
Untitled - Limousin Cattle Breeders' Society of South Africa

JOURNAL OF AVIAN BIOLOGY 34: 198–210, 2003
Increased survival and breeding performance of double breeders inlittle penguins Eudyptula minor, New Zealand: evidence forindividual bird quality?
Edda Johannesen, Dave Houston and Janis Russell
Johannesen, E., Houston, D. and Russell, J. 2003. Increased survival and breedingperformance of double breeders in little penguins Eudyptula minor, New Zealand:evidence for individual bird quality? – J. Avian Biol. 34: 198–210.
The little penguin Eudyptula minor is unique among penguin species in being able tofledge chicks from two clutches in one breeding season. Pairs laying two clutches ina given season make a higher reproductive investment, and may be rewarded by ahigher reproductive success as they may raise twice as many chicks as pairs layingone clutch. The higher effort made by pairs laying two clutches could correlatenegatively with survival, future reproductive performance or offspring survival,indicating a cost of reproduction. Conversely, a positive relationship between thenumber of clutches produced in a given breeding season and survival, futurereproductive performance or offspring survival would indicate that birds laying twoclutches belonged to a category of birds with higher fitness, compared to birds layingonly one clutch in the season. In this study we used a long-term data set taken froman increasing population of little penguins in Otago, SE New Zealand. We modelledthe relationship between the number of clutches laid in a breeding season andsurvival probability, reproductive performance in the next breeding season and firstyear survival of offspring using capture-recapture modelling.Birds laying two clutches produced 1.7 times more fledglings during a breedingseason than pairs laying one clutch. We found that birds laying two clutches had ahigher probability of breeding in the following breeding season, a higher probabilityof laying two clutches in the following breeding season and a higher survivalprobability. There was no overall difference in post-fledging survival between theyoung of birds producing one clutch and the young of birds producing two clutches.However, the survival of young of single clutch breeders declined with laying date,whereas the young of double clutch breeders had the same survival rate irrespectiveof laying date. For a subset of data with birds of known age, we found evidence thatthe probability of laying two clutches increased with age. However, there were alsoindications for differences among birds in the tendency to lay two clutches that couldnot be attributed to age. We tentatively interpret our results as evidence of qualitydifference among little penguin breeders.
E. Johannesen (correspondence), Di�ision of Resource Ecology, Institute of MarineResearch, P.O. Box 1870 Nordnes, 5817 Bergen, Norway. E-mail: [email protected]. D.Houston, Department of Conser�ation, Box 388, Oamaru, New Zealand. J. Russell,Oamaru Blue Penguin Colony, 1 Thames St, Oamaru, New Zealand.
Life history theory (e.g. Stearns 1992) predicts thereshould be a trade off between different fitness compo-nents. This trade off is affected by population growthrate so that in increasing populations individualsshould invest in current reproduction at the expense offuture survival and reproduction. Trade offs should bemanifested as negative correlations between fitness
components, so that, for example, current reproductiveoutput should be negatively correlated with survival,future reproductive output or offspring survival. How-ever, in empirical studies from wild populations, apositive association between different fitness compo-nents is often found (e.g. Forslund and Part 1995,Viallefont et al. 1995, Cam et al. 1998, 2002, Cam and
© JOURNAL OF AVIAN BIOLOGY
JOURNAL OF AVIAN BIOLOGY 34:2 (2003)198

Monnat 2000a, b). Such positive correlations imply thatsome individuals have more resources than others. Ifthis cannot be attributed to differences in the environ-ment of different individuals, it can be due to thatdifferent individuals belong to categories that differaccording to fitness. One categorisation that can ex-plain positive correlations in fitness traits is age orexperience, as young, inexperienced individuals tend toperform less well than older individuals (e.g. Forslundand Part 1995, Cam and Monnat 2000a). However,several studies have found that age differences cannotalone account for all fitness variation among individu-als. When controlling for age and experience severalstudies have shown that some individuals consistentlyperform better than others, indicating that they are ofbetter ‘‘quality’’ (e.g. Reilly and Cullen 1981, Dann andCullen 1990, Sydeman and Eddy 1995, Hipfner et al.1999, Cam and Monnat 2000a, Cam et al. 2002).
In avian populations, age and quality has also beenadvocated as an explanation of the effect of laying dateon reproductive performance, i.e. that early breeders(higher quality or older) have a higher reproductivesuccess than late breeders (lower quality or younger;Dann and Cullen 1990, Sydeman and Eddy 1995,Hipfner et al. 1999, Lepage et al. 1999, Sandercocket al. 1999, Hansson et al. 2000, Nilsson 2000,Prevot-Julliard et al. 2001, Christians et al. 2001, Wik-tander et al. 2001). However, a direct effect of layingdate on reproductive performance caused by a seasonalchange in food availability or other environmental vari-ables, can also explain the correlation between repro-ductive success and laying date. The two types ofexplanation (i.e. age/quality versus environmental ef-fects) can only be disentangled through experimentsthat manipulate laying date (e.g. Hipfner et al. 1999,Lepage et al. 1999, Nilsson 2000, Christians et al. 2001,but see Hansson et al. 2000). However, indications ofthe direct effect of laying date versus the effect ofindividual quality on breeding performance can also beobtained by studying multibrooded species (e.g.Hochachka 1990). In multibrooded species, the con-founding of seasonal changes and quality of late andearly breeders are broken as the same pair (i.e. of thesame quality) has clutches that experience differentenvironmental conditions (early and late in the breed-ing season).
Our study is conducted on an increasing populationof little penguins Eudyptula minor. We use an eight-yeardata set from Otago, SE New Zealand to test forcorrelations between demographic parameters. The lit-tle penguin is the smallest of the penguin species and isfound breeding in colonies along the coast of southernAustralia and New Zealand (Williams 1995). Littlepenguins nest during the southern summer, althoughlaying dates within a season are not strongly synchro-nised and can vary by up to six months (Reilly andCullen 1981). The clutches of the little penguin consist
of two similar sized eggs (Williams 1995). Little pen-guins are unique among penguins in that they can raisetwo clutches during a breeding season (see e.g. Gales1985, Williams 1995). Individuals that lay two clutches(double breeders) make twice the reproductive invest-ment as individuals laying one single clutch (singlebreeders), and can be rewarded by twice the reproduc-tive success of single breeders as they can raise up tofour chicks in a season, whereas single breeders canonly raise two. The proportion of pairs laying twoclutches varies among years and geographic location. InOtago the probability of raising two clutches to com-pletion is particularly high (Gales 1985, Rogers et al.1995, Bull 2000), at other locations most birds laying asecond clutch do so because their first clutch failed.Double breeding leads to an extended breeding seasonfor a given pair (incubation takes 32 days and chickrearing a minimum of 6 weeks), especially if bothclutches are successful (Reilly and Cullen 1981).
Using our data set on little penguins, we first explorethe relationship between fitness parameters by testing ifindividuals laying one clutch or two clutches in abreeding season differ in reproductive success, survivalprobability, probability of breeding in the next seasonand probability of laying two clutches in the nextbreeding season. We model the relationships by usingmultistate capture-recapture models (Brownie et al.1993, Cam et al. 1998), taking into account that recap-ture rate might vary among groups of individuals asonly breeders in the colony are observable. Secondly,we look at the fate of the fledged young of the doubleand single breeders. Chicks from the first and secondclutches of double breeders will experience differentenvironmental conditions, and after fledging (when theyleave for sea), the young are independent of theirparents (Williams 1995) and have to rely on fat reservesuntil they acquire foraging skills and are able feedefficiently. If there is an environmental effect on postfledgling survival of little penguins, young fledged latein the season should have a lower survival probabilitythan young fledged early in the season, whereas ifdifferences in offspring survival are determined by theinvestment (e.g. parents with two clutches trade off thenumber of clutches with the effort used on each clutch)or quality of the parents (parents laying two clutchesare better parents or have better genes than parentslaying one clutch), there should be a difference insurvival of young of double and single breeders. Wetest these ideas by using Barker’s joint live dead en-counter model with individual covariates (Barker 1997).The inclusion of dead recovery information is particu-larly useful when estimating first year survival ofseabirds, because young are not regularly seen at thecolony before they start breeding (e.g. Frederiksen andBregnballe 2000). Finally, for a subset of breeders forwhich age is known, we explore whether there is an agedifference between double breeders and single breedersand further examine the repeatability in the probability
JOURNAL OF AVIAN BIOLOGY 34:2 (2003) 199
of laying two clutches. Our study colony is well suitedto study these trade-offs as all individuals breed in nestboxes, thus limiting environmental variation in breed-ing conditions among individuals, and ensuring a highprobability of observing breeders at the colony. Also,we look at trade-offs with reproductive investment(number of clutches laid), rather than the outcome ofthe investment (e.g. breeding success) that can be influ-enced by chance events (Cam and Monnat 2000b).Furthermore, since dispersal in this species seems lim-ited and because we combined several sources of data,we can study trade-offs with survival without the poten-tial confounding influence of dispersal.
Our study provides an exhaustive analysis consider-ing several aspects of trade-offs and correlations inlife-history traits. We are studying both intra-and inter-generational trade-offs as we consider offspring survivalas well as adult demographic parameters. Furthermore,we are studying correlations between fitness parametersin a non-equilibrium situation. In a growing populationlike ours, individuals are expected to trade off repro-duction at the expense of survival.
Methods
Study area and monitoring
We used data collected from Oamaru (45°07�S,170°58�E), Otago, New Zealand. Here the wildlifetourist operation ‘‘The Oamaru Blue Penguin Colony’’has been running since 1993. Penguins coming ashoreto the colony at dusk pass spectators on viewing standswith lights before reaching their breeding colony, theQuarry site. There is no difference between breedingperformance at the Quarry site and that of a controlsite (Creek) 900 m away (Houston and Russel unpubl.data), so it is unlikely that there is any negative effect oftourists on breeding performance. The Quarry site hasbeen provisioned with wooden nest boxes since 1993,when the entire area was cleared and renovated, so thequality of the different nests within the area should behomogenous. Efforts have been taken to prevent nest
predation by rats, ferrets, cats and dogs. In 1993, aferret depredated two chicks, the ferret was killed be-fore it killed any other birds. No other evidence of nestpredation of either eggs, chicks or breeders have beenobserved at the colony for the years when breeding datawas included in this study (1993–1997), although insubsequent years predation by ferrets and dogs wasobserved. The number of nest boxes has increased since1993 (Table 1) together with an increase in the numberof breeding pairs. The number of natural nests withinthis colony is believed to be zero.
The monitoring at the Quarry colony was done bychecking the nest boxes weekly. At each visit the con-tent of the nest was recorded and chicks, if present,were weighed, and banded with flipper bands whentheir flippers were properly developed. A chick wasassumed to have fledged (i.e. gone to sea) when itdisappeared from the nest after being at least 6 weeksold. The fledgling weight of a given chick was taken tobe the weight of the chick upon the last visit beforefledging. Unbanded adults were banded using flipperbands upon their first encounter. Only adult birdsincubating eggs or guarding small chicks were consid-ered breeders. Chicks and juvenile birds could not besexed. Breeding birds could be sexed using pair affilia-tions and beak morphology. However, it should benoted that this method is not 100% accurate (Rennerand Davies 1999, Hocken and Russel 2002). Onlybreeders banded as chicks could be aged, the propor-tion of breeders that could be aged increased as thestudy progressed, from 2% in 1993 to 39% in 1997. Ina given year, all breeding birds within the colony couldbe identified due to the easy access to the nest boxesand the frequent monitoring.
Apart from the Quarry site, the Creek colony (seeabove) and colonies at Taiaroa Head (80km furthersouth) have been regularly monitored since the early1990’s, together with a few irregularly visited sites alongthe Otago coastline (Dann 1994, Perriman and Steen2000). Recoveries of dead banded birds are recordedfrom all along the Otago coastline, but most dead birdshave been found around the intensively studiedcolonies. Dispersal seems limited in the little penguin:
Table 1. The number of boxes and double and single breeding pairs at the Quarry little penguin colony from 1993 to 1997. Thedouble breeding pair groups also included 7 pairs laying 3 clutches (two in 1994, four in 1995 and one in 1996). None of thesepairs managed to raise all three broods.
Single breedersDouble breedersBoxesYear
Failed first clutch Total Total
Egg stage Chick stage
1993 387 16170871994 5 1 22 21
1995 169 6 2 31 311996 169 7 1 33 491997 165 11 0 41 57
200 JOURNAL OF AVIAN BIOLOGY 34:2 (2003)

only ca. 1% of young fledged at the Quarry site andlater found breeding, bred in one of the other moni-tored colonies (Steen et al. unpubl. data), and nodispersal of breeding birds has been observed betweenbreeding sites in Otago (Johannesen et al. 2002).
Analysis
Grouping of data
Our analysis focused on the difference in fitnessparameters of two groups of little penguin breeders:double and single breeders. By using two groups we gotsample sizes that were about the same size (overall 45%double breeders) and for which the proportion of birdsbelonging to each group did not vary from year to year(Table 1; however, from 1998 to 2001, there were nodouble breeders in the population, Russell and Houstonunpubl. data). We could have refined our classificationby dividing the double breeders into those raising asuccessful first brood and those laying replacementclutches, and further divided the breeders laying re-placement clutches by the stage of clutch failure, as thetime spent on the first clutch and thus the reproductiveeffort differs between these groups. However, usingonly two groups prevented splitting up the data toomuch and we do not believe that using only two groupsbias our results.
The chance of making wrong classifications, i.e. clas-sifying birds as single breeders although they actuallylaid two clutches was calculated to be less than 4%(daily nest survival rates was 99.3%), using the Mayfieldestimator (e.g. Klett and Johnson 1982) implemented inthe program MARK (White and Burnham 1999), sincefailing to observe a clutch would mean that the clutchhad to be laid and fail between the weekly nest boxchecks.
Model selection
According to Burnham and Anderson (1998), whenanalysing observational data where many factors couldbe important, such as for capture-recapture data, modelselection using Akaikes Information Criterion, (AICvalues, or AICc, adjusted for small sample sizes) isbetter than hypothesis testing using P-values (AICc isdefined by −2 log likelihood+2K(n/(n−K−1))where K is the number of parameters in the model andn is the sample size). The model with the lowest AICc
value is the most parsimonious model that best explainsthe variation in the data. For a given study, a limitedset of models with a priori biological interpretationsshould be considered and the focus should be onparameter estimation with appropriate measures of un-certainty (Burnham and Anderson 1998). In this paper,we tried to follow this philosophy.
Breeding success and probability of double breeding
We tested whether birds laying two clutches in a givenyear were rewarded by a higher reproductive successthan birds laying only one clutch. We also tested forage differences between double and single breeders fora subset of breeders of known age. These analyses werecarried out using repeated measurements (Proc Mixedand Glimmix macro, with logistic link and binomialerror, using a compound symmetry covariance structureimplemented in SAS, Littell et al. 1996).
Correlations of fitness parameters using multistatecapture-recapture models
We used the program MARK and multistate capturerecapture models to model the correlation betweendouble breeding, survival and breeding in the next year(Brownie et al. 1993, Viallefont et al. 1995, Cam et al.1998). The breeding seasons in the five years weretreated as equivalent to five capture occasions. In agiven year, being captured was defined as being ob-served breeding in the colony. Breeding birds could bein one of two states: double breeder and single breeder(i.e. a breeder that laid one single clutch). Multistatemodels have three parameters: survival probability,transition probability, which is the probability ofchanging state, and recapture probability. All of theseparameters are modelled as dependent on the state theindividual belongs to. With the multistate approach wecould test if survival probability until the next year andthe probability of laying two clutches in the next yearwas different for birds that laid one or two clutches ina given year. As all individuals breeding in the colonycould be observed in a given year, recapture probabili-ties were considered equivalent to the probability ofbreeding the next year. We could therefore test whetherbirds laying one clutch in a given year had a differentprobability of breeding the next year than birds thatlaid two clutches in a given year.
The models used included three variables (year, sexand state) for each parameter (survival, recapture andtransition probability). Combining all three variableswith interactions for all three parameters leads to anextremely high number of possible models. We there-fore performed modelling in two steps. First, we testedfor the effect of year and sex on survival and recapturerate using capture-recapture models. We then modelledsurvival, recapture and transition probability as depen-dent on group while retaining the best sex and timestructure according to AICc for survival and recapturefrom the first step.
First year sur�i�al using Barker’s combined recaptureand reco�ery model
First year survival of fledged young was analysed usingBarker’s model implemented in the program MARK
JOURNAL OF AVIAN BIOLOGY 34:2 (2003) 201

(Barker 1997). Like the models by Burnham (1993) andCatchpole et al. (1993), Barker’s model combines liverecapture and dead recovery information. In addition,Barker’s model also allows the incorporation of liverecaptures between capture occasions. Barker’s modelhas six nuisance parameters to be estimated in additionto survival probability (recapture probability, recoveryprobability, fidelity, return rate, the probability of re-capture between capture occasions for a bird that eitherdied or stayed alive between the capture occasion;Barker 1997, White and Burnham 1999). The six nui-sance parameters were modelled according to age andyear in a separate analysis (Steen et al. unpubl. data).Survival of older age classes was treated as variable byyear, whereas first year survival was modelled accord-ing to individual co-variates and year.
We included recapture and recovery information upuntil the breeding season of 2001 for the five cohorts ofyoung that fledged in 1993–1997. This allowed at leastthree years of recovery and recapture of all cohorts.Most penguins start to breed when they are betweentwo and four years old (Reilly and Cullen 1981, Table2). As in the multistate analysis, each capture occasioncorresponds to a particular breeding season, and anindividual was considered captured when observedbreeding in the Quarry colony. Observations of non-breeding birds in the colony or at other colonies wereincluded as captures between the capture occasions. Wemodelled first year survival according to four individualco-variates for each bird previously found importantfor juvenile survival in several species of birds (e.g.Kremetz et al. 1989, Magrath 1991, Harris et al. 1994,Ringsby et al. 1998) including little penguins (Reillyand Cullen 1981): fledgling weight (W), clutch type (T;if the young originated from a double breeder or froma single breeder), laying date (D) and number offledglings in the clutch (F; 1 or 2). We also included the
effect of year (Y) in the models. We included oneinteraction term, the interaction between clutch typeand laying date, since if young of double and singlebreeders are of different quality, their survival could beaffected qualitatively different by laying date (Ringsbyet al. 1998).
In our analysis of first year survival, we treatedindividual young from the same brood as independentin the analysis of first year survival. This is reasonableif only the conditions after fledgling affects first yearsurvival, since there is no evidence that young from thesame broods stay together after fledgling. However, ifparental effects are important for first year survival, weshould have taken this dependency into account in ourmodelling. Unfortunately, taking such effects into ac-count is not currently possible when using capturerecapture models.
Results
Onset of breeding
The mean onset of breeding (laying date of the firstclutch of double and single breeders) was the 10th ofSeptember. The onset of breeding differed somewhatbetween years (F4,317=2.399, P=0.05). In 1993 layingdate was 17 days earlier (SE: 7.7 days), whereas themean laying date the other years did not differ signifi-cantly from the mean or from each other (1994: sevendays later, SE: 7.0 d, 1995: one day later, SE: 6.2 d,1996: five days earlier, SE: 5.7 d, 1997: three days later,SE: 3.9 d). No other year effects on breeding parame-ters were found from 1993–1997 (Perriman et al. 2000).
The onset of breeding, i.e. laying dates of firstclutches for single and double breeders were different,and the distribution of laying dates showed a three
Table 2. Models with AICc values, delta AICc values and number of parameters (cPar) for survival, breeding and transitionprobabilities. Additivity is denoted with + and interaction terms with ×. State has two modalities: single or double breeder.
cParDelta AICc Deviance
a) Modelling survival and recapture probabilityTransition probability: sex×state×year
1: Survival: state+year, Recapture: state 175.12621.17 232222.652: Survival: state+year, Recapture: constant 178.837
23.563: Survival: year, Recapture: state 179.751224: Survival: state×year, Recapture: state 173.0852625.95: Survival: state×year, Recapture: constant 27.11 25 176.5466: Survival: year, Recapture: constant 27.26 21 185.673
b) Modelling transition probabilitySurvival: state+year, Recapture: state7: Transition: state 0 9 184.269
10 184.2422.068: Transition: state+sex8 188.6132.259: Transition: constant
181.695123.7410: Transition: state+year11: Transition: sex 4.31 9 188.58712: Transition: sex +state+year 5.84 13 181.666
186.668116.613: Transition: year14:Transition: sex+year 8.69 12 186.641
202 JOURNAL OF AVIAN BIOLOGY 34:2 (2003)
peak distribution (Fig. 1): first the first clutches ofdouble breeders (average lay date: 31st of July, SE: 3.24d), then the clutches of single breeders (average laydate: 17th of Sept., SE: 3.24 d) and then the secondclutches of double breeders (average lay date: 3rd ofOct., SE: 3.24 d).
Causes of failure and difference in breedingsuccess between double and single breeders
Most of the eggs resulted in fledged chicks (approxi-mately 70%, 615 out of 904 eggs laid). Out of the 289eggs that did not result in a fledged young, 8% wereabandoned, 29% failed to hatch, 11% broke, 31% ofeggs disappeared, 12% of chicks disappeared beforethey were large enough to fledge, 8% of the chicks werefound dead in the nests with undetermined cause ofdeath, less than 1% were found depredated (2 chicks,see Methods) or starved (1 chick).
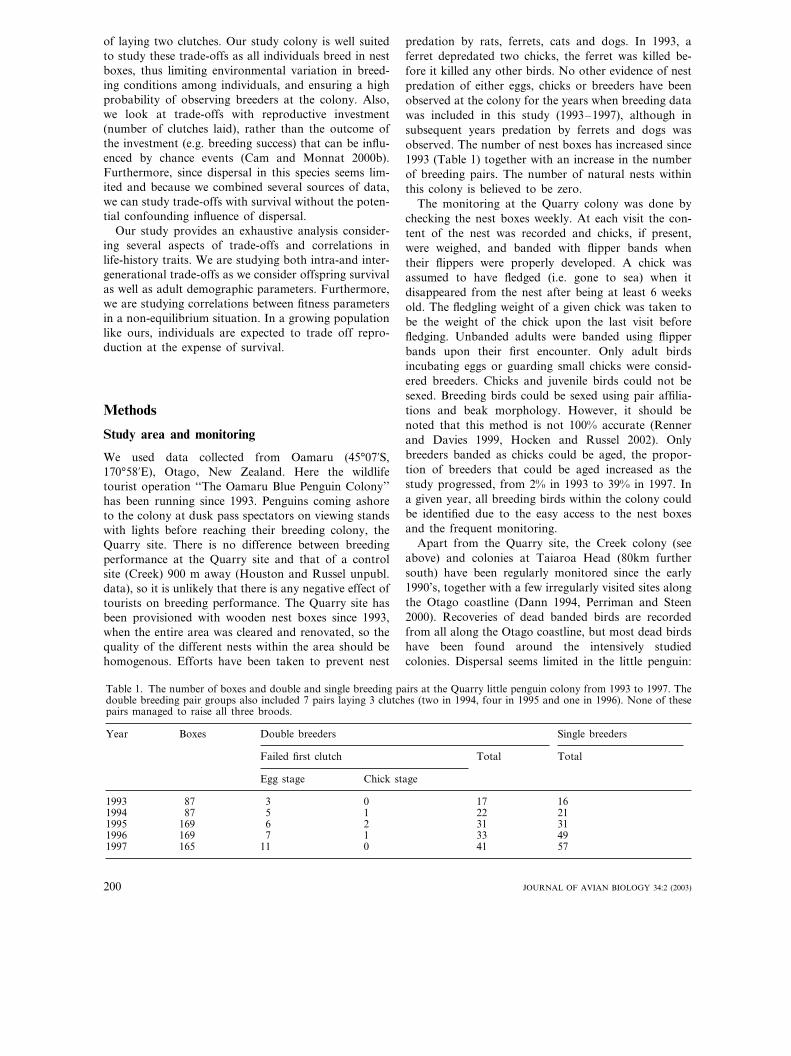
The number of fledged chicks for a double breedingpair was about 2.5 and for a single breeding pair 1.5,i.e. double breeders produced 1.7 more fledglings thansingle breeders. We found a significant difference be-tween reproductive success of double and single breed-ers using repeated measurements, with breeding pair asthe subject factor, (�2=32.7, df=1, P�0.001, n(pairs)=318; Fig. 2).
Fig. 2. The number of fledglings produced per pair as afunction of laying date. The LOESS (local linear regression)line describes the overall relationship between laying date andthe number of broods produced. Open circles represent singlebreeders and filled circles double breeders.
Correlations of fitness parameters using multistatecapture-recapture models
We first tested for sex and year variation in survivaland recapture probability (i.e. breeding probability) ofbreeders. The best model did not include any effect ofsex, but there was an effect of year on survival proba-bility (results not shown). Therefore, a model withconstant recapture and survival probability varyingwith year was used as a basis for the multistatemodelling.
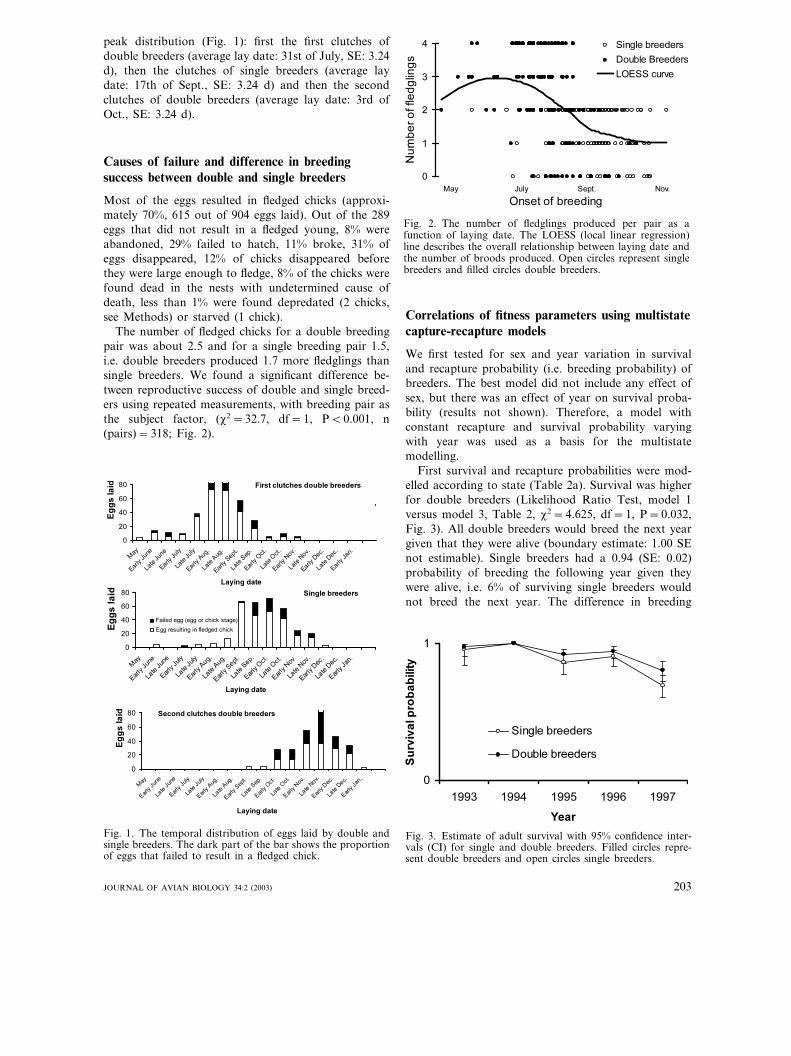
First survival and recapture probabilities were mod-elled according to state (Table 2a). Survival was higherfor double breeders (Likelihood Ratio Test, model 1versus model 3, Table 2, �2=4.625, df=1, P=0.032,Fig. 3). All double breeders would breed the next yeargiven that they were alive (boundary estimate: 1.00 SEnot estimable). Single breeders had a 0.94 (SE: 0.02)probability of breeding the following year given theywere alive, i.e. 6% of surviving single breeders wouldnot breed the next year. The difference in breeding
Fig. 1. The temporal distribution of eggs laid by double andsingle breeders. The dark part of the bar shows the proportionof eggs that failed to result in a fledged chick.
Fig. 3. Estimate of adult survival with 95% confidence inter-vals (CI) for single and double breeders. Filled circles repre-sent double breeders and open circles single breeders.
JOURNAL OF AVIAN BIOLOGY 34:2 (2003) 203

probability between single and double breeders was notquite significant (Likelihood Ratio Test, model 1 versusmodel 2, Table 2, �2=3.711, df=1, P=0.054).
Transition probability, i.e. the probability of chang-ing state, was modelled using the best model for recap-ture and survival (Table 2b). The probability of adouble breeder laying only one clutch the consecutiveyear was 0.21 (SE: 0.03). The probability for a singlebreeder laying two clutches the following year, given itwas alive and bred, was 0.30 (SE: 0.03). This result hastwo implications. First, double breeders have a higherprobability of remaining a double breeder (79%) in thefollowing year, than have single breeders of becomingdouble breeders (30%). The probability of doublebreeding for a bird that was a double breeder in theprevious year is also higher than the population mean(45%). Secondly, there was a significantly lower proba-bility for a double breeder to become a single breederthan for a single breeder to become a double breeder(Likelihood Ratio Test, model 9 versus model 7, Table2, �2=4.344, df=1, P=0.037). This means that moresingle breeders will become double breeders over timethan the reverse, implying that more individuals becamedouble breeders as they aged. The Goodness of Fit ofthe best model was: �2=36.372, df=34, P=0.35 (pro-gram MSSURVIV, Hines 1994).
First year survival of offspring from double andsingle breeders
Description of the data and the explanatory �ariables
Out of the 615 chicks fledged from 1993 to 1997, 606were banded and had been weighed just before fledging.The fate of the 606 fledged chicks included in theanalysis are summarised in Table 3.
We used four individual covariates, together withyear, when analysing first year survival (see methods).Of these, year and laying date was correlated sincebreeding began earlier in 1993 compared to the other
years. Year was unrelated to the other variables (Johan-nesen unpubl. data). Laying date differed betweenclutch types (see Fig. 1). The number of chicks pro-duced (one or two) did not differ between clutch types(probability of fledging both chicks in a successfulclutch was for single breeders 68.2% (SE: 3.7) firstclutch double breeders 74.8% (SE: 4.1), and for secondclutch double breeders 72.3% (SE: 4.2). The probabilityof fledging one chick versus two varied with layingdate, average laying date of clutches that fledged onechick was 2nd Oct. (SE: 4.6, n=103), average layingdate of clutches that fledged two chicks was 23rd Sept.(SE: 2.9, n=256). Fledgling weight did not vary ac-cording to laying date (slope estimate: 0.155, SE:0.131). The average fledgling weight for clutches whereboth chicks fledged was lower (1061.05g, SE: 7.12g,n=355, within brood averages used) than the fledglingweight for one chick clutches (1109g, SE: 13.5g,F1,355=12.75, P�0.001, see also Reilly and Cullen1981), that is, chicks from one chick clutches wereapproximately 5% heavier at fledging. Young fromdifferent clutch types had very similar fledgling weights(single clutch: 1072 g, SE: 9 g, first double: 1076 g, SE:12 g, second double: 1078 g, SE: 12 g). Fledglingweights were comparable to others studies on the samespecies (e.g. Gales 1987).
Factors determining first year sur�i�al
Year was the most important determinant of first yearsurvival as can be seen from the large jump in AICc
when including year as a variable (Table 4). The bestmodel according to AICc included an effect of year inaddition to the number of chicks fledged in the clutch,clutch type, laying date and an interaction betweenlaying date and clutch type (Table 3). However, themodels were not very well differentiated according toAICc, as models differing with less than 2 AICc unitsshould all be taken into account (Burnham and Ander-son 1998). There was an interaction between laying date
Table 3. The number of fledged chicks from 1993 to 1997 at the Oamaru Quarry colony, with the subsequent fate: the numberfound dead either younger or older than one year, the number seen alive older than one year, but not observed reproducing atthe colony, and finally the number observed reproducing at the colony. The number found reproducing later in the colony isgiven with the age distribution at first breeding in parenthesis. Note that one bird bred in its first year of life, whereas most birdsbred when two years old.
Cohort Fledged Found dead Seen alive notFound dea�1 Reproducingreproducing �1 year�1 year year
1993 67 1 4 6 22 (0, 20, 1, 0, 1, 0, 0)1994 86 0 2 9 30 (0, 25, 4, 1, 0, 0, –)
33 (0, 26, 7, 0, 0, –, –)104011819951996 157 0 5 4 31 (0, 26, 3, 2, –, –, –)1997 178 4 5 12 22 (1, 14, 7, –, –, –, –)Total 606 5* 19** 41 148
* Out of the five birds that were found dead in their first year of life, three were found away from the colony, one at the colony,and for one, the place of recovery was unrecorded.** Out of the nineteen birds found dead after their first year of life, seven were found away from the colony, eleven at thecolony, and for one, the place of recovery was unrecorded.
204 JOURNAL OF AVIAN BIOLOGY 34:2 (2003)
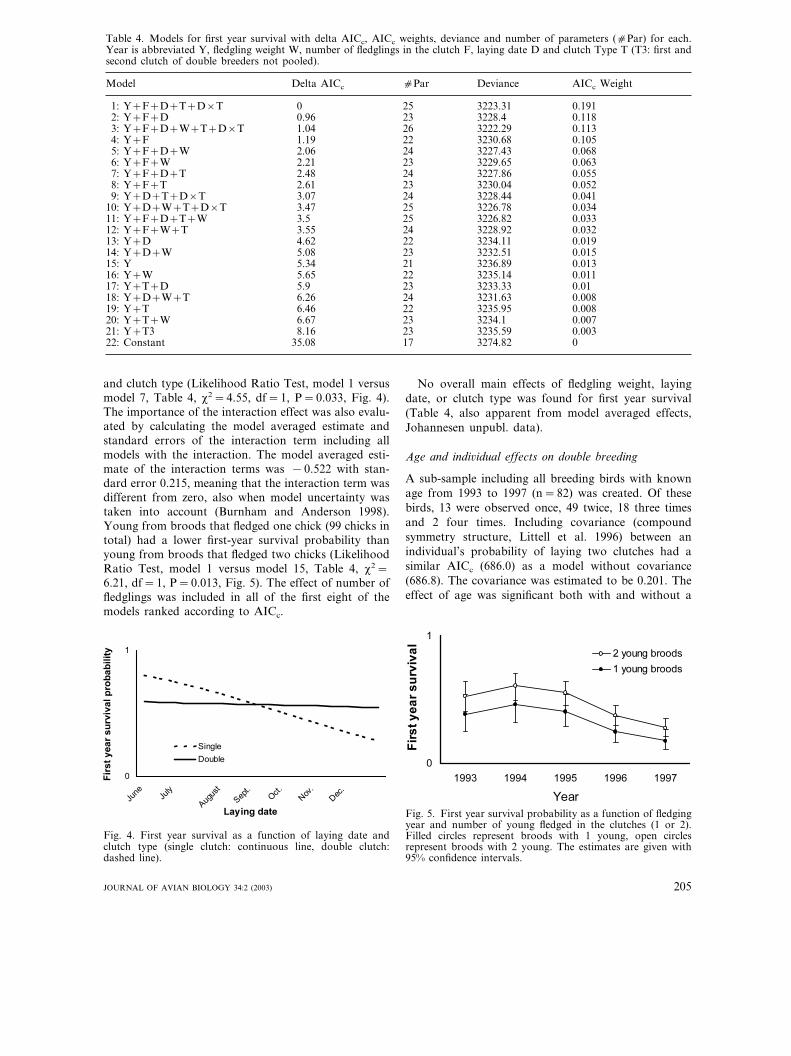
Table 4. Models for first year survival with delta AICc, AICc weights, deviance and number of parameters (cPar) for each.Year is abbreviated Y, fledgling weight W, number of fledglings in the clutch F, laying date D and clutch Type T (T3: first andsecond clutch of double breeders not pooled).
AICc WeightModel Delta AICc cPar Deviance
1: Y+F+D+T+D×T 0 0.19125 3223.310.1182: Y+F+D 0.96 23 3228.4
3: Y+F+D+W+T+D×T 1.04 0.11326 3222.294: Y+F 0.1051.19 22 3230.685: Y+F+D+W 2.06 0.06824 3227.43
0.0636: Y+F+W 2.21 23 3229.657: Y+F+D+T 2.48 0.05524 3227.868: Y+F+T 0.0522.61 23 3230.049: Y+D+T+D×T 3.07 0.04124 3228.44
10: Y+D+W+T+D×T 0.0343.47 25 3226.7811: Y+F+D+T+W 3.5 0.03325 3226.8212: Y+F+W+T 0.0323.55 24 3228.9213: Y+D 4.62 0.01922 3234.1114: Y+D+W 0.0155.08 23 3232.5115: Y 5.34 0.01321 3236.8916: Y+W 0.0115.65 22 3235.1417: Y+T+D 5.9 0.0123 3233.3318: Y+D+W+T 0.0086.26 24 3231.6319: Y+T 6.46 0.00822 3235.9520: Y+T+W 0.0076.67 23 3234.121: Y+T3 8.16 0.00323 3235.5922: Constant 35.08 17 03274.82
and clutch type (Likelihood Ratio Test, model 1 versusmodel 7, Table 4, �2=4.55, df=1, P=0.033, Fig. 4).The importance of the interaction effect was also evalu-ated by calculating the model averaged estimate andstandard errors of the interaction term including allmodels with the interaction. The model averaged esti-mate of the interaction terms was −0.522 with stan-dard error 0.215, meaning that the interaction term wasdifferent from zero, also when model uncertainty wastaken into account (Burnham and Anderson 1998).Young from broods that fledged one chick (99 chicks intotal) had a lower first-year survival probability thanyoung from broods that fledged two chicks (LikelihoodRatio Test, model 1 versus model 15, Table 4, �2=6.21, df=1, P=0.013, Fig. 5). The effect of number offledglings was included in all of the first eight of themodels ranked according to AICc.
No overall main effects of fledgling weight, layingdate, or clutch type was found for first year survival(Table 4, also apparent from model averaged effects,Johannesen unpubl. data).
Age and indi�idual effects on double breeding
A sub-sample including all breeding birds with knownage from 1993 to 1997 (n=82) was created. Of thesebirds, 13 were observed once, 49 twice, 18 three timesand 2 four times. Including covariance (compoundsymmetry structure, Littell et al. 1996) between anindividual’s probability of laying two clutches had asimilar AICc (686.0) as a model without covariance(686.8). The covariance was estimated to be 0.201. Theeffect of age was significant both with and without a
Fig. 5. First year survival probability as a function of fledgingyear and number of young fledged in the clutches (1 or 2).Filled circles represent broods with 1 young, open circlesrepresent broods with 2 young. The estimates are given with95% confidence intervals.
Fig. 4. First year survival as a function of laying date andclutch type (single clutch: continuous line, double clutch:dashed line).
JOURNAL OF AVIAN BIOLOGY 34:2 (2003) 205

covariance structure in the model (both P-values�0.001).
We looked in detail at the age specific probability ofdouble breeders by looking at the 16 males (to avoiddependence between partners in the same pair, pairfidelity was 82%) fledged in 1993 that started reproduc-ing in 1995 and reproduced in the following two years(i.e. birds with the same age and experience). Out ofthese, 12.5% were double breeders in their first year ofbreeding and second year of life (CI: 2.8–41.3). 44.8%were double breeders in their second year of breeding(CI: 21.2–69.3) and 56.3% were double breeders intheir third year of breeding (CI: 30.6–78.8), whichimplies that the probability of double breeding increasethroughout the life span, especially between the firstand second year of breeding. Also, the probability ofdouble breeding is significantly lower than the overallpopulation average (45%) in the first year of breeding.
Further, we looked in detail at the first cohort ofmale breeders. This cohort was suited for testing forrepeatability in double breeding probability becauseeach individual had five breeding opportunities duringthe years of study. Out of the 31 males breeding in1993, 23 bred every year from 1993–1997. The averageyearly probability of double breeding based on thissub-sample of birds was 61.7%. Of these males, eightwere double breeders all five years of study (three weresingle breeders all years). We found that such a highnumber of consistent double breeders would not beexpected by chance: in 1000 randomised samples of 23birds, each with five breeding opportunities and a61.7% probability of double breeding, none had eightconsistent double breeders or more.
Discussion
Positive correlations between fitness parameters oflittle penguin breeders
Confounding factors
We found a positive correlation between the number ofclutches laid, and survival until the next year, and theprobability of breeding the next year and the probabil-ity of laying two clutches the next year. A positivecorrelation between the number of clutches produced(or any other measure of breeding success) and survivalcould be an artefact of mortality during the breedingseason: if a breeder dies during the breeding season theconsequence would be reduced breeding success. Therewas little evidence for this in the present study; onlyone failed breeder disappeared during the breedingseason. Furthermore, winter is the most trying seasonfor little penguins. In winter, shorter days (little pen-guins are visual predators), together with severeweather conditions, colder water and lower foodavailability, lead to a negative energy balance (Gales
and Green 1990) and most of the mortality of littlepenguins occurs during winter (Reilly and Cullen 1979,Stahel and Gales 1987, Dann 1992, Harrigan 1992,Norman et al. 1992, Dann et al. 2000, Johannesen et al.2002).
Age and the probability of double breeding werecorrelated in our study. As more single breeders wereyoung, the correlation between the number of clutcheslaid and survival could be due to lower survival ofyounger birds. We tested for different survival of birdsaged two years and older birds using a subset of thedata (using only one year, survival from 1996 to 1997,because the proportion of young birds increased in thepopulation as the study progressed and the survivalvaried by year). There was no indication of a significantage differences in survival (�2=0.297, df=1, P=0.98,n=157, survival of birds aged two years: 92.6% andolder: 89.2%). Similarly, the only important age differ-ence in survival of little penguins is between the firstyear of life and the rest of the life span (Sidhu et al.unpubl. data from a 30 year study of little penguins,Phillip Island, Australia). Therefore, the positive corre-lations between double breeding and survival shouldnot be due to age specific survival and double breedingprobability.
Positi�e correlations between fitness parameters of littlepenguin breeders: the effect of indi�idual bird quality?
Reilly and Cullen (1981) and Dann and Cullen (1990)studied breeding success of little penguins from PhillipIsland, Australia using other methods than multi statemodels. They found positive correlations betweenfitness parameters, also when age and experienced wasaccounted for, and interpreted their results as evidencefor differences in individual quality.
Our results mirror those found by multistate mod-elling by Cam et al. (1998), see also Cam et al. 2002) fora kittiwake population. In their study the categoriesused were breeders and non-breeders, and positive cor-relations were found between present breeding perfor-mance, breeding performance the next year andsurvival. Using data from the same kittiwake popula-tion, Cam and Monnat (2000a) and Cam et al. (2002)found positive correlations between fitness parametersalso when controlling for age and experience, i.e. con-sistent differences in fitness between individuals, inter-preted as differences in individual quality. Based ontheir results, Cam et al. (2002) argued support for thehypothesis that the apparent effect of age on breedingperformance found for many animal species is due toselection (the selection hypothesis, sensu Curio 1983).According to this hypothesis, lower performance ofyounger individuals found at the population level, isdue to that lower quality individuals are removed fromthe older age classes in the population due to selectivemortality. In our study, we found evidence for age
206 JOURNAL OF AVIAN BIOLOGY 34:2 (2003)

effects on double breeding, and also some evidence forconsistent individual effects, even though the samplesize was small. We cannot easily evaluate the relativeimportance of age effects and individual quality effectsin the present study. First, the number of breeders withknown age was limited. Second, birds with known agestarted breeding toward the end of the study, so thenumber of breeding events per birds with known agewas low, making it hard to detect any individual effects.More data is needed before we draw firm conclusionsabout the relative importance of individual effects andage effects on double breeding and other correlates ofquality in the little penguins.
Cam and Monnat (2000b) discussed the problem offinding good correlates of individual quality, preferen-tially correlates that remain constant throughout thelife of an individual and divide individuals into a fewhomogenous groups. Double breeding as a correlate ofquality has some weaknesses. First, it is confoundedwith age within an individual. Second it changed withthe environmental conditions, e.g. there were no doublebreeders in the populations from 1998 to 2001 (Russelland Houston unpubl. data). Also, the two categoriesare heterogeneous, double breeders consist of bothindividuals that were successful and that failed withtheir first clutch, and furthermore, the ones that failed,failed at different stages of breeding. The decline insurvival of young of single breeders with laying date(Fig. 4) suggests heterogeneity within single breedercategory as well.
First year survival of little penguins
Determinants of first year sur�i�al
There was a lack of overall difference in first yearsurvival among young from double and single clutchesas well as an overall lack of an effect of laying date.For many single brooded species, post-fledging survivalhas been found to be influenced by laying date (Harriset al. 1994). This has also been found in penguins(Reilly and Cullen 1982, Olsson 1997). Double breedingbreaks the correlation between parent ability and layingdate, as successful double breeders have two broodsthat are fledged more than two months apart. Thesurvival of young of double breeders was not affectedby laying date. The decline with season for singlebreeders might be due to lower age or quality of latesingle breeders compared to early single breeders.
First year survival was higher for young fromclutches where both young fledged than for clutcheswhere only one young fledged. This result is similar towhat was found on shags by Harris et al. (1994), seealso references therein). Their interpretation of theirresult was that the quality of the parents differed andthat this was reflected in offspring survival. This inter-
pretation fits well with our overall results as well,although we cannot distinguish age and individualquality effects properly in our present study (seeabove).
Positive effects of fledging weight on post fledgingsurvival have been found in many bird studies, but notall (see Magrath 1991 for a review, Sagar and Horning1997 for a study on sea birds, and Ringsby et al. 1998).Differences in first year survival between early and latefledged young are often mediated through higher bodyweights of young produced early in the season (van derJeugd and Larsson 1998). We did not see a significanteffect of fledgling weight on survival, and no seasonaleffects on fledgling weight. The higher weight and lowerfirst year survival of young from clutches where onlyone single young fledged underlines the lack of impor-tance of fledgling weight on first year survival. The lackof an effect of fledgling weight on first year survival is,however, surprising, since newly fledged little penguinsmust rely on their fat reserves before learning to forageefficiently without help from the parents.
Comparison of our results on first year sur�i�al withother studies on little penguin
To our knowledge, the only other studies on first yearsurvival of little penguins are from Phillip Island, Aus-tralia. In contrast to our results, there was an effect ofboth fledgling weight and laying date in the Australianpopulation (Reilly and Cullen 1982). Chiaradia andKerry (1999) found that late-hatched chicks at PhillipIsland, Australia, did not fledge due to starvation. Incontrast, we found that most failures occurred at theegg stage (regardless of season). It is thus possible thatseasonal effects are more important for the PhillipIsland population than for the Oamaru population, inthat resources gets more limited towards the end of theseason in Phillip Island compared to Oamaru. Alterna-tively, the age or quality of parents is determiningsurvival of young at Phillip Island, but because there isno double breeding, laying date and parent quality isconfounded in a way that the result from Phillip Islandresembles our results on single breeders (see Fig. 4).Lack of an effect of laying date on first year survivaldoes not mean that season is irrelevant for biology oflittle penguin biology in the present study. Before theenergetically challenging winter, both breeding activitiesand moult (requiring two weeks of storage and two tothree weeks fasting ashore) has to be completed(Williams 1995). This whole process takes about sevenmonths for a successful double breeder. In this study,double breeders were found to be able to complete thiscycle without sacrificing either their own survival or thesurvival of their young.
Reilly and Cullen (1982) compared fledgling weightsand laying dates for young recovered alive after theirfirst year than for young recovered dead during their
JOURNAL OF AVIAN BIOLOGY 34:2 (2003) 207

first year. Their method imposes fewer constraints onthe relationship between survival, fledgling weight andlaying date than in the present study where the relation-ships were constrained to be linear. Studies on otherseabirds (e.g. Sagar and Horning 1997) have found thatthere was a critical lower threshold weight that had tobe attained for fledglings to survive, rather than a linearrelationship between survival and weight. The deadrecovery data in the present study was limited, butcomparing average weights and laying dates of youngseen again at the colony with those never seen again didnot reveal any relationship with fledgling weight, andthe proportion of birds seen again of the lightest birdsat fledging was as high as the proportion of the heavierbirds at fledging (Johannesen unpubl. data). We there-fore conclude that the different effect of fledglingweight on first year survival is not due to differentmethods applied but to differences among the studypopulations.
Our population differs in two ways from the Aus-tralian population. First, the Oamaru colony was char-acterised by a population increase (Table 1) during theyears of study. Second, the conditions were probablybeneficial around the colony. Just one chick was foundstarved, so most likely, the food was plentiful aroundthe colony throughout the breeding season. Due to theprovision of nest boxes, there was an excess of highquality breeding sites. This can also be seen from thedemographic performance of the birds. There were ahigh proportion of double breeders, high juvenile sur-vival (this study, Sidhu et al unpubl. data), and a highaverage number of fledglings produced per pair (onaverage 1.92 fledged young par pair per breeding sea-son; in fact the highest recorded for the species, seeFortescue 1999). Also, most young bred at the Quarrysite when they were two to three years old (Table 3)compared to three to four years old at Phillip Island,Australia (Reilly and Cullen 1981, see also Oro andPradel 2000 for a similar result on an increasing popu-lation of Audouins gull). The high survival of young,low age at first breeding and high reproductive perfor-mance found in this study has lead to an increase inpopulation size that can be explained by in situ recruit-ment alone. However, not only has high breeding per-formance lead to a population increase, but thepopulation increase could have selected for the highreproductive output, because fitness is increased byinvesting in present over future reproduction in increas-ing populations (Stearns 1992). We could not detectany short-term effects of the high demographic perfor-mance in this colony, since only positive correlationswere found between fitness parameters. More years ofstudy are needed before we can test for any long-termeffects of the high investment in reproduction forbreeding little penguins in the Quarry colony from1993–1997. However, in the later years first-year sur-vival has declined (Fig. 5). Similarly, also, the probabil-
ity of double breeding has declined, with no doublebreeding in the breeding seasons 1998–2001 (Houstonand Russell unpubl. data). This could indicate a long-term trade-off, or alternatively be due to density depen-dent depletion of resources around the colony as itincreased in size.
Conclusion
We found evidence for intrinsic differences amongbreeding birds and no apparent trade-off betweenfitness parameters in this increasing population of littlepenguins. As density increased, overall demographicperformance (i.e. the proportion of breeders laying twoclutches and first year survival) declined. As morecohort move through the population, further light canbe shed on the relationship between density, individualquality differences, age specific demography and indi-vidual trade-offs in fitness parameters.
Acknowledgements – Oamaru Blue Penguin Colony monitor-ing team are greatly acknowledged for their weekly checks onthe penguins. T. Boulinier, M. Frederiksen, R. Julliard, K.McKoy, A.C. Prevot-Julliard and L. Sidhu are thanked fordiscussions and comments on the paper. Stephen Davies isthanked for correction of the English. E.J. was funded by theNorwegian Research Council and the French Ministry ofResearch.
ReferencesBarker, R. J. 1997. Joint modeling of live–recapture, tag–re-
sight and tag–recovery data. – Biometrics 53: 666–677.Brownie, C., Hines, J. E., Nichols, J. D., Pollock, K. H. and
Hestbeck., J. B. 1993. Capture-recapture studies for multi-ple strata including non-Markovian transitions. – Biomet-rics 49: 1173–1187.
Bull, L. 2000. Factors influencing little penguin Eudyptulaminor egg success on Matiu-Somes Island, New Zealand. –Emu 100: 199–204.
Burnham, K. P. 1993. A theory for combined analysis of ringrecovery and recapture data. – In: Lebreton, J.-D. andNorth, P. M. (eds). Marked individuals in the study of birdpopulations. Birkhauser Verlag, Basel, pp. 199–214.
Burnham, K. P. and Anderson, D. R. 1998. Model selectionand inference: a practical information theoretic approach.– Springer Verlag, New York.
Cam, E. and Monnat, J.-Y. 2000a. Apparent inferiority offirst-time breeders in the kittiwake: the role of heterogene-ity among age classes. – J. Anim. Ecol. 69: 380–394.
Cam, E. and Monnat, J.-Y. 2000b. Stratification based onreproductive state reveals contrasting patterns of age-re-lated variation in demographic parameters in the kittiwake.– Oikos 90: 560–574.
Cam, E., Hines, J. E., Monnat, J.-Y., Nichols, J. D. andDanchin, E. 1998. Are adult nonbreeders prudent parents?The kittiwake model. – J. Anim. Ecol. 79: 2917–2930.
Cam, E., Link, W. A., Cooch, E. G., Monnat, J.-Y. andDanchin, E. 2002. Individual covariation in life-historytraits: seeing the trees despite the forest. – Am. Nat. 159:96–105.
Catchpole, E. O., Freeman, S. N. and Morgan, B. J. T. 1993.On boundary estimation in ring recovery models and theeffect of adding recapture information. – In: Lebreton,
208 JOURNAL OF AVIAN BIOLOGY 34:2 (2003)

J.-D. and North, P. M. (eds). Marked individuals in thestudy of bird populations. Birkhauser Verlag, Basel, pp.215–229.
Chiaradia, A. F. and Kerry, K.R. 1999. Daily nest attendanceand breeding performance in the little penguin Eudyptulaminor at Phillip Island, Australia. – Marine Ornithol. 27:13–20.
Christians, J. K., Evanson, M. and Aiken, J. J. 2001. Seasonaldecline in clutch size in European starlings: a novel ran-domization test to distinguish between the timing andquality hypotheses. – J. Anim. Ecol. 70: 1080–1087.
Curio, E. 1983. Why do young birds reproduce less well? –Ibis 125: 400–404.
Dann, P. 1992. Distribution, population trends and factorsinfluencing the population size of little penguins Eudyptulaminor on Phillip Island, Victoria. – Emu 91: 263–272.
Dann, P. 1994. The abundance, breeding distribution and nestsites of blue penguins in Otago, New Zealand. – Notornis41: 157–166.
Dann, P. and Cullen, J. M. 1990. Survival, patterns of repro-duction, and lifetime reproductive output in little bluepenguins Eudyptula minor on Phillip Island, Victoria, Aus-tralia. – In: Davies, L. S. and Darby, J. T. (eds). Penguinbiology. Academic Press, San Diego, pp. 63–84.
Dann, P., Norman, F. I., Cullen, J. M., Neira, F. J. andChiaradia, A. 2000. Mortality and breeding failure of littlepenguins, Eudyptula minor, in Victoria, 1995–96, followinga widespread mortality of pilchard, Sardinops sagax. –Marine Freshw. Res. 51: 355–362.
Forslund, P. and Part, T. 1995. Age and reproduction in birds– hypotheses and tests. – Trends Ecol. Evol. 10: 374–378.
Fortescue, M. 1999. Temporal variation in breeding success ofthe little penguin Eudyptula minor on the east coast ofAustralia. – Marine Ornithol. 27: 21–28.
Frederiksen, M. and Bregnballe, T. 2000. Diagnosing a declinein return rate of 1-year-old cormorants: mortality, emigra-tion or delayed return? – J. Anim. Ecol. 69: 753–761.
Gales, R. 1985. Breeding seasons and double brooding of thelittle penguin Eudyptula minor in New Zealand. – Emu 85:127–130.
Gales, R. 1987. Growth strategies in blue penguins Eudyptulaminor. – Emu 87: 212–219.
Gales, R. and Green, B. 1990. The annual energetics cycle oflittle penguin Eudyptula minor. – Ecology 71: 2297–2312.
Hansson, B., Bensch, S. and Hasselquist, D. 2000. The qualityand timing hypothesis evaluated using data on great reedwarblers. – Oikos 90: 575–581.
Harrigan, K. E. 1992. Causes of mortality of little penguinsEudyptula minor in Victoria. – Emu 91: 273–277.
Harris, M. P., Buckland, S. T., Russel, S. M. and Wanless, S.1994. Post fledging survival to breeding age of shags Pha-lacrocorax aristolelis in relation to year, date of fledgingand brood size. – J. Avian Biol. 25: 268–274.
Hines, J. E. 1994. MSSURVIV – Users Manual. – NationalBiological Service, Patuxent Wildlife Research Center,Laurel, USA.
Hipfner, J. M., Gaston, A. J., Martin, D. L. and Jones, I. L.1999. Seasonal declines in replacement egg-layings in along-lived, Arctic seabird: cost of late breeding or variationin female quality? – J. Anim. Ecol. 68: 988–998.
Hochachka, W. 1990. Seasonal decline in reproductive perfor-mance of song sparrow. – Ecology 71: 1279–1288.
Hocken, A. G. and Russel, J. J. 2002. A method for determin-ing of sex by bill measurements in blue penguins Eudyptulaminor. – New Zealand J. Zool. 29: 63–69.
Johannesen, E., Steen, H. and Perriman, L. 2002. Seasonalvariation in survival, weights and population counts ofblue penguins (Eudyptula minor) in Otago, New Zealand. –New Zealand J. Zool. 29: 213–219.
Klett, A. T. and Johnson, D.H. 1982. Variability in nestsurvival rates and implications to nesting studies. – Auk
99: 77–87.Kremetz, D. G., Nichols, J. D. and Hines, J. E. 1989. Post-
fledging survival of European starlings. – Ecology 70:646–655.
Lepage, D., Desrochers, A. and Gauthier, G. 1999. Seasonaldecline in growth and fledging success in snow geese Ansercaerulescens : an effect of date or parent quality? – J. AvianBiol. 30: 72–78.
Littell, R. C., Milