
initiation of the distention-induced descending peristaltic reflex ...
114
INITIATION OF THE DISTENTION-INDUCED DESCENDING PERISTALTIC REFLEX IN THE OPOSSUM ESOPHAGUS: ROLE OF MUSCLE CONTRACTILI'IY by Ahmad Muinuddin A thesis submitted to the Department of Physiology in conîomity with the requirements for the degree of Master of Science Queen's University Kingston, Ontario, Canada December, 1998 copyright 8 Ahmad Muinuddin
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of initiation of the distention-induced descending peristaltic reflex ...

INITIATION OF THE DISTENTION-INDUCED DESCENDING
PERISTALTIC REFLEX IN THE OPOSSUM ESOPHAGUS:
ROLE OF MUSCLE CONTRACTILI'IY
by
Ahmad Muinuddin
A thesis submitted to the Department of Physiology
in conîomity with the requirements for the degree of
Master of Science
Queen's University
Kingston, Ontario, Canada
December, 1998
copyright 8 Ahmad Muinuddin

National Library Bibliothèque nationale du Canada
Acquisitions and Acquisitions et Bibliographie Services services bibliographiques
395 Wellington Street 395. rue Wellingtm OtrawaON K 1 A W OttawaON K1AON4 Canada canada
The author has granted a non- exclusive licence allowing the National Library of Canada to reproduce, loan, distriiute or selI copies of this thesis in microform, paper or electronic formats.
The author retains ownership of the copyright in this thesis. Neither the thesis nor substantial extracts fiom it may be printed or othenvise reproduced without the authoc's permission.
L'auteur a accordé une licence non exclusive permettant à la Bhfiotheque nationale du Canada de reproduire, prêter, distriiuer ou vendre des copies de cette thèse sous la fome de micr&che/nlm, de reproduction sur papier ou sur format électronique.
L'auteur conserve la propriété du droit d'auteur qui protège cette thèse. Ni la thèse ni des extraits substantiels de celle-ci ne doivent ê e imprimés ou autrement reproduits sans son autorisation.

Distedon-mduced or secondary peristalgs is the physiological mec-
wfiereby refluxed gestric contents or food left behind &a primary peiistalsis are propefled
hto the stomach, Previous studies using a triple-chamber organ baîh have demonstrateci
that the balloon distention PD)-mduced descendhg peristaltic reflex k the opossum
smooth muscle esophagus mvoives long descendhg intramural neurons (Paterson anci
1ndrakrishna.n 1995). However, this reflex is abolished when a Ca2+-k Krebs solution is
placed at the site of mention, suggesting that either -tic transmission occurs at the
origin of the refleex, or initiation of the reflex requires the development of muscie tension in
response to BD.
To test the latter possiiiiity, the BD-mducd descending peristaltic reflex was
studied ushg an 8- 10 cm length of srnooth mus& esophagus in a dual chamber organ
bath before and d e r the addition of agents that affect smooth muscle c~ntractiiity~ The
preparation was such that the p r o d (orai) and distal (abord) segments of the snooth
rn-le esophagus were in chemicaUy W lated chambers, leaving the neuromuscular.
apparatus between segments intact. Foliowing BD in the oral chamber* "O££" contractions
were recordai m both the oral and abord chamber. Addition to the oral chamber (Le. site
of distention) of either nifedipine or cheleryhke chionde, both inhibitors of saooth
muscle contractility, aboiïshed the BD-mduced contractioos m both the oral and abrai
chambers. The abord response to electricai field stimulation was not inhieci by
nifidipine demonstrating that it did not alter the neiiral pathways hvohred m this reflex m
our preparation

To examine the e h of these agents on tonk an isobaric barostat
with a cornpliant 8 cm long poiyethylene bag was eniployed. Neither Ca2*-free Krebs
solution, chelerythrine chloride, nifwip'he nor the neurotoxin tetrodotoxin, dtered
pressure-vohrme c w e s suggesting that restiog esophageal body tone and resistance to
initial stretch are not dependent on n e d h o r s or snooth muscle contmdity.
However, both nifedipine and chelerythaine chloride, as wel as CaZ* -fke Krebs solution
mark* decreased phasic con;traCtions over top of the distendmg bag7 hduced by ramp
distestion wiîh the barostat.
Taken togeth.r7 these observations suggest that local stretch-induced phasic
muscle contracton plays an miportant role m the Siitiation of the BD-mduced descending
peristaitic reflex.
iii

Statement of CeAuthorship
Uoder the guidance of Dr. W.G. Paterson, Ahmad Muin* designed,
performed end adyzed the exp&mts descri'bed in this tfiesis. Onginal drafts of aU text
were written by Ahmaù Muinuddin The finishad produci Hicludes editorial suggestions by
Dr. Patersoril

This thesis is dedicated m loving memory of my aunt, Zahida Jabeen Shariff(1942 -1997).
1 em gmtefbi to severai mdividuals, each ofwhom have played cm miportant part m enabhg
me to reach this stage in my aireer. First, 1 wouid Wre to thaak my nipavisor, Dr. W.G.
Paterson, for giving me the opportimity to participate m the excitmg universe of medical
research- His guidance, patience and confidence in my abilities over the past two years have
contributed to my graduate Srpenence beyond measure. 1 am gmtefid to Rofessor Wd.
MacKay, my mentor fiom the University of Toronto, who has aiways supporteci my
endeavours. Hats off to my good niends, 'Doc' Dave Miller a technologist pur excellent-
whom I have had the privilege of working with and leamhg h m and my iab buddy, Rob
White -the fbture Possum King. Special thanks to DTS. Vanner, Hill, Moore, Barajas-Lopez,
Nakatsu and Faguson for givmg constructive feedback and Drs. Beck and Ying for their
contiming encouragement during tough times. Thanks to Margi and ïla for sharing their
equipment and expertise, Pat and CLinm the GI Function Lab for aiways accommodatmg me
into their busy schedules and Brenda and Sandra for aU of their administrative assûtance and
for aIways wearing a d e . The schoiariy environment provided by the fàcuhy and N b the
department of physiology was much appreciated. Thdcs to the St. Lawrence College
Piacement Students -Beverly, Mary, MicheHe, AI& Christme, Martha, Liz, Lmda and Lisa-
for helping m the lab and a s h g good questions. I am indebted to my h i i y -Mom, Dad and
Tariq- for their never endhg love and unwaveringgsupport -in particula. my fàther, fm always
challenging me to do my best, and have fun, and for waking me up m the morning. To my
Wong Eends - f i e r and Tabassum, Carl, Ilan, h a n , Jamal, Khurram, Masha, Mcma,
Mustafa, Romana, Saba, Sabeen, Super Jen, and Suraya and Mhi -may God bless you di.
Financial support fhm Queen's University and Procter and Gamble was gma@ appreciated.
v

Table of Contents
. Abstract .........................................................*............................... ....................Il
.................................................... Staîernent of Co-Authorshq, ................... ... N ............................................................................................ Achowledgements v
............................................................................................... TabIe of Contents vi .................................................................................................... List of Figures x
List of Abbreviaîions ........................................................................................... xi
Introduction
I) Esophageal Anatomy and Physiology O The Fundamentals ....................... ... 1
Upper Esophageal Sphincter ............................................................ 4 Esophageai Body ............................................................................. 4 Lower Esophageal Sphmcter ................................................................. 5 Esophageal Motor Physiology ..................... ..... .............................. 6
............*.......... ..................................................... Primary Peristalsis ... 6 Secondary Peristalsis ................... .. ....................................................... 8
........................... II) OveMew of the Contml of Esophageal Motor Function 10
............................................................... Innervation of the Esophagus 10 Swdowing Center .................... ... ....... .. ............................................. 10 i)Merent Input .......................... .. .. .. .................................................. 1 1 iiiCoordinatÏng Region .................... ... ..... ....................... ............ 1 1 iiiiEfferenî Output ................................................................................... 12 Extrinsic Innervation .................... ,.. ............. .. ............................ 1 2 OMotor Innervation ................... ................ ......................................... 12 hiSensory Innervation ............................................................................. 14 Intrinsic Innervation ................................................................................. 16 i)Enteric Nervous System ....................................... ................................ 16
Control of Esophageal PeristPlsis ............. .. ..... .. ............................... 20 central CoIltrol ..................................................*.................................... 20 i) S triated Muscle Esophagus ................................................................. 2 0 iiiSxnooth Muscie Esophagus .............................................................. 2 0 Peripheral Control ....................... ... .................................................... 22 ï)Intrinsic Neural Control ................... .. ........................................... 2 2 iiIntrinSic Myogenic Contml .................... .. ........................................ -26

iia)Sigd Transduction Pathways in Esophageal Circular Muscle ............. 28 Integraîïon of Control Mechanismic ......................................................... 31
.......................... ...*. ........ III) The Dbtention-Indaced Peristiitic R e k .. ....... 32
Thesis Objectives ................................................................................................ 37
Chapter 2
Methods
Rationale for Method Used ................................................................................. 39 Animal Models ................... .... ................................................................. 39
......................................... Assesment of Esophageal Motor Function 4 0 ï)Manometry ........................................................................................... 40 ii)Electronic Barostat ...................... ..... .......O.................................. 4 1 Pharmacological Agents ...................................................................... 42
.................... ......................................... ................... Specinc Protocols .. ... 43 Removal of the Esophagus ..................................................................... 43
......................................... Dose Response Curves ...................... .... 44 Descendhg Peridtic Reflex: Double-Cbambered Bath Experiments ....... 46
................... ...................... Esophageal Cornpliance: Barostat Studies .. 5 0
.......................................................... ..................... Data Anaiysis ........... 52 Dose Response C w e s ..................................... .. ................................... 52
................................................... Descendhg Peristaltic Reflex Studies 5 4 Barostat Studies ............................ ......................................................... 54 Sutkt îcd Methods ........... .........,. ....................................................... 55
Chapter 3
Dose Determination ......................................................................................... 58
Descendhg Peristaltic Reflex Studies ................... .. ......................................... 58 .... ...................... Effect of Inhi'bitors of Smooth Muscle Contraction .,., ,.. 62
ï)Effect of O ca2' 20 mM ~ g " Krebs ................. .......... ...................... 62 QEffect of Niedipine .............................................................................. 62
.................... ......*.*....*........... üiiEffect of Chelerythrine Qloride ..... 65
............ ......................... EBxt of Blockade of Synaptic Transmission .. 65 i)Abod Chamber .................................................................................... 65

Esophageai C o m p W Studies .......................................................................... 67 E f k t of Smoth M i d e Relaxants and Neural Blockade on: i)Esopbaged Compliance ................................................................... 67 ii)Resistance of Esophageal Wall to Inaial Smch .................................... 69 k3Phasic Contractions Over Top of the Distendhg Bag ........................... 69
Chapter 4
Discussion
Descendhg Pexkkthic Reflex Studies .................................................................. 76 ..... .................... ï)Role of Local Smooth Muscle Contractiüty .... ......... 76
6)Role of Synaptic Transmission: Ef£êct of co-Conotoxin GMA ............... 78
Barostat Studies .............................................................................................. 80 ï)Esop&eal Body Resting Tone ............................................................. 80 iiPhasic Muscle ContractÜay ............................................................... 82
VITA .............. ..................... ............................................................................... 102

List of Figures
Figure 1 . Figure 2 .
Figure 3 .
Figure 4 . Figure 5 . Figwre 6 . Figure 7 . Figure 8 . Figure 9 . Figure 10 . Figure 11 . Figure 12 .
Figure 13 .
Figure 14 .
Figure 15 .
Figure 16 .
Figure 17 .
Neuromuscular anatomy of the esophagus .................................... 3
htracelhdar rnechanism of contraction of esophaged body muscle in respome to ACh or nare stimulation ......................... 30
Schematic representation of (A) possible i n t r a m d mural pathways nsponsble for distention-Hrduceû descending peristaltic reflex in the opossum esophegus and (B) triple-chamber organ
................................................................................... bath set-up 35
.......................... Sche-tic representation of muscle strip set-up 45
............... ......................... In vitro double chamber set-up ... 47
......................... In vitro barostat set-up ................................. ....... 51
.............. Er-vivo pressure-vohnne relationship of the baiostat bag 53
........................... .................... Barostat distention protocol ... 53
Barostat data collection .................................. .,.,. ................. 56
...... Dose response curves of Sedipine d chelerytbrk chloride 5 9
..... ...................... Response of esophagus to BD and EFS ......... 60
Effect of TfX on BD & EFS induced peristattic contraction amplitudes .................................................................................. 61
Effect of ~ a 2 ' - f k e Krebs on BD & EFS induced perisialtic contraction amplitudes ................................................................ 63
Effect of nifiedipine on BD & EFS induced -ic ...... contraction amplitudes .................... ........................... 64
Effect of chelerythrine chloride on BD & EFS induceci peristaltic contraction amplitudes ................................................................. 66
Effêct of wCTX on BD-mduced peristahic contraction amplitudes .................... .. .... .. .............................................. 68
Effect of -0th muscle relaxants and 'ITX on esophaged cornphce .............................................................................. 7 0

Figure 18 . E&* of -0th muscle netaxants and TTX on response to initial stretch ............................................................................... 71
Figure 19 . Profile of phasic contractiüty over top of the bstat bag ............ 72
Figure 20 . Effect of smooth muscle relaxants and m[ on contraction Iadex .. 74
Figure 2 1 . Effèct of distendhg barostat bag pressure on contracton index ..... 74

List of Abbreviations
O c
Ach ANOVA BD ca2' CC
chele CM CI CNS 0-CTX DAG EFS ENS Gr Hz ICC LES LM M mA M C ml mM =Hg ms MP NANC NO VIP NT P PKC PNS SEM
de- Celsius acetylcholm analph of variance balloon distention calcium ion cubic catimeter chelerythrine chioride circuiar muscle contracton index central navous system omega-conotoxh GVIA diacyigiycerol eIectricai field stimulsition enteric nervous system gastrointestinal hertz InterStifiaf Celis of Cajal lower esophageal sphincter 1ongmiM muscle molar müliamp rnagnesium ion mülditre millimolar mülimews of mercury mülisecond myenteric plexus wnadrenergic nonchoiinergic nitnc oxide v a s o h e mtestinal peptide neuroneansmitter probabiiÏty that nuii hypothesis is true protein kinase C peripheral nemous system standard error of the mean tmdotoxin upper esophageal sphincter voit

Chapter 1
Introduction
The esophagus is a hoiiow m u d a r tube wbich connects the pharynx to the
stomach. It is closed at the p r o d end by the upper esophageal sphincter (UES) and at
the distal end by the lower esophageal sphincter (LES). The primary fiinction of the
esophagus is to propel a food or fluid bolus ifom the pharynx into the stornach. This is
accomplished by the deglutition or swallowing reflex and gravity. The UES prevents entry
of air into the esophagus duruig respiration, and also the retrograde rnovement of
esophageal contents into the hypopharynx. The LES prevents reflux of gasaic metions
h o the esophagus. Cornplex and incompletely understood interactions between the
central nervous system (CNS), entenc nervous system (ENS) and muscular componems of
the esophagus underlie the motor reflexes of this organ.
The experimental focus of this thesis was to investigate the role of muscle
contractiiity in the initiation of the distention-induced descendhg peristaltic reflex in the
smooth muscle portion of the opossum esophagus. Hence, a description of the hctional
anatomy and control mechanisms of esophageai motor fùnction, as wel as the cellular
events leading to smooth muscle contraction, is presented in this section.
1) Esophageai Anatomy and Physiology -The Fundamentais
Hisoiogp
The esophageal waii, like other regions of the gastrointestinal (GI) tract, is
composed of three distinct layers: the mucosa. submucosa, and musnilaris propria.
1

Unlike the remainder ofthe GI tract, thae is no serosal layer to the esophagus. Instead.
the esophagus is covered by a loose layer of co~mective tissue d e d the adventitia, which
blends into the surrounding tissue (Netter 1971). The mucosa is dniided histologically
into three layers: an epithelial 1-g (stratidied squamous epithelium in aU regions of the
esophagus except the LES, where both squamous and wlumnar epithelium may coexist), a
supporting wnnective tissue lamina propria, and a thin smooth muscle layer, the
muscularis muwsae, which produces local movements and foldhg of the mucosa (Al
Yassin and Toner 1977, DeNardi and RiddeIl 199 1). in the lamina propria of the region
near the stornach are groups of glands d e d esophaged cardiac glands that secrete mucus
(Fenogiio-Preiser et 01. 1989). The mucosal layer of the esophagus has no absorptive
bct ion, but plays a major protective role against refluxed stomach acid and other
swallowed agents. The submucosa comprises a dense network of comective tissue within
which are blood vessels, lymphatic channels, esophageal glands, and a nerve plexus cdled
the submucous or Meissner's plexus. The muscularis propna, the muscular wall proper,
consists of muscle which is divided into two layers. The inner circular muscle (CM) layer,
with fibers oriented dong the circderence of the tube, serves to constrict the esophageal
lumen, and the outer longitudinal muscle (LM) layer, with fibers oriented dong its axis,
serves to shorten the esophagus (Lerche 1950). The combined action of these smooth
muscle layers, opposed at right angles to one another, is the bais of peristaltic
contraction. Located between the CM and LM layen lies a nerve plexus called the
myenteric or Auerbach's plexus (Fig. 1). Both the mynteeric and submucosal plexuses are
continuous around the cirderence of the esophageal wall (Sobotta and Hammersen
1985).

Auerbach's ple
Figue 1. Neuromuscular anatomy of the esopha-S. The imer circular muscle iayer. uith fibers oriented along the circumference of the tube, serves to connrict the esoghageal lumén and the outer longitudinal muscle. with fibres oriented alone its axis. serves to shonen the esophaps. Located between the CM and LAM layers lies the -4uerbach.s or myentenc plexus. Meissner's or submucous plexus is found in the submucosa. (From KafinIas 1992.)

From a iùnctional perspective the esophagus can be dMded into three sections, the
E S , esophageal body, and LES
Uppr Esophageui Sphincter
The esophagus begins at the UES, a purely striated muscle structure, which
consists predomulantfy of the cricopharyngeus and caudal fiben of the inferior pharyngeal
constrictor muscles. CM fibers fiom the p r o d esophagus also conm'bute to the
sphincter (Netter 197 1, Asoh and Goyal 1978, Welch et al- 1979). The UES is
functiondy denned as a zone of high in tra lha l pressure that separates the pharynx
from the upper esophagus. ui humans, the axiai Iength of the UES, as measured by
intralurninal manometry, is between 2 and 4 cm (Goyal et al. 1970, Ellis 197 1).
Esophcrgeal Body
in the adult human, th e length of th e esophageal body varies with trunk length, and
on average is 23 cm (Li et al. 1994). It extends fiom the Iowa border of the UES to the
upper border of the LES. While in the resting state, the esophagus is coilapsed. In
humans, it is capable of distending up to 2 cm anteroposteriorly and 3 cm lateraliy to
accommodate the ingesteci bolus (Long and Orlando 1998). In the proXimai one-quarter
to one-third of the esophagus, the muscle is striated in both the h e r CM layer and the
outer LM layers of the muscdaris propria. Striated muscle begins to be replaceci by
smooth muscle about 4 cm below the UES. This marks the beginnllig of the transition
zone (Meyer et al. 1986). The transition zone is of variable length; striated muscle and
smooth muscle fibers continue to be UiteTmingied up to 10-13 cm below the
4

cncopharyngeus. The distal one-halfto one-third of the esophageal body is composed
entirely of smooth muscle in both layers (Meyer et ol. 1 986). Occasionally, single bundles
of b t e d muscle extend within the smooth muscle aii the way to the caudal end of the
esophageal body (Arey and Tremaine 1933).
b e r Esophageal Sphincrr
The LES is located at the junction between the esophagus and the stomach
(gastroesophageal junction). It can be identined with intraluminal manometry as a 2-4 cm
zone of high pressure. The LES is composed of smooth muscle in moa species, including
humans, dogs, cats and opossums. Anatomic evidence of a sphincter has been ciifficuit to
obtain. The moa detailed description of the muSCUIar anatomy of the human sphincter
was given by Liebermann-Meffert and colleagues (1979). They found an asymmetric
thickening of the CM in the terminal esophagus, just above the angle of His (the junction
of the tubular esophagus and the saccular stomach). In experimental aaimals, the
relationship between anatomic structure and function has been more precisely studied.
Sunilar thickening of circular, but not longitudinal, muscle has been observed in the
opossum and cat (Seelig and Goyai 1978, Bimcani et al. 1982). In the opossum,
ultrastructural studies have demonstrateci several merences between LES muscle celis
and those of the esophageal body. These différences may help to explain some of the
mechanisms responsible for the bctional Werences between LES and esophageai body
contradity. Muscle ceUs of the LES are larger in diameter and fom fewer gap junctions
(Daniel and Posey-Daniel 1984). However, neuronal cell bodies in the LES are smaller
than those in the esophageai body, perhaps due to the tonically wntracted state of the

sphincter (Sengupta et al. 1987). Mitochondnal and smooth endoplasniic retidwn mass
is greater in the LES than in the esophageal body (Christensen and Roberts 1983).
Morphologically, the LES can also be disthguished fiom the esophageal body by the
presence of numerous intermusaiar spaces containhg comective tissue and blood vessels
(Seelig and Cmyal 1978).
Esophageai Motor Physiology
The major tiinction of the esophagus is to propel food and fluid hto the stomach.
This is accomplished by way of sequential or cbperistaltic" contractions of the esophageal
body in concert with an appropriately timed relaxation of the UES and LES. The
esophagus also clears any reflwced gastric contents back into the stomach and takes part in
mch reflex activities as vomithg and belching. N o d peristalsis is categorized into
either prirnary or secondary peristalsis, depending on its mode of initiation.
P n m q PeristaIlsis
Rimary peristalsis in the esophagus is a coordinated motor pattern initiated by the
act of swallowing. Continuous neuronal discharge of vagal lower motor neurons to the
UES mates a high pressure mne through tonic contraction of the UES muscles (Asoh
and Goyal 1978, Yoshida et al. 198 1). During swdowing, this newonai discharge
temporarily ceases aiiowing relaxation of the UES to occur (Van Overbeck et al. 1985).
UES opening also requires the elevation and anterior displacement of the larynx. which is
mediated by contraction of the suprahyoid muscles (McComeI 1988, Cook et al. 1989).
Relaxation of the UES lasts for one second and is folowed by a postreiaxation contraction

(Kahrilas et 02- 1992). Mer the botus passes through the UES, the UES closes and
retums to its tonically contracted state, and a rapidy progressive CM contraction
proceeds distaiiy through the esophagus starting in the upper esophagus and moWig
towards the LES. Each location dong the esophageal a i s matracts with a latency that
increases graddy f?om the upper esophagus to the LES. In the upper third of the
esophagus, contractions occurs Within 1-2s after swafiowing whereas in the lower third
this latency inmeases to betwecm 5-8s (Biancani and Behar 1995).
There is a coordinated contraction of the LM and CM layers of the smooth muscle
esophagus d u ~ g peristafsis, though each layer conm%utes to the process differently. CM
contraction produces the lumen-occluding propulsive wave front, whereas LM contraction
is associated with esophageal shortening. Sugarbaker and wlleagues (1 984) directly
recorded local contractile and electrical activity of opossum CM and LM in vivo. They
found a sequentid contraction of both layers in response to swaliowing. With vagal
stimulation, simultaneous LM short ening and sequential CM contractions were observed.
The LM contraction in each segment of the smooth muscle esophagus preceded and
outlastecl the CM contraction. Since LM contraction has the effect of shortening the
segment, this temporal arrangement wuld be expected to increase the CM ceil density at
the site of CM contraction, hence augmenting the amplitude of the propulsive contraction
(Sugarbaker et al. 1984). The LM contraction is associated with membrane
depolarization, whereas CM contraction is accompanied by a bimodal membrane potential
response -an initiai hyperpolarization (inhi'bition) foilowed by depoiarization The LES
relaxes during swaüowing and thus dows the aboraliy directed contraction wave to pass

the bolus fkom the esophagus to the stomach. Mer passage of the bolus through the LES
into the stomach, the LES closes with a prolonged contraction
Motility studies with dry swaiiows reveal that contractions in the striated muscle
segment are shorter in duration (1-2s) than those in the smooth muscle segment (4-7s).
The velocity of the peristaitic wave is 3 cm/s in the upper esophagus, accelerates to 5 cm/s
in the mid-region, and then slows again to 2.5 cmk just above the LES (Humphries and
Casteil 1977). Though the force of contractions can Vary from segment to segment and
even fkom swallow to swallow, contractions in the distai one third of the esophagus are
also usually stronger (50-1 50 mm Hg) tban those in the upper one third (40-123 mm Hg)
(J3iancani et al. 1975). However, contractions in the proximai and distal esophagus are
stronger than those in the rniddle third (20-80 mm Hg), perhaps because of the transition
between striated and smooth muscle esophagus in this region.
The physical characteristics of the ingested bolus also affect the speed and force of
esophageai phasic contractions. Increasing the size of the bolus tends to slow its
propagation velocity while increasing the force of contractions (Janssens et al. 1973,
Hollis 1975). An increased viscosity also increases the force of contractions and reduces
the speed of penstaisis (Dodds et al. 1973, Dooley et al. 1988). Bolus temperature cm
also modulate esophageal peristalsis; warm boluses increase peristaltic velocity, and cold
boluses d u c e it (Wimhip et ai. 1970, Decarle et al. 1 977).
Secomby Peridlsis
Distedon-induced or secondary peristalsis is the physiological mechanism
whereby refluxed gastric contents or food lefi behind d e r primary peristalsis are propeiied
8

into the stomach. Distention-induced peristalsis is triggered by stimulation of sensory
receptors in the esophageal body.
Bailoon distention (BD) is a commoniy used method to induce secondary
peristalsis under experimental conditions. Several studies in both animais and humans
have utilized intraiuminaI BD to characterize the esophageai responses to distention.
Localized transient BD of the smooth muscle esophagus in vivo, as weU as the
extrinsically denewated in vitro smooth muscle esophagus has been shown to produce
t h e separate responses (Christensen and Lund 1969, Christensen 1970). The dation of
the distending baiioon produces a brief localized contraction that occurs at and just
p r o d to the rostral rnargin of the distending bailoon -the "on" response. During the
maintenance of distention, there is shortening of the esophagus, due to longitudinal muscle
contraction, that is sustained for the fiill duration of the penod of distention -the
"duration" response. Whiie distendeci, the circular smooth muscle esophagus below the
BD site remains inhiited because of smooth muscle membrane hyperpolarization.
Balloon deflation causes a bnef caudal circumferential contraction -the "off' response *
(Christensen and Lund 1969). Thus the esophageal response to imralwRinal distention
consists of p r o d excitation and distal inhiition, which serves to ensure aboral
propulsion of the intrduminaI bolus.
Early studies suggested that the esophageal response to swallowing resembled that
of baiIoon distention (Craemer and Schlegel 1957, Fieshier et al. 1959). However more
recent quantitative work by Paterson and colieagues (1 99 la) has demonstrated that
contrary to earier reports secondary peristalsis differs sigdicantly fkom prirnary
peristalsis. The peristaitic wave velocity in the midesophagus is taster in BD-induced

peristalsis than m primary pe&tdsk. Fiirthemiore, at dl &es aboral to a disteading
balloon, evoked contraaions are of lower q W e tha. the swallow-mduced response,
whereas at sites orad to the bdoon, contraction amplitudes are of grerater ampütude when
induced by BD compared wah d o w i n g .
II) Overinew of the Control of Esophageal Motor Function
Innervation of the Esophagus
The wntrol r n e c u of esophageal paisralsis include those of the CNS, ENS,
and myogenic properties of the esophageal musculature. The esophageai innervation
provides the mechanism for the excitation and inhibition of the muscle at all these Ievels
and also provides vital sensory feedback via afEerent pathways which play miportant roles
in reflex modulation of the esophageal body and sphincter motor activities.
Swallowing Centre
CNS control of esophageal motor fiinction resides in the swdowiug centre - a
group of interconnected brainstern nuclei that receive cranial as weîi as peripheral inputs.
It is composed of two intimately co~~nected halfcentm and is located m the m e d a and
pons (Jean 1984). The d o w i n g centre is linked to the vomithg and respiratory centres.
Primary peristalns can be triggered either voIuntady, via higher brain centres, or
reflexively, by stimulation of peripheral afkents. Stimulation of the swallowing centre
activates vagai e f f i nerves to the oropharyngeal and the esophageal muscle. The vagal
nerves mediate primary peristalsis m the striateci and snooth mus& segments of the
esophagus. ConceptuaUy, the d o * centre has three fùnctional components: an

afferent reception system, a compiar orgenipiig qstem of neutons. and an efferent system
of motor neurons @iamant 1989).
i) Afferent Input
Mixent information fiom the periphery enta h o the solitary tract, the afferent
reception portion of the swdowing centre. This sensory idormation serves to initiate the
swaliowhg sequence, as weii as to aiter previously iaitiated 4 0 w i n g centre activity and
therefore modifL ongoing motor aaMty. The sensory information fiom the oropharyngeal
ana as well as other information invohred with this aspect of the d o w i n g mecbanism
enters the solitary tract through vagal nerve pathways and extravagal craniai m e s
(trigemïd, niciai, hypoglossal, and glossopharyngeal) @oty 1968, Carpenter 1989).
Sensory mformation fiom the esophageal body and sphincters is carried in the vagus
nerve, with the ceU bodies located in the nodose gangiion This semry pathway phys an
miportant role in monitoring and modulating esophageal motor activity. There is evidence
that sensory mformation is also carrieci by the sympathetic nemes to the spinal cord
segments C 1 to L3 (Sengupta et al. 1990, C o h et al. 1 99 1). This pathway is believed
to mediate nociception
ii) Coordinatrnatrng Region
The nucleus tractus solitarii and the neighboring reticulat substance d e up the
portion of the swdowing centre thaî is considered responsible for the progranminig of the
entire SwaUowïng sequence (Jean 1984). One level ofintegration (dorsal) wahin this
centre is mvohed m both the initiation and organization of the swaüowing sequence. A
11

second level of orgaiiization (ventral) appears to act as a connection pathway to the
various motor neuron pools Uivohred in the swdowing act, hc1uding the integration of
the deglutition reflex with actRrity in other medullary centces, such as the respiratory
centre.
ïhe motor neurons involveci in the swdowing sequence lie mainly in the
trigeminai, facial and hypogiossal nuclei, the nucleus ambiguus (NA) of the vagus (for
esophageal striated muscle), and the dorsal motor nucleus of the vagus (DMNV) (for
smooth muscle esophagus) (Fryscak et al. 1984, Carpater 1989).
Extrinsic Innervation
i) M o m lnnelyatratron
The primary extrinsic innervation of the esophagus is via the vagus nerve. Axons
of the lower motomeurons that supply pharyngeal muscles travel via the vagus into the
niperior pharyngeai nerves, which arise fiom the vagal tninks at the level of the nodose
ganglion. These nerves form the pharyngeal plexus, which innemates pharyngeal muscles,
including the cricopharyngeus muscle and the upper portion of the esophagus (Goyal and
Paterson 1989).
Motor fibers to the striated muscle esophagus aise nom the vagus, originating in
the upper portion of the neck. Hence, electrical stimulation of the vagus nerve in the
midportion of the neck produces no response in the striatecl muscle esophagus. Lower
motomeurons are myeünated and make direct contact with the striated muscle fibers

through the motor end plate (Toyama et al. 1 975). Acetylcholuie (ACh) is the
neuromuisminer (NT) invohred at the motor end plate, exerting its effécts through
stimulation of nicotinic chohergic receptors on the striated muscle (Toyama et al. 1975).
The vagus nerves also carry preganglïonic fibers to the srnooth rmiscle portion of
the esophagus (including the LES) where they branch out to fonn the esophageai plexus,
entering the esophagus at different leveis (Peden et al. 1950). Preganglionic fibers within
the esophageal waU travel for severd centimeta before reaching the postganpiionic
neurons in the intfamurai plexus, where they release ACh that activates muscarinic and
nicotinic recepton of myentenc nerve cells (Kravitz et al. 1978).
Sympathetic efferent pathways play a minor role in esophageal motor hction
compared to vagal (parasympathetic) efferents. The smooth muscle esophagus and LES
receive sympathetic nerve innervation that arises fiom the ce11 bodies in the
intermediolateral ce11 columns of spinal segments T4L2 (Collman et al. 1992).
Preganglionic fibers enter the cervical sympathetic gangiia, ganglia in the thoracic
sympathetic chain, and possibly the cetiac ganglia. The majority of the fibers to the lower
esophagus travel via the greater splanchnic nerves to enter the ceiiac ganglia where they
synapse with postganglionic neurons (Baumgarten and Lange 1969). The postgangiionic
branches accompany blood vessels, and a few fibers join the vagus to reach the esophagus.
The majority of these postganglionic axons terminate in the myenteric plexus and
submucosai plexus (Foumet et al. 1979). In the srnooth muscle esophagus, sympathetic
nerves affect muscle contractility mainly through modulation of other neurons (Seelig and
Goyal 1978). The density of adrenergic innemation varies in the esophagus and is lower
in the LES than in the more proximal esophagus (Baumgarten and Lange 1969).

ii) S e m t y Imentonon
In the digestive tract, sensory receptors are divided into three groups: mucosal,
muscle and serosai receptors (Leek 1977, Clerc and Mei 1983). This approach is based
on the findings that receptor endings within and outside the gut w d is largely responsible
for detennining receptor characteristics. Several modalities of sensory receptors are found
in the GI tract, including mechanoreceptors, chemoreceptors (acid receptors,
osmoreceptors) and themoreceptors. Through these receptors, information regarding the
physical and chernical environment within the lumen is available to the CNS via afferent
idow, and so is idormation regarding the amplitude and wavefom of every contraction
that occurs (Gnindy 1993). This in ni enables the various motor and SecTetory activities
to be adjusted to the specific needs of the individual.
The activation of tension-sensitive mechanoreceptors in the esophagus evokes
spike discharges in vagal (parasympathetic) and sympathetic afferent axons. Vagal
sensory pathways have been demonstrated in the rat (Andrew 1956), cat (Mei 1970) and
more recently in the opossum (Sengupta et al. 1989). A series of experiments aimed at
investigating stimulus/response characteristics of both vagal and spinal aferents supplying
the smooth muscle portion of the esophagus were recently performed in the opossum. In
these studieq single-unit vagal afferent actMty was recorded in response to distention in
the smooth muscle portion of the esophagus. Wee types of aifiirent fibres were
disthguished based on their responses to hcreasing levels of bdoon distention (BD).
These were low threshold, high threshold and wide-dynamic range mechanoreceptors
(Sengupta et al. 1990). Vagal afEerents were low threshold while hi@ threshold and wide
dynamic range affierents projected to the spinal cord. Sengupta and wileagues concludeci

that spinal afferents signal noxious wents white vagai fibres operate in the physiological
m g e to monitor peristalsis and act as a trigger for secondary peristalsis (Sengupta et al.
1989, Sengupta et al. 1990).
Three types of themoreceptors have been hctionaiiy i d d e d in the dista1
esophagus and gasnic fûndus and corpus of the cat based on electrophysiological
recordings made fiom vagal afEerent units with electrodes implanted in the nodose
ganglion (El-- and Mei 1982). These include w m receptors which respond only
to temperatures between 30°C and 50°C, cold receptors which respond between 10°C and
35°C and mixeci receptors which respond to temperatures in both ranges. These
themoreceptors were not sensitive to mechanical stimulation. Stimulation of wann
receptors inhibiteci nerve-induced contractions in the proximal esophagus, whiie
stimulation of cold receptors enhanceci them. Thus, esophageal themoreceptors can
modulate esophageal rnotor function.
Though numerous physiologîcal -dies have demonstrated that throughout the gut
there are neural receptors that cm respond to specific mechanid, themial and chernicd - stimulation, considerable controversy exists as to the anatomy of the specific neural
structures that represent these receptors.
In the striated musculature ofthe dog esophagus, Secfions stained with toluidine
blue and silver demonstrated the presence of muscle spindles in both the muscularis
propria and the intermuscular space of the distal third of the esophagus (Asaad et al.
1983). These richly innemateci spindles contained betwea five to seven intraftsai fibers.
In somatic striated muscle, muscle spindes act as mechanoreceptive sensors, and hence it

is assumed that these structures pedorm a similar hct ion in the striated esophagus. They
are, however, not present in smooth muscle.
Structures thought to be mechanoreceptors have been identifieci in both the
striated and smooth muscle portions of the esophagus. These structures have been caiied
intraganglionic laminar newe endings (IGLEs) by Rodrigo and wlleagues (1975). The ceil
bodies that give rise to vagal esophageal affirent fibers are located in the nodose ganglion,
and a number of these are immunoreactive for calcitonin gene-related peptide (CGRP) or
substance P (SP) (Rodrigo et al. 1982, Rodrigo et ai. 1985, Green and Dockray 1987).
The IGLEs in ganglia in the p r o d esophagus arise from fibers distributecl through the
superior laryngeal nerve, whereas those in the ganglia of the distal esophagus are the
endings of fibers that traverse the thoracic vagus. IGLEs found in the middle of the
esophagus originate from fibers traveling through both pathways (Christensen 1984).
Intrinsic Innervation
Enrenc Nervms System
In humans, the ENS is estimated to contain nearly 100 million neurons (Fumess
and Costa 1987). This system controls motility, exocrine and endocrine secretions and the
microcirdation of the GI tract. It is also involved in regdating h u n e and infiammatory
processes (Goyal and Hirano 1996).
There are two major networks of nerve fibers that are intrùisic to the gut: the
myenteric (Auerbach's ) plexus and the submucous (Meissner's) plexus. The myenteric
plexus provides primarily motor innemation to the circuiar and longitudinal muscle layen
and is involved in the control of peristaltic activity. The submucous plexus contains

secretory newons that play a crucial role in exocrine and endocrine ceil secretory wntrol
in the gut (Costa and Brookes 1994). Sensory neurons that respond to stretch, tonicity
and temperature are also found in the enteric plexuses.
The basic structure of the ENS is consistent throughout the GI tract. It is a
crissscrssing network of nerve fiber bundles with neurons grouped hto gangiia at moa
intersections (Christensen el d. 1983, Christensen 1993). The nerve budes contain the
axons of both intrinsic (intramural) neurons and extrhsic &irent and efEerent fiers
(Singaram and Sengupta 1996). Since only a smd percentage of nerve endings
degenerate after extrinsic denervation of the gut, the large majority of nerve fibers in this
layer are of intrinsic origin. Esophageal intramural neurons are fewer in number and more
loosely ananged than elsewhere in the gut (Christensen and Robinson 1982, Christensen el
. al. 1987, Sengupta et al. 1987).
The myentenc plexus is f o n d in both the striated and the smooth muscle portions
of the esophagus, though it is less developed in the striated muscle esophagus (Christensen
and Robinson 1982). However, since striated muscle fibers receive direct motor input
fiom extrinsic neurons, the function of the plexus here is unclear. It is possible that entenc
neurons of this region promote sensory output or innemate esophageal glands (Whitmore
1983). The submucosa3 plexus of the esophagus is exceedingiy sparse. In the cat it is only
a loose network of nerve fibers, devoid of ganglion cells; however, a few ganglion ceUs
are present in the submucous plexus of the opossum (Christensen and Rick 1985).
There are two important types of effector newons within the myenteric plexus of
the smooth muscle esophagus. One is capable of mediating cholinergie excitation of bath
circula. and longitudinal layers of smooth muscle through M2 muscarinic rewptors in the

esophageal body and M3 receptors in the LES (Gilbert and Dodds 1986, Goyal 1989,
Sohn et al. 1993), whüe the other is capable of mediating nonadrenergic nonchobergic
(NANC) inhiion mainly in the CM layer (Crist et al. 1984, Gilbert and Dodds 1986).
Cholinergie excitation o f the excitatory neurons is nicotinic, whereas that of the inbibitory
neuron can be muscarinic (MI) as weli. Histochemid studies have demonstrated the
existence of intrinsic cholinergic neurons in the esophagus (Seeiig et al. 1984).
AcetyIcholine is the major excitatory transmitter releaseà by postgangiionic enteric
neurons during peristalsis. The NANC neurotransmitter responsible for the initial
inhibitory response and the subsequent rebound excitation and contraction in the circular
smooth muscle layer of the esophagus is nimc oxide (NO) (Anand and Paterson 1994, Ny
et al. 1995).
The terminal motor innemation to the srnooth muscle of the esophagus arises
fiom ce1 bodies in the myentenc ganglia, making them postganglionic nerves. Motor
axons to the muscle layers travel fiom the ganglia in srnd nerve bundles that penetrate the
muscle layers (Furness and Costa 1986). These nerve axons contact smooth muscle celis
difEerently fiom the motor end-plates found in the striated muscle. Once they penetrate
the muscle layer, axons arborize, spreading between and among the bundies of smooth
muscle. The axon terminais are studded with characteristic swehgs called varicosities.
It is via these varicosities that axons of the intramurai neurons synapse with effector cells.
In the ENS, these variwsities serve as nodules containing packets of
neurotrafl~mitters, which are stored as vesicles. Several vesîcle types can be found in the
same nerve varicosity, irnplying that one neuron can release multiple types of transmitters
(Burnstock 1982). Interactions between presynaptic membrane proteins and those of the

synaptic vesicle are essential for organbhg the exocytotic apparatus and for eaabiing
neurotransmitter release in both the CNS and PNS.
Another group of ceus, the interstitial cells of Cajal (ICCs), are probably of major
hctionai importance. The interstitial ce11 was discovered 18% by Cajai, who suggested
that it was intercaiated anatomidy and bctionaliy between autonornic nerves and
effector cds. ICCs are mononuclear polymorphic ceus tint branch out extensively, the
branches intersecting to fom a neîwork throughout which the axons extend. These
branches form gap junctions with one another and with smooth muscle ceUs and newe
varicosities (Christensen 1993). ICCs bear a certain ultrastructural resemblance to both
smooth muscle ceUs and fibroblasts, suggesting that they have a mesenchymal origln.
However, some observers suggest that ICCs may also be neural or glial in nature
(Christensen 1993). The distribution of ICCs varies throughout the GI tract. In the
opossum esophagus, ICCs are unifomily distniuted in the thickness of the circular smooth
muscle layer, but are progressively more numerous in a craniocaudd distribution dong the
smooth muscle esophagus. Furthemore, ICCs are absent in the LM layer and relatively
sparse in the MP (Christensen et al. 1987a). However, in hurnans and cats, ICCs are
found in both the circular and longitudùial layers of the esophageal smooth muxle
(Faussone-Pelle* 1987, Faussone-Pellegrini and Cortiesini 1987). The finding that
ICCs have gap junction contacts to smooth muscle and are closely associateci with nerves
is consistent with the view that ICCs are intercalated between the nerves and muscles and
serve to modulate neuromuscular responses (Irnainimi and Hama 1969, Daniel and Posey-
Danie1 1984).

Control of Esophageal PeristilsY
Autonomie control of esophageal peristalsis hvolves a combination of centrai
(SaNisic) and penpheral ( i i c ) neuronal mechanisms using an array of putative
transmitter substances, as weii as myogenic W o n , varying regionaIly within the organ
because of variations in muscular composition Both neurogenic and myogenic models
have been proposed to explain the control of esophageai peristalsis.
Central Control
i ) Sm-ated M d e Esophagrrs
As discussed previously, the striateci muscle esophageai motor fiuiction is under
the direct control of sequentidy firing vagal fibers programmeci by the swallowing centre.
Both primary and secundary peristaisis in the striated muscle esophagus are abolished after
extrinsic denervation (Carveth et al. 1962). Other types of esophageal responses in the
stnated muscle portion also appear to depend on central wntrol. These include the
inhibition evoked by frequent swailows, and reverse peristalsis observed in ruminants
@oty 1968, Stevens and Sellers 1968).
îi) Smooth Muscle Esophogus
The neuromudar mechanisms responsible for peristalsis in the smooth muscle
esophagus are dl not completely understood. It is weU established that swailow-induced
primary peristalsis is dependent on the activation of the swaiiowing centre and the va@
pathways to the esophagus. This is demonstrated by the fact that bilateral cervical
vagotomy or vagal cooling abolishes primary peristalsis in the esophagus (Ryan et al.
20

1977, Reynolds et ai. 1984). Uniilce the striatecl muscle esophagus, sequential fking of
vagai aerent neurons destined for progressive1y more caudal smooth muscle esophageal
segments is not essentiai to activate peristdsis. Though bilateral vagotomy aboiishes
esophageal motor activity in the esophagus, includuig the distai ssnooth muscle portion in
both the ait and monkey, with the, recovery of some aspects of motor bction takes
place (Binder et al. 1968, Burgess et al. 1972). Moreover, vagal efferent stimulation of
the distai stump of the severed vagus, which activates a i l vagal fibres ~Unultaneously,
results in sequentiai, rather than simultaneous, contraction of the smooth muscle portion
(Dodds et al. 1978% Dodds et al. 1978b).
Experiments by Gidda and Goyal (1 984) recorded swailow-evoked potentials fiom
single ceMcal vagaf efferent fibres of the opossum and disthguished two types of
preganglionic efferent fibres destined for the smooth muscle esophagus based on their
latency distributions. Short-latency fibres began firing within 1s of the onset of
swalloving, as recorded by myohyoid activity. Long-latency fibres had latencies to onset
of discharge that ranged between 1 and 5s. The respective latency distributions suggest 4
that short-latency discharges correlate with the initial inhibition, and the long-latency fibers
correlate with the p e n d t i c contractions.
Taken together, these results suggest a crucial role for the swaiiowing centre in the
initiation of peristalsis in the smooth muscle portion of the esophagus, but not necessarily
for its organization into a progressing contraction wave fiont. This role of the swaiiowing
centre is f.urtIler demonstrated by the fact that swdlows induced in close succession inhibit
or forestall anticipateci contractions in the smooth and striated muscle segments (Dodds et
al. 1976). For example, wmplete propagation of the peristdtic contraction takes place

der a swaiiow if a second swaliow does not occur within at least 10 seconds after the
first one. However, &er repeated swallows a - shorter intemals, no contractions are
observeci in the entire esophagus until &er the last swallow, which is foiiowed by a high-
amplitude contraction. The inhibition of peristaisis by repeated swaiiows is refmed to as
degiutitive inhtiition (Meyer et al. 198 1).
Peripberal Control
The peripheral mechanisrns involved in controlling peristdsis of the smooth muscle
esophagus have been rwealed by studying the responses to 1) intralumioal bailoon
distention in the extrînsically denervated esophagus, 2) elecaical vagal efferent nerve
stimulation, and 3) eledcal stimulation of intramural nerves in CM and LM strips of
esophaged smooth muscle. Two proposed models of peripheral control are the neural and
myogenic models.
i') Intrinsic N a r d Connuf
Several lines of evidence converge to indicate that local enteric circuitry controls
progressive CM contraction in the smooth muscle esophagus. Christensen and Lund
(1 969) demonstrated that in vitro preparations of opossum esophagus responded to
distention and low ftequency EFS with abordy propagated contractions which were
qualitatively similar to swailow-induceci peristdsis. T'IX abolished the EFS-induced
response demonstrating that such stimulation selectively excites intramufal enteric nerves
and intact vagal temiinals.

Investigations using reduced preparations such as muscle strip preparations and
intraceiluiar recording fkom singe ceils have demonstrateci that smooth muscle saips
exhibit characteristic contrade and membrane potential responses. In generai, LM strips
respond to EFS with contractions which last throughout the stimulation period, whereas
CM strips or rings respond with Vanable irïtra-stimulus and post stimulus contractions
foUowing a latency period (Eslami et al. 1994). Weisbrodt and Christensen examined the
Muence of short train duration (0.25s) EFS on the contrade response of CM strips.
They found that CM strips responded by contraction &er the end of the stimulus. This
response was hence narned the ccOff' response. Crist and colleagues (1984a) demonstrated
that when esophageai CM strips are stimulated with long train stimuli (5- 1 Os), both intra-
stimulus and post stimulus contractions occur. These responses were termeci as "'on7' and
"'off" contractions respectively. Both on- and off-contractions are neurogenic, because
they are abolished by TTX. The delay fkom the cessation of the stimulus to the onset of
the "ofl" contraction was different among different levels of the esophagus. The delay
hcreased progressively down the esophagus. This suggested that local neurornuscular
mechanisms in each regional esophageai segment had built-in mechanisms that could
potentiaily produce peristaltic-like contractions without influence ffom neighbouring
segment S.
Sidtaneous electrical activation of di vagal eEerent nerves to the esophagus
produces a delayed peristaltic contraction, rather than an immediate simultaneous
contraction, indicating that intrinsic neural mechanisms can mediate the peristaltic
response. Work by Gidda and Goyal (1985) has demonstrated that esophageal
contraction in response to va@ efkent stimulation is preceded by an active inhibition

that begins promptly with the onset ofthe vagal sbmulatim (id inhibition). The
duration of this initial preceding the contraction inaeases distaDy dong the
esophagus and corresponds to the laîency of esophageal contraction to vagal stllmilation.
This initial &Vition is associated with hyjmpolarization ofthe CM (Rattan et al. 1983).
Two types of ciradar smooth muscle contraction have aiso been dernonstrate.
using long train (8s) vagal efferent stimulation in the anesthetized opossum (Dodds et al.
1978). " A type? contractions occurred between 1-6s fier stimulus onset, depending on
the esophageai level they were recording fiom "B type" contractions occuned
approximately 1s after cessation of the stimulus. A and B waves diffa fforn each other in
several ways. A waves are more fiequently evoked by low-frequency stimulation and
have an apparent propagation velocity that is sirnilar to that of swallow-induced penstalsis.
B contractions occur more fkequently at high-fiequency stimulation and have a much more
rapid propagation velocity that is sirnilar to that of the post-stimulus off contraction
recorded £tom opossum circular smooth muscle strips. Both A and B wave contractions
are abolished by TTX and are therefore neurogenic. A waves are chohergic in nature as
they are abolished by atropine, whereas B waves are atropine-resistant and hence are
noncholinergic.
It is generdy accepted that the on-contractions seen in vitro are similar to the A
waves of vagai stimuiation, since both are inhiiited by atropine and have prominent
aborally increasing latency gradients dong the esophagus. OfFantractions in the CM
strips in vitro are siimiar to B contractions evoked by vagal stimulation in vivo, in that
both are atropine resistant and the latency gradient is minimal.

The cholinergic iduence on esophageal motor fitnction is most prominent in cats
and humans. In these species, swallow-induceci peristalsis is atropine-sensitive (Dodds et
al. 198 1, Paterson et al. 199 1) and augmentesi by cholinergic agonists and cholinesterase
inhibitors (Hofis and Casteii 1976). Similar to these models, the cholinergic on response
mechanimi is more prominem proximally and decreases distally in the opossum (Crist et
al. 1984b), whereas the NANC off contraction is more prominent distally (Gilbert and
Dodds 1986). However, the NANC mechanism predomiaates in the opossum. Taken
together, these data suggest that esophageai peristalsis is a result of '0lended" activation
of cholinergic and NANC contractions.
Esophageal pexistalsis is characterimi by a gradient of inaeasing latency of
contractions dong the length of esophagus as one proceeds distally. This latency gradient
is the bais for smooth muscle esophageai peristalsis. IntramUral neurons release a NANC
neurotransmitter (NT) that causes initial hyperpolarization and inhibition of esophageal
smooth muscle. Several studies have demonstrated that NO is the NANC inhibitory NT in
the gut (Yamato et al. 1992). NO is produced nom Larginine by the enzyme NO-
synthase (NOS), which is present in myentenc neurons (Kontuek and Konturek 1995).
Furthemore, histological staining for NADPH diaphorase, which colocaiizes with NOS,
indicated that approximately half of the myenteric ganglion cells in the diaal human
esophagus are nitrergic (Singaram et al. 1994). NOS can be reversibly inhiiited by
analogs of L-arginine, such as N-nitro-L-arginine methyl ester (L-NAME) and N-nitro-L-
arginine (LNNA). In the opossum, these NOS iahibitors have been show to: 1) decrease
swaiiow-induced contraction amplitudes in the distai esophagus, and when combined with
atropine, to abolish primary peristalsis; 2) dimuiish swdow-induced contraction latemies,

especially in the distal esophagus; 3) aboiïsh vagal-stimulation-induced, end-of-stimulus B
contractions; and 4) inhi'bit membrane hyperpolarization and the subsequent NANC
depolarization induced by BD, swdowing and vagal-stimuiation (Murray et al. 199 1,
Yamata et al. 1992, Anand and Paterson 1994). Taken together, these studies
demonstrate that NO is the endogemus NT that is responsi'ble for NANC
neurotransmission in the body of the opossum esophagus, which involves initial inhiition
of the srnooth musde foliowed by ''rebound" excitation.
Several other neuropeptides that have been shown to affect gastrointestinai
motility have dso been identifieci in esophagd neural tissues. These include vasoactive
intestinal peptide (VIP), substance P (SP), calcitonin-gene related peptide (CGRP),
galanin (GAL), neuropeptide Y (N'Y), peptide histidine isoleucine (PHI) and enkephalins
(Christensen et al. 1987b, Wattchow et al. 1987, Christensen et al. 1989, Biancani et al-
1989, Singaram et d 1994). Many of these have clear-cut eEects on esophageal body
and LES contractiiity when used in a phannacologicai setting; however, th& physiological
sigiuficance is unclear.
zi) In-c Myogenic Control
The basis for myogenic control of phasic contractil@ in the GI tract is maRifest by
two fùndamental characteristics: (1) the electrical control actMty, an omnipresent
oscillation of membrane potential C'slow wave") that controls smooth muscle excitabiiity;
and (2) the abilîty of the smooth muscle ceus to communicate with each other (coupling)
in such a way that the entire tissue cm operate as a fimctionai unit.

Esophageal contractions occur when the membrane potentiai depolarizes above its
excitation threshold. Unlike other GI smooth muscle, however, esophageai smooth
muscle is electrically quiescent (Le. no eleceical control aaMty or slow wave) and does
not contract spontan8ousIy (Crist et al. 11.987). Rezent studies in muscle stxip and whole
organ in vitro preparations have demonstrated that a latent myogenic oscïiiatov
mechanisns for control of phasic contractions exists in esophageal smooth muscle and that
it may be activated by nonspecific excitation of the smooth muscle membrane (Helm et al.
i 99 1, Helm et al. 1 992).
The most direct evidence that the mechanisrn for generation of phasic contractions
Lies within the mooth muscle itselfis the finding that in in vin0 esophageal preparations
treated with TTX, the smooth muscle esophagus is capable of generating propulsive
movemems in response ro direct muscle stimulation using electrical square waves of long
pulse duration ( S m a et al. 1977). The myogenic mechanism of tbis propagation is
believed to operate partiaily through ekctrotonic spread of activation by gap junctions
present between smooth muscle cells. 4
Gap junctions are symmetrical patches of the membrane of two cells, occupied by
special intracellula. channels, that facilitate the movement of intracellular regdatory
molecules and ions ffom cell to ceil (Perachia 1980). When an action potentid occurs
across the membrane of a single-unit smooth muscle, it is rapidiy propagated via these gap
junctions to the entire group of interconnected muscle celis that function electncally and
mechanically as a hctional syncytium.
The mechanimis involved in the integration of the myogenic and neural control
systems of peridaisis in the smooth muscle esophagus have yet to be detemineci. The role

of intramufal inhiiitory nemes under this mode1 could be to inhibit retrograde peristalsis,
mediate descending iahriion in advance of the peristaitic wave, and to modulate the
velocity of myogenic propagation of contractions (Hehn et al. 1992). Furthmore, ifthe
ICCs senie as transducers for nerve to muscle signaling, and as conduction paths for
muscle to muscle communication, then they are iikely integral components of any
myogenic control system.
zia) Signal T r d c t i o n Pathways in Esophageal Circulm Muscle
Smooth muscle is categorîzed into muIti- and single-unit types, based on
Merences in how the muscle fibers become excited. Most smooth muscle, including that
found in the GI tract, is of the single-unit variety and is also referred to as viscerd smooth
muscle. Muscle fibers that make up this type of muscle become excited and contract as a
single unit. Excitation of smooth muscle celis results in an increase of f i e cytosolic
calcium. This increase is due to influx of extraceiluiar ca2' into the cytosol via L-type
voltage-gated calcium channels, ligand gated ca2' channels or f?om release of calcium in
the cytosol fiom intracelldar stores. Calcium release initiates a cascade of intracellular
events which leads to force generation through cross-bndge cycling between the
contractile füarnents -a& and myosin (reviewed by Walsh et al. 1996). As the calcium
concentration increases in srnooth muscle, more cross bridges are formed resuiting in
greater tension development.
The myogenic mechanisnu respomible for esophageal smooth muscle contraction
in the esophageal body are different from those responsible for LES tone. The smooth
muscle of the esophageal body is reiaxed at rest and contracts in response to vagal

stimulation or field stimulation with a bnef and forcefui phasic contraction However, the
LES exhibits spontaneous tone and myoel&cal actRrity which is characterized by
wntinuous spike burst activity with or without the presence of phasic contractions. LES
tone is maintaineci following neural blockade with tetrodotoxin, indicating its' myogenic
origin (Goyal and Rattan 1976). Current data suggest that esophageal body and LES CM
utilUe distinct ca2' sources, phospholipid pools, and signal transduction pathways to
contract in response to ACh (Hülemeier et al 1991, Sohn et al. 1994, Biancani et al.
1997).
The intracelldar mechanisms of contraction of erophaged body muscle in response
to ACh or nerve stimulation has been extensively studied in the cat model. Unlike feline
LES muscle which utilizes primady intracelidar sources of ca2' for tonic contractions7
. phasic contractions of the esophageal body circular smooth muscle utilize primarily extra-
cellular calcium (Biancani et al. 1987). According to the model put forward by Biancani
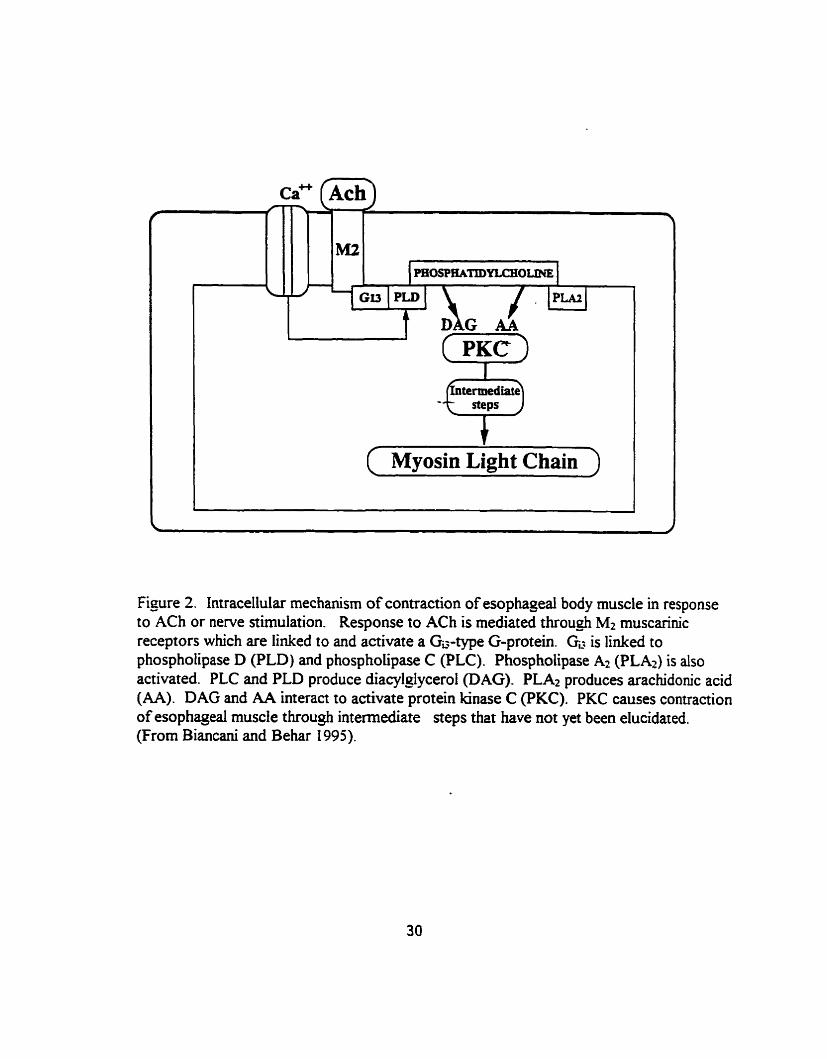
and colleagues (1997), response to ACh is mediated through Mt muscarhic receptors
which are linked to and activate a Ga-type G-protein (Fig. 2). Gd is linked to
phospholipase D (PLD) and phospholipase C (PLC). Phosphoiipase Az (PLAI) is also
activatecl, but the types of muscarinic receptors and G-proteins responsible for PLA2
activation are not yet known. PLC and PLD produce diacylglycerol @AG). PLA2
produces arachidonic acid (AA). DAG and AA interact to activate protein kinase C
(PKC). DAG-induced activation of PKC is ca2'-independent suggesting that the influx of
ca2' is r e q W ody to activate the phospholipases involved in DAG production. PKC
causes contraction of esophageai muscle through intermediate steps that have not yet been
elucidated (Biancani et ai. 1997).
t
termediate -:7 ( Myosin Light Chain )
Figure 2. Intracellular mechanism of contraction of esophageal body muscle in response to ACh or nerve stimulation. Response to ACh is mediated throu& M2 muscarhic receptors which are linked to and activate a GG-type G-protein. GL. is linked to phospholipase D (PLD) and phospholipase C (PLC). Phospholipase A2 (PLA*) is also activated. PLC and PLD produce diacykJycerol @AG). PLA2 produces arachidonic acid (AA). DAG and AA interact to activate protein kinase C (PKC). PKC causes contraction of esophageal muscle through intemediate seps that have not yet been elucidated. (From Biancani and Behar 1995).

UnWre the esophageal body, spontaneous LES tone depends on spontaneous, low-
level activity of phospholipase C, resulting in the production of submrurimal concentrations
of IP3 and DAG (Biancani et aï. 1994). In permeabüized LES muscle ceus, submaxïmal
doses of Il?, and DAG act synergistically, and their interaction depends on ca2* release
and is mediated through a PKC-dependent pathway resuiting in submaximal tonic
contraction. In the cat, it has been shown that LES contraction is mediated by the calcium-
dependent PKC-P II isofom, whereas contraction of the esophageal body is mediated by
calcium-independent PKCI isofonn (Sohn et al. 1997).
Integration of Centrai and Penpheral Control Mechanisrns
It is clear that centrai and peripheral levels of control act in a highly wordinated
fashion to regulate peristalsis. Studies to date have not established how this integration
occurs or which level of control is no rd ly dominant during esophageai peristalsis. In
primary peristaisis, initiation of the reflex and coordination of the upper striated
musculature is under direct central control. The fundamental role of peripheral control
mechanisms in esophageai motor finction is demonstrated by the fact that diaention-
induced peristalsis is always propagated in the caudal direction after extrinsic denervation
with cervical vagotomy and even in the isolated esophagus rnounted in an organ bath.
Hence, the direction and velocity of the peristaitic contraction appear to depend on the
balance between excitatory and inhibitory neural influences dong the esophagus. The
mechanism whereby central discharge integrat es with the peripheral neuromuscuiar control
mechanisms in the smooth muscle esophagus has not yet been cladied. If the nnal
propagation of the peristaltic wave also involves a myogenic contribution, the neural
31

mechanisms, either extrinsic or imrinsic, would need tu provide convolleci activation of
the muscle in such a way as to pennit passage of a single coatraction aboraîiy dong the
esophagus. How muscle properties contribute to peristaisis directeci by either central or
intramurai neural mechanisms is not yet clear. These multiple levels of control make it
possible to conceive of local intramUral mechanisms for penstalsis operating independently
if centrai mechanisms are defective or absent.
III) The Distention-Induced Descending Peristaltic Reflex
Distention of the intestine evokes a aereotyped reflex in which CM contracts on
the oral side of the stimulus and relaxes on the anal side (Bayliss and Starling 1899). This
reflex has been most extensively studied in the guinea-pig srnaII intestine and rat colon.
The neuronal circuitry responsible for the reflex is large& contained within the rnyentenc
plexus (Smith et al. 1990) and includes sensory neurons, interneurons, and both excitatory
and Uihibitory motor neurons (Fmess and Costa 1987, Miftakhov and Wmgate 1996).
The efferent motor iimb of the penstaltic reflex evoked by stretch appiied to
circular muscle consists of orad (ascending) contraction, accompanied by excitatory
junction potentials (EJPs), and caudad (descending) relaxation, accumpanied by inhibtory
junction potentials (IJPs) (Costa and Fumess 1976, Grida and Makhlouf 1986, Smith et
al. 1990, Yuan et ai. 1991). The components reflect release of the excitatory
neurotrammitters ACh, the tackykinins SP and neurokinin 4 and the inbibitory
neurotransmitters VIP and NO (Grider and Makhlouf 1986, Grider 1993). Release of
these transminers is regulated by cholinergie and noncholinergic internewons coupled to
sensory neurons (Tonini and Costa 1990, Grider 1993).

In vitro studies using Bat-sheet prepdons to study the co~eIation between
mechanical and electrical responses of the paistaItic r d e x show that the reflex can be
evoked by mucosal stroking or muscle stretch. However, the reflex evoked by muscle
stretch is not affècted by the removai of the mucosa, implying that mucosal stimulation
and muscle stretch activate distinct populations of sensory neurons (*el 1979, Smith et
al. 1990). Measurernents of E P s and LJPs niggest that the reflex evoked by mucosal
stimulation and muscle stretch is mediated by the same population of enteric motor
neurons (Smith and Fumess 1988, Smith et al. 1991, Yuan et al. 199 1). Hence, the
sensory neurons activated by muscle stretch and mucosal stimulation are distinct,
converging on the sarne motor neurons (Bomstein et al. 199 1, Smith et al. 1992).
The distention-induced peristaltic reflex response in the smooth muscle esophagus
has also been extensively studied, because it provides an opportunity to explore the
intrinsic neuromuscdar mechanism responsible for peristalsis. Similar to the smd
intestine, the esophageal response to intrduminal distention consias of proximal excitation
and diaal inhibition, which serves to ennire aboral propulsions of the ùitduminai bplus. 4
However, unlike the smaü intestine, the proximal excitation reflex in the esophagus is
mediated by the vagus nerve through muscarinic-cholinergic transmission. BD studies by
Paterson (1991) in the anesthetized opossum demonstrateci that contractions proximal to
the BD site were abolished by bilateral cervical vagotomy. When the BD was carrieci out
in the mid-esophagus, proxhnd contractions were abolished by atropine. With BD in the
distal esophagus, the evoked proximai contractions were less atropine sensitive. In
contrast to the excitation observed proximal to a distendhg balioon, the response diaal to
the BD site consists of inhibition that is foiiowed upon balloon deflation by the "off'

contraction These distal respollses persist after bilaterai cervical vagotomy and are
minimdly & i e d by atropine (Paterson et al. 1988, Paterson 1989), but are abolished by
blockade of NO synthase (Anand and Paterson 1994, Ny et al. 1995).
E a r k studies by Paterson (1989) examlned the electrical correlates of esoplugeal
peristalsis using suction electrode and intduminai pressure recordings eoxn CM at sites 1
and 5 cm above the LES. Unlike the 5 cm site, where initial hyperpolarization in response
to BD was foiiowed upon balloon defiation by prompt membrane depolarization, spike
bursts and muscle contraction, the BD-related membrane hypexpolarization at the 1 cm
site was foilowed by a subthreshold depoiarization and another hyperpolaritation-
depolarization sequence. The characteristic spike burst and esophageal CM contraction
occur with this second depolarization and thus the contraction at the 1-cm site is markedly
deiayed relative to the 5-cm site. The proposed explamtion for this phenornena is that
proximal contraction might reactivate intramwai inhibitory neurons just as the diaai site is
recove~g f?om the initiai hyperpolarization, thus causing either a suppression of the
subsequent depolarkation or a further hyperpolarization-depolarization sequence. These
findings suggest that esophageal contractions per se can activate descending inhibition.
Recent studies (Paterson and Indrakrishnan 1995) were carried out to characterize
the intramural neural pathways involved in the distention-induced descendhg peristaltic
reflex in the opossum smooth muscle esophagus. Prior to beginning these studies it was
hypothesized that three possible pathways (Fig. 3a) could explain the observations made
to date: Mode1 1) the mechanoreceptor that responds to distention wuld send multiple
processes aborally that synapse with and release NO directly ont0 the circular smooth
muscle of the esophageal body (Le. the mechanosensitive neuron has a combined sensory

Mode1 t
Model 2
Model 3
Mechanoreceptor 4 Combined Sensory-Motor Neuron
Mononeuronal
Polysynaptic
Smooth Muscle Cell
Recording Cham ber
Oral Intermediate Aboral
F i p r e 3. (A) Schematic representation of possible intramural neural pathways responsible for dinention-induced descendin3 penaaltic reflex in the opossum esophagus. Model 1) combined sensory-motor neuron: the mechanoreceptor that responds to diaention could send multiple processes aborally that synapse with and release NO direct- onto the circular smooth muscle of the esophaseai body; Model2) mononeuronal pathway: the mechanoreceptor cell activated by distention excites a descending inhibitory motomeuron, which in mm releases NO ont0 the circular smooth muscle; Model 3) polysynaptic pathway: the mechanoreceptor activates an interneuron t hat, via a polysynaptic pathway, Ieads to activation of the NO neurons. (B) Schematic representation of the triple-charnber organ bath set-up which pennitted different sections of the smooth muscle esophagus to be chemically isoiated, while keeping the neuromuscular apparatus intact (Adapted fiom Paterson and Indrakrishnan 1995).

and motor function); Model 2) the mechanoreceptor ceil activaîed by distention excites a
descendhg inhibitory motomeuron, which in tum releases NO ont6 the circular smwth
muscle; and Model 3) the mechanoreceptor activates an intemeuron that, via a
polysynaptic pathway, l a d s to activation of the NO neurons (i.e. similar to descending
peristaltic rdex in the intestine). ïhey used a mple chamber organ bath which permitteci
different sections of the smooth muscle esophagus to be chemically isolated, wMe keeping
the neuromdar appanuus intact (Fig. 3b). BD in the oral chamber evoked membrane
hyperpolarization in the abord chamber, which was folîowed, upon baüoon deflation, by
depolarkation, spike burst, and circula snooth muscle contraction. Addition of TTX to
any of the chambers abolished this reflw indicating its neurogenic nature. In these
studies, it was found that synaptic blockade using O ca2+-20 m M M ~ ~ ' Krebs abolished the
BD-induced abord c'off' contractions when placed in either the abord or oral chamber,
but not when placed in the intermediate chamber. These results demonstrated that the
neural pathways mediating the distention-induced descending peristaltic reflex in the
opossum esophagus involve long descending intramural neurons. This contrasts with the
small intestine where most entenc reflex pathways are polysynaptic and include
interneurons which connect to other intemewons fomiing ascending and descending
chains (Fmess et al. 1993, Costa and Brookes 1994). The NOS inhibitor L-NAME,
when placed in the aboral chamber, abolished this reflex, indicating that NO is the final
mediator. The results obtained using O ca2*-20 m . MC Krebs solution to block synaptic
transnission at the ongin of the reflex are l e s clear-cut. Though O ca"-20 rnM ~ g "
Krebs signincantly attenuated the reflex (suggesting that synaptic transmission is occurring
at the origin of the reflex), none of the putative NT antagonists tested were able to

significantly alter the response. This inchideci addition to the oral chamber of antagonists
of the sensory neurotransmitters SP and CGRP.
Thesis Objective
Though O ca2'-20 mM M~*+ Krebs inhibited the distention-induced descendhg
peristaltic reflex, the mechanism by which it exerted its e f f i is not clear. One
explanation proposed is that a synapse does exist between a distention-sensitive
mechanoreceptor and the long descending intramd neurons, though it involves a NT not
yet tested for. However, stimulation of sensory neurons with capsaicin or addition of the
putative sensory NT substance P or CGRP to the oral chamber fded to eiicit the
descendhg peristaltic reflex in this preparation (Paterson and Indrakrishnan 1995). In
excitable ceiis, ca2+ influx via voltage dependent calcium channels mediates a variety of
functions including synaptic transmission between neurons and excitation-contraction
couphg in muscles. Hence, influx of extracellular Ca 26 into smooth muscle ceiis is
necessary for smooth muscle contraction to occu. 1s the inhibitory effect of
O ca2'-20 mM ~ g * ' Krebs in the oral chamber really due to the attenuation of synaptic
transmission at the origin of the reflex, or couid it be due to decreased muscle tension
development in response to stretch ? In other words could the distention-induad
descending peristaltic reflex actually be initiateci by muscle contraction in response to
stretch, rather than by the stretch itself?
Recent investigations by Brookes and coiieagues (1997) were aimed at i d e n m g
the mechanisrns underlying the initiation ofperistdsis in the guinea-pig ileum. In these in
Mbo expeximents, segments of intestine were radially stretched using a microprocessor-
37

controIIed stepper motor with an in-series force transducer to record CM contraction.
W~th addition to this preparation of agents which reduce the a<&ity of smooth muscle,
an increase in the threshold stretch r@ed to initiate peristalsis was obsewed. Addition
of agents which selectively increase the excitabiiity of CM r d t e d in a reduction in
threshold stretch required for peristaltic initiation. Furthemore, these arperiments
demoll~trafed that when the ileum is close to threshold stretch, peristalsis can be triggered
simply by Uicreasing circular muscle tension. These results suggest that the initiation of
peristaisis rnay be due to initial contraction of CM activatiag neuronal circuitry via
contraction-sensitive mechanoreceptors (Brookes et ai. 1997).
The aim of this thesis was to examine how the addition of O ca2' -20 mM M ~ ~ *
Krebs to the oral chamber inhibits the BD-induced descending peristaltic reflex in the
opossum smooth muscle esophagus. The working hypothais wu that locai smooth
muscle contractility in response to BD is necessary to initiate the distention-induced
descending peristaltic reflex, and that addition of O ~ a % mM MC Krebs
solution to the oral chamber inhibited this locai musde contractüity. This thesis
investigated the role of smooth muscle contractility in the initiation of the distention-
induced descending peristaltic reflex.

Chapter 2
Methods
Rationale for Methods Used
Animal Models
Our m e n t understanding of esophageai rnotor fùnction is derived primarily fkom
animal studies. Due to species variation among different animal models, caution mus be
exercised when applying these observations to humans. The esophagus of humans, other
primates, and opossums is composed predominantly of smooth muscle in both layers
(Marklin et al. 1979). In contrast, dogs, rabbits, rats, guinea pigs and sheep have an
esophageal body composed completely of striated muscle. Striated muscle comprises only
the proximal third of the esophagus in primates and opossums. Cats also have a mixed
musculature esophagus; ..&wever only the distd few centirneters consia of smooth
muscle, making the opossum a better mode1 for studying long intramural reflexes within
the smooth muscle segment. In primates, cats and mamipials, there exias a cornplex
interplay of both centrai and intrinsic neural mechanisms, Zmilar to what is found in 4
humans. Nonetheless, differences in the balance between choluiergic excitatow and
NANC inhibitory Muences dong the esophagus exist between these animal models. The
CM off contraction is only rninimally sensitive to atropine in the opossum (Cnst et al.
1984) but is abolished in the cat (Biaacani et ol. 1989) and partially inhibiteci in humans
(Tottn~p et al. 1990). NOS inhiiitors abolish the off contraction in opossum (Yamato et
al. 1992) and human (Sugarbaker et al. 1992) esophageal CM stnps. In the opossum, the
cholinergie on-response mechanism is more prorninent proximaüy and decreases distaily,
whereas the NANC off contraction is more prorninent distally. Therefore, similar to the
39

humm, in the opossum, both intramd mecham'sms appear to contribute to peristalsis.
For these reasons, the North Amencan opossum, Di&@his vir@ziona. an extensively
studied animal mode4 was chosen for these investigations as it has an esophagus that is
both anatomically and physio1ogidly simiiar to that of humans.
Assessrnent of Esophageai Motor Fuoction
i ) Mcmometry
Intraluminal rnanometry is a fiequentiy employai quantitative technique which is
used in the clinical and basic research laboratory to study the physiology and
pharmacoIogy of the esophagus in both health and disease (Casteil and Casteil 1994). A
typical rnanometry set-up includes a pneumohydraulic pump powered by cornpressed
nitrogen, pressure transducers, amplifier, polygraph and a rnanometry catheter (Castell and
Dalton 1994). The rnanometry catheter consists of an assembly of soft, thin polyvinyl
tubes that have side or diaai openings. Bubble-fke distilleci water is constantly perfiised
through the catheter system. When placed in the lumen of the esophagus, occlusion of the
individual catheter tip (port) -either by a contracthg LES or a contractile wave in the
esophageal body- results in an increase in the pressure within the water-filied catheter
tubing. Since water is essentidy non-mmpressible, it transmits pressure changes weii.
As the esophageal wall CM contracts, a localized inaease in intrarnural pressure impedes
water flow, which results in measurable pressure changes in the assembly of the tube and
is an indication of peristaltic pressure. nie pressure recorded in the esophagus is due to a
direct squeeze by the esophageal muscle on the pressure-sensitive catheter tip (Goyal and
Paterson 1989). The displacement of the diaphragm in the pressure transducer is

converteci into an electricai signal, which is then amplified and recordeci by a cornputer
data acquisition system. The data coiiected include amplaude, duration and latency to
onset of esophageal cirnilat muscle contractions. From tbis information, the rate of
peristalsis can be caldated.
ii) Electronic Barostat
The most widely used technique to study the biomechanical and visco-elastic
propertïes of the GI tract in both the clinical and research setting is the isobaric
measurement method employing the electronic barostat. The barostat was initidy
developed for use in "tone studies" by Azpiroz and Malagelada (1985) at Mayo Clinic. It
consists of a cornpliant bag which is ~ ~ e c t e d to a pressure transducer module linked
through an automateci feed-back mechanism to an air injection-aspiration p m p driven by
an electric rnotor. Output of air tiom the injection pump to the barostat bag lads to
inflation of the bag to a pre-selected volume or pressure. When the barostat bag is placed
into the esophageal lumen, the barostat can respond to changes in intraesophageal bag
pressure by injecting or withdrawing air to maintain a pre-selected pressure. Hence, at a
selected distendhg pressure, any change in intraesophageal bag volume is indicative of a
change in the compliance of the esophageal w d . Thus, an increase in bag volume
indicates an increase in compliance, whereas a decrease in bag volume indicates a decrease
in compliance.

Phaumacoiogid Agents
As d i d earlier, though O Ca "-20 mM Mg *+ Krebs solution inhiiits the
distention-induced descending peristaltic rdex, it is not clear how it acts. Two potentid
e f f i s of this soIution at the site of distention are 1) the impediment of synaptic
transmission between mechanosensitive neurons and internewons andlor effector neurons
due to lack of srhacellular ca2* (Katz et d. 1995), and 2) the inhibition of wnooth muscle
contradty due to lack of extrace1Iuiar ca2+. To examine the role of smooth muscle
contractility in this reflex, two inhibitors of smooth muscle contraction which act via
different mechanisms were chosen. The dihydropyridine, nifédipine, was employed to
selectively block L-type ca2- channels which are responsible for infiwc of extfaceflular
ca2- in smooth muscle celis of the GI tract. Nikdipine inhibits both phasic and tonic
smooth muscle contractility in the esophagus (Blackwell et al. 1981) and midies have
demonstrated that nifedipine does not affect synaptic transmission in enteric neurons of the
guinea pig ileum (Barajas-Lopez et al. 1996, Izza et al. 1997). The bernophenanthridine
alkaIoid, chelerythrine chloride, is a potent inhibitor of PKC (Herbert et al. 1990), that has
been shown to inhibit phasic contraction in smooth CM ceUs taken from the feline
esophageal body and LES (Sohn et al. 1997). To examine the role of synaptic
transmission at the site of distention, the a-conotoxin GVIA fiom Comrs geo+grqhus
venom was chosen (Olivera et ai. 1990). This conotoxin selectively blocks N-type, but not
L-type calcium channels, and thus prevents the influx of extracellular CC h o neuronal
ceils (McCleskey et ai. 1987). In the opossum. it has been used as an inhibitor of synaptic
transmission in the LES (Saha et al. 1992). The p s e r fish poison, tevodotoxin (m, was employed to seiectiveiy inhibit ne~rotrans~ssion of imemated srnooth muscle
42

prqarations while not a f f i g smooth muscle conrractilicy (Gershon 1967). TTX
abolishes action potentials in nentes by prwenting the rapid entry of sodium through the
depolarued ceU membrane (Gersbon 1967). Niidpine and TTX were supplied by Sigma
Chernicals (St. Louis, MO). Chelerythrgie chloride was obtained fiom RB1 (Naticlq MA)
and M T X GVIA was supplied by Alomone Labs (Jerusalem, Isael).
Specific Rotocols
AU procedures were approved by the Queen's University Animal Care Committee.
F i opossums (Di&iphzs virgnzam) of either sex weighmg between 1.4 and 6.0 kg were
used in these studies.
Removd of Esophagus
The opossum was anaesthetized with a taii vein injection of 4Omgkg body weight
of pentobarbitai sodium (Somnotol, MTC Pharmaceuticais, Cana& Packers, Cambridge,
Canada). ï h e thoracic cavity and abdomen were opened dong the mid-line and the,length 6
of esophagus requUed was measured in situ, whereafter the entire smooth muscle
esophagus and part of the aomach was removed by thoracotomy and Iaparotomy. M e r
removal of the esophagus, the animal was euthanized with an intracardiac injection of
pentobarbital sodium. The esophagus was imrnediately placed in a dissecting dish
containing modified Krebs solution. The composition of the Krebs solution was the
fouowing (in mM): 1 15 NaCl, 4.75 KCi, 1 Na&PO4 1.2 MgSOc 25 NaHCQ, 2.5
CaCIz, and 11 glucose, dissolved in distilleci water. In all of the experiments presented in
this thesis, the Krebs solution was constantly bubbled with a gas mixture of 5%C& -
43

95%& and maintaineci at 34OC (Le. the physiological core body temperature of the
opossum) and pH between 7.2 and 7.4.
Dose Respoose Ciuves
To establish which concentration of drug to use in our in vitro experirnents, dose
response curves of both the L-type calcium chamel blocker nifedipine and the PKC-
inhibitor c h e l m e chloride were performed on circular smooth muscle esophageal
strips. The esophagus was splayed open with an incision dong its longitudinal axis and
pinned, mucosal d a c e up, to the silicone gel block. After visuaily locating the LES, the
mucosal layer was carefbîly rernoved by sharp dissection. Smooth muscle from the distal
esophagus was then cut into transverse strips (0.5 cm x 2.0 cm) taken fkom between 2 cm
and 6 cm above the LES. One end of each stnp was secured by silk thread to a hook
placed at the bottom of the recording chamber filied with oxygenated (95% 4 -5% COz)
Krebs solution. Silk thread was also used to attach the other end of the muscle sîrip to a
Harvard Isomemc Force Transducer (Mode1 50-7905, Edenbndge, Kentucky). The
muscle strips were placed under an initial tension of -5g and allowed to equiliirate for 60
minutes before recordings were obtained. Isometric changes in tension were measured via
a Harvard T r d u c e r Internice (Mode1 50-8861) and were displayed and recorded on a
Harvard Universai OsciIiograpb The selected agent was added to the muscle bath in
cumulatively increasing concentrations. Electncal Field Stimulation (EFS) was delivered
three separate times at each concentration using a Grass S88 (Grass Instniment Co.,
Quhcy, Mass.) stimdator through a stimulus isolation unit (SIUS) via two Pt-Ir.
electrodes embedded 10 mm apart in the wd of the recording chamber (Fig. 4). The EFS

MUSCLE
ISOMETRIC FORCE C
TRANSDUCER
I
MUSCLE 'Sm
RETORT STAND
Figure 4. Schernatic representation of muscle strip set-up. Excitation of the muscle strip with EFS (5 s train, 1 ms puise duration, 20 6OV-IOOV) re& in the phasic muscle contractions which are detected by the isometric force tninsducer and recorded on an oscilograph.

consisteci of a 5s train of stimulation at ktween 60V to 1ûûV (supramawnal voltage)
with a &equency of 20 Ht and puise duration of 1 rns. Previous studies have shown that
these parameters are optimal for stimuiating intramma1 neurons and eliciting the
neurogenic offresponse (Crist et al. 1984). The preparation was allowed to equilr'brate
for 5 minutes at each darerent drug concentration before EFS was commenced. The
inteml between two successive stimuli was maintained greater than 1 minute to prevent
over stimulation of the muscle strip.
The Descending Peristaitic Reflex: Double-Chambered Bath Experirnents
The double-chamber bath used was a modified version of that described by
Paterson and Indrakrishnan (1 995). It consisted of a small block of silicone and chambers
that were created using movable Plexiglas dividers (Fig. 5). The set-up consisted of an
oral chamber (10 ml in volume) and an abord chamber (1 50 ml in volume) which allowed
the proximal and distai segments of the esophagus to be chemicdy isolated while keeping
the neuromuscular apparatus intact.
The distal 8-10 cm length of opossum smooth muscle esophagus was threaded
horizontally through 8-mm holes in each Plexiglas divider. Inert silicone grease @ow
Coniing, Miciland, MI) was appiied around the esophagus-Plexiglas interface to create a
seal beîween chambers. The effdveness of this seal was tested by adding Trypan Blue
solution (Sigma Chemid Co. St. Louis, MO) to the oral chamber anà visuaiîy checking
for leakage of this dye into the aboral chamber. Experirnents ody proceeded if no leaicage
of dye was observeci. Previous studies in our laboratory (Paterson and Indrakrishnan
1995) employed hctional tests to c o n f b the effectiveness of these seals. Addition of

EFS PERFUSED MANOMETRY CATHETER
ORAL CHAMBER
ABORAL CHAMBER
Figure 5. Schematic representation of the double-chamber organ bath used to study the descendhg peristahic reflex Both transient intralumnial BD (-5 s) and EFS (3 s train, 1 ms puise duration, 10 Hz, 60 V) were performed m the oral chsunber. Esophageal contractile actMty was recorded fiom both the oral and aboral chambers with a mhindly perfused manometory catheter (0.15 d m h ) .

T ï X (1 0%) to one chamber did not &ect nerve-mediateci responses evoked in the
adjacent chamber.
Intraluminal manometry was @ormeci using a d e t e r systern that consisteci ofa
distending bdoon, and five polyvinyl recordhg tubes (0.8 mm inner diameter, 1.1 6 mm
outer diameter) that were glued together with tetrahydrof'wan (BDH Inc. Ontario). A
one-millimeter diameter side-hole orifice was cut in each of 4 recording tubes and the tube
distal to the o%ce was d e d , such that pressure recording ports were created at sites 1,
2,3, and 4 cm fiom the distal end of the baiioon. The fifth tube was used to infiate and
deflate the balloon. hiring pressure recordin& each catheter lumen was minimaliy
perfùsed with distilled water at a rate of 0.15 d m i n using a Mui Scientinc
pneurnohydraulic capillary infusion system (Mode1 PIP-3, Mui Enterprises Inc.,
Mississauga, ON). When secured in the chambered organ bath, intraluminal pressures
were recorded at a site 1 cm distal to the bdoon (i-e. oral chamber) and at sites 2,3 and 4
cm distal to the baiioon (i-e. aboral chamber). The catheters were connected to pressure
aansducers (Transpack II; Sorenson Research, Abbott, Salt Lake City, UT). AN data were
coUected by the WinDaq'200 data acquisition system (Dataq Insauments Inc., Akron,
Ohio) and stored on hard disk. A latex balioon, 2 cm in length, and made from a finger cot
(size medium 28030, Realmont LTD, Cowande, Quebec), was placed over top of the
catheter and then foldeù back on itseif The ends were sealed with sik suture and
Vetbond Tissue Adhesive No. 1469 (3M Ltd., St. Pad, MN). Wtth this set-up, the
bdoon produced cirderentiai distemion of the esophagus aromd the manometry tube
when inflated with air via a syringe.

EFS imrolved pashg 30s trains of square wave ment puises of 10 Hz fiequency,
1 ms puise duration and 60 V stimulus strength to the adventitial surfàce ofthe ewphageal
segment in the bath via a bipolar silver chloride stimulating eiectrode. These electrodes
were comected to a Grass Stidator (S88). To correlate esophageal contractile activity
with EF4, the EFS adfiict was mea~u~ed by two recording electrodes which were placed
in the oral chamber- Any response beginning d e r the termination of the stimulus was
defineci as an "off contraction The Krebs soIution in the abord c h b e r was changed
eveq 20-30 minutes throughout the duration of the protocol.
The protocol was as follows:
I After positioning the esophagus into the double chamber organ bath, a 20-25
minute stabilization period ensueci.
2. To estabiish baseline control responses for both the BD and EFS-induced
descendmg peristdtic reflex, transient 2cc BD (-5 s) or EFS was performed at
Ieaa three times each, with at least three minutes dowed to elapse between each
a stimulus.
3. Selected agents were then added to either the oral or aboral chamber. With
inhibitors of smooth muscle wntractiiity (nifidipine (1 o4 M), cheleythrine
chloride (104 M)) or O Ca "-20 mM M g *' Krebs, the agents were added to the
oral c h b e r and dowed 20 minutes to equilibrate with the tissue. The N-type
ca2+ channe1 antagonist, a-conotoxh GVIA (lad M), was added to either the oral
or aboral chamber, followed by a 20 minute equil'bration period.
4. Three post drug BD and EFS were perfonned.

5. To determine whether or not the actions of the agents were reversiile, the
preparation was washed several times with normal Krebs and allowed up to 1 hour
to remver.
Esophageai Cornpliance: Bmstat Studies
To M e r investigate the mechariism of action of these inhibitors of smooth
muscle contractility and neurotransmission, esophageal compliance was assessed using an
automated barostat syam (SVS Barostat, Synectics Viscerd Stirnulator Module). Data
acquisition and measurements were taken ushg Polygram for Wmdows V LOB6 and
stored on hard disk (Fig. 6).
A cornpliant 8 cm long polyethylene bag (maximum diameter of 30 mm, maximum
capacity 50 mi) was employed to measure in Mfro changes in esophageal compliance in
response to agents that affect smooth muscle contractility and neurotransniission. Prior
to each experiment, the intraesophageal bag was fkst tested for leaks by placing it in a
beaker mled with water and examinhg for changes in intra-bag volume or the presence of
air bubbles during a test run at pressures of up to 40 mm Hg (twice the pressure employed
in Our experimental protocol). The barostat flow rate was set at 15 d s . Before s t h g
the experiment protocol, the barostat was caiiibrated rnanually using a
sphygrnomanomet er.
The cornpliance of the bag ex vivo was measured to ensure reliabilif~ of our
measurements in the low-compliance portion of the curve. This confirmeci that with
operating volumes of less than 20 ml, the elastic remil of the bag did not contribute to the
resistance to inflation. This allowed us to be certain that any change in volume for a given

COMPUTER CONTROL SYSTEM
PRESSURE AIR TRANSDUCEFt RESERVOIR
PRESSURE LINE
Figure 6. Schematic representation of the electronic barostat set-up. To maintain pressure inside the system at a pre-seiected level, air is mjected d o or withdrawn fiom the mtraesophageal polyethykne bag through the inflation line. The bag is c o ~ e ~ t e d to the kostat by two catheters, one to monitor htrabag pressure and the other to move air in and out of the hag.

pressure change refïected primarily the esophageal wali properties and not those of the
distendhg polyethylene bag -7).
After the distal esophagus was removed nom the opossum, it was placed in an m
Mtro bath. The intraesophageal bag was insexted into the esophagus where it was secured
into place with a tie at both the prolomal and distal ends. The tissue was then left for a 20
minute stablzization period. The selected agent (dedipine (104 M), chelerythrine chloride
(1 o4 M), O Ca 2+ - 20 mM Mg *+ Krebs or tetrodotoxin (1 o4 M)) or control (normal Krebs
solution) was then added to the bath and the protocol commenceci. The inmiesophageai
bag was idated to the selected distending pressure for 30 seconds, followed by a 60
second deflation period. The distending pressures employed were in the foliowing order
0, 1,2,3,4,8, 12, 16,20 mm Hg. This distention cycle took twelve minutes and was
foUowed by a three minute rest period (Fig. 8). The protocol was repeated 4 thes an hour
for 2 hours. During bag distention, the pressure in the bag is maintained at a constant level
while changes in barostat bag volume are monitored. Hence, at a selected distending
pressure, any change in intraesophageal bag volume is indicative of a change in the
compliance of the esophageal wall. Thus, an increase in bag volume indicates an increase
in compliance, whereas a decrease in bag volume indicates a decrease in compliance.
Data Anaiysis
Dose Response Curves
The percent change in EFS-induced off contraction amplitude in response to
increasing concentrations of selected smooth muscle relaxants was assessed. Al1 changes
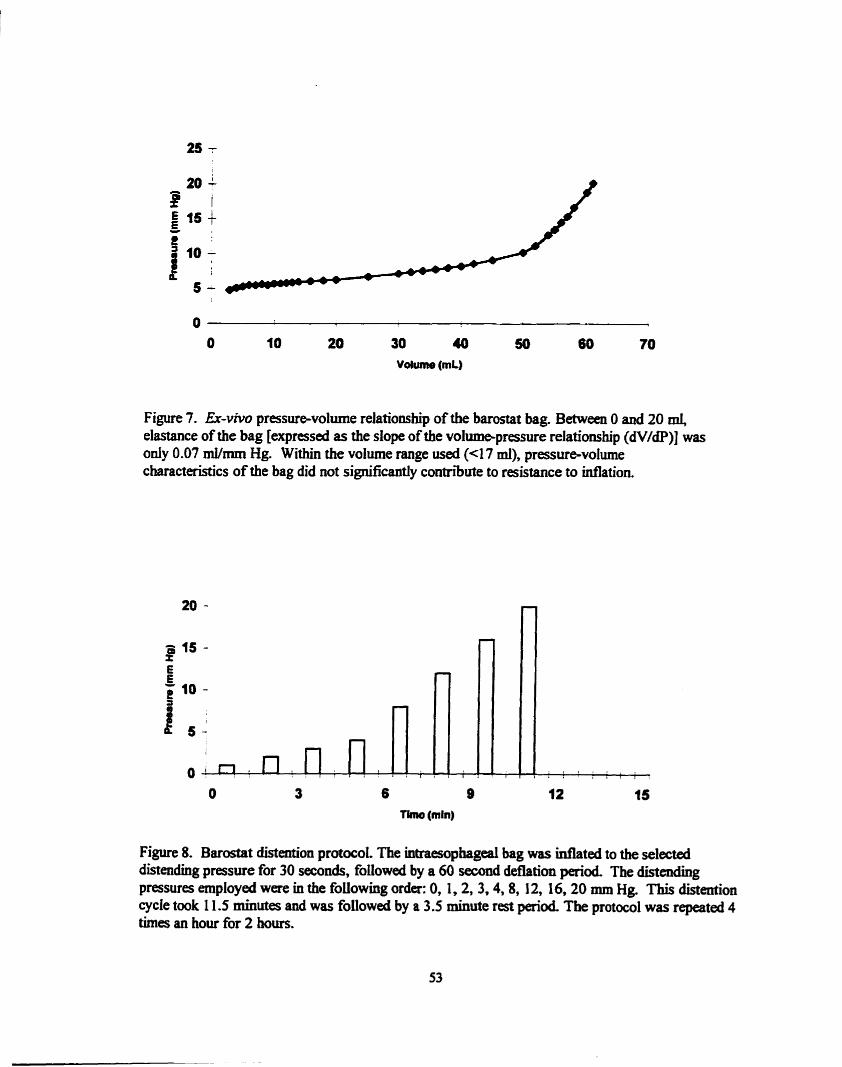
Figure 7. Er-viw pressuravolume reiationship of the barostat bag. Between O and 20 mi, elastance of the bag [expressal as the dope of the volumepressure relatioaship (dV/dP)] was only 0.07 mVmm Hg. Within the volume range used (4 7 ml), pressure-volume characteristics of the bag did not significantly contribute to raistance to inflation.
T h o (min)
Figure 8. Barostat distmtion protocol. The intraesophageal bag was inflated to the ~Iected distendhg pressure for 30 seconds, foiiowed by a 60 second deflation perid The distendiug pressures employed were in the foilowing orda: O, 1,2, 3.4,8, 12, 16.20 mm Hg. This distention cycle took 1 1.5 minutes and was foiiowed by a 3.5 minute r a t period The protocol was repeated 4 tirnes an hou for 2 hom.

are expressed with respect to the control dues of the tissue before the addition to the
chamber of selected agents.
Dacending Peristaltic Reflex Studies
The amplitude of the off response was assessed to determine the effect of selected
agents on the descending peristaltic reflex. B a d on data from previous studies (Patmon
and Indraknshnan 1995) and pilot studies, experiments only proceeded if the contraction
amplitudes of the control responses were greater than 10 mm Hg in both the oral and
abord chamber. To ensure representative results, at least three pre- and three post-
intervention observations were made in each animal. Responses at all recordlng sites were
analyzed in both chambers. For clarity, data presented fiom the abord chamber is f?om
the 3 cm site as this is representative of the 2 cm and 4 cm sites.
Barostat Studies
For each experimental condition a volume-pressure c w e was established and
analyzed using regression analysis. As described by Mayrand and Diamant (1 993), the
two aspects of tone that were assessed were 1) cornpliance of the esophageal wall,
expressed as the slope of the volume-pressure relationship (dV/dP), and 2) resistance of
the esophageal wall to initial stretch, which can be expressed as the x-intercept ofthe
volume-pressure graph. The equation of a linear hct ion cm be represented in the
foiiowing fom: y = A + Bx; B is the slope and A is the y-ùitercept. The x-axis intercept
(when y=G) is defined by the ratio -NB. The maximum volume recorded at the selected

distending pressure was used to plot the pressuevolume w e s in aii of the experiments
(Fig- 9).
Phasic contractions over top of the barostat bag can also be identifid with the
barostat. Phasic contractitity results in a rapid increase in the pre-set intraesophageal bag
pressure. The barostat is equipped with feedback sewors which act to osset pressure
variations by adjusting the volume of air injecteci into the intraesophageal bag. These real-
tirne barostat volume adjustments serve to maintain constant distending pressure on the
esophageal wall. A contraction index (CI) was dehed to quant* the phasic contractile
activity over top of the barostat bag during one complete distentioo cycle. The amplitude
of the contraction as measured by volume displacement of the barostat bag (ml) x the
duration of the contraction (s) (Fig. 9) is a measure of phasic contracblity. The CI is the
cumulative mm of the contractile activity for aii the selected distending pressures in one
distention cycle.
Statisticd Methods - In the double-chambered organ bath studies, cornparison of the magnitude of
contraction amplitudes before and &er addition of selected agents was carried out using a
two-tailed paired Student's t test. When signifiant changes were detected in both
chambers, the Mann-Whitney test was used to compare ciBiences in magnitude of
change between the 1 cm and 3 cm sites. In the barostat studies, the equations defining
the hear relationship between volume and pressure were calculateci by lin= regession
analysis using the least-squares principle (Microsofi Excel for Wmdows 95 Version 7).
Merences between treatrnent groups were determined by one-way analysis of variance

Figure 9. Baronat data collection. The maximai volume recorded during the entire penod of a selected dinending pressure (eg. 12 mm Hg) was used to plot the pressure-volume curves in al1 of the experirnents (mow). The amplitude of the contraction as measured by volume displacement of the baronat bag (mi) x the duration of the contraction (s) was a measure of phasic contractility. The contraction index (CI) is the cumulative nim of the phasic contrade activity for al1 the selected distendiq pressures in one entire diaention cycle.

(ANOVA). When significant variation was detected, the Student-Newman-Keuls post test
for multiple-sample cornparison was d e d out. A P value <O.OS was accepteci as
indicative of a statistidy sigmficam difference between two sets of observations. AU
values reporteci are means +/- standard error of the mean (SEM). AU staîisticai
calculations were done using the GraphPad Instat V. 2.04a statistical package (GraphPad
Software, CA).

Chapter 3
ResuIts
Dose Determination
CM strips nom the smooth muscle esophageal body did not develop spontaneous
tone, but responded with the characteristic post-stimulus off contractions following EFS.
Both nifidipine and chelerythrùie chloride causeci dose dependent inhibition of the off
contraction (Fig. 10). This inhibition appeared to be irreversible with nifiedipine but
partiaily revenible following chelerythrine chloride &er several washes with fkesh Krebs
solution. B a d on these studies, a concentration of lo4 M dedipine and chelerythrine
chloride was chosen for use in subsequent studies as this concentration caused near
maximal (>80%) inhibition of the off contraction.
Descending Peristaltic Reflex Studies
A typical response to ûnramural BD and EFS in the opossum smooth muscle
esophagus in Mtro is depicted in Fig. 1 la- 1 1 b. CM o f contractions are seen at sites distal
to the baiïoon foliowing balloon deflation or EFS. This reflex was usually reproducible for
several hours, although the magnitude of the response tended to graduaiiy decrease over
time. The addition of TTX (1 0%') to the oral chamber abolished both the BD- and EFS-
induced descendhg penstaltic refiex (n=5) (F4g. 12).
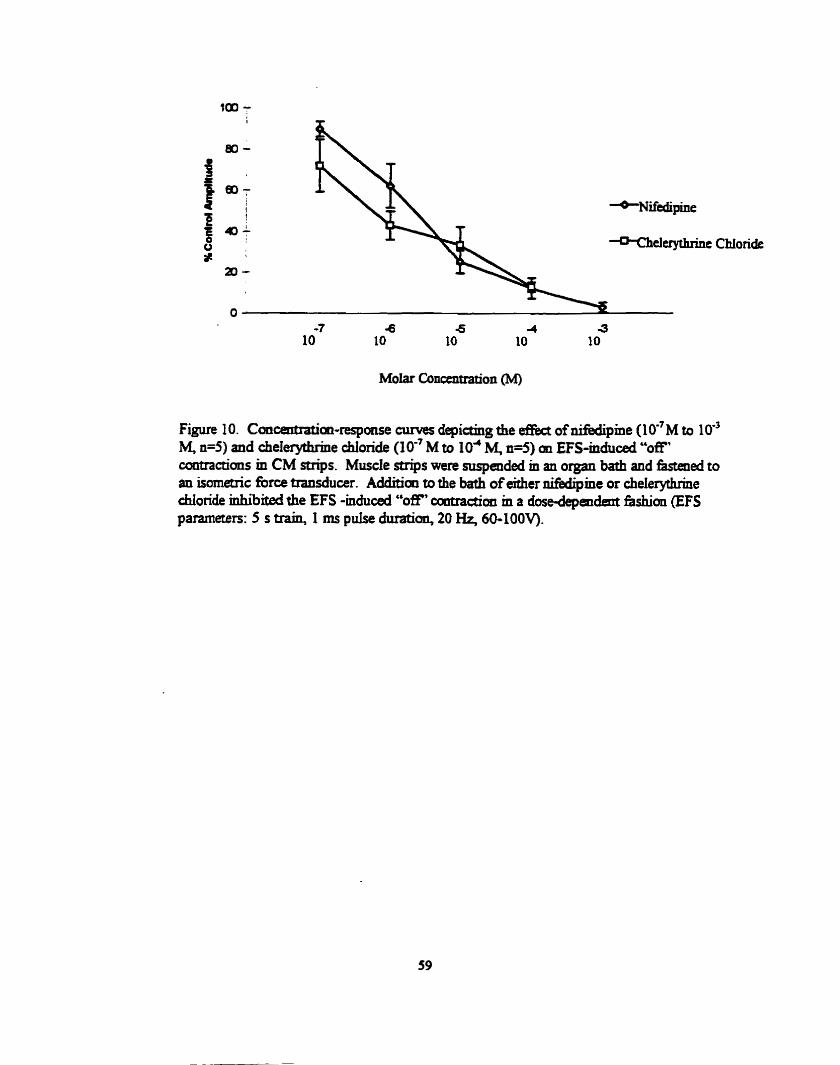
Figure 1 0. Concentmian-respans~ n i ~ s àepicthg the e f f i s of nifedpine (1 W' M to 1 v3 M, n=5) and cheleqduine chloride (IO-' M to 104 M, n=5) cm EFS-inducecl "OS' cmtraccicms in CM strips. Muscle mips were suspended in an ogan bath and fastaied to an isomerric force tmnducer. Addition to the bath of either niledipine or chelerythnne chloride mhi'bited the EFS -mduced "off' contnibim m a dosedepaident fishion (EFS parameten: 5 s trah, 1 ms puise durafian, 20 Hr, 60-100V).


A) Effect of Tetrodotoxin on B Wnduced Descendhg
h
Peristaltic Reflex Contraction Amplitudes CID s E 4)- E Y
8 30- D 3 I C
I Control - E a D - E :
m m Q: g 10 ' - C O L O - C o C) Oral
Chamber Aboral
Chamber
CI
Peristaitic Reflex Contraction Anplitudes m I
Figure 12. Addition of tetrodotoxin (lo4 M) to the oral chamber rnarkedly inhibiteci both the BD(A)- and EFS (B) -inducd off contraction in both the oral and aboral chambers. (n=5, two-tailed paired Student's t test, #P<0.05, *P<0.02, ** Pc0.0 1 vs. control observations).

Effect of Inhicbitors of Smooth Muscle Contraction
OEflect of O ~d+-20 m ~ ~ ' j - Krebs
Replacement of normal Krebs with O Ca *+-20 m M M g " Krebs solution (n=5) to
the oral chamber (Fig. 13A) markdy f i i i t e d the BD-induced off contractions in the oral
chamber (61.2 +/- 15.1 mm Hg vs. 2.0 +/- 1.2 mm Hg, M . 0 1 8 ) and abord chamber
(55.0 +/- 7.1 mm Hg vs. 10.8 +/- 6.8 mm Hg, W.014). Moreover, larger BD volumes of
3 cc were unable to initiate this inhibitecl reflex response. Though EFS-induced
contractions (n=5) were m k e d l y atteuuated in the oral chamber (39.6 +/- 9.5 mm Hg vs.
8.5 +/- 3.0 mm Hg; M.049), the contractions were not significantly inhriited in the
abord chamber (50.0 +/- 7.2 mm Hg vs. 32.8 +/- 18.7 mm Hg; P= 0.44) (Fig. 13B). The
inhibitory effect of O ~ 8 - 2 0 mM M ~ ~ ' Krebs solution was reversible, with post-wash
contraction amplitudes retuming to control levels ( ~ 5 ) .
i(lE#eccr of Nijedïpzne
Addition to the oral chamber of the L-type calcium channel blocker nifidipine (IO-< a
M; n=5) vimially abolished the BD-induced off contractions in both the oral chamber
(55.1 +/- 7.3 mm Hg vs. 0.4 +/- 0.2 mm Hg, W.0016) and aboral chamber (42.8 +/- 8.9
mm Hg vs. 1.7 +/- 0.9 mm Hg, W -0096) (Fig. 14A). Furthemore, larger BD volumes
(Le. 3 cc) were unable to initiate this Uihibited reflex response following Nfedipine. The
inhibitory effect of nifedipine was irreversible as the off response did not retum after
several washes of the tissue with fiesh Krebs solution. Wah EFS (n=5), the off
contractions were markedly inhi'bited in the oral chamber (50.9 +/- 5.1 mm Hg vs. 9.0 +/-
3.3 mm Hg, W-OOS), but d e BD, the EFS-indu& off contractions in the aboral
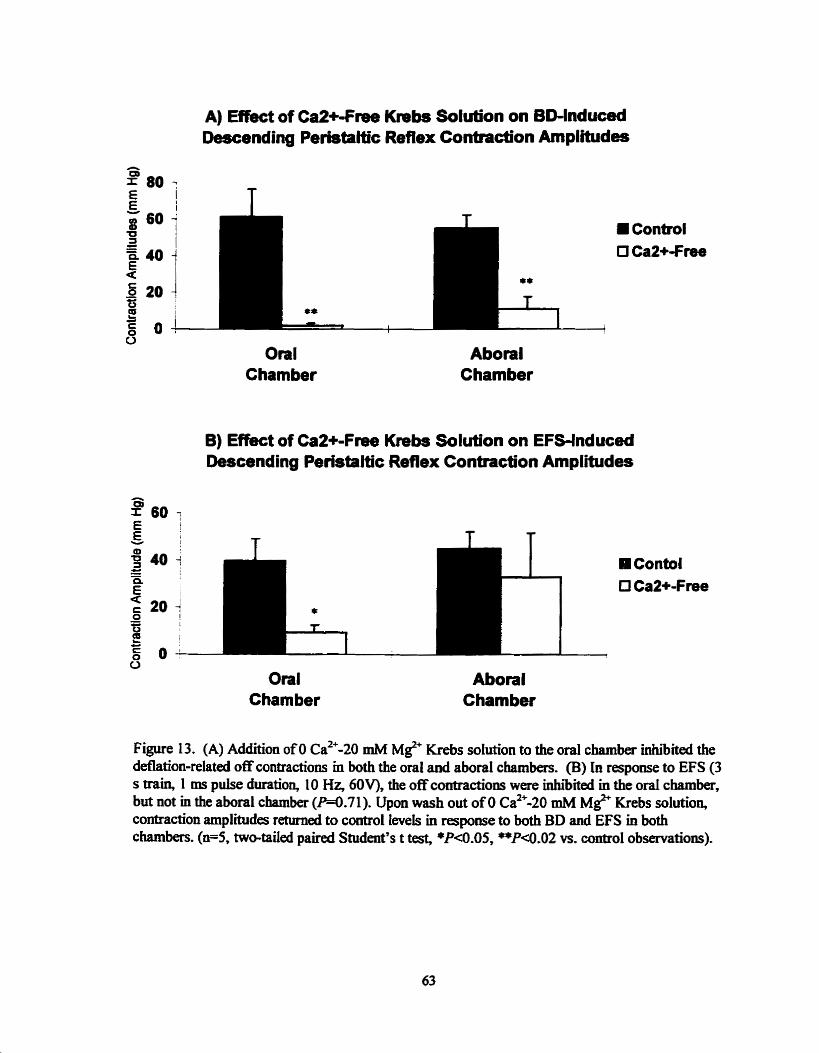
A) ERed of Ca2+-Free Descendhg Peristaltic
Krebs Solution on BD-lnduced Reflex Contraction Amplitudes
O Oral
Chamber Aborai
Chamber
B) Effect of Ca2+-Free Krebs Solution on EFS-lnduced Descending Peristaltic Reflex Contraction Amplitudes
a Oral
Cham ber Aborai
Chamber
Figure 13. (A) Addition of O cas-20 mM MC Krebs solution to the oral chamber inhiibited the deflation-related off contractions in both the oral and aboral chambers. (B) In response to EFS (3 s train, 1 ms pulse duration, 10 Hz, 60V). the off contractions were inbibiteci in the oral chamber, but not m the aboral chamber (W.71). Upon wash out of O ~ 8 - 2 0 rnM MC Krebs solution, contraction amplmides returned to control levels in response to both BD and EFS in both chambers. ( ~ 5 , two-tailed paired Student's t test, *PQ).05, **Pa.02 vs. control observations).
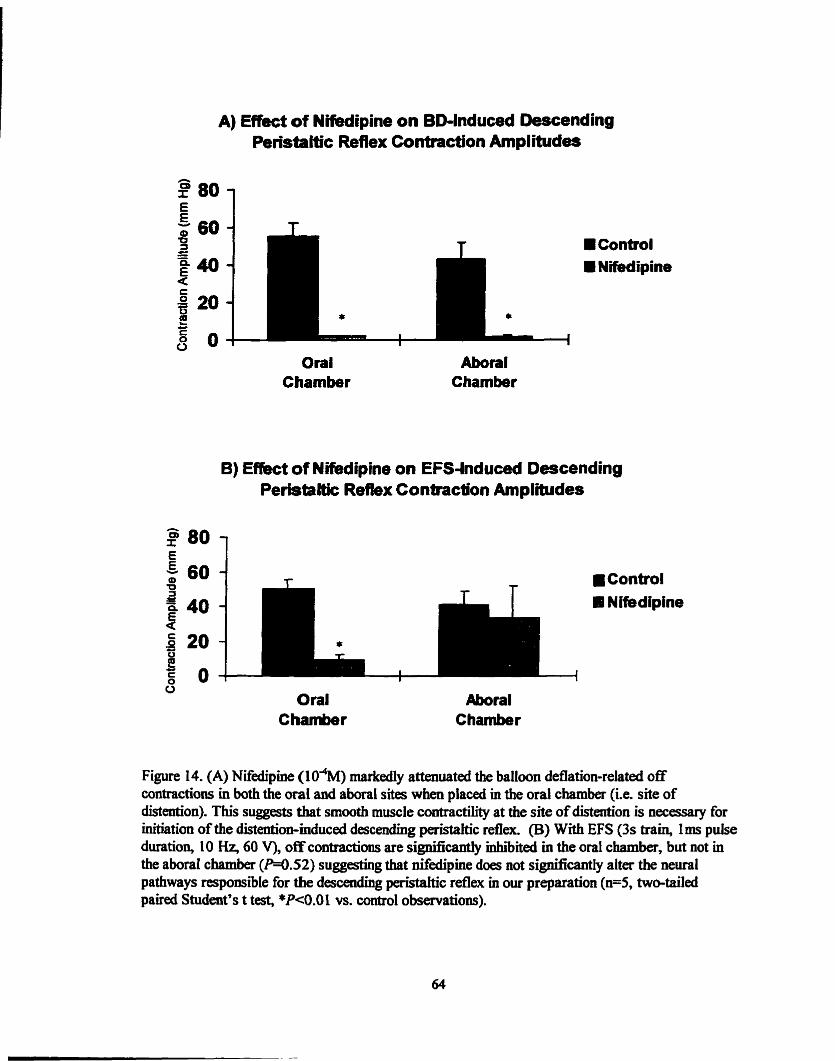
A) Effect of NMipine on BD-lnduced ûescending Peristaltic Reflex Contraction Amplitudes
E 60 -
O 3 C - II Control E40- Nifedipine 4
20 - P * 8 O - I
Oral Chamber
Aboral Chamber
B) E î k t of Nifedipine on EFS-lnduced Descending Peristalt- Reflex Contraction Amplitudes
Conbol U Nifedipine
1 I i
Oral C harnbe r
Figure 14. (A) Nifedipine (10%) markedly attenuated the baiioon deflation-related off contractions in both the oral and aboral sites when placed in the oral chamber (Le. site of distention). This suggests that srnooth muscle contractility at the site of distention is uecesssry for initiation of the distariion-mduced descending peristaltic refiex. (B) With EFS (3s &ab, lais pulse duration, 10 I - 4 60 V), off contractions are significaxrtiy inhïibited in the oral chamber, but not in the abord cbamk (W.52) suggsting that nifédipine does not signincantly alter the neural pathways responsible for the descending paistaltic reflex m our preparation ( ~ 5 , twwtailed paued Student's t test, *P<O.O 1 vs. control observations).

charnber (Fig. 14B) were not inhiïied (41.4 +/- 7.3 mm Hg vs. 33.4 +/- 18.5 mm Hg,
W -52).
zii)Eaect of Chderythrine Chloride
When p l a d in the oral chamber, the PKC-ùihi'bitor cheleythruie chloride (104 M;
n=5) sigdïcantly inhiiited the BD-induced off contraction in the oral chamber (46.0 +/-
11.7 mm Hg vs. 1.5 +/- 0.6 mm Hg, W.019) and abord chamber (49.5 +/- 4.0 mm Hg
vs. 2.4 +/- 1 -6 mm Hg, W -0002) (Fig. 1 1 c, Fig. 1 SA). Larger BD volumes (i. e. 3 cc)
were also unable to initiate this inhibited reflex response. Unlike nifedipine, the inhibitory
effects of chelerythrine chloride were partially reversible following repeated washings with
fresh Krebs solution. In those experiments where reversibiiity data was obtained, the off
response returned to -70% of control values ( ~ 3 ) (Fig. 1 le*. In response to EFS
(n=5), the off contractions were significantly inhibited in both the oral chamber (36.5 +/-
12.9 mm Hg vs. 10.2 +/- 4.6 mm Hg, W.041) and abord chamber (32.4 +/- 3.6 mm Hg
vs. 2 1 -2 +/- 5 -2 mm Hg, M . 0 3 1) (Fig. 1 1 d, Fig. 1 SB), dthough the magnitude of
inhibition was signifimtly greater in the oral charnber (73.9 +/- 5.7% decrease in
amplitude in oral chamber vs. 30 +/- 8.9% decrease in aboral chamber, M.016, Mann-
Whitney test).
Effe* of Blockade of Synaptic Transmission
i)Aborul Chamber
Previous studies (Paterson and Indrakrishnan 1995) have clearly donimented that
the off contraction following balloon deflation involves synaptic transmission between

PerWaItic Reflex Contadion Pnplihides
Oral Chaniber
B) Effect d Chekrythrlne of EFS-lnduced Descendhg Peristaftic Reflex Contraction Anplitudes
Oral Aboral
Figure 15. (A) Chelexythrine chioride (lo4M) when placed m the oral chamber ngni f idy inhi'bited the ballooa de£iatian-related off contractions m buth the orai and abonil chambers. (B) In response to EFS (3 s train, Ims puise duraîïon, 10 H z , 60V), the mean amplitude ofthe off cmtmctiolls were significantly Hihibaed in bath the oral chamber and aborai chamber. (n=5, two- taiied paired Student's t tesf #P<0.05. *P<O.O2, **P<0.00 1 vs. cam-01 observations). Statistical analysis with the Mann-Whitney test revealed that with EFS, îhere was a signifiant difIéreince beîween the median perceut inhi'biticm in the oral chamber and the aboral chamber (73.9 +/- 5.7% VS. 30.0 +/- 8.9% W.016).

nerve and muscle. Therefore, to determine the optimal dose of Q-CTX to use in our
experiments, three pilot studies were Camed out in which the &ect of d T X in
concentrations ranging between 104 M to IO-' M on the BD-induced off contraction in the
abord chamber was detennined. AU three concentrations used produced greata than
70% inhiaition of these contractions. Because of the expense of this drug and the
apparent near ma>timal inhibition at lo4 M, this conceniration was chosen for subsequent
experiment S.
i i ) W C h b e r
Addition of o-CTX (1 o4 M) to the oral chamber (Fig. 16) did not resuit in
statistically signifiant inhibition of the BD-induced off contraction in either the ord
chamber (76.4 +/- 36.3 mm Hg vs. 36.0 +/- 10.3 mm Hg; n=5; m.36) or abord
chamber (96.7+/- 63.2 mm Hg vs. 43.4 +/- 26.7 mm Hg; n=4; W.19). However, the
results of these experiments were highiy inconsistent f?om one preparation to the next.
This included one experiment where off contractions in the oral chamber (which involves a
synapse between NO containing i n t ~ s i c ne- and smooth muscle) were a d y
enhanceci following addition of 0-CTX to the oral chamber.
Esophageai Cornpliance Studies
Effect of Smooth Muscle Relaxants snd Neural Blockade on:
i ) EKphuged CornpZiance
Pressure-volume m e s of the smooth muscle esophagus were constructeci fiom
tracings such as Fig. 9. Linear regression analysis revealed that the pressure-volume

E f b t of Conotoxin in Oral Chamber on BWnduced Descendhg Perirtaloc Reflex Contraction Amplitudes
Oral Cham ber
Aborat Chamber
Figure 16. Addition of a€TX ( 1 0 ~ M) to the oral chamber had no sipnificant inhibitory efféd ai the BD-mduced off contraction m eÏther the o d chamber (n=5, W . 3 6 ) or aboral chamber (n=4, W. 19) (twbtailed paired Student 's t test).

airves were hi* linear in all experimental treatment groups (Fig. 17) and had the
following slopes: corn01 expaiments (0.50 +/- 0.04 d m m Hg; h . 9 5 , n=5); nifidipine
(0.45 +/- 0.03 mVmm Hg; h . 9 5 , n4); chelerythrhe chbride (O -46 +/- 0.03 mVmm Hg
?=0.96, n=4); O car*-20 mM MC Krebs (0.48 +/- 0.04 d m m Hg; h . 9 5 , n=4); TTX
(0.49 +/- 0.04 mVmm Hg; h . 9 5 , fi). As determineci by ANOVA, there were no
significant differences (M.91) between the slopes of the pressurevolume m e s in any
of these treatment groups.
ii) Resistmce of Esophageal Wall to Initia2 Saetch
The values of the x-axis intercept, which represent the resistance of the esophageal
w d to initial stretch, were extrapolated from the pressure-volume cuwes of the treatment
groups and are shown in Fig. 18. As determined by ANOVA, there were no significant
differences ( M . 7 1 ) arnong the x-axis intercepts between any of treatment groups:
controis (-2.33 +/- 0.48 mm Hg); aifedipine (-2.43+/-0.3 5 mm Hg); chelerythrine chloride
(-2.17+/-0.89 mm Hg); O ca2--20 mM M ~ " Krebs (-2.3 1+/-0.90 mm Hg); TTX (-3 -29 +/-
0.54 mm Hg).
iii) PhPric Conb=actiom Uver Top of the Distench'ng Bag
With inflation of the barostat bag to pressures of at least 3 mm Hg, phasic
contractile activity was induced in the esophagus. As can be seen in the typical control
tracing (taken at a distendhg barostat bag pressure of 8 mm Hg), phasic contractile
advity occurred on top of the barostat bag throughout most of the period of bag

Emct of Inhibitors of Sm- Muscle Contractility and NeumtransmîssIon on Esophageal Body Pressure-
Volume Curves
O 5 I O 15 20
Di-nding Preanim (mm Hg)
Figure 17. The dope of the pressure-volume cumes as established with the electronic barostat were no c i B i between any of the treatment groups m cornparison to controls ( M . 9 1, ANOVA).

Effbct of Inhibitors of Sm>oth Muilde Conttactility and NeufotraiwnJsùon on Resistance of Esophageal Wall to
Initial Stretch
Figure 18. The resistance to initiai stntch, as exîrapoked 6mn the x-intercept fhm the pressure- volume curves, is a reflection of tone in the resting state. TXis point represents the level at which the distending pressure inside the barostat bag is e q d to the pressure outside the bag, and correspb~1ds to the minunum amouut of pressure required to distend (or open) the esophagus. There were no signifiant difnrrences m x-intercept values or resistance to initial stretch ammg any ofthe treatmm groups m cornparkm to wutrois (W.71, ANOVA).
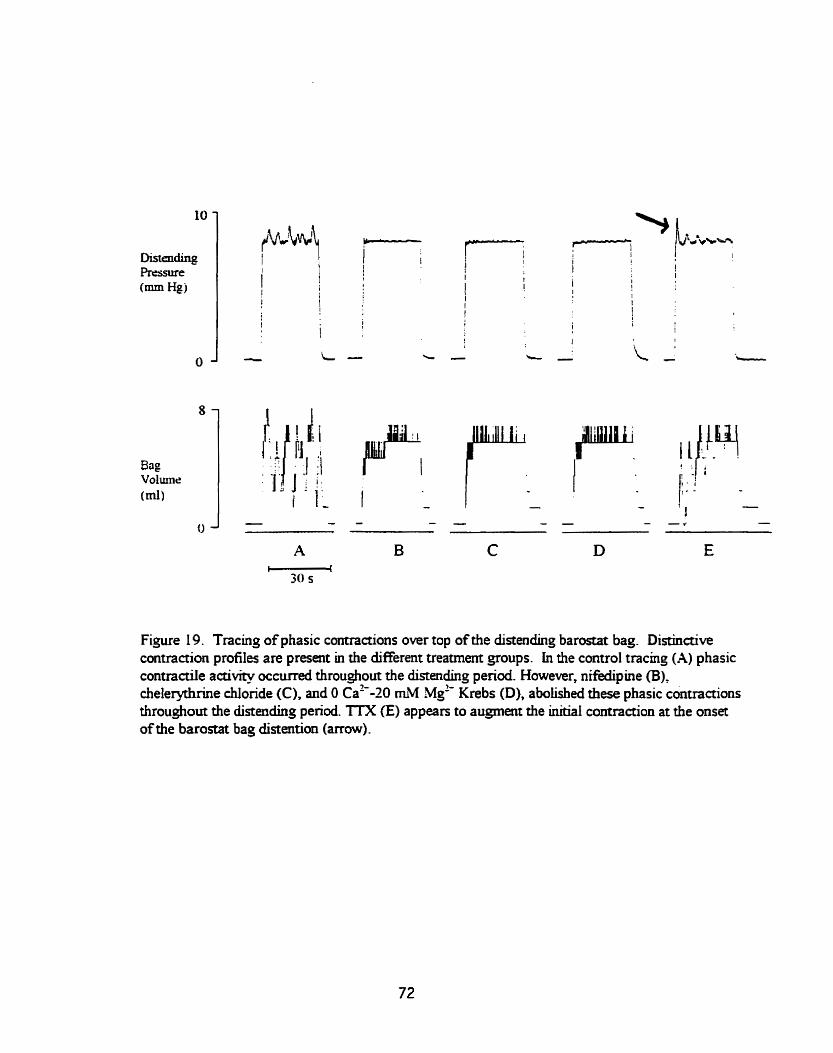
Figure 19. Tracing of phasic contractions over top of the distendhg b a r o ~ t bag. Distinctive contraction profiles are present in the different treatmem groups. in the control tracinp (A) phasic contractile activity occurred throughout the distmding period. However, nifedipine (B)_ cheleryrhrine chlonde (C), and O ca2--20 mM ~ g - Krebs (D), abolished these phasic contractions throughout the distendhg period. TTX fE) appean to augment the indial contraction at the onset of the barostat bag distation (arrow).

distention (Fig- 19A). This contractile aaivity was M y abolished with Nfedpine,
chelerythrine chloride or O ca2'-20 m . MC Krebs solution (Fig. 19B-19D).
To quantify the inhibition of phasic contractility in response to these agents, both
the contraction fkquency and contraction index were examineci. The contraction index,
as dehed in the methods section, was calculated for ail treatment groups. ANOVA
detected significant variation amoog treatment groups (P<0.0001). The Student-
Newman-Kds multiple-sample comparison post test was carrieci out to Merentiate
among this variation. As depicteci in Fig. 20, with respect to the control group (132 +/- 24
ml x s), the contraction index was significantly inhibited by nifedipine (1 +/- O ml x s,
PKO.00 1 ), chelerythruie chloride (1 5 +/- 10 ml x s, P<0.00 l), and O ca2* -20 mM ~ g * '
Krebs solution (1 +/- O ml x s, P<0.00 1). However, n w a l blockade with TTX (9 1 +/- 14
ml r s, -0.05) did not significantly alter the overd contraction index. The frequency of
contractions during a complete distention cycle were aiso investigated. ANOVA detected
si@cant variation among treatment groups (P<0.0005). The Student-Newman-Keuis
multiple-sample comparison post test was carried out to differentiate among this variation.
With respect to mntrol group (1 2.8 +/- 2.0 contractions per distention cycle), the
contraction fiequency was s i sn i f idy ïnhibited by nifedipine (0.3 +/-0.0, P<O.O 1 ),
cheleryuuine chionde (2.8 +/- 1 -7, PC0.0 1 ), and O ca2+-20 mM M~'' Krebs solution (0.0
+/O 0.0, P<0.01). However, neural blockade with TTX did not signincantly alter the
o v d contraction fkquency (9.2 +/- 2.9, Dû.05).
Visual inspection of the contraction profile during distention suggested that TZX
augmenteci the contraction amplitude of the initial contraction over top of the barostat bag
(see arrow) (Fig. 19E). Quantitative d y s i s revealed that there was a signincant inmease
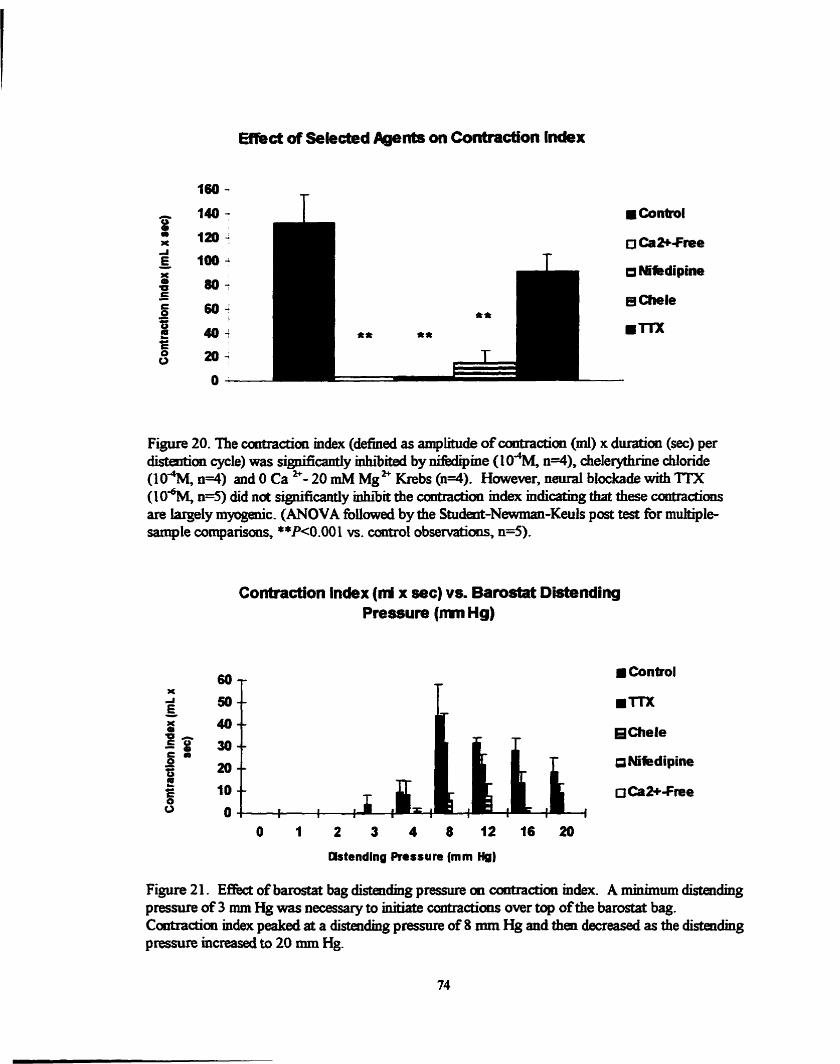
E-ct of Sekded Agents on Contraction lndex
- 140- 8 1 " * lmJ A E t0o-I - X Q)
a -dipine u E
80- - g 6 0 ;
s Chele L " 40; m m o 20-
O
Figure 20. The contraaiou mdex (definecl as amplitude of contraction (ml) x dufation (sec) pet distention cycle) was signikmtiy inhiibited by -me (10"M, n4), chelerythrine chloride (104M, 0=4) and O Ca "- 20 mM M~ Krebs (n=4). Hawever, neurai blockade with TIX (10% n4) did na significantiy &'bit the contraction m k mdidng that these c ~ n t r a c î i ~ l l ~ are largely myogeiic. (ANOVA foiiowed by the Studesit-Newman-Keuls post test for multiple- sample comparisons, **P<0.00 1 vs. contr01 observations, ~ 5 ) .
Contraction lndex (nrl x sec) v a Barostat Dbdending Preswre (mn Hg)
x A E 1IC
50 -- mTTX
3 40 -- Chele I g 30-- e - - 20 -- a Mfedipine s 2 w C O
10 -- O Ca*-Free O O t 1 1 I
1 I I I I
Wtending Pressure (mm Hg)
Figure 21. Effed of bamstat bag disteidmg pressure m contraction mdex. A minimum distendmg pressure of 3 mm Hg was neces- to initiate contractions over top of the barostat bag. Contraction index peaked at a distendmg pressure of 8 rnm Hg and then decfeased as the distendhg pressure increased to 20 mm Hg.

in Uiitial contraction amplitude between control and TTX arperiments, as demonstrateci by
barostat bag displacement volume at distending bag pressures of both 8 mm Hg (4.6 +/-
0.7 mi vs. 6.6 +/- 0.7 ml; ~ 5 ; W.01) and 12 mm Hg (4.2 +/- 0.4 mi vs. 6.8 +/- 0.9 ml;
~ 5 ; W.007). However, at these distendhg pressures, there were no différences
between wntrol and m( experirnents in either the duraton of the initial contraction (8
mm Hg: 3.8 +/- 0.5 s VS. 3.6 +/- 0.4 S, W.77; 12 mm Hg: 3.8 +/- 0.4 s VS. 3.3 +/- 0.3 S,
W -3 5 ) or iatency to omet of the initial contraction (8 mm Hg: 1 -2 +/- 0.1 s vs. 2.7 +/-
0.7 s, M . 1 0 ; 12 mm Hg: 12 =Hg: 1.5 +/-0.3 S V ~ . 2.5 +/- 0.4 s, M . 0 8 ) .
A minimum distending pressure of 3 mm Hg was required to initiate phasic
contractions over top of the barostat bag (Fig. 2 1). Both contraction fiequency and
contraction index peaked at a barostat bag distendhg pressure of 8 mm Hg and then
decreased as the distending pressure increased to 20 mm Hg.

Chapter 4
Discussion
The distentfo11-induced descendhg peristaltic reflex is a b@iy complex motor
activity which is irifluenced by central and peripheral levels of controt as weil as myogenic
propeaies of the musculature. Though this rdex has been extensiveb studied m the
snall intestine, M e is known about the 8itramural rnechanisms responsiile for the
initiation and propagation of this reflex m the srnooth muscle esophagus. The primary
objective of this thesis was to bvestigate the role of muscle coneaaility m the initiaton of
the distention-induced descendhg peristakic reflex.
As previously docunrented 51 the triple-chamber organ bath studies of Paterson
and Indrakrishnan (1995), addition of O Ca " -20 mM Mg '' Krebs to the oral chamber
(Le. site of distention) abolished the distention-induced descmdmg pristahic reflex
However, it was not clear from these earlïer studies whether this Inhrion was due to
synaptic blockade or mipairment of muscle contractiiity. The current experiments support
the latter.
Descending Peristaltic Reflex Studies
i ) Role of Local Smooth M d e Contractility
Calcium charnel blockers, mcluding nifiedipine, are widely used to treat spastic
disorden of the esophagus hcluding nutcracker esophagus and diffw esophageal spana.
In humans, they have been shown to d u c e LES tone and the amplihide and d d o n of
pristahic esophageal contractions (Blackweii et al. 198 1, Smout et al. 1992). In our
studies, we employed nifiedipine to selectheiy impair Si&ur of extracellular calcium into
76

the srnooth muscle cells, and hence i n h i esophageal srnooth muscle contractiiity without
affecting synaptic traasmission Though no studies could be found that investigated the
effects of nifiedipine on synaptic transmission in the opossum, studies in the mea-pig
ileum have demonstratecl that blockade of L-type calcium chaunels with either verapamil
or nifedipine does not impede synaptic transmission (Izza et al. 1997).
In our preparation, addition of nEkdipine to the oral chamber Whially abolished
the baiioon deflation-related off contractions in both the oral and abord chambers, and the
EFS related off contractions in the oral chamber. This inhihitory efféct of nifidipine was
not revenible as responses did not return afkr severai washes with Krebs solution, even
d e r 1 hour. Though we did not attempt to reverse the effect of nifedipine by iocreasing
the calcium ion content in our preparation, previous in vitro studies in the smaü intestine
- have demonstrated that the inhibitory effect of Sedipine on intestinal wntractility is not
r e v d by increasing calcium ion concentration in the organ bath (Grbovic and
Radmanovic 1987). It is unlikely that nifedipine affecteci neurotransrnission in the
descendhg reflex pathway since EFS of intrarnuraj nerves in the ord chamber was stiU
able to initiate the reflex as demonstrated by off contractions in the abord chamber.
Previous studies have demonstrated that both phasic CM contractiiity in the
esophageal body and LES tone are mediated by PKCdependent imrace1lula.r transduction
pathways (Biancani et al. 1994, HiIiemeier et al. 1996, Biancani et al. 1997). In the
experiments in this thesis, the PKC-inhi'bitor chelerythrine chloride significamly inhibited
the descendhg peristaltic refiex when added to the site of distemion. As expected, the
EFS-induced contraction amplitudes wen markedly inhibitecl in the oral chamber.
However, EFS-induced contraction amplitudes were also inhibited in the abord chamber,

although the niagmhide of this mhi'bition was ggniscady less than m the oral c)wnber.
This raises the possibility that chekrytbrine chloride bas some mimi non-specific effect on
neurotransmission. It is also possible, however, thaî the signifiant decline in response m
the aboral chamber with EFS was at least par@ due to mn-speciiïc Eatigue of the
prrparation
Thus, nifiedipine and chelerytbrhe chloride, agents which &'bit s~llooth muscle
conûactility by Mixent mechanisns, both aboîished the BD-induced descending
pe&tahic reflex when placed m the oral chember. These results mdicate that local snooth
muscle contraction at the site of distention is an miportant component of the distention-
mduced descendhg peristahic reflex E the esophagus.
ii) Role of Synaptic Tr~lf~l l t ission: Effect of mConotoxin GVLP
Although the results with nifedipme and chelerythrine chloride m the oral chamber
strongiy suggested that the previousiy documnted efkt of O ~a"-20 mM ~ g ' + Krebs
solution m this chamber was due to impairment of muscle contraction rather than an effect
on synaptic transmission, confimatory d e s were attempted by determinhg the effect of
synaptic blockade with a>-CTX on this reflex
mCTX blocks N-type, but not L-type Ca" channels, hence preventing the idlm
of extracellular calcium into presynaptic nerve terminais whüe aiiowing -0th muscle
ceiis to utilize extracellular calcium necessary for tension development in response to
stretch. ûur results with this toxh were hconclusive. Shce synaptic transmission
between nitrergic neurons and the snooth muscle ceii is the final step in the descending
peristaltic reflex (Paterson and In- 1995), t was expected that the addition of
78

mCTX to the aboral chamber wodd &%Ît this synaptic transmission and hence abolish
the off contractions in this chamber. Indeed in püot studies, addition of o-CTX to the
aboral chamber r d t e d in marked decrease of aborai wntracaons. Howwer, i f d T X
was &&g synaptic transmission in our preparation, then wheo placed in the oral
chamber, it should also clearly inhibit the defiation-related ~Ecuntractions in that chamber
(Le. 1 cm site). Howwer, addition of (D-CTX to the oral chamber had inconsistent effects
on the distention-ùiduced descending peristaltic reflar. There was no signincant inht'bition
of oral or aboral contraction amplitudes. It is possible that wCTX in the concentrations
used is not selective in our tissue preparation, or that it has some unknown non-specinc
effects.
In smooth CM stnps fiom the opossum esophagus, Cayabyab and Daniel (1995)
have demonstrated that aXTX (300 nM) slowly inhibiteci ï J P s by 2040%, reachg
maximal significant inhibition at 40-60 min. In our pilot studies t appeared that I O ~ M
mCTX was having a profound effect on synaptic transmission as reflected by the
decreased amplitude of the off response. However, this was not reproducible with our
subsequent experiments. Ifthe degree of inhiition caused by aFCTX was less than those
in the pilot studies (Le. 2040% vs. over 70%), then we estirnatecl that a much Iarger
sample size (n>20) would be required to maintain the probability of an @ha (Type I)
error at 5% and a beta error (Type II) at 1W. Our experiments with mCTX wdd not be
pursued m e r as the cost of this agent and number of animals required was prohibitive.

Barostat Studies
i) h p h a g e d Body Resthg Tone
The objective of the barostat studies in the smooth muscle esophagus was to assess
the mechanism by which the selected agents were inhiiiting the descending perist;iltic
reflex. W e initidy hypothesized that tonic muscle contraction ocamkg in response to
esophageal distention was criticai to the initiation ofthe descending peristaltic reflex and
that this tone was inhiited by nifedipine7 chelerythrhe chloride and O ca2+-20 mM M~''
Krebs solution.
IntereSnngly, we could not detect any sigdicant différences in esophageal
cornpliance or resistance of the esophageal wali to initial stretch in any of the treatment
groups. This finding suggests that these agents do not alter resting tone in the esophaged
body and that the &ect of these agents on the distention-induced descending peristaltic
reflex was not due to inhiiition of tonic esophageal contraction in response to stretch.
These redts are consistent with other barostat studies which have demoastrated that
dedipine does not alter resting esophageal body tone in vivo in humans (Smout et al.
1992)'. This is in contrast to the LES where both nifidipine and chelaythrine chloride
inhibit tonic wntractility in a dose dependent mamer (Blackweil et aï. 1991, Biancani et
al. 1994). It is possible that there is nrinimal active tone in the esopbagus and that it is
dependent on intraceiiuk calcium stores. However, these intracelIular stores should be
depleted over time. Our protocol was conducted over a 2 hr period. Even &a 1 hr,
more than adequate t h e for the dmgs to take &&ct, and in the absence of extraceiidar
calcium, there were no signifiant Merences between the control and treatment groups.

These r d t s suggests that there may be no active tonic contraction of the
esophageal body of the opossum in Mtro either at rest or in response to distedon This is
consistent with recent shidies in humam (Onnir et al. 1993) where no measurable tone
was found in the esophageal body at rest. Furthermore, T@tmp and colleagues (1990b)
have demonstrated that in vitro CM strip preparations firom the cat esophageal body show
little resting tone under isometric conditions. However, recent in vivo barostat stuciies in
the human have demonstrateci that smooth musde tone (both cornpliance and resistance to
initial metch) has an active component that can be reduced by the snooth muscle
relaxant, amyl nitrate (Mayrand et al. 1 994, G o d e z et ai. 1997). We did not confirm
active tone in the opossum esophagus using other phannacological methods of relaxation.
One explanation for our results is that the resistance to stretch in the esophagus is
due primarily to passive visco-elastic properties of the esophageai wall. For instance,
there are localized différences in composition of smooth muscle in the esophagus, such as
vaIied levels of coilagen and elastic fibers, and these factors may be contributhg factors to
the compliance of the esophagus (Gregersen et al. 1992, Gregersen and Kassab 1996). 4
It is uniikely that the visco-elastic properties inherent in the barostat bag
contributed to these results. We employed an ultra thin polyethylene bag with high
compliance over the range of volumes used ensuring that the elastic recoil of the bag was
vimiaily nil. This set-up peRnitted the measurement of the intraesophageal volume under
a @en set of isobaric conditions that t d y rdected the esophageal wall properties
exclusive of the barostat bag properties.

ii) Phûnc Musce ContractiIity
Thou& tonic muscle actbhy was not altered in these experimentq phasic
contractions over top of the distending barostat bag were signifïcantly iahibited by
dedipine, chelerythrhe chloride and O ca2'-20 mM M ~ ~ + Krebs solution. However, TTX
did not siBnificantly inhibit these phasic contractions, indicathg that they are largely
myogenic in origin Prewious BD studies in the opossum esophagus Nt Mvo recorded
smooth muscle contractions over top of the distending baiioon during the period of
baiioon distedon (Paterson 199 1). The amplitude of these contractions increased with
exposure to TTX. This not only suggests that these contractions are myogenic in ongin,
but also that under n o r d circumstances, the muscle receives some tonic inhibitory input
that is removed when TTX is appîied. Christensen (1970) also observai similar TTX-
resistant contractions at a d o r just proximd to a distendmg bdloon It is important to
recognize that uniike the intraesophageal bdoons used by Christensen (1970) and
Paterson (1 99 1 ), which were 2 cm in length and inflated for approximately 5 seconds the
phasic contractions recorded in this current protowl were from a 8 cm long bag which
was uinated for 30 seconds. Hence the contractile activity observed in these experiments
represents a composite (Le. both myogenic and nauogenic) of esophageal contractions Ui
response to prolonged bag distention over a long (8 cm) segment.
Though the types of phasic contractions were not assessed, it is interestkg to note
that the initial contraction in response to barostat distention was si@camly enhanceci in
the presence of TTX suggesting that this contrade response is fbnctionally simiiar to the
contractions observed on top of the distending balioon as d e s c n i by Paterson (1991).

The threshold for distention induced peristalsis might be an important determinant
for the development of reflux esophagitis. Impairment or ioss of sensi- of s-ry
thresholds in the esophagus may r d t in a gr-er amount of esophageal contents being
required to imtiate secondary peristalsis. This constant exposure of rduxed contents
could lead to gastroesophaged r&ux disease. To ensure that the aboiishment of the
descendhg -tic renat was not due to receptor de sensitizatoq kger BD v o h e s (3
cc) were ais0 used after addition of the agent being teste4 however, the results were
consistent with the iower BD volumes (i-e. siBnificant inhibition of the reflex). In the
barostat studies, it was observed that a minimum distending pressure of3mm Hg was
necessary to initiate contractions over top of the barostat bag. The CI peaked at a
distending pressure of 8 mm Hg and then decreased as the distendhg pressure increased
to 20 mm Hg. This suggests that a lower limit of smooth muscle tension is required to
initiate phasic contractiiiity. This result is consistent with findings by Brookes and
wlleagues (1997) who demo~l~trated that in segments of guinea-pig ileum, when close to
threshold stretch, peristalsis cm be initiated sirnply by increasing circular muscle tension.
Furthermore, esophsgeal CM "off' contraction amplinides Vary in relation to the degree of
muscuiar tissue stretch prior to EFS stimulation, suggesting that myogenic factors also
contniute to bolus modulation of peristalsis (Sama et al. 1977, T+ttnip et al. 1990b).
Summary
The aim of this thesis was to examine the relationship between local smooth
muscle contractility in response to stretch and the distention-induced dexending peristaltic
reflex. These experiments demonstrate that the descendhg penstaltic reflex is markediy

mhîited when agents afEecting smooth muscle contractility, but not synaptic trammision,
are placed at the site of distentioa These agas do not alter the w-mpliance of the
smooth muscle esophageal body, or its resistance to initial stretcb, but ïnhiiit phasic
coniractions over top of the distendhg barostat bag. Taken together, these observations
suggest that local stretch-induced phasic muscle contractions are involveci in the initiation
of the baiioon distention-induced descendhg perïstaitic reflex in the opossum smooth
muscle esophagus.
These fïndings, when combined with the previous results of Paterson and
Indrakrishnan (1 995), support the hypothesis (Mode1 1-see Fig. 3a) that the distention-
induced descending peristaitic reflex involves long descending intramural neurons that may
have both mechano-sensory and motor fictions. That is, local stretch-induced phasic
muscle contraction may directly activate long descending intramural neurons, which then
release NO ont0 ciradar smooth muscle dom stream This in tum produces
hyperpolarization (inhibition) of the muscte foilowed by "rebound" contraction
(excitation). Confirmation of this hypothesis however, must await anatomid
demonstration of such neurons.

Al Yassïn, T.M. and P.G. Toner. Fie structure of squamous epithehm and submucosal glands of human esophagus. J. Amt. 123: 705,1977-
Anand, N. and W.G. Paterson. Role of nitnc oxide in esophageal peristalsis. Am. J. P&sioZ. 266 (Gastrointest. Liver Physiol. 29): G123-G13 1, 1994-
Andrew, B.L. The newous control of the cervical oesophagus of the rat during s~dowing. J. P&si01. Lond. 134: 729-740,1956.
Arey, L.B. and M-J. Tremaine. The muscle content of the lower esophagus in man. And Rec. 56:315, 1933.
Asaad, K., Abd-El Rahman, S., Nawar, N.N.Y. and Y. Mikhail. Intrinsic innewation of the oesophagus in dogs with special reference to the presence of muscle spindles. Acta Anaf. 115: 91-96, 1983.
Asoh, R and RK. Goyal. Manometry and electromyography of the upper esophageal sphincter in the opossum. Gartroenterology 74: 5 14-520, 1978.
Azpiroz, F. and J.R Malagelada. Physiological variations in canine gastric tone measured by an electronic barostat. Am. J. Physol. 248 (Gastrointest. LNer Physiol. 1 1): G2290G237, 1985.
Barajas-Lopez, C., Peres, A. L. and R Espinosa-Luna. Ceiiular mechanisrns underlying adenosine actions on chohergic transmission in enteric neurons. A r J. Physiof. 27 2 :C264-C275, 1996.
Baumgarten, M.G. and W. Lange. Adrenergic innervation of the oesophagus in the cat (Fefis h e M c a ) and rhesus monkey (Macacus rrhess). Z ZeIIorsch. Mikrosk And. 95: 529-545, 1969.
Bayliss, W.M. and E.H. Starling. The movemems and innemation of the small intestine. J. Physiof. 24:99, 1899.
Bianami, P., Sohn, U.D., Rich, H.G., Haniett, KM. and J. Behar. Signal Transduction Pathways in Esophageal and Lower Esophageal Sphincter Cudar Muscle Ain. JMed 103(5A):23S-28S, 1997.
B i a n w P. and J. Behar. Esophageal Motor Function. In: Textbook of Gastroenterology, Section B. Motility, Second edition, Ed. Tadataka Yamada. J.B. Lippincott Company, Phüadelphi~ 1995.

Biancani, P., Harnett, KM., Sohn, U.D., Rhirn, B.Y., Behar, J., Wemeier, C. and KN. B k . Merential signal transduction patbways in LES tone and response to Ach. Am. J . P h y s ~ i - 266:G767-G774,1994.
Biancani, P., Baafed, MC, HilIemeier, C. and J. Behar. Role of peptide isolacine in relaxation of cat lower esophageal sphincter. Gastroenteroiogy 97: 1083-9, 1 989.
Biancarù, P., Hillemeier, C., Bitar, ICN. and G.M. Makhlouf. Conûaction mediated by ca2' ïnûux in esophageai musde and by Ca ** release in the LES. Am. J . Phj~sioI. 253 (Gastrointest. Liver Physiol. 16): G760-766, 1987.
Biancani, P., Walsh, J.H. and J. Behar. Vasoactive intestinal peptide: a neurotransmitter for lower esophageal sphincter relaxation. J. C h Illvesi. 73: 963-967, 1984.
Biancani, P., Zabinski, M.P., Kernstein, U and J. B e k Lower esophageal sphincter mechanics: anatomic and physiologic relationships of the esophagogastric junction of cat. GcrstroeritetoIogy 82: 468-475, 1982.
Biancani, P., Zabinski, M. and J. Behar. Pressure, tension, and force of closure of the human lower esophageai sphincter and esophagus. J. Ciin I imst . 56476, 1975.
BlackweU, J.N., Holt, S. and R C . Heading. Effect of nifedipine on oesophageal motiüty and gastric emptying. Digestron 2 1 : 50-56, 198 1.
Blank, E.L., Greenwood, B. and W.$. Dodds. Cholinergie control of smooth muscle peristalsis in the cat esophagus. Am. J. Physoi. 257: GS l7-GS23, 1989.
Brookes, S. J.H., Zafiorodnyuk, V., Chen, B.N., Humphreys, C.M.S. and M. Costa. Mechanism of Initiation of Peristalsis in the Guinea-Pig ileum. [Abstract] GustroenteroIogy 1 22: 4, 1 997.
Brookes, S.J.H., Song, 2. -M., Chen, B.N. and M. Costa. Neuronal and rnudar mechanisms interact to tngger peristalsis. [Abstract] Gatwenterdogy 1995.
Burgess, J. N., Schlegel, J.F. and F. H EEs. Eff- of denmation on feline esophageal nuiction and morphology. 4 Surg. Res. 12:24,1972.
Bumtock, G. Uhstruchiral identification of neurotransmitters. In Basic Science in Gastroenterology: Structure of the Gut, edited by J.M. Polak, S.R Bloom, N-A Wright and M. J. Daly. Norwich, Engiand, pp. 1-1 O, 1982.
Carpenter, D.O. Central navous system mechani- in deglutition and emesis. In: Handbook of Physiology. Section 6: The Gastroïntestinal System, edited by S.G. Schultz, J.D. Wood and B.B. Rauner. Bethesda, Maryland: Arne- Physiological Society, pp. 685-714,1989.

Carveth, S.W., Schlegeb J.F. and CF. Code. Esophageal motility after vagotomy, phrenicotomy, myotootomy, and myomectomy in dogs. Surg. Gpecol. Obstet 114: 3 1-42, 1 962.
Castell, D.O. and J.A Casteii. Esophageal Motility Testing. 2nd Edition. Appleton & Lange. 1994.
Cayabyab, F-S. and E.E. Daniel. P charme1 opening mediates hyperpolarizations by nitric oxide donors and UPs in opossum esophagus. A m J. Physiol- 268: G83 1-G842, 1995.
Christenseq J. Mechanisms of Secondary Esophageai Peristalsis. Am. J. Med 103(5A):44S46S, 1997.
Christensen, J. The entenc nervous system. In: An iiiustrated guide to gastrointestllial motility. D. Kumar and D. Wmgate (eds). New York: Churchill Livingstone, pp. 10-31, 1993.
Chnaensen, J., Williams, T.H., Jew, J. and T.M. O'Donsio. Distribution of hunoreactive substance P in opossum esophagus. Dig. Dis. Sci. 34: 5 13-520, 1989.
Christensen, J., Rick, G. A and D. J. SON. Intrammil nerves and interstitial cells revealed by the Champy-Maillet stain in the opossum esophagus. J. Auton. Nem. Sys. 19: 137-151, 1987a.
Christensen, J., Williams, T.H., Jew, J., and T.M. O'Dorisio. Distribution of vasoactive intestinal polypeptide-immunoreactive structures in the opossum esophagus. Gmoenterology 92: 1007-1 O 18, 198%
Christensen, J. and G.A. Rick. Nerve ce11 density in subrnuwus plexus throughout the gut of cat and opossum. Gastruen~eroIogy 89: 10644069,1985.
Christensen, J. Ongin of sensation in the esophagus. [ReMew]. An. J . PhysioI. 246G221- G225, 1984.
Christensen, J., Eck, G.A, Robinson, B.A., Stiles, M.J. md M A Wur Arrangement of the myenteric plexus throughout the gastrointestinal tract of the opossum. Ga~lroenteroIogy 85: 890-899, 1983.
Christensen, J. and RL. Roberts. Differences between esophageai body and lower esophageal sphincter in mitochondria of smooth muscle in oposnim. Gastroenterology 85: 650456, 1983.

C b r i s t ~ J. and B.A R o b * i n . Anatorny of the myenteric plexus of the opossum esophagus. Gastroenterology 83 : 1033-42, 1982.
Christensen, J. Patterns and origin of some esophageal responses to =ch and ektrical stimulation. ~ o e n t e r o I o g y Sg(6): 909-16, 1970.
Christensen, I. and G.F. Lund. Esophageal responses to distension and electrical stimulation. J. C h Invest. 48: 408-4 19,1969.
Christensen, J. and E.E. Daniel. Effects of some autonomie dmgs on ciradar esophageal smooth muscle. J. Pharmacol. W. Zkr. 159:243-249,1968.
Clerc, N. and N. Mei. Vagal mechanoreceptors located in the lower esophageal sphincter of the cat. J. Phyno. Lod- 336: 487-498, 1983.
Clouse, RE. and N.E. Diamant. Motor Physiology and Motor Disorden of the Esophagus. In: Gastroiritestinal and Liver Disease. 6" ed. Feldman, M., Scharschmidt, B.F. and M.K Sleisenger (eds). Philadelphia, W.B. Saunders, pp. 467-497, 1998.
Cobb, B. W., Gidda, J. S. and RK. Goyal. Mechanism of esophageal peristalsis: Latency gradients and slow electrical response due to vagal and intramurai stimulation. Gasttoen~eroZogy 78: 1 1 50, 1 980.
Cohm, P.I., Tremblay, L. and N.E. Diamant. The disiribution of spinal and vagai sensory neurons that innervate the esophagus of the cat. Gustruenteroiogy 1 O3 18 17-822, 1992.
Collman, PL, Tremblay, L. and NE. Diamant. Distri'bution of vagal and spinal sensory neurons to the esophagus of the cat [Abs~act]. Gostroenteroiogy 100:A432, 1991.
Cook, LJ., Dodds, W.J., Dantas, RO., Massey, B., Kem, M.K., Lang, I.M., Brasseur, J.G. and W.J. Hogan. Opening mechanism of the human upper esophageal sphincter. Am. J. Physd. 257 (5 Pt 1):G748-59, 1989.
Costa, M and S.J.H. Brookes. The Enteric Nervous System. Am. A Gmîruen~er. 89(8): S 129-S 137, 1994.
Costa, M. and J.B. Fmess. The peristaltic reflex: an anaiysis of nerve pathways and their pharmacology. Naunyn ScImriedkbergs Arch PhannoCoII 294:4760,1976.
Craemer, B. and J. Schlegel. Motor responses of the esophagus to distention. J. A& P&~oL 10: 498-504, 1957.

Crist, J., Surprenant, A and RK Goyal. Intraceilular Studies of Electrid Membrane Properties of Opossum Esophageal C i d a r Smooth Muscle. Ga~aoenterology 92:987-92, 1987.
Crist, J., Gidda, J.S. and RK Goyai. Characteristics of "on" and "off contractions in esophageal cirCU1SLi muscle in vitro. Am. J. Physioi. 224 (Gastroimest. Liver Physiol. 9): G137-Gl44,1984a
Crist, J., Gidda, J. S. and RK. Goyal. Inframurai mechanism of esophageal peristalsis: roles of cholinergie and nonchohergic nemes. Proc. Nil. A d Sci. USA. 8 1 ~3595-9, l984b.
Csendes, A, Henriquez, A and P. Csendes. Length of the humas esophagus in vivo during rnanometric studies. Diseases of the Ewphgus 6: 1 7- 1 9, 1993.
Daniel. E.E. and V-Posey-Daniel. Neuromuscular structures in opossum esophagus: roie of interstitial ceiis of Cajal. Am. J. Physiol. 246 (Gastroimest. Liver Physiol. 9): G305-G3 15, 1984.
Davison, J.S. and D. Gmdy. Modulation of singie vagal efferent fibre discharge by gastrointestinal afferents in the rat. J. PhysioZ. 28469-82, 1978.
Decarle, D.J., Szabo, AC. and J. Christensen. Temperature dependance of responses of esophageal smooth muscle to electrical field stimulation. Am. J PhysoL 232:E432-E436,1977.
Decktor, D.L. and J.P. Ryan. Trammembrane Voltage of Opossum Esophageal Smooth Muscle and Its Response to Electrical Stimulation of I a t ~ s i c Nerves. Gatroenterology 82:3O 1-8, 1 982.
DeNardi, F.G. and RH. RiddeU. The Normal Esophagus. Am J Surg. Pafhoi. 1 S(3):296- 309, 1991.
Desai, KM., Sessa, W.C. and J.R Vane. Involvem~ of niîric oxide in the reflex relaxation of the stomach to accommodate food or fluid. Ndure L o d . 351:477- 479,1991.
Diamant, N.E. Neuromuscular Mechanisms of Primary Peristalsis. Am. J. Med 1 O3(5A):4OS43 S, 1997.
Diamant, NE. Physiology of Esophaged Motor Function GatroenteeoL C h N Am. 18(2): 179-194, 1989.
Diamant, N.E. and T.Y. El-Sharkawy. Neural control of esophageal peridsis. A conceptuai anaiysis. [Review]. Gasiroenferobgy 72: 546-556, 1977.

Dodds, W. J., Dent, J., Hogan, W.J. and RC. Arndorfer. Effkct of atropine on esophageai motor fimction in h m . Am. J . Physoi- 240: G29eG296, 198 1.
Dodds, WJ., Christensen, J., Dent, J., Amdorfa, R.C. and JD. Wood. Pharmacdogical investigation ofprimary -sis in smooth muscle portion of opossum esophagus Am. J. PhysioL 237: E561-E566,1979.
Dodds, W.J., Christensen, J., Dent, J. et al. Esophageal contractions induced by vagal stimulation in the opossum Am. J. P's iol . 235: E392-E4O 1, 197841.
Dodds, W.J., S t a J.J., Stewart, E.T. et. al. Respomes of feline esophagus to &cd vagal stimulation Am. J. Physioi. 235: E63, 1978b..
Dodds, W. J., St* J. J. and W. J. Hogan. Factors determining pressure measurement a m c y by intrahiminal manometry. ~ ~ e r o l o g y 70: 1 17, 1976.
Dodds, W.J., Hogan, W.J., Reid, D.P., Stewart, E.T. and RC. Arndorfier. A cornparison between primary esophageal pexistalsis following wet and dry swdows. J. Appl. PbsioI. 35:851-857, 1973.
Dooley, C.P., Schlossrnacher, B. and J.E. Valenniela. Effects of alterations in bolus viscosity on esophageal peristalsis in humans. A m J. Physol. 254:G8, 1988.
Doty, RW. Neural organization of degiutition. In: Code CF. (ed) Handbook of Physiology, Section 6: Alimentaxy C d . Washington, DC, Amencan Physiological Society, pp. 1 86 1 - 1902, 1968.
Ellis, F.H. Upper esophageal sphincter in health and disease. Surg= Ch. N. Am. 5 1553- 565,197 1.
El-Ouazzani, T. and N. Mei. Electrop$siological properties and role of vagal themoreceptors of lower esophagus and stomach of cat. G~l~troenterolqy 83: 995- 1 O0 1, 1982.
Eslami, M.H., Richards, W.G. and D.J. Sugarbaker. Esophageal Physiology. Chest Surg. Clin. AL Am. 4(4):635-52, 1994.
Faussone-Peiiegrbi, M.-S. Comparative study of Interstitial Celis of CajaL Acta AM. 130: f 09-1 26, 1987.
FaussonePeUegrini, M.-S. and C. Cortiesim. Ultrastructural fmtures and localization of interstitiai ceiis of Cajal in the smooth muscle mat of the human esophagus J. Submimosc. C " l . 17: 1 87- 197, 1987.

Fenopiio-Reiser, C M , Lantz, P-E-, Listram, MB., Davis, M- and F.O. m e . Gastrointestinai pathology. An atlas and text. New York: Raven, 1989.
Fleshler, B., Hendrix, T.R, Kramer, P. and F.J. Ingeffiger. The characteristics and siniilarity of primary and secondary peristalsis in the esophagus. J. C h Zmest. 38: 110-1 16, 3959.
Foumet, J., Snape Jr., W.J. and S. Cohen. Sympathetic cuntrol of lower esophageal sphincter hct ion in the cat. Action of direct cervical and splanchnic neme stimulation J. Ch. Imestst 63562-570, 1979.
Fryscak, T., Zenker, W. and D. Kantner. mirent and &emt innervation of the rat esophagus. A tracing study with horseradish peroxidase and nuclear yeiiow. A m . EmbrjmIlogy 170: 63-70,1984.
Fumess, J.B. and J-C. Bernstein. The Enteric Nervous System and Its Extrinsic Connections. In: Textbook of Gastroenterology, second edition, (ed) T. Yamada. J.B . Lippincott Company, Philidelphia, pp . 2-23, 1 995.
Fumess, J.B., Johnson, P.J., Pompolo, S. and J.C. Bonistein. Evidence that enteric moùlity refiexes can be initiated through entirely intrinsic mechanisrns in the guinea-pig d intestine. Neurogastroented. Mot. 7: 89-96, 1 995.
Fumess J.B., Bomsteïn, J.C., Pompolo, S., Young, H.M., Kunze, W.A.A., Yuan, S.Y. and H. Kelly. The Nerve Circuits for Motibty Control in the Gastrointestinal Tract J. Smooth Mucle Res. 29: 143- 145, 1993.
Fumess, J.B. and M. Costa. The enteric nervous system. Edinburgh: Churchilt Livingstone, 1987.
GabeUq G. Structure of Muscles and Nerves in the Gastrointestinal Tract. In: Physiology of the GastrointestUial Tract. 3" Ed. L.R Johnson (ed.) Raven Press, New York, pp. 751-793, 1994.
Gabella, G. and D. Blundefi. Gap junctions of the muscles of the small and large intestine. Cell Tissue Res. 2 1 9:469, 1 98 1.
Gershon, M.D. Effects of tetrodotoxin on innervateci smooth muscle preparations Br. J. Phannacol C ' o t h e r . 29:259, 1967.
Gidda, J. S., Cobb, B. W. and RK Goyal. Modulation of esophaged peristalsis by vagal aerent stimulation in opossum. J. Clin Inwst. 68: 141 1-1419, 1989.
Gidda, J.S. and RK. Goyal. Regional gradient of initial inhiWhion and refiactoriness in esophaged smooth muscle. GastroenteroZogy 89:843-85 1, 1985.

Gidda, J.S. and RK. Goyal. Swaüowcvoked action potentiais in vagal preganglionic efferents. J. Neurophysioi. 52: 1 169,1180,1984.
Gilbert, RJ. and W.J. Dodds. EEect of selective muscarùllc mtagonists on peristaitic contractions in opossum smooth muscle. Am. J. Physoi. 250: G5û-G59.1986.
Ginzel, KH Investigations concerning the initiation of the peristaltic reflex in the guinea- pig ileum. J. Physioi. 148:75P, 1979.
G o d e z , M., Marin, F., Vasconex, C., Annengol, J.-R and J.-R. Malagelada Oesophageal toae in patients with achalask Gur 41 : 29 l-Z96,1997.
Goyai, RK and 1. Hirano. The Entaic Nervous System N. Engl. J. Med 334( 17): 1 106- 1115,1996.
Goyal, RK. Muscaxinic receptor subtypes, physiology and ciinical implications. N. h g l . J . Med. 321:1022-1029, 1989.
Goyal, RK. and W.G. Paterson. Esophageai motility. In: Handbook oflhysiology. Section 6: The Gastrointestinal System, (Eds) S.G. Schultz, J.D. Wood, and B.B. Rauner. Bethesda, Maryland: American Physiological Society, pp. 865-908, 1989.
Goyai, RK. and S. Rattan. Genesis of basal sphincter pressure: effe* of tetrodotoxin on lower esophage. sphincter pressure in opossum in vivo. Gastroenterology 7 1: 62- 67, 1976.
Goy& RK., Sangree, M.K and T. Hersh. Pressure inversion point at the upper high pressure zone and its genesis. Gaslioenteroio&y 59:754-759, 1970.
Grbovic, L. and B.Z. Radrnanovic. The effe* of nifidipine on the contractile responses of the longitudinal and circuiar muscle of the isolated guinea-pig ileum to various agonists. Arch int. Phucod)m. 289: 165- 176, 1987.
Gregersen, H. and G. Kassab. Biomechanics of the gastrointestinal tract. Neurqgastreenteroi. Mot- 8:277-297, 1 996.
Gregersen, H. and L.L. Cbristeasen RessureCross-Sectionai Area Relations and Elasticity in the Rabbit ûesophagus in vivo. Digestion 57: 174-179, 1996.
Gregersem, H., Giversen, LM., Rasmussen, LM. and k Tonnip. Biomechanid wall properties and collagen content in the partially obstnicted opossum esophagus. G~str0enteroIog-y 1 O3 : 1 547-5 1, 1 992.

Grider, J.R and J-G. Ji Distinct Populations of Sensory Neurons Mediate the PeristaItic Refiex Elicited by Muscle Stretch and Mucosal Stimulation. J. Neuroscience. 14(5):2854-2860,1994.
Grider, J.R Interplay of VIP and nitric oxide in the reguiation of the descendhg r e l d o n phase of peristalsis. Am. J. Physioi. 264: 6334-340, 1993.
Grider, JR and G U Makhlouf. Colonic peristaltic refiex identification of VIP as mediator of descending relaxation Am J. P@sioI. 25 1 G4O-W 15, 1986.
Grundy, D. Mechanoreceptors in the Gastrointestinal Tract. J. Smooth Muscle Res. 29: 37-46, 1993.
Heirn, J.F., Bro, S.L., Dodds, W.J., Sania, S.K. and RG. Hoffmana. Myogenic mechanisrn for peristaisis in opossum srnooth muscle esophagus. Am. J. Physol. 263 (Gastrointest. Liver Physiol. 26): G953-G959, 1992.
He& J.F., Bro, S.L., Dodds, W. J., Sama, S K , Hoflhm, RG. and RC. Arndorfer. Myogenic oscillatory rnechanism for opossum esophageal smooth muscle contractions. Am. J. Physioi. 261 (Gastrointest. Liver Physiol. 24): G377-G383, 1991.
Herbert, J.M., Augereau, J.M. and J.P. MaflJand. Chelerythrine is a potent and specific inhibitor of Protein Kinase C. Bi~chem. Biophys. Res. Corn. 1 72:993-999, 1990.
Hillemeier, C., Bitar, K.N., Soho, U. and P. Biaocani. Protein Kùuise C Mediates Spontaneous Tone in the Cat Lower Esophageal Sphincter. J. Phmm. Ep. mer. 277(1):144-149, 1996.
HïIl&neier, C., Bitar, K.N., Marshai, J.M. and P. Biancani. IntraceIfdar pathways for contraction in gastroesophageal smooth muscle cells. Am. J. Physiol. 260G770- G775, 1991
Hoiüs, J.B. and D.O. Castell. Effects of cholinergie stimulation on human esophaged peristaisis. J. Appl. Physioi- 40:40, 1976.
Hollis, J.B. and D.O. Casteli. Effect of dry and wet swdows of Merent volumes on esophageal peristalsis. J. AppL Physioi. 28: 1 16 1-1 164, 1975.
Holzer, P., Lippe, LTH., Lotfi Tabrizi, A, Lenard Jr., L., and L. Bartho. hial Excitatory and Inhiiitory E f f i of Nhic &ide on Peristalsis in the Guinea Pig Intestine. J. P h . Exp. mer. 280 (1):154-161,1997.

Hmphries, T.J. and D.O. CasteIl. Pressun profile of esophaged peristalsk in nonaal humans as mes~sufed by direct intraesophageai transducers. Am. J . Dig. Dis. 22: 641-645,1977.
hmhmi, M and K. Hama An electron microscope study on the interstitial cells of the *d of the lovebird (Uronloncha domestica). Z Zèï~orsch Mibosk A m . 97:35 1-357, 1969.
W AA, Mascolo, N., Costa, M. and F. Capassu. Effect of papverine on synaptic transmission in the guinea-pig ileum. Br. J. P h . 12 l(4): 768-772, 1997.
hssens J. and G. Vantrappen. Ambulatory rnanometxy [Review]. Minena Chi-ca 46(7 Suppl):53-5, 1991.
lanssens, J., DeWever, I., VanTrappen, G. and J. Heiiemans. Peristalsis in smooth muscle esophagus d e r m o n and bolus deviation. WoenteroIoay 7 1 : 1004, 1976.
Janssens, J., Valembois, P., Vantmppen, G., Hellemans, J. and W. Pelernans. 1s the pNaary peristaltic contraction of the canine esophagus bolus-dependent? Gastroenteroiogy 65: 750-756, 1973.
Jean, A Brainstem organization of the swallowing network. Brain Beha. Evol. 25: 109, 1984.
Kahrilas, P.J., Longemann, J.A, Lin, S. and G.A. Ergun. Phaiyngeal clearance during swdowing: a combined manomeaic and videofiuoroscopic study. Gclstroenterology 103: 128, 1992.
Kahrilas, P.J. Functional anatomy and physiology of the esophagus. In: Caste& D.O., ed. n e fiophagus. Boston: Little, Brown and Company, pp. 1-27, 1992.
Ka& E., Ferro, P.A, Cherksey, B.D., Sughnori, M., Llinas, R and O.D. Uchitel. Effect of ca2* channel blockers on transmitter release and presynaptic currents at the frog neuromuscular junction. 1 Physoi. (Zond). 486(Pt 3):695-706, 1995.
Konturek, S.K. and P.C. Konturek. Role of nitric oxide in the digestive system. Digesth 56(1):1-13, 1995.
h v i t z , J.J., Snape, W.J. and S. Cohen. EEect of thoracic vagotomy and vagal stimulation on esophageal fiinction Am. J. PIysioI. 234: E359-E364, 1978.
Kunze, W A A , Bernstein, J-C. and J.B. Fumess. Identification of sensory nerve cds in a penpheral organ (the intestine) of a m d . Neut.oscience 66(1): 1-4, 1995.

Le& B.F. Abdominal and pelvic visceral receptors. Br. Med Bull. 33 3: 163-168, 1977.
Lerche, W. The esophagus and pharynx in action. Springf~eld, IL: Charles C. Thomas, 1950.
Li, Q., Caste& J.A. and D.O. Casteli. Manometric determination of esophageai length. Am. J . Gastroenterol. 89:722,1994.
Liebermami-Meffert, D., Allgower, M., Schmid, P. and AL. Blum. Musailas equivalent of the lower esophageal sphincter. GastjwnteroIogy 76: 3 1-3 8, 1 979.
Long, J.D. and RC. Orlando. N o r d Anatomy and Developmentai Anomalies. In: Gastrointestinal and Liver Disease. 6& ed. Feldman, M., Scharrchmidt, B.F. and M.H. Sleisenger (eds). Philadelphia, W.B. Saunders, pp. 457-466, 1998.
MacGdchrist, A.J., Christensen, J. and G A Rick. The d i s t r i i o n of myeiinated nerve fibers in the mature opossum esophagus. J. Auton. Nerv. Syst 3 5:227-236, 1987.
M a k h l o ~ G.M. Smooth Muscle of the Gut. In: Textbook of Gastroenterology, 2' ed. Tadataka Yamada (ed. ). Lippincott Company, Philadelphia, pp. 86-1 10, 1995.
Marklin, G.F., Krause, W.J. and J.H. Cutts. Structure of the esophagus in the addt opossum, Di&@his virginm. Anai. Am. 145: 249- 16 1, 1979.
Mayrand, S., Tremblay, L. and N. Diamant. In vivo measurement of fehe esophageai tone. Am. J. PhysioI- 267: G914G921, 1994.
Mayrand, S. and N.E. Diamant. Measwement of Human Esophageal Tone in =vo. Gustroenteroiogy 1 O5 : 1 4 1 1 - 1 420, 1 993.
McCleskey, E.W., Fox, AP., Feldmn, D.H., Cruz, L. J., Olivera, B.M., Tsien, R W. and D. Yoshikami. a-Conotoxj.n: direct and persistent blockade of specific types of calcium channels in neurons but not muscle. Prm. Natl. A d Sei. USA 84: 4327433 1,1987.
-UcComeI, F.M. S. Analysis of pressure generation and bolus transit during pharyngeal swallowing. Lqqgoscop 98: 71-78, 1988.
Mei, N. Disposition anatomique et proprietes electrophysi010giques des neurones sensitifs vagaux chez le chat. Exp. Braïn Res. 1 1:465-479, 1970.
Meyer, O. W., Austin, R.N., Brady, C.E. and D.O. Casteil. Muscle anatomy of the human esophagus. J. Clin. Gastroenterol. 8(2): 13 14, 1986.

Meyer, G.W., Gerhard, D.C. and D.O. Casteii. Hurnan esophageai response to rapid d o w i n g : muscle refkactory period or neural inhiition? Am J. PhysoZ. 241:G129-G136,1981.
Miftakhov, RN. and D.L. W w e . E1eçbica.i actMty of the sensory Serent pathway in the enteric nerwus system. Biol cybem. 75,471-483. 1996.
Murray, J., Du, C., Ledlow, A, Bates, J.N. and J.L. Coakliii. Nitrc oxide: mediator of nonadrenergic noncholinergic response to opossum circuiar muscle Am. J. P.sioZ.26 1 : G401, 199 1.
Neuhuber, W.L. Sensory vagd innervation of the rat esophagus and cardia: a iight and electron microscope anterograde traciug study. J. Auton New. Syst. 20243-255, 1987.
Netter, F.H. Digestive system: upper digestive tract. In: The CIBA Coilection of Medid Illustrations, Ed. E. Oppenheimer. New York: CIBA Publishing, 1971.
Nguyen, P. and D.O. Casteii. Stimulation of esophageai mechanoreceptors is dependent on rate and duration of distention. Am. J. Physiol. 267 (Gastrointest. Liver Physiol. 30): G115-G118, 1994.
Ny, L., Alm, P., Larsson, B., Ekstrom, P. and K.E. Andersson. Nitric oxide pathway in cat esophagus: localization of nient oxicie synthase and fhctiond ef f is . Am. J. Phyno. 268:G59-70, 1995.
Olivera, B .M., Rivier, J-, Clark, C., Ramilo, C.A., Corpuz, G.P., Abogadie, F.C., Mena, E.E., Woodward, S-R, HiIlyard, D.R. and L. J. C u . Diversity of C m s Neuropeptides. Science 249: 257-263, 1990.
Orvar, K.B., Gregersen, H. and J. Christensen. Biomechanid Characteristics of the Human Esophagus. Dig. Dis. Sci. 3 8(2): 1 97-205,1993.
Paterson, W.G. and B. Indrakrishnan. Descendhg perïstaltic reflex in the opossum esophagus. Am. J. PhysioI. 269 (Gastrointest. Liver Physiol. 32): G2 19-G224, 1995.
Paterson, W.G. Neuromuscular mechanisms of esophageai responses at and proximal to a disteadhg baiIoon. Am. J. PhysoZ. 260 (Gastrointest. Liver Physiol. 23): G148- G155,1991.
Paterson, W.G., Hynna-Liepert, T.T., and M. Selucky. Cornparison of primary and secondary esopbigeal peristalsis in humans: Effêct of atropine. Am. J. PhysoZ. 260: GS2, 199 1 a-

Paterson, W-G-, Selucky, M and T.T. Hynna-Liepert. Effect of Intraesophageal location and muscarinic blockade on bdoon distention-induced chest pain. Dig. DD Ski. 36(3):282-288, 199 1 b.
Paterson, W.G. Eiectncal Correlates of Peristaltic and Nonperistaltic Contractions in the Opossum Smooth Muscle Esophagus. GastrwnteroI' 97:665-75, 1989.
Paterson, W.G., Rattan, S. and RK. Goyai. Esophageal responses to transient and sustained esophageal distension. Am. J . Physioi. 255 (Gastrointest. Liver Physiol. 18): G587-G595,1988.
Peden, IX, Schneider, M.D. and RD. Bickel. Anatornic relations of the vegus nerves to the esophagushagus Am J. Surg. 80: 32-34,1950.
Perachia, C. S t r u d correlates of gap junction permeation. Int. Rw- C'ol . 66: 8 1, 1980.
Preiksaitis H.G. and N.E. Diamant. Phasic contractions of the muscuiar components of human esophagus and gastroesophageal junction in vitro. Cm. J PhysioZ. Phmmo~oi. 73: 3 56.363, 1995.
Rao, S.S.C., Hayek, B. and R-W. Summers. Impedance Planimetry: An Integrated Approach for Assessing Sensory, Active, and Passive Biomechanical Properties of the Human Esophagus. Am. J. GatroentetoL 90(3), 1995.
Rattan, S., Gidda, J.S. and RK. Goyal. Membrane potential and mecahnical responses to vagal stimulation and swallowing. Gastroenterology 859220928, 1983.
Rattan, S., Grady, M. and RK. Goyal. Vasoactive intestinal peptide causes peristaltic contractions in the esophageal body. Lue Sci. 30: 1557-1563, 1982.
Reynolds, R P. E., El-Sharkaway, T.Y. and N.E. Diamant. Oesophageal penstalsis in the cat: the role of central innemation assessed by transient vagal blockade. Am. J. Physiol. 246: 66666674, 1984.
Richards, W.G. and D.J. Sugarbaker. Neuronal CoritroI of Esophageal Function Chesi Surg. Clzn N. Am. 5(1), 1995.
Rodrigo, J., Pol* J.M., Fenuuidez, L., Ghatei, MA, Muldberry, P., and S.R Bloom. Calcitonin gene-related peptide immunoreactive sensory and motor nerves of the rat, cat, and monkey esophagus. G~z~roeteroIogy 88:Wl-M 1,1985.

Rodrigo, J., de Filipe, J., Robles-Chillida, E.M., Perez Anton, J.A and A Gomez. Sensory vagal nature and anatomid access pgths to esophageal Iaminar nerve endings in myenteric ganglia Det ermination by sufgïcai degaieration methods. Acta A m 1 12147-57, 1982.
Rodngo, J., Heraandez, C-J., Vida2 M-A and J.A Pedrosa. Vegetative innervation of the esophagus. II. Intraganglïonic lamlliar endings. Ac& AMt. 92: 79-100, 1975.
Ryan, J.P., Snape Jr., W. J. and S. Cohen Inhience of vagal cooliag on esophageal f k t i o n Am. J. PhysioZ. 232: El 59-El 64,1977.
Saha J.K., Sengupta, J.N. and RK. Goyal. Role of chlonde ions in lower esophageai sphincter tone and relaxation, Am. J. PhysioZ. 263(1 Pt 1):G115-26, 1992
Sarna, S.K., Daniel, E.E. and W.E. Waterfiill. Myogenic and neural control systems for esophageal motiIity. G&oenteroZogy 73 : 1345- 13 52, 1977.
Seelig, L.L., Jr., Doody, P., Brainard, L., Gidda, J.S. and RK. Goyal. Acetylcholinesterase and choline acetyltraasfera~e staniùig of neurons in the opossum esophagus. Anat. Rec. 209: 125- 130,1984.
Seelig, L.L., Jr. and RK. Goyal. Morphological evaluafion of opossum lower esophageal sphincter. G~uenterology 75: 5 1 -58, 1 978.
Sengupta, J.N., Saha, J.K. and RK. GoyaI. Stimulus-Response Function Studies of Esophageal Mechanosensitive Nociceptors in Sympathetic Merents of Opossum J. Neurop@s. 64(3): 796-8 12, 1990.
Sengupta, J.N., Kauvar, D. and RK- Goyal. Characteristics of Vagal Esophageal Tension- Sensitive AfEerent Fibers in the Opossum. J. of Neurophys. 6 l(5): 1 O0 1 - 1 O 1 O, 1989.
Sengupta, A., Paterson, W.G. and RK. Goyal. Atypical Localization of Myenteric Neurons in the Opossum Lower Esophageal Sphincter. Amen'can Jamml of Anaiomy l80:342-348, 1987.
Shelton Jr., RL., Bosma, J.F. and B.V. Sheeîs. Tongue, hyoid and larymc displacement in swaüow and phonation. J Appï. PhysioZ. 15(3):283-288, 1960.
Sims, S.M., Jiao, Y. and HG. Preiksaitis. Regdation of intraceiidar calcium in human esophageai smooth muscles. Am J. Physiol. 273 (Celi Physiol. 42):Cl679-C 1689, 1997.

Singaram, C. and A Sengupta. fIistopathology ofthe enteric neuropathies: From silver staining to Mmunohistochemimy. Gastroenterd C h N. Am. Vol. 25, No. 1, 1996.
Singamm, C., Sengupta, A, Sweet, M.A, Sugarbaker, D.J. and RK Goyd. Nitrinergic and peptidergic innervation of the hurnan oesophagus. G& 3 5: l690-1696,1994.
Singaram, C., Sengupta, A, Sugarbaker, D. J., and RK. Goyal. Peptidergic innervation of the human esophageal smooth muscle. Ga~troenterology 101 : 1256-1 263, 199 1.
Srnith, T.K., Bornstein, J.C. and J.B. Fumess. Convergence of reflex pathways exciteci by distension and mechanical stimulation of the mucosa on to the same myenteric neurons of the guinea-pig s d intestine. J. Nmosc i . l2(4): 1502, 1992.
Smith, T.K., Bomstein, J.C. and J.B. Fumess. Interactions between refiexes evoked by distention and mucosal stimulation: electrophysiological -dies of guïnea-pig ileum. J. Auton. Nem. Syst. 34:69-76, 1991.
Smith, T.K., Bornstein, J-C. and J.B.Furness. Distension-evoked escenduig and descendhg reflexes in the Circuiar muscle of the guinea-pig ileum. J. Auton. Nerv. Sys. 29:203, 1990.
Smith, T.K. and J.B. Fumess. Reflex changes in cimilar muscle activity eiicited by stroking the mucosa. an electrophysiological anaiysis in the isolated guinea-pig ileum. J. Auton. Nen? Syst 22:205-2 18, 1988.
Smout, AJ.P.M, Devore, M.S., Dalton, C.B. and D.O.Castell. Effects of Nifedpine on Esophageal Tone and Perception of Esophageai Distension. Dig. Dis. Sci.
a 37(4): 598-602, 1992.
Sobotta, J. and F. Hammersen. Histology, Color Atlas of Mîcroscopic hatomy. 3rd ed. Munich: Kastner & Cdwey, 1985.
Sohn, U.D-, Zoukhri, D., Dartt, D., Sergheraert, C., Hatmett, KM., Behar, J. and P. Biancaai. Diffièrem Protein Kinase C Isozymes Mediate Lower Esophageal Sphincter Tone and Phasic Contraction of Esophageal Circular Smooth Muscle Mole& P h 5 1 :462-470,1997.
Sohn, U.D., Chiu, T.T., Bitar, K.N., Eliliemeier, C., Behar, J. and P. Biancani. Calcium requirernents for acetyicholine-indu& coutraction of cat esophageai c irdar muscle celis. Am 1 Physol. 266: G33043338, 1994.
Sohn, U.D., Harnett, KM., De Petris, G., Behar, J. and P. Biancani. Distinct Muscarhic Receptors, G Roteins and Phosphoiipases in Esophageal and Lower Esophageal Sphincter Circular Muscle. J. P b Exp. mer. 267 (3): 1205- 12 14, 1993.

Somlyo, AP. Excitation contraction couphg and the ultrastructure of smooth muscle. Circ. &S. 57:497, 1985.
Song, 2.-M., Brookes, S.J.H and M Costa. Projections of specifïc morphological types of neurons within the myenteric plexus of the d intestine of the guinea-pis Cell TiweRes 285:149-156, 1996.
Stanley, E.F. The calcium c h e l and the organization of the presynaptic transmitter release fice- Trends NWOSCI~. 20: 404-409,1997.
StendaI, C. Practicai Guide to Gastrointestinal Function Testing. Biackweii Science Ltd, pp. 225-23 1, 1997.
Stevens, C.E. and AF. Sellers. Rumination- Ln: Handbook of Physiology. Afimentary Canai. Bile; Digestion; Ruminal Physiology, edited by CF. Code. Washingîon, DC: American Physiological Society, sect. 6, vol. V, pp. 2699-2704, 1968.
Sugarbaker, D.J. and W.G. Richards. Effects of nitric oxide synthesis inhibitors on circuiar muscle of human lower esophageal sphincter in vitro. [Abmct] Gastroenterology 102: AS2 1, 1992.
Sugarbaker, D. J., Rattan, S. and RK. Goyal. Mechanical and elecaical activity of esophageal miooth muscle during peristalsis. Am. J. Physol. 246 (Gastrointest. Liver Physiol. 9): G145-G1 50, 1984.
Surprenant, A. Control of the Gastrointestinal Tract by Enteric Neurons A m Rev. Physiol. 56: 1 17-40, 1994.
Toma, T.P., Zighelboim, J., Phillips, S.F. and N.J. Tdey. Methods for studying intestinal sensitivity and cornpliance: in vitro studies of baibons and a barostat Neurog~stroenteroi. Mot. 8: 1 9-28, 1 996.
Tonini, M. and M. Costa. A pharmacologicd analysis of the neuronal circuitry involved in distension-evoked enteric excitatory r e f l a Neuroscience 38(3): 787-795, 1990.
T@trup, A, Svane, D. and A Forman. Nitrix oxide mediating NANC inhibiion in the opossum lower esophageal sphincter. Am. J. PhysioI. 260: G385-G389, 199 1.
T4ttmp, A, Forman, A, Funch-Jensen, P., Raundahl, U. and K.E. Andersson. Effects of t r a n s m d field stimulation in isolateci muscle stnps fhm human esophagus. Am. J. Physioi- 258 (3 Pt 1): G344-51,1990.

Toyama, T., Yokoyam, 1. and K Nishi. EffecfS of hexamethonnim and other gangiionic blocking agents on dectricai activity of the esophagus induced by vagal stimulation in the dog. Eur. J PiaarmacoJ. 3 1 : 63-7lYl97S.
Van Overbeck, J.J.M., W~tt, KP., Paping, RKL. and HM Segenhout. Sirnuitanmus manometry and electromyography in the pharyngoesophageal segment. Lmyngoscope 95582, 1985.
Walsh, M.P., H o r o e A, Clemeat-Chornieme, O., Andrea, J-E, Men, B.G. and KG. Morgan. Protein kinase C mediation of ca2+-independent contractions of vascuiary smooth muscle. Biochem Cell Biol. 74:485-502, 1996.
Wattchow, D.A, Fumess, J.B., Costa, M., O'Brien, PI. and M. Peacock. Dis t r i i on of neuropeptides in the human esophagus. GastruenteeoZo~y 93: 1363- 137 1, 1987.
Welch, R W., Lukennann, K., Ricks, P.M., Drake, S.T. and GA Gates. Manometry of the normal upper esophageal sphincter and its alterations in laryngestomy. J. Ch. Invest. 63(5): 1036-41, 1979.
Whitehead, W.E., Delvaux, M. and The Working Tearn. Standardization of Barostat Procedures for Testing Smooth Muscle Tone and Sensory Thresholds in the Gastrointestinal Tract. Dig. Dis. Sci. 42(2):223-24 1, 1997.
Whitmore, 1. The ultrasmichiral characteristics of oesophageal striated muscle in the guinea pig and mannoset. Cell Tissue Res. 234: 365, 1983.
WinSip, D.H., Deandrade, S.R and F.F. Zboralske. Influence of bolus temperature on hurnan esophageal motor fùnction. J. Clin. Iwesr. 49: 243-250,1970.
Yamato, S., Spechler, S.J. and RK. Goyd. Role of nitrix oxide in esophageal peristalsis in the opossum. Gastroentedogy 1 O3 : 197-2O4, 1992.
Yoshida, Y., Miyadd, T., Hirano, M., S b , T., Totoki, T. and T. Kanaseki. Localization of efferent neurons innervating the pharyngeal constfictor muscles and the ceNical esophagus muscle in the cat by means of the horseradish peroxidase method. N-ci Le!?. Z(2):g 1-5, 198 1.
Yuan, S.Y., Fumess, J.F., Bomsteh, J-C. and T.K. Smith. Mucosal compression elicits polarized rekes and enhances responses of the ciradar muscle to distension in the s m d intestine. J. Auron Nem. Sys. 3 5:2 19,199 1.
IMAGE EVALUATION TEST TARGET (QA-3)
APPLlED 1 IMAGE. Inc 1653 East Main Street - -. - Rochester. NY 14609 USA
3 -- - - Phone: 71 6/482-0300 -- -- - - Fax 7t 6i288-5989
O 1993. Appüed Image. Inc.. AU RlghEI Reserved




















