosis and
Arterial Stiffness in Apo E-/- Mice and Rabbits Fed a High Fat and
Cholesterol Diet
Running title: Chatterjee et al.; Glycosphingolipid inhibition ameliorates atherosclerosis
Subroto Chatterjee, PhD1; Djahida Bedja, MS2,3; Sumita Mishra, PhD1; Christine Amuzie, BS1;
Alberto Avolio, PhD3; David Kass, MD2; Dan Berkowitz, MBBCh4; Mark Renehan, BS1
1Dept of Pediatrics; 2Dept of Medicine; 4Dept of Anesthesiology and Critical Care Medicine,
Johns Hopkins University School of Medicine, Baltimore, MD; 3Australian School of Advanced
Medicine, Macquarie University, Sydney, Australia
Address for Correspondence:
Subroto Chatterjee, PhD
Johns Hopkins University Hospital
1383 Blalock Bldg.
600 N Wolfe St.
Baltimore, MD 21287
Tel: 410-614-2518
Fax: 410-955-3247
E-mail: [email protected]
Journal Subject Codes: Vascular biology:[96] Mechanism of atherosclerosis/growth factors, Atherosclerosis:[90] Lipid and lipoprotein metabolism, Treatment:[118] Cardiovascular pharmacology
Alberto Avolio, PhD3; David Kass, MD2; Dan Berkowitz, MBBCh4; Mark RRRenennehehehanann,, BSBSBS11
1DeDeDeptptpt ooof f f PePePediatatatririr ccs; 2Dept of Medicine; 4Depttt oooff f AnA esthesiologygy andndd CrCC itical Care Medicine,
JJJohhnhns Hopkkinins s UnUnUnivvveree sisiitytyty SSSchchchooool l ofofof MMMeededicicicinine, BBBaalltimomomorrere,, MDMDMD;; 3AuAuAustststrarar lliannn SSSchhchooooooll l ofofof AAAdvdvdvananancecc d
MMeMedidicicic nnene,, MaMaMacqcqquuauaririeee UUnUnivivvererrsisitytty,,, SySyydnnneyeyey,, AuAuAuststrraralilia a
AdAdddress ffor CCorrespo dndence:
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
2
Abstract
Background—Glycosphingolipids are integral components of the cell membrane and have been
shown to serve as messengers, transducing growth factor initiated phenotypes. Here we have
examined whether inhibition of glycosphingolipid synthesis could ameliorate atherosclerosis and
arterial stiffness in transgenic mice and rabbits.
Methods and Results—Apolipoprotein E-/- mice (12 weeks of age, n = 6) were fed regular chow or
a western diet (1.25% cholesterol, 2% fat). Mice were fed 5mg/kg (mpk) or 10mpk of an inhibitor of
glycosphingolipid synthesis, D-threo-1-phenyl-2-decanoylamino-3-morpholino-1-propanol
(D-PDMP), solubilized in vehicle (5% Tween-80 in PBS) and the placebo group received vehicle
only. At 20 and 36 weeks of age, serial echocardiography was performed to measure aortic intima
medial thickening (IMT). Aortic pulse wave velocity (PWV) measured vascular stiffness. Feeding
mice a western diet markedly increased aortic PWV, IMT, oxidized LDL, Ca2+ deposits, and
glucosyl- and lactosylceramide synthase activity. These were dose-dependently decreased by
feeding D-PDMP. In liver, D-PDMP decreased cholesterol and triglyceride levels by raising the
expression of SREBP2, LDL-r, HMGCo-A reductase, and cholesterol efflux genes (e.g., ABCG5,
ABCG8). D-PDMP affected VLDL catabolism by increasing the gene expression for LPL and
VLDLr. Rabbits fed a western diet for 90 days had extensive atherosclerosis accompanied by a
17.5-fold increase in total cholesterol levels and a 3-fold increase in lactosylceramide levels. This
was completely prevented by feeding D-PDMP.
Conclusions—Inhibition of glycosphingolipid synthesis ameliorates atherosclerosis and arterial
stiffness in ApoE-/- mice and rabbits. Thus, inhibition of glycosphingolipid synthesis may be a
novel approach to ameliorate atherosclerosis and arterial stiffness.
Key words: APOE, atherosclerosis, imaging, glycosphingolipids, sphingolipids
medial thickening (IMT). Aortic pulse wave velocity (PWV) measured vascular stifffnesess.s Feeedidinng
mice a western diet markedly increased aortic PWV, IMT, oxidized LDL, Ca2+ deppeposssiitss,s, aandndnd
glucosyl- and lactosylceramide synthase activity. These were dose-dependently decreased by
feedinng g D-D PDPDMPP.. InIn liver, D-PDMP decreased cchoholelesterol and triglycereridide levels by raising the
exexxprprpreesessis onn ooof f f SRREBEBP2P2, , LDLDL-L r,r, HHMGMGCoC -AA rredducucttaassee, annd d chcholessteteterrol efefflfluxu ggeneneses ((e.g.g.,, ABA CGCG5,
AABCCCG8). D-PDPDMPMPMP aafffffececctetedd d VLVLVLDLDLL ccaattabbbolisssmmm byy innncreeeaasasiningg ththt e e geeene eeexpppreresssssiioion n fofofor r LPLPPL L L anandd d
VLVLVLDLDLDLr.r. RRababbbibib tsts ffeeded aa wwesesterrnrn dddieiet t fofofor 90900 ddayayayss hahaddd eexexteteennssiviveee aatatheheerorooscsclelerrorosisis s acacccocompmpmpaaanieieedd d byby a
17.55-ffolold inincreae se iin n tototal l chc ololesesteeror l levevelsls aandnd aa 3-folold d inincrcreaeases iin n lalactctosylyly ceeraramimidde lleve elels.s TThiiss
was comppleleteteelylyly ppprereveveentntedeed bbby y fefeededdiining g D-DD PDPDP MPMPMP.
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
3
Introduction
Atherosclerosis contributes to nearly one half of the mortality in the western world and is growing
in epidemic proportions in rapidly developing countries in Asia. High levels of blood cholesterol,
high blood pressure, obesity, diabetes, stress, and lifestyles including smoking are among the risk
factors contributing to this alarming increase in this disease. While several therapeutic modalities
including cholesterol synthesis inhibitors (family of statins), cholesterol absorption inhibitor
(ezetimibe), blood pressure lowering drugs, platelet adhesion inhibitors and many more are either
prescribed alone and/or in combination, there is a need for novel approaches to mitigate the
initiation and progression of atherosclerosis1. However, these therapeutic modalities do not
decrease atherosclerotic plaque burden. A relationship between high blood levels of cholesterol
and glycosphingolipids was suggested several decades ago2-7. Subsequently, a tighter correlation
between the load of LDL cholesterol and GSL/lactosylceramide shedding in urinary proximal
tubular cells in patients with the homozygous form of familial hypercholesterolemia was reported3.
D-PDMP is an analog of glucosylceramide and was synthesized to inhibit glucosylceramide
synthesis in patients with Gaucher’s disease. However, we observed that this compound could
directly inhibit the activity of purified lactosylceramide synthase7. D-PDMP is a small molecular
weight compound and is well tolerated by experimental animals e.g. mice, rats, rabbits up to 10
times the effective dose. In mice, the effective dose of D-PDMP is 10 mg per kg (mpk) body
weight when given orally compared to the use of 100mpk of various adamanate derivatives used
by other investigators8, 9. Since D-PDMP turnover time is a short ~52 min in mice10, it is rapidly
detoxified and excreted with little or no side effects. Delivery of D-PDMP by oral gavage, or
intraperitoneal injection, has no effect on appetite and overall wellbeing of the experimental
animals. We report here that feeding a western diet to ApoE-/- mouse and normal rabbits leads to
decrease atherosclerotic plaque burden. A relationship between high blood levellsss ofoff chohoholelelestststerere ool
and glycosphingolipids was suggested several decades ago2-7. Subsequently, a tighter correlation
beetwtwweeeeeenn thththe e e lloloaddd ooof f LDL cholesterol and GSL/laaacctc ooosylceramide sshehh ddddininingg in urinary proximal
uubuuulal r cells inn ppatatieientntts s wiwiwiththth ttthehhe hhomomomoozzyyggouus ffformmm oof ffafammimililiialall hhyyypeererchchholoo eesesteteeroroolelemimimia a wawawas rrerepopoortrtrted
D-D-PDPDPDMPMPM iisss aanan aananaalologg g ofoff ggluuccoc sysysylclclceereramammididideee ananandd d wwwasss sysyyntntthehehessizezezed d tooo iiinhnhnhibibitit ggglulul cococ sysysylclclcerramamamidide ee
ynthesis in pppatatatieiei ntntnts s s wiwiw ththt GGGauauauchhhererer’sss ddisisseaeaeasesese.. HoHoHowewewevevev r, wwwee e obobobssserererveveed d d thththatatat ttthihihis s cococompmpmpououound could
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
4
extensive atherosclerosis, vascular wall thickness and stiffness and increase in the arterial levels of
glucosylceramide and lactosylceramide. Feeding these animals D-PDMP dose-dependently
ameliorates atherosclerosis and vascular stiffness.
Materials and Methods
Animals and Treatments
Apolipoprotein E-deficient (ApoE-/-) male mice aged 11 weeks (Jackson Labs, Bar Harbor, Me)
were purchased and baseline physiological parameters were measured prior to further
experimentation. At the age of 12 weeks, the ApoE-/- mice were started on a high fat and high
cholesterol diet (HFHC) of 4.5kcal/g, 2.0% fat, and 1.25% cholesterol (D12108C, Research Diet
Inc., New Brunswick, NJ) for 20 to 36 weeks with and without treatment of D-PDMP (5mpk, 10
mpk) and compared to control mice fed only chow diet and placebo fed HFHC plus vehicle. Food
was rationed once a week to estimate the weekly growth rate and food intake. Physiological
studies were performed at around 12, 20 and 36 weeks. Tissues were harvested at 12, 20 and 36
weeks of age for molecular and histopathological studies.
D-PDMP was purchased from Matreya LLC (Pleasant Gap, PA). All other chemicals were
purchased from Sigma-Aldrich (St. Louis, MO) unless mentioned otherwise. Animals were
subject to anthropometric measurements (body weight, percent body fat) and physiological
measurements [blood pressure; ultrasound to measure aortic intima-media thickening (AoIMT);
pulse wave velocity (PWV), a measure of arterial stiffening and arteriosclerosis)]. A group of mice
(n=5) were euthanized to obtain baseline values for aortic tissue and blood samples were collected.
