
Induction of DNA Hypomethylation by Tumor Hypoxia
7
www.landesbioscience.com Epigenetics 119 ABSTRACT In cancer, the extensive methylation found in the bulk of chromatin is reduced, while the normally unmethylated CpG islands become hypermethylated. Regions of solid tumors are transiently and/or chronically exposed to ischemia (hypoxia) and reperfusion, conditions known to contribute to cancer progression. We hypothesized that hypoxic microenvironment may influence local epigenetic alterations, leading to inappropriate silencing and re-awakening of genes involved in cancer. We cultured human colorectal and melanoma cancer cell lines under severe hypoxic conditions, and examined their levels of global methylation using HPLC to quantify 5-methylcytosine (5-mC), and found that hypoxia induced losses of global methylation. This was more extensive in normal human fibroblasts than cancer cell lines. Cell lines from metastatic colorectal carcinoma or malignant melanoma were found to be markedly more hypomethylated than cell lines from their respective primary lesions, but they did not show further reduction of 5-mC levels under hypoxic conditions. To explore these epigenetic changes in vivo, we established xenografts of the same cancer cells in immune deficient mice. We used Hypoxyprobe™ to assess the magnitude of tissue hypoxia, and immunostaining for 5-mC to evaluate DNA methylation status in cells from different regions of tumors. We found an inverse relationship between the presence of extensive tumor hypoxia and the incidence of methylation, and a reduction of 5-mC in xenografts compared to the levels seen in the same cancer cell lines in vitro, verifying that methylation patterns are also modulated by hypoxia in vivo. This suggests that epigenetic events in solid tumors may be modulated by microenvironmental conditions such as hypoxia. INTRODUCTION Seventy to 80 percent of all CpG sites in human DNA are methylated at cytosine residues, generating the modified nuclide 5-methylcytosine (5-mC) 1 (Fig. 1). The meth- ylation process depends on the sum balance of several factors. These factors include several DNA methyltransferase enzymes (DNMTs), de-methylases, and methylation centers that trigger DNA methylation and are likely related to unusual tertiary DNA structures. 2-8 Classical models propose that, once established during development, DNA methylation patterns are fixed and then perpetuated through the maintenance activity of a DNMT enzyme that recognizes newly synthesized DNA as hemi-methylated and rapidly converts it to fully methylated form. 9 In cancer, the tight regulation of DNA methylation breaks down, and the distribution of 5-mC changes. Despite the hypermethylation of normally unmethylated promoter regions and the well-documented presence of increased DNMT1 activity and expression in cancer, 10-13 all types of cancer examined to date display global hypomethylation. 14-16 The processes of CpG island hypermethylation and global genomic hypomethylation seem to be very different. Researchers have characterized the functional and site-specific effects of aberrant methylation at specific CpGs in important cancer-related genes, and reported on the consequences of hypermethylation of promoter regions, however the effect of global hypomethylation and its role in cancer progression remains unclear. A global decrease in 5-mC levels is commonly observed in neoplasia, where it is an early event, as well as in aging cells. Functionally, hypomethylation may contribute to chromosomal instability in cancer and perhaps, to increased expression of selected affected genes such as the cdk inhibitor p16INK4a in primary and metastatic breast carcinoma 17 and to ectopic expres- sion of g-synuclein in cancers outside the CNS. 18 Hypomethylation has also been found to affect repeat and unique sequences in parallel, and detection of partially hypomethylated XIST alleles in prostate cancer tissues was suggested to be useful for the identification of Siranoush Shahrzad Kelsey Bertrand Kanwal Minhas Brenda L. Coomber* Department of Biomedical Sciences; University of Guelph; Guelph, Ontario Canada *Correspondence to: Brenda Coomber; Department of Biomedical Sciences; Ontario Veterinary College; University of Guelph; Guelph, Ontario, N1G 2W1 Canada; Tel.: 519.824.4120; Fax: 519.767.1450; Email: [email protected] Original manuscript submitted: 02/12/07 Revised manuscript submitted: 06/11/07 Manuscript accepted: 06/14/07 Previously published online as an Epigenetics E-publication: http://www.landesbioscience.com/journals/epigenetics/abstract.php?id=4613 KEY WORDS 5-methylcytosine, cancer, hypomethylation, hypoxia, tumor microenvironment, ischemia, HPLC ABBREVIATIONS 5-mC 5-methylcytosine HPLC high performance liquid chromatography DNMT DNA methyltransferase enzyme CRC colorectal cancer LOI Loss of imprinting ACKNOWLEDGEMENTS We are grateful for financial support from the Canadian Cancer Society/National Cancer Institute of Canada (CCS/NCIC) and the Cancer Research Society (CRS) for this study. HPLC facilities and technical support were kindly provided by Investigative Science Incorporated (ISI), Burlington, ON, Canada. We would also like to thank Alyssa Foulkes and Barb Mitchell, University of Guelph, for assistance with mouse husbandry, and Dr. Senji Shirasawa, International Medical Center of Japan, for providing the Dks-8 cell line. [Epigenetics 2:2, 119-129; April/May/June 2007]; ©2007 Landes Bioscience Research Paper Induction of DNA Hypomethylation by Tumor Hypoxia
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Induction of DNA Hypomethylation by Tumor Hypoxia

www.landesbioscience.com Epigenetics 119
AbstrActIn cancer, the extensive methylation found in the bulk of chromatin is reduced, while
the normally unmethylated CpG islands become hypermethylated. Regions of solid tumors are transiently and/or chronically exposed to ischemia (hypoxia) and reperfusion, conditions known to contribute to cancer progression. We hypothesized that hypoxic microenvironment may influence local epigenetic alterations, leading to inappropriate silencing and re-awakening of genes involved in cancer. We cultured human colorectal and melanoma cancer cell lines under severe hypoxic conditions, and examined their levels of global methylation using HPLC to quantify 5-methylcytosine (5-mC), and found that hypoxia induced losses of global methylation. This was more extensive in normal human fibroblasts than cancer cell lines. Cell lines from metastatic colorectal carcinoma or malignant melanoma were found to be markedly more hypomethylated than cell lines from their respective primary lesions, but they did not show further reduction of 5-mC levels under hypoxic conditions. To explore these epigenetic changes in vivo, we established xenografts of the same cancer cells in immune deficient mice. We used Hypoxyprobe™ to assess the magnitude of tissue hypoxia, and immunostaining for 5-mC to evaluate DNA methylation status in cells from different regions of tumors. We found an inverse relationship between the presence of extensive tumor hypoxia and the incidence of methylation, and a reduction of 5-mC in xenografts compared to the levels seen in the same cancer cell lines in vitro, verifying that methylation patterns are also modulated by hypoxia in vivo. This suggests that epigenetic events in solid tumors may be modulated by microenvironmental conditions such as hypoxia.
