
In vitro control of prolactin (PRL) and growth hormone secretion of neonatal rat pituitary glands:...
11
Life Sciences, Vol. 52, pp. 259-269 Pergamon Press Printed in the USA IN VITRO CONTROL OF PROLACTIN (PRL) AND GROWTH HORMONE SECRETION OF NEONATAL RAT PITUITARY GLANDS: EFFECTS OF OVINE PRL, SALMON CALCITONIN, ENDOTHELIN-3, ANGIOTENSIN I], BROMOCRYPTINE AND SOMATOSTATIN B. Kacs6h*, B. E. T6th, L. M. Avery and C. E. Grosvenor The Pennsylvania State University, Department of Molecular and Cell Biology, University Park, PA 16802, USA (Received in final form November 6, 1992) Summarv Bromocryptine potently decreased prolactin (PRL) secretion of pituitary glands of 2-day-old rats in vitro (up to 85% inhibition; ED5o between 0.1 and 1.0 nM) without altering the bioactivity to immunoreactivity (B/I) ratio. Bromocryptine tended to suppress growth hormone (GH) secretion although the effect did not reach statistical significance. Angiotensin-II (A-H; 1-1000 nM) stimulated PRL secretion in a dose-dependent manner without affecting secretion of GH. The B/I ratio of PRL secreted in response to A-II was increased. Somatostatin (SRIF) had no effect on PRL secretion but inhibited GH secretion in a dose-dependent manner; significant inhibition (50%) was observed at 100 nM. A 6-h exposure to ovine PRL (oPRL) in concentrations equipotent with 1.2-120 ng/ml rat PRL (rPRL) in the Nb2 bioassay had no effect on immunoreactive rPRL secretion. Salmon calcitonin (sCT) and endothelin-3 (ET-3; 0.1-100 nM) failed to inhibit secretion of PRL or GH. PRL secretion was slightly stimulated by sCT with no apparent dose-response relationship. The present findings suggest that neonatal pituitary glands do not display autoregulation of PRL secretion, and sCT and ET-3 (either endogenous or milk-derived) may not function as PRL inhibiting factors in 2-day-old pups. Thus, the receptors of PRL, sCT and ET-3 on lactotropes, or their functional coupling with inhibition of basal PRL secretion, occur at a later stage of development. The specificity of the PRL releasing factor (PRF) activity of A-II at this age is unique for established PRFs and might reflect a physiological function of PRL in osmoregulation. The increased B/I *Corresponding author;, present address: Mercer University School of Medicine, Division of Basic Medical Sciences, Macon, GA 31201 0024-3205/93 $6.00 + .00 Copyright © 1992 Pergamon Press Ltd All rights reserved.
Transcript of In vitro control of prolactin (PRL) and growth hormone secretion of neonatal rat pituitary glands:...

Life Sciences, Vol. 52, pp. 259-269 Pergamon Press Printed in the USA
IN VITRO CONTROL OF PROLACTIN (PRL) AND GROWTH HORMONE SECRETION OF NEONATAL RAT PITUITARY GLANDS: EFFECTS OF OVINE PRL, SALMON
CALCITONIN, ENDOTHELIN-3, ANGIOTENSIN I], BROMOCRYPTINE AND SOMATOSTATIN
B. Kacs6h*, B. E. T6th, L. M. Avery and C. E. Grosvenor
The Pennsylvania State University, Department of Molecular and Cell Biology, University Park, PA 16802, USA
(Received in final form November 6, 1992)
Summarv
Bromocryptine potently decreased prolactin (PRL) secretion of pituitary glands of 2-day-old rats in vitro (up to 85% inhibition; ED5o between 0.1 and 1.0 nM) without altering the bioactivity to immunoreactivity (B/I) ratio. Bromocryptine tended to suppress growth hormone (GH) secretion although the effect did not reach statistical significance. Angiotensin-II (A-H; 1-1000 nM) stimulated PRL secretion in a dose-dependent manner without affecting secretion of GH. The B/I ratio of PRL secreted in response to A-II was increased. Somatostatin (SRIF) had no effect on PRL secretion but inhibited GH secretion in a dose-dependent manner; significant inhibition (50%) was observed at 100 nM. A 6-h exposure to ovine PRL (oPRL) in concentrations equipotent with 1.2-120 ng/ml rat PRL (rPRL) in the Nb2 bioassay had no effect on immunoreactive rPRL secretion. Salmon calcitonin (sCT) and endothelin-3 (ET-3; 0.1-100 nM) failed to inhibit secretion of PRL or GH. PRL secretion was slightly stimulated by sCT with no apparent dose-response relationship. The present findings suggest that neonatal pituitary glands do not display autoregulation of PRL secretion, and sCT and ET-3 (either endogenous or milk-derived) may not function as PRL inhibiting factors in 2-day-old pups. Thus, the receptors of PRL, sCT and ET-3 on lactotropes, or their functional coupling with inhibition of basal PRL secretion, occur at a later stage of development. The specificity of the PRL releasing factor (PRF) activity of A-II at this age is unique for established PRFs and might reflect a physiological function of PRL in osmoregulation. The increased B/I
*Corresponding author;, present address: Mercer University School of Medicine, Division of Basic Medical Sciences, Macon, GA 31201
0024-3205/93 $6.00 + .00 Copyright © 1992 Pergamon Press Ltd All rights reserved.

260 PRL and GH Secretion in Neonatal Rats Vol. 52, No. 3, 1993
ratio of PRL secreted in response to A-H may be due to the release of specific PRL variants, and might be a sign of functional heterogeneity among lactotropes. The differential sensitivity of PRL and GH to the applied secretagogues suggests that the intracellular regulation of PRL and GH are compartmentalized in the mammosomatotrope cell.
Pituitary secretion of prolactin (PRL) and growth hormone (GH) are under the influence of releasing and release inhibiting factors (1,2). Besides dopamine (DA) a catecholamine acting on D2 dopaminergic receptors (3), several peptide/protein PRL inhibiting factors (PIFs) have been described including salmon calcitonin (sCT), endothelin-3 (ET-3) and PRL itself as an autoregulator at the pituitary level (4-8). While some information is available on the development of the inhibitory control of PRL secretion by DA in vitro and in vivo (9,10), the ontogeny of peptidergic PIF activity is unknown. The period of suckling and especially the early postpartum days during ontogeny are unique in several aspects. Proteins and peptides can be absorbed intact from the gut of the neonate (11). Thus, biologically active compounds in milk may serve as a supplementary, exogenous source of regulatory factors in the neonate (12). These milk-borne factors are effectively delivered to the neonate when the hypothalamo-hypophyseal vascular connection is not fully developed (13). Moreover, whereas GH-secreting cells are abundandant in the pituitary gland of the perinatal rat, the development of PRL-secreting cells is delayed and occurs mainly after birth. Original prolactin secretors also secrete GH and are therefore termed mammosomatotropes (14). There are indications that the development of PRL- secreting cells is under the influence of a yet unidentified milk-borne factor(s) (15). PRL is known to have several molecular variants due to posttranscriptional and (mainly) posttranslational modifications. The biological and immunological activities of PRL variants depend on the type of bioassay and antibody, respectively; using the same bioassay and the same antiserum the bioactivity and immunorectivity of PRL variants do not change parallel with each other, therefore the bioactivity-to- immunoreactivity (B/I) ratio is characteristic for each variant and may be substantially different from 1 (16). In the present experiments the potential function of peptide/protein PIFs were characterized in pituitary glands of neonatal rats in vitro on day 2 postpartum. In addition, the PRL releasing factor-like (PRF-like) activity of angiotensin-[[ (A-H) (17,18) was assessed. The effects of the putative PIFs and PRFs were evaluated on PRL-like bioactivity as well as on PRL-like immunoreactivity.
Materials and Methods
Adult virgin female Sprague-Dawley rats, obtained from Harlan Laboratories, Inc. (Indianapolis, IN), were maintained on a 14-h light, 10-h dark schedule in a temperature controlled vivarium (20-22 °C). Animals were bred in the laboratory, housed individually from day 19-20 of gestation, and checked for parturition twice a day. Rat pups were used in the experiments on day 2 postpartum. The pups were decapitated, and pituitary glands were collected in culture medium. For each litter trunk blood of male and female pups, respectively, was pooled, collected in 1.5 ml polypropylene tubes, allowed to clot at room temperature for 1 h then overnight at 4 °C. Serum was collected after microfuging the samples and stored at -20 °C until assayed for PRL.

Vol. 52, No. 3, 1993 PRL and GH Secretion in Neonatal Rats 261
Culture of neonatal oituitarv alands The pituitary glands were rinsed twice with excess culture medium (Dulbecco's
modified Eagle's medium [DMEM] supplemented with 10 mM HEPES, 100 U/ml penicillin, 100 I~g/ml streptomycin, 20 I~g/ml bacitracin, 20 lig/ml L-ascorbic acid and 0.1% RIA-grade BSA). Male and female pituitaries were handled separately and placed individually in the wells of flat bottomed 24-well culture dishes (Corning, New York). To stabilize basal secretion, each gland was incubated in 1 ml medium for 3 h at 37 °C in a humidified 95% air/5% CO2 incubator. After the stabilization period, each well was rinsed twice with 1 ml culture medium, and the glands were further incubated in 1 ml/well medium for 2-10 hours as indicated later. The secretion rate of each individual gland was monitored during two subsequent incubation periods: the first incubation was with culture medium (DMEM) followed by an incubation with a potential PIF or PRF diluted in DMEM. The long (6-h) incubation used for putative PIFs was necessary to prevent PRL-concentrations falling below the detection limit of the assays upon exposure to the test substances. In addition, certain PIFs (e.g. ET-3) act as PIFs only during a prolonged exposure (7), whereas the same PIF may have a transient PRF-like activity during a short exposure in a perifusion system (19). A shorter (2-h) exposure was chosen for A-H (a potential PRF) because basal secretion is detectable with this incubation time. Furthermore, a short incubation is preferable for releasing factors because of potential receptor desensitization/downregulation. To establish a dose-response relationship, the glands were exposed to a concentration range of the factors as indicated later. At the end of each incubation the media were collected and stored at -20 °C until assayed for hormone concentrations.
Drugs/DeDtides Bromocryptine mesylate and ovine PRL (oPRL) were obtained from Sigma (St.
Louis, MO). Bromocryptine was dissolved in absolute ethanol and diluted in DMEM as indicated later. Ovine PRL was dissolved in 10 mM NaHCO3 and diluted in DMEM for pituitary culture or in Fisher's medium for Nb2 assay. The dose-response curves obtained with oPRL and rat PRL (rPRL) B-6 in the bioassay were parallel; thus, the concentration of oPRL is expressed in terms of rPRL standard and reflects equipotent concentrations in the Nb2 lymphoma bioassay. All peptides (A-H, somatostatin [SRIF], sCT, ET-3) were purchased from Peninsula Laboratories, Inc. (Belmont, CA), dissolved in distilled water, aliquoted and lyophilized. The aliquots were dissolved and diluted in DMEM immediately before exposing the pituitaries to the peptides.
Hormone assays Immunoreactive rGH and rPRL was detected in RIA using kits generously
provided by Dr. A.F. Parlow and the National Hormone and Pituitary Program, NIDDK, as described earlier (20,21). Hormone levels are expressed in terms of the RP-2 and RP-3 reference preparation for GH and PRL, respectively. PRL-like bioactivity was measured by the Nb2 lymphoma assay using a method reported by Tanaka et al. (22) and Russell et al. (23) with modifications, as described elsewhere (21). The assay relies on PRL-receptor-mediated cell proliferation monitored by the uptake of [3H]thymidine (ICN, Costa Mesa, CA). Due to a mutant PRL receptor (24) that displays an extremely high affinity to PRL (25), this bioassay is even more sensitive than RIA and is the only bioassay suitable to detect PRL in the low concentrations that occur in neonatal rats. Since the rPRL RP-3 contains thimerosal as a preservative, it cannot be used directly in the bioassay. Therefore, biological grade (B-6) rPRL was used as a standard. The B-6 standard gives a displacement curve parallel with that of RP-3 in

262 PRL and GH Secretion in Neonatal Rats Vol. 52, No. 3, 1993
RIA; the bioassay values were converted to the RP-3 standard. Dilution series of serum and culture medium samples gave parallel dose-response curves with the standard hormone preparations in the RIAs and Nb2 assay in the dilution range used in the study. All samples from each experiment were assayed in the same RIA and Nb2 assay to avoid interassay variability.
Statistical analvsis Statistical analysis was performed using the SuperANOVA and Statview
programs (Abacus Concepts, Inc., Berkeley, CA) using a Macintosh Plus personal computer (Apple Computer, Inc., Cupertino, CA). The effect of the releasing or release inhibiting factors was calculated both by direct comparison of hormone concentrations in the samples and by comparison of the change in secretion rate in response to the treatment (ratio of the second and first incubations). The effect of the drugs on bioactive and immunoreactive levels of PRL were also compared (bioactivity to immunoreactivity [B/I] ratios). The values were compared by one-way ANOVA followed by Bonferroni-Dunn's and Student-Newman-Keuls multiple range tests comparing each value with the DMEM control. The effects were considered statistically significant if p < 0.05.
Results
To elucidate whether the secretion rate of the hormones was a linear function of time during the experiment, secretion rates of immunoreactive (ir) PRL and GH were calculated from a 3-h and a subsequent 10-h incubation period. Neonatal pituitary glands secreted 0.45 + 0.08 ng/h PRL and 15.7 + 2.9 ng/h GH during the first 3-h incubation, whereas 0.32 + 0.05 ng/h PRL and 16.3 + 4.6 ng/h GH (n=8) during the subsequent 10-h incubation. Thus, the secretion rate of irPRL displayed approximately a 30% decrease while that of GH was reasonably steady during the first 13 hours after the stabilization period. The bioactivity to immunoreactivity (B/I) ratio of PRL was not significantly affected by the length of incubation (3h: 1.619 + 0.886 vs. 10h: 2.294 + 0.643).
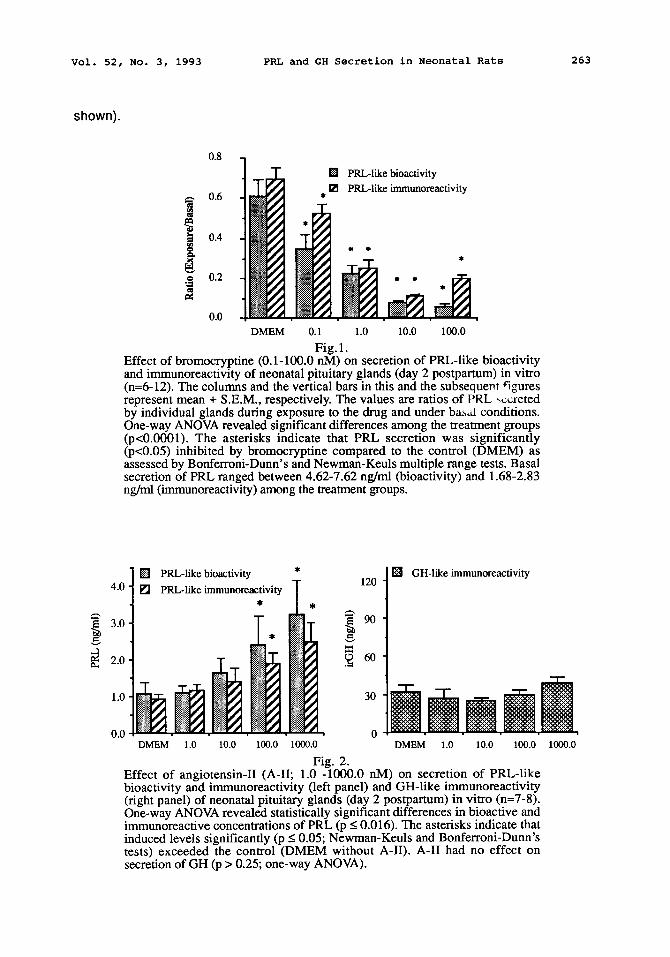
In order to evaluate if the test system was suitable to detect secretory responses, the effect of the well-established D2 receptor agonist bromocryptine mesylate was first evaluated. PRL secretion was highly sensitive to the inhibitory action of bromocryptine: ED5o was between 0.1 and 1.0 nM and the maximal inhibition was approximately 85%. The effect of bromocryptine on PRL-like bioactivity and immunoreactivity was similar (Fig. 1). Thus, the B/I ratio of PRL secreted by the neonatal pituitary glands was not altered. Bromocryptine had no significant effect on the secretion of irGH (not shown).
A-H stimulated PRL secretion in a dose-dependent manner without affecting GH secretion (Fig. 2). Its PRL-releasing effect was higher for bioactivity than for immunoreactivity (p_<0.01, two-way ANOVA) with no apparent dose-response relationship (Fig. 3); thus, the B/I ratio of PRL increased in response to A-~.
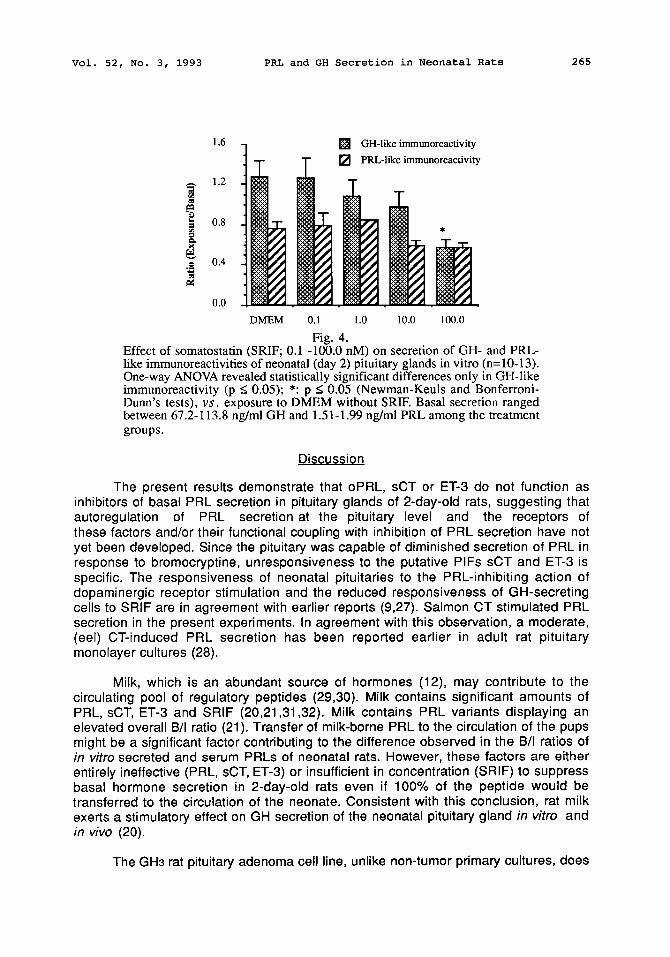
SRIF which is a physiological release inhibiting hormone for GH (26) significantly inhibited secretion of irGH; however, its effect on irPRL secretion did not reach statistical significance (Fig. 4). Bioactive levels of PRL were also unaffected (not
Vol. 52, No. 3, 1993 PRL and GH Secretion in Neonatal Rats 263
shown).
0.8
0.6
0.4
o 0.2
0.0
[ ] PRL-like bioactivity
• [ ] PRL-like immunoreactivity
DMEM 0.1 1.0 10.0 100.0
Fig. 1. Effect of bromocryptine (0.1-100.0 nM) on secretion of PRL-like bioactivity and immunoreactivity of neonatal pituitary glands (day 2 postpartum) in vitro (n=6-12). The columns and the vertical bars in this and the subsequent figures represent mean + S.E.M., respectively. The values are ratios of PRL ,~:reted by individual glands during exposure to the drug and under ba~al conditions. One-way ANOVA revealed significant differences among the treatment groups (p<0.0001). The asterisks indicate that PRL secretion was significantly (p<0.05) inhibited by bromocryptine compared to the control (DMEM) as assessed by Bonferroni-Dunn's and Newman-Keuls multiple range tests. Basal secretion of PRL ranged between 4.62-7.62 ng/ml (bioactivity) and 1.68-2.83 ng/ml (immunoreactivity) among the treatment groups.
4.0
3.0
2.0
1.0
0.0
[] PRL-like bioactivity [] PRL-like immunoreactivity
DMEM 1.0 10.0 100.0
120
90
30
0 1000.0
Fig. 2.
[ ] GH-like immunoreactivity
DMEM 1.0 10.0 100.0 1000.0
Effect of angiotensin-II (A-II; 1.0 -1000.0 nM) on secretion of PRL-like bioactivity and immunoreactivity (left panel) and GH-like immunoreactivity (right panel) of neonatal pituitary glands (day 2 postpartum) in vitro (n=7-8). One-way ANOVA revealed statistically significant differences in bioactive and immunoreactive concentrations of PRL (p < 0.016). The asterisks indicate that induced levels significantly (p < 0.05; Newman-Keuls and Bonferroni-Dunn's tests) exceeded the control (DMEM without A-II). A-II had no effect on secretion of GH (p > 0.25; one-way ANOVA).
264 PRL and GH Secretion in Neonatal Rats Vol. 52, No. 3, 1993
16
12
~ 8
..~ 4
[ ] PRL-Iike bioactivity
[ ] PRL-like immunoreactivity
DMEM 1.0 10.0 100.0 1000.0
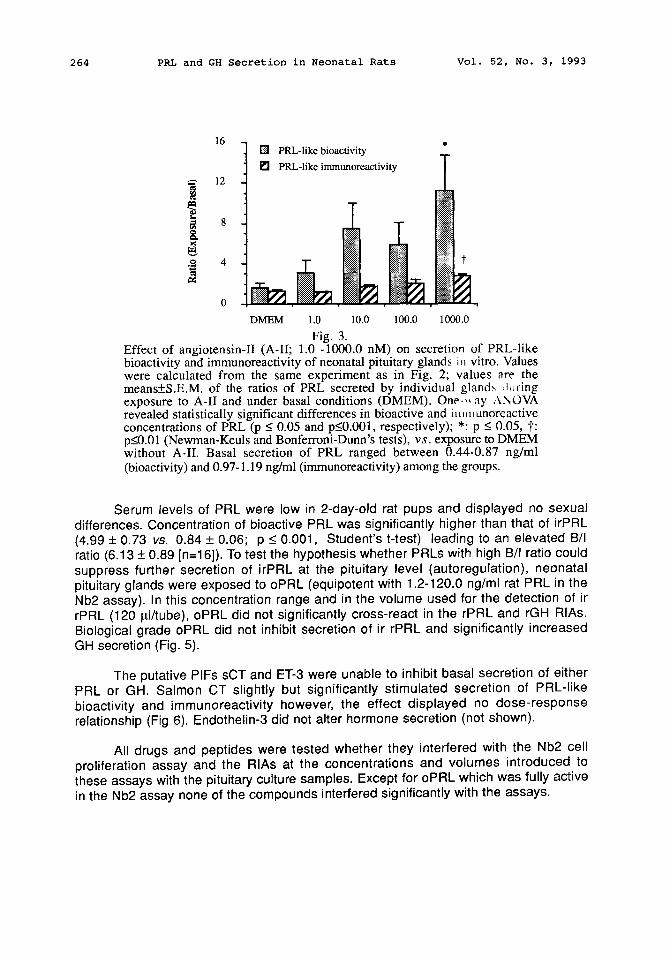
Fig. 3. Effect of angiotensin-II (A-II; 1.0 -1000.0 nM) on secretion of PRL-like bioactivity and immunoreactivity of neonatal pituitary glands iu vitro. Values were calculated from the same experiment as in Fig. 2; values are the means+S.E.M, of the ratios of PRL secreted by individual gland~ Juring exposure to A-II and under basal conditions (DMEM). One ,, ay ,,\NOVA revealed statistically significant differences in bioactive and im, mnoreactive concentrations of PRL (p _< 0.05 and p_<0.001, respectively); *: p -< 0.05, t: p_<0.01 (Newman-Keuls and Bonferroni-Dunn's tests), v s . exposure to DMEM without A-II. Basal secretion of PRL ranged between 0.44-0.87 ng/ml (bioactivity) and 0.97-1.19 ng/ml (immunoreactivity) among the groups.
Serum levels of PRL were low in 2-day-old rat pups and displayed no sexual differences. Concentration of bioactive PRL was significantly higher than that of irPRL (4.99 + 0.73 vs. 0.84 + 0.06; p < 0.001, Student's t-test) leading to an elevated B/I ratio (6.13 + 0.89 [n=16]). To test the hypothesis whether PRLs with high B/I ratio could suppress further secretion of irPRL at the pituitary level (autoregulation), neonatal pituitary glands were exposed to oPRL (equipotent with 1.2-120.0 ng/ml rat PRL in the Nb2 assay). In this concentration range and in the volume used for the detection of ir rPRL (120 pJ/tube), oPRL did not significantly cross-react in the rPRL and rGH RIAs. Biological grade oPRL did not inhibit secretion of ir rPRL and significantly increased GH secretion (Fig. 5).
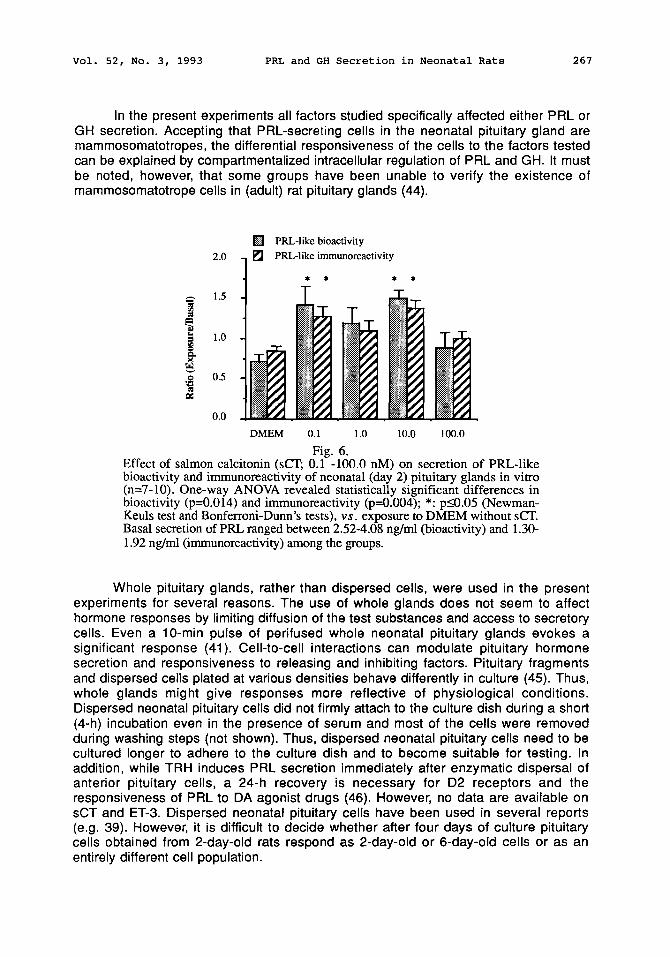
The putative PIFs sCT and ET-3 were unable to inhibit basal secretion of either PRL or GH. Salmon CT slightly but significantly stimulated secretion of PRL-like bioactivity and immunoreactivity however, the effect displayed no dose-response relationship (Fig 6). Endothelin-3 did not alter hormone secretion (not shown).
All drugs and peptides were tested whether they interfered with the Nb2 cell proliferation assay and the RIAs at the concentrations and volumes introduced to these assays with the pituitary culture samples. Except for oPRL which was fully active in the Nb2 assay none of the compounds interfered significantly with the assays.
Vol. 52, No. 3, 1993 PRL and GH Secretion in Neonatal Rats 265
1.6
1.2
0.8
0.4
0.0
o
DMEM 0.1
[ ] GH-like immunoreactivity
[ ] PRL-like immunoreactivity
1.0 10.0 100.0
Fig. 4. Effect of somatostatin (SRIF; 0.1 -100.0 nM) on secretion of GH- and PRL- like immunoreactivities of neonatal (day 2) pituitary glands in vitro (n=10-13). One-way ANOVA revealed statistically significant differences only in GH-like immunoreactivity (p < 0.05); *: p < 0.05 (Newman-Keuls and Bonferroni- Dunn's tests), v s . exposure to DMEM without SRIF. Basal secretion ranged between 67.2-113.8 ng/ml GH and 1.51-1.99 ng/ml PRL among the treatment groups.
Discussion
The present results demonstrate that oPRL, sCT or ET-3 do not function as inhibitors of basal PRL secretion in pituitary glands of 2-day-old rats, suggesting that autoregulation of PRL secretion at the pituitary level and the receptors of these factors and/or their functional coupling with inhibition of PRL secretion have not yet been developed. Since the pituitary was capable of diminished secretion of PRL in response to bromocryptine, unresponsiveness to the putative PIFs sCT and ET-3 is specific. The responsiveness of neonatal pituitaries to the PRL-inhibiting action of dopaminergic receptor stimulation and the reduced responsiveness of GH-secreting cells to SRIF are in agreement with earlier reports (9,27). Salmon CT stimulated PRL secretion in the present experiments. In agreement with this observation, a moderate, (eel) CT-induced PRL secretion has been reported earlier in adult rat pituitary monolayer cultures (28).
Milk, which is an abundant source of hormones (12), may contribute to the circulating pool of regulatory peptides (29,30). Milk contains significant amounts of PRL, sCT, ET-3 and SRIF (20,21,31,32). Milk contains PRL variants displaying an elevated overall B/I ratio (21). Transfer of milk-borne PRL to the circulation of the pups might be a significant factor contributing to the difference observed in the B/I ratios of in vitro secreted and serum PRLs of neonatal rats. However, these factors are either entirely ineffective (PRL, sCT, ET-3) or insufficient in concentration (SRIF) to suppress basal hormone secretion in 2-day-old rats even if 100% of the peptide would be transferred to the circulation of the neonate. Consistent with this conclusion, rat milk exerts a stimulatory effect on GH secretion of the neonatal pituitary gland in vitro and in vivo (20).
The GH3 rat pituitary adenoma cell line, unlike non-tumor primary cultures, does

266 PRL and GH Secretion in Neonatal Rats Vol. 52, No. 3, 1993
not display autoregulation by PRL (33). Considering that tumor cells are dedifferentiated and share several characteristics with embryonic cells, and that pituitary lactotropes differentiate during the perinatal period and postpartum (14), the lack of autoregulation of PRL secretion in the neonatal pituitary gland is not surprising. In addition, while PRL receptors are expressed in the adult pituitary gland (34), they appear late during ontogeny in the rat (35). The reason for the oPRL-induced GH secretion is not clear but is most likely due to a contamination in the biological grade oPRL preparation.
l l~ PRL-like immunoreactivity 1.6 [] GH-like immunoreactivity t
~ 1.2
0.4
0.0 DMEM 1.2 12.0 120.0
Fig. 5. Effect of biological grade ovine PRL (oPRL; 1.2 -120.0 ng/ml) on secretion of PRL- and GH-like immunoreactivities of neonatal (day 2) pituitary glands in vitro (n= 10-12). One-way ANOVA revealed statistically significant differences only in GH-like immunoreactivity (p < 0.025); ?: p<0.05 (Newman-Keuls test), *: p < 0.05 (Bonferroni-Dunn's test), v s . exposure to DMEM without oPRL. Basal secretion ranged between 1.84-2.30 ng/ml PRL and 50.4-72.0 ng/ml GH among the treatment groups.
Stimulation of PRL secretion by A-I] resulted in a more pronounced elevation of bioactive than immunoreactive PRL. Secretion of PRL with an elevated B/I ratio may reflect the release of specific PRL variants in response to a PRF(s). Release of PRL variants might be due to a change in the secretory process of stored vs. newly synthesized PRL (3,36,37) and/or to a subpopulation of (somato)mammotropes displaying a differential responsiveness to a particular PRF as reported for thyrotropin releasing hormone (TRH) (38). The pituitary gland of the neonatal rat displays numerous unique features. GH secretion of neonatal pituitaries is stimulated by classic PRFs such as TRH (1,39), oxytocin (40,41) or vasoactive intestinal polypetide (unpublished observation). In contrast with hemipituitaries of adult female rats (42), GH secretion of neonatal pituitary glands was not stimulated by A-I[. So far A-[I is the only PRF that does not stimulate GH secretion but is an active PRF for the neonatal rat pituitary gland in vitro. This might reflect an in vivo function for A-II in the regulation of PRL secretion in the neonatal rat. For example, since the principal function of A-l] is osmoregulation, A-l]-regulated PRL could serve as an osmoregulator in the perinatal rat. Phylogenetic and ontogenetic correlations would also suggest such a function because PRL is the major osmoregulator in fish (43).
Vol. 52, No. 3, 1993 PRL and GH Secretion in Neonatal Rats 267
In the present experiments all factors studied specifically affected either PRL or GH secretion. Accepting that PRL-secreting cells in the neonatal pituitary gland are mammosomatotropes, the differential responsiveness of the cells to the factors tested can be explained by compartmentalized intracellular regulation of PRL and GH. It must be noted, however, that some groups have been unable to verify the existence of mammosomatotrope cells in (adult) rat pituitary glands (44).
2.0
1.5
- ~ 1.0
,~ 0.5
0.0
[ ] PRL-like bioactivity [ ] PRL-like immunoreactivity
DMEM 0.1 1.0
Fig. 6. 10.0 100.0
Effect of salmon calcitonin (sCT; 0.1 -1000 nM) on secretion of PRL-like bioactivity and immunoreactivity of neonatal (day 2) pituitary glands in vitro (n=7-10). One-way ANOVA revealed statistically significant differences in bioactivity (p=0.014) and immunoreactivity (p--0.004); *: p_<0.05 (Newman- Keuls test and Bonferroni-Dunn's tests), v s . exposure to DMEM without sCT. Basal secretion of PRL ranged between 2.52-4.08 ng/ml (bioactivity) and 1.30- 1.92 ng/ml (immunoreactivity) among the groups.
Whole pituitary glands, rather than dispersed cells, were used in the present experiments for several reasons. The use of whole glands does not seem to affect hormone responses by limiting diffusion of the test substances and access to secretory cells. Even a 10-min pulse of perifused whole neonatal pituitary glands evokes a significant response (41). Cell-to-cell interactions can modulate pituitary hormone secretion and responsiveness to releasing and inhibiting factors. Pituitary fragments and dispersed cells plated at various densities behave differently in culture (45). Thus, whole glands might give responses more reflective of physiological conditions. Dispersed neonatal pituitary cells did not firmly attach to the culture dish during a short (4-h) incubation even in the presence of serum and most of the cells were removed during washing steps (not shown). Thus, dispersed neonatal pituitary cells need to be cultured longer to adhere to the culture dish and to become suitable for testing. In addition, while TRH induces PRL secretion immediately after enzymatic dispersal of anterior pituitary cells, a 24-h recovery is necessary for D2 receptors and the responsiveness of PRL to DA agonist drugs (46). However, no data are available on sCT and ET-3. Dispersed neonatal pituitary cells have been used in several reports (e.g. 39). However, it is difficult to decide whether after four days of culture pituitary cells obtained from 2-day-old rats respond as 2-day-old or 6-day-old cells or as an entirely different cell population.

268 PRL and GH Secretion in Neonatal Rats Vol. 52, No. 3, 1993
It remains to be elucidated whether sCT and ET-3 are capable of inhibiting PRF- (e.g. TRH- or A-H-) stimulated PRL secretion in neonatal rat pituitary glands on day 2 postpartum, and when these factors and (o)PRL become capable of inhibiting basal secretion of PRL during ontogeny.
Acknowledaement
The authors express their thanks to Dr. A. F. Parlow and the National Hormone and Pituitary Program, NIDDK, for providing the rGH and rPRL RIA kits and the biological grade rPRL preparation (B-6).
References
1. J.D. NEILL, Prolactin Secretion and Its Control. In: The Phvsioloav of Reproduction, E. Knobil and J.D. Neill (eds), 1379-1390, Raven-Press, New York (1988).
2. A. ARIMURA and M.D. CULLER, Regulation of Growth Hormone Secretion. In: The Pituitarv Gland. H. Imura (ed), 221-259, Raven Press, New York (1985).
3. N. BEN-JONATHAN, Endocr. Rev. 6 564-589 (1985). 4. G.V. SHAH, R.M. EPAND and R.C. ORLOWSKI, J. Endocrinol. 11 6 279-286
(1988). 5. A.M. JUDD, T. KUBOTA, S.I. KUAN, W.D. JARVIS, B.L. SPANGELO and
R.M. MacLEOD, Endocrinology 127 191-199 (1990). 6. W.K. SAMSON, K.D. SKALA, B.D. ALEXANDER and F.-L.S. HUANG, Biochem.
Biophys. Res. Commun. 169 737-743 (1991). 7. B. KANYICSKA, T.P. BURRIS and M.E. FREEMAN, Biochem. Biophys. Res.
Commun. 174 338-343 (1991 ). 8. L.S. FRAWLEY and C.L. CLARK, Endocrinology 11 ~ 1462-1466 (1986). 9. O. KHORRAM, LoR. DePALATIS and S.M. McCANN, Endocrinology 11,5 1698-
1704 (1984). 10. S.R. OJEDA and S.M. McCANN, Endocrinology 95 1499-1505 (1974). 11. E. TELEMO, B.R. WESTROM, G. EKSTROM and B.W. KARLSSON, Biol.
Neonate 52 141-148 (1987). 12. D.W. WEST, Exp. Clin. Endocrinol. 8:145-156 (1989). 13. K. SZABO and K CSANYI, Cell Tissue Res. 224 563-577 (1982). 14. L.S. FRAWLEY and F.R. BOOCKFOR, Endocr. Rev. 12 337-355 (1991 ). 15. T.E. PORTER, L.E. CHAPMAN, F.M. VAN DOLAH and L.S. FRAWLEY,
Endocrinology 12~ 792-796 (1991 ). 16. Y.N. SINHA, Trends Endocrinol. Metab. 3100-106 (1992). 17. G. AGUILERA, C.L. HYDE and K.J. CAIT, Endocrinology 111 1045-1050 (1982). 18. C. SCHRAMME and C. DENEF, Neuroendocrinology 36 483- (1983). 19. W.K. SAMSON, K.D. SKALA, B. ALEXANDER and F-L.S. HUANG,
Endocrinology 128 1465-1473 (1991 ). 20. B. KACSOH, L.C. TERRY, J.S. MEYERS, W.R. CROWLEY and
C.E. GROSVENOR, Endocrinology 125 1326-1336 (1989). 21. B. KACSOH, B.E. TOTH, L.M. AVERY, Y. YAMAMURO and C.E. GROSVENOR,
Endocrine Regulations 25 98-110 (1991). 22. T. TANAKA, R.P.C. SHIU, P.W. GOUT, C.T. BEER, R.L. NOBLE and H.G.

Vol. 52, No. 3, 1993 PRL and GH Secretion in Neonatal Rats 269
FRIESEN, J. Clin. Endocrinol. Metab. 511058-1063 (1980). 23. D.H. RUSSEL, A.R. BUCKLEY, D.W. MONTGOMERY, N.A. LARSON, P.W. GOUT,
C.T. PUTNAM, C.F. ZUKOSKI and R. KIBLER, J. Immunol. 138276-284 (1987). 24. S. ALl, I. PELLEGRINI and P.A. KELLY, J. Biol Chem 26620110-20117 (1991 ). 25. R.P.C. SHIU, H.P. ELSHOLTZ, T. TANAKA, H.G. FRIESEN, P.W. GOUT, C.T.
BEER and R.L. NOBLE, Endocrinology 113159-165 (1983). 26. Y.S. PATEL and C.B. SRIKANT, Ann. Rev. Physiol. 48551-567 (1986). 27. O. KHORRAM, L.R. DePALATIS and S.M. McCANN, Endocrinology 113720-
728 (1983). 28. Y. IWASAKI, K. CHIHARA, J. IWASAKI, H. ABE and J. FUJITA, Life Sci. 251243-
1248 (1979). 29. P. MOSINGER, H. PLACER, O. KOLDOVSKY, Nature 184:1245-1246 (1959). 30. P.A. GONELLA, P. HAMARTZ nad W.A. WALKER, J. Cell. Physiol. 140138-149
(1989). 31. G.V. SHAH, B. KACSOH, R. SESHADRI, C.E. GROSVENOR and W.R.
CROWLEY, Endocrinology 12~561-67 (1989) 32. H-C. LAM, K. TAKAHASHI, M.A. GHATEI, S.R. BLOOM, FEBS Letters 261184-
186 (1990). 33. T.W.C. HO, J.R. GREENAN and A.M. WALKER, Endocrinology 1241507-1514
(1989). 34. S. CHIU, R.D. KOOS and P.M. WISE, Endocrinology 1301747-1749 (1992). 35. P.A. KELLY, J. DJIANE, M-C. POSTEL-VlNAY and M. EDERY, Endocr. Rev. 1._22
235-251 (1991). 36. M.G. FARQUHAR, Membrane Traffic in Prolactin and Other Secretory Cells. In:
Prolactin. Basic and Clinical Correlates, R.M. MacLeod, U. Scapagnini and M.O. Thorner (eds), 3-16, Springer-Verlag, New York (1985).
37. I. PELLEGRINI, G. GUNZ, F. GRISOLI and P. JAQUET, Endocrinology 1261087- 1095 (1990).
38. J. ARITA, Y. KOJIMA and F. KIMURA, Endocrinology 1303167-3174 (1992). 39. J.B. WELSH, L. CUTTLER and M. SZABO, Endocrinology 1192368-2375
(1986). 40. M. MORI, S. VIGH, A. MIYATA, T. YOSHIHARA, S. OKA and A. ARIMURA,
Endocrinology 125 1009-1013 (1990). 41. Z. ACS, B. KACSOH, Z. VERESS and B.E. TOTH, Life Sci. 4._22 2315-2321
(1988). 42. M.K. STEELE, A. NEGRO-VILAR and S.M. McCANN, Endocrinology 109 893-
899 (1981 ). 43. D. SWENNEN, F. RENTIER-DELRUE, B. AUPERIN, P. PRUNET, G. FLIK, S.E.
WENDELAAR BONGA, M. LION and J.A. MARTIAL, J. Endocrinol. 131219-227 (1991 ).
44. D. WYNICK, M.S. VENETIKOU, R. CRITCHLEY, J.M. BURRIN and S.R. BLOOM, J. Endocrinol. 126261-268 (1990).
45. H. SUGIMOTO, M. SUZUKI, T. TAKEUCHI and K. ISHIKAWA, J. Endocrinol. 1 :~1 237-244 (1991).
46. P.F. SMITH, E.H. LUQUE and J.D. NEILL, Methods in Enzymology 124443-465 (1986).




















