Influencia de la conexion rio-laguna sobre la ictiocenosis lacustre en la varzea del rio Ichilo...
16
ARTÍCULOS CIENTÍFICOS - TÉCNICOS Rev. Bol. Ecol. 17: 33- 48, 2005 33 RESUMEN El río Ichilo es un sistema “río-llanura de inundación” de la Cuenca Amazónica boliviana. En las lagunas de su várzea se establece una diversa ictiocenosis, que es afectada por el intercambio de aguas que ocurre entre el río y las lagunas durante la época de inundación. El presente trabajo presenta una primera descripción de los efectos que se observan en la composición y abundancia de la ictiocenosis lacustre, tanto a nivel taxonómico como de gremios tróficos y de estrategias de vida. El estudio se llevó a cabo en tres lagunas de várzea conectadas de diferente manera al río Ichilo (por cárcava, por canal y de forma indirecta). Las capturas fueron realizadas mediante una batería de siete redes de diferentes aberturas de malla durante los períodos de aguas bajas (octubre 2000) y aguas altas (marzo 2001). Se capturaron 56 especies de peces pertenecientes a 5 órdenes y 16 familias. Las familias Characidae y Curimatidae fueron las más importantes en las tres lagunas. El gremio trófico dominante fue el de los omnívoros en la laguna conectada por cárcava, los detritívoros en la laguna conectada por canal y los carnívoros en la laguna de conexión indirecta. La estrategia de vida dominante en las tres lagunas, independientemente del período de captura, fue la estrategia periódica. Palabras claves: peces, río Ichilo, várzea, conexión río – laguna ABSTRACT Ichilo River is a “river-floodplain system” located in Bolivian part of the Amazon basin. A diverse fish community is stablished in the várzea lakes of this system, community which is affected by the exchange of water that occurs between the main channel of the river and its várzea lakes during the flood pulse. We present a preliminary description of the effects of the flood pulse on the composition and abundance of the community in taxonomic, trophic and life traits terms. The study was carried out in three várzea lakes with different degrees of connection to the river (direct and indirect connection). Fish were caught with a battery of seven gillnets with different mesh size, during the low-water (October 2000) and high-water (March 2001) periods. Fifty-six species belonging to 5 orders and sixteen families were caught. The families Characidae and Curimatidae were the most abundant in the three lakes. The omnivores were the dominant trophic guild in the lake with direct connection to the river; detritivorous fishes were dominant in the lake with a connecting channel; and carnivores were dominant in the lake with indirect connection. The dominant life trait in all lakes was the periodic strategy. Key words: fish community, Ichilo river, river-lake connection, floodplain INFLUENCIA DE LA CONEXIÓN RÍO – LAGUNA SOBRE LA ICTIOCENOSIS LACUSTRE EN LA VÁRZEA DEL RÍO ICHILO (COCHABAMBA-BOLIVIA) INFLUENCE OF RIVER-LAKE CONNECTION OVER THE FISH COMMUNITY IN THE RIVER ICHILO FLOODPLAIN Fernando Carvajal 1* y Mabel Maldonado 1 1 Unidad de Limnología y Recursos Acuáticos, Universidad Mayor de San Simón. Casilla 992. Cochabamba, Bolivia. * E-mail para correspondencia : [email protected]
Transcript of Influencia de la conexion rio-laguna sobre la ictiocenosis lacustre en la varzea del rio Ichilo...

ARTÍCULOS CIENTÍFICOS - TÉCNICOS Rev. Bol. Ecol. 17: 33- 48, 2005
33
RESUMEN
El río Ichilo es un sistema “río-llanura de inundación” de la Cuenca Amazónica boliviana. En las lagunas de suvárzea se establece una diversa ictiocenosis, que es afectada por el intercambio de aguas que ocurre entre elrío y las lagunas durante la época de inundación. El presente trabajo presenta una primera descripción de losefectos que se observan en la composición y abundancia de la ictiocenosis lacustre, tanto a nivel taxonómicocomo de gremios tróficos y de estrategias de vida. El estudio se llevó a cabo en tres lagunas de várzea conectadasde diferente manera al río Ichilo (por cárcava, por canal y de forma indirecta). Las capturas fueron realizadasmediante una batería de siete redes de diferentes aberturas de malla durante los períodos de aguas bajas (octubre2000) y aguas altas (marzo 2001). Se capturaron 56 especies de peces pertenecientes a 5 órdenes y 16 familias.Las familias Characidae y Curimatidae fueron las más importantes en las tres lagunas. El gremio trófico dominantefue el de los omnívoros en la laguna conectada por cárcava, los detritívoros en la laguna conectada por canal ylos carnívoros en la laguna de conexión indirecta. La estrategia de vida dominante en las tres lagunas,independientemente del período de captura, fue la estrategia periódica.
Palabras claves: peces, río Ichilo, várzea, conexión río – laguna
ABSTRACT
Ichilo River is a “river-floodplain system” located in Bolivian part of the Amazon basin. A diverse fish communityis stablished in the várzea lakes of this system, community which is affected by the exchange of water that occursbetween the main channel of the river and its várzea lakes during the flood pulse. We present a preliminarydescription of the effects of the flood pulse on the composition and abundance of the community in taxonomic,trophic and life traits terms. The study was carried out in three várzea lakes with different degrees of connectionto the river (direct and indirect connection). Fish were caught with a battery of seven gillnets with different meshsize, during the low-water (October 2000) and high-water (March 2001) periods. Fifty-six species belonging to 5orders and sixteen families were caught. The families Characidae and Curimatidae were the most abundantin the three lakes. The omnivores were the dominant trophic guild in the lake with direct connection to the river;detritivorous fishes were dominant in the lake with a connecting channel; and carnivores were dominant in thelake with indirect connection. The dominant life trait in all lakes was the periodic strategy.
Key words: fish community, Ichilo river, river-lake connection, floodplain
INFLUENCIA DE LA CONEXIÓN RÍO – LAGUNA SOBRE LAICTIOCENOSIS LACUSTRE EN LA VÁRZEA DEL RÍO ICHILO
(COCHABAMBA-BOLIVIA)
INFLUENCE OF RIVER-LAKE CONNECTION OVER THEFISH COMMUNITY IN THE RIVER ICHILO FLOODPLAIN
Fernando Carvajal1* y Mabel Maldonado1
1 Unidad de Limnología y Recursos Acuáticos, Universidad Mayor de San Simón. Casilla 992. Cochabamba, Bolivia.
* E-mail para correspondencia : [email protected]

REVISTA BOLIVIANA DE ECOLOGÍA Y CONSERVACIÓN AMBIENTAL
34
(SAINT-PAUL et al., 2000), afectando su composición,abundancia, estructura trófica y las historias de vida delos peces.
