
Inflammation-Induced Enhancement of the Visceromotor Reflex to Urinary Bladder Distention:...
10
O RIGINAL R EPORTS Inflammation-Induced Enhancement of the Visceromotor Reflex to Urinary Bladder Distention: Modulation by Endogenous Opioids and the Effects of Early-in-Life Experience With Bladder Inflammation Jennifer DeBerry,* Timothy J. Ness, † Meredith T. Robbins, † and Alan Randich* *Department of Psychology and † Department of Anesthesiology, University of Alabama at Birmingham, Birmingham, Alabama. Abstract: Abdominal electromyographic (EMG) responses to noxious intensities of urinary bladder distention (UBD) are significantly enhanced 24 hours after zymosan-induced bladder inflammation in adult female rats. This inflammation-induced hypersensitivity is concomitantly inhibited by endogenous opioids because intraperitoneal (i.p.) naloxone administration before testing significantly increases EMG response magnitude to UBD. This inhibitory mechanism is not tonically active because naloxone does not alter EMG response magnitude to UBD in rats without inflammation. At the dose tested, naloxone does not affect bladder compliance in rats with or without inflammation. The effects of i.p. naloxone probably result from blockade of a spinal mechanism because intrathecal naloxone also significantly enhances EMG responses to UBD in rats with inflammation. Rats exposed to bladder inflammation from P90-P92 before reinflammation at P120 show similar hypersensitivity and concomitant opioid inhibition, with response magnitudes being no different from that produced by inflammation at P120 alone. In contrast, rats exposed to bladder inflammation from P14-P16 before reinflammation at P120 show markedly enhanced hypersensitivity and no evidence of concomitant opioid inhibition. These data indicate that bladder inflammation in adult rats induces bladder hypersensitivity that is inhibited by an endogenous opioider- gic mechanism. This mechanism can be disrupted by neonatal bladder inflammation. Perspective: The present study observed that bladder hypersensitivity resulting from acute bladder inflammation is suppressed by an opioid-inhibitory mechanism. Experiencing bladder inflammation during the neonatal period can impair the expression of this opioid inhibitory mechanism in adulthood. This suggests that bladder insults during development may permanently alter visceral sensory systems and may represent 1 cause of painful bladder disorders. © 2007 by the American Pain Society Key words: Bladder, opioids, inflammation, neonatal, visceromotor reflex, pain. M ycotic and bacterial infections of the urinary tract often produce urinary bladder inflamma- tion, but our understanding of the neural sub- strates that are altered by infectious processes and result in bladder pain is limited. Recently, we showed that brief intravesical administration of zymosan in adult rats led to urinary bladder inflammation that became progres- sively more severe in a time-dependent manner. 25 This treatment also led to a significant increase in arterial blood pressure and abdominal muscle electromyo- graphic (EMG) responses to urinary bladder distention (UBD) when rats were tested 24 hours after zymosan administration. 25,26 These zymosan-induced increases in responses were observed primarily at suprathreshold intensities of UBD in the range believed to be noxious, and thus this hypersensitivity to UBD could be represen- tative of a hyperalgesic state. 25,26 In separate studies, we Received March 21, 2007; Revised June 7, 2007; Accepted June 15, 2007. Supported in part by grants R21 DK066027 and R01 DK51419. Address reprint requests to Dr. Jennifer DeBerry, Department of Psychol- ogy, University of Alabama at Birmingham, 901 19th Street South, Room 270, Birmingham, AL 35205. E-mail: [email protected] 1526-5900/$32.00 © 2007 by the American Pain Society doi:10.1016/j.jpain.2007.06.011 The Journal of Pain, Vol 8, No 12 (December), 2007: pp 914-923 Available online at www.sciencedirect.com 914
-
Upload
ua-birmingham -
Category
Documents
-
view
1 -
download
0
Transcript of Inflammation-Induced Enhancement of the Visceromotor Reflex to Urinary Bladder Distention:...

IUa
J*B
Msi
RSAo21©d
The Journal of Pain, Vol 8, No 12 (December), 2007: pp 914-923Available online at www.sciencedirect.com
9
ORIGINAL REPORTS
nflammation-Induced Enhancement of the Visceromotor Reflex torinary Bladder Distention: Modulation by Endogenous Opioidsnd the Effects of Early-in-Life Experience With Bladder Inflammation
ennifer DeBerry,* Timothy J. Ness,† Meredith T. Robbins,† and Alan Randich*Department of Psychology and †Department of Anesthesiology, University of Alabama at Birmingham,irmingham, Alabama.
Abstract: Abdominal electromyographic (EMG) responses to noxious intensities of urinary bladderdistention (UBD) are significantly enhanced 24 hours after zymosan-induced bladder inflammation inadult female rats. This inflammation-induced hypersensitivity is concomitantly inhibited by endogenousopioids because intraperitoneal (i.p.) naloxone administration before testing significantly increases EMGresponse magnitude to UBD. This inhibitory mechanism is not tonically active because naloxone does notalter EMG response magnitude to UBD in rats without inflammation. At the dose tested, naloxone doesnot affect bladder compliance in rats with or without inflammation. The effects of i.p. naloxone probablyresult from blockade of a spinal mechanism because intrathecal naloxone also significantly enhances EMGresponses to UBD in rats with inflammation. Rats exposed to bladder inflammation from P90-P92 beforereinflammation at P120 show similar hypersensitivity and concomitant opioid inhibition, with responsemagnitudes being no different from that produced by inflammation at P120 alone. In contrast, ratsexposed to bladder inflammation from P14-P16 before reinflammation at P120 show markedly enhancedhypersensitivity and no evidence of concomitant opioid inhibition. These data indicate that bladderinflammation in adult rats induces bladder hypersensitivity that is inhibited by an endogenous opioider-gic mechanism. This mechanism can be disrupted by neonatal bladder inflammation.Perspective: The present study observed that bladder hypersensitivity resulting from acute bladderinflammation is suppressed by an opioid-inhibitory mechanism. Experiencing bladder inflammationduring the neonatal period can impair the expression of this opioid inhibitory mechanism in adulthood.This suggests that bladder insults during development may permanently alter visceral sensory systemsand may represent 1 cause of painful bladder disorders.
© 2007 by the American Pain Society
Key words: Bladder, opioids, inflammation, neonatal, visceromotor reflex, pain.itstbg(aria
ycotic and bacterial infections of the urinarytract often produce urinary bladder inflamma-tion, but our understanding of the neural sub-
trates that are altered by infectious processes and resultn bladder pain is limited. Recently, we showed that brief
eceived March 21, 2007; Revised June 7, 2007; Accepted June 15, 2007.upported in part by grants R21 DK066027 and R01 DK51419.ddress reprint requests to Dr. Jennifer DeBerry, Department of Psychol-gy, University of Alabama at Birmingham, 901 19th Street South, Room70, Birmingham, AL 35205. E-mail: [email protected]/$32.002007 by the American Pain Society
toi:10.1016/j.jpain.2007.06.011
14
ntravesical administration of zymosan in adult rats ledo urinary bladder inflammation that became progres-ively more severe in a time-dependent manner.25 Thisreatment also led to a significant increase in arteriallood pressure and abdominal muscle electromyo-raphic (EMG) responses to urinary bladder distentionUBD) when rats were tested 24 hours after zymosandministration.25,26 These zymosan-induced increases inesponses were observed primarily at suprathresholdntensities of UBD in the range believed to be noxious,nd thus this hypersensitivity to UBD could be represen-
ative of a hyperalgesic state.25,26 In separate studies, we
azifhgmm
tpaagtbtafm
pvizcttaeac(wbbmd
M
A
AaTwfthltpta
A
t
gvigvwbmmt
A
td2bttiBw
taifptwaTdfi
U
traot(qbmcmi
C
rtsalb
915ORIGINAL REPORT/DeBerry et al
lso found that the bladder hypersensitivity produced byymosan treatment as an adult was markedly enhancedn rats that were also administered intravesical zymosanor 3 consecutive days as neonates (P14-P16).26 This en-anced bladder response was paralleled by a morelobal bladder hypersensitivity, manifested as increasedicturition frequency26 and decreased thresholds foricturition reflexes during cystometry.8
Other studies of primary hyperalgesia and inflamma-ion involving either the knee joint31 or the hindaw28,39 have shown that descending inhibitory systemsre progressively engaged during the first 24 hours ofcute inflammation and act to suppress somatic hyperal-esia.11,27,32-36 These data suggested the possibility thathe urinary bladder hypersensitivity produced by acuteladder inflammation in the adult rat may be concomi-antly suppressed by inflammation-induced activation ofn inhibitory system37,38 and that the enhancement ef-ect we observed with neonatal exposure to zymosanight be due to impairment of this inhibitory system.21
To investigate these possibilities, and testing the hy-othesis that an opioidergic inhibitory system was in-olved, we examined whether the bladder hypersensitiv-ty produced by a single, acute exposure to intravesicalymosan in adult rats was augmented if the opioid re-eptor antagonist naloxone was present during UBDesting (experiments 1, 4, and 5). We performed cys-ometrograms (CMG) to determine whether naloxoneffected urinary bladder compliance, either in the pres-nce or absence of bladder inflammation (experiment 2),nd we examined whether the opioid system was lo-ated spinally by examining the effects of intrathecali.t.) naloxone (experiment 3). Finally, we examinedhether the effects of naloxone on responses to acuteladder inflammation were influenced by a prior bout ofladder inflammation occurring either in adult rats 1onth before UBD testing (P90-P92; experiment 4) oruring the neonatal period (P14-P16; experiment 5).
aterials and Methods: General
nimals and Animal CareAdult female Sprague-Dawley rats (Harlan, Prattville,L) were maintained in separate cages and were eitherge P90 or P120 at the time of their initial treatments.imed-pregnant female Sprague-Dawley rats (Harlan)ere also maintained in separate cages. After birth, only
emale pups were maintained and were age P14 at theime of their initial treatments. The rat pups remainedoused with their mothers until weaning at �P21. The
ight-dark cycle for all rats was 6:00 AM to 6:00 PM. At theime of adult testing, no attempt was made to control forhase of estrous cycle.16 All studies were approved byhe University of Alabama at Birmingham Animal Carend Use Committee.
dult Zymosan Pretreatment ProcedureTwenty-four hours before adult testing, all rats with
he exception of groups AN and AV in experiment 1 and u
roup Z-N in experiment 4 were pretreated with intra-esical zymosan. Rats were anesthetized with inhaledsoflurane (5% induction, 2% maintenance), and a 22-auge angiocatheter was placed into the urinary bladderia the urethra. Intravesical zymosan (1% solution in sterileater; 0.5 mL) was administered and left to dwell in theladder for 30 minutes and drained before catheter re-oval. Each animal was administered ampicillin (100g/mL; 0.2 mL; subcutaneous [s.c.]) and awakened. These
reatments typically occurred beginning at �7:30 AM.
dult Surgical PreparationOn the day of adult testing with either UBD or CMG,
he trachea was cannulated for artificial respiration un-er deep isoflurane anesthesia (4% in oxygen) and a2-gauge angiocatheter was placed into the urinaryladder via the urethra and held in place by a tight su-ure around the distal urethral orifice. The animal washen moved to a recording area and maintained on 4%soflurane until surgical procedures were completed.ody temperature was maintained throughout testingith a heating pad.The timing of the UBD or CMG test session began when
he animal was placed in the recording area. The leftbdominal skin was incised and silver wires were placednto the left external oblique musculature of each animalor differential amplification of EMG activity. After thisrocedure the anesthesia was reduced to 1% and main-ained at this level for the duration of testing. All dataere saved on a computer using Spike-2 software andssociated hardware (Micro 1401; CED, Cambridge, UK).he EMG response was defined as (rectified EMG activityuring UBD–rectified baseline EMG before UBD)/recti-ed baseline EMG.
rinary Bladder DistentionThe UBD trials and the timing of UBD trials were iden-
ical across all experiments. Initially, three 20-second-du-ation, 60 mm Hg distentions of the urinary bladder weredministered (data not presented) to overcome a periodf bladder sensitization that occurs before demonstra-ion of vigorous and reliable visceromotor reflexesVMRs).3 These distentions were followed by a fixed se-uence of 20-second-duration graded distentions of theladder at pressures of 10, 20, 30, 40, 50, 60, 70, and 80m Hg, respectively (3-minute intertrial interval). UBD
onsisted of distending the bladder via air pressure andonitoring intravesical pressure via an in-line, pneumat-
cally linked, low-volume pressure transducer.
ystometrogramThe timing of the CMG session was identical for each
at that underwent CMG testing. The bladder was emp-ied using abdominal palpation before the onset of thealine infusion, which always occurred exactly 10 minutesfter a naloxone or vehicle injection was given (see be-ow). Room-temperature saline was infused into theladder at a slow, steady rate (0.05 mL/min) for 40 min-
tes through polyethylene tubing threaded through a
Tttd
S
E
T
bacgzeattdtwttp
E
C
cofltttmstwsaa
E
I
tcapao(mw(l
usfsUccae
E
IP
aamemibPp(gtpaAw(tpeal(pbdgtcwb
E
IP
atmsTtw
916 Inflammation-Induced Enhancement of the Visceromotor Reflex to UBD
-port extension and into the transurethral catheter. In-ravesical pressure was measured through a sidearm ofhe T-port connected to a low-volume pressure trans-ucer.
pecific Methods
xperiment 1
wenty-Four-Hour Adult PretreatmentThis experiment was performed to determine if theladder hypersensitivity observed in the normal adult ratfter acute exposure to intravesical zymosan was beingoncomitantly suppressed by opioid inhibition. Tworoups of rats at age P120 were administered the adultymosan pretreatment procedure described in the Gen-ral Methods, and 2 additional groups received the samenesthesia protocol but in the absence of an intravesicalreatment. Twenty-four hours later, all rats underwenthe adult surgical preparation and UBD testing proce-ure described in the General Methods. At the time ofesting, half of the rats in each condition were injectedith either naloxone hydrochloride (1 mg/kg, intraperi-
oneal [i.p.]) or sterile water (1 mg/kg, i.p.; vehicle con-rol). Ten minutes after the injection, the UBD testingrocedure was initiated.
xperiment 2
MG Testing of Passive ComplianceCMGs were performed to determine whether passive
ompliance of the urinary bladder was affected by nal-xone administration in the presence or absence of in-ammation. Two groups of rats at age P120 underwenthe adult zymosan pretreatment procedure described inhe General Methods. Twenty-four hours later, the pre-reated groups and an additional 2 groups of age-atched, untreated rats underwent CMG testing as de-
cribed in the General Methods. Before the onset of CMGesting, half of the rats in each condition were injectedith either naloxone hydrochloride (1 mg/kg, i.p.) or
terile water (1 mg/kg, i.p.; vehicle control). Ten minutesfter the injection, the CMG testing procedure was initi-ted.
xperiment 3
ntrathecal NaloxoneThis experiment was performed to determine whether
he effect of i.p. naloxone observed in experiment 1ould be due to a spinal locus of action. Two groups ofdult rats at age P120 underwent the adult zymosanretreatment procedure and 24 hours later received thedult surgical procedure described in the General Meth-ds. At the time of testing, a 7.8-cm intrathecal catheterPE10) was inserted through a slit in the atlantooccipitalembrane and threaded down the spinal column. Ratsere administered either naloxone hydrochloride
1 �g/�L; 10 �g; intrathecal [i.t.]) or vehicle (10 �L) fol-
owed by a 7 �L flush with vehicle. Two vehicles were ased in separate groups of rats; physiological saline orterile water. There were no significant differences in theunctions generated with the 2 vehicles and so the dataets were combined. Ten minutes after the injection, theBD testing procedure was initiated. The intrathecalatheters were targeted to the L5-S2 region of the spinalord and catheter placement was sampled and verified insubset of rats (N � 4) with Evans blue injections at thend of the testing procedure.
xperiment 4
nflammation at P90-P92 and 24-Hour AdultretreatmentThis experiment was performed to determine whetherhistory of exposure to bladder inflammation during
dulthood one month before testing would affect theagnitude of the opioid inhibitory effect observed in
xperiment 1. Specifically, the adult zymosan pretreat-ent procedure and UBD testing procedure were admin-
stered to adult rats (P120) with or without exposure toladder inflammation for 3 consecutive days at age P90-92. Groups of rats were administered the adult zymosanretreatment procedure from P90-P92. One month laterP120), 2 of the pretreated groups and an additional 2roups of age-matched, untreated rats were anesthe-ized and once again administered the adult zymosanretreatment procedure followed 24 hours later by thedult surgical preparation and UBD testing procedure.t the time of testing, half of the rats in each conditionere injected with either naloxone hydrochloride
1 mg/kg, i.p.) or sterile water (1 mg/kg, i.p.; vehicle con-rol). Ten minutes after the injection, the UBD testingrocedure was initiated. Thus, the zymosan conditions ofxperiment 1 were replicated exactly, and the effects ofdult (P90-P92) intravesical zymosan administration fol-owed by re-exposure to zymosan 24 hours before UBDP120) were also examined. A remaining group that wasretreated from P90-P92 was left untreated 24 hoursefore UBD testing and was administered naloxone hy-rochloride (1 mg/kg, i.p.) on the day of testing. Thisroup was included to determine whether opioid inhibi-ion could be permanently engaged after the intravesi-al zymosan treatments from P90-P92 or if this inhibitionas evident only after re-exposure to zymosan 24 hoursefore UBD testing.
xperiment 5
nflammation at P14-P16 and 24-Hour AdultretreatmentThis experiment was performed to determine whetherhistory of exposure to bladder inflammation during
he neonatal period of development would affect theagnitude of the opioid inhibitory effect produced by a
ubsequent acute exposure to bladder inflammation.he adult zymosan pretreatment procedure and UBDesting procedure were administered to adult rats (P120)ith or without prior exposure to bladder inflammation
s neonates. For 3 consecutive days from age P14-P16, 4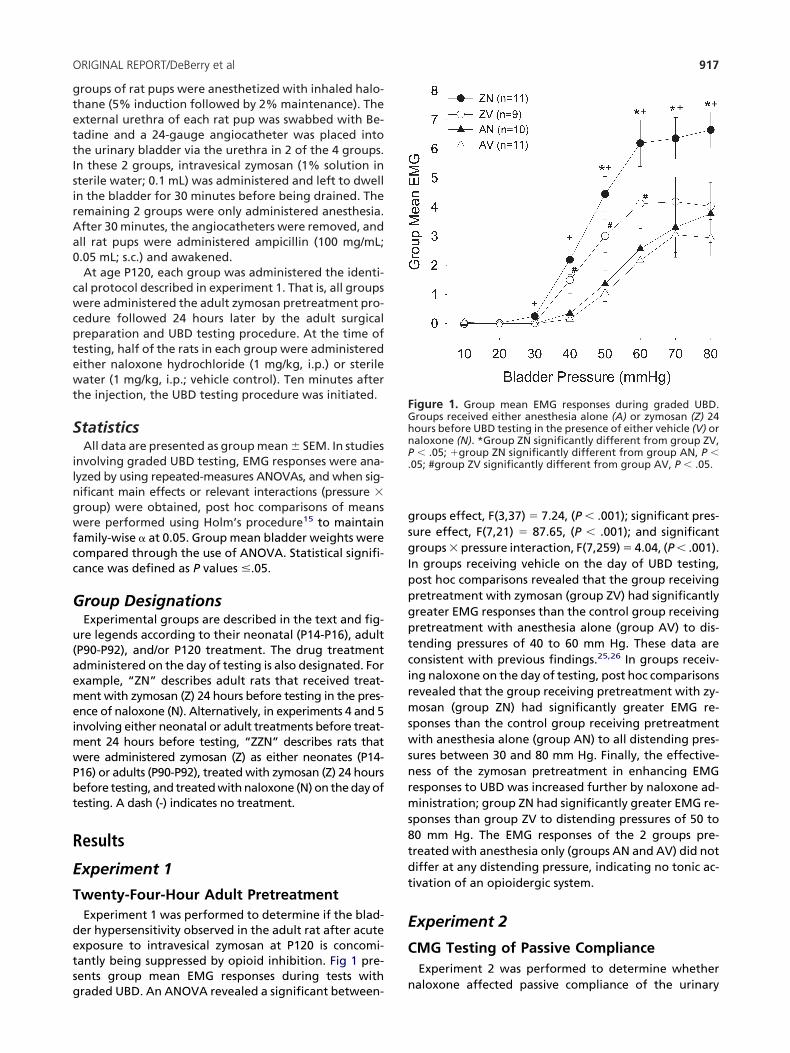
gtettIsirAa0
cwcptewt
S
ilngwfcc
G
u(aemeimwPbt
R
E
T
detsg
gsgIppgptcirmswsnrms8tdt
E
C
FGhnP.
917ORIGINAL REPORT/DeBerry et al
roups of rat pups were anesthetized with inhaled halo-hane (5% induction followed by 2% maintenance). Thexternal urethra of each rat pup was swabbed with Be-adine and a 24-gauge angiocatheter was placed intohe urinary bladder via the urethra in 2 of the 4 groups.n these 2 groups, intravesical zymosan (1% solution interile water; 0.1 mL) was administered and left to dwelln the bladder for 30 minutes before being drained. Theemaining 2 groups were only administered anesthesia.fter 30 minutes, the angiocatheters were removed, andll rat pups were administered ampicillin (100 mg/mL;.05 mL; s.c.) and awakened.At age P120, each group was administered the identi-
al protocol described in experiment 1. That is, all groupsere administered the adult zymosan pretreatment pro-
edure followed 24 hours later by the adult surgicalreparation and UBD testing procedure. At the time ofesting, half of the rats in each group were administeredither naloxone hydrochloride (1 mg/kg, i.p.) or sterileater (1 mg/kg, i.p.; vehicle control). Ten minutes after
he injection, the UBD testing procedure was initiated.
tatisticsAll data are presented as group mean � SEM. In studies
nvolving graded UBD testing, EMG responses were ana-yzed by using repeated-measures ANOVAs, and when sig-ificant main effects or relevant interactions (pressure �roup) were obtained, post hoc comparisons of meansere performed using Holm’s procedure15 to maintain
amily-wise � at 0.05. Group mean bladder weights wereompared through the use of ANOVA. Statistical signifi-ance was defined as P values �.05.
roup DesignationsExperimental groups are described in the text and fig-re legends according to their neonatal (P14-P16), adultP90-P92), and/or P120 treatment. The drug treatmentdministered on the day of testing is also designated. Forxample, “ZN” describes adult rats that received treat-ent with zymosan (Z) 24 hours before testing in the pres-
nce of naloxone (N). Alternatively, in experiments 4 and 5nvolving either neonatal or adult treatments before treat-
ent 24 hours before testing, “ZZN” describes rats thatere administered zymosan (Z) as either neonates (P14-16) or adults (P90-P92), treated with zymosan (Z) 24 hoursefore testing, and treated with naloxone (N) on the day ofesting. A dash (-) indicates no treatment.
esults
xperiment 1
wenty-Four-Hour Adult PretreatmentExperiment 1 was performed to determine if the blad-er hypersensitivity observed in the adult rat after acutexposure to intravesical zymosan at P120 is concomi-antly being suppressed by opioid inhibition. Fig 1 pre-ents group mean EMG responses during tests with
raded UBD. An ANOVA revealed a significant between- nroups effect, F(3,37) � 7.24, (P � .001); significant pres-ure effect, F(7,21) � 87.65, (P � .001); and significantroups � pressure interaction, F(7,259) � 4.04, (P � .001).
n groups receiving vehicle on the day of UBD testing,ost hoc comparisons revealed that the group receivingretreatment with zymosan (group ZV) had significantlyreater EMG responses than the control group receivingretreatment with anesthesia alone (group AV) to dis-ending pressures of 40 to 60 mm Hg. These data areonsistent with previous findings.25,26 In groups receiv-ng naloxone on the day of testing, post hoc comparisonsevealed that the group receiving pretreatment with zy-osan (group ZN) had significantly greater EMG re-
ponses than the control group receiving pretreatmentith anesthesia alone (group AN) to all distending pres-
ures between 30 and 80 mm Hg. Finally, the effective-ess of the zymosan pretreatment in enhancing EMGesponses to UBD was increased further by naloxone ad-inistration; group ZN had significantly greater EMG re-
ponses than group ZV to distending pressures of 50 to0 mm Hg. The EMG responses of the 2 groups pre-reated with anesthesia only (groups AN and AV) did notiffer at any distending pressure, indicating no tonic ac-ivation of an opioidergic system.
xperiment 2
MG Testing of Passive ComplianceExperiment 2 was performed to determine whether
igure 1. Group mean EMG responses during graded UBD.roups received either anesthesia alone (A) or zymosan (Z) 24ours before UBD testing in the presence of either vehicle (V) oraloxone (N). *Group ZN significantly different from group ZV,� .05; �group ZN significantly different from group AN, P �
05; #group ZV significantly different from group AV, P � .05.
aloxone affected passive compliance of the urinary
btocnTFttaaboc
E
I
ittootmr13tsZdc
iro
E
IP
hitponaa3rtScpdgPa
FC(tv
FGpo.natmto
918 Inflammation-Induced Enhancement of the Visceromotor Reflex to UBD
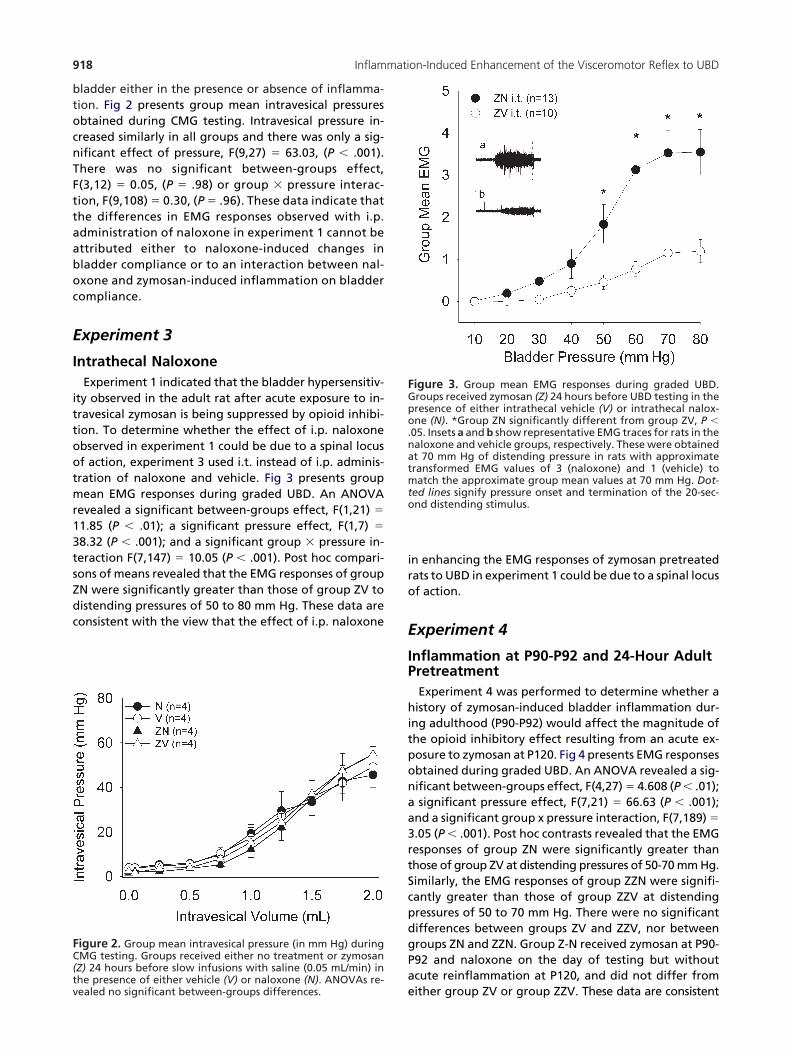
ladder either in the presence or absence of inflamma-ion. Fig 2 presents group mean intravesical pressuresbtained during CMG testing. Intravesical pressure in-reased similarly in all groups and there was only a sig-ificant effect of pressure, F(9,27) � 63.03, (P � .001).here was no significant between-groups effect,(3,12) � 0.05, (P � .98) or group � pressure interac-ion, F(9,108) � 0.30, (P � .96). These data indicate thathe differences in EMG responses observed with i.p.dministration of naloxone in experiment 1 cannot bettributed either to naloxone-induced changes inladder compliance or to an interaction between nal-xone and zymosan-induced inflammation on bladderompliance.
xperiment 3
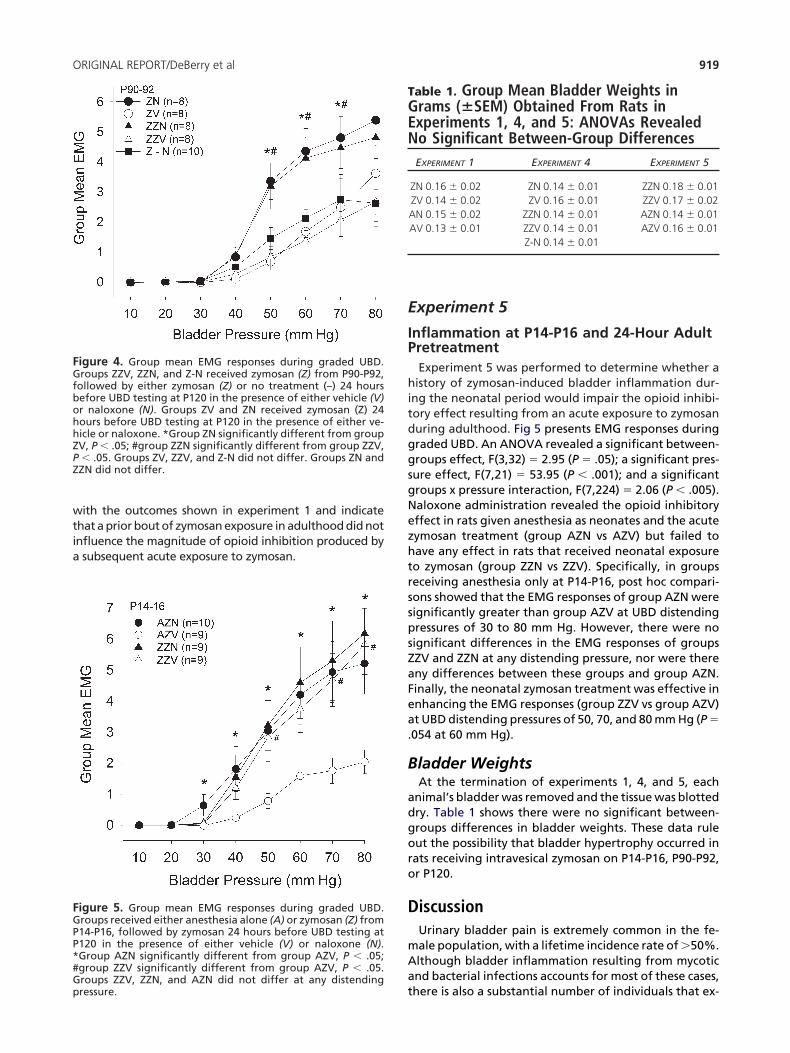
ntrathecal NaloxoneExperiment 1 indicated that the bladder hypersensitiv-
ty observed in the adult rat after acute exposure to in-ravesical zymosan is being suppressed by opioid inhibi-ion. To determine whether the effect of i.p. naloxonebserved in experiment 1 could be due to a spinal locusf action, experiment 3 used i.t. instead of i.p. adminis-ration of naloxone and vehicle. Fig 3 presents groupean EMG responses during graded UBD. An ANOVA
evealed a significant between-groups effect, F(1,21) �
1.85 (P � .01); a significant pressure effect, F(1,7) �
8.32 (P � .001); and a significant group � pressure in-eraction F(7,147) � 10.05 (P � .001). Post hoc compari-ons of means revealed that the EMG responses of groupN were significantly greater than those of group ZV toistending pressures of 50 to 80 mm Hg. These data areonsistent with the view that the effect of i.p. naloxone
igure 2. Group mean intravesical pressure (in mm Hg) duringMG testing. Groups received either no treatment or zymosanZ) 24 hours before slow infusions with saline (0.05 mL/min) inhe presence of either vehicle (V) or naloxone (N). ANOVAs re-
eealed no significant between-groups differences.
n enhancing the EMG responses of zymosan pretreatedats to UBD in experiment 1 could be due to a spinal locusf action.
xperiment 4
nflammation at P90-P92 and 24-Hour AdultretreatmentExperiment 4 was performed to determine whether aistory of zymosan-induced bladder inflammation dur-
ng adulthood (P90-P92) would affect the magnitude ofhe opioid inhibitory effect resulting from an acute ex-osure to zymosan at P120. Fig 4 presents EMG responsesbtained during graded UBD. An ANOVA revealed a sig-ificant between-groups effect, F(4,27) � 4.608 (P � .01);significant pressure effect, F(7,21) � 66.63 (P � .001);
nd a significant group x pressure interaction, F(7,189) �.05 (P � .001). Post hoc contrasts revealed that the EMGesponses of group ZN were significantly greater thanhose of group ZV at distending pressures of 50-70 mm Hg.imilarly, the EMG responses of group ZZN were signifi-antly greater than those of group ZZV at distendingressures of 50 to 70 mm Hg. There were no significantifferences between groups ZV and ZZV, nor betweenroups ZN and ZZN. Group Z-N received zymosan at P90-92 and naloxone on the day of testing but withoutcute reinflammation at P120, and did not differ from
igure 3. Group mean EMG responses during graded UBD.roups received zymosan (Z) 24 hours before UBD testing in theresence of either intrathecal vehicle (V) or intrathecal nalox-ne (N). *Group ZN significantly different from group ZV, P �
05. Insets a and b show representative EMG traces for rats in thealoxone and vehicle groups, respectively. These were obtainedt 70 mm Hg of distending pressure in rats with approximateransformed EMG values of 3 (naloxone) and 1 (vehicle) toatch the approximate group mean values at 70 mm Hg. Dot-
ed lines signify pressure onset and termination of the 20-sec-nd distending stimulus.
ither group ZV or group ZZV. These data are consistent
wtia
E
IP
hitdggsgNezhtrsspsZaFea.
B
adgoro
D
mAa
FGPP*#Gp
FGfbohhZPZ
TGEN
919ORIGINAL REPORT/DeBerry et al
ith the outcomes shown in experiment 1 and indicatehat a prior bout of zymosan exposure in adulthood did notnfluence the magnitude of opioid inhibition produced bysubsequent acute exposure to zymosan.
igure 5. Group mean EMG responses during graded UBD.roups received either anesthesia alone (A) or zymosan (Z) from14-P16, followed by zymosan 24 hours before UBD testing at120 in the presence of either vehicle (V) or naloxone (N).Group AZN significantly different from group AZV, P � .05;group ZZV significantly different from group AZV, P � .05.roups ZZV, ZZN, and AZN did not differ at any distending
igure 4. Group mean EMG responses during graded UBD.roups ZZV, ZZN, and Z-N received zymosan (Z) from P90-P92,
ollowed by either zymosan (Z) or no treatment (–) 24 hoursefore UBD testing at P120 in the presence of either vehicle (V)r naloxone (N). Groups ZV and ZN received zymosan (Z) 24ours before UBD testing at P120 in the presence of either ve-icle or naloxone. *Group ZN significantly different from groupV, P � .05; #group ZZN significantly different from group ZZV,� .05. Groups ZV, ZZV, and Z-N did not differ. Groups ZN andZN did not differ.
tressure.
xperiment 5
nflammation at P14-P16 and 24-Hour AdultretreatmentExperiment 5 was performed to determine whether aistory of zymosan-induced bladder inflammation dur-
ng the neonatal period would impair the opioid inhibi-ory effect resulting from an acute exposure to zymosanuring adulthood. Fig 5 presents EMG responses duringraded UBD. An ANOVA revealed a significant between-roups effect, F(3,32) � 2.95 (P � .05); a significant pres-ure effect, F(7,21) � 53.95 (P � .001); and a significantroups x pressure interaction, F(7,224) � 2.06 (P � .005).aloxone administration revealed the opioid inhibitoryffect in rats given anesthesia as neonates and the acuteymosan treatment (group AZN vs AZV) but failed toave any effect in rats that received neonatal exposureo zymosan (group ZZN vs ZZV). Specifically, in groupseceiving anesthesia only at P14-P16, post hoc compari-ons showed that the EMG responses of group AZN wereignificantly greater than group AZV at UBD distendingressures of 30 to 80 mm Hg. However, there were noignificant differences in the EMG responses of groupsZV and ZZN at any distending pressure, nor were thereny differences between these groups and group AZN.inally, the neonatal zymosan treatment was effective innhancing the EMG responses (group ZZV vs group AZV)t UBD distending pressures of 50, 70, and 80 mm Hg (P �054 at 60 mm Hg).
ladder WeightsAt the termination of experiments 1, 4, and 5, each
nimal’s bladder was removed and the tissue was blottedry. Table 1 shows there were no significant between-roups differences in bladder weights. These data ruleut the possibility that bladder hypertrophy occurred inats receiving intravesical zymosan on P14-P16, P90-P92,r P120.
iscussionUrinary bladder pain is extremely common in the fe-ale population, with a lifetime incidence rate of 50%.lthough bladder inflammation resulting from mycoticnd bacterial infections accounts for most of these cases,
able 1. Group Mean Bladder Weights inrams (�SEM) Obtained From Rats inxperiments 1, 4, and 5: ANOVAs Revealedo Significant Between-Group DifferencesEXPERIMENT 1 EXPERIMENT 4 EXPERIMENT 5
ZN 0.16 � 0.02 ZN 0.14 � 0.01 ZZN 0.18 � 0.01ZV 0.14 � 0.02 ZV 0.16 � 0.01 ZZV 0.17 � 0.02AN 0.15 � 0.02 ZZN 0.14 � 0.01 AZN 0.14 � 0.01AV 0.13 � 0.01 ZZV 0.14 � 0.01 AZV 0.16 � 0.01
Z-N 0.14 � 0.01
here is also a substantial number of individuals that ex-

pdcsmgsiaednmtdpiwt
tptatbctes
ihioneidhw
dcipCor2lUinbS(ra
capsfsbiprnrrou
nsemeglrtaet
tsOflaoiobbwpmarcte
ofsPttoimt
920 Inflammation-Induced Enhancement of the Visceromotor Reflex to UBD
erience bladder pain from diseases such as painful blad-er syndrome (PBS)/interstitial cystitis (IC), when no mi-robial infection is present. In previous and presenttudies, we demonstrated that briefly exposing adult fe-ale rats to intravesical zymosan, a mycotic inflammo-en, resulted in modest but significant bladder hyper-ensitivity during graded UBD 24 hours later,25 and wenterpreted this hypersensitivity as possibly representing
hyperalgesic state. Bladder hypersensitivity was mark-dly enhanced when rats also experienced 3 consecutiveays of exposure to intravesical zymosan during the neo-atal period (P14-P16) but not if that treatment was ad-inistered during the adolescent period (P28-P30).26 We
heorized that neonatal inflammation “primes” or “pre-isposes” a rat to develop a hypersensitive bladder, es-ecially when it is re-exposed to an additional bladder
nsult as an adult, and we suggested that experienceith neonatal bladder inflammation may be one poten-
ial cause for disorders such as PBS/IC.26
Independent studies of primary hyperalgesia involvinghe knee joint31 or hind paw28,39 have demonstratedrogressive engagement of descending inhibitory sys-ems during the first 24 hours of acute inflammation thatct to suppress somatic hyperalgesia. This prompted uso consider whether bladder hypersensitivity producedy acute bladder inflammation in the adult rat may beoncomitantly suppressed by inflammation-induced ac-ivation of an inhibitory system and whether neonatalxposure to zymosan might, in some fashion, impair thisystem from functioning effectively in adulthood.Experiment 1 demonstrated that a brief exposure to
ntravesical zymosan in the adult rat resulted in a bladderypersensitivity that was being suppressed by an opioid
nhibitory system because administration of i.p. nalox-ne before UBD testing significantly increased the mag-itude of EMG responses to tests of graded UBD. Thisffect was replicated in experiments 4 and 5. The opioidnhibition was triggered by bladder inflammation andoes not appear to be tonically active, since naloxonead no effect on EMG responses to UBD as comparedith vehicle in the noninflamed condition.Although naloxone has been reported to affect blad-er capacity in chloralose- or ketamine-anesthetizedats,1 our results cannot be explained by naloxone-nduced changes in bladder compliance. Specifically, ex-eriment 2 showed that intravesical pressures duringMG testing were not differentially affected by nalox-ne or vehicle administration in zymosan-treated rats orats without inflammation. We intentionally infused.0 mL of volume to achieve distending pressures equiva-ent to the maximal pressures tested in studies of gradedBD. The fact that i.t. administration of naloxone in exper-
ment 3 produced similar findings to i.p. administration ofaloxone also suggests that naloxone-induced changes inladder compliance are unlikely to account for our results.himizu et al30 also failed to find any influence of naloxone0.5 mg/kg, intravenous) alone on various cystometric pa-ameters and similar negative results have been reported in
conscious rat preparation.23 2Cruz and Downie6 have shown that abdominal muscleontractions are present during spontaneous voidingnd during voiding evoked by saline infusion within thehysiological range of distending pressures in both con-cious and urethane-anesthetized rats. It should there-ore be acknowledged that the VMR used in the presenttudy is not necessarily specific for nociceptive events,ut this response does increase in a graded fashion to
ntensities of UBD that are above the nonpainful, normalhysiological range of intensities. This suggests that it isepresentative of primary afferent processing related tooxious intensities of UBD. The demonstration of opioid-elated effects further supports the use of this model asepresentative of nociceptive sensory processing, as opi-ids have limited conscious effects apart from pain mod-lation.3,22
Experiment 3 suggested that the effects of systemicaloxone were likely due to blockade of a spinal opioidystem, since i.t. administration of naloxone significantlynhanced the EMG responses to UBD in a manner re-arkably similar to i.p. naloxone. The length of i.t. cath-
ters was intended to deliver naloxone to the L5-S2 re-ion of the cord, and we believe this to be the primary
ocus of action. However, we cannot rule out possibleostral spread of naloxone to other spinal regions impor-ant for bladder control (eg, T12-L1). In addition, therelso could be a peripheral opioid component becausenkephalins are present in parasympathetic ganglia onhe bladder.7,14,17
The effects observed in the present studies suggesthat 2 opposing processes contribute to bladder hyper-ensitivity resulting from acute bladder inflammation.ne process is enhanced input, perhaps mediated by in-ammation-induced sensitization of primary nociceptivefferents12,13,19 and/or central sensitization of second-rder spinal dorsal horn neurons.4,40 A second process
nvolves inflammation-induced activation of a reactivepioid system that opposes enhanced input from theladder. In our studies of adult rats treated with a singlerief bout of bladder inflammation, opioid inhibitionas present, but it was not sufficient to completely sup-ress the enhanced input resulting from bladder inflam-ation. Modest bladder hypersensitivity was the result-
nt state. This spinal opioid inhibitory influence mayeflect a system of interneurons localized to the spinalord, since enkephalinergic neurons are prevalenthroughout the regions of hypogastric and pelvic affer-nt spinal input.18
We then considered whether prior activation of thispioid inhibitory system might alter how it was mani-ested if reactivated later in life. We examined whether 3uccessive days of urinary bladder inflammation at P90-92, followed by re-exposure to brief bladder inflamma-ion at P120, influenced the magnitude of opioid inhibi-ion. We found no influence of the P90-P92 treatmentver and above that produced by exposure to bladder
nflammation 24 hours before UBD testing at P120. Theagnitude of bladder hypersensitivity and opioid inhibi-
ion were no different from that produced by the adult
4-hour bladder inflammation pretreatment alone.
npettnh(tae
eicfosgpEowtaasttifotaca
Pttntlsa1ssflllt
mttsna
�
ctcnitattftdtsesthes
vmtdddskjdsntisi
tasfdfnraapcdosttg
921ORIGINAL REPORT/DeBerry et al
Finally, we examined whether 3 successive days of uri-ary bladder inflammation at P14-P16 impaired the ca-acity to activate this opioid inhibitory system after re-xposure to brief bladder inflammation in adulthood,hus contributing to the enhanced bladder hypersensi-ivity observed in our previous studies.26 Naloxone hado effect on EMG responses to UBD over and above ve-icle in neonatal zymosan-treated rats in experiment 5groups ZZN and ZZV did not differ). These data supporthe view that neonatal bladder inflammation, unlikedult bladder inflammation, impairs the functioning orffectiveness of an opioid-inhibitory system.We considered the possibility that the failure to see an
nhancement of EMG responses after naloxone admin-stration in neonatal zymosan-treated rats was due to aeiling effect in our EMG response measure, but severalactors make this interpretation unlikely. First, we havebtained substantially larger EMG responses in previoustudies than those obtained in the present studies,26 sug-esting that a response ceiling was not attained in theresent studies. Second, there were no differences in theMG responses of groups ZZN and ZZV in the lower rangef graded distending pressures (eg, 30 to 60 mm Hg),here the EMG responses were approximately 50% of
he maximal responses observed. If naloxone was havingn effect in group ZZN, then it should have been observ-ble at these submaximal response levels. We also con-idered the possibility that neonatal bladder inflamma-ion produced histological changes that contributed tohese outcomes. However, we have performed histolog-cal analyses on a limited number of bladders obtainedrom adult rats treated with either intravesical zymosanr anesthesia alone from P14-P16 in a manner identicalo experiment 5 but in the absence of any additionaldult treatments. We have observed no major histologi-al differences between animals treated in this mannert this point in our analyses.Finally, we considered whether failure of the adult
90-P92 zymosan treatment to alter the functioning ofhe opioid inhibitory system was due to the possibilityhat 3 consecutive days of inflammation at P90-P92 mayot be comparable to 3 consecutive days of inflamma-ion at P14-P16; that is, the adult treatment resulted iness severe inflammation. Data from our previous studiesuggest that this is unlikely. We have demonstrated thatdolescent rats treated in a similar fashion for either 3 or4 consecutive days both fail to show bladder hypersen-itivity in adulthood after brief re-exposure to zymo-an.26 The primary focus of administering 14 days of in-ammation in those studies was to determine whether a
onger duration of exposure to zymosan was requiredater during development to produce similar outcomeso 3 days of inflammation during the neonatal period.The issue of how neonatal exposure to bladder inflam-ation impairs opioid inhibition is a matter of specula-
ion. First, it may interfere with the development of ei-her opioid inhibitory interneurons or a descendingystem that utilizes spinal opioids in modulating spinalociceptive transmission from bladder afferents. For ex-
mple, neonatal bladder inflammation might reduce u-opioid or -opioid receptor expression in the spinalord. Similar views have been advanced to account forhe loss of spinal morphine suppression of somatic noci-eption and tactile allodynia after tight ligation of L5/L6erves.24 There also is abundant evidence that descend-
ng inhibitory systems derived from either the rostroven-ral medulla (RVM)11,32 or the locus coeruleus (LC)33-36
re recruited after inflammation of somatic tissue. Al-hough these descending systems are present at birth,hey are only partially functional by P10-P12 and becomeunctionally mature by P21.2,9,10 Perhaps the neonatalreatment impaired the development of one of theseescending inhibitory systems. Alternatively, it may behe case that the neonatal treatment enhances the den-ity and spinal segmental distribution of bladder affer-nts, as has been demonstrated for somatic afferents intudies of neonatal hind paw inflammation.29 If nocicep-ive input was markedly enhanced and a developing in-ibitory system was not altered to compensate for thisnhanced input, then the effectiveness of the inhibitoryystem would be functionally reduced.An alternative that is functionally equivalent to these
iews is that neonatal bladder inflammation might aug-ent the operation of a descending system whose role is
o inhibit a spinal opioid inhibitory circuit, for example,isinhibition or facilitation. This notion is consistent withata of MacArthur et al,20 which showed that removal ofescending spinal influences by mid-thoracic spinal tran-ection resulted in increases in both dynorphin and en-ephalin mRNA in rats receiving unilateral hind paw in-ection of CFA. They concluded that the net effect ofescending afferents on spinal nociceptive circuits was touppress the response of opioid-containing neurons tooxious stimulation from the periphery. Hence, augmen-ation of this descending influence via neonatal bladdernflammation would functionally operate to produce theame outcome as simple impairment of a descendingnhibitory system.
Coutinho et al5 have provided an interesting parallelo the present findings in their studies of maternal sep-ration in producing visceral hypersensitivity. They mea-ured the VMR to graded colorectal distention andound that intraperitoneal naloxone administration pro-uced a parallel leftward shift in the stimulus-responseunction of adult rats that were not handled or mater-ally separated as neonates but had no effect in adultats that had previously undergone maternal separations neonates. The maternally separated rats also showeddiminished stress-induced cutaneous analgesia com-
ared with nonhandled control animals, suggesting aompromised ability of the former rats to activate en-ogenous opioidergic pain inhibitory systems. Althoughur studies controlled for maternal separation, thesetudies more generally demonstrate that opioid-inhibi-ory mechanisms can be impaired in rats by exposinghem to a neonatal challenge and are consistent with areater activation of these systems in rats that do not
ndergo neonatal challenges.
icieptc
flratbpe
R
1up
2ns
3ut
4c
5BacP
6vR
7Cot
8lmS
9s7
1oo
1ai
1mm4
1tfl4
1S
922 Inflammation-Induced Enhancement of the Visceromotor Reflex to UBD
In conclusion, our data indicate that urinary bladdernflammation induces a bladder hypersensitivity that isoncomitantly opposed by spinal opioid inhibition. Moremportantly, this inhibition can be disrupted by neonatalxposure to bladder inflammation, suggesting that someainful bladder disorders, such as those triggered by ei-her mycotic inflammation or PBS/IC, may in part be a
onsequence of early-in-life experiences with bladder in- daid S, Goldstein M: Peptide neurons in peripheral tissues
iA
1c
1ti2
1Ssc
1kvv
1oA
2Dng
2Sai
2ptJ
2Af
2of
2ap2
2ns
2nJ
2
ammation, as we have argued elsewhere.26 Since a va-iety of other disorders, such as irritable bowel syndromend fibromyalgia, also have been attributed to a diminu-ion in the effectiveness of inhibitory systems, it woulde interesting to see whether epidemiologic data sup-ort the occurrence of some similar type of neonatalvent that contributed to the underlying cause of those
isorders.eferences
. Booth AM, Hisamitsu T, Kawatani M, De Groat WC: Reg-lation of urinary bladder capacity by endogenous opioideptides. J Urol 133:339-342, 1985
. Boucher T, Jennings E, Fitzgerald M: The onset of diffuseoxious inhibitory controls in postnatal rat pups: A C-Fostudy. Neurosci Lett 257:9-12, 1998
. Castroman P, Ness TJ: Vigor of visceromotor responses torinary bladder distension in rats increases with repeatedrials and stimulus intensity. Neurosci Lett 306:97-100, 2001
. Cervero F: Visceral pain: Mechanisms of peripheral andentral sensitization. Ann Med 27:235-239, 1995
. Coutinho SV, Plotsky PM, Sablad M, Miller JC, Zhou H,ayati AI, McRoberts JA, Mayer EA: Neonatal maternal sep-ration alters stress-induced responses to viscerosomatic no-iceptive stimuli in rat. Am J Physiol Gastrointest Liverhysiol 282:G307-G316, 2002
. Cruz Y, Downie JW: Abdominal muscle activity duringoiding in female rats or irritated bladder. Am J Physiolegul Integr Comp Physiol 290:R1436-R1445, 2005
. De Groat WC, Kawatani M, Hisamitsu T, Lowe I, Morgan, Roppolo J, Booth AM, Nadelhaft I, Kuo D, Thor K: The rolef neuropeptides in the sacral autonomic reflex pathways ofhe cat. J Auton Nerv Syst 7:339-350, 1983
. DeBerry J, Uzzell T, Ness TJ, Randich A: Effects of early-in-ife bladder inflammation on spontaneous micturition and
icturition reflexes during cystometry (CMG) in adult rats.ociety for Neuroscience Abstract, 2005
. Fitzgerald M, Jennings E: The postnatal development ofpinal sensory processing. Proc Natl Acad Sci U S A 96:7719-722, 1999
0. Fitzgerald M, Koltzenburg M: The functional developmentf descending inhibitory pathways in the dorsolateral funiculusf the newborn rat spinal cord. Brain Res 389:261-270, 1986
1. Guan Y, Terayama R, Ren K, Dubner R: Plasticity in excitatoryminoacid receptor-mediateddescendingpainmodulationafter
nflammation. J Pharmacol Exp Ther 300:513-520, 2002
2. Habler H-J, Janig W, Koltzenberg M: Activation of un-yelinated afferent fibres by mechanical stimuli and inflam-ation of the urinary bladder in the cat. J Physiol (Lond)
25:545-562, 1990
3. Habler HJ, Janig W, Koltzenberg M: Receptive proper-ies of myelinated primary afferents innervating the in-amed urinary bladder of the cat. J Neurophysiol 69:395-02, 1993
4. Hokfelt T, Schultzberg J, Elde R, Nilsson G, Terenius L,
ncluding the urinary tract: Immunohistochemical studies.cta Pharmacol Toxicol 43(Suppl 2):79-89, 1978
5. Holm S: A simple sequentially rejective multiple test pro-edure. Scand J Stat 6:65-70, 1979
6. Johnson OL, Berkley KJ: Estrous influences on micturi-ion thresholds of the female rat before and after bladdernflammation. Am J Physiol Regul Integrative Comp Physiol82:R289-R294, 2002
7. Kawatani M, Lowe IP, Booth AM, Backes MG, ErdmanL, De Groat WC: The presence of leucine-enkephalin in theacral preganglionic pathway to the urinary bladder of theat. Neurosci Lett 39:143-148, 1983
8. Llewellyn-Smith IJ, Dicarlo SE, Collins HL, Keast JR: En-ephalin-immunoreactive interneurons extensively inner-ate sympathetic preganglionic neurons regulating the pel-ic viscera. J Comp Neurol 488:278-289, 2005
9. McMahon SB: Neuronal and behavioural consequencesf chemical inflammation of the rat urinary bladder. Agentsctions 25:231-233, 1988
0. MacArthur L, Ren K, Pfaffenroth E, Franklin E, Ruda MA:escending modulation of opioid-containing nociceptiveeurons in rats with peripheral inflammation and hyperal-esia. Neurosci 88:499-506, 1999
1. Morin M, Lafrenaye S, Goffaux P, Paul-Savoie E, Marchand: Effect of early pain experience on pain modulation mech-nism of adolescents. American Pain Society Annual Meet-
ng Abstract 643, 2007
2. Ness TJ, Lewis-Sides A, Castroman P: Characterization ofressor and visceromotor reflex responses to bladder disten-ion in rats: Sources of variability and effect of analgesics.Urol 165:968-974, 2001
3. Pandita RK, Pehourson R, Christoph T, Friderichs E,ndersson K-E: Actions of tramadol on micturition in awake,
reely moving rats. B J Pharmacol 139:741-748, 2003
4. Porreca F, Tang QB, Bian D, Riedl M, Elde R, Lai J: Spinalpioid mu receptor expression in lumbar spinal cord of ratsollowing nerve injury. Brain Res 795:197-203, 1998
5. Randich A, Uzzell T, Cannon R, Ness TJ: Inflammationnd enhanced nociceptive responses to bladder distensionroduced by intravesical zymosan in the rat. BMC Urol 6:2-8,006
6. Randich A, Uzzell T, DeBerry JJ, Ness TJ: Neonatal uri-ary bladder inflammation produces adult bladder hyper-ensitivity. J Pain 7:468-479, 2006
7. Ren K, Dubner R: Enhanced descending modulation ofociception in rats with persistent hindpaw inflammation.Neurophysiol 76:3025-3037, 1996
8. Ren K, Ruda MA: Descending modulation of Fos expres-

s7
2Ae
3pl1
3Cwt
3pfl
3tpB
3t
i8
3fln
3bu3
3rg2
3t3
3td1
4
923ORIGINAL REPORT/DeBerry et al
ion after persistent peripheral inflammation. Neuroreport:2186-2190, 1996
9. Ruda MA, Ling QD, Hohmann AG, Peng YB, Tachibana T:ltered nociceptive neuronal circuits after neonatal periph-ral inflammation. Science 289:628-631, 2000
0. Shimizu I, Kawashima K, Ishii D, Oka M: Effect of (�)-entazocine and 1,3-di-o-tolylguandidine (DTG) sigma (�)
igands, on micturition in anesthetized rats. Br J Pharmacol31:610-616, 2000
1. Schaible HG, Neugebauer V, Cervero F, Schmidt RF:hanges in tonic descending inhibition of spinal neuronsith articular input during the development of acute arthri-
is in the cat. J Neurophysiol 66:1021-1032, 1991
2. Teryama R, Guan, Y, Dubner R, Ren K: Activity-inducedlasticity in brain stem pain modulatory circuitry after in-ammation. NeuroReport 11:1915-1919, 2000
3. Tsuruoka M, Willis WD Jr: Bilateral lesions in the area ofhe nucleus locus coeruleus affect the development of hy-eralgesia during carrageenan-induced inflammation.rain Res 726:233-236, 1996
4. Tsuruoka M, Willis WD Jr: Descending modulation from
he region of the locus coeruleus on nociceptive sensitivity in rat model of inflammatory hyperalgesia. Brain Res 743:6-92, 1996
5. Tsuruoka M, Maeda M, Inoue T: Persistent hindpaw in-ammation produces coeruleospinal antinociception in theon-inflamed forepaw of rats. Neurosci Lett 367:66-70, 2004
6. Tsuruoka M, Matsutani K, Inoue T: Coeruleospinal inhi-ition of nociceptive processing in the dorsal horn duringnilateral hindpaw inflammation in the rat. Pain 104:353-61, 2003
7. Vanegas H: To the descending pain-control system inats, inflammation-induced primary and secondary hyperal-esia are two different things. Neurosci Lett 361:225-228,004
8. Vanegas H, Schaible HG: Descending control of persis-ent pain: inhibitory or facilitatory? Brain Res Rev 46:295-09, 2004
9. Wei F, Ren K, Dubner R: Inflammation-induced Fos pro-ein expression in the rat spinal cord is enhanced followingorsolateral or ventrolateral funiculus lesions. Brain Res 782:36-141, 1998
0. Woolf CJ: Evidence for a central component of post-
njury pain hypersensitivity. Nature 306:686-688, 1983



















