
Inflammatory response to Dentitruncus truttae (Acanthocephala) in the intestine of brown trout
8
Inflammatory response to Dentitruncus truttae (Acanthocephala) in the intestine of brown trout Bahram S. Dezfuli a, *, Giancarlo Giovinazzo b , Alice Lui a , Luisa Giari a a Department of Biology and Evolution, University of Ferrara, St. Borsari 46, 44100 Ferrara, Italy b Department of Cellular and Environmental Biology, University of Perugia, St. Elce di Sotto, 06123 Perugia, Italy Received 6 September 2007; revised 5 November 2007; accepted 18 November 2007 Available online 5 December 2007 KEYWORDS Salmo trutta; Acanthocephalan; Inflammation; Mast cells, Rodlet cells Abstract Brown trout, Salmo trutta L., were infected with the acanthocephalan Dentitrun- cus truttae with the most affected areas being the anterior (near the pyloric caeca) and mid- dle intestine. The parasite attached with a proboscis which usually penetrated the mucosa, lamina propria, stratum compactum, stratum granulosum and, sometimes, the muscularis layer. Around the parasite’s body was an area of inflammatory tissue. At the point of attach- ment the lamina propria was thickened and the stratum compactum, stratum granulosum and muscularis layer were disrupted by proboscis penetration. Rodlet cells were more numerous in infected fish (P < 0.01), and were found in the epithelial layer away from the worm. Infected intestines had larger numbers of mast cells (P < 0.01), often in close proximity to, and inside, the blood capillaries and associated with fibroblasts of the muscularis layer and the stratum granulosum. Their migration toward the site of infection was suggested. Intense degranulation of mast cells was encountered in all intestinal layers especially near the parasite’s body. Immunohistochemical tests were conducted on sections of intestinal tissue of uninfected and infected fish revealing the presence of met-enkephalin and serotonin (5-HT) in immuno- related cells of the intestine wall. Infected trout had larger numbers of elements positive to met-enkephalin and serotonin antisera. These data provided evidence for the role of the immune system of brown trout in the modulation of the inflammatory response to D. truttae. Results are discussed with respect to host immune response to an intestinal helminth. ª 2007 Elsevier Ltd. All rights reserved. Introduction Intestinal helminths of mammals induce alterations in the structure of the gut tissue and these changes affect its normal function [1,2], moreover, these worms often induce intestine inflammation and elicit host immune reactions, mainly in gut-associated lymphoid tissue [3]. The inflammation * Corresponding author. Tel.: þ39 0532 455 701; fax: þ39 0532 455 715. E-mail address: [email protected] (B.S. Dezfuli). 1050-4648/$ - see front matter ª 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.fsi.2007.11.013 available at www.sciencedirect.com journal homepage: www.elsevier.com/locate/fsi Fish & Shellfish Immunology (2008) 24, 726e733
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Inflammatory response to Dentitruncus truttae (Acanthocephala) in the intestine of brown trout

Fish & Shellfish Immunology (2008) 24, 726e733
ava i lab le a t www.sc iencedi rec t .com
j ourna l homepage : www.e lsev ie r . com/ loca te / f s i
Inflammatory response to Dentitruncus truttae(Acanthocephala) in the intestine of brown trout
Bahram S. Dezfuli a,*, Giancarlo Giovinazzo b, Alice Lui a, Luisa Giari a
a Department of Biology and Evolution, University of Ferrara, St. Borsari 46, 44100 Ferrara, Italyb Department of Cellular and Environmental Biology, University of Perugia, St. Elce di Sotto, 06123 Perugia, Italy
Received 6 September 2007; revised 5 November 2007; accepted 18 November 2007Available online 5 December 2007
KEYWORDSSalmo trutta;Acanthocephalan;Inflammation;Mast cells, Rodlet cells
* Corresponding author. Tel.: þ39 0455 715.
E-mail address: [email protected] (B.S.
1050-4648/$ - see front matter ª 200doi:10.1016/j.fsi.2007.11.013
Abstract Brown trout, Salmo trutta L., were infected with the acanthocephalan Dentitrun-cus truttae with the most affected areas being the anterior (near the pyloric caeca) and mid-dle intestine. The parasite attached with a proboscis which usually penetrated the mucosa,lamina propria, stratum compactum, stratum granulosum and, sometimes, the muscularislayer. Around the parasite’s body was an area of inflammatory tissue. At the point of attach-ment the lamina propria was thickened and the stratum compactum, stratum granulosum andmuscularis layer were disrupted by proboscis penetration. Rodlet cells were more numerous ininfected fish (P < 0.01), and were found in the epithelial layer away from the worm. Infectedintestines had larger numbers of mast cells (P < 0.01), often in close proximity to, and inside,the blood capillaries and associated with fibroblasts of the muscularis layer and the stratumgranulosum. Their migration toward the site of infection was suggested. Intense degranulationof mast cells was encountered in all intestinal layers especially near the parasite’s body.Immunohistochemical tests were conducted on sections of intestinal tissue of uninfectedand infected fish revealing the presence of met-enkephalin and serotonin (5-HT) in immuno-related cells of the intestine wall. Infected trout had larger numbers of elements positive tomet-enkephalin and serotonin antisera. These data provided evidence for the role of theimmune system of brown trout in the modulation of the inflammatory response to D. truttae.Results are discussed with respect to host immune response to an intestinal helminth.ª 2007 Elsevier Ltd. All rights reserved.