Las ictiocenosis de las lagunas de la llanura de inun-dación del río Ichilo son poco conocidas aún. MALDO-NADO et al. (1997) indican que en ellas los Chara-ciformes y Siluriformes son los grupos mas diversos yabundantes; desde el punto de vista trófico, los de-tritívoros y carnívoros son los grupos mas importantes(AYALA et al., 2000).
Varias especies de peces de estos ambientes sustentanactividades de pesca de subsistencia regional y sopor-tan la segunda pesquería comercial más importante deBolivia (Puerto Villarroel), a pesar de ello, aún no existeel conocimiento científico-técnico necesario para me-jorar o conservar este recurso. Por ello, la informaciónbiológica sobre los peces y su ecología en la planiciede inundación es necesaria para planear esquemasde utilización de los recursos ícticos.
En este sentido, el presente estudio pretende aportarelementos ecológicos básicos sobre los factores quepodrían regular los cambios de la ictiocenosis, entérminos de su composición taxonómica y trófica, y delas estrategias de vida de los peces, en relación con elintercambio río-laguna en la várzea del río Ichilo.
ÁREA DE ESTUDIO
El río Ichilo es el límite natural entre los departamen-tos de Cochabamba y Santa Cruz, ubicado entre los15˚57’ y 17˚50’ latitud sur y los 65˚21’ y 63˚56’ de longi-tud oeste.
Según MALDONADO et al. (1996), el río Ichilo pertenecea los sistemas “rio-planicie de inundación”, ubicado muycerca de las cabeceras fluviales, por lo que el pulso deinundación al que está sometido, es notoriamenteirregular condicionando inundaciones de tipo polimo-dal y de corta duración.
La región presenta bioclima pluvial (NAVARRO YMALDONADO, 2002), con una precipitación pluvialanual de 2 497 mm y una temperatura media anualde 23,6 ˚C. Los meses de abril y noviembre tienenuna distribución de precipitación regular siendo eneroy febrero los meses más lluviosos y con mayor esco-rrentía (CUMAT, 1985).
INTRODUCCIÓN
El río Ichilo es el sistema fluvial más importante del De-partamento de Cochabamba, tanto por su extensión ycaudal, como por la actividad socio-económica a la queestá vinculado. Numerosas lagunas originadas enmeandros abandonados forman parte fundamental delsistema (MALDONADO et al., 1996), y proveen un im-portante hábitat de cría para un gran número, posible-mente para la mayoría, de las especies de peces deríos de las tierras bajas. Ellos explotan las condicionesbenignas de la planicie de inundación, puesto que tien-de a ser menos turbulenta, más productiva y contienemenos depredadores que los canales de los ríos (SAINT-PAUL & PETER, 1979; JUNK, 1980; ARAUJO-LIMA etal., 1986; MACHADO-ALLISON, 1987; MAGO-LECCIA,1994; WINEMILLER & JEPSEN, 1998).
De esta manera, las lagunas de la planicie de inundaciónconectadas con el río son sistemas abiertos que puedenintercambiar organismos. Por lo tanto, niveles altos deagua incrementan el tamaño del ambiente acuático ytambién inyectan nutrientes que estimulan el rápidocrecimiento de microorganismos, invertebrados y plan-tas, proveyendo abundante alimento y protección paralos peces durante el nivel hidrológico más alto (WEL-COMME, 1979).
Los grados y tipos de conexión río-laguna dependende la geomorfología de la llanura aluvial en un áreadeterminada y del origen y movimiento de las aguas.DRAGO (1981) reconoce dos grados de conexión:directo e indirecto. La conexión directa se presenta enlas áreas menos evolucionadas del valle aluvial, exis-tiendo tres tipos de conexión directa: por una boca(extremo de cauce abandonado que aún no ha sidocolmatado), por una cárcava de erosión (cortes de al-bardones producidos por el flujo de inundación) y porcauces temporarios. El grado de conexión indirecta esel resultado de la existencia de los tipos anteriores enzonas geomorfológicamente más evolucionadas, don-de cárcavas de erosión o cauces temporarios han sidoprácticamente colmatados y su superficie se hallageneralmente cubierta por vegetación acuática y pa-lustre. En los sectores más alejados de la llanura alu-vial los cursos temporarios han desaparecido quedan-do las lagunas aisladas de la influencia de las corrien-tes principales, por lo que reciben aportes cuando losniveles hidrométricos son altos y solamente en formade aguas de desborde no encauzadas.
De esta manera, la inundación llega a ser el factor crí-tico responsable de las fluctuaciones en las ictiocenosis

35
CARVAJAL, F. y M. MALDONADO: Conexión río-laguna e Ictiocenosis en El Río Ichilo
Figura 1. Ubicación de las lagunas estudiadas en la várzea del río Ichilo
Según MALDONADO et al. (1997) en las lagunas de lallanura de inundación del río Ichilo se reconoce elsiguiente calendario hidrológico: entre noviembre y ene-ro se produce el llenado de estos ambientes; pos-teriormente durante el mes de febrero toma lugar lainundación; a continuación marzo y julio son los me-ses del drenaje de estas lagunas y durante los mesesde agosto a octubre ocurre el aislamiento del canal delrío.
El estudio se llevó a cabo en tres lagunas de origenmeándrico en la várzea del río Ichilo (Fig. 1), a una al-titud promedio de 200 m.s.n.m. La laguna Río Viejo(16˚ 50’ S y 64˚ 47’ O) se ubica frente a Puerto Villa-rroel y posee una conexión directa (cárcava de ero-sión) con el río. La laguna Bufeos (16˚ 25’ S y 64˚ O)también posee conexión directa a través de un caucetemporario, y por último, la laguna Redonda (16˚ 25’ Sy 64˚ 42’ O) es de comunicación indirecta, solamente através de la llanura. Estas dos últimas lagunas están ubi-cadas aproximadamente a 50 Km de Puerto Villarroel.

REVISTA BOLIVIANA DE ECOLOGÍA Y CONSERVACIÓN AMBIENTAL
36
MÉTODOS
Las capturas se llevaron a cabo en dos épocas hidro-lógicas extremas: aguas bajas (octubre y diciembre de2000) y de aguas altas (marzo de 2001).
Se utilizó una batería de 7 redes agalleras de fondocon 10, 20, 40, 50, 70, 80 y 100 mm de abertura demalla. Las dos primeras con longitudes de 10 m, latercera de 20 m y las restantes con 30 m de largo (to-das con 1.5 m de alto).
La pesca se realizó por un lapso de 3 días en cadalaguna, excepto en la laguna Río Viejo en la cual setrabajó solamente dos días durante la época de aguasaltas por motivos ajenos a nuestro control.
Todas las redes fueron colocadas perpendicularmen-te a la orilla de las lagunas y usualmente en lugaresbajos (1.5-2.5 m de profundidad). La red de 10 mm secolocó durante un período de 20 min, la red de 20 mmdurante 15 min y la de 40 mm por 90 minutos (estosperíodos se establecieron para evitar la saturación yexcesiva mortandad de los peces), las redes con ma-yor abertura de malla permanecieron extendidas las24 horas del día.
El grupo de las tres primeras redes se colocaron entrelas 10:00 – 12:00 y 17:00 – 19:00 horas, y las de mayortamaño de malla se revisaron a las 8:00, 17:00 y 20:00horas de cada día.
Estos horarios se eligieron en base a los resultados ob-tenidos por REJAS Y MALDONADO (2000), quienesestudiaron la composición y estructura de la comuni-dad de peces de laguna Bufeos con casi idénticos mé-todos de pesca.
Los peces se pesaron y luego fueron fijados en forma-lina al 10% para su traslado e identificación taxonómi-ca con claves especializadas tales como GOSLINE(1940), MENEZES (1969), MEES (1974), FINK &MACHADO-ALLISON (2001), GERY (1977), WAIS &CASTELLO (1984), LAUZANNE & LOUBENS (1985),REIS (1989), VARI (1984, 1989, 1991, 1992), MAGO-LECCIA (1994), MACHADO-ALLISON & FINK (1995).
Para cada época y laguna se evaluó la captura porunidad de esfuerzo (CPUE), expresada como númerode individuos y peso fresco en gramos de pecescapturados durante una hora por 45 m2 de red, el grupode las tres primeras redes fue extrapolado a esta su-perficie.
Los grupos tróficos se identificaron basados en el tra-bajo de AYALA et al. (2000) y apoyándose en otros au-tores tales como JUNK et al. (1983), BAYLEY (1988),
WINEMILLER (1989), GALETTI et al. (1990), ARAUJO-LIMA et al., (1995), MACHADO-ALLISON & FINK (1995),FUGI et al. (1996), AGOSTINHO et al. (2000).
Se establecieron 4 categorías: detritívoros, omnívoros,carnívoros y piscívoros.
Las estrategias de vida de cada especie fueron asigna-das según estudios realizados por WINEMILLER &TAPHORN (1990), WINEMILLER (1992), WINEMILLER& ROSE (1992) respaldados por otros investiga-dores tales como VAZZOLER & MENEZES (1992), A-GOSTIHNO et al. (2000) y observaciones personalesde campo.
La mayoría de las estrategias de vida en las que seclasificaron las especies capturadas, fueron asignadasde la misma manera que en los trabajos mencionadosarriba; otras fueron categorizadas según algunos ras-gos distintivos de su reproducción, y por último para ungrupo muy reducido de especies se utilizó el tamañode los individuos de acuerdo a los criterios de WINE-MILLER (1989).
Se reconocieron tres estrategias de vida:
• Equilibrio (K): en ella se incluyen especies queposeen cuidado paternal, época de reproduc-ción larga, reproducción repetitiva varias vecesen la época (lo que da lugar a períodos de re-producción acíclica o no controlada por las llu-vias), distribución de edades uniforme a travésdel año, alta sobrevivencia de juveniles comoresultado del cuidado paternal, una generaciónrelativamente larga, cuerpo grande, tamaño depuesta pequeño, diámetro del oocito grande.
• Oportunismo (r1): Incluye especies de tallaspequeñas, no tienen (o tienen poca) protecciónpaternal de la cría, época de reproducción pro-longada, repetidos episodios de reproducción,una distribución uniforme de edades a travésde la época lluviosa (y en sequía también paraalgunas especies), generaciones cortas,pequeñas nidadas, oocitos pequeños y fluctua-ciones poblacionales intermedias. Estos pecesposeen una alta capacidad de colonización.
• Periódico (r2): Tienen muy poca o ningunaprotección paternal de la cría, épocas de repro-ducción cortas, un solo episodio de reproduc-ción cada año o reproducción estacional o cíclica,predominancia de juveniles en la época lluviosa,generaciones largas, cuerpo grande o mediano,tamaños de puestas grandes, pequeños oocitosy fecundidades altas.

37
CARVAJAL, F. y M. MALDONADO: Conexión río-laguna e Ictiocenosis en El Río Ichilo
Cuadro 1. Lista sistemática de los peces capturados en las tres lagunas y en las dos épocas estudiadas. AB – aguas bajas, AA – aguas altas
RESULTADOS
Composición taxonómica de la ictiocenosis
Se capturó un total de 56 especies pertenecientes a 5órdenes y 16 familias, 46 especies durante la épocade aguas bajas y 37 durante la época de aguas altas.Los Characiformes estuvieron representados por 36especies, los Siluriformes por 15 , los Perciformes por3, los Gymnotiformes y los Clupeiformes por 1 especiecada uno. El cuadro 1 indica la presencia de las espe-cies, familias y órdenes capturados en las tres lagunas.
En la laguna Río Viejo se capturó un total de 24 especiesde peces en las dos épocas hidrológicas, en la lagunaBufeos 41 y en la laguna Redonda 25. Las especiescomunes a todas las lagunas y épocas fueron Mylossomaduriventre (Characidae), Serrasalmus rhombeus(Characidae), Triportheus angulatus (Characidae) yPotamorhina altamazonica (Curimatidae) (Cuadro 1).
En términos de número de especies la familia Characi-dae y Curimatidae fueron las más diversas. Un total de34 especies fueron capturadas para ambas familias y26 y 8 especies para cada una de ellas.
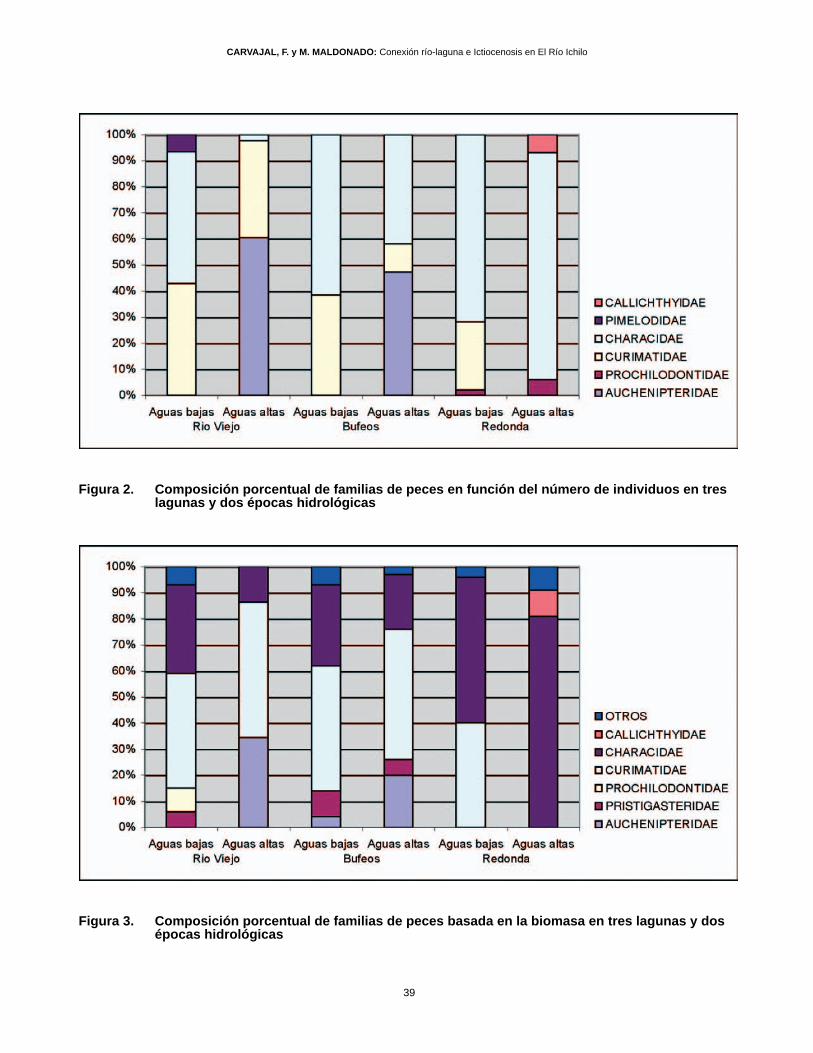
Las diferencias entre épocas y lagunas en la composi-ción porcentual de las familias, en base al número deindividuos, se pueden apreciar en la figura 2. En aguasbajas los Characidae son dominantes en las lagunasalcanzando desde el 46 hasta el 72% en la lagunaRedonda, los Curimatidae representaron entre un 30y 40 %. En aguas altas se observan cambios drásticosen las lagunas Río Viejo y Bufeos, pues la proporciónde Characidae disminuye a favor de los Auchenipteridaeque representan hasta 52 y 44% respectivamente, encambio en la laguna Redonda, la dominancia de

REVISTA BOLIVIANA DE ECOLOGÍA Y CONSERVACIÓN AMBIENTAL
38
Cuadro 1. (Cont.)
Characidae se acentúa hasta un 87%, siendo los Cu-rimatidae reemplazados por Callichthydae (7%) y Pro-chilodontidae (6%).
Considerando el número de individuos por especies, seencontró que las especies dominantes en las capturasfueron los carácidos Triportheus angulatus y Cteno-brycon spirulus, el auqueniptérido Auchenipterus am-byacus, y los curimátidos Potamorinha latior , Psectro-gaster rutiloides y P. curviventris.
Cuando se analiza la composición porcentual en térmi-nos de biomasa por órdenes, se observa que estuvodominada por los Characiformes (80%), seguida porlos Siluriformes (14%), Clupeiformes (5%) y Percifor-mes (1%). Con respecto a las familias capturadas, lacomposición porcentual en biomasa muestra que lafamilia dominante fue la de los Curimatidae con el 43%seguida por los Characidae con el 34%.
La biomasa de peces en la laguna Río Viejo (Fig. 3),fue dominada porcentualmente en ambas épocas porla familia Curimatidae (44 - 50%, aguas bajas y altasrespectivamente), seguida por los Characidae (34%)
en aguas bajas y Auchenipteridae (33%) en aguas al-tas. En esta época tanto los Prochilodontidae como losPristigasteridae están ausentes.
La laguna Bufeos mostró un comportamiento similar,siendo la familia más importante Curimatidae (48 - 50%,aguas bajas y altas respectivamente), seguida porCharacidae (31- 21%) y Pristigasteridae (10%) en aguasbajas y Auchenipteridae (20%) en aguas altas (Fig. 3).
Al igual que la composición en número de individuos,la composición en biomasa se mostró diferente en lalaguna Redonda en comparación a las otras lagunas(Fig. 3). Los Characidae son dominantes en ambasépocas (56 % en aguas bajas y 81 % en aguas altas),pero mucho más en aguas altas; los Curimatidae soloaparecen importantes en aguas bajas con 40%, casidesapareciendo en aguas altas, época en la cualaparece un notorio incremento de Callichthydae (10%).
Considerando la composición porcentual de especiesen la biomasa, las especies más abundantes a lo lar-go de todo el estudio fueron P. latior, T. angulatus, A.ambyacus, P. curviventris y P. rutiloides.
39
CARVAJAL, F. y M. MALDONADO: Conexión río-laguna e Ictiocenosis en El Río Ichilo
Figura 2. Composición porcentual de familias de peces en función del número de individuos en tres lagunas y dos épocas hidrológicas
Figura 3. Composición porcentual de familias de peces basada en la biomasa en tres lagunas y dos épocas hidrológicas

REVISTA BOLIVIANA DE ECOLOGÍA Y CONSERVACIÓN AMBIENTAL
40
Composición trófica de la ictiocenosis
La composición de grupos tróficos presentes en las treslagunas estudiadas muestra que hay 3 grupos conproporciones más o menos equilibradas: los detritívo-ros con el 33%, los omnívoros (31%), carnívoros (27%),siendo los piscívoros los menos representados (9%).En proporciones absolutas dominan los carnívoros(27 + 9). El grupo más importante numéricamente enla época de aguas bajas fue el de los carnívoros (42%)y de los detritívoros (30%). Mientras que en la épocade aguas altas los grupos más sobresalientes fueronlos omnívoros (44%) y los piscívoros (36%).
Los grupos tróficos más importantes en las lagunas RíoViejo y Bufeos lo conformaron los omnívoros (44 y 29%respectivamente) y los detritívoros (33 y 39% res-pectivamente). En cambio en la laguna Redonda loscarnívoros (35%) y los omnívoros (31%) fueron losgrupos más abundantes porcentualmente.
