
Changes in plasma vitellogenin, sex steroids, calcitonin, and thyroid hormones related to sexual...
11
GENERAL AND COMPARATIVE ENDOCRINOLOGY 75, 316-326 (1989) Changes in Plasma Vitellogenin, Sex Steroids, Calcitonin, and Thyroid Hormones Related to Sexual Maturation in Female Brown Trout (Salvo trutta) BIRGITTA NORBERG,*~'BJ~RN TH. BJbRNSSON,*'t CHRISTOPHER L. Bnowtu,?* ULF-PETERWICHARDT,S LEONARD J. DEFTOS,§ AND CARL HAUX* *Department of Zoophysiology, P.O. Box 250 59, WOO 31 Giiteborg, Sweden; fDepartment of Zoology, University of California at Berkeley, California 94720; SSalmon Research Institute, S-810 70 &vkarleby, Sweden; and SDepartment of Medicine, San Diego Veterans Administration Medical Center and University of California San Diego, La Jolla, California 92161 Accepted December 20, 1988 Female brown trout (Salmo trutta) from a wild strain (Baltic sea trout) and a cultured strain were sampled individually for blood plasma at regular intervals during the period around final sexual maturation. The plasma samples were analyzed for vitellogenin (VTG), estradiol-17S, testosterone, 17a,20g-dihydroxy-4-pregnen-3-one (17,2O@P), calcitonin, tri- iodothyronine (Ts), thyroxine (T,,), and total and free plasma calcium. In the wild fish, VTG, estradiol-17l3, and testosterone peaked 30 days before ovulation, while 17,2O@P had a sharp peak at ovulation. Both Ts and T, declined at the beghming of the sampling period, reached minimal levels 30 days before ovulation, and rose sharply at the time of ovulation. Calcitonin levels were elevated during final maturation. Total plasma calcium correlated with plasma VTG levels. In the cultured strain, sampling was started 2 weeks before ovulation. The levels of VTG, estradiol-17g, and testosterone decreased throughout the sampling period. 17,2Og-P and calcitonin concentrations were high during the period close to ovulation. Plasma thyroxine remained at basal levels in cultured trout. The discrepancies observed between wild and cultured females may be due to differences in stress susceptibility, environmental conditions, life cycles, or to genetic divergence between the strains. Q1989 Academic Press, Inc. Liver synthesis of the yolk’ protein pre- cursor vitellogenin (VTG) is induced by es- trogens, particularly estradiol-178 (Bailey, 1957; Chen, 1983; Norberg and Haux, 1985). The protein is rapidly secreted and transported to the ovaries where it is se- questered into the growing oocytes and transformed into yolk proteins during the period of rapid gonadal growth (cf. Wal- lace, 1985). Changes in plasma VTG levels reflect the different stages of the gonadal growth cycle in salmonid fish (So et al., r Current address: Directorate of Fisheries, Insti- tute of Marine Research, Austevoli Marine Aquacul- ture Station N-5392 Storebii, Norway. * Current address: Hawaii Institute of Marine Biol- ogy, University of Hawaii at Manoa, Coconut Island, Kaneohe, HI 96744. 1985; Copeland et al., 1986; Dye et al., 1986; Tam et al., 1986). A period of slow but steady growth of the gonads, with little incorporation of yolk proteins (yolk vesicle formation), is followed by a rapid growth phase due to the incorporation of vitello- genin (true vitellogenesis; Wallace and Sel- man, 1981). Sexual maturation in female salmonids involves typical changes in the plasma lev- els of the major ovarian steroids estradiol- 17/Z!, testosterone, and 17cr,20@OH-4- pregnen-3-one (17,208-P). Synthesis of these steroids is under pituitary control and is induced by gonadotropins (cf. Naga- hama, 1987a). Both estradiol-17/3 and tes- tosterone levels rise several months before sexual maturation, but while estradiol-17P 316 001~6480/89 $1.50 Copyright Q 1989 by Academic Press, Inc. All rig& of nproduction in my form re.servcd.
Transcript of Changes in plasma vitellogenin, sex steroids, calcitonin, and thyroid hormones related to sexual...

GENERAL AND COMPARATIVE ENDOCRINOLOGY 75, 316-326 (1989)
Changes in Plasma Vitellogenin, Sex Steroids, Calcitonin, and Thyroid Hormones Related to Sexual Maturation in Female Brown
Trout (Salvo trutta)
BIRGITTA NORBERG,*~'BJ~RN TH. BJbRNSSON,*'t CHRISTOPHER L. Bnowtu,?* ULF-PETERWICHARDT,S LEONARD J. DEFTOS,§ AND CARL HAUX*
*Department of Zoophysiology, P.O. Box 250 59, WOO 31 Giiteborg, Sweden; fDepartment of Zoology, University of California at Berkeley, California 94720; SSalmon Research Institute,
S-810 70 &vkarleby, Sweden; and SDepartment of Medicine, San Diego Veterans Administration Medical Center and University of California San Diego, La Jolla, California 92161
Accepted December 20, 1988
Female brown trout (Salmo trutta) from a wild strain (Baltic sea trout) and a cultured strain were sampled individually for blood plasma at regular intervals during the period around final sexual maturation. The plasma samples were analyzed for vitellogenin (VTG), estradiol-17S, testosterone, 17a,20g-dihydroxy-4-pregnen-3-one (17,2O@P), calcitonin, tri- iodothyronine (Ts), thyroxine (T,,), and total and free plasma calcium. In the wild fish, VTG, estradiol-17l3, and testosterone peaked 30 days before ovulation, while 17,2O@P had a sharp peak at ovulation. Both Ts and T, declined at the beghming of the sampling period, reached minimal levels 30 days before ovulation, and rose sharply at the time of ovulation. Calcitonin levels were elevated during final maturation. Total plasma calcium correlated with plasma VTG levels. In the cultured strain, sampling was started 2 weeks before ovulation. The levels of VTG, estradiol-17g, and testosterone decreased throughout the sampling period. 17,2Og-P and calcitonin concentrations were high during the period close to ovulation. Plasma thyroxine remained at basal levels in cultured trout. The discrepancies observed between wild and cultured females may be due to differences in stress susceptibility, environmental conditions, life cycles, or to genetic divergence between the strains. Q1989 Academic Press, Inc.
Liver synthesis of the yolk’ protein pre- cursor vitellogenin (VTG) is induced by es- trogens, particularly estradiol-178 (Bailey, 1957; Chen, 1983; Norberg and Haux, 1985). The protein is rapidly secreted and transported to the ovaries where it is se- questered into the growing oocytes and transformed into yolk proteins during the period of rapid gonadal growth (cf. Wal- lace, 1985). Changes in plasma VTG levels reflect the different stages of the gonadal growth cycle in salmonid fish (So et al.,
r Current address: Directorate of Fisheries, Insti- tute of Marine Research, Austevoli Marine Aquacul- ture Station N-5392 Storebii, Norway.
* Current address: Hawaii Institute of Marine Biol- ogy, University of Hawaii at Manoa, Coconut Island, Kaneohe, HI 96744.
1985; Copeland et al., 1986; Dye et al., 1986; Tam et al., 1986). A period of slow but steady growth of the gonads, with little incorporation of yolk proteins (yolk vesicle formation), is followed by a rapid growth phase due to the incorporation of vitello- genin (true vitellogenesis; Wallace and Sel- man, 1981).
Sexual maturation in female salmonids involves typical changes in the plasma lev- els of the major ovarian steroids estradiol- 17/Z!, testosterone, and 17cr,20@OH-4- pregnen-3-one (17,208-P). Synthesis of these steroids is under pituitary control and is induced by gonadotropins (cf. Naga- hama, 1987a). Both estradiol-17/3 and tes- tosterone levels rise several months before sexual maturation, but while estradiol-17P
316 001~6480/89 $1.50 Copyright Q 1989 by Academic Press, Inc. All rig& of nproduction in my form re.servcd.

PLASMA HORMONES IN TROUT 317
levels decrease some weeks before final maturation and are low at ovulation, testos- terone remains elevated and starts to de- crease only during the last week preceding ovulation (Scott et al., 1983). Plasma 17,20@P increases sharply during final mat- uration and ovulation, but is otherwise low (Wright and Hunt, 1982; Scott et al., 1983; cf. Goetz et al., 1987). Experimental stud- ies on the events that accompany final oocyte maturation and ovulation in vivo and in vitro have confirmed the role of 17,2Op-P as the maturation-inducing steroid in salmonid fish (cf. Nagahama, 1987b).
There is ample reason to suspect involve- ment of thyroid hormones in the reproduc- tive processes of salmonids. Cyclic changes in thyroid function, in particular plasma thyroid hormone levels, appear to be syn- chronized with gonadal maturation and/or spawning in a variety of salmonid species (cf. Leatherland, 1982; Brown, 1988). At present, specific actions of thyroid hor- mones in salmonid reproduction have not been established.
The physiological function of calcitonin in teleost fish is a matter of speculation. A calcium regulatory function is possible, but available data are conflicting, as calcitonin induces hypocalcemia in some species but not in others (cf. Bjiimsson ef al., 1986; Fenwick and Lam, 1988). Histological and physiological data indicate that calcitonin has a female-specific role during salmonid reproduction (Deville and Lopez, 1970; Watts et al., 1975; Yamane and Yamada, 1977; Bjiirnsson and Deftos, 1985; Bjomsson et al., 1986). In female rainbow trout, calcitonin levels show a distinct peak immediately prior to ovulation and then rapidly fall again, while no plasma calcito- nin changes occur in males (Bjomsson et al., 1986).
Recent studies on the regulation of repro- duction in wild sahnonids have been made on Oncorhynchus species (Ueda et al., 1984; Yamauchi et al., 1984; Dye et al., 1986; Fitzpatrick ef al., 1986), while avail- able information on other salmonids fo-
cuses on cultured fish, mainly the rainbow trout. Data on the reproductive endocrinol- ogy in the brown trout are limited and con- cern the levels of gonadotropin, estradiol- 17B, and VTG during different phases of vitellogenesis in wild females (Crim and Idler, 1978) and the annual variations of estradiol-17B and gonadotropin in cultured trout (Breton et al., 1983).
The present study attempts to increase the understanding of the endocrine changes that occur during sexual maturation and ovulation in female brown trout. The hor- mone profiles were followed during vitello- genesis and final maturation in wild trout, while a short-term study of the changes that occur during final maturation was made in cultured females. The sampling protocols were designed to get a high resolution of the various hormone profiles since rapid changes in plasma hormone levels may take place.
MATERIALS AND METHODS
Fish, holding conditions, and sampling. Wild Baltic sea trout (S. trutta) were captured during their spawn- ing migration up the river Dal%lven in central Sweden, in July and August. The average body weight was 3.0 f 0.2 kg (n = 21). No significant sex differences in body weight were observed. The fish were individually tagged and kept at the Salmon Research Institute, Alvkarleby (60.6” N), in a 63-m* outdoor tank (volume ca 30 m3) supplied with 800 liters . mitt- ’ of unfiltered river water. No food was given during the sampling period. The water temperature was 18.4” on July 25 and decreased throughout the sampling period to 0.1” on November 28. Before blood sampling, the fish were anesthetized for 5 min in MS-222. In October, the fe- males were checked for ovulation at each sampling occasion by gentle pressure on the abdomen. Blood samples were taken by heart puncture, at lO- to 14-day intervals from the beginning of August until the end of November. The females ovulated by the end of Octo- ber. Eleven of the females were sampled in May of the following year. Although both male and female fish were kept together in the tank, only females (n = 16) were included in the present study.
Sexually maturing female brown trout (S. trutta) (n = 10) of a cultured strain (Bonashamn trout), that spend their entire life cycle in fresh water, were kept under a natural photoperiod in a 9-m* tank with a wa- ter volume of 3 m3 at the broodstock fish farm of the Indals&.lvens Vattenregleringsaktiebolag at Bo-

318 NORBERG ET AL.
niishamn in central Sweden (63.4” N). The tank was supplied with running fresh water and the water tem- perature was 7.0” at the beginning of the sampling pe- riod on October 15 and 5.0” at the end of the sampling period on November 3. The fish were not fed during the sampling period. On October IS, the average weight of the fish was 0.90 ? 0.03 kg (n = 10). On November 3, all the fish were spent and their average weight was 0.80 2 0.05 kg (n = IO). The fish were individually marked by fin clipping. Blood samples were collected at 3-day intervals during the ovulatory period in the month of October. At each sampling oc- casion, the fish were anesthetized in 0.1% 2- phenoxyethanol (Kebo AB, Sweden). The anesthe- tized fish were checked for ovulation and blood was withdrawn from the duct of Cuvier into a heparinized syringe (Lied et al., 1975). All blood samples were immediately centrifuged and the plasma was deep fro- zen on dry ice. The plasma samples were stored at - 20” until analyzed.
Radioimmunoassays. The levels of plasma VTG were measured by radioimmunoassay (Norberg and Haux, 1988). Samples with VTG concentrations over 20 ng . ml-’ were diluted in order to fall within the range of the standard curve (l-20 ng VTG . ml-‘).
Steroids were extracted from 50 pl of plasma ac- cording to Castellanos Perez (1979): 2 ml of a mixture of diethylether (Merck) and heptane (Merck) (4: 1) was added to the plasma sample in a pyrex tube with a tight screw cap. The tube was shaken for 1 min and dipped in liquid nitrogen for 5 sec. The organic phase was poured off into a glass tube and evaporated at 35”. The residue was dissolved in 1 ml of phosphate-buffered saline (0.05 M Na-phosphate buffer, pH 7.6; 0.9% NaCl) and stored at - 20” until analyzed. The extrac- tion yields for testosterone, estradiol-17S, and 17,2Op-P, respectively, were assessed by measuring the recovery of known amounts of ‘H-labeled steroids (Amersham) from juvenile brown trout plasma.
Steroid levels were determined by previously de- scribed tadioimmunoassays for estradiol-17B (Scott et al., 198Oa), testosterone (Scott ef al., 1980b), and 17,2OB-P (Scott et al., 1982), as modified by Scott and Sumpter (1983). Testosterone, estradiol-17g, and 17,2Og-P were purchased from Sigma and ‘H-labeled steroids were purchased from Amersham. Labeled 17,2Og-P was prepared from 3H-labeled 17a-proges- terone as described by Scott et al. (1982).
Plasma levels of thyroxine (T.,) and triiodothyronine (Ts) were determined by radioimmunoassay (Specker and Richman, 1984).
Plasma calcitonin levels were determined by a ra- dioimmunoassay for salmon calcitonin I (Deftos et al.,
1974). Plasma and ultrafiltrate analyses. About 0.3 ml of
each plasma sample from the cultured brown trout was ultrafiltered (Amicon, MPS-1 System) in order to ob- tain a protein-free ultrafiltrate. Subsequently, the
plasma and ultratiltrate samples were analyzed by atomic absorption spectroscopy (Perkin Elmer 403) for total (Ca.r) and free (Car) plasma calcium, respec- tively. Plasma from the sea trout was similarly ana- lyzed for Car.
RESULTS
Vitellogenin
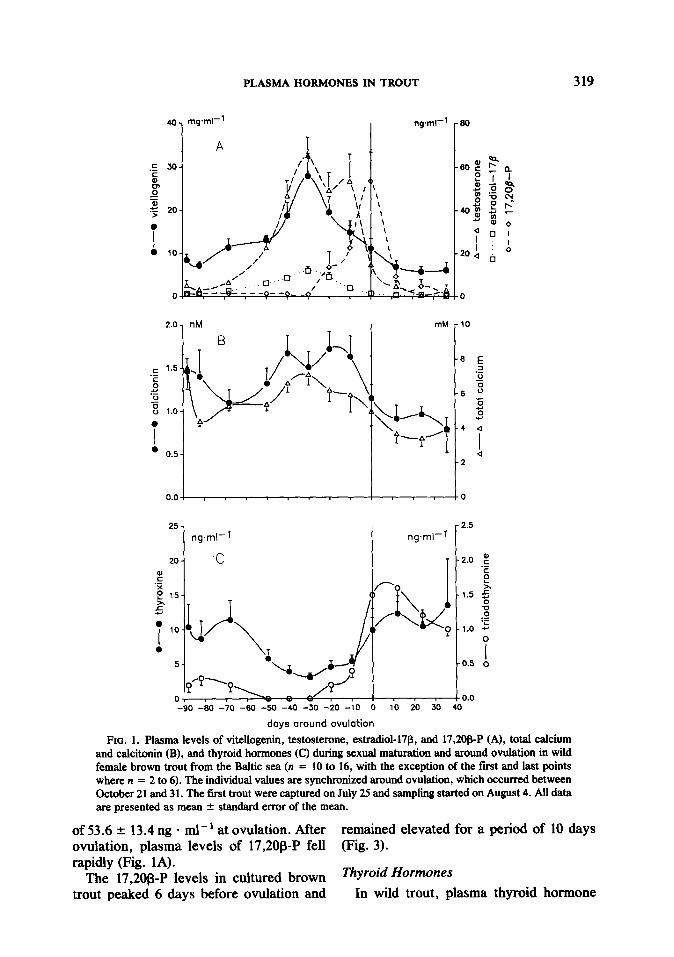
The plasma VTG levels of wild sea trout females increased and reached a peak value of 28.0 r 5.2 mg . ml-’ 30 days prior to ovulation (Fig. 1A). The VTG levels then gradually declined, but remained elevated throughout the sampling period. In May of the following year, the VTG levels had de- creased to 1.1 + 0.3 mg . ml-‘.
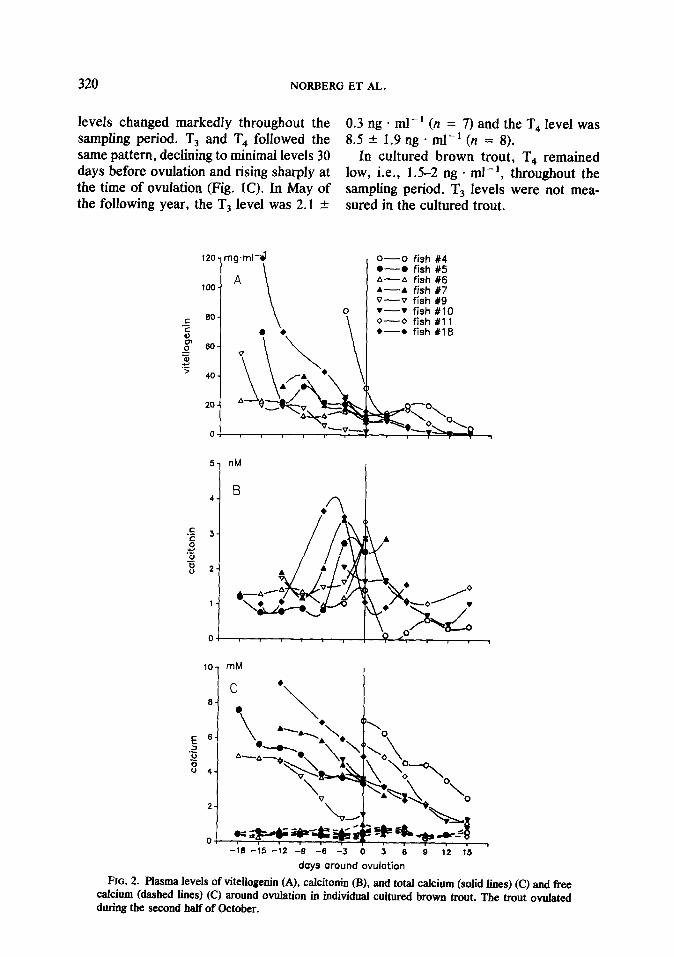
In cultured brown trout, the levels of VTG were the highest (>lOO mg . ml-’ in some fish) at the beginning and declined throughout the sampling period. At ovula- tion, the average VTG level was ca. 20 mg + ml-‘, while the average concentration 15 days after ovulation was 2 mg * ml - ’ (Fig. 2A).
Steroids
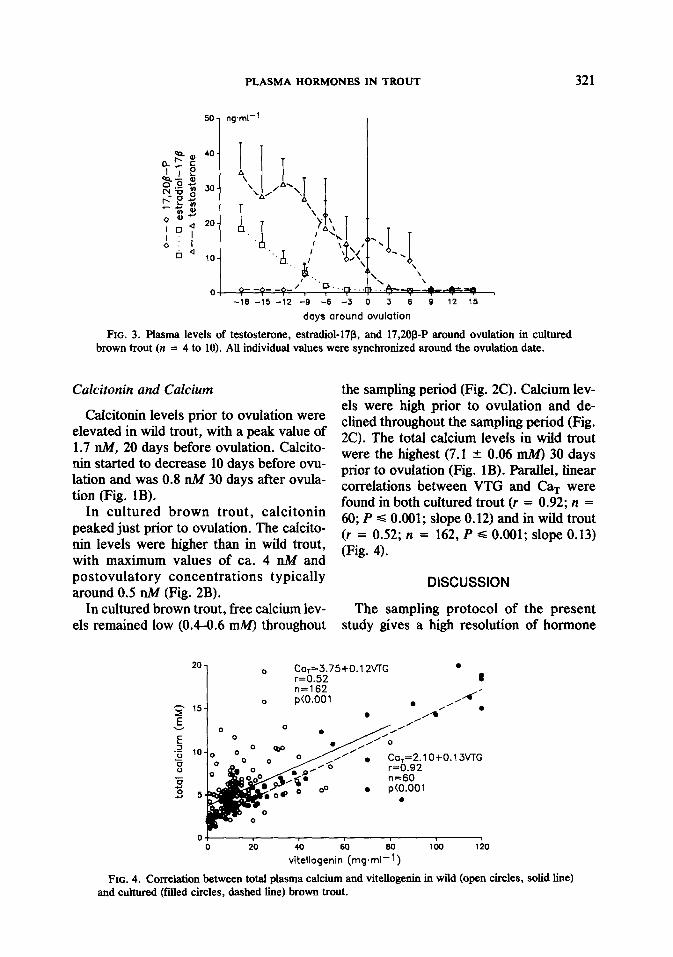
The levels of testosterone and estradiol- 17p in wild trout increased and reached peak values 30 days before ovulation (Fig. IA). The maximum testosterone concentra- tion was five times higher than the maxi- mum estradiol concentration (65.2 + 8.5 ng . ml-’ and 12.0 ? 2.0 ng * ml-‘, respec- tively). The changes in estradiol-17p fol- lowed the blood VTG concentration, while testosterone levels remained elevated until ovulation. Two weeks after ovulation, estradiol-17B and testosterone had de- creased to levels of ca. 0.5 ng * ml- ’ (Fig. 1A). In cultured fish, testosterone and estradiol-17l3 levels were the highest at the beginning and declined throughout the sam- pling period (Fig. 3).
In wild trout, levels of 17,20@P were sta- ble at ca. 1 ng - ml-’ until 20 days before ovulation and reached a distinct peak value
40
.E 30 % ET
=
.: 20
i l 10
i
i 0 d
PLASMA HORMONES IN TROUT
mg.ml-’ I
ng.ml-’
319
25
1
ng.ml-l
20 ,C
-90 -80 -70 -60 -50 -40 -30 -20 -10 0 10 20 30 40
days around ovulation
FIG. 1. Plasma levels of vitellogenin, testosterone, estradiol-17J3, and 17,208-P (A), total calcium and calcitonin (B), and thyroid hormones (C) during sexual maturation and around ovulation in wild female brown trout from the Baltic sea (n = 10 to 16, with the exception of the fast and last points where n = 2 to 6). The individual values are synchronized around ovulation, which occurred between October 21 and 31. The first trout were captured on July 25 and sampling started on August 4. All data are presented as mean + standard error of the mean.
of 53.6 -+ 13.4 ng - ml-’ at ovulation. After remained elevated for a period of 10 days ovulation, plasma levels of 17,20@P fell (Fig. 3). rapidly (Fig. 1A).
The 17,20@P levels in cultured brown ThYroid Hormones trout peaked 6 days before ovulation and In wild trout, plasma thyroid hormone
320 NORBERG ET AL.
levels changed markedly throughout the sampling period. T, and T4 followed the same pattern, declining to minimal levels 30 days before ovulation and rising sharply at the time of ovulation (Fig. 1C). In May of the following year, the T3 level was 2.1 -t
120 mg.ml-i)
100 A
1 \
0.3 ng . ml-’ (n = 7) and the T, level was 8.5 + 1.9 ng * ml-’ (n = 8).
In cultured brown trout, T, remained low, i.e., 1.5-Z ng . ml-‘, throughout the sampling period. TS levels were not mea- sured in the cultured trout.
o-o fish %4 O-O fish 15 A-A fish X6 A-A fish 67 v--v fish #9
0 v--v fish AI10
\
0-O fish #ll a---* fish X18
10 mM 1
days around ovulation
FIG. 2. Plasma levels of vitellogenin (A), calcitonin (B), and total calcium (solid lines) (C) and fkee calcium (dashed lines) (C) around ovulation in individual cultured brown trout. The trout ovulated during the second half of October.
PLASMA HORMONES IN TROUT 321
-18 -15 -12 -9 -6 -3 0 3 6 9 12 15
days around ovulation
FIG. 3. Plasma levels of testosterone, estradiol-17@, and 17,208-P around ovulation in cultured brown trout (n = 4 to 10). All individual values were synchronized around the ovulation date.
Calcitonin and Calcium
Calcitonin levels prior to ovulation were elevated in wild trout, with a peak value of 1.7 IN, 20 days before ovulation. Calcito- nin started to decrease 10 days before ovu- lation and was 0.8 nM 30 days after ovula- tion (Fig. 1B).
In cultured brown trout, calcitonin peaked just prior to ovulation. The calcito- nin levels were higher than in wild trout, with maximum values of ca. 4 nA4 and
the sampling period (Fig. 2C). Calcium lev- els were high prior to ovulation and de- clined throughout the sampling period (Fig. 2C). The total calcium levels in wild trout were the highest (7.1 & 0.06 mM) 30 days prior to ovulation (Fig. 1B). Parallel, linear correlations between VTG and Car were found in both cultured trout (r = 0.92; n = 60; P s 0.001; slope 0.12) and in wild trout (r = 0.52; n = 162, P c 0.001; slope 0.13) (Fig. 4).
postovulatory concentrations typically around 0.5 nA4 (Fig. 2B).
DISCUSSION
In cultured brown trout, free calcium lev- The sampling protocol of the present els remained low (0.4-0.6 mM) throughout study gives a high resolution of hormone
20
1
0 CaT=3.75+0.1 2VTG r=0.52 n=162
0 p(O.001
. 8
_. . A-+-
/’ .
Ca,=2.1 O+O.lJWG r=0.92 n=60 p(O.001
.
1 0 20 40 60 60 100 120
vitellagenin (mg.ml-l)
FIG. 4. Correlation between total plasma calcium and vitellogenin in wild (open circles, solid line) and cultured (flied circles, dashed line) brown trout.

322 NORBERG ET AL.
and VTG profiles during final sexual matu- ration of female brown trout. In wild trout, VTG levels were elevated at the beginning of August and reached a peak level at the end of September. During the month pre- ceding ovulation, plasma VTG decreased. The cultured females had higher maximal plasma VTG concentrations than wild fe- males, with the highest observed concen- tration being 2120 mg * ml-‘. A higher nu- tritional status of the cultured trout, due to a better food supply, is one possible cause of the higher VTG levels. The levels of estradioL17B 2 weeks before ovulation were also higher in the cultured trout than in the wild trout.
The data indicate that an overshoot of VTG is present in the plasma of spent fish, which is cleared from the circulation rela- tively slow. A similar pattern was seen in the rainbow trout (Scott and Sumpter, 1983). In pink salmon (0. gorbuscha; Dye et al., 1986) and chum salmon (0. keta; Ueda et al., 1984), VTG fell more rapidly and to lower levels during the late phase of spawning migration than in the Baltic sea trout of the present study. Most Oncorhyn- thus species reach maturity and spawn only once during their life cycle. There would be little use for “leftover” VTG in spent fish since they die shortly after spawning. In species that spawn annually, like the trout, VTG that is not sequestered by the oocyte may be resorbed and serve as an energy source for the adult fish after spawning. Plasma levels of yolk proteins in rainbow trout are elevated after ovulation, due to the resorption of yolk during follic- ular atresia (Babin, 1987). The same may be true for the brown trout. Oocytes from On- corhynchus species may also be able to se- quester VTG more efficiently and during a period closer to maturation than oocytes from brown trout and rainbow trout.
Total and protein-bound plasma calcium levels were highly correlated with VTG lev- els in cultured trout. In the wild trout, the correlation between total calcium and VTG
was also highly significant. This suggests that determinations of total calcium may be a good indicator of VTG in brown trout. However, estimations of VTG content are more accurately carried out with other di- rect methods. Elliot et al. (1979) measured serum concentrations of protein-bound phosphorus and total calcium in estradiol- treated rainbow trout. Based on the corre- lation between protein-bound phosphorus and total calcium, they suggested a 1: 1 ratio between these components in rainbow trout VTG. Given the phosphorus content in brown trout VTG to be 0.58% (Norberg and Haux, 1985), the average molar ratio of phosphorus and protein-bound calcium was 1.1 -’ 0.1 in the cultured trout. Thus, the ratio between phosphorus and calcium ap- pears to be the same in brown trout as in rainbow trout.
Estradiol-17B induces the synthesis of VTG both in viva (Bailey, 1957; Chen, 1983; Norberg and Haux, 1985) and in vitro (Maitre et al., 1986). VTG followed the lev- els of estradiol-17B and decreased 1 month before ovulation. In the cultured trout, estradiol-17B decreased throughout the sampling period. This indicates a similar pattern in both wild and cultured fish. Estradiol- 17B levels reported in female brown trout (Crim and Idler, 1978; Breton et al., 1983; this study) are lower than in other Salmo species, but are similar to estradiol-l7B levels in Oncorhynchus spe- cies and Salvelinus fontinalis (cf. Goetz et al., 1987). Confinement and handling stress suppress plasma androgen levels in male brown trout (Pickering et al., 1987), and cortisol may reduce the levels of vitello- genin and estradiol-17B in female brown trout and rainbow trout (Carragher, Sump- ter, Pottinger, and Pickering, unpublished results). Thus, the lower vitellogenin and estradiol-17p levels in wild females com- pared to cultured females in the present study may be due to a higher susceptibility to stress in the wild trout.
Testosterone increased concomitantly

PLASMA HORMONES IN TROUT 323
with VTG and remained elevated until the week before ovulation in the wild females. When 17,208-P started to rise, testosterone decreased, but average levels of testos- terone were still 20 times higher than es- tradiol-178 at ovulation. The physiological function of testosterone in female fish is un- clear at present. Testosterone is used as a substrate in estradiol-178 synthesis (Scott et al., 1983). The role of testosterone as a substrate for estradiol-178 may not be the sole function in view of the high levels of testosterone in both wild and cultured fe- males. Fitzpatrick et al. (1987) found that female coho salmon (0. kisutch) showing an ovulatory response upon treatment with gonadotropin-releasing hormone (GnRH) had significantly higher testosterone levels before hormone administration than fe- males that did not respond to GnRH treat- ment. A high testosterone concentration may thus be a prerequisite for final oocyte maturation. This hypothesis is supported by in vitro studies of follicles from amago salmon (Young et al., 1982) and rainbow trout (Jalabert and Fostier, 1984) that have shown that testosterone enhances germinal vesicle breakdown in response to gonado- tropin treatment.
Around the time of ovulation, 17,208-P was elevated in both wild and cultured trout, with a sharp peak at ovulation in the wild fish. The profile of 17,2Op-P in the brown trout is somewhat different from that in the rainbow trout where the levels of 17,20@P rose sharply between 8 and 4 days before ovulation but only decreased slowly and remained elevated for as long as 28 days postovulation (Scott et al., 1983). However, the peak levels of 17,208-P in the rainbow trout seem to be considerably higher than in other salmonids (cf. Goetz et al., 1987; this study), with the exception of the chum salmon (Ueda et al., 1984). This may be due to a slower elimination of 17,2Op-P from the plasma of the rainbow trout as compared to the brown trout.
Plasma thyroid hormones in the wild
trout were lowest 50-30 days before ovula- tion, as vitellogenin, testosterone, estra- diol- 17p, calcitonin, and plasma calcium all increased. One plausible explanation for this pattern is that thyroid hormones were rapidly being deposited in the growing oocytes at that time (see Kobuke et al., 1987; Brown and BerfiT1989). Some of the earlier reports are compatible with this hy- pothetical explanation. A decline in plasma thyroid hormone levels during the late stages of sexual maturation has been re- ported in some salmonids. In several of these studies the decline has been more pronounced in female than in male fish (Sower and Schreck, 1982; Biddiscombe and Idler, 1983; Dickhoff et al., 1989).
Thyroid hormone levels increased two- to threefold in wild trout at the time of spawning and remained elevated. These data are in agreement with earlier observa- tions on cultured female brown trout (Pick- ering and Christie, 1981). T, injections ac- celerate the final maturation of salmonid eggs. The injections were the most effective when given in conjunction with GnRH, sug- gesting the induction of enhanced ovarian sensitivity to GtH by thyroid hormones (Dickhoff et al., 1989). Thus, it is possible in the present study that the increase in plasma thyroid hormones around the time of ovulation had an influence on the final maturation of oocytes. The absence of any appreciable change in the profile of circu- lating T4 of the cultured trout implies that large increases in plasma thyroid hormones are not necessary for ovarian maturation.
Calcitonin levels were elevated before ovulation and decreased following ovula- tion in both strains of female brown trout as in rainbow trout (Bjiimsson et al., 1986). These observations, together with earlier histological and physiological data (Deville and Lopez, 1970; Watts et al., 1975; Ya- mane and Yamada, 1977, Bjomsson and Deftos, 1985; Bjiimsson et al., 1986), thus strongly suggest that the function of calci- tonin in salmonids is related to some event

324 NORBERG ET AL.
late in female sexual maturation. It is un- likely that this female-specific role is linked to calcium regulation. First, although large changes in calcium turnover must take place during vitellogenesis, the increase and subsequent decrease of calcitonin lev- els occur at a later stage in the maturation process, around ovulation. Second, if a pre- sumptive hypocalcemic role of calcitonin is accepted, a decrease in calcitonin levels during vitellogenesis should be postulated, as the physiological demand for calcium during vitellogenesis requires an increase in the flux of calcium ions from reservoirs to the circulation (Fleming et al., 1964; Mugiya and Watabe, 1977; Bjiirnsson et al., 1986). Such a change in calcium turnover would be hindered by increased levels of a hypocalcemic hormone.
In conclusion, the present study provides important information on the physiology and endocrinology of final sexual matura- tion in female brown trout. The overall pat- terns were similar in both wild and cultured fish, although some differences are appar- ent: Plasma levels of both estradiol-17P and VTG were considerably higher in cultured fish than in wild females. On the other hand, plasma levels of 17,20@P were higher in the wild trout. Distinct changes in plasma thyroid hormones were evident in the wild trout, but not in the cultured trout. The hypothesis of calcitonin as a reproduc- tive hormone in salmonid fish is supported. The dissimilarities between the wild and the cultured females may be due to differences in stress susceptibility, environmental con- ditions, or life cycles. In addition, the ge- netic divergence between the two different strains may contribute to the observed vari- ations.
ACKNOWLEDGMENTS
We gratefully acknowledge Dr. A. P. Scott for gen- erously providing antisera to steroid hormones. We also thank Ms. Aina Stenborg, Ms. Inga Maj &born, MS. Cheryl Chalbetg, and Mr. Brian Williams for ex- cellent technical assistance. We are indebted to Mr.
Ame Klitgaard for providing excellent facilities at the Boniishamn broodstock farm. The study was financed by Stiftelsen Lam Hiertas Minne, Anna Ahrenbergs Fond, Kungliga och Hvitfeldtska Stipendieitittnin- gen, and Wilhelm och Martina Lundgrens Vetenskaps- fond. Further, this work is a result of research spon- sored in part by NOAA, National Sea Grant College Program, Department of Commerce, under Grant NA80AA-DO0120, through the California Sea Grant College Program, and in part by the California State Resources Agency, project No. m-101. The U.S. Government is authorized to reproduce and to distrib- ute for governmental purposes.
REFERENCES
Babin, P. J. (1987). Apolipoproteins and the associa- tion of egg yolk proteins with plasma high density lipoproteins after ovulation and follicular atresia in the rainbow trout (Salmo gairdneri). J. Biol. Chem. 242,4290-42%.
Bailey, R. E. (1957). The effect of estradiol on serum calcium, phosphorus and protein of goldfish. J. Exp. Zool. 136, 455-469.
Biddiscombe, S., and Idler, D. R. (1983). Plasma lev- els of thyroid hormones in sockeye salmon (On- corhynchus nerka) decrease before spawning. Gen. Comp. Endocrinol. 52, 467-470.
Bjomsson B. Th., and Deftos, L. J. (1985). Plasma calcium and calcitonin in the marine teleost, Ga- dus morhua. Camp. Biochem. Physiol. A 81,593- 5%.
Bjdmsson, B. Th., Haux, C., Forlin, L., and Deftos, L. J. (1986). The involvement of calcitonin in the reproductive physiology of the rainbow trout. 1. Endocrinol. 108, 17-23.
Breton, B., Fostier, A., Zohar, Y., Le Bail, P. Y ,, and Billard, R. (1983). Gonadotropine glycoproteique maturante et oestradiol-17fl pendant le cycle re- producteur chez la truite fario (Salmo trutru) femelle. Gen. Comp. Endocrinol. 49,220-231.
Brown, C. L., (1988). Photoperiod effects on thyroid function in fish. In ‘Processing of Environmental Information in Vertebrates” (M. H. Stetson, Ed.), pp. l-17. Springer Verlag, New York.
Brown, C. L., and Bern, H. A. (1989). Thyroid hor- mones in early development, with special refer- ence to teleost fishes. In “Development, Matura- tion, and Senescence of the Neuroendocrine System” (M. P. Schreibman and C. G. Scanes, Eds.). Academic Press, New York, in press.
Castellanos Perez, R. (1979). “Evaluation of a Proges- terone Radioimmunoassay and Its Application in Cuban Cattle.” Rapport Sveriges lantbruksuni- versitet, Inst. for klinisk kemi, 034&8659; 1.
Chen, T. T. (1983). Identification and characterization of estrogen-responsive gene products in the liver

PLASMA HORMONES IN TROUT 325
of rainbow trout. Canad. J. Biochem. Ceil Biol. 61, 802-810.
Copeland, P. A., Sumpter, J. P., Walker, T. K., and Croft, M. (1986). Vitellogenln levels in male and female rainbow trout (Salmo gairdneri Richard- son) at various stages of the reproductive cycle. Comp. Biochem. Physiol. B 83,487-493.
Grim, L. W., and Idler, D. R. (1978). Plasma gonado- tropin, estradiol and vitellogenin and gonad phos- vitin levels in relation to the seasonal reproduc- tive cycle of female brown trout. Ann. Biol. Anim. Biochim. Biophys. 18, 1081-1005.
Deftos, L. J., Watts, E. G., Copp, D. H., and Potts, J. T., Jr. (1974). A radioimmunoassay for salmon calcitonin. Endocrinology 94, 155-160.
Deville, I., and Lopez, E. (1970). Le corps ultimo- bran&al du saumon, Salmo salar: Etude histo- physiologique a diverses &apes de son cycle vital en eau deuce. Arch. Anat. Microsc. 59, 393-482.
Dickhoff, W. W., Yan, L., Plisetskaya, E., Sullivan, C. V., Swanson, P., and Bernard, M. (1989). Re- lationship between metabolic and reproductive hormones in salmonid fish. In “Proceedings of I. International Symposium on Fish Endocrinology, Edmonton, 1988” (R. Peter, P. K. T. Pang, and M. P. Schreibman, Eds.), in press.
Dye, H. M., Sumpter, J. P., Fagerlund, U. H. M., and Donaldson, E. M. (1986). Changes in repro- ductive parameters during the spawning migration of pink salmon, Oncorhynchus gorbuscka (Wat- baum). J. Fish Biol. 29, 167-176.
Elliot, J. A. K., Bromage, N. R., and Whitehead, C. (1979). Effects of oestradiol-178 on serum calcium and vitellogenin levels in rainbow trout. J. Endo- crinol. 83, 54P-55P.
Fenwick, J. C., and Lam, T. J. (1988). Effects of cai- citonin on plasma calcium and phosphate in the mudskipper, Periophtalmodon schlosseri (Teleo- stei), in water and during exposure to air. Gen. Comp. Endocrinol. 70.224-230.
Fitzpatrick, M. S., Redding, J. M., Ratti, F. D., and Schreck, C. B. (1987). Plasma testosterone con- centration predicts the ovulatory response of coho salmon (Oncorhynchus kisutch) to gonado- tropin-releasing hormone analog. Canad. J. Fish. Aquat. Sci. 44, 1351-1357.
Fitzpatrick, M. S., Van Der Kraak, G., and Schreck, C. B. (1986). Profiles of plasma sex steroids and gonadotropin in coho salmon, Oncorhynchus kisutch, during final maturation. Gen. Comp. En- docrinol. 62, 437-451.
Fleming, W. R., Stanley, J. G., and Meier, A. H. (1964). Seasonal effects of external calcium, es- tradiol, and ACTH on the serum calcium and so- dium levels of Fundulus kansae. Gen. Comp. En- docrinol. 4, 61-67.
Goetz, F. W., Fostier, A. Y., Breton, B., and Jala-
bert, B. (1987). Hormonal changes during meiotic maturation and ovulation in the brook trout (Salvelinus fontinalis). Fish Physiol. B&hem. 3, 203-211.
Jalabert, B., and Fostier, A. (1984). The modulatory effect in vitro of oestradiol-17B, testosterone or cortisol on the output of 17u-hydroxy-ZO@dihy- droprogesterone by rainbow trout (Salmo gaird- nerd) ovarian follicles stimulated by the matura- tional gonadotropin s-GtH. Reprod. Nutr. Dev. 24, 127-136.
Kobuke, L., Specker, J. L., and Bern, H. A. (1987). Thyroxine content in eggs and larvae of coho salmon, Oncorhynchus kisutch. J. Exp. Zool. 242, 89-94.
Leatherland, J. F. (1982). Environmental physiology of the telecstean thyroid gland: A review. Envi- ron. Biol. Fishes 7, 83-110.
Lied, E., Gjerde, J., and Braekkan, 0. R. (1975). Sim- ple and rapid technique for repeated blood sam- pling in rainbow trout (Saimo gairdnert). 1. Fish. Res. Board. Canad. 32, 699-701.
Maltre, J. L., Valotaire, Y., and Guguen-Guillouzo, C. (1986). Estradiol stimulation of vitellogenin synthesis in primary culture of male rainbow trout hepatocyes. In Vitro Cell. Dev. Biol. 22,337-362.
Mugiya, Y., and Watabe, N. (1977). Studies on fish scale formation and resorption. II. Effect of es- tradiol on calcium homeostasis and skeletal tissue resorption in the goldfish, Carassius auratus, and the killiish, Fundulus heteroclitus. Comp. Bio- them. Physiol. A 57, 197-202.
Nagahama, Y. (1987a). Gonadotropin action on game- togenesis and steroidogenesis in teleost gonads. Zool. Sci. 4, 289-222.
Nagahama, Y. (1987b). 17a,208-dihydroxy-4-preg- nene-3-one: A teleost maturation-inducing hor- mone. Dev. Growth Differ. 29, 1-12.
Norberg, B., and Haux, C. (1985). Induction, isolation and a characterization of the lipid content of plasma vitellogenin from two Salmo species: Rainbow trout (Salmo gairdnen) and sea trout (Salmo trutta). Comp. Biochem. Physiol. B 81, 869-876.
Norberg, B., and Haux, C. (1988). An homologous radioimmunoassay for brown trout (Salmo trutta) vitellogenin. Fish Physiol. Biochem. 5, 59-68.
Pickering, A. D., and Christie, P. (1981). Changes in the concentrations of plasma cortisol and thyrox- ine during sexual maturation of the hatchery- reared brown trout, Salmo trutta L. Gen. Comp. Endocrinol. 44,487-4%.
Pickering, A. D., Pottinger, T. G., Carragher, J., and Sumpter, J. P. (1987). The effects of acute and chronic stress on the levels of reproductive hor- mones in the plasma of mature male brown trout,

326 NORBERG ET AL.
Salmo trutta L. Gen. Comp. Endocrinol. 68,249 259.
Scott, A. P., Bye, V. J., and Baynes, S. M. (198Oa). Seasonal variations in sex steroids of female rain- bow trout (Salmo gairdneri Richardson). J. Fish Biol. 17, 587-592.
Scott, A. P., Bye, V. J., Baynes, S. M., and Spring- ate, J. R. C. (198Ob). Seasonal variation in plasma concentrations of 1 I-ketotestosterone and testos- terone in male rainbow trout (Salmo gairdneri Richardson). J. Fish Biol. 17, 495-505.
Scott, A. P., Sheldrick, E. L., and Flint, A. P. F. (1982). Measurement of 17a20B-dihydroxy-4- pregnen-3-one in plasma of trout (Salmo gairdneri Richardson): Seasonal changes and response to salmon pituitary extract. Gen. Comp. Endocrinol. 46, -51.
Scott, A. P., and Sumpter, J. P. (1983). A comparison of the female reproductive cycles of autumn- spawning and winter-spawning strains of rainbow trout (Salmo gairdneri Richardson). Gen. Comp. Endocrinol. 52, 79-85.
Scott A. P., Sumpter J. P., and Hardiman P. A. (1983). Hormone changes during ovulation in the rainbow trout (Salmo gairdneri Richardson). Gen. Comp. Endocrinol. 49, 126134.
So, Y. P., Idler, D. R., and Hwang, S. J. (1985). Plasma vitellogenin in landlocked Atlantic salmon (Salmo salar Ouananiche): Isolation, homologous radioimmunoassay and immunological cross- reactivity with vitellogenin from other teleosts. Comp. Biochem. Physiol. B 81, 63-71.
Sower, S. A., and S&reck, C. D. (1982). Steroid and thyroid hormones during sexual maturation of coho salmon (Oncorhynchus kisutch) in seawater or fresh water. Gen. Comp. Endocrinol. 47, 42- 53.
Specker, J. L., and Richman, N. H. (1984). Environ- mental salinity and the thyroidal response to thy- rotropin in juvenile coho salmon (Oncorhynchus kisutch). J. Exp. Zool. 230, 329-333.
Tam, W. H., Roy, R. J. J., and Makaran, R. (1986). Ovarian cycle and plasma concentrations of estro- gen and vitehogenin in brook trout (Salvelinus fontinalis, Mitchill). Canad. J. Zool. 64,744-751,
Ueda, H., Hiroi, O., Hat-a, A., Yamauchi, K., and Nagahama, Y. (1984). Changes in serum concen- trations of steroid hormones, thyroxine, and vi- tellogenin during spawning migration of the chum salmon, Oncorhynchus keta. Gen. Comp. Endo- crinol. 53, 203-211.
Wallace, R. A. (1985). Vitellogenesis and oocyte growth in nonmammalian vertebrates. In “Devel- opmental Biology” (L. W. Browder, Ed.), Vol. 1, pp. 127-177. Plenum, New York.
Wallace, R. A., and Selman, K. (1981). Cehular and dynamic aspects of oocyte growth in teleosts. Amer. Zool. 21, 325-343.
Watts, E. G., Copp, D. H., and Deftos, L. J. (1975). Changes in plasma calcitonin and calcium during the migration of salmon. Endocrinology %, 214- 218.
Wright, R. S., and Hunt, S. M. W. (1982). A radioim- munoassay for 17a20$-dihydroxy-4-pregnen-3- one: Its use in measuring changes in serum levels at ovulation in Atlantic salmon (Salmo salar), coho salmon (Oncorhynchus kisutch), and rain- bow trout (Salmo gairdnert). Gen. Comp. Endo- crinol. 47, 475-482.
Yamane, S., and Yamada, J. (1977). Histological changes of the ultimobranchial gland through the life history of the Masu salmon. Bull. Japan. Sot. Sci. Fish. 43, 375-386.
Yamauchi, K., Kagawa H., Ban M., Kasahara N., and Nagahama Y. (1984). Changes in plasma estradiol-17B and 17a,20@dihydroxydpregnen- 3-one levels during final oocyte maturation of the Masu salmon Oncorhynchus masou. Bull. Japan. Sot. Sci. Fish. 50, 2137.
Young, G., Kagawa, H., and Nagahama, Y. (1982). Oocyte maturation in the amago salmon (Onco- rhynchus rhodurus): In vitro effects of salmon go- nadotropin, steroids and cyanoketone (an inhibi- tor of 3B-hydroxy-AT-steroid dehydrogenase). J. Exp. Zool. 224,265-275.




