The rest of the mice were divided into several groups. These were: Placebo (treated with vehicle
only 5%Tween-80 in phosphate buffered saline), 5mpk of D-PDMP solubilized in vehicle, and
cholesterol diet (HFHC) of 4.5kcal/g, 2.0% fat, and 1.25% cholesterol (D12108CCC,, ReReR sesesearararchchch DDiet
nc., New Brunswick, NJ) for 20 to 36 weeks with and without treatment of D-PDMP (5mpk, 10
mpmppk)k)k) aaandndnd cccomomompaaarereredd to control mice fed only chooow ww dddiet and placebobob fededd HHHFHC plus vehicle. Food
wwass s rar tioned ooncncce a wwweekekek ttto o esesestitimmamattete ttthhee weeeeekkkly ggrrrowwtwthhh raratete aandndd ffooood dd innntaakekeke.. PhPhyysysioioiololologigiicacal ll
ttudududieieesss wewererere ppererffoformrmr eeed aat t arrououo ndndnd 11122,2, 2220 00 anananddd 363636 wwweeeekkss.s TTTisisissusuuesese wwwererereee hhharrvvesesstetet dd d atatt 1112,2, 2220 0 ananddd 33636
weeks of agege fffororor mmmololo ecece uluu ararr aaandndnd hhhisisi tototopaaathththololologogogicicicalall ssstutuudid eseses.
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
5
10mpk of D-PDMP solubilized in vehicle. Vehicle and D-PDMP was delivered daily by oral
gavage. The physiological measurements were repeated at 20 and 36-week intervals and then the
mice were euthanized. New Zealand white male rabbits (7lbs) were fed rabbit chow supplemented
with 0.2% cholesterol and 14% coconut oil with and without 10mpk D-PDMP for 90 days. At 1, 2,
3 and 4 months, 5mL of blood was drawn from the ear vein. Serum was prepared and total
cholesterol levels were measured at each time point. Lactosylceramide levels were measured at 3
months. All experimental protocols were approved by the Committee for Animal Care and Use at
the Johns Hopkins University.
Glycosyltransferase Assays
Aortic tissue was homogenized in Tris buffer (pH 7.8) and centrifuged at 10,000 rpm for 10
minutes. The supernatant was used as a source for enzyme. [3H]UDP-Gal (American
Radiolabeled Chemicals, St. Louis, MO) served as the nucleotide sugar donor in LacCer synthase
assay and [3H] UDP-Glc served as the nucleotide sugar donor in GlcCer synthase assay. The
details of these assays have been described previously11, 12. All assays were conducted in triplicate
from 3 to 5 aortic samples from mice in each group.
Measurement of Atherosclerotic Lesions
Trans-thoracic echocardiography was performed in conscious mice using the 2100 Visualsonic
ultrasound device (Toronto, Ontario, Canada), equipped with a 40MHz linear transducer
respectively13, 14. The aorta was viewed in the two-dimensional (2D) mode and LV viewed along
the parasternal long axis. The intima-media thickness (IMT) was measured from the ascending
aortic wall and computed as the difference between external (Ao-ex) and internal (Ao-int)
diameters.
Inner ascending aortic diameter was measured from the inner-to-inner edge and the
Aortic tissue was homogenized in Tris buffer (pH 7.8) and centrifuged at 10,000 0 rprpm mm fofofor rr 101010
minutes. The supernatant was used as a source for enzyme. [3H]UDP-Gal (American
RaRadididiolololabababelelelededed Chehehemmmicals, St. Louis, MO) served d asaa tthe nucleotidee sus gagar rr dddonor in LacCer synthase
assaaay y and [3H]H] UUUDDPDP-G-G-Glclcc ssserererveveved d asasas ttthehee nnnucleleeootidee ssuugaaar r dodonononor r ininn GGlGlcCcCCerr ssynynynthththasaseee asasssasaay.. TTTheheh
dedetatatailillsss ofof tthehehesese aasssayayss hahaaveve bbbeeeennn dededescscririibebeedd d ppprevevevioiooussslyly11,11, 1212. . AlAllll l asasssasaaysysy wwweerere e ccocondndn ucucucteteted iinin ttririppplicccata e
from 3 to 5 aoaoortrtrticici sssamamamplplples fffroroom m m mimim cecece iin n n eaeaeachchch gggrororoupupup..
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
6
external was measured from the external edge to the external edge of the ascending aortic wall. All
measurements were performed according to the guidelines set by the American Society of
Echocardiography. For each mouse, three to five values for each measurement were obtained and
averaged for evaluation.
Blood pressure and Pulse wave velocity measurements
Systolic, diastolic and mean arterial blood pressure and heart rate were measured non-invasively in
conscious mice using the CODA tail cuff blood pressure system (Kent Scientific Corporation
USA). Data were recorded for later analysis.
Pulse wave velocity (PWV) measurement was performed non-invasively using the high
frequency and high-resolution Doppler spectrum analyzer (DSPW). Mice were placed supine on a
temperature and ECG controlled (Indus Instruments) plate and under anesthesia with 1.5%
isoflurane. Core temperature was maintained at 37 °C. A 10-MHz Doppler probe was used to
measure blood flow velocity signals at the thoracic and abdominal aorta sequentially. PWV was
calculated by the thoracic-abdominal distance divided by the pulse transit time between flow
pulses recorded at the thoracic and abdominal aortic sites. Pulse transit time was determined by the
time delay between the foot of the proximal and distal aortic flow waves in reference to the R wave
of the ECG. The sequential flow measurements in the aorta were taken a short time apart and there
was no detectable difference in physiological parameters (e.g. heart rate) between the two
measurements. Heart rate was maintained at a normal physiologic heart rate of approximately 500
(b/min)15-17. Estimated effect of mean pressure (MP (mmHg) on aortic PWV (PWVp, m/sec) was
calculated using the following equation (Avolio A et al, unpublished observations):
PWVp = 0.0006(MP)2 - 0.0564MP+3.9711
This relationship was used to correct for changes in PWV due to mean pressure between the
frequency and high-resolution Doppler spectrum analyzer (DSPW). Mice were pplalalaceceed d sususupipipinenene oon a
emperature and ECG controlled (Indus Instruments) plate and under anesthesia with 1.5%
ssofofflululurararanenee.. CoCoCoree ttteememperature was maintained at 33377 °°C. A 10-MHzHzz Dopoppplplplere probe was used to t
mmeaasasure blood d flflflowww vvele oococititity y y sisisigngnaaalslss aattt tthhe ththhoooraciiic anddd aaabdbddommmininnall aoaortrtta sesequququenentitialala lylyly. PWPWPWVV wwwas
caalclclculululatata eded bbby yy ththeee ththhororacacicici --abdbdbdomomminininaalal dddissstatatanncnce e didid vvviddededd d bybyy ttthhehe pppulullseee tttrarar nnnsiitit ttimimimeee bebeetwtwtweeeenn flflf owoww
pulses recordedeed d d atatt tttheheh ttthohoh raaaciciciccc ananand d d abababdooomimiminananalll aoaoortrtrticicc sssiti esess.. PuPuPulslslse e e trtrtrannnsisisitt t tititimememe wwwasasa dddetetetererermim ned by thhhe
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
7
treated and control groups.
Histopathology
Masson trichrome staining was performed on 5μm thin slices of the ascending aorta from mice 36
weeks after treatment and HFHC diet. These samples were photographed using a Nikon 80I
Eclipse equipped with Nikon DS-EI1 camera and the NIS-Elements software (Nikon, Japan) was
used for image analysis.
Measurement of Triglyceride and Cholesterol Levels
The serum level of triglycerides, LDL cholesterol and HDL cholesterol were measured using
commercially available kits from Wako Chemicals (Richmond, VA). Liver levels of these lipids
were quantified by HPTLC followed by densitometric scanning of charred plates.
Measurement of oxidized LDL Levels
The serum level of oxidized LDL (oxLDL) was measured using an ELISA assay and monoclonal
antibody against human oxLDL (Avanti Polar Lipids, Alabaster, AL). The ApoE-/- mouse serum
was plated in a 96-well microtitration plate at a 1:100 dilution in PBS containing 0.27 mM EDTA
and 20uM BHT overnight at 4 C. The oxLDL antibody was added after washing at a concentration
of 5ug/mL in a solution of 0.27 mM EDTA, 0.02% sodium azide in PBS and incubated overnight
at 4 C. The binding of the secondary antibody of goat anti-mouse IgM conjugated to alkaline
phosphatase (Sigma-Aldrich, St. Louis, MO) was quantified using the chemiluminescent substrate
pNPP (New England Biolabs, Ipswich, MA). Following a 4-hour incubation with the secondary
antibody, the reaction was stopped with NaOH and the plate was read at 495 nm on a microplate
reader (Bio-Rad Laboratories, Hercules, CA).
HPLC Analysis of Glycosphingolipid Levels in Liver Tissue in ApoE (-/-) Mice
Approximately 10 mg of liver tissue was homogenized in chloroform-methanol (2:1, v/v) and
were quantified by HPTLC followed by densitometric scanning of charred plateses..
Measurement of oxidized LDL Levels
Thhe e e seseerrurum mm lelelevvvel l ofofof ooxidized LDL (oxLDL) was mmmeeasured using ananan ELILILISSASA assay and monoclona
anntiiibbody againnststt hhumumaanan ooxLxLxLDLDLDL ((AvAvAvananntii Polllarrr Lipppidds, AlAlA ababbasssteterr,, AAAL)L)). Thhhe e ApApApoEoE-/-/- - momomousususee ssserrrum
wawaass s plplplatata eded iiin nn a a 99696--w-weelll mimim crcrototo itittrararatititioonon ppplalaatetete aaat aa a 1:1::10000 didilululutititiononn iinn n PBPBBS S S ccocontntn aiaiainininingngng 000 2.2.27 mMmMmM EEEDDTDTA
and 20uM BHTHTHT oovevevernrnrnigigighth aaatt t 4 44 C.C.C. TTThehehe oooxLxLxLDLDLDL aaantntn iiibobob dydyd wwwasasas aaadddddededed aaaftftftererer wwwasasashihih ngngng aaat tt aa a cococ ncentratioonn
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
8
lipids were extracted according to the Bligh and Dyer method18. The total lipid extracts were dried
in nitrogen and subject to deacylation using sphingosine ceramide N-deacylation19. Following
deacylation, lipids underwent o-phthalaldehyde derivatization and quantification of the levels of
glucosylceramide and lactosylceramide by reverse-phase high performance liquid
chromatography (RP-HPLC). A C18 column was used with an isocratic organic mobile phase
(methanol-water, 88:12, v/v) and calibrated with standard glycosphingolipids of known chemical
structure and column affinity. All samples were analyzed in triplicate and a representative quantity
(n = 3) of liver tissue samples was used for each treatment from control, placebo, 5mpk and 10mpk
D-PDMP–treated ApoE-/- mice.