Las variaciones entre épocas para cada laguna seobservan en la figura 4. En la laguna Río Viejo, lasproporciones de grupos se mantienen muy similaresentre ambas épocas: omnívoros (45% y 43% aguasbajas y altas respectivamente) junto con los detritívo-ros (33% y 33%) representaron los grupos tróficos másnumerosos. En la laguna Bufeos los grupos más im-portantes durante la época de aguas bajas lo con-formaron los carnívoros (44%) y los detritívoros (32%),en cambio durante la época de aguas altas lo fuerondetritívoros (43%) y los omnívoros (36%). La lagunaRedonda mostró como grupos importantes en aguasbajas a los carnívoros (45%) y los detritívoros (29%),en tanto que en aguas altas el grupo dominante fue elde los omnívoros (70%) (Fig. 4).
Las tendencias de la CPUE para los grupos tróficos enlas tres lagunas y en las dos épocas se muestran en lafigura 5. Entre las lagunas, los valores más bajos detodo el estudio se dieron en la laguna Río Viejo tomandoen cuenta el número de individuos, y los más altos enla laguna Bufeos. Comparando entre las épocas, en lalaguna Río Viejo, no se muestran diferencias importan-tes entre las mismas, en la laguna Bufeos los valoresde CPUE fueron mayores en aguas altas, al contrariode la laguna Redonda que mostró mayores valoresdurante las aguas bajas.
Por su lado la laguna Bufeos presentó el valor de capturamás alto de los detritívoros (57.6 ind/malla/h), omnívoros(43.6 ind/malla/h) y piscívoros (16.4 ind/malla/h) a lolargo de todo el estudio. De manera diferente, la laguna
Redonda presentó el valor más alto de captura de to-das las lagunas para los carnívoros (43.5 ind/malla/h),seguidos en orden de importancia para esta mismalaguna, por los omnívoros (36.9 ind/malla/h) y losdetritívoros (30.2 ind/malla/h) (Fig. 5).
Por otro lado, la laguna Bufeos presenta mayor valorde captura de detritívoros (19.2 ind/malla/h), omnívo-ros (32.0 ind/malla/h) y piscívoros (14.2 ind/malla/h) enla época de aguas altas. Existe un incremento de losdetritívoros (de 19.2 a 38.4 ind/malla/h), omnívoros(de 11.6 a 32.0 ind/malla/h) y piscívoros (de 3.2 a13.2 ind/malla/h) en la época de aguas altas, respec-to a la época de aguas bajas, y una disminución de loscarnívoros de la época de aguas bajas a la épocade aguas altas (de 25.5 a 4.0 ind/malla/h) (Fig. 5).
Por último, la laguna Redonda presentó el valor máxi-mo de las tres lagunas en la captura de los carnívorosdurante las aguas bajas. El grupo de los omnívoros semantiene prácticamente constante en las dos épocasde muestreo, los piscívoros presentan una leve dis-minución, mientras que los detritívoros (de 26.3 a 3.8ind/malla/h) como los carnívoros (41.5 a 1.9 ind/ma-lla/h), presentaron cambios drásticos entre épocashidrológicas.
En general se puede observar que para las dos épocashidrológicas existe una tendencia a un incremento delos carnívoros a medida que la laguna esta más alejadadel río (laguna Redonda), en cambio los detritívoros,omnívoros y piscívoros tienden a ser más abundan-tes en lagunas conectadas al río por el canal como esel caso de la laguna Bufeos.
Realizando un análisis por la biomasa capturada, te-nemos de un modo general que los detritívoros repre-sentaron el 46%, los omnívoros el 26%, los carnívorosel 11% y los piscívoros el 17% de todas las capturasrealizadas.
La composición porcentual en biomasa para las treslagunas y épocas estudiadas se muestra en la figura 6,no se observan cambios notables, a excepción delperíodo de aguas altas en la laguna Redonda en quelos piscívoros se vuelven más abundantes.
En términos de CPUE, los mayores valores se pre-sentaron en la laguna Bufeos en ambas épocas(2 342,6 y 4 114,0 g/malla/h para ambas épocas),valores menores se capturaron en laguna Río Viejo ylaguna Redonda que fluctuaron entre 923.1 y 1 618.8g/malla/h, dándose los mayores en época de aguasbajas.

CARVAJAL, F. y M. MALDONADO: Conexión río-laguna e Ictiocenosis en El Río Ichilo
41
Figura 4. Composición relativa de grupos funcionales de peces en base al número de individuos en tres lagunas y dos períodos hidrológicos
Figura 5. Abundancia relativa expresada como CPUE de los grupos funcionales de peces en base al número de individuos

REVISTA BOLIVIANA DE ECOLOGÍA Y CONSERVACIÓN AMBIENTAL
42
Figura 7. Abundancia relativa como CPUE de los grupos funcionales de peces expresada en biomasa
Figura 6. Composición porcentual de grupos funcionales de peces basada en la biomasa en tres lagunasy dos épocas hidrológicas
CARVAJAL, F. y M. MALDONADO: Conexión río-laguna e Ictiocenosis en El Río Ichilo
43
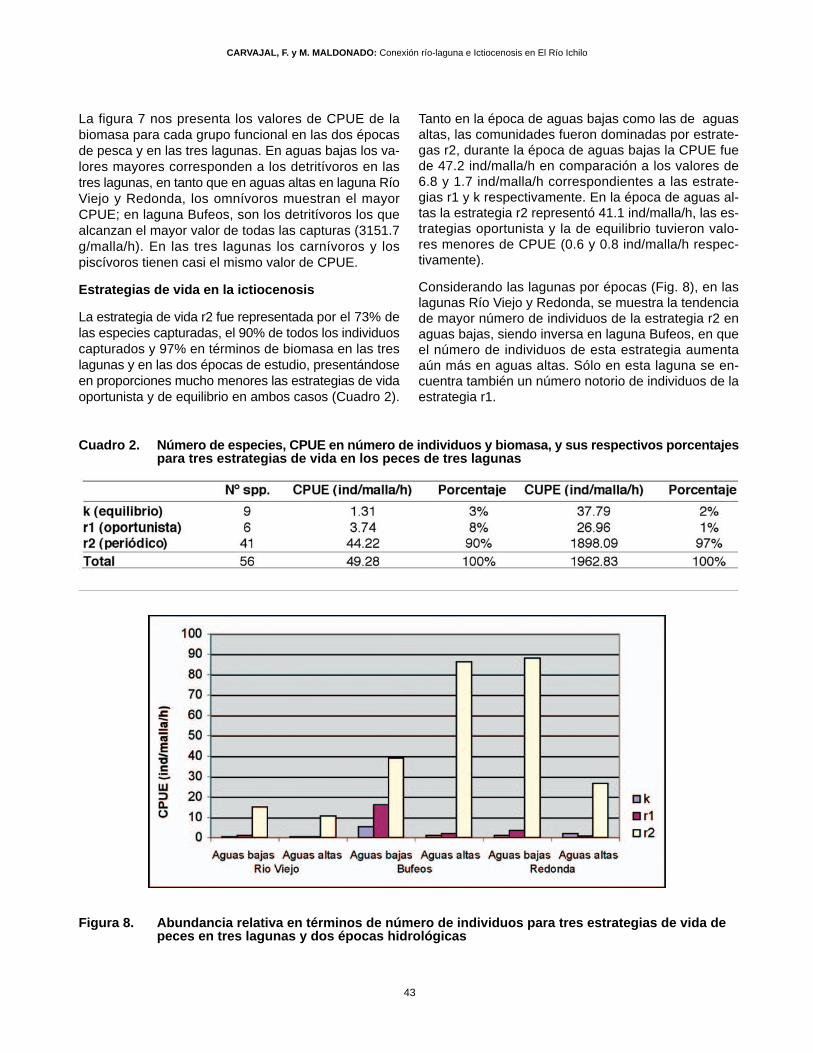
Tanto en la época de aguas bajas como las de aguasaltas, las comunidades fueron dominadas por estrate-gas r2, durante la época de aguas bajas la CPUE fuede 47.2 ind/malla/h en comparación a los valores de6.8 y 1.7 ind/malla/h correspondientes a las estrate-gias r1 y k respectivamente. En la época de aguas al-tas la estrategia r2 representó 41.1 ind/malla/h, las es-trategias oportunista y la de equilibrio tuvieron valo-res menores de CPUE (0.6 y 0.8 ind/malla/h respec-tivamente).
Considerando las lagunas por épocas (Fig. 8), en laslagunas Río Viejo y Redonda, se muestra la tendenciade mayor número de individuos de la estrategia r2 enaguas bajas, siendo inversa en laguna Bufeos, en queel número de individuos de esta estrategia aumentaaún más en aguas altas. Sólo en esta laguna se en-cuentra también un número notorio de individuos de laestrategia r1.
Figura 8. Abundancia relativa en términos de número de individuos para tres estrategias de vida de peces en tres lagunas y dos épocas hidrológicas
Cuadro 2. Número de especies, CPUE en número de individuos y biomasa, y sus respectivos porcentajespara tres estrategias de vida en los peces de tres lagunas
La figura 7 nos presenta los valores de CPUE de labiomasa para cada grupo funcional en las dos épocasde pesca y en las tres lagunas. En aguas bajas los va-lores mayores corresponden a los detritívoros en lastres lagunas, en tanto que en aguas altas en laguna RíoViejo y Redonda, los omnívoros muestran el mayorCPUE; en laguna Bufeos, son los detritívoros los quealcanzan el mayor valor de todas las capturas (3151.7g/malla/h). En las tres lagunas los carnívoros y lospiscívoros tienen casi el mismo valor de CPUE.
Estrategias de vida en la ictiocenosis
La estrategia de vida r2 fue representada por el 73% delas especies capturadas, el 90% de todos los individuoscapturados y 97% en términos de biomasa en las treslagunas y en las dos épocas de estudio, presentándoseen proporciones mucho menores las estrategias de vidaoportunista y de equilibrio en ambos casos (Cuadro 2).

REVISTA BOLIVIANA DE ECOLOGÍA Y CONSERVACIÓN AMBIENTAL
44
MALDONADO et al. (1996) indican que el río Ichilo porsu cercanía a las cabeceras fluviales de donde nace,posee un pulso de inundación irregular y corto, por loque podríamos suponer que las especies que llegarían a dominar el sistema, serían aquellas con estrategiade vida oportunista (r1), como lo proponen REJAS &MALDONADO (2000). En este sentido WINEMILLER(1989) indica que si un ambiente se mantiene largosperíodos de tiempo sin perturbaciones mayores, losestrategas periódicos por virtud de su gran fecundidad,eventualmente tienen episodios de fuerte almace-namiento. Si viven mucho, el efecto de acumulación lespermite dominar la comunidad en la ausencia deperturbaciones mayores (p.e. desecación completa) oreducción y limitación de recursos que disminuyenseveramente subsecuentemente al almacenaje. Estapodría ser una explicación a la gran dominancia de losestrategas r2 en las lagunas del río Ichilo, puesto queeste sistema parece no presentar cambios drásticoscuando hay una transición de la época de aguas bajasa la época de aguas altas.
A manera de conclusión podemos comentar que losresultados presentados en este trabajo, evidencian laposible influencia de la relación río-laguna sobre laictiocenosis lacustre, tal como ha sido demostradoen otros sistemas río-planicie de inundación. Así, lavariedad específica y la abundancia de peces semostraron mayores en el ambiente con conexión direc-ta por un canal, mediante el cual se produce periódicay regularmente un intercambio entre el río y la laguna,frente al conectado por cárcava y al desconectado.
Esta relación río-laguna también parece producir unajuste en la composición de los grupos tróficos y taxo-nómicos, ya que fueron más similares entre sí las lagu-nas con conexión directa, siendo la laguna desconecta-da muy disímil a las anteriores.
De esta manera, en las lagunas que se conectan demanera directa al río, lo que significa que su condiciónhidrométrica y físico-química es afectada de formainmediata, habita una comunidad de peces similar a lade los ríos dominada por las familias Characidae yCurimatidae y los grupos tróficos de los omnívoros ydetritívoros. Las variaciones entre las épocas hidrológi-cas también son similares, siendo la familia Characidaedominante durante el periodo de aguas bajas y la fami-lia Auchenipteridae durante el periodo de aguas altas.
En la laguna que no posee conexión directa con el ríoy que recibe agua mediante los desbordes de lainundación, la comunidad de peces es altamente
DISCUSIÓN
Los resultados obtenidos en este trabajo constituyen laprimera descripción comparativa de la ictiocenosis enlagunas de la várzea del río Ichilo en función al gradode intercambio río-laguna en este tipo de sistemas. Conestos resultados se espera aportar a la generación dehipótesis que permitan explicar los complejos me-canismos ecológicos que controlan su diversidad yfuncionamiento.
Los resultados de este trabajo deben considerarse conlimitaciones impuestas por la técnica de pesca, (selec-tiva), lo que significa que solo se capturaron una fracciónde las especies, y la carencia de repeticiones en el tipode lagunas (una por tipo de conexión) debido a proble-mas de accesibilidad.
Sin embargo, algunas tendencias pueden tomarse enconsideración, como en el caso de la composición delas capturas en términos de los grupos taxonómicos,particularmente a nivel de órdenes y familias quemostraron un patrón de abundancia similar al observa-do para otros ambientes amazónicos y para la mis-ma llanura de inundación del río Ichilo, por ejemploCASTELLÓ (1987) en lagunas del Beni, GALETTI et al.(1990) en el río Alto Paraná, WINEMILLER (1990) enVenezuela, ARAUJO-LIMA et al. (1995) en Brasil,MALDONADO et al. (1997) en el río Ichilo, POUILLYet al. (1999) en el río Mamoré, REJAS & MALDONA-DO (2000) en otras lagunas de la várzea del río Ichilo.
En el caso de la composición trófica, de igual manerauna composición con altos porcentajes de detritívo-ros y piscívoros - carnívoros, es un hecho común en laAmazonía (BAYLEY, 1973; ARAUJO-LIMA et al., 1986;MACHADO-ALLISON, 1987), debiendo hacer notar quela mayoría de especies de estos grupos pertenecen alos Characiformes y Siluriformes. Muchos de los abun-dantes peces detritívoros en los ríos de Sud Américaestán contenidos en dos familias relacionadas cerca-namente: Prochilodontidae y Curimatidae (BOWEN,1983; FORSBERG et al., 1993).
De los resultados encontrados sobre las estrategias devida de los peces, podemos decir sin tomar en cuentatodas las posibles interacciones reproductivas y eco-lógicas con el medio ambiente en las lagunas de várzea,que los peces en su mayoría poseen la estrategiade vida r2. Según WINEMILLER & ROSE (1992) estaestrategia es favorecida en ambientes estacionales ode variación espacial a gran escala por la maduracióntardía, alta fecundidad y turnos reproductivos pulsados.

45
CARVAJAL, F. y M. MALDONADO: Conexión río-laguna e Ictiocenosis en El Río Ichilo
dominada por la familia Characidae y predominan lasespecies con hábitos alimenticios carnívoros y omní-voros.
Por útlimo, respecto a las estrategias de vida, la estrate-gia periódica o r2 se manifiesta en la mayoría de laiciofauna lacustre en la várzea del río Ichilo proba-blemente debido a que les permite ajustarse a esteambiente periódicamente fluctuante.
AGRADECIMIENTOS
Este trabajo fue posible gracias al financiamiento delPrograma de Cooperación Interuniversitaria entre laUMSS (Bolivia) y el VLIR (Bélgica). Agradecemosa Danny Rejas por su apoyo en el trabajo de campo .
BIBLIOGRAFÍA
AGOSTINHO, A. A., S. M. THOMAZ, C. V. MINTE-VERA& K. O. WINEMILLER. 2000. Biodiversity in theHigh Paraná river floodplain In: Biodiversity inWetlands: assessment, function and conser-vation. Vol. 1. pp. 89-118. B. GOPAL, W. S. JUNK& J. A. DAVIS (Eds.). Bakhuys Publishers, Leiden,The Netherlands.
ARAUJO-LIMA, C. A., A. A. AGOSTINHO & N. N. FABRÉ.1995. Trophic aspects of fish communities inBrazilian rivers and reservoir. pp. 105-136. In:TUNDUSI, J. G., C.E. BICUDO & T. MATSU-MURA-TUNDUSI (Eds). Limnology in Brazil.Brazilian Academy of Science/Brazilian Limno-logical Society, Rio de Janeiro.
ARAUJO-LIMA, C. A., B. R. FORESBERG, R. VICTORIA& L. MARTINELL. 1986. Energy sources for de-tritivorous fishes in the Amazon. Science 234:1256-1258.
AYALA, G., K. ZAMBRANA & M. MALDONADO. 2000.Estructura trófica de la ictiocenosis en lagunasde la llanura inundable de los ríos Ichilo y Chapare(Bolivia). Rev. Bol. Ecol. 7:25-35.
BAYLEY, P. B. 1973. Studies on the migratory characin,Prochilodus platensis Homberg 1889 (Pisces,Characoidei) in the river Pilcomayo, South Ame-rica. J. Fish. Biol. 5:25-40.
BAYLEY, P. B. 1988. Factors affecting growth rates ofyoung tropical floodplain fishes: seasonality anddensity dependence. Environmental Biology ofFishes 2(2):127-142
BOWEN, S. H. 1983. Detritivory in neotropical fishcommunities. Environmental Biology of Fishes9(2):137-144.
CASTELLÓ, V. 1987. Composición y evolución espacio-temporal de la comunidad de peces de los llanosde Moxos, Beni-Bolivia. Tesis Doctoral. Universi-dad de Córdoba. España. 181 p.
CUMAT. 1985. Estudio de la Capacidad de Uso Mayorde Tierra Río Ichilo - Río Isarsama, Departamentode Cochabamba. USAID. La Paz, Bolivia. 85 p.
DRAGO, E. C. 1981. Grados de conexión y faseshidrológicas en ambientes de la llanura aluvialdel río Paraná (Argentina). Ecología Argentina6:27-33.

REVISTA BOLIVIANA DE ECOLOGÍA Y CONSERVACIÓN AMBIENTAL
46
FINK, W. L. & A. MACHADO-ALLISON. 2001.Serrasalmus hastatus, a new species of pirañafrom Brazil, with comments on Serrasalmus altu-vei and Serrasalmus compressus (Teleostei:Characiformes). Occasional papers of the Muse-um of the University of Michigan 70(3):11-18.
FORESBERG, B. R., C. A. ARAUJO-LIMA, L. A.MARTIRELLI, R. L. VICTORIA & J. A. BONSÁIS.1993. Autotrophic carbon sources for fishes ofthe central Amazon. Ecology 74(3):643-652.
FUGI, R., N.S. HAHN & A.A. AGOSTINHO. 1996.Feeding styles of five species of bottom feedingfishes of the high Paraná river. EnvironmentalBiology of Fishes 46(3):297-307.
GALETTI, Jr. P. M., K. E. ESTEVES, N. R. LIMA, C. A.MESTRINER, M. M. CAVALLINI, A. C. CESAY &C. S. MIYAZAWA. 1990. Aspectos comparativosda ictiofauna de duas lagoas marginais do rioMogi-Guaçu (Alto Paraná-Estaçao Ecologíado Jatai, sp). Acta Limnol. Brasil. 3:865-885.
GERY, J. 1977. Characoids of the World. T. F. H.Publications. U.S.A. pp 95-550.´
GOSLINE, W. A. 1940. A revision of the Neotropicalcatfishes of the Family Callichthydae. StanfordIchthyological Bulletin. Natural History Museumof Stanford University 2(1): 1-29.
JUNK, W. J. 1980. Áreas inundáveis. Um desafio paraa Limnologia. Acta Amazônica 10(4):775-795.
JUNK, W. J., G. M. SOARES & F. M. CARVALHO. 1983.Distribution of fish species in a lake of the Amazonriver floodplain near Manaus (Lago Camaleao),with special reference to extreme oxygen con-ditions. Amazoniana 7(4):397-431.
LAUZANNE, L. y G. LOUBENS. 1985. Peces del ríoMamoré. ORSTOM –CORDEBENI – UTB. Paris.115 p.
MACHADO-ALLISON, A. 1987. Los peces de los llanosde Venezuela: un ensayo sobre su historia natural.Universidad Central de Venezuela. CDC. 141 pp.
MACHADO-ALLISON, A. & W. FINK. 1995. Sinopsis delas especies de la sub-familia Serrasalminaepresentes en la cuenca del Orinoco. UniversidadCentral de Venezuela. Museo de Biología. Pp. 6-7.
MAGO-LECCIA, F. 1994. Electric fishes of thecontinental waters of America. Biblioteca de laAcademia de Ciencias Físicas Matemáticas yNaturales XXIX:17-19.
MALDONADO, M., E. GOITIA, F. ACOSTA, M. CADIMAy D. CASTELLÓN. 1996. Caracterizaciónlimnológica de lagunas en la llanura aluvial delrío Ichilo, Cochabamba (Bolivia). Rev. Bol. Ecol.1 (1): 29-37.
MALDONADO, M., E. GOITIA, F. ACOSTA, D.CASTELLÓN y M. CADIMA. 1997. Bases parael Manejo de los Recursos Hidrobiológicos en elDepartamento de Cochabamba. Informe FinalTécnico. Universidad Mayor de San Simón.Facultad de Ciencias y Tecnología, Unidad deLimnología y Recursos Acuáticos. 138 p.
MEES, G. F. 1974. The Auchenipteridae and Pime-lodidae of Suriname (Pisces,Nematognathi).Zoologische Verhandelingen. Uitgegeven doorhet Rijksmuseum Van Natururlijke historie teLeiden 32: 15-16, 116-117.
MENEZES, N. A. 1969. Systematic and evolution ofthe tribe Acestrorhynchini (Pisces, Characi-dae). Archivos de Zoología 18(1-2): 1-150.
NAVARRO, G. y M. MALDONADO. 2002. GeografíaEcológica de Bolivia. Ed. Centro de EcologíaSimón I. Patiño-Departamento de Difusión.Cochabamba, Bolivia. 719 p.
POUILLY, M., C. IBAÑEZ, M. GUTIERREZ & T. YUNOKI. 1999. Funcionamiento ecológico de las lagunasde la zona de inundación del río Mamoré (Beni-Bolivia). Rev. Bol. Ecol. 6:41-54.
REIS, R. 1989. Systematic Revision of the NeotropicalCharacid Sub-Family Stethaprioninae (Pisces,Characiformes). Comun. Mus. Ciênc. PUCRS,Sér. Zool. 2(6):3-86.
REJAS, D. y M. MALDONADO. 2000. Variacionesespacio-temporales de la composición de lacomunidad de peces de una laguna de la várzeadel río Ichilo (Cochabamba, Bolivia). Rev. Bol.Ecol. 7: 37-46.

47
CARVAJAL, F. y M. MALDONADO: Conexión río-laguna e Ictiocenosis en El Río Ichilo
WINEMILLER, K. O. & D. B., JEPSEN. 1998. Effects ofseasonality and fish movement on tropical riverfood webs. Journal of Fish Biology 53 (SuplementA):267-296.
WINEMILLER, K. O. & D. C. TAPHORN. 1990. Laevolución de las estrategias de vida en los pecesde los llanos occidentales de Venezuela. Biollania6:77-100.
WINEMILLER, K. O. & K. A. ROSE. 1992. Patterns oflife history diversification on North American fishes.Implications for population regulation. CanadianJournal of Fisheries and Aquatic Sciences 49:2196-2218.
WINEMILLER, K.O. 1989. Patterns of variation in lifehistory among South America fishes in seasonalenvironments. Oecología 81:225-241.
SAINT-PAUL, V. & U. PETER. 1979. A situacao da pescana Amazonia Central. Supl. Acta Amazonica 9(4):109-114.
SAINT-PAUL, V., M. ZUANON, A. VILLACORTACORREA, M. GARCIA & N. N., FABRE. 2000.Fish communities in Central Amazonian whiteand black water floodplains. Enviromental Biolo-gy of Fishes 57:235-250.
VARI, R. P. 1984. Systematics of the NeotropicalCharaciform Genus Pothamorhina Pisces: Cha-raciformes). Smithsonian Contributions to Zoology400:36-37.
VARI, R. P. 1989. Systematics of the NeotropicalCharaciform Genus Psectrogaster Eigenmannand Eigenmann (Pisces: Characiformes). Smith-sonian Contributions to Zoology 481:43.
VARI, R. P. 1991. Systematics of the NeotropicalCharaciform Genus Steindachnerina Fowler(Pisces: Ostariophysi). Smithsonian Contributions to Zoology. 507-118.
VARI, R. P. 1992. Systematics of the NeotropicalCharaciform Genus Curimatella Eigenmann andEigenmann (Pisces: Ostariophysi), with sum-mary coments on the Curimatidae. SmithsonianContributions to Zoology 533:48.
VAZZOLER, A. E. DE M. & N. A. MENEZES. 1992.Síntese de conhecimentos sobre o comportamentoreproductivo dos Characiformes da Ámerica doSul (Teleostei, Ostariophysi). Rev. Brasil. Biol.52(4):627-640.
WAIS, I. R. & H. P. CASTELLO. 1984. Auchenipterusnuchalis, única especie del género presente enel río Paraná, Argentina (Pisces, Auchenipteridae).Revista del Museo Argentino de Ciencias Naturales“Bernardino Rivadavia”. Zoología XIII(5):58-70.
WELCOMME, R. 1979. Fisheries ecology of flooplainrivers. Longman Ed. London. 317 p.
WINEMILLER, K. O. 1990 Spatial and temporal variationin tropical fish trophic networks. Ecologicalmonographs 60:331-367.
WINEMILLER, K. O. 1992. Life history strategies andthe effectiveness of sexual selection. Oikos62:318-327.

REVISTA BOLIVIANA DE ECOLOGÍA Y CONSERVACIÓN AMBIENTAL
48