IntroductIonSeventy to 80 percent of all CpG sites in human DNA are methylated at cytosine
residues,generatingthemodifiednuclide5-methylcytosine(5-mC)1(Fig.1).Themeth-ylationprocessdependsonthesumbalanceofseveralfactors.ThesefactorsincludeseveralDNAmethyltransferaseenzymes(DNMTs),de-methylases,andmethylationcentersthattrigger DNA methylation and are likely related to unusual tertiary DNA structures.2-8Classicalmodelsproposethat,onceestablishedduringdevelopment,DNAmethylationpatterns are fixedand thenperpetuated through themaintenance activityof aDNMTenzymethatrecognizesnewlysynthesizedDNAashemi-methylatedandrapidlyconvertsittofullymethylatedform.9
Incancer,thetightregulationofDNAmethylationbreaksdown,andthedistributionof 5-mC changes. Despite the hypermethylation of normally unmethylated promoterregionsandthewell-documentedpresenceofincreasedDNMT1activityandexpressionincancer,10-13alltypesofcancerexaminedtodatedisplayglobalhypomethylation.14-16TheprocessesofCpGislandhypermethylationandglobalgenomichypomethylationseemtobeverydifferent.Researchershavecharacterizedthefunctionalandsite-specificeffectsofaberrantmethylationatspecificCpGsinimportantcancer-relatedgenes,andreportedontheconsequencesofhypermethylationofpromoterregions,howevertheeffectofglobalhypomethylationanditsroleincancerprogressionremainsunclear.Aglobaldecreasein5-mClevelsiscommonlyobservedinneoplasia,whereitisanearlyevent,aswellasinaging cells. Functionally, hypomethylation may contribute to chromosomal instabilityincancerandperhaps,toincreasedexpressionofselectedaffectedgenessuchasthecdkinhibitorp16INK4ainprimaryandmetastaticbreastcarcinoma17andtoectopicexpres-sionofg-synucleinincancersoutsidetheCNS.18Hypomethylationhasalsobeenfoundtoaffectrepeatanduniquesequencesinparallel,anddetectionofpartiallyhypomethylatedXISTallelesinprostatecancertissueswassuggestedtobeusefulfortheidentificationof
siranoush shahrzad Kelsey bertrand Kanwal Minhas brenda L. coomber*
Department of Biomedical Sciences; University of Guelph; Guelph, Ontario Canada
*Correspondence to: Brenda Coomber; Department of Biomedical Sciences; Ontario Veterinary College; University of Guelph; Guelph, Ontario, N1G 2W1 Canada; Tel.: 519.824.4120; Fax: 519.767.1450; Email: [email protected]
Original manuscript submitted: 02/12/07 Revised manuscript submitted: 06/11/07Manuscript accepted: 06/14/07
Previously published online as an Epigenetics E-publication:http://www.landesbioscience.com/journals/epigenetics/abstract.php?id=4613
Key words
5-methylcytosine, cancer, hypomethylation,hypoxia, tumormicroenvironment, ischemia,HPLC
AbbrevIAtIons
5-mC 5-methylcytosineHPLC highperformanceliquid chromatographyDNMT DNAmethyltransferaseenzymeCRC colorectalcancerLOI Lossofimprinting
AcKnowLedgeMents
WearegratefulforfinancialsupportfromtheCanadian Cancer Society/National CancerInstitute of Canada (CCS/NCIC) and theCancerResearchSociety(CRS)forthisstudy.HPLC facilities and technical support werekindly provided by Investigative ScienceIncorporated(ISI),Burlington,ON,Canada.We would also like to thank Alyssa FoulkesandBarbMitchell,UniversityofGuelph,forassistance with mouse husbandry, and Dr.SenjiShirasawa,InternationalMedicalCenterofJapan,forprovidingtheDks-8cellline.
[Epigenetics2:2,119-129;April/May/June2007];©2007LandesBioscience
Research Paper
Induction of DNA Hypomethylation by Tumor Hypoxia

120 Epigenetics 2007;Vol.2Issue2
caseswithpronouncedhypomethylation,whichtendto be more aggressive.19 Decreased methylation ofDNAcanleadtolossofimprinting(LOI)andthiscandrivecellularproliferationincancer.Theclearestexample of this phenomenon is the LOI at theIGF2/H19regionasaresultofhypomethylationatthedifferentiallymethylatedregionofIGF2,aneventseeninabout40%ofcolorectalcarcinomas.20.21
The presence of both methylated and unmethylated DNAsequences within the same tumor sample suggests that epigen-etic changes in the tumor cells may differ based on exposures todifferent tumor microenvironmental conditions such as hypoxia/ischemia.22,23Ithaslongbeenknownthatassolidtumorscontinuetoexpand,therateofangiogenesisisusuallyinadequatetoprovideuniformperfusiontothegrowingmass.24Tumorvasculatureisalsoabnormal, highly disorganized and irregular, with poor mural cellrecruitment, disrupted hierarchical branching, and inconsistentflowincompressedoroccludedsegments.25,26Theneteffectisthatregions of solid tumors are transiently and/or chronically exposedto ischemia and reperfusion, with all of its attendant sequelaeof hypoxia/anoxia, followed by relative hyperoxia, fluctuation innutrient(especiallyglucose)levels,acidosisandotherdisruptionsinpH, and toxic reactive oxygen species generation.Aswell as beingmutagenic,23,27 such microenvironmental conditions may disruptDNA methylation patterns, providing a potential link betweenextracellularenvironment,epigeneticalterations,andcancerprogres-sion.Wethereforehypothesizedthatprogressionofsolidcancersis,atleastinpart,influencedbylocalepigeneticalterations(changeinDNAmethylationpatterns)relatedtovascularproximityandtumorperfusion,leadingtoinappropriatesilencingandawakeningofgenesinvolvedincancer.Hereweemployedcellcultureandexperimentaltumor systemsand investigated towhatextentglobalhypomethyl-ationisrelatedtothepresenceofhypoxiainthemicroenvironmentofsolidtumors.
MAterIALs And MetHods Cell lines. Humancolorectalcancer(CRC)celllines,HCT116,
DLD-1, SW480, and SW620 and melanoma cell lines, WM115andWM239wereobtained fromATCC.Dks-8wasaK-raswild-typesublineofDLD-1[28].TheSW480celllinewasderivedfroma primary lesion (Dukes stage B colon carcinoma) from a 50-yearold Caucasian male patient. The SW620 cell line was culturedfrom a lymph node metastasis in the same patient at a later time[29,30].WM239celllinewasestablishedfromametastatichumanskin lesion,whereas theWM115cell linewas isolated froma lateverticalgrowthphaselesion[31-33].HumandermalfibroblastswerepurchasedfromATCCandusedasnon-cancercontrolcells.
In vitro exposure to microenvironmental conditions. Cellsweremaintainedinstandardcultureconditions:DMEM(Sigma,Canada)supplementedwith10%heat-inactivatedfetalbovineserum,50µg/mlgentamicin,1mMsodiumpyruvate,and1.25µg/mlamphotericinBas5µl/mlofFungizoneAntimycotic(Invitrogen/Gibco,Canada).Cellswereculturedat37˚Cinahumidifiedatmospherecontaining
5% CO2. Hypoxic conditions were achieved using a ModularIncubatorChamber(Billups-RothenbergInc.,CA,USA)modifiedtopermitcontinuousflushingofthechamberwithahumidifiedmixtureof95%N2and5%CO2.Theoxygencontentinthechamberwaskeptatlessthan0.1%inallhypoxiaexperiments.Thus,theinvitroexperimentswereinfactperformedinseverehypoxia,whichmightbe considered anoxia.Confluentmonolayerswere trypsinized, and2x106cellswereseededinto100mmplates,whichwereincubatedundernormalcellcultureconditionsovernight.Thereafter,theplateswere assigned to twogroups, control andhypoxia, and exposed totheseconditionsfor24h.Aftertheincubation,cellsweretrypsinized,pelleted,andwashedtwiceusingalargevolumeofPBS,pelletedandkeptat-80˚CuntilDNAisolation.Eachexperimentwasperformedinduplicateandrepeatedatleastthreetimes.
In vivo evaluation in human tumor xenograft model.Allinvivoprocedureswereperformedaccordingtotheguidelinesandrecom-mendations of the Canadian Council of Animal Care (CCAC),under the supervision of the local Animal Care Committee. Celllines HCT116, DLD-1, SW480, SW620, Dks-8, WM115 andWM239 were used for parallel cell culture and subcutaneousimplantation of 1–2 x 106 cells (in 100µl of 0.1% BSA in PBS)into the flank of immune deficient RAG1- mice.34Tumor growthwasmonitoredusingVerniercalipersandvolumedeterminedbythestandard formula (length x width2 x 0.5);23 tissues were harvestedonce tumor volume reached approximately 100–300 mm3. Micewere euthanizedbyCO2asphyxia followedbycervicaldislocation.Forhypoxiadetection,micewereinjectedi.p.with150mg/kgbodyweightHypoxyprobe-TM(ChemiconInternationalInc.,Temecula,CA) one hour prior to euthanasia.Tumors were removed and cutintopieces,whichwere then snap-frozen in cryomatrix and storedat-80˚Cfor latersectioning,orsnap-frozenin liquidnitrogenandstoredat-80˚CforsubsequentDNAisolation.
Cell sorting.The tumorcellsmostproximal toperfusedvesselswere labeled by a cardiac injection of 0.25 mg/mouse Hoechst33342 dye in 200µl volume, five minutes before euthanizing thexenograftbearingmice.35,36Thisprocedureyieldshighlyfluorescentcells immediatelysurroundingthevasculatureandlowfluorescenceintensityinmoredistalhypoxicareas.Afterenzymaticdisaggregationofeachtumorintoasingle-cellsuspension,samplesweresortedbyfluorescence-activated cell sorting (FACS), and the cells displayingthe 5% highest and 5% lowest Hoechst fluorescence intensitieswerecollectedandstainedforhypoxiaand5-mClevelsasdescribedbelow.
Immunofluorescence double staining for 5-mC and hypoxic adducts.WemodifiedtheprotocoldescribedbyBenchaibetal.37toimmunostainsectionsofcancercellxenografts,andisolatedsuspen-sionsofcells.SixµmcryosectionsofHCT116,DLD1,WM115andWM239 tumors were air-dried at room temperature (RT) for 15minutes,thenfixedin4%paraformaldehydefor12minutesatRT
Figure 1. Molecular Structure of 5-methylcytosine paired with guanine, each connected to their backbone made of sugar and phosphate groups.
TumorHypoxiaLeadstoChangesinDNAMethylation
www.landesbioscience.com Epigenetics 121
TumorHypoxiaLeadstoChangesinDNAMethylation
andtreatedwith1%TritonX(inPBS)for20min.ToensurethatmethylatedDNAwasaccessibletoantibodies,theDNAwasdena-tured insituwith4NHClatRTfor15min.Afterdenaturation,thesampleswereblockedwith0.4%BSAovernightat4˚C,andthenincubated with mouse anti-5-mC primary antibody (Calbiochem,1:1000) for threehoursat37˚C.Thiswas followedbyanti-mouseCy3 secondary antibody incubation (1:200) (Sigma-Aldrich) forone hour at RT. Anti-HypoxyprobeTM-1 antibody (ChemiconInternationalCorp.)wasdirectlylabeledwithAlexaFluor488usingZenonAlexaFluor488MouseIgG1LabelingKit,accordingtothemanufacturer’s instructions (Invitrogen Canada Inc., Burlington,ON)andused at1:25dilution.Cell suspensions from four sortedpopulations(asdescribedabove,Section2.4)wereusedtogeneratecytospin slides, which were stained using the same protocol astissue cryosections. Images were captured using a Leica Opti-Techepifluorescence microscope equipped with appropriate excitationandemissionfilters.ImagesweremergedusingAdobePhotoshop5.0(Adobe).ControlslidesreceivedPBSinplaceofprimaryantibody.The immunostained cells were classified into three groups: strong,weak,andnostainingfor5-mC,andanaverageofatleast25cellsfromeachexperimentwerescoredinablindedfashion.
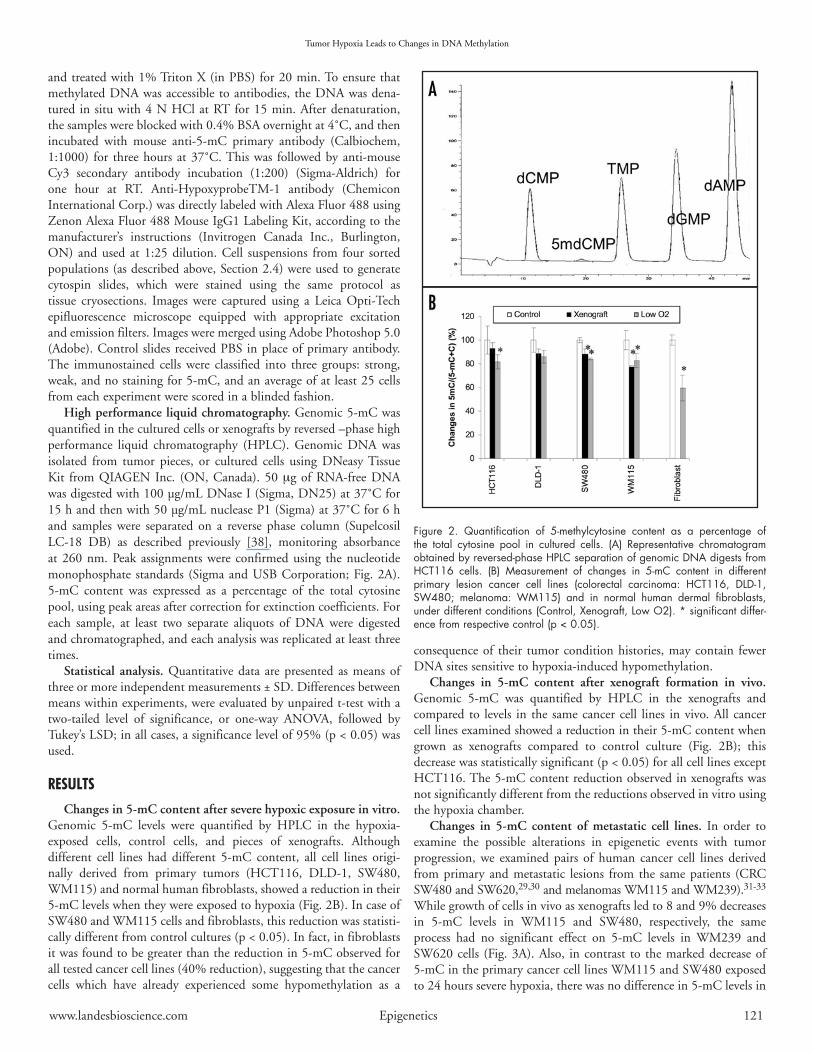
High performance liquid chromatography.Genomic5-mCwasquantifiedintheculturedcellsorxenograftsbyreversed–phasehighperformance liquid chromatography (HPLC). Genomic DNA wasisolated from tumor pieces, or cultured cells using DNeasyTissueKit fromQIAGENInc. (ON,Canada).50µgofRNA-freeDNAwasdigestedwith100µg/mLDNaseI(Sigma,DN25)at37˚Cfor15handthenwith50µg/mLnucleaseP1(Sigma)at37˚Cfor6hand samples were separated on a reverse phase column (SupelcosilLC-18 DB) as described previously [38], monitoring absorbanceat260nm.Peakassignmentswere confirmedusing thenucleotidemonophosphate standards (SigmaandUSBCorporation;Fig.2A).5-mC content was expressed as a percentage of the total cytosinepool,usingpeakareasaftercorrectionforextinctioncoefficients.Foreach sample, at least two separate aliquots of DNA were digestedandchromatographed,andeachanalysiswasreplicatedatleastthreetimes.
Statistical analysis.Quantitativedata arepresented asmeansofthreeormoreindependentmeasurements±SD.Differencesbetweenmeanswithinexperiments,wereevaluatedbyunpairedt-testwithatwo-tailed level of significance, or one-way ANOVA, followed byTukey’sLSD;inallcases,asignificancelevelof95%(p<0.05)wasused.
resuLts
Changes in 5-mC content after severe hypoxic exposure in vitro. Genomic 5-mC levels were quantified by HPLC in the hypoxia-exposed cells, control cells, and pieces of xenografts. Althoughdifferent cell lines had different 5-mC content, all cell lines origi-nally derived from primary tumors (HCT116, DLD-1, SW480,WM115)andnormalhumanfibroblasts,showedareductionintheir5-mClevelswhentheywereexposedtohypoxia(Fig.2B).IncaseofSW480andWM115cellsandfibroblasts,thisreductionwasstatisti-callydifferentfromcontrolcultures(p<0.05).Infact,infibroblastsitwasfoundtobegreaterthanthereductionin5-mCobservedforalltestedcancercelllines(40%reduction),suggestingthatthecancercells which have already experienced some hypomethylation as a
consequenceof their tumorconditionhistories,maycontain fewerDNAsitessensitivetohypoxia-inducedhypomethylation.
Changes in 5-mC content after xenograft formation in vivo. Genomic 5-mC was quantified by HPLC in the xenografts andcompared to levels in the samecancercell lines invivo.All cancercelllinesexaminedshowedareductionintheir5-mCcontentwhengrown as xenografts compared to control culture (Fig. 2B); thisdecreasewasstatisticallysignificant(p<0.05)forallcelllinesexceptHCT116.The5-mCcontentreductionobservedinxenograftswasnotsignificantlydifferentfromthereductionsobservedinvitrousingthehypoxiachamber.
Changes in 5-mC content of metastatic cell lines. In order toexamine the possible alterations in epigenetic events with tumorprogression, we examined pairs of human cancer cell lines derivedfromprimary andmetastatic lesions from the samepatients (CRCSW480andSW620,29,30andmelanomasWM115andWM239).31-33Whilegrowthofcellsinvivoasxenograftsledto8and9%decreasesin 5-mC levels in WM115 and SW480, respectively, the sameprocess had no significant effect on 5-mC levels in WM239 andSW620cells (Fig.3A).Also, incontrast to themarkeddecreaseof5-mCintheprimarycancercelllinesWM115andSW480exposedto24hoursseverehypoxia,therewasnodifferencein5-mClevelsin
Figure 2. Quantification of 5-methylcytosine content as a percentage of the total cytosine pool in cultured cells. (A) Representative chromatogram obtained by reversed-phase HPLC separation of genomic DNA digests from HCT116 cells. (B) Measurement of changes in 5-mC content in different primary lesion cancer cell lines (colorectal carcinoma: HCT116, DLD-1, SW480; melanoma: WM115) and in normal human dermal fibroblasts, under different conditions (Control, Xenograft, Low O2). * significant differ-ence from respective control (p < 0.05).

TumorHypoxiaLeadstoChangesinDNAMethylation
122 Epigenetics 2007;Vol.2Issue2
themetastasisderivedcells(WM239andSW620)betweenhypoxia-exposedandcontrolcultures(Fig.3B).Nevertheless,5-mClevelsinthemetastasis-derivedcellswereonly60%and78%oftheirrespec-tiveprimarycelllines,WM115andSW480.
Dual immunofluorescence staining of 5-mC and hypoxic adducts.Weuseddualimmunofluorescencestainingtoexaminethepatterns of 5-mC levels in relation to hypoxic areas of the tumorxenografts in both tissue cryosections, and in cells sorted fromhypoxic and well-oxygenated regions. As demonstrated in Figure4, protein adducts are onlydetected inpatches that correspond toregions of tissue hypoxia. In contrast, 5-mC is detectable in allregionsof tumor sections,but shows lower intensityof staining inthehypoxicareasofthetumors.
TheHoechst33342injectionproceduredescribedaboveproducedahighlyfluorescentpopulationofcellsimmediatelysurroundingthevasculature(Brightcells),andweaklyfluorescentpopulationofcellsin more distal tumor regions (Dim cells).The cells displaying the5% highest and 5% lowest Hoechst fluorescence intensities werecollected from tumors, digested into single cell suspensions andsortedbyFACS(Fig.5)andthenco-stainedforhypoxicadductsand5-mClevels.Theoverlayofthetwoimagesconfirmedaninverserela-tionshipbetweenglobal5-mCpresenceandcellspositiveforhypoxicadducts(Fig.5A-D),reflectingthefindingsofinsitustainedtumorpiecesandof5-mClevelsincellsexposedtohypoxiainvitro.
Quantificationof stainingpatterns in these sorted cells demon-strateda significantlyhigherproportionofcellswithnodetectable5-mC in theDim sort compared to theBright sort, and a signifi-cantlyhigherproportionof cellswith strong5-mCstaining in theBrightsortcomparedtotheDimsort(p<0.05;Fig.5E).
dIscussIon
GlobalDNAhypomethylation throughout the cell genome is acommoncharacteristicofcancercellDNA,withthedegreeofhypo-methylation increasing from normal through benign, primary and
secondarymalignancyinsometypesofcancer.39-41GlobalDNA hypomethylation in many cases affects the genomeat large and often results in a decreased overall contentof5-mC.It isoftennotclearexactlywhichsequencesaresubjecttoglobalhypomethylation,norarethemechanismsthat trigger this event well understood. Nutritional statusis known to influence global DNA hypomethylation. Forexample, dietary methyl deficiency causes DNA hypo-methylation,whichincaseofshort-termexposures,canbemostlyreversedbyphysiologicalintakesofamethyldonorsuchasfolicacid.42-44However,theinfluenceofnutritionongene-specificDNAmethylationinhumansisnotaswellstudied.
In cancer, the tumor microenvironment promotesincreasedmutagenesisaswellasselectingforcellswhichareresistanttothesephysiologicalstresses,therebycontributing
Figure 3. Measurement of 5-methylcytosine content as a percentage of the total cytosine pool in primary cancer cell lines, melanoma WM115 and colorectal SW480 and their metastatic counterparts WM239 and SW620 under different conditions (A) Control and xenograft and (B) Control and Low O2. * significant difference from respective control (p < 0.05).
Figure 4. Immunofluorescence staining of cryosections from colorec-tal carcinoma (Dks-8) and melanoma (WM115) xenografts. (A and D) Immunofluorescence staining of 5-methylcytosine; (B and E) Detection of hypoxic areas (green) by anti-HypoxyprobeTM-1 immunostaining in the same field; (C and F) overlay of the two channels demonstrating an inverse relationship between global 5-methylcytosine presence and hypoxic regions of the tumor; aster-isks mark regions of hypoxia within each section.

www.landesbioscience.com Epigenetics 123
TumorHypoxiaLeadstoChangesinDNAMethylation
totumorprogression.23,27UsingBigBlue®ratderivedcelllines;45,46wepreviouslyshowedthatmutationrateintheλ-shuttlecIItrans-geneincreasedincellsrecoveredfromxenografts,comparedtotheirinvitrocounterparts.47Furthermore,numerousstudieshavedemon-strated that hypoxia is an especially important mutagenic tumormicroenvironmentalcondition.23,47,48Nevertheless,theinfluenceoftumor microenvironment in general and hypoxia in particular, onDNAmethylationpatternsisstillunknown.
This study is therefore the first toexamine the linkagebetweenabnormal DNA methylation in solid tumors and under ischemia-likeconditionsinvitro.DNA5-mClevelswereexaminedinhumanCRC and melanoma cell lines exposed to hypoxic environments,eitherincellculture,orinregionsoftumorsxenograftedtoimmunedeficientRAG-1nullmice.Bothgrowthasxenograftsandinvitroexposuretolowoxygenledtodecreasesof5-mClevelsintheprimarycancercells.Inagreementwiththeseobservationshypoxiaalsoledtodecreaseof5-mClevelof fibroblasts inculture indicating that thiseffect is general and independent of other abnormalities of cancercells.
Chawla et al.49 tested the effect ofhypoxiaonhepaticDNAmethylationinrat.Theymaintainedyoungratsin10%oxygenatmospherefortendaysandmeasuredDNAmethylation in terms of incorporation of[3H] methyl of S-adenosyl-L-methionine(SAM). The methylases mediated incor-poration of radioactivity (a measure ofCpG specificity) in hypoxic DNA wasabout nine fold more than in the controlliver DNA. This finding is in agreementwith our results, although the conditionsemployedintheirstudyforinvivohypoxiacan be considered chronic, mild, hypoxia.Our experiments were acute (24 h) andperformedunderseverehypoxicconditions(lessthan0.1%oxygen)inordertomimicthe highly hypoxic conditions present insolid tumors.50-54 There is evidence that
substantial yet highly heterogeneous changes in tumor blood flowand pO2 can occur with different periodicities and magnitudes indifferenttumortypesandindifferentregionsofthesametumor.Theseverity of hypoxia (the absolute level of oxygen)within tumors ishighlyvariable,rangingfromnormoxictocompletelyanoxic.55
Ithasbeendemonstrated that impairment in theproductionofSAM,themajormethylatingagent,alsooccursduringhypoxia.49,56-58SAMisanabundantmethyldonorinhumanmetabolism,involvedinmorethan100methyltransferreactions,includingDNAmethyla-tion, and is synthesized frommethionine andATPby the enzymemethionine adenosyltransferase (MAT).59,60 It has been reportedthat in rat hepatocytes cultured under low oxygen levels, MAT isinactivated in an NO synthase-dependent fashion.57 While this isatbestanindirectapproachtoevaluatethelevelsofDNAmethyla-tionunderhypoxia,thesefindingssuggestthatareductioninSAMlevels in hypoxic conditions may be at least partly responsible forthesignificantlylowerlevelsof5-mCobservedinourreport.Thus,limited availability of SAM during hypoxia may be a cause of theincreaseinunmethylatedsitesonDNA.
Figure 5. Hypoxia and 5-methylcytosine in cells sorted from xenografted tumors. I) Histogram of UV Hoechst 33342 intensity FACS analysis of unsorted cell suspension of a Dks-8 xeno-graft. II) Histogram of sorted “Dim” cells, and III) Histogram of sorted “Bright” cells; A-D) Immunostaining of sorted “Dim” cells for DNA (DAPI) to indicate nuclei (A); hypoxic adducts (green) by anti-HypoxyprobeTM-1 (B); immuno-fluorescence staining of 5-mC (red) (C); overlay of hypoxic adduct and 5-mC images (D). Arrows indicate hypoxic cells with reduced 5-mC stain-ing. E) Quantification of 5-mC staining (mean ± SD) of counts performed on immunostained “Dim” and “Bright” cells for 4 different sorted tumors. *significantly higher number of cells with undetectable 5-mC in the Hoechst “Dim” population compared to the “Bright” population (p< 0.05). § significantly higher number of cells with high levels of 5-mC in “Bright” population vs “Dim” (p < 0.05).

TumorHypoxiaLeadstoChangesinDNAMethylation
124 Epigenetics 2007;Vol.2Issue2
Hypoxia exposure increased expression of IGF2 in xenograftedtumors and in cultured cell lines.61 This report and the findingsof Cui et al.,20 who demonstrated that the IGF2 gene undergoeshypomethylationandlossofimprintingincolorectalcancer,maybelinkedthroughourfindingsthathypoxiainducesDNAhypometh-ylation.Furthermore,hypomethylationwasfoundtoberesponsiblefor the increased expression of the cdk inhibitor p16INK4a inprimary and metastatic breast carcinoma,17 and hypoxia has alsobeenreportedtostimulatep16expression.62Ourresultssuggestthathypoxia-inducedhypomethylationofp16genemayplayamajorroleincancerprogression.
Inourexperiments,theprimarycancercelllinesshowlesssensi-tivitytolowoxygen(inregardtomethylationchanges)comparedtothenormalhuman fibroblasts.Thisobservation, togetherwithourresults thathypoxiahadnoeffecton theglobalmethylation statusof metastatic cancer cells, suggests that there is a limited pool ofCpGsitessensitivetohypoxiainduceddemethylation.Ifthosesiteshavebeenalreadyaffectedduringcancerprogression(suchasinthecaseofprimaryversusmetastaticcancercells),theywouldnotshowsensitivity tohypoxia.Dual immunofluorescencestainingof frozensectionsorsuspensionsortedcellsfromxenograftsalsosupportedourhypothesisthatthereisaninverserelationshipbetweenhypoxiaand5-mCcontent.
Incontrast tothemarkeddecreaseof5-mCcontent inprimarycancercellsafterxenograft formation, thisconditiondidnotaffectthe 5-mC content of metastatic cells, perhaps for reasons outlinedabove.
Interestingly, there are some cells in our fluorescence-sortedsamples that show robust hypoxic adduct staining despite beingobtainedfromtheBrightpopulationoftumorcellsuspensions.ThisisareflectionofthefactthatHoechststainingisaveryrapidevent,whileformationanddestructionofhypoxicadductstakesconsider-ably longer. Since vascular flow in solid tumors is heterogeneouswithrapidfluctuations,50-54re-establishmentofflowtoapreviouslyischemic region just prior to injection of Hoechst would have theapparentlyparadoxicaleffectof labelinghypoxiccellsbrightlywiththis perfusion marker. Conversely, stasis in a blood vessel duringthefewminutesofHoechstcirculationcouldproducetheoppositeeffect-acellfreeofhypoxicadductstainingfromthedimlylabeled(poorlyperfused)population.However,theseeventsarerare,andthequantificationof5-mCstaining incells fromeither sortedpopula-tion, combined with the in-situ co-staining of 5-mC and hypoxicadductsrevealsthatthereisashiftin5-mCcontentofcellsbetweenhypoxicandnormoxiccompartmentsofxenograftedtumors.
Inconclusion,ourstudieshaveuncoveredevidencethatoneofthepossiblemechanismsforregulationofDNAmethylationincanceristheischemic/hypoxicconditionfoundinsolidtumors.Sinceoursisthefirststudytodemonstratethat5-mClevelsalterwithhypoxia,wehavenoinformationatpresentonthekineticsofsuchchanges.Itispossiblethathypomethylationoccursviaanactive(andpresumablyrapid)processor through lackofmaintenancemethylation (whichwouldpresumablybemuchslower)orsomecombinationof these.Thisaspectoftheepigeneticresponseiscurrentlyunderinvestigationin our laboratory. Extensive demethylation of DNA during tumorprogression couldbe a source of the continually generated cellulardiversityassociatedwithcancer.Modulationofthesetissueenvironmentsmayprovideapromisingapproachtocontrollingepigeneticeventsinprogressionofsolidtumorsandcarriesimportantimplicationsfor
applying anticancer therapies involving tumor hypoxia (e.g., anti-angiogenesis)incancers.
References 1. EhrlichM,Gama-SosaMA,HuangLH,MidgettRM,KuoKC,McCuneRA,GehrkeC.
Amountanddistributionof5-methylcytosineinhumanDNAfromdifferenttypesoftis-suesofcells.NucleicAcidsRes1982;10:2709-21.
2. BirdA.DNAmethylationpatternsandepigeneticmemory.GenesDev2002;16:6-21. 3. SmithSS,KaplanBE,SowersLC,NewmanEM.Mechanismofhumanmethyl-directed
DNAmethyltransferaseandthefidelityofcytosinemethylation.ProcNatlAcadSciUSA1992;89:4744-8.