Analysis of gene expression by Quantitative Real-Time PCR
A 50-mg piece of liver tissue was homogenized from each subject and total RNA was isolated
using TRIzol reagent according to the manufacturer’s instruction (Invitrogen, Camarillo, CA).
Two micrograms of RNA were reverse-transcribed with SuperScript II using random primers.
Real-time PCR was performed using SYBR Green PCR Master Mix (Applied Biosystems, Foster
City, CA, USA) in an Applied Biosystems Step one Real time PCR system with the following
thermal cycling conditions: 10 min at 95 °C, followed by 40 cycles of 95 °C for 15 s and 60 °C for
1 min for denaturation, annealing and elongation. Relative mRNA levels were calculated by the
method of 2-DDCt. Data were normalized to GAPDH mRNA levels. To determine the speci city of
ampli cation, melting curve analysis was applied to all nal PCR products. All samples were
performed in triplicate. Primers used in the present study (Supplemental Table S1) were
synthesized by Integrated DNA Technologies (Coralville, USA). Expression suite software
(Applied Biosystems) was used to analyze the data.
Western blot analysis
Approximately 90–100 mg of frozen liver tissue from each animal was homogenized in 1 mL of
Analysis of gene expression by Quantitative Real-Time PCR
A 50-mg piece of liver tissue was homogenized from each subject and total RNA was isolated
ussininngg g TRTRTRIzIzIzoolol reaaagegegentn according to the manufactutuurerer rr’s instructionn (((Inviviitrtrtrooogen, Camarillo, CA).
TTwoo o micrograamsmss ooof f RNRNR A AA wewewerrere rreveveveeersrse-e-ttrannscscribeeedd wiiththth SSuuupererSSccrriiptpt III II uuusininngg rarandnddomomm pppriiimmemerrrs..
ReRealala -t-ttimimi ee PCPCPCR R wawawas s pepeperfrfforormemeed d usususinining g SYSYSYBRBRBR GGGrerer eenen PCPCCR RR MMaMaststterere MMMixixix ((ApApApplplp ieieied dd BiBiosososyysysteteemmsms, , FoFoFostste
City, CA, USSA)A)A) iin n n anana AAAppppplililiededed BBBioioosysysysttemememsss StStStepepep ooonenen RReaeaal ll tititimememe PPPCRCRCR ssysysystetetemmm wiwiiththth ttthehehe fffolo lowing
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
9
buffer (50 mM Tris, 150 mM NaCl, 1 mM EDTA, 1% Triton X-100, 0.5% sodium deoxycholate,
0.1% SDS, pH 7.4) containing protease inhibitor cocktail (Roche). After protein quantitation using
Bradford protein assay reagent (PIERCE), equal amount of homogenate proteins were resolved by
SDS-PAGE. Respective proteins were detected by immunoblotting. Immunoreactive bands of
predicted molecular mass were visualized using an ECL plus kit (GE Healthcare Life Sciences)
and quantified with the KODAK Molecular Imaging Software (Kodak, New Haven, CT). GAPDH
was used as a loading control. The data represent the mean ± SD of three independent experiments.
Statistical Analysis
All values are expressed as dot plots alongside mean± SEM. Comparison between groups was
performed with Kruskal-Wallis test and Dunn’s multiple comparison post-test. For repeated
measures in mice and rabbits, a two-way repeated measures ANOVA (RM-ANOVA) was
performed with Bonferroni’s multiple comparison test for comparisons between groups. * p 0.05,
** p 0.01, *** p 0.001, **** p 0.001. GraphPad PRISM and Excel statistical software were
used.
Results
Aortic wall thickness, vascular stiffness and remodeling with and without treatment
Ultrasound studies revealed that, as compared to control mouse aorta (Fig 1A), feeding a western
diet to mice from age 12 weeks to 20 weeks contributed to a marked increase in aortic wall
thickening (indicated by arrows) (Fig 1B) in placebo mice. This was not observed in mice fed
HFHC diet plus 10mpk of D-PDMP (Fig 1C). At 36 weeks of age the control mice show some
Ca2+ deposit, perhaps due to aging (Fig 1D). However, extensive Ca2+ deposits were noted in
placebo mice (Fig 1E) and this was prevented by treatment with 10mpk of D-PDMP (Fig 1F).
performed with Kruskal-Wallis test and Dunn’s multiple comparison post-test. Foor rer pepeatateded
measures in mice and rabbits, a two-way repeated measures ANOVA (RM-ANOVA) was
perfforormmedd wiwith BBoonferroni’s multiple comparisonon teest for comparisisons bebetween groups. * p 0.05
** pp 0.01, **** p 00.000101, ******* pp 00.0001. GGraphPhPadd PPRISMSM andnd Exccell staatisisticaal sos ftf wwaree wwere
useded.
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
10
Further quantitative analysis revealed that aortic intima thickening progressed rapidly in
placebo mice fed HFHC diet from 12 weeks to 36 weeks of age as compared to control mice.
However, D-PDMP exerted a dose-dependent decrease in aortic intima thickening. In fact, the use
of 10mpk of D-PDMP maintained the aortic intima-media thickening in HFHC fed mice
comparable to control mice (Fig 1G). Likewise, pulse wave velocity measurement revealed a
similar pattern, i.e., suggesting that the thickening of the aortic wall was associated with arterial
stiffening. This was lowered to control levels upon treatment with 5 and 10mpk D-PDMP in a
dose-dependent manner (Fig 1H). Since blood pressure did not change significantly with
treatment, the increase in arterial stiffness is largely independent of blood pressure.
Body weight for baseline (29.4±0.93 g) the control (31.13±1.61 g) and the placebo
(34.5±0.88 g) groups were not significantly different. At 36 weeks, the 5 and 10 mpk plus HFHC
diet fed mice showed a dose -dependent increase in body weight (36.53 ±1.77 g and 41.03±0.55 g)
compared to other groups. This was mainly because of an increase in bone mass density and
muscle mass. These mice were also physically active and less aggressive than the placebo group
on HFHC diet plus vehicle only. The latter instead showed an increase in percent body fat and
were fragile. This suggests that HCHF may have an association with bone loss and calcified aortic
wall shown in Fig 1E and no association with obesity.
Assessment of Atherosclerosis
Masson trichrome staining of 5μm thin slices of the aorta in control mice revealed normal aortic
wall thickness with no fibrosis (Fig 2A). In contrast, 36-week-old placebo mice fed HFHC diet
alone and daily dose of vehicle exhibited narrowed lumen volume, occupied largely by plaques
loaded with cholesteryl esters (needle-like white crystals), amorphous Ca2+ deposits within the
plaque and extensive fibrosis (Fig 2B).
Body weight for baseline (29.4±0.93 g) the control (31.13±1.61 g) and thheee plplp acccebebebooo
34.5±0.88 g) groups were not significantly different. At 36 weeks, the 5 and 10 mpk plus HFHC
diietett fffededed mmmicicicee showowowede a dose -dependent increaseee iiin n bbody weight (3(336.53533 ±±±1.1 77 g and 41.03±0.55 g
coommpmpared to ototheheher r grroououppsps. ThThThisiis wwasasas mmmaiainnly beeecauuusee off f aanan iiincnccrereaaaseee inin bbbonne e mamamassss dddenennsisiittyty aaandndnd
mumuuscscs lelele mmasasass.ss. TThhehessese mmmiccce e weweererr aaalslslso oo phphhysysysicicicaalallyyy aaccttivvve aaandndnd llleess ss agagggrgrresesessisiivevee tthahahan n ththt ee e plpplaccebebboo grgrgrououupp
on HFHC dietett pppluluusss vevevehihihiclc e e e onononlyyy.. ThThThe ee laaattttt ererer iiinsnsnsteteteadadad ssshohoh wewewed d d ananan iiinnncrcc eaeaeasesese iiin n n pepepercrccenenent t bobobodydd fat and
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
11
Treatment with 5mpk of D-PDMP significantly improved lumen volume; cholesteryl
esters were expunged from the plaque (Fig 2C) compared to placebo (Fig 2B), reduced fibrosis,
and elastin fragmentation. The most unexpected result was the observation that treatment with
10mpk of D-PDMP completely prevented atherosclerosis in these mice at age 36 weeks (Fig 2D).
Notably, coronary artery disease, the principle cause of morbidity and mortality due to
atherosclerosis, was drastically prevented by the treatment. A clear lumen was observed at 36
weeks for control mice, while a narrowed lumen volume was observed for placebo mice fed HFHC
diet (Fig 2E). This was mitigated by treatment with 5mpk and 10mpk D-PDMP (Fig 2E).
Histopathological analysis of the left coronary artery and quantification of lumen area revealed
that D-DPMP reduces obstruction and plaque deposition in apoE-/- mice (Supp. Fig. 1).
Treatment with D-PDMP dose-dependently decreases the activity of glycosyltransferases in
the aorta in ApoE-/- mice
High activity of glucosylceramide synthase and lactosylceramide synthase in the aorta in ApoE-/-
mice was observed in 20-week-old mice fed HCHF diet (Fig 3A, 3B). When these mice were
given a daily supply of D-PDMP by oral gavage during this period (at age 20 weeks) it dose
-dependently decreased the activity of these enzymes. We noted a 10-fold increase in the activity
of LacCer synthase in 36-week-old mice ApoE-/- mice fed the western diet (Fig 3C). However, up
to the age of 36 weeks did not exhibit any further increase in the activity of GlcCer synthase
compared to 20-week-old mice (Fig 3D). Notwithstanding these observations, some of this
increase may be attributed to effects of aging. D-PDMP dose-dependently decreased the activity of
both GlcCer synthase and LacCer synthase in 36 week old mice, as indicated by the mass of
LacCer (Fig 3E) and GlcCer (Fig 3F).
Treatment with D-PDMP ameliorates hyperlipidemia in ApoE-/- mice
Feeding a high fat and cholesterol diet markedly increased the serum level of oxidized LDL
hat D-DPMP reduces obstruction and plaque deposition in apoE-/- mice (Supp. . FiFiFig.gg 11).).).
Treatment with D-PDMP dose-dependently decreases the activity of glycosyltransferases in
hhe e aoaoaortrtrtaa ininin AAApopooEE-E-/- mice
HHigghgh activity ofoff gglluucocosysyylclclceereramamamidide e e sysysynntthhaase annnd laacacttosylylylcecerarammimiddee ssynynthththaasse e ininin tthehe aaaororrtata in n n ApApApoEoEE-/
mimiicecec wwwasas ooobsbsbserervvvedd d inin 200-0-weweekekek-o-o-oldldld mmicicce ee fefefeddd HCHCHCHFHFHF diieiet t (((FiFiFig g 3A3A3A,, 3B3B3B).). WWWhehenn n thththesessee e mmmiccce wwererre
given a daily y y sususupppppplylyly ooof f f D-D-D PDPDPDMPMPMP bby y y ororralala gggavavavaaagegege dddurururinini g g g thththisisis ppperererioioiod d (a(a(att t agagage e e 20202 wwweeeeeeksksks) ) ) it dose
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
12
measured using an ELISA assay. In contrast, feeding the glycosyltransferase inhibitor
dose-dependently decreased the serum levels of ox-LDL level to below baseline levels in the
ApoE-/- mice serum (Fig 4A). Similarly, the serum level of LDLc (Fig 4B) and triglycerides (Fig
4C) were increased in western diet fed mice serum. This was also decreased to baseline values in
mice fed D-PDMP. Interesting, although feeding the western diet decreased the serum levels of
HDLc (Fig 4D), treatment with D-PDMP raised HDLc level in the serum. Thus D-PDMP may be
cardioprotective.
Effect of D-PDMP on the expression of genes involved in LDL metabolism
As shown in Fig 5A, treatment with 10mpk D-PDMP increased the mRNA levels of
3-hydroxy-3-methylglutaryl-Coenzyme A reductase and sterol regulatory element binding
transcription factor 2 up to approximate 5.2-fold (P < 0.0001) and 3.6-fold (P < 0.0001),
respectively. The mRNA level of Low density lipoprotein receptor was also elevated by D-PDMP
respectively.
Effect of D-PDMP on the expression of transporter genes involved in cholesterol efflux, bile
synthesis and triglyceride metabolism
Real time PCR showed that the expression level of CD36, SRB-1 and ABCA1 genes were
significantly upregulated by D-PDMP treatment (Fig 5B). These transporters play a major role in
cholesterol efflux from peripheral tissues. As shown in Fig 5C, treatment with D-PDMP dose
dependently increased the mRNA levels of lipoprotein lipase and very low-density lipoprotein
receptor to approximate 4.8-fold (P < 0.0001) and 2.4-fold (P < 0.0001) in liver samples of drug
treated mice. The level of Cyp7A1 was also significantly up regulated by the drug treatment (Fig
5D), indicating cholesterol catabolism and bile acid synthesis upon drug treatment.
Effect of D-PDMP on the expression of proteins involved in LDL metabolism
Western immune-blot assays revealed that placebo mice liver tissue had significantly lower
3-hydroxy-3-methylglutaryl-Coenzyme A reductase and sterol regulatory elemeennnt bbbinindididingngng
ranscription factor 2 up to approximate 5.2-fold (P < 0.0001) and 3.6-fold (P < 0.0001),
eespsppececectititiveveelylyly. ThThe e e mmRmRNA level of Low density lipipipopopprotein receptoor rrf waasss aalalso elevated by D-PDMP
eespppece tively.
EfEfffefefectctct oooff D-D-D-PDPPDMPMPMP oonnn tththee exexxprprpresesessisisiononn oooff f trtrtrannnspspsporororttet rrr gegegenenenes s inininvvovolvlvlvededed innn chchholololesese teteterrorol eefeffflfluxuxx, bbibile
ynthesis annd d d trtrtrigigglylyyceceerirr deee mmmetttabababolololisi mmm
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
13
expression of SREBP2 protein as well as LDL receptor protein mass (Fig 6A,B). In contrast,
treatment with D-PDMP markedly increased the expression of these proteins. These findings are
in tandem with the gene expression data above, suggesting that drug treatment prevented
atherosclerosis by increasing cholesterol metabolism via the LDL receptor pathway.
Treatment with D-PDMP ameliorates atherosclerosis in normal rabbits fed a western diet
Treatment with the glycosphingolipid glycosyltransferase inhibitor D-PDMP prevented
atherosclerosis in New Zealand white rabbits. Rabbits fed HFHC diet showed a marked increase in
aortic lactosylceramide levels (Fig 7A) accompanied by extensive atherosclerosis and a 17-fold
increase in serum cholesterol (Fig 7B). In contrast to rabbits fed HFHC diet alone, rabbits treated
with D-PDMP showed a prevention of atherosclerosis as measured by lactosylceramide and
cholesterol levels which were similar to the control (Fig 7).
Discussion
The following major findings emerged from the present study. D-PDMP, an inhibitor of
glucosylceramide synthase and Lactosylceramide synthase dramatically and dose-dependently
ameliorated atherosclerosis in ApoE-/- mice and normal rabbits fed a western diet. In mice this
was accompanied with complete reversal of aortic intima-media thickening, and pulse wave
velocity, an index of vascular stiffness. Our mechanistic studies revealed that D-PDMP decreased
serum levels of cholesterol by way of regulating the expression of genes implicated in the
biosynthesis, egress, and conversion of cholesterol to bile acids. The triglyceride levels were also
dramatically reduced in the D-PDMP -treated mice by an increase in VLDL receptor and
lipoprotein lipase gene expression.
A close association between glycosphingolipids and atherosclerosis was suggested by us3,
with D-PDMP showed a prevention of atherosclerosis as measured by lactosylcererrammmidddee e ananand d d
cholesterol levels which were similar to the control (Fig 7).
DDisccscussion
ThThhe ee fofofolllll owowwinining g mmmajojoorr fffinndndiningsgss eemememergrgrgededd fffrororommm ththhe e e pppreeseseennt t stststuuudy.y.y. DDD-PPPDMDMDMPPP, , , anann iinhnhn ibibbitittoor ooof f
glucosylceraamimim deded sssynynynthththasa e e e ananand LaLaLactcttoso ylylylcececerararamimimidedede sssynynynthhhasasa ee e drdrd amamamata icicicalalallylyly aaandndnd dddososose-e-e dededepepp ndently
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
14
7 and others2, 20, 21. However, few attempts have been made to examine the effects of
inhibiting/depleting the level of GSLs in atherosclerosis in experimental animal models. Recently,
Bietrix et al. used an iminosugar, N-5--adamatane -1 '-ul-methoxy)-pentyl1- deoxynojirimycin
(AMP-DNM), an inhibitor of glucosylceramide synthase, which markedly reduced plasma
cholesterol and inhibited atherosclerosis development in ApoE*3 Leiden and LDL receptor -/-
mice. Additional studies have made use of this inhibitor in enhancing insulin responsiveness in a
rat model of diabetes22 and in liver steatosis23. Another inhibitor of GlcCer synthase, Zavesca (1,5-
butylimino) 1,5-dideoxy-D- glucitol and Genz-112638 / miglustat is already in use for substrate
inhibition therapy24 in patients with Gaucher's disease. D-PDMP inhibits both GlcCer synthase
and LacCer synthase activity in cultured mammalian cells and can directly inhibit the activity of
pure LacCer synthase7. Our in vivo studies have shown its efficacy in mitigating
VEGF/FGF-induced angiogenesis, restenosis in rabbits following balloon angioplasty25 and
lowering renal tumor volume by 50%26. Since arteriosclerosis is a multi-factorial disease, we
hypothesized that a compound such as D-PDMP could be efficacious in lowering the GSL load
and the phenotypes above.
An anticipated observation in this study was the remarkable filling of the lumen with
atherosclerotic plaques in ApoE -/- mice fed high fat and cholesterol diet which progressively
worsened from 20 weeks to 36 weeks of age. This was accompanied with extensive arterial wall
thickening and an associated increase in the arterial wall stiffness measured by ultrasound.
Consequently, pulse wave velocity also increased. This was ameliorated upon treatment with
D-PDMP. An unexpected observation was that from week 12 to week 20 there were no visible Ca
2+ deposits in the arterial wall. However, at 36 weeks of age, even mice fed mice chow alone had a
few Ca2+ deposits. Whereas, mice fed HFHC diet had extensive Ca2+ deposits, and this was
and LacCer synthase activity in cultured mammalian cells and can directly inhibbititit tthehh aaactctctivivivititity y of
pure LacCer synthase7. Our in vivo studies have shown its efficacy in mitigating
VEVEGFGFGF/F/F/FGFGFGF i-i-indnducucuceeded angiogenesis, restenosis inn rrrababbbits followinggg bballoloooonon angioplasty25 and
oowwewering renalal ttumumumoror vvolololumummeee bbyby 5550%0%%2262 . Sinncceee artteerriiossclclclereroosiiss iis s aa mumultltl i--fafactctctoororiaial l dididiseses aaase,e,e, wwwe e
hyhypopopothththesesizizzededed tthahaat aa a cocoompmpmpoounndnd sssucucuch h h asas DDD-P-P-PDMDMDMP P P cccoouulddd bbbeee eeeffifificacac cicic ouououss iiin llowowwererrinini g g g ththhe GSGSGSL L lolooaadad
and the phennotototypypypeseses aaabobob vevv ..
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
15
prevented by treatment with D-PDMP. These observations were made using 5 and 10mpk of
D-PDMP in a dose- dependent manner. Not only are these drug concentrations 10 -fold lower than
a previous report8, but also these mice did not accumulate subcutaneous fat. There was also a
higher bone density27 as compared to the placebo over the 36-week study, and no overt obesity was
observed. As expected, D-PDMP treatment not only decreased the activity of GlcCer synthase but
also LacCer synthase in the aorta in these mice. This was accompanied by a decrease in the level of
GlcCer, LacCer as well as ceramide in the liver tissue. D-PDMP also decreased the level of serum
cholesterol and triglycerides via recruiting multiple genes/pathways in lipid metabolism. For
example, we noted that D-PDMP increased LDL receptor and SREBP2 gene expression and
decreased HMG-CoA reductase gene expression. These findings are suggestive of inhibition of
cholesterol biosynthesis and increased LDL uptake. It also increased the expression of genes
responsible for cholesterol efflux by way of increasing the expression of ABCA1 and increased
expression of ABCG 5, ABCG 8 proteins responsible for pumping cholesterol out from liver
and intestine for excretion. In a previous study it was shown that GSL accumulation in particular
lactosylceramide in cultured cells can inhibit the efflux of cholesterol via the
ABCA1/Apolipoprotien A-1 pathway. In contrast, D-PDMP was found to serve as a cholesterol
efflux accelerator by inducing the expression of these genes. Our findings are in agreement and
expand these observations to an animal model of atherosclerosis.
A 7-hydroxylase enzyme, coded by the gene Cyp7A1, can convert cholesterol to bile acid.
The expression of this gene was increased in mice treated with D-PDMP. This gene plays a pivotal
role in the expression of an enzyme required for the conversion of cholesterol to bile acids. In a
previous study the iminosugar N sugar AMP- DNM was also shown to increase the expression of
this gene in liver tissue in ApoE*3 Leiden mice and facilitate the lowering of cholesterol load by
decreased HMG-CoA reductase gene expression. These findings are suggestive ooof f inininhhihibibibitititiononon oof
cholesterol biosynthesis and increased LDL uptake. It also increased the expression of genes
eespspponononsisisiblblblee e fofofor chchhoololesterol efflux by way of incrrreaeae ssing the expresssssion n ofofof AABCA1 and increased fff
exxprprese sion of ABABABCGCGCG 55, ABABABCGCGCG 88 prprprooto eeeinnns reeesppponsssibbble fffooror pppumummppiinnggg chchhoolo eesteteerororoll ouout tt frfrfromomm llliviivereer
anndd d ininntetet ststininne ee fofor r exxxcrcreeetioioon.n. IInn n a a prprprevevevioioususus ssstututudyyy iit t wawawas shshshowowownn thththatatt GSGSGSL L accccucumumumulalal titiiononon innn papapartrtiiccuululara
actosylceramimimideded iiin n n cucucultltturu ededed cccellllslsls ccanana inininhihihibibibit t thththe e efefefflflfluxux ooof f f chchcholololesesesteterororol l l vivivia a a thththee
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
16
conversion to bile acids and their subsequent excretion8. Moreover, in our study HDL cholesterol
was increased about 2-fold in the serum of D-PDMP treated mice. Thus a combination of genes
implicated in the lipid and lipoprotein metabolic pathways contributed to the complete reversal of
a marked increase in cholesterol level and atherosclerosis in the ApoE-/- mice fed HFHC diet.
Previous studies have shown that an enzyme lipoprotein lipase plays a crucial role in
linking up VLDL, the major triglyceride carrying lipoprotein to VLDL receptors28. We observed
that the liver tissue in D-PDMP treated mice had increased expression of gene required for the
expression of lipoprotein lipase1 as well as VLDL receptors. Such observations could explain why
treatment with D-PDMP could reduce the markedly increased serum levels of triglycerides in
ApoE-/- mice a HFHC diet for 36 weeks (Fig 4C).
We have previously shown that oxidized LDL can dose- dependently increase the activity
of LacCer synthase in cultured human arterial smooth muscle cells by phosphorylating the serine,
threonine and tyrosine residues in this enzyme. This was mitigated by pre- treatment of cells with
D-PDMP29. The activation of this enzyme generated LacCer that, in turn, activated a signaling
cascade involving ROS generation, p44MAPK activation, nuclear transcription factor c-Fos and
cyclin A to induce cell proliferation29. In the present study we show that feeding a western diet
significantly increases the level of Ox-LDL in the serum of ApoE-/- mice. This may have
contributed to the activation of LacCer synthase in the arterial wall, thus contributing to an
increase in the levels of LacCer. In contrast, in mice fed western diet plus D-PDMP the level of
Ox-LDL was decreased to basal levels (Fig 4A). This reduction in LacCer level might have
contributed to the resumption of cholesterol efflux at the normal level (Fig 8). An important
observation made in our present study is that fibrosis contributes greatly towards arterial wall
thickness in our western diet fed model of atherosclerosis and this too was prevented in mice fed
ApoE-/- mice a HFHC diet for 36 weeks (Fig 4C).
We have previously shown that oxidized LDL can dose- dependently increase the activity
off LLLacacacCeCeCerrr sysys nnnthahaasesese in cultured human arterial smmomooooth muscle celllslsl by y phphphoso phorylating the serine
hhhreeeono ine and d tytyroror ssinenee rreesesidididueueuess ininn tthhihiss ennnzymememe. Thhhiiss wwwasass mmmittigigi atattededd bbyy prpre-e- tttrereeatatmememenntnt oof ccecelllllsss wwwith
D-D-PDPDPDMPMPM 2929.. ThTThee accctitivavatitiionon oof f f ththhisisis eeennznzymymymee e gggeneneneraraatetedd LLaLacCcCcCeeer ttthahah tt,t iiinn n tutuurnrn,,, acacctitivavav teteed d d a ssiigngnnalaliininggg
cascade invoolvlvvininng g g ROROROSS gegenenenerarr titiiononon,, p4p4p 4M4M4MAPAPAPK K K acacctititivavavatit ononon,,, nununuclclcleaeaear trtrtranananscscscririr ptptp ioioon n n fafafactctctororo c-Fos andd
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
17
D-PDMP.
In sum, we conclude that there is a tight relationship between glycosphingolipid
metabolism, and lipoprotein metabolism, which heavily impacts upon aortic intima media
thickening via fibrosis and lipid deposition. Such multigeneic changes bring about arteriosclerosis
and its pathological sequelae. Herein, we demonstrate that several genes responsible for the
biosynthesis of cholesterol, its efflux, absorption, bile acid production, and excretion were
improved upon treatment with a glycosphingolipid glycosyltransferase inhibitor. Additionally,
genes responsible for triglyceride metabolism were also improved due to treatment resulting in
decreased triglyceride levels in apoE-/- mice. An important observation was the increased
expression of genes responsible for apoA1, the major protein in HDL, thus contributing to an
increase in the plasma levels of HDL in treated mice.
Limitations
The glycosphingolipid inhibitor used in this study inhibits the synthesis of glucosylceramide
synthase as well as lactosylceramide synthase and therefore is not specific for a particular enzyme
in the biosynthetic pathway. However, this compound is not toxic to animals when given for the
duration of this study -6 months. Clearly, further studies are required using relatively more specific
inhibitors in the near future. PWV depends on arterial pressure, but in these experiments PWV was
measured using external Doppler transducers so it was not possible to obtain PWV-pressure
relationships by altering pressure with vasoactive agents. However, the changes in pressure
between the groups was not large as assessed by tail-cuff measurements, and when there was a
change the PWV values were adjusted for pressure differences using relationships obtained in
mice from other studies. Results showed that pressure adjustments did not produce any major
alterations, hence this is not considered to be a significant limitation in these studies.
expression of genes responsible for apoA1, the major protein in HDL, thus contrtriibibututu innng gg tototo aaan n na
ncrease in the plasma levels of HDL in treated mice.
LLiLimmimitationss
ThThhe glgg ycycosphphphinini ggogolllipipiddd iinnhiibbiitooor r ususededed iinn ththisis stttududyyy iinnhihiibibiitsts tttheee syynyntthesiis ofof gggluuucocosysyylcccerramammidideee
yyyntntnthahahasesese aaasss wewewellllll aaasss lalalactctctosososylylylcececerararamimimidedede sssynynynthththasasaseee anananddd thththerererefefeforororeee isisis nnnototot ssspepepecicicififificcc fofoforrr aaa papapartrtrticiciculululararar eeenznznzymymyme
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
18
Acknowledgments: We thank Sara Kimiko Suzuki, Maya Hernandez, Fraulein Li, Bryan
Brensinger and Jennifer Hou for assistance during various phases of this research.
Funding Sources: This study was funded by NIH grant’s P01HL107153 and
3PO1HL107153-03S1 (SC).
Conflict of Interest Disclosures: None.
References:
1. Kwiterovich P. The johns hopkins textbook of dyslipidemia. Philadelphia: Wolters Kluwer Health/Lippincott Williams & Wilkins; 2010. 2. Dawson G, Kruski AW, Scanu AM. Distribution of glycosphingolipids in the serum lipoproteins of normal human subjects and patients with hypo- and hyperlipidemias. J Lipid Res. 1976;17:125-131. 3. Chatterjee S, Kwiterovich PO. Glycosphingolipids of human plasma lipoproteins. Lipids. 1976;11:462-466. 4. Bhunia AK, Arai T, Bulkley G, Chatterjee S. Lactosylceramide mediates tumor necrosis factor-alpha-induced intercellular adhesion molecule-1 (icam-1) expression and the adhesion of neutrophil in human umbilical vein endothelial cells. J Biol Chem. 1998;273:34349-34357. 5. Gong N, Wei H, Chowdhury SH, Chatterjee S. Lactosylceramide recruits pkc / and phospholipase a2 to stimulate pecam-1 expression in human monocytes and adhesion to endothelial cells. Proc Natl Acad Sci U S A. 2004;101:6490-6495. 6. Glaros EN, Kim WS, Quinn CM, Wong J, Gelissen I, Jessup W, Garner B. Glycosphingolipid accumulation inhibits cholesterol efflux via the abca1/apolipoprotein a-i pathway: 1-phenyl-2-decanoylamino-3-morpholino-1-propanol is a novel cholesterol efflux accelerator. The J Biol Chem. 2005;280:24515-24523. 7. Chatterjee S, Ghosh N. Oxidized low density lipoprotein stimulates aortic smooth muscle cell proliferation. Glycobiology. 1996;6:303-311. 8. Bietrix F, Lombardo E, van Roomen CP, Ottenhoff R, Vos M, Rensen PC, Verhoeven AJ, Aerts JM, Groen AK. Inhibition of glycosphingolipid synthesis induces a profound reduction of plasma cholesterol and inhibits atherosclerosis development in apoe* 3 leiden and low-density lipoprotein receptor / mice. Arterioscler Thromb Vasc Biol. 2010;30:931-937. 9. Kamani M, Mylvaganam M, Tian R, Rigat B, Binnington B, Lingwood C. Adamantyl glycosphingolipids provide a new approach to the selective regulation of cellular
2. Dawson G, Kruski AW, Scanu AM. Distribution of glycosphingolipids in the seeruum mipoproteins of normal human subjects and patients with hypo- and hyperlipidemimiiasasas.. J J J LiLiLipipipid d d ReReR s.
1976;17:125-131.
3. Chatterjee S, Kwiterovich PO. Glycosphingolipids of human plasma lipoproteins. Lipids.1976766;1;1;11:1:1:4646462-2-2-46466.6.6.
44.. BBBhuh nia AKAK, , ArAAraaai TTT,, BuBuBulklkkleleey y y G,G,G CCChahahatttererjejejee e S.S. LLLacctosysysylclclcerere amamamididide e mememeddidiaaatess tttumumumororo nnnecececroror sisisisssfaafactttoro -alpha-inducucuced iintntterceceellluuular adadheheessiion mmmoolecccuulle-11 (i(i(icacacammm-1)) eexxpreessssiionnn aaand thhhe aaddhheeesionnn ooof neneeututu roror phphill iinn n huhuh mamaman n umummbibiliiccacall vveieinn n enendodooththeeleliaaal l ceceelllss. JJJ BBBioiolll CChCheemm.. 19199998;2;277373:3:334343494949--344435757..
555. GGGonongg NNN, WWW ieiei HHH, , ChChChowowdhdhdhururyy SHSHSH,, ChChCh tatatttete jrjrjeeee SSS. LLaLa tctctosos lylylcecerara imimiddede rrececruruitititss kpkpkcc /// aa dndnd php osphp olippasasse e e a2a2a2 ttto o ststtimimmulullatatate e pepecacac mm-11 exeexprpresessisiionoon iiin nn huhumamam nn n momononocycytetetes s anand d adaddheheessisionoon to
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
19
glycosphingolipid metabolism. J Biol Chem. 2011;286:21413-21426. 10. Radin NS, Shayman J, Inokuchi J-I. Metabolic effects of inhibiting glucosylceramide synthesis with pdmp and other substances. Adv Lipid Res. 1993;26:183. 11. Chatterjee S. Assay of lactosylceramide synthase and comments on its potential role in signal transduction. Meth Enzymol. 2000;311:73-81. 12. Basu M, De T, Das KK, Kyle JW, Chon HC, Schaeper RJ, Basu S. Glycolipids. Meth Enzymol. 1987;138:575-607.
13. Habashi JP, Holm TM, Doyle JJ, Aziz H, Bedja D, Dietz HC. At2 signaling is a positive prognostic and therapeutic modifier of marfan syndrome: Lessons on the inequality of acei and arbs. Circulation. 2009;120:S963-S963. 14. Olson LE, Bedja D, Alvey SJ, Cardounel A, Gabrielson KL, Reeves RH. Protection from doxorubicin-induced cardiac toxicity in mice with a null allele of carbonyl reductase 1. Cancer Res. 2003;63:6602-6606.
15. Jun J, Reinke C, Bedja D, Berkowitz D, Bevans-Fonti S, Li J, Barouch LA, Gabrielson K, Polotsky VY. Effect of intermittent hypoxia on atherosclerosis in apolipoprotein e-deficient mice. Atherosclerosis. 2010;209:381-386. 16. Sussan TE, Jun J, Thimmulappa R, Bedja D, Antero M, Gabrielson KL, Polotsky VY, Biswal S. Disruption of nrf2, a key inducer of antioxidant defenses, attenuates apoe-mediated atherosclerosis in mice. PLoS One. 2008;3. 17. Sussan TE, Rangasamy T, Blake DJ, Malhotra D, El-Haddad H, Bedja D, Yates MS, Kombairaju P, Yamamoto M, Liby KT, Sporn MB, Gabrielson KL, Champion HC, Tuder RM, Kensler TW, Biswal S. Targeting nrf2 with the triterpenoid cddo-imidazolide attenuates cigarette smoke-induced emphysema and cardiac dysfunction in mice. Proc Natl Acad Sci U S A. 2009;106:250-255. 18. Bligh EG, Dyer WJ. A rapid method of total lipid extraction and purification. Can J Biochem. 1959;37:911-917. 19. Zama K, Hayashi Y, Ito S, Hirabayashi Y, Inoue T, Ohno K, Okino N, Ito M. Simultaneous quantification of glucosylceramide and galactosylceramide by normal-phase hplc using o-phtalaldehyde derivatives prepared with sphingolipid ceramide n-deacylase. Glycobiology. 2009;19:767-775. 20. Garner B, Priestman DA, Stocker R, Harvey DJ, Butters TD, Platt FM. Increased glycosphingolipid levels in serum and aortae of apolipoprotein e gene knockout mice. J Lipid Res. 2002;43:205-214. 21. Glaros EN, Kim WS, Quinn CM, Jessup W, Rye K-A, Garner B. Myriocin slows the
2003;63:6602-6606.
15. Jun J, Reinke C, Bedja D, Berkowitz D, Bevans-Fonti S, Li J, Barouch LA, GaGaabrbrbrieielslslsononon KKK, Polotsky VY. Effect of intermittent hypoxia on atherosclerosis in apolipoprotein e-deficient miceAtherosclerosis. 2010;209:381-386.
16166. SSuSussan TTTEEE, JJununun JJ, , ThThThimimmmumum lalaappppppa a R,R,, BBededjajaj DD, , AAnAntteroo MMM,,, GaGG brbrrieieiellslsonon KKKL,LL PPololotototsksks y y VYVYY,, BiBiiswswswala SDDisrrsrupu tion of nrnrff2f ,, a a kekek y y y ininindududuccecer r oofof aaantnttioooxiddaannnt deefeeenseeess,s, aatttttennnuauateess apapoooe-m-mededediiaiatetedd d atatatheheheroooscsclelelerroosinnn mmmici e. PLoSoS OOnenee. 20000088;33.
17. SuSussanan TTE,E RRanangagasamymy TT, BlBlakake DJJ, MaMalhlhototrar DD, ElEl-H-Hadaddad d d H,H, BBede jaa DD, YaYatet s MSM ,,Kombairaju PPP, , , YaYaYamamam momomotoo MMM,,, LiLiLibybyb KKKT,T,T, SSSpopopornrnrn MBMBMB,, GaGaG brbrbrieieelslssononn KKKL,L,L, CCChahahampmpmpioion nn HCHCHC,,, TuTT der RM,KeKensnslelerr TWTW BiBiswswalal SS TaTargrgetetiningg nrnrf2f2 wwitithh ththee trtrititererpepenonoidid ccddddoo-imimididazazololididee atattetenunuatateses ccigigararetetttee
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
20
progression of established atherosclerotic lesions in apolipoprotein e gene knockout mice. J Lipid Res. 2008;49:324-331.
22. Aerts JM, Ottenhoff R, Powlson AS, Grefhorst A, van Eijk M, Dubbelhuis PF, Aten J, Kuipers F, Serlie MJ, Wennekes T. Pharmacological inhibition of glucosylceramide synthase enhances insulin sensitivity. Diabetes. 2007;56:1341-1349.
23. Bijl N, Sokolovi M, Vrins C, Langeveld M, Moerland PD, Ottenhoff R, van Roomen CP, Claessen N, Boot RG, Aten J. Modulation of glycosphingolipid metabolism significantly improves hepatic insulin sensitivity and reverses hepatic steatosis in mice. Hepatology. 2009;50:1431-1441.
24. McEachern KA, Fung J, Komarnitsky S, Siegel CS, Chuang W-L, Hutto E, Shayman JA, Grabowski GA, Aerts JM, Cheng SH. A specific and potent inhibitor of glucosylceramide synthase for substrate inhibition therapy of gaucher disease. Molec Genet Metab. 2007;91:259-267. 25. Chatterjee S. Methods for treatment of angiogenesis. US Patent application #20090202439. Issued Aug 13, 2009.
26. Chatterjee S, Alsaeedi N, Hou J, Bandaru VVR, Wu L, Halushka MK, Pili R, Ndikuyeze G, Haughey NJ. Use of a glycolipid inhibitor to ameliorate renal cancer in a mouse model. PLoS One. 2013;8:e63726. 27. Hyder JA, Allison MA, Barrett-Connor E, Detrano R, Wong ND, Sirlin C, Gapstur SM, Ouyang P, Carr JJ, Criqui MH. Bone mineral density and atherosclerosis: The multi-ethnic study of atherosclerosis, abdominal aortic calcium study. Atherosclerosis. 2010;209:283-289. 28. Takahashi S, Suzuki J, Kohno M, Oida K, Tamai T, Miyabo S, Yamamoto T, Nakai T. Enhancement of the binding of triglyceride-rich lipoproteins to the very low density lipoprotein receptor by apolipoprotein e and lipoprotein lipase. J Biol Chem. 1995;270:15747-15754. 29. Chatterjee S, Bhunia AK, Snowden A, Han H. Oxidized low density lipoproteins stimulate galactosyltransferase activity, ras activation, p44 mitogen activated protein kinase and c-fos expression in aortic smooth muscle cells. Glycobiology. 1997;7:703-710.
Figure Legends:
Figure 1. Aortic wall thickening and pulse wave velocity in atherosclerotic mice is ameliorated by
treatment with D-PDMP. Aortic ultrasound imaging: 2DB-mode ultrasound images of the aorta
from ApoE-/- mice fed mice chow. Top panel (A, B, C) shows 20 week old mice. Bottom panel (D,
25. Chatterjee S. Methods for treatment of angiogenesis. US Patent application #2000090202439. ssued Aug 13, 2009.
26. Chatterjee S, Alsaeedi N, Hou J, Bandaru VVR, Wu L, Halushka MK, Pili R, Ndikuyeze G, Haugghey y NJJ. Usse of a glycolipid inhibitor to ameliorate renal cancer in a mouse model. PLoS One200131313;8;8;8:e:e6363637272726.
2277. HHyder JA, AAlAlliiisoon nn MAMAMA, , BaBaBarrrretettt-t-CoCoConnnnor EE,,, Deetrtraanooo RR,R, WWWoononggg NNDND, SiSS rrrlininn CCC,, GaGaapspsstuturrr SMSMSM,OOOuyyayang P, CaCarr JJJJJ,, Crriiqquui MHMHMH. BoB nenen mmmiineraaal dennsiitty y ananand d d atata hhherooscclerossisss: TTThhee mmululltii-e-ettthnnnic ssstuuudyofff aaaththherere ososclclclereerososiiss, ababddodommiminanall l aoaoortrtrticicic ccaalalciciiumumum sstututudydydy. AtAtheheherororosscsclelelerororosisisss. . 202001000;2;209090 :2:2: 83833-2-2-28999.
28. Takahashhi i i S,S,S, SSSuzuzuzukukukiii J,J, KKKohoho nonono MMM, OiOiO dadada KKK,, TaTaTamamamaii i T,T,, MMMiyiyiyababbo o o S,S, YYYamamamamamamototo o o T,T,T, NNNakaka ai T.EnEnhahancncememenentt ofof tthehe bbinindidingng ooff trtrigiglylyceceriridede r-ricichh lilipopoprprototeieinsns ttoo ththee veveryry llowow ddenensisityty llipipopoproroteteinin
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
21
E, F) shows 36 week old mice. Control (A) ApoE-/- mice fed mice chow. ApoE-/- mouse fed high
fat, high cholesterol (HFHC) diet plus vehicle (B), HFHC plus 10mpk of D-PDMP (C). Thirty six
week old control ApoE-/-mice fed mice chow (D); placebo fed HFHC chow (E) and HFHC fed
mice treated with 10mpk D-PDMP (F). Note the arrows indicate marked increase in aortic wall
thickening in HFHC plus vehicle fed; placebo (B mice) mice at 20 weeks of age. This was
followed by a marked increase in Ca2+ deposits at 36 weeks of age (marked by asterisk) in this
group of mice as compared to control mice. This was prevented by feeding D-PDMP (F). Aortic
wall thickness and aortic wall stiffness in atherosclerotic mice is ameliorated by treatment with
D-PDMP shown in graphs (G) and (H). Quantitation of intima-media thickness (AoIMT) (G) and
pulse wave velocity (PWV) respectively (H), in ApoE-/- mouse on a mice chow diet (control 0.63
± 0.04 mm/4.37±0.26 m/s), HFHC diet plus vehicle (Placebo 1.21 ± 0.06 mm/6.38±0.89 m/s) (H),
HFHC diet plus treatment with 5 and 10mpk of D-DPMP (1.04 ± 0.04mm/6.07±0.5 m/s and 0.73 ±
0.04 mm/4.24±0.15 m/s). Note that both AoIMT and PWV increase continuously in placebo mice
from 12 weeks to 36 weeks and this was ameliorated upon treatment with D-PDMP in a
dose-dependent manner. A two-way RM ANOVA using the Bonferroni multiplo comparisons
post-test was performed.. * p 0.05, ** p 0.01, *** p 0.001; n = 3-5.
Figure 2. D-PDMP treatment ameliorates atherosclerotic plaque buildup and lumen volume in
ApoE-/- mice fed a western diet. Masson Trichrome stained ascending aortic rings of ApoE-/-
mouse: Control mice fed regular mice chow (A), mice fed high fat, high cholesterol (HFHC) diet
consisting of 2% fat and 1.25 cholesterol plus vehicle (Placebo) (B), HFHC plus 5mpk,
D-PDMP(C), and HFHC plus 10mpk, D-PDMP (D). Bar = 50 μm. Elastin fibers (E) decreased
following treatment with D-PDMP. Quantification of lumen area (F) reveals a decrease in lumen
pulse wave velocity (PWV) respectively (H), in ApoE-/- mouse on a mice chow dddieett t (cccononontrtrtrololol 00.63
± 0.04 mm/4.37±0.26 m/s), HFHC diet plus vehicle (Placebo 1.21 ± 0.06 mm/6.38±0.89 m/s) (H)
HFHFHCHCHC dddieieiett t plplpluus tttrerereata ment with 5 and 10mpk of DDD-D-D-DPMP (1.04 ± 0.0.0 04mmmmmm/6/ .07±0.5 m/s and 0.73 ±
00..0444 mm/4.244±0±00.15 mm/m s)s)). NoNoNotetee tthahaatt bobothth AooIIMMMT aaannnd PPPWVWVWV inncncrereeassse cocontntininuuououususlyly iin nn plpllacaccebebebo oo mmmice
frromomom 111222 weweeekekekss totoo 3336 6 weweweekeks aananddd thththisisis wwasasas aaammmelililiororo aaateeded uuupopoonn n trtreaeaeatmtmmenenent t t wwwiththh DDD-P-P-PDMDMDMPP P innn a
dose-dependedentntnt mmmanananneneer.r.r AAA tttwowow -w-wwayayy RRM M M ANANANOVOVOVA A A usususining g g ththhe e e BoBoBonfnn ererrrororoninini mmmululu titiplplplo oo cococompmm arisons
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
22
volume in the mice fed HFHC. Lumen area is significantly reduced due to increased plaque
accumulation in placebo mice aorta (Black arrow, B). Treatment with D-PDMP significantly
reduced and/or delayed medial thickening, elastin fibers, and plaque accumulation fragmentation
in a dose-dependent manner. A nonparametric one-way ANOVA using the Kruskal-Wallis test
and Dunn’s multiple comparison post-test were performed. * p 0.05, ** p 0.01, *** p 0.001;
n = 3-5.
Figure 3. Glycosphingolipid glycosyltransferase activity in ApoE-/- mice fed high fat and high
cholesterol diet with and without D-PDMP. Freshly harvested aorta tissue was stored in Tris HCl
buffer supplemented with a protease inhibitor cocktail, homogenized and the activity of
glucosylceramide synthase and lactosylceramide synthase were measured. Lactosylceramide
synthase activity significantly decreased at 20 weeks (A) and 36 weeks (C) following treatment
with D-PDMP. A significant change in glucosylceramide synthase activity was also observed at 20
weeks (B) and 36 weeks (D). In (E) and (F), treatment with D-PDMP decreases the mass of
glycosphingolipids in the liver of ApoE-/- mice fed a western diet. Glycosphingolipids were
extracted from 10 mg of harvested liver tissue and homogenized in CHCl3-MeOH, 2:1).
Following treatment with the enzyme SCDase and derivatization by OPA, mass was calculated via
liquid chromatography and tandem mass spectrometry against glucosylceramide and
lactosylceramide standards. A significant decrease from the placebo in the mass of
glucosylceramide was seen in the 5mpk treated and 10mpk treated mice fed the HFHC diet. A
nonparametric one-way ANOVA using the Kruskal-Wallis test and Dunn’s multiple comparison
post-test were performed. * p 0.05, ** p 0.01, *** p 0.001; n = 3.
buffer supplemented with a protease inhibitor cocktail, homogenized and the actitiivivvitytyt ooof f f
glucosylceramide synthase and lactosylceramide synthase were measured. Lactosylceramide
yyntntnthahahassese aaactctctivivivity y y sisisigng ificantly decreased at 20 wwweeeee kkks (A) and 36 weww ekksss ((C(C) following treatment
wwithhh D-PDMP.P AAA sssiggninin fifiicacacantntnt ccchhahangnggee e ininn gglucooosyyylceeraaamiidedede ssyyynththt asasse acctitiivivv ttyy wwwaass alalsososo ooobsbsbserrrvvevedd d aatat 20
weweeekekeksss (B(B( )) ) ananandd 3366 wwweeeeekskss ((D)D).. InInn (E(E(E))) anannd d (F(F(F)),), ttrerereatattmemem ntntt wwwititithh h D-D-D-PDPDDMPMPMP dddeeccrereasasa esese tthehehe mmaaasss s ofoff
glycosphingogolililipipipidsdsds iiin n thththe lililivevever ooof f f ApApApoEEE-/-//-- mimimicecece fffededed aaa wwwesesesteteterrrr n nn dididietee . GlGlGlycycycosososphphp innngogogolililipipipidsdd were
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
23
Figure 4. Plasma levels of oxidized LDL, cholesterol, triglycerides, and HDL-c in ApoE-/- mice
fed a high fat and high cholesterol diet with and without D-PDMP. Serum extracted from ApoE-/-
mice was analyzed for the presence of oxLDL (A), LDLc (B), triglycerides (C), and HDLc (D) on
microtiter plates. Serum concentrations of oxLDL were determined using an
immunohistochemical ELISA assay with an antibody against oxLDL. Concentrations of LDLc
triglycerides and HDLc concentrations were taken from microtiter readings following Wako kit
assays. Values are means ± SEM. * p 0.05, ** p 0.01, *** p 0.001; n = 3.
Figure 5. Effect of D-PDMP on the expression of hepatic genes, which play roles in cholesterol
and lipid metabolism. Expression of hepatic genes involved in LDL metabolism;
3-hydroxy-3-methylglutaryl-Coenzyme A reductase (Hmgcr), Low density lipoprotein receptor
(Ldlr), Sterol regulatory element binding transcription factor 1(Srebf1) , Sterol regulatory element
binding transcription factor 2(Srebf2) (A) HDL metabolism; apolipoprotein A-I(Apoa1),
scavenger receptor class B (SRB-1), CD36 antigen (B) VLDL and Triglyceride metabolism;
lipoprotein lipase (Lpl), very low density lipoprotein receptor (Vldlr) (C) and Cholesterol efflux
and uptake; ATP-binding cassette sub-family ABCA1(Abca1), cholesterol 7
alpha-hydroxylase(Cyp7A1) (D) as assessed by quantitative real-time PCR . Expressions of these
genes were significantly up regulated in mice treated with D-PDMP in a dose dependent manner.
Values are means ± SEM. A nonparametric one-way ANOVA using the Kruskal-Wallis test and
Dunn’s multiple comparison post-test were performed. * p 0.05, ** p 0.01, **** p 0.0001; n
= 3.
Figure 6. Western blot analysis of liver Srebp-2, LDLR, ABCG5 and ABCG8 proteins. Western
and lipid metabolism. Expression of hepatic genes involved in LDL metabolismm;;
3-hydroxy-3-methylglutaryl-Coenzyme A reductase (Hmgcr), Low density lipoprotein receptor
LLdldldlr)r)r), , StStSterereroolol regggulululata ory element binding transcriririppptiion factor 1(Srrebee f11))) , , SStS erol regulatory elemen
bbiindddini g transccririippptiioon n faff cctctororor 222(S(S(Srerebfbfbf22)2 ((AAA(( ) HHDDLL mmmetttaboooliliismsmm; apapoololippopoprrootteeininn AAA-I-I(A(AApopooa1a1a1),,
ccavavavenenengegegerr rererecceceptpttorrr cclalaassss BB (SSSRBRBRB-1-1-1),),) CCCD3D3D3666 ananantititigegeen (((BBB)) ) VLVLVLDLDLDL aanndnd TTTririggglyycycererrididideee mememettatabobooliiismsm;;;
ipoprotein liipapapaseses (((LpLpLpl)l)l ,, veeeryryry llowoww dedeensnsititity y y lilil popopoprprprotototeieie n n n rerecececeptptptororo (((VlVlV dldllr)r)r) (((CCC) ) ) ananand d ChChCholololesesestet rol efflux CCC
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

DOI: 10.1161/CIRCULATIONAHA.113.007559
24
blots illustrate D-PDMP induces the expression of Srebp-2 (A), LDL receptor (B), ABCG5 (C)
and ABCG8 (D) proteins in liver tissues. Values are means ± SEM. A nonparametric one-way
ANOVA using the Kruskal-Wallis test and Dunn’s multiple comparison post-test were performed.
* p 0.05, **** p 0.0001; n = 3.
Figure 7. Inhibiting glycosphingolipid synthesis inhibits atherosclerosis in rabbits. New Zealand
white rabbits (n=3) were fed a diet with and without cholesterol (2%) and fat (14%) and with and
without D-PDMP (10mpk) for three months. Marked increase in lactosylceramide levels in the
aorta tissue was observed (A) accompanied by extensive atherosclerosis and marked increase
(17-fold) in serum cholesterol levels (B) were found in rabbits fed the hyperlipidemic diet in
comparison to control and drug-treated rabbits at 3 months. This was prevented by treatment with
D-PDMP. Statistical analysis: In 7A, a nonparametric one-way ANOVA using the Kruskal-Wallis
test and Dunn’s multiple comparison post-test were performed. In 7B, a two-way repeated
measures ANOVA was performed with Bonferroni’s multiple comparisons post-test. * p 0.05,
**** p 0.0001; n = 3.
Figure 8. D-PDMP inhibits ox-LDL induced LCS activity and atherosclerosis development.
Oxidized LDL (ox-LDL) activates LacCer synthase (LCS) to produce large quantities of LacCer,
which stimulates the activity of NADPH oxidase. Concomitantly, there is an increase in the
generation of superoxide radicals (ROS). ROS mediates p44MAPK phosphorylation thereby
stimulating c-fos expression, promoting cell proliferation of vascular-smooth-muscle cells further
to angiogenesis and generation of atherosclerosis. D-PDMP, a potent inhibitor of LCS, impaired
ox-LDL mediated induction of LCS activity thereby inhibiting the above pathway.
17-fold) in serum cholesterol levels (B) were found in rabbits fed the hyperlipiddemememicic dddieieiet tt ininin
comparison to control and drug-treated rabbits at 3 months. This was prevented by treatment with
D-D-PDPDPDMPMPMP. StStStaaatisstititiccacal analysis: In 7A, a nonparamememetrrric one-way ANANNOVVVA A A uusing the Kruskal-Walli
eestt aand Dunn’n’ss mummultltipipipleee cccomomomppaparirisososonn n ppoposst-teeesttt weereee peererffoformrmmeeded. IInIn 77B,B a ttwowoo-w-wwayay repepepeaeaateeeddd
memeeasasa ururureses AAANONON VAVAVA wawwass peperfffororo mememed d d wiwiththt BBBoononfefeferrrrr onoonii’s mumumultltltiipiplelele cccomommpapap rrrissoonsnss pppososo t-t-tetetesst. ** pp p 00.0.005,5
**** p 0.000000001;1;1; nnn == 333..
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

GH
Figure 1
ControlPlacebo5mpk10 mpk
***
***
***
***
*** ******
***
**
***
***
******
***
*
***
***
*
GHH
CoCo tntrorollPl b
*****
****
***
****
*** ******
***
**
****
***
******
*****
*
***
*******
*
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

LumenLumen
E
Figure 2
*
*
*LLLLLLLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLume
LLLLLLLLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumenLumen
E
**
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

Figure 3
C E
FD
A
B FFFDDBBB
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from
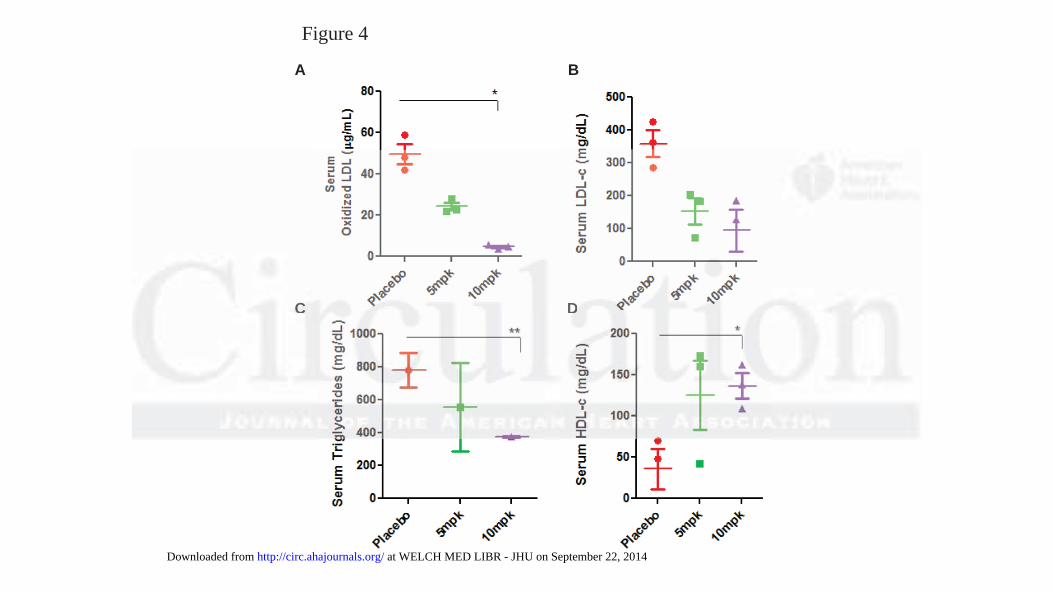
Figure 4
A B
C DCCC DDDDDD
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

Figure 5
A B
DC DDDCC
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

Figure 6
A
LDLr
GAPDH
LDLr
GAPDH
B
GAPDHH
LDLr
GAPDH
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

Figure 6, cont’d
ABCG5
GAPDH
ABCG5
GAPDH
C
ABCG8
GAPDH
ABCG8
GAPDH
D
ABCG5
GAPDH
GAPDH GAPDH
ABCG8
GAPDH
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from
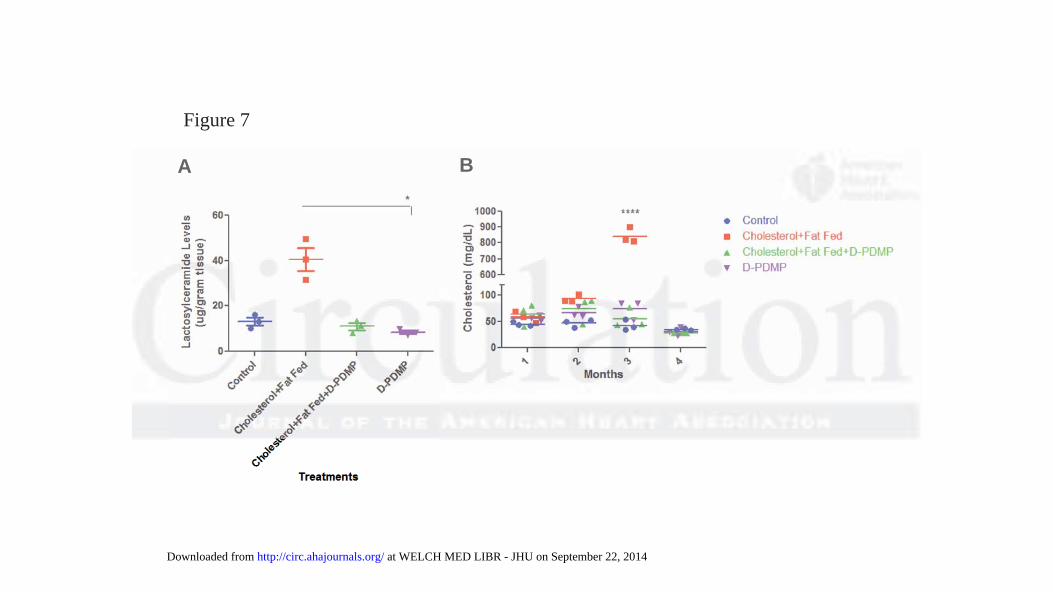
Figure 7
A BA B
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

Figure 8
at WELCH MED LIBR - JHU on September 22, 2014http://circ.ahajournals.org/Downloaded from

SUPPLEMENTAL MATERIAL

Supplemental Table 1. Primers used in the present study.
Gene Forward primer (5′-3′) Reverse primer (5′-3′)
LDLR CAGACTGCAAGGACAAGTCA GAG CCA TCT GCA CAC TGG AA
VLDLR CCACAGCAGTATCAGAAGTC GCCATCACTAAGAGC AAGAG
LPL AGTGTTTGTGAAATGCCATGA CA CGGATGCTTTCTTCTCTTGTTTG
APOA1 GTGGCTCTGGTCTTCCTGAC ACGGTTGAACCCAGAGTGTC
CD36 GCC AAG CTA TTG CGA CAT GA AGA ATC TCA ATG TCC GAG ACT
SREBP1 CCG TGG GCT GAG GAA GGA TGT GTA CTT GCC CAT GGC A
SREBP2 CATCCCTTGGGCCAGAAGTT TCCTTGGCTGCTGACTTGATC
Abca1 AACAGTTTGTGGCCCTTTTG AGTTCCAGGCTGGGGTACTT
HMGR TCTGGCAGTCAGTGGGAACTATT CCTCGTCCTTCGATCCAATTT
CYP7A1 CAGTGAAGGTCATGCTGGAG CGCAATGAAGAAGGTGACAA
SR-BI TCCCTCATCAAGCAGCAGGT TTCCACATCCCGAAGGACA

Supplemental Figure 1.
Supplemental Figure 1. Histopathological analysis showing Masson’s Trichrome stained left
coronary artery (LCA) of control ApoE-/- mouse on a chow diet (A), Apo E-/- on high fat high
cholesterol (HFHC) diet plus vehicle (Placebo) (B), Apo E-/- on HFHC diet plus treatment with
5 (C )and 10mpk of D-DPMP (D) respectively. Quantification of the cross sectional area of LCA
(E). The placebo mice (E) show a significant increase in the cross sectional area (CSA), loss of
smooth muscle cells, perivascular fibrosis, damaged wall, thrombosis and occlusion as compared
to control and treated mice. The 5 and 10 mpk of D-DPMP treated mice show and dose
dependent decline in cross sectional area and not significantly different to control mice.
** **
** **
*** ***
***
E F
C D
Copyright © 2022 FDOKUMEN




