532 455 701; fax: þ39 0532
Dezfuli).
7 Elsevier Ltd. All rights reserved
Introduction
Intestinal helminths of mammals induce alterations in thestructure of the gut tissue and these changes affect its normalfunction [1,2], moreover, these worms often induce intestineinflammation and elicit host immune reactions, mainly ingut-associated lymphoid tissue [3]. The inflammation
.

Brown trout’s inflammatory response to D. truttae 727
comprises a complex series of homeostatic mechanismsinvolving the immune, circulatory, and nervous systems inresponse to tissue injury or infection [4]. Several reports onfish address the essential role of immune and enteric neuro-modulators in the inflammation process caused by helminths[5e8].
In this study of brown trout infected with Dentitruncustruttae two inflammatory cell types were abundant, rodletcells (RCs) and mast cells. Rodlet cells are closely linked toother piscine inflammatory cells [9,10]. Mast cells, also
Figure 1 Parasitized intestine of Salmo trutta. (a) Sagittal secttruttae. Contact between trunk spines (arrows) and mucosa and laboscis penetrated to the stratum compactum (arrow head); bar Zseveral rodlet cells (arrow heads) are present; bar Z 30 mm. (c) Numthocephalan trunk and mucosa; bar Z 10 mm. (d) Mast cells (arrowulosum; arrow shows stratum compactum, bar Z 30 mm. (e) Largranulosum, asterisk shows stratum compactum; bar Z 30 mm. (f)inside muscularis layer; bar Z 10 mm. (g) Degranulation of several m
known as eosinophilic granule cells (EGCs), possess cyto-chemical and distribution features that have led to the sug-gestion that these cells are analogous to mammalian mastcells [10] and considered to be a component of mammal in-nate immune system [11]. In rainbow trout it was reportedthat mast cells are motile [12] and react to parasitic infec-tions [13,14] with release of their contents by degranula-tion [15,16]. Migration of these cells in fish in response toinjection of bacterial toxins and mast cell degranulationagents has been reported [17e19].
ion through brown trout intestine with attached Dentitruncusck of intestinal folds in this region (asterisk) was evident. Pro-250 mm. (b) Intact epithelia distant from parasite attachment,erous mast cells (arrow heads) in contact zone between acan-
heads) around the proboscis (P) embedded in the stratum gran-ge number of mast cells (arrow heads) within the stratumMast cells (arrow heads) surrounding a blood vessel (asterisk)ast cells (arrows) within the stratum granulosum; bar Z 10 mm.

728 B.S. Dezfuli et al.
The presence of intestinal helminths in vertebrates hasbeen associated with a higher level of serotonin (5-HT) infish [20], in mammals [2,21] and met-enkephalin in fish[6,20]. One of the major mediators secreted by mammalianmast cells is serotonin, but, little is known about the role of5-HT in the immune system of fish [6]. Met-enkephalin func-tion in the regulation of gut contractile activity is wellknown in mammals [2], whereas, in fish a great deal re-mains to be discovered. The studies referring to alterationsin neuroendocrine system of fish alimentary canal due tothe presence of parasites are rare and quite recent. Thus,herein, a comparison on the distribution of serotonin(5-HT) and met-enkephalin in S. trutta infected with Denti-truncus truttae and in uninfected conspecifics will beprovided.
There is no previous ultrastructure report on theco-occurrence of mast cells and RCs at the site of infection
Figure 2 Electron micrographs of infected intestine. (a) Two rodbasal nuclei (asterisks) and rodlets (arrow heads); bar Z 3.45 mm.fibroblast-like ensheathing cells (asterisks); note numerous electrmast cells and a blood vessel in the muscularis layer, a mast cell (outside, empty arrows show outer part of the endothelia; bar Z 3.other cell inside a vessel, fibroblast-like ensheathing cells (asteriskendothelia; bar Z 3.28 mm. (e) A mast cell degranulating, reticularound several granules (arrow heads) and collagen fibres (asterishalo (arrow) around the granule and two mitochondria (arrow hea
of an acanthocephalan in a salmonid. The present studywas designed to provide data describing: (a) histopathologyand ultrastructure of cells involved in the trout immunereaction; (b) presumed migration of the mast cells towardsthe site of infection and their degranulation; and (c) therole of met-enkephalin and serotonin (5-HT) againstinflammation caused by the acanthocephalan.
