
Sphingosine Forms Channels in Membranes That Differ Greatly from Those Formed by Ceramide
Upload
independentCategory
view
1download
0
ANALYTICALBIOCHEMISTRY
Analytical Biochemistry 331 (2004) 275–282
www.elsevier.com/locate/yabio
Analysis of Xuorescently labeled glycosphingolipid-derived oligosaccharides following ceramide glycanase digestion
and anthranilic acid labeling
David C.A. Neville,¤ Virginie Coquard,1 David A. Priestman, Danielle J.M. te Vruchte, Daniel J. Sillence, Raymond A. Dwek, Frances M. Platt, and Terry D. Butters
Oxford Glycobiology Institute, Department of Biochemistry, University of Oxford, South Parks Road, Oxford OX1 3QU, UK
Received 13 February 2004
Abstract
Interest in cellular glycosphingolipid (GSL) function has necessitated the development of a rapid and sensitive method to bothanalyze and characterize the full complement of structures present in various cells and tissues. An optimized method to characterizeoligosaccharides released from glycosphingolipids following ceramide glycanase digestion has been developed. The procedure usesthe Xuorescent compound anthranilic acid (2-aminobenzoic acid; 2-AA) to label oligosaccharides prior to analysis using normal-phase high-performance liquid chromatography. The labeling procedure is rapid, selective, and easy to perform and is based on thepublished method of Anumula and Dhume [Glycobiology 8 (1998) 685], originally used to analyze N-linked oligosaccharides. It isless time consuming than a previously published 2-aminobenzamide labeling method [Anal. Biochem. 298 (2001) 207] for analyzingGSL-derived oligosaccharides, as the Xuorescent labeling is performed on the enzyme reaction mixture. The puriWcation of 2-AA-labeled products has been improved to ensure recovery of oligosaccharides containing one to four monosaccharide units, which wasnot previously possible using the Anumula and Dhume post-derivatization puriWcation procedure. This new approach may also beused to analyze both N- and O-linked oligosaccharides. 2004 Elsevier Inc. All rights reserved.
Keywords: Glycosphingolipids; HPLC; Anthranilic acid; Oligosaccharide analysis
Glycosphingolipids (GSLs)2 are found on alleukaryotic cell surfaces where they play important roles
0003-2697/$ - see front matter 2004 Elsevier Inc. All rights reserved.doi:10.1016/j.ab.2004.03.051
¤ Corresponding author. Fax: +44-1865-275216.E-mail address: [email protected] (D.C.A. Neville).1 Present address: E.S.T.B.B. II, Faculté des Sciences, Université Catholiq2 Abbreviations used: 2-AA, 2-aminobenzoic acid/anthranilic acid; 2-AB
gangliotetraglycosylceramide; Gal, galactose; GalNAc, N-acetylgalactosamine; �-Di-N-acetylneuraminylgangliotetraglycosylceramide; GD1b, II3-�-N-acetylne�-N-acetylneuraminyl�-2,8-N-acetylneuraminylgangliotetraglycosylceramide; ceramide; GD3, II3-�-N-acetylneuraminyl�-2,8-N-acetylneuraminyllactosylceramGM1a, II3-�-N-acetylneuraminylgangliotetraglycosylceramide; GM1a-Gc, II3-�neuraminylgangliotetraglycosylceramide; GM2, II3-�-N-acetylneuraminylgangsylceramide; GM3, II3-�-N-acetylneuraminyllactosylceramide; GM3-Gc, II3
�-2,8-N-acetylneuraminyl-IV3-�-N-acetylneuraminyl�-2, 8-N-acetylneuraminylgneuraminyl-IV3-�-N-acetylneuraminyl�-2, 8-N-acetylneuraminylgangliotetraglyIV3-�-N-acetylneuraminylgangliotetraglycosylceramide; GU, glucose unit; HPLlactotriglycosylceramide; Lc4, lactotetraglycosylceramide; LNT, lacto-N-tetraosglycolylneuraminic acid; IV3-�-Neu5Ac-nLc4, IV3-�-N-acetylneuraminylneolacphase; NPC1, Niemann–Pick disease, Type C; PBS, phosphate-buVered saline;
in mediating cell-signaling events [1]. The synthesis ofGSLs in the endoplasmic reticulum/Golgi is coordi-
ue de Lyon, 25 rue du Plat, 69288 Lyon, Cedex 02. France., 2-aminobenzamide; Cg3 (GA2), gangliotriglycosylceramide; Cg4 (GA1),Gb3, globotriaosylceramide; Gb4, globotetraosylceramide; GD1a, II3, IV3-�,uraminyl�-2,8-N-acetylneuraminylgangliotetra-glycosylceramide; GD1c, IV3-GD2, II3-�-N-acetylneuraminyl�-2,8-N-acetylneuraminylgangliotriglycosyl-
ide; Glc, glucose; GlcCer, glucosylceramide; GlcNAc, N-acetylglucosamine;-N-glycolylneuraminylgangliotetraglycosylceramide; GM1b, IV3-�-N-acetyl-
liotriglycosylceramide; GM2-Gc, II3-�-N-glycolylneuraminylgangliotriglyco--�-N-glycolylneuraminyllactosylceramide; GQ1b, II3-�-N-acetylneuraminylangliotetraglycosylceramide; GSL, glycosphingolipid; GT1a, II3-�-N-acetyl-cosylceramide; GT1b, II3-�-N-acetylneuraminyl�-2, 8-N-acetylneuraminyl-C, high-performance liquid chromatography; LacCer, lactosylceramide; Lc3,e; LNnT, lacto-N-neotetraose; NeuAc, N-acetylneuraminic acid; NeuGc, N-totetraglycosylceramide; nLc4, neolactotetraglycosylceramide; NP, normal-
SPE, solid-phase extraction; TFA, triXuoroacetic acid.

276 D.C.A. Neville et al. / Analytical Biochemistry 331 (2004) 275–282
nated by a series of cell-type and tissue-speciWc glyco-syltransferases, resulting in the generation of over 300diVerent species [2,3]. Although synthesized via commonprecursors, the ceramide moiety can vary in the alkyland acyl chain lengths and in saturation and hydroxyl-ation status, but it is the monosaccharide type, linkage,and anomericity that deWnes the species of GSL. Tocharacterize the composition of GSLs in eukaryotic cellswhere expression can be modulated by growth, diVeren-tiation, oncogenesis, and genetic deWciencies in both bio-synthetic and catabolic enzymes, the analysis of GSLoligosaccharides provides a useful approach. The avail-ability of endoglycosidases that release the oligosaccha-ride intact from GSLs coupled with sensitiveXuorescence labeling and high-resolution separation byHPLC has permitted the development of a robust buttime-consuming analysis of the major family of GSLs,the glucosphingolipids [4]. However, the limitations inthe current label of choice, 2-aminobenzamide, hasdemanded both improvements in Xuorescence labelingmethods and reWnement of separation techniques toimprove the analysis of GSLs from mouse tissue at thefemtomole level. The method that we describe here hasbeen optimized using ceramide glycanase and 2-anthra-nilic acid to allow oligosaccharide release followed byselective labeling of GSL glycans in the presence of saltsand detergent using a “one-pot” reaction strategy withimproved sensitivity.
Materials and methods
Reagents
Macrobdella decora ceramide glycanase was pur-chased from Calbiochem (Nottingham, Notts, UK). AllGSLs (at least 95% pure), anthranilic acid, and sodiumcyanoborohydride were purchased from Sigma (Poole,Dorset, UK). HPLC-grade acetonitrile, methanol, chlo-roform, boric acid (Aristar), and sodium acetate trihy-drate (Analar) ammonium acetate (Analar) werepurchased from VWR International (Lutterworth, Leics,UK). Oasis-HLB (10 mg, 1 cc) and SepPak C18 (100 mg,1 cc) solid-phase extraction (SPE) columns were pur-chased from Waters (Elstree, Herts, UK). DiscoveryDPA-6S (50 mg, 1 cc) solid-phase extraction columnswere purchased from Supelco (Poole, Dorset, UK).Peptide-N4-(N-acetyl-�-glucosaminyl)asparagine ami-dase (PNGase F) was purchased from NEB (Hitchin,Herts, UK) and used according to the manufacturersinstructions.
Glycosphingolipid extraction
Brain tissue from a symptomatic, 10-week-old NPC1mouse [5] was homogenized using a tight Dounce in
25 mM 2-(N-morpholino)ethanesulfonic acid, pH 6.5,containing 150 mM NaCl and then Bligh and Dyer [6]extracted with 3.2 volumes of chloroform/methanol(1:2.2, v/v) overnight at 4 °C. The phases were separatedby the addition of 1 volume of chloroform and 1 volumeof PBS. After spinning at 1000g for 10 min, the lowerphase was taken and dried down before being redissolvedin 50 �l chloroform/methanol (1:3, v/v). GSLs were puri-Wed using a SepPak C18 column as follows: the redis-solved lower phase was applied to a SepPak C18 columnthat had been preequilibrated with 2 £ 1 ml methanoland 2 £ 1 ml water and corresponding upper phases wereadded to the SepPak C18 column before washing with5 ml of water. Total GSLs were eluted in 5 ml chloro-form/methanol (1:3, v/v) and 1 ml of methanol.
Liver and brain samples from C57Bl/6 mice and abrain sample from a symptomatic (16-week-old) Sand-hoV [7] mouse were homogenized with a Polytron at aconcentration of 100 mg wet tissue weight/ml of Milli-Qwater. GSLs were then extracted using a modiWcation ofthe Svennerholm and Fredman [8] method that gavemaximum recovery of both neutral and charged GSLs;1 ml of tissue sample extract was mixed with 4 ml ofchloroform/methanol (1:2, v/v) to give a Wnal solventratio of chloroform/methanol/water of 4:8:3. The tubeswere mixed gently for 3 h at room temperature. Afterspinning at 1000g for 10 min at room temperature, 4 mlsupernatant was removed for further processing. This4 ml was separated into two phases by the addition of1 ml chloroform and 1 ml PBS. After spinning at 1000gfor 10 min, the lower phase was removed to a new tube,dried down under a stream of nitrogen and redissolvedin 50 �l chloroform/methanol (1:3, v/v). The upper phasewas added to and mixed with the redissolved lowerphase. GSLs were then further puriWed using a SepPakC18 column. The lower and upper phase mix wasapplied to a C18 column that had been preequilibratedwith 2 £ 1 ml methanol and 2 £ 1 ml water. The columnwas washed with 3 ml of water and total GSLs wereeluted with 1 ml chloroform/methanol (98:2, v/v), 2 mlchloroform/methanol (1:3, v/v), and 1 ml of methanol.Typically, 40 �l of total GSLs (1 mg wet weight tissueequivalent) was taken for ceramide glycanase digestion.
Ceramide glycanase digestion
Total GSLs from mouse tissues and GSL standards(10 �l, 0.5–2 mg/ml in chloroform/methanol (2:1, v/v))were dried down under a stream of nitrogen beforedigestion with 50 mU ceramide glycanase as describedpreviously [4]. Essentially, the GSLs, whether obtainedas a commercial standard or tissue-derived, were takento dryness in a small conical tube and redissolved, withmixing, in 10 �l incubation buVer (50 mM sodium ace-tate, pH 5.0, containing 1 mg/ml sodium cholate orsodium taurodeoxycholate). To this was added, with

D.C.A. Neville et al. / Analytical Biochemistry 331 (2004) 275–282 277
gentle mixing, 0.05 U ceramide glycanase in a further10 �l incubation buVer (giving a Wnal concentration of2.5 U/ml). One unit is deWned as the amount of enzymethat will hydrolyze 1.0 nmol of ganglioside, GM1, perminute at 37 °C. Incubations were performed at 37 °C for18 h.
Carbohydrate Xuorescent labeling
The ceramide-glycanase-released oligosaccharideswere labeled with anthranilic acid essentially asdescribed by Anumula and Dhume [9] but with the fol-lowing modiWcations. Anthranilic acid (30 mg/ml) wasdissolved in a solution of 4% sodium acetate trihydrate(w/v) and boric acid (2% w/v) in methanol. This solutionwas added to solid sodium cyanoborohydride (Wnal con-centration 45 mg/ml) and mixed to give the Wnal labelingmixture. The ceramide glycanase digestion mixture(20 �l) was made up to 30 �l with water; 80 �l of labelingmixture was added and the mixture was placed at 80 °Cfor 45–60 min. The reaction was allowed to cool to roomtemperature, 1 ml acetonitrile/water (97:3, v/v) wasadded, and the mixture was vortexed. Labeled oligosac-charides were puriWed following chromatographythrough Discovery DPA-6S columns. The columns werepreequilibrated with 2£ 1 ml acetonitrile. The sampleswere loaded using gravity Xow and allowed to dripthrough the column. The column was washed with 4£1 ml acetonitrile/water (99:1, v/v) followed by 0.5 ml ace-tonitrile/water (97:3, v/v). The labeled oligosaccharideswere eluted with 2£ 0.6 ml water and stored at either 4or ¡20 °C.
Carbohydrate analysis by HPLC
PuriWed 2-AA-labeled oligosaccharides were sepa-rated by normal-phase high-performance liquid chroma-tography (NP-HPLC) using a 4.6 £ 250-mm TSK gel-Amide 80 column (Anachem, Luton, Beds, UK) withslight modiWcations to the published method [10]. Thechromatography system consisted of a Waters Alliance2695 separations module and an in-line Waters 474 Xuo-rescence detector set at Ex� 360 nm and Em� 425 nm. Allchromatography was performed at 30 °C. Solvent A wascomposed of 20% 100 mM ammonium acetate, pH 3.85,in Milli-Q water and 80% acetonitrile. Solvent B wascomposed of 20% 100 mM ammonium acetate, pH 3.85,in Milli-Q water, 60% Milli-Q water, and 20% acetoni-trile. Gradient conditions for GSL analysis were as fol-lows: time D 0 min (t D 0), 86% solvent A (0.8 ml/min);t D 6, 86% solvent A (0.8 ml/min); t D 35, 54.7% solvent A(0.8 ml/min); t D 37, 5% solvent A (1 ml/min); t D 39, 5%solvent A (1 ml/min); t D 41, 86% solvent A (1 ml/min);t D 42, 86% solvent A (1.2 ml/min); t D 54, 86% solvent A(1.2 ml/min); t D 55, 86% solvent A (0.8 ml/min). Sampleswere injected in Milli-Q water/acetonitrile (2:8, v/v). All
chromatography was controlled and data were collectedand processed using Waters Millennium or Empowersoftware. Glucose unit values were determined, follow-ing comparison with a 2-AA-labeled glucose oligomerladder (derived from a partial hydrolysate of dextran)external standard, using Peak Time software (developedin-house). When analyzing N-linked oligosaccharidesthe following gradient conditions were used: t D 0, 86%solvent A (0.8 ml/min); t D 6, 86% solvent A (0.8 ml/min);t D 40, 47.6% solvent A (0.8 ml/min); t D 42, 5% solvent A(1 ml/min); t D 44, 5% solvent A (1 ml/min); t D 46, 86%solvent A (1 ml/min); t D 47, 86% solvent A (1.2 ml/min);t D 59, 86% solvent A (1.2 ml/min); and t D 60, 86% sol-vent A (0.8 ml/min).
Results and discussion
Fluorescent labeling with anthranilic acid (2-amino-benzoic acid; 2-AA) has previously been used to analyzeN-linked oligosaccharides released by PNGase F [9–11].The labeling reaction was performed directly on the reac-tion mixture, without any prior glycan cleanup to removeprotein, salts, and detergent. This same study demon-strated a superior sensitivity for 2-AA over 2-AB labelingand the eVect of time on the derivatization reaction.However, recovery of mono- to tetrasaccharides was notpossible due to the post-labeling processing methodemployed in the puriWcation of 2-AA labeled oligosac-charides. Labeling with 2-AA has also been used formonosaccharide analysis and the reaction kinetics formono- and disaccharide labeling investigated [12].Monosaccharide analysis required no sample puriWca-tion and, therefore, no losses were found prior to HPLC.We have modiWed the postlabeling cleanup procedure toenhance recovery of all saccharides and optimized HPLCconditions to separate GSL-derived oligosaccharides.
A number of experiments were performed to maxi-mize the recovery of mono- to tetrasaccharides following2-AA labeling. The use of a constant volume andamount of dextran hydrolysate (30 �l of a 0.25-mg/mlsolution in water) was chosen to compare resultsobtained when optimizing the puriWcation of labeledoligosaccharides, concentrations of labeling reagents,used and ratio of labeling solution to sample (data notshown). This led to the formulation of the labelingmethod outlined under Materials and methods. Ofparticular note was that the DPA-6S columns gavesigniWcantly better recovery of Glc1–4-2-AA-labeledoligosaccharides than Oasis-HLB columns when thepostlabeling puriWcation protocol is followed (data notshown). As many of the GSL-derived oligosaccharidesreleased following ceramide glycanase digestion werefound to elute between Glc1-2AA and Glc5-2AA (seeTable 1) DPA-6S columns were chosen as the preferredpostlabeling recovery matrix. There was, however, little

278 D.C.A. Neville et al. / Analytical Biochemistry 331 (2004) 275–282
diVerence in recovery of higher Glc oligomers whenusing either column matrix. This agreed with observa-tions in the original published method for N-linked oli-gosaccharide analysis [9].
The reaction conditions used for Xuorescent labelingof free oligosaccharides were essentially identical tothose Wrst proposed for N-linked oligosaccharides [9–11]and monosaccharides [12], where little or no desialyla-tion of mono- to tetra-sialylated oligosaccharides wasobserved. Labeling of commercially available ganglio-sides (GM1a, GD1a, GD1b, and GT1b), following cera-mide glycanase digestion, conWrmed these observations(data not shown) with no desialylated species detected.The pH of the labeling solution had previously beendetermined as approximately pH 5.3 [12], which is withinthe reported optimal pH range of 4–10 for reductive-ami-
Table 1Glycosphingolipid analysis using ceramide glycanase: NP-HPLCretention of GSL-derived 2-AA labeled oligosaccharides
Note. Glucose unit (GU) values were determined following com-parison with a 2-AA-labeled glucose oligomer ladder (derived from apartial hydrolysate of dextran) external standard. The determined GUvalues for N-acetylneuraminic acid (NeuAc)- and N-glycolylneurami-nic acid (NeuGc)-containing glycosphingolipid species were subject toslight variation (0.02–0.04 GU per NeuAc/NeuGc residue) dependenton individual NP-HPLC column batch numbers. GU values for neu-tral GSL-derived oligosaccharides were not aVected by diVerent col-umn batches.
a GSL nomenclature according to IUPAC-IUB Joint Commissionon Biochemical Nomenclature [3].
b Not determined.c The nature of the sialic acid designation was conWrmed following
digestion with Arthrobacter ureafaciens sialidase and/or TFA hydroly-sis.
d Human milk sugars, prepared in-house.
Glycolipid-derived oligosaccharidea
GU SD n
1 Lac 2.04 0.011 1202 Cg3 (GA2) 2.57 0.006 1153 Lc3 2.67 NDb ND4 Gb3(ceramide trihexoside) 2.82 0.010 945 GM3 2.93 0.010 936 GM2 3.29 0.014 557 GM3-Gcc 3.32 0.011 868 Gb4 (globoside) 3.45 0.008 899 Lc4 (LNTd) 3.53 ND ND
10 nLc4 (paragloboside, LNnTd) 3.57 ND ND11 GM2-Gcc 3.70 0.011 3412 Cg4 (GA1) 3.72 0.009 6513 GalNAc-Cg4 (GalNAc-GA1) 4.00 0.011 7614 GM1a 4.12 0.012 9015 GD3 4.26 0.014 1416 IV3-�-Neu5Ac-nLc4
(sialylparagloboside)4.32–4.35 ND ND
17 GM1b 4.56 0.013 6418 GM1a-Gcc 4.62 0.010 1519 GD1a 4.96 0.021 8720 GD1b 5.62 0.015 2321 GT1b 6.36 0.023 2222 GloboLex9, GL-Y [17] 6.93 0.034 1123 GQ1b 7.22 0.056 12
nation of aldehydes with sodium cyanoborohydride [13]and close to the optimal pH of 5.7 determined when label-ing monosaccharides in totally aqueous solution [14].
The conditions used for HPLC analysis were adaptedfrom those published by Yuen et al. [10] for the separationof N-linked oligosaccharides following 2-, 3-, or 4-amino-benzoic acid labeling where the mobile phase contained2.5 mM triXuoroacetic acid (TFA) and 10 mM ammo-nium acetate, pH 4.2 (Wnal pH 3.5). However, it was foundthat the presence of TFA was not necessary and thatchanging the pH of the ammonium acetate solution to3.85 facilitated oligosaccharide separation. Additionally,it was necessary to increase the concentration of ammo-nium acetate to 20 mM to separate Gb3 and GM3.
Previous experiments in this laboratory demonstratedthe applicability of using ceramide glycanase from theleech M. decora for the release of oligosaccharides fromboth commercially available GSLs and biologicallyderived material [4]. The major rate-limiting step in bothprocedures is the time taken to obtain GSLs from eithertissue or cell samples. However, once the free oligosac-charides have been released from the puriWed GSLs, the2-AB labeling, recovery, and HPLC analysis run timesare very time consuming. A comparison of the timesrequired for either 2-AB or 2-AA labeling, postGSL puri-Wcation, is shown in Fig. 1. This demonstrates that with
Fig. 1. Comparison of the time taken to process GSL-derived oligosac-charides using either 2-AB or 2-AA Xuorescent labeling. GSLs wereisolated as described under Materials and methods and subjected toceramide glycanase digestion. The minimum times required to processand Xuorescently derivatize the GSL-derived oligosaccharides toobtain either 2-AB- or 2-AA-labeled oligosaccharides are shown.

D.C.A. Neville et al. / Analytical Biochemistry 331 (2004) 275–282 279
increasing numbers of samples there would be a signiW-cant time saving using the 2-AA as opposed to the 2-ABexperimental procedure. Performing the labeling reactionhad not, however, been applied to a ceramide glycanasedigest mixture without prior sample cleanup [4].
The applicability of this method was demonstratedinitially on a standard mix of commercially availableGSLs. Fig. 2 shows the spectrum obtained using a stan-dard mix of neutral and mono- to tri-sialylated GSLs.
Fig. 2. HPLC separation of 2-AA-labeled oligosaccharides obtainedfrom ceramide glycanase digestion of commercially available GSLs.Commercially available GSLs were digested overnight with ceramideglycanase, labeled with 2-AA, and puriWed following DPA-6S columnchromatography as described under Materials and methods. The GUvalues for each species are shown in Table 1. The elution position ofeach 2-AA-labeled oligosaccharide relative to the GU values(obtained from a 2-AA-labeled dextran hydrolysate external standard)is shown.
The GU values obtained for these GSLs and a numberof other GSL-derived oligosaccharides are shown inTable 1. Glc is set to 1 GU for all analyzes. These dataare similar to those reported by Wing et al. [4] and dem-onstrate that this method can be substituted for the 2-AB method. The GU values obtained are loosely basedon the number of monosaccharides present and on thenumber of charged residues present in each structure.However, there are no hard and fast rules that might beapplied to calculate the theoretical size of any particularGSL-derived oligosaccharide unlike those applied to N-linked oligosaccharides [15]. The relationship betweenthe various biosynthetic pathways for GSL formation,starting with GlcCer and pertaining to the GSLs identi-Wed in Figs. 2 and 4, is shown in Fig. 3. Unlike thesequential addition of monosaccharides observed in thebiosynthesis of N-linked oligosaccharides, a number ofdiVerent pathways are invoked in the formation of thediVerent series of GSLs. These pathways are dependenton the action of speciWc glycosyltransferases at deWnedpoints in GSL biosynthesis [2,16] with the major branchpoints being postLacCer (O, globo, and lacto/neolactoseries) and GM3 (a and b series).
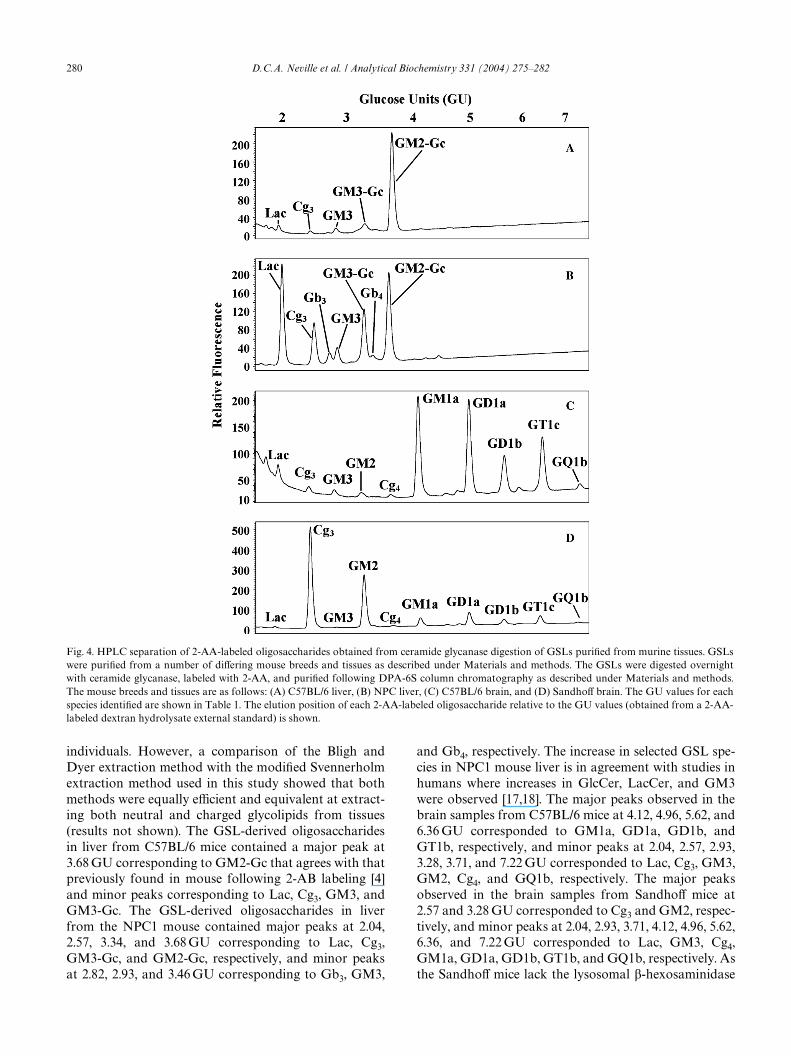
A number of diVerent mouse tissues from diVeringmouse strains were chosen to validate the experimentalapproach using tissue-derived samples. GSLs isolatedfrom mouse livers (C57BL/6 and NPC1, Figs. 4A and B,respectively) and brains (C57BL/6 and SandhoV; Figs.4C and D, respectively) were analyzed. Two diVeringextraction methods were employed in this study due tosamples being processed at diVerent times and by diVerent
Fig. 3. GSL biosynthetic pathway. The major pathways involved in GSL biosynthesis are delineated and demonstrate the relationships between thediVerent GSL series. Only the GSLs derived from GlcCer are shown. GSL-derived oligosaccharides were found for all except GD1c and GD2. As thelacto/neolacto-series GSLs may be further processed to possess larger oligosaccharides only the initial steps are shown. GSL nomenclature accordingto IUPAC-IUB Joint Commission on Biochemical Nomenclature [3].
280 D.C.A. Neville et al. / Analytical Biochemistry 331 (2004) 275–282
individuals. However, a comparison of the Bligh andDyer extraction method with the modiWed Svennerholmextraction method used in this study showed that bothmethods were equally eYcient and equivalent at extract-ing both neutral and charged glycolipids from tissues(results not shown). The GSL-derived oligosaccharidesin liver from C57BL/6 mice contained a major peak at3.68 GU corresponding to GM2-Gc that agrees with thatpreviously found in mouse following 2-AB labeling [4]and minor peaks corresponding to Lac, Cg3, GM3, andGM3-Gc. The GSL-derived oligosaccharides in liverfrom the NPC1 mouse contained major peaks at 2.04,2.57, 3.34, and 3.68 GU corresponding to Lac, Cg3,GM3-Gc, and GM2-Gc, respectively, and minor peaksat 2.82, 2.93, and 3.46 GU corresponding to Gb3, GM3,
and Gb4, respectively. The increase in selected GSL spe-cies in NPC1 mouse liver is in agreement with studies inhumans where increases in GlcCer, LacCer, and GM3were observed [17,18]. The major peaks observed in thebrain samples from C57BL/6 mice at 4.12, 4.96, 5.62, and6.36 GU corresponded to GM1a, GD1a, GD1b, andGT1b, respectively, and minor peaks at 2.04, 2.57, 2.93,3.28, 3.71, and 7.22 GU corresponded to Lac, Cg3, GM3,GM2, Cg4, and GQ1b, respectively. The major peaksobserved in the brain samples from SandhoV mice at2.57 and 3.28 GU corresponded to Cg3 and GM2, respec-tively, and minor peaks at 2.04, 2.93, 3.71, 4.12, 4.96, 5.62,6.36, and 7.22 GU corresponded to Lac, GM3, Cg4,GM1a, GD1a, GD1b, GT1b, and GQ1b, respectively. Asthe SandhoV mice lack the lysosomal �-hexosaminidase
Fig. 4. HPLC separation of 2-AA-labeled oligosaccharides obtained from ceramide glycanase digestion of GSLs puriWed from murine tissues. GSLswere puriWed from a number of diVering mouse breeds and tissues as described under Materials and methods. The GSLs were digested overnightwith ceramide glycanase, labeled with 2-AA, and puriWed following DPA-6S column chromatography as described under Materials and methods.The mouse breeds and tissues are as follows: (A) C57BL/6 liver, (B) NPC liver, (C) C57BL/6 brain, and (D) SandhoV brain. The GU values for eachspecies identiWed are shown in Table 1. The elution position of each 2-AA-labeled oligosaccharide relative to the GU values (obtained from a 2-AA-labeled dextran hydrolysate external standard) is shown.

D.C.A. Neville et al. / Analytical Biochemistry 331 (2004) 275–282 281
necessary to remove the nonreducing terminus GalNacresidue, the storage of Cg3 and GM2 is diagnostic of thisdisease’s phenotype [7]. The GSL-derived oligosaccha-ride structures were conWrmed following enzyme diges-tions (single and/or combination) with sialidase, �-hexosaminidase, �-galactosidase, or �-galactosidase(data not shown).
The sensitivity of detection when labeling knownamounts of standard sugars is between 20 and 50 fmol.This is comparable to that obtained previously, whenusing ceramide glycanase followed by 2-AB labeling,where 160 fmol of GSL-derived oligosaccharide wasdetected [4]. However, there are a number of other fac-tors that can inXuence method sensitivity such as (a)substrate concentration (there is a lack of linearity abovesubstrate concentrations of 150 �M) [4], (b) amount of
protein (typically 1 mg wet tissue equivalent is taken forGSL analysis but consistent results have been achievedwith cellular protein amounts above 100 �g) and (c)known diVerences in GSL expression when analyzingdiVerent species and tissue samples [19–22].
The procedure outlined above has also been appliedto N-linked oligosaccharides derived from �1-acid gly-coprotein and transferrin sugars following release usinghydrazinolysis (Figs. 5A and B, respectively) and toribonuclease B and fetuin oligosaccharides followingPNGase F digestion (Figs. 5C and D, respectively). Tooptimally separate N-linked oligosaccharides, the gradi-ent conditions for HPLC analysis were extended by afurther 5–42 min prior to washing and reequilibration.This demonstrates that both large and small oligosac-charides can be analyzed using this method. Preliminary
Fig. 5. HPLC separation of 2-AA-labeled N-linked oligosaccharides. N-linked oligosaccharides released from (A) ovalbumin and (B) transferrin fol-lowing hydrazinolysis and from (C) ribonuclease B and (D) fetuin following PNGase F digestion were labeled with 2-AA and puriWed followingDPA-6S column chromatography as described under Materials and methods. The elution position of each 2-AA-labeled oligosaccharide relative tothe GU values (obtained from a 2-AA-labeled dextran hydrolysate external standard) is shown.

282 D.C.A. Neville et al. / Analytical Biochemistry 331 (2004) 275–282
work has shown that free, reducing O-linked oligosac-charides, obtained following release from glycoproteinsusing an optimized hydrazinolysis procedure [23], mayalso be analyzed using this approach (data not shown).
Conclusions
This study demonstrated that analysis of GSL-derived oligosaccharides is possible by use of 2-AA Xuo-rescent labeling followed by NP-HPLC. The method hasa number of advantages over the 2-AB approach. First,processing samples using the 2-AA approach is muchquicker. This is clearly shown in Fig. 1, where the totalamount of time from digest to HPLC can be as little as2 h for 2-AA. With the 2-AB method, the time fromdigest to HPLC can be as great as 6.5 h. Second, theHPLC run time is 55 min when using 2-AA and either 2or 3 h with the 2-AB approach. The methodology canalso be applied to N- and O-linked oligosaccharides.
References
[1] M.L. Allende, R.L. Proia, Lubricating cell signaling pathways withgangliosides, Curr. Opin. Struct. Biol. 12 (2002) 587–592.
[2] S. Ichikawa, Y. Hirabayashi, Glucosylceramide synthase and gly-cosphingolipid synthesis, Trends Cell Biol. 8 (1998) 198–202.
[3] M.A. Chester, IUPAC-IUB Joint Commission on BiochemicalNomenclature (JCBN). Nomenclature of glycolipids. Recommen-dations 1997, Eur. J. Biochem. 257 (1998) 293–298.
[4] D.R. Wing, B. Garner, V. Hunnam, G. Reinkensmeier, U. Anders-son, D.J. Harvey, R.A. Dwek, F.M. Platt, T.D. Butters, High-per-formance liquid chromatography analysis of gangliosidecarbohydrates at the picomole level after ceramide glycanasedigestion and Xuorescent labeling with 2-aminobenzamide, Anal.Biochem. 298 (2001) 207–217.
[5] P.G. Pentchev, A.D. Boothe, H.S. Kruth, H. Weintroub, J. Stivers,R.O. Brady, A genetic storage disorder in BALB/C mice with ametabolic block in esteriWcation of exogenous cholesterol, J. Biol.Chem. 259 (1984) 5784–5791.
[6] E.G. Bligh, W.J. Dyer, A rapid method of total lipid extractionand puriWcation, Can. J. Biochem. Physiol. 37 (1959) 911–917.
[7] K. Sango, S. Yamanaka, A. HoVmann, Y. Okuda, A. Grinberg, H.Westphal, M.P. McDonald, J.N. Crawley, K. SandhoV, K. Suzuki,et al., Mouse models of Tay-Sachs and SandhoV diseases diVer inneurologic phenotype and ganglioside metabolism, Nat. Genet. 11(1995) 170–176.
[8] L. Svennerholm, P. Fredman, A procedure for the quantitative isola-tion of brain gangliosides, Biochim Biophys Acta 617 (1980) 97–109.
[9] K.R. Anumula, S.T. Dhume, High resolution and high sensitivitymethods for oligosaccharide mapping and characterization bynormal phase high performance liquid chromatography followingderivatization with highly Xuorescent anthranilic acid, Glycobiol-ogy 8 (1998) 685–694.
[10] C.T. Yuen, C.K. Gee, C. Jones, High-performance liquid chro-matographic proWling of Xuorescent labeled N-glycans on glyco-proteins, Biomed. Chromatogr. 16 (2002) 247–254.
[11] K.R. Anumula, P. Du, Characterization of carbohydrates usinghighly Xuorescent 2-aminobenzoic acid tag following gel electro-phoresis of glycoproteins, Anal. Biochem. 275 (1999) 236–242.
[12] K.R. Anumula, Quantitative determination of monosaccharides inglycoproteins by high-performance liquid chromatography withhighly sensitive Xuorescence detection, Anal. Biochem. 220 (1994)275–283.
[13] R.F. Borch, M.D. Bernstein, H.D. Durst, The cyanohydridoborateanion as a selective reducing agent, J. Am. Chem. Soc. 93 (1971)2897–2904.
[14] K. Sato, A. Okuba, S. Yamazaki, Optimization of derivatizationwith 2-aminobenzoic acid for determination of monosaccharidecomposition by capillary electrophoresis, Anal. Biochem. 262(1998) 195–197.
[15] G.R. Guile, P.M. Rudd, D.R. Wing, S.B. Prime, R.A. Dwek, Arapid high-resolution high-performance liquid chromatographicmethod for separating glycan mixtures and analyzing oligosaccha-ride proWles, Anal. Biochem. 240 (1996) 210–226.
[16] A. Varki, Biological roles of oligosaccharides: all of the theoriesare correct, Glycobiology 3 (1993) 97–130.
[17] M.T. Vanier, Biochemical studies in Niemann-Pick disease. I.Major sphingolipids of liver and spleen, Biochim. Biophys. Acta750 (1983) 178–184.
[18] M.T. Vanier, G. Millat, Niemann-Pick disease type C, Clin. Genet.64 (2003) 269–281.
[19] M. Sekine, T. Ariga, T. Miyatake, R. Kase, A. Suzuki, T. Yamakawa,An interspecies comparison of gangliosides and neutral glycolipidsin adrenal glands, J. Biochem. (Tokyo) 97 (1985) 1219–1227.
[20] A. Markotic, A. Marusic, J. Tomac, J. Muthing, Gangliosideexpression in tissues of mice lacking beta2-microglobulin, Clin.Exp. Immunol. 128 (2002) 27–35.
[21] M. Saito, H. Kitamura, K. Sugiyama, Liver gangliosides of vari-ous animals ranging from Wsh to mammalian species, Comp. Bio-chem. Physiol. B Biochem. Mol. Biol. 129 (2001) 747–758.
[22] J. Muthing, U. Maurer, K. Sostaric, U. Neumann, H. Brandt, S.Duvar, J. Peter-Katalinic, S. Weber-Schurholz, DiVerent distribu-tions of glycosphingolipids in mouse and rabbit skeletal muscledemonstrated by biochemical and immunohistological analyzes, J.Biochem. (Tokyo) 115 (1994) 248–256.
[23] A.H. Merry, D.C. Neville, L. Royle, B. Matthews, D.J. Harvey,R.A. Dwek, P.M. Rudd, Recovery of intact 2-aminobenzamide-labeled O-glycans released from glycoproteins by hydrazinolysis,Anal. Biochem. 304 (2002) 91–99.
Copyright © 2022 FDOKUMEN




