4. XieS,WangZ,OkanoM,NogamiM,LiY,HeWW,OkumuraK,LiE.Cloning,expres-sionandchromosomelocationsofthehumanDNMT3genefamily.Gene1999;236:87-95.
5. FremontM,SiegmannM,GaulisS,MatthiesR,HessD,JostJP.DemethylationofDNAbypurifiedchickembryo5-methylcytosine-DNAglycosylaserequiresbothproteinandRNA.NucleicAcidsRes1997;25:2375-80.
6. BhattacharyaSK,RamchandaniS,CervoniN,SzyfM.AmammalianproteinwithspecificdemethylaseactivityformCpGDNA.Nature1999;397:568-9.
7. MummaneniP,WalkerKA,BishopPL,TurkerMS.Epigeneticgeneinactivationinducedbyacis-actingmethylationcenter.JBiolChem1995;270:788-92.
8. Wilkins JF. Genomic imprinting and methylation: epigenetic canalization and conflict.TrendsGenet2005;21:356-65.
9. Turker MS, BestorTH. Formation of methylation patterns in the mammalian genome.MutatRes1997;386:103-5.
10. Robertson KD, Keyomarsi K, Gonzales FA, Velicescu M, Jones PA. Differential mRNAexpressionofthehumanDNAmethyltransferases(DNMTs)1,3aand3bduringtheG(0)/G(1)toSphasetransitioninnormalandtumorcells.NucleicAcidsRes2000;28:2108-13.
11. EadsCA,DanenbergKD,KawakamiK,SaltzLB,DanenbergPV,LairdPW.CpGislandhypermethylationinhumancolorectaltumorsisnotassociatedwithDNAmethyltransfer-aseoverexpression.CancerRes1999;59:2302-6.
12. DeMarzoAM,MarchiVL,YangES,VeeraswamyR,LinX,NelsonWG.Abnormalregu-lationofDNAmethyltransferase expressionduring colorectal carcinogenesis.CancerRes1999;59:3855-60.
13. Issa JP, Vertino PM, Wu J, Sazawal S, Celano P, Nelkin BD, Hamilton SR, Baylin SB.IncreasedcytosineDNA-methyltransferaseactivityduringcoloncancerprogression.JNatlCancerInst1993;85:1235-40.
14. HermanJG,BaylinSB.Genesilencingincancerinassociationwithpromoterhypermeth-ylation.NEnglJMed2003;349:2042-54.
15. JonesPA,BaylinSB.Thefundamentalroleofepigeneticeventsincancer.NatRevGenet2002;3:415-28.
16. Dunn B K. Hypomethylation: One Side of a Larger Picture. Ann N Y Acad Sci 2003;983:28-42.
17. VanZeeKJ,CalvanoJEandBisognaM.Hypomethylationandincreasedgeneexpressionofp16INK4ainprimaryandmetastaticbreastcarcinomaascomparedtonormalbreasttissue,Oncogene1998;16:2723-27.
18. YanagawaN,TamuraG,HondaT,EndohM,NishizukaS,andMotoyamaT.DemethylationofthesynucleingammageneCpGislandinprimarygastriccancersandgastriccancercelllines.ClinCancerRes2004;10:2447-51.
19. LanerT,SchulzWA,EngersR,MullerM,FlorlAR.HypomethylationoftheXISTgenepromoterinprostatecancer.OncolRes2005;15:257-64.
20. Cui H, Onyango P, Brandenburg S, Wu Y, Hsieh CL, Feinberg AP. Loss of imprintingin colorectal cancer linked to hypomethylation of H19 and IGF2. Cancer Res 2002;62:6442–6.
21. WongJJ,HawkinsNJ,WardRL.Colorectalcancer:amodelforepigenetictumorigenesis.Gut2007;56:140-8.
22. Yu JL, Rak JW, Carmeliet P, Nagy A, Kerbel RS, Coomber BL. Heterogeneous vasculardependenceoftumorcellpopulations.AmJPathol2001;158:1325-34.
23. ShahrzadS,QuayleL,StoneC,PlumbC,ShirasawaS,RakJW,CoomberBL.Ischemia-induced K-ras mutations in human colorectal cancer cells: role of microenvironmentalregulationofMSH2expression.CancerRes2005;65:8134-41.
24. FolkmanJ.Tumorangiogenesis:therapeuticimplications.NEnglJMed1971;285:1182-6. 25. CarmelietP,JainR.Angiogenesisincancerandotherdiseases.Nature2000;407:249-57. 26. JainRK.Molecularregulationofvesselmaturation.NatMed2003;9:685-93. 27. Papp-SzaboE,JosephyPD,CoomberBL.Microenvironmentalinfluencesonmutagenesis
inmammaryepithelialcells.IntJCancer2005;116:679-85. 28. ShirasawaS,FuruseM,YokoyamaN,SasazukiT.Alteredgrowthofhumancoloncancercell
linesdisruptedatactivatedKi-ras,Science1993;260:85-8. 29. FoghJ,FoghJMandOrfeoT.Onehundredandtwenty-sevenculturedhumantumorcell
linesproducingtumorsinnudemice.JNatlCancerInst1977;59:221-26. 30. Leibovitz A, Stinson JC, McCombs WB III, McCoy CE, Mazur KC and Mabry ND.
Classificationofhumancolorectaladenocarcinomacelllines.CancerRes1976;36:4562-69. 31. HerlynM,ThurinJ,BalabanG,BennicelliJL,HerlynD,ElderDE,BondiE,GuerryD,
NowellP,ClarkWH,KoprowskiH.Characteristicsofculturedhumanmelanocytesisolatedfromdifferentstagesoftumorprogression.CancerRes1985;45:5670-76.
32. HerlynM,KathR,WilliamsN,Valyi-NagyI,RodeckU.Growthregulatoryfactorsfornor-mal,premalignant,andmalignanthumancellsinvitro.AdvCancerRes1990;54:213-34.

www.landesbioscience.com Epigenetics 125
TumorHypoxiaLeadstoChangesinDNAMethylation
33. Rodeck U, Herlyn M, Menssen HD, Furlanetto RW, Koprowski H. Metastatic but notprimarymelanomacelllinesgrowinvitroindependentlyofexogenousgrowthfactions.IntJCancer1987;40:687-90.
34. ChenJ,ShinkaiY,YoungF,AltFW.Probing immune functions inRAG-deficientmice,CurrOpinImmunol1994;6:313-19.
35. Yu JL, Rak JW, Carmeliet P, Nagy A, Kerbel RS, Coomber BL. Heterogeneous vasculardependenceoftumorcellpopulations.AmJPathol2001;158:1325-34.
36. Yu JL, Rak JW, Coomber BL, Hicklin DJ, Kerbel RS. Effect of p53 status on tumorresponsetoantiangiogenictherapy.Science2002;295:1526-8.
37. BenchaibM,AjinaM,LornageJ,NiveleauA,DurandPandGuerinJF.QuantitationbyimageanalysisofglobalDNAmethylationinhumanspermatozoaanditsprognosticvalueininvitrofertilization:apreliminarystudy.FertilSteril2003;80:947–53.
38. RamsahoyeBH.Measurementofgenome-wideDNAcytosine-5methylationbyreversed-phasehigh-pressureliquidchromatography.MethodsMolBiol2002;200:17-27.
39. Gama-SosaMA,SlagelVA,TrewynRW,OxenhandlerR,KuoKC,GehrkeCW,EhrlichM.The5-methylcytosinecontentofDNAfromhumantumors.NucleicAcidsRes1983;11:6883-94.
40. GoelzSE,VogelsteinB,HamiltonSR,FeinbergAP.HypomethylationofDNAfrombenignandmalignanthumancolonneoplasms.Science1985;228:187-90.
41. KimYI,GiulianoA,HatchKD,SchneiderA,NourMA,DallalGE,SelhubJ,MasonJB.GlobalDNAhypomethylationincreasesprogressivelyincervicaldysplasiaandcarcinoma.Cancer1994;74:893-9.
42.PufuleteM,Al-GhnaniemR,KhushalA,ApplebyP,HarrisN,GoutS,EmeryPW,SandersTA.Effectof folic acid supplementationongenomicDNAmethylation inpatientswithcolorectaladenoma.Gut2005;54:579-81.
43. ChristmanJK,SheikhnejadG,DizikM,AbileahS,WainfanE.Reversibilityofchangesinnucleicacidmethylationandgeneexpressioninducedinratliverbyseveredietarymethyldeficiency.Carcinogenesis1993;14:551-7.
44. PogribnyIP,RossSA,WiseC,PogribnaM,JonesEA,TryndyakVP,JamesSJ,DraganYP,PoirierLA.IrreversibleglobalDNAhypomethylationasakeystepinhepatocarcinogenesisinducedbydietarymethyldeficiency.MutatRes2006;593:80-7.
45. McDiarmid HM, Douglas GR, Coomber BL, Josephy PD. Epithelial and fibroblast celllines cultured from the transgenic BigBlue rat: an in vitro mutagenesis assay. Mutat Res2001;497:39-47.
46. McDiarmid HM, Douglas GR, Coomber BL, Josephy PD. 2-Amino-1-methyl-6pheny-limidazo[4,5-b]pyridine (PhIP)-induced mutagenesis in cultured Big Blue rat mammaryepithelialandfibroblastcells.EnvironMolMutagen2002;39:245-53.
47. Papp-Szab—,E,Josephy,PD,Coomber,BL.Microenvironmentalinfluencesonmutagen-esisinmammaryepithelialcells.IntJCancer2005;116:679-85.
48. YuanJ,NarayananL,RockwellS,GlazerPM.DiminishedDNArepairandelevatedmuta-genesisinmammaliancellsexposedtohypoxiaandlowpH,CancerRes2000;60:4372-76.
49. Chawla RK, Watson WH, Jones DP. Effect of hypoxia on hepatic DNA methylationand tRNAmethyltransferase in rat: similarities toeffectsofmethyl-deficientdiets. JCellBiochem1996;61:72-80.
50. Kimura H, Braun RD, Ong ET, Hsu R, Secomb TW, Papahadjopoulos D, Hong K,Dewhirst MW. Fluctuations in red cell flux in tumor microvessels can lead to transienthypoxiaandreoxygenationintumorparenchyma.CancerRes1996;56:5522–8.
51. DewhirstMW,BraunRD,LanzenJL.TemporalchangesinpO2ofR3230ACtumorsinFischer-344rats.IntJRadiatOncolBiolPhys1998;42:723–6.
52. BraunRD,LanzenJL,DewhirstMW.FourieranalysisoffluctuationsofoxygentensionandbloodflowinR3230Actumorsandmuscleinrats.AmJPhysiol1999;277:H551-68.
53. BrurbergKG,GraffBA,RofstadEK.Temporalheterogeneityinoxygentensioninhumanmelanomaxenografts.BrJCancer2003;89:350–6.
54. BrurbergKG,GraffBA,OlsenDR,RofstadEK.Tumor-linespecificpO2fluctuationsinhumanmelanomaxenografts.IntJRadiatOncolBiolPhys2004;58:403–9.
55. ChaplinDJ,DurandRE,OlivePL.Acutehypoxiaintumors:implicationsformodifiersofradiationeffects.IntJRadiatOncolBiolPhys1986;12:1279–82.
56. Perez-MatoI,CastroC,RuizFA,CorralesFJ,MatoJM.MethionineadenosyltransferaseSnitrosylationisregulatedbythebasicandacidicaminoacidssurroundingthetargetthiol.JBiolChem.1999;274:17075-9.
57. AvilaMA,CarreteroMV,RodriguezEN,MatoJM.Regulationbyhypoxiaofmethionineadenosyltransferaseactivityandgeneexpressioninrathepatocytes.Gastroenterology1998;114:403-7.
58. BlaiseS,AlbertoJM,NedelecE,AyavA,PourieG,BronowickiJP,GueantJL,DavalJL.Mild neonatal hypoxia exacerbates the effects of vitamin-deficient diet on homocysteinemetabolisminrats.PediatrRes2005;57:777-82.
59. KotbM,GellerAM.Methionine adenosyltransferase: structure and function.PharmacolTher1993;59:125-43.
60. Mato JM, Alvarez L, Ortiz P, Pajares MA. S-adenosylmethionine synthesis: molecularmechanismsandclinicalimplications.PharmacolTher1997;73:265-80.
61. HedborgF,UllerasE,GrimeliusL,WassbergE,MaxwellPH,HeroB,BertholdF,SchillingF,HarmsD,SandstedtB,FranklinG.Evidenceforhypoxia-inducedneuronal-to-chromaf-finmetaplasiainneuroblastoma.FASEBJ2003;17:598-609.
62. ZygmuntA,TedescoVC,UdhoE,KrucherNA.Hypoxia stimulatesp16expressionandassociationwithcdk4.ExpCellRes2002;278:53-60.




