Materials and methods
A sample of 32 Salmo trutta (mean total length �SE Z 34.5 � 0.8 cm) from 3 gill net samples (July andSeptember 2006, April 2007) taken from Lake Piediluco(Province of Terni, Italy). Fish were anaesthetized usingMS222 (Sandoz) and their spinal cords severed. The diges-tive tracts were removed, opened longitudinally and
let cells among the epithelial cells, cell cortex (white arrows),(b) Intact mast cells inside the muscularis layer encircled by
on-dense granules; bar Z 3.43 mm. (c) Close contact betweenarrow head) inside the capillary and another mast cell (arrow)40 mm. (d) Muscularis layer, a mast cell (arrow) in contact withs) around the capillary, empty arrows indicate outer part of theated aspects of some granules (arrows), electron-lucent halosk); bar Z 1.78 mm. (f) Higher magnification of electron-lucentds); bar Z 0.67 mm.
Brown trout’s inflammatory response to D. truttae 729

730 B.S. Dezfuli et al.
examined for helminthes which were recorded by numberand location. Pieces of gut tissue measuring up to15 � 15 mm in area, with attached parasites, were excisedfrom 20 infected fish and fixed in chilled (4 �C) Bouin’s for8 h. The samples were then transferred to 70% ethanol, de-hydrated through an ethanol series, and prepared forparaffin embedding. Sections (5 mm) were stained with hae-matoxylin-eosin, alcian blue/PAS, or Toluidine Blue. For im-munohistochemical analysis, Salmo trutta-D. truttae tissuesections were processed using the indirect immunohisto-chemical method (peroxidase-anti-peroxidase immunocom-plex) [20]. The sections were incubated with the primaryantisera (met-enkephalin and serotonin raised in rabbit, co-des AB 1975 and AB 938, Chemicon Int., Temecula, CA, USA)for 24 h at room temperature. Controls for the specificity ofthe immunohistochemical reactions were performed bypreabsorption of each antiserum with the correspondingantigen (met-enkephalin, code H 2785, Bachem AG, Buben-dorf, Switzerland; Serotonin, code H 9523, Sigma Chemi-cals, St. Louis, MO, USA). Mammalian (swine, rat) tissuesections were used as positive controls.
For light and electron microscopy, host tissues measur-ing up to 7 � 7 mm in area were fixed for 2.5 h in chilled(4 �C) 2% glutaraldehyde solution, in 0.1 M sodium cacody-late pH 7.2. After 2.5 h the tissues were rinsed for 12 h in0.1 M sodium cacodylate buffer containing 5% sucrose.The specimens were then post-fixed in 1% osmium tetroxidein the same buffer for 2 h, dehydrated in graded ethanols,transferred to propylene oxide, and embedded in an Epoxy-Araldite� mixture. Semi-thin sections (1.5 mm) were cut ona Reichert Om U 2 ultramicrotome and stained with Tolui-dine Blue or basic fuchsin. Ultrathin sections (90 nm)were stained with a solution of 4% uranyl acetate in 50%ethanol and Reynold’s lead citrate and examined usinga Hitachi H-800 transmission microscope. For comparison,tissues from 6 uninfected trout were processed with theparasitized samples.
Light photomicrographs were taken using a Nikon micro-scope Eclipse 80i. Images of intestinal sections showing theoverall distribution of the mast cells and RCs, as well as cellimages and dimensions (30 mast cells and 30 RCs) wereobtained using computerised image analysis software (LuciaG 4.8, Laboratory Imaging, Praha, Czech Republic).
To quantify changes in mast cell and rodlet cell numbersin intestine in both healthy and parasitized fish, thick resinsections were analysed at 400� magnification. Two in-testinal sections were observed from each of six uninfectedand 20 infected trout. Cell counts were based on the same
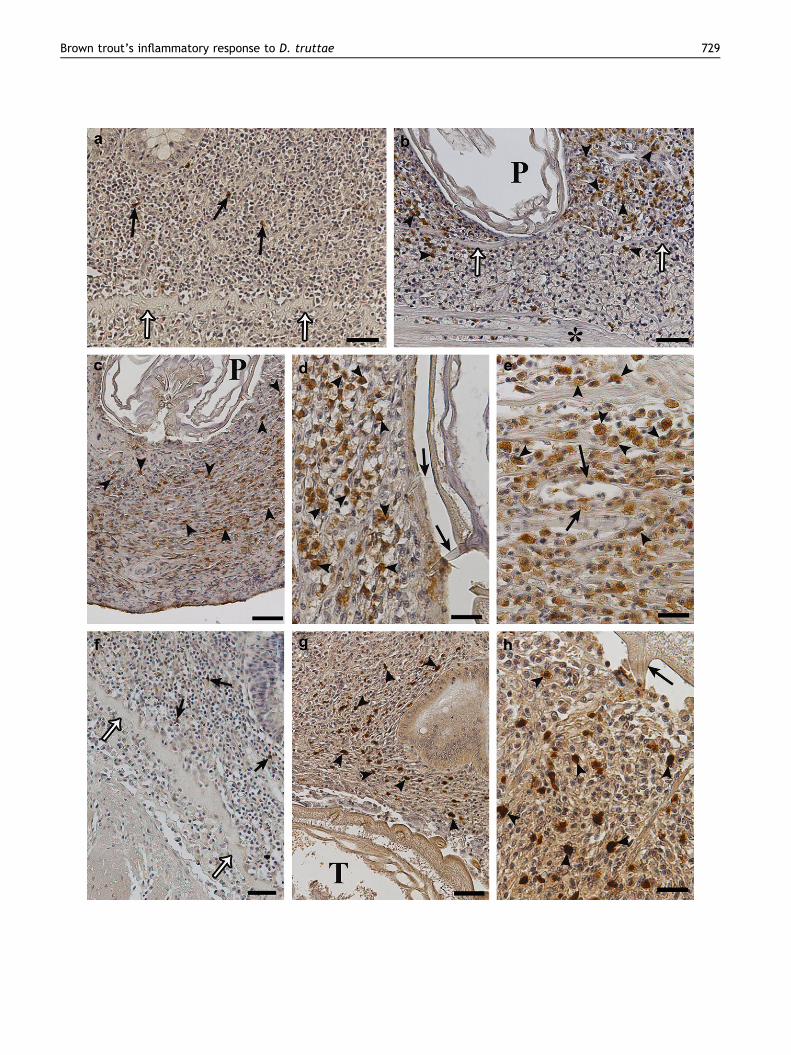
Figure 3 Immunohistochemical tests on uninfected and infected S(arrows) are positive to met-enkephalin antiserum, empty arrowscephalan proboscis (P) inside the lamina propria-stratum granulomet-enkephalin antiserum; asterisk shows muscularis layer and emboscis (P) in the muscularis layer, numerous elements (arrow heads)truttae proboscis hooks (arrows) embedded in muscularis layer tantiserum; bar Z 30 mm. (e) Mast cells (arrow heads) positive to mcontact with blood vessels; bar Z 30 mm. (f) Uninfected intestine,arrows show stratum compactum; bar Z 60 mm. (g) Acanthocepha(arrow heads) around the parasite trunk (T) are positive to anti-senear trunk spine (arrow) appear immunoreactive to serotonin anti
size tissue areas (58,000 mm2) of the same region of theintestine. Results were analysed using Student’s two tailedt-test, where significance was set at P < 0.05.
Results
In a sample of 32 Salmo trutta, 26 specimens (81.2%) wereinfected with the acanthocephalan D. truttae (Fig. 1a).A total of 1478 worms were recorded (mean in-tensity � SE Z 46.2 � 11.7). Most acanthocephalans werein the anterior (near the pyloric caeca) and middle intes-tine (Fig. 1a). The majority of the worms did not crossthe stratum granulosum (Fig. 1a), but in several cases theproboscis penetrated the muscularis layer (Fig. 3c).
D. truttae had numerous trunk spines making contactwith the mucosal folds and damaging the villi, whichshowed detachment of epithelial cells and reduced num-bers of mucosal folds near the acanthocephalan body(Fig. 1a). Folds more distant from the worm body remainedintact, and in their epithelial layer a large number of rodletcells (RCs) were observed (Figs. 1b and 2a). The RCs (majoraxis 17.46 � 2.19 mm, mean � SD) were characterized bya distinctive cell cortex, several inclusions (rodlets) anda basal nucleus (Fig. 2a). Rodlet cells counted in 52 micro-scopic fields showed that the intestine of parasitized browntrout had a significantly higher number of RCs (8.40 � 3.00,mean � SD, n Z 40) than those of uninfected fish(4.75 � 1.91, mean � SD, n Z 12) (t-test, P < 0.01).
The mucosa, lamina propria, stratum granulosum, andmuscularis layer were disrupted at the point of attachmentof the parasite proboscis. Numerous mast cells were notedin the tissues near the parasite trunk/body (Fig. 1c) andaround the proboscis (Fig. 1d). Comparison of the numberof mast cells in uninfected (25.33 � 9.60, mean � SD,n Z 12) and infected trout (81.00 � 18.36, mean � SD,n Z 40) in 52 microscopic fields showed that the intestineof parasitized fish had a significantly higher number ofmast cells (t-test, P < 0.01). Mast cells (major axis12.02 � 1.06 mm, mean � SD, n Z 30) were irregular inshape with an eccentric, polar nucleus, and cytoplasmcharacterized by numerous large, electron-dense,membrane-bounded granules (1.24 � 0.57 mm averagesize � SD) (Figs. 1c,f and 2b). The cytoplasm typically con-tained two to three mitochondria (Fig. 2f) and an inconspic-uous Golgi apparatus. Mast cells were frequentlysurrounded by collagen fibres of the stratum granulosum(Fig. 2e) or by fibroblast-like ensheathing cells (Fig. 2b,d).
. trutta. (a) Micrograph showing uninfected intestine, few cellsshow stratum compactum; bar Z 60 mm. (b) Around acantho-sum, note numerous cells (arrow heads) immunoreactive to
pty arrows indicate stratum compactum; bar Z 60 mm. (c) Pro-are positive to anti-met-enkephalin serum; bar Z 60 mm. (d) D.issue, arrow heads show immunoreactivity to met-enkephalinet-enkephalin antiserum, some mast cells (arrows) are in closefew cells (arrows) are positive to serotonin antiserum, empty
lan embedded in the stratum granulosum, numerous elementsrotonin serum; bar Z 60 mm. (h) Numerous cells (arrow heads)serum; bar Z 30 mm.

Brown trout’s inflammatory response to D. truttae 731
In both infected and uninfected brown trout the stratumgranulosum was rich in mast cells but numbers were higherin parasitized trout (Fig. 1e). In the stratum granulosum(Fig. 2e) and in the muscularis layer, numerous mast cellswere in close contact with the capillaries (Figs. 1f and2c,d), and the cells were observed in the outer layer ofthe endothelia (Figs. 1f, 2c and 3e) as well as inside theblood vessels (Fig. 2c,d).
In infected intestines, numerous mast cells were incontact with and/or inside the blood vessels, mainly withinthe muscularis layer. The cells were scattered throughoutthe muscle fibres (Fig. 2b). Degranulation of the mast cellswas common in the lamina propria and the stratumgranulosum (Fig. 2e). The cells exhibited higher rates ofdegranulation in areas near the acanthocephalan body. De-granulation was visible by light microscopy (Fig. 1e,g) andcharacterized by conspicuous swelling of granules andoften by the presence of free granules adjacent to themast cells (Fig. 1e,g). Transmission electron microscopyrevealed several stages of mast cell degranulation. The ma-trix of the granules had an extensively reticulated aspect,and prominent electron-lucent halos encircled the granules(Fig. 2e,f). In some instances, granules were fused (notshown).
Immunohistochemical tests with the antiserum againstmet-enkephalin revealed a low presence of positive cells inuninfected S. trutta (Fig. 3a). In contrast, a larger numberof immunoreactive elements was observed near the D. trut-tae proboscis in the lamina propria-stratum granulosum(Fig. 3b) and in the muscularis layer (Fig. 3c,d). Further,in all intestinal layers of the host, mast cells appeared tobe positive to met-enkephalin antiserum (Fig. 3e). In unin-fected fish, few serotonin (5-HT) immunopositive endocrinecells were observed (Fig. 3f), whereas a larger number ofcells positive to anti-serotonin serum was found in laminapropria, mainly around the trunk of D. truttae (Fig. 3g,h).Some mast cells in infected intestine and uninfected gutwere found to be positive to anti-serotonin serum.
Discussion
Most often lesions of the digestive tract of mammals are theexpression of biochemical and physiological alterationsinduced by intestinal parasites [1]. Some records on the ef-fects of parasitic helminths on the alimentary canal andassociated organs of fishes appeared in [6,8,13]. Most intes-tinal helminths do not severely damage the fish’s intestine,mainly due to their superficial relationship with the tissues.Acanthocephalan pathogenecity is due to two factors:density of parasite burden and depth of penetration intothe host tissues [7]. Dentitruncus truttae is a species thatpenetrates deeply and creates extensive damage to thedigestive tract. However, there have been few reports onthe identification of inflammatory cells involved in thisand other fish species [10,20].
In S. trutta parasitized with D. truttae, a greater pres-ence of two inflammatory cell types was observed, RCsand mast cells. Occurrence of RCs in reactive foci as a resultof infection by micro- and macro-parasites has beenreported [10,22,23]. Rodlet cells represent an inflammatorycell type closely linked to other piscine inflammatory cells,
such as mast cell/EGCs [10,24], epithelioid and mesothelialcells [9]. Rodlet cells have an important role in hostdefence [10,23,25].
The second inflammatory cell type encountered in largernumbers in all intestinal layers of the infected brown troutwere mast cells. They were numerous at the site ofinfection, although they were present in smaller numbersin uninfected fish. Several types of piscidins (antimicrobialpeptides) have been identified recently from mucosal tissueand immune cells (e.g. mast cells and RCs) of a number ofteleosts [24]. Mammal mast cells have been shown to playa critical role in defence against bacterial, viral, and fungalinfections [11,26], moreover, in mammals, mast cell prod-ucts are pivotal in mediating leukocyte recruitment intoinflammatory sites. They are capable of presenting antigensto T cells, interact directly with and affect the function ofcells of the adaptive immune system, and mediate tissuemodelling [27].
In all vertebrates mast cells may be strategicallypositioned at perivascular sites to regulate inflammatoryresponses [27]. In our study, mast cells were associatedwith capillaries, in the stratum granulosum and the muscu-laris layer of infected intestines. In the muscularis layer thevessels have larger diameters (Fig. 1f). Mast cells in boththe above sites were in contact with fibroblasts. In rainbowtrout this relationship between the two cell types also hasbeen described [28e30]. Furthermore, a mast cell-fibroblast association was observed in a lesion in which anunusual mast cell proliferation was reported in Oncorhyn-chus kisutch [31]. In mammals [32] and in fish [33] it iswell known that fibroblasts can influence mast cell motilityand proliferation and in these two classes of vertebratesseveral lines of evidence suggest that mast cells are in-volved in fibrotic processes and in tissue remodelling[34,35]. This extensive literature and the results of thisstudy might support the idea that unusual proliferation ofmast cells at the site of D. truttae infection in the stratumgranulosum and lamina propria, suggests that these cellsmigrate there from the muscularis layer. In summary, thepresence of large numbers of mast cells in infected fishand their association with fibroblasts seem to be necessaryto mediate intestine remodelling after extensive tissueinjury caused by the acanthocephalan.
In fish considerable descriptive data exist showing howmast cells degranulate in response to exposure to a varietyof pathogens and/or known degranulating agents [15e19].Mannose-binding protein from Schistosoma mansoni in hu-man has been demonstrated to induce mast cell activation,usually assessed through mast cells degranulation [11].There is no reference in the literature on mannose-bindingprotein to acanthocephalans. In the present study, abun-dant degranulation of mast cells at the site of acanthoceph-alan infection was observed. The excessive degranulationespecially near the D. truttae body might be related tothe presence of the worm. Evidence for mast cell involve-ment in response to parasitic helminths in fishes is growing[9,10,13,14 and present study].
Immunohistochemical staining demonstrated thepresence and accumulation of elements positive to met-enkephalin and serotonin antisera in close proximity to theD. truttae body. The same finding was reported in S. truttanaturally infected with the tapeworm Cyathocephalus

732 B.S. Dezfuli et al.
truncatus [6], with the acanthocephalan Pomphorhynchuslaevis [20] and in chub Leuciscus cephalus (L.) infectedwith P. laevis [8]. In the present study, mast cells appearedto react positively to met-enkephalin, the first definitivereport of this finding. Met-enkephalin function in the regu-lation of gut contractile activity is well known in mammals[36], whereas in fish a great deal remains to be discovered.Enkephalins belong to the endogenous opiate system and inmammals play an important role in the modulation of theinflammatory process [37,38]. The presence of a large num-ber of cells positive to met-enkephalin antiserum ininfected S. trutta individuals makes it reasonable to pre-sume that by increasing the amount of the met-enkepha-lin-like substance secreted, inflammation caused byD. truttae may be modulated in the brown trout.
Fish immune cells produce several defensive substancessuch as piscidins [39], pleurocidin [19] and serotonin[6,20,40]. With reference to serotonin (5-HT) in mammals,it has been established that this biogenic amine modulatesimmune function at a variety of levels [41]. D. truttae mayinduce the recruitment of cells, which secrete serotonin.However, much work still has to be done to characterizebiochemically and pharmacologically the role of serotoninin the functioning of the fish immune system [40]. Accord-ing to Castro [1] and Fairweather [2] worms provoke intenseinvolvement of vertebrate immune and neuroendocrinesystems. The present results support this concept.
Acknowledgments
We thank E. Simoni University of Ferrara for technicalassistance. Thanks are due to The Lucidus Consultancy forhelp with our English and to Alan Pike for providing usefulsuggestions. This study was supported by grants from theItalian Ministry of the University and Scientific Research andTechnology.
References
[1] Castro GA. Immunophysiology of enteric parasitism. ParasitolToday 1989;5:11e9.
[2] Fairweather I. Peptides: an emerging force in host response toparasitism. In: Beckage NE, editor. Parasites and pathogens:effects on host hormones and behaviour. New York: Chapman& Hall; 1997. p. 113e39.
[3] Rothwell TLW. Immune expulsion of parasitic nematodes fromthe alimentary tract. Int J Parasitol 1989;19:139e68.
[4] Sharkey KA. Substance P and calcitonin gene-related peptide(CGRP) in gastrointestinal inflammation. Ann NY Acad Sci1992;664:425e42.
[5] Maifrino LBM, Liberti EA, De Souza RR. Vasoactive intestinalpeptide- and substance P-immunoreactive nerve fibres inthe myenteric plexus of mouse colon during the chronic phaseof Trypanosoma cruzi infection. Ann Trop Med Parasitol 1999;93:49e56.
[6] Dezfuli BS, Arrighi S, Domeneghini C, Bosi G. Immunohisto-chemical detection of neuromodulators in the intestine ofSalmo trutta Linnaeus naturally infected with Cyathocephalustruncatus Pallas (Cestoda). J Fish Dis 2000;23:265e73.
[7] Dezfuli BS, Giari L, Simoni E, Bosi G, Manera M. Histopathol-ogy, immunohistochemistry and ultrastructure of the intestineof Leuciscus cephalus (L.) naturally infected with Pompho-rhynchus laevis (Acanthocephala). J Fish Dis 2002;25:7e14.
[8] Bosi G, Domeneghini C, Arrighi S, Giari L, Simoni E, Dezfuli BS.Response of neuroendocrine system of the intestine of Leucis-cus cephalus (L., 1758) naturally infected with Pomphorhyn-chus laevis Muller, 1776 (Acanthocephala). Histol Histopathol2005;20:509e18.
[9] Dezfuli BS, Simoni E, Rossi R, Manera M. Rodlet cells and otherinflammatory cells of Phoxinus phoxinus infected withRaphidascaris acus (Nematoda). Dis Aquat Org 2000;43:61e9.
[10] Reite OB. Mast cells/eosinophilic granule cells of salmonids:staining properties and responses to noxious agents. FishShellfish Immunol 1997;7:567e84.
[11] Marshall JS, Jawdat DM. Mast cells in innate immunity.J Allergy Clin Immunol 2004;114:21e7.
[12] Vallejo AN, Ellis AE. Ultrastructural study of the response ofeosinophilic granule cells to Aeromonas salmonicida extracel-lular products and histamine liberators in rainbow trout,Salmo gairdneri Richardson. Dev Comp Immunol 1989;13:133e48.
[13] Sharp GJE, Pike AW, Secombes CJ. The immune response ofwild rainbow trout Salmo gairdneri Richardson to naturallyacquired plerocercoid infections of Diphyllobothrium dendri-ticum (Nitzsch 1824) and D. ditremum (Creplin 1825). J FishBiol 1989;35:781e94.
[14] Reite OB. The rodlet cells of teleostean fish: their potentialrole in host defence in relation to the role of mast cells/eosi-nophilic granule cells. Fish Shellfish Immunol 2005;19:253e67.
[15] Paulsen SM, Sveinbjørnsson B, Robertsen B. Selective stainingand disintegration of intestinal eosinophilic granule cells inAtlantic salmon after intraperitoneal injection of the zincchelator dithizone. J Fish Biol 2001;58:768e75.
[16] Schmale MC, Vicha D, Cacal SM. Degranulation of eosinophilicgranule cells in neurofibromas and gastrointestinal tract in thebicolor damselfish. Fish Shellfish Immunol 2004;17:53e63.
[17] Ellis AE. Eosinophilic granular cells (EGC) and histamine re-sponses to Aeromonas salmonicida toxins in rainbow trout.Dev Comp Immunol 1985;9:251e60.
[18] Matsuyama T, Iida T. Influence of tilapia mast cell lysate onvascular permeability. Fish Shellfish Immunol 2001;11:549e56.
[19] Murray HM, Leggiadro CT, Douglas SE. Immunocytochemicallocalization of pleurocidin to the cytoplasmic granules of eo-sinophilic granular cells from the winter flounder gill. J FishBiol 2007;70:336e45.
[20] Dezfuli BS, Pironi F, Giari L, Domeneghini C, Bosi G. Effect ofPomphorhynchus laevis (Acanthocephala) on putative neuro-modulators in the intestine of naturally infected Salmo trutta.Dis Aquat Org 2002;51:27e35.
[21] Terenina NB, Asatrian AM, Movsessian SO. Neurochemicalchanges in rats infected with Trichinella spiralis and T.pseudospiralis. Dokl Biol Sci 1997;355:412e3.
[22] Leino RL. Reaction of rodlet cells to a myxosporean infectionin kidney of the bluegill, Lepomis macrochirus. Can J Zool1996;74:217e25.
[23] Dezfuli BS, Giari L, Shinn AP. The role of rodlet cells in theinflammatory response in Phoxinus phoxinus brains infectedwith Diplostomum. Fish Shellfish Immunol 2007;23:300e4.
[24] Silphaduang U, Colorni A, Noga EJ. Evidence for widespreaddistribution of piscidin antimicrobial peptides in teleost fish.Dis Aquat Org 2006;72:241e52.
[25] Mazon AF, Huising MO, Taverne-Thiele AJ, Bastiaans J,Verburg-van Kemenade BML. The first appearance of rodletcells in carp (Cyprinus carpio L) ontogeny and their possibleroles during stress and parasite infection. Fish ShellfishImmunol 2007;22:27e37.
[26] Penissi AB, Rudolph MI, Piezzi RS. Role of mast cells ingastrointestinal mucosal defence. Biocell 2003;27:163e72.
[27] Mekori YA. The mastocyte: the ‘‘other’’ inflammatory cell inimmunopathogenesis. J Allergy Clin Immunol 2004;114:52e7.

Brown trout’s inflammatory response to D. truttae 733
[28] Flano E, Lopez-Fierro P, Razquin BE, Villena A. In vitrodifferentiation of eosinophilic granular cells in Renibacteriumsalmoninarum-infected gill cultures from rainbow trout. FishShellfish Immunol 1996;6:173e84.
[29] Ezeasor DN, Stokoe WM. A cytochemical, light and electronmicroscopic study of the eosinophilic granule cells in the gutof the rainbow trout, Salmo gairdneri Richardson. J Fish Biol1980;17:619e34.
[30] Powell MD, Briand HA, Wright GM, Burka JF. Rainbow trout(Oncorhynchus mykiss Walbaum) intestinal eosinophilicgranule cell (EGC) response to Aeromonas salmonicida andVibrio anguillarum extracellular products. Fish ShellfishImmunol 1993;3:279e89.
[31] Kent ML, Powell MD, Kieser D, Hoskins GE, Speare DJ,Burka JF. Unusual eosinophilic granule cell proliferation incoho salmon (Oncorhynchus kisutch). J Comp Pathol 1993;109:129e40.
[32] Galli SJ. New insights into ‘‘The riddle of the mast cells’’:microenvironmental regulation of mast cells developmentand phenotypic heterogeneity. Lab Invest 1990;62:5e33.
[33] Temkin RJ, McMillan DB. Gut-associated lymphoid tissue(GALT) of the goldfish, Carassius auratus. J Morphol 1986;190:9e26.
[34] Metcalfe DD, Baram D, Mekori YA. Mast cells. Physiol Rev 1997;77:1033e79.
[35] Rocha JS, Chiarini-Garcia H. Mast cell heterogeneity betweentwo different species of Hoplias sp. (Characiformes:Erythrinidae): response to fixatives, anatomical distribution,histochemical contents and ultrastructural features. FishShellfish Immunol 2007;22:218e29.
[36] Venkova K, Davidoff M, Radomirov R. Effects of Met-enkepha-lin on the mechanical activity and distribution of Met-enkeph-alin-like immunoreactivity in the cat large intestine. Peptides1992;13:193e201.
[37] Radulovic J, Mancev Z, Stanojevic S, Vsiljevic T, Kovacevic-Jovanovic V, Pesic G. Modulation of humoral immune responseby central administration of leucine-enkephalin: effects ofmu, delta and kappa opioid receptor antagonists. J Neuroim-munol 1996;65:155e61.
[38] Wu SX, Li YQ, Shi JW. Temporal changes of preproenkephalinmRNA and leu-enkephalin-like immunoreactivity in theneurons of the caudal spinal trigeminal nucleus and uppercervical cord after noxious stimuli. J Hirnforschung 1998;39:217e22.
[39] Silphaduang U, Noga E. Peptide antibiotics in mast cells offish. Nature 2001;414:268e9.
[40] Khan NA, Deschaux P. Role of serotonin in fish immunomodu-lation. J Exp Biol 1997;200:1833e8.
[41] Lee TDG, Swieter M, Befus AD. Mast cells responses tohelminth infection. Parasitol Today 1986;2:186e91.




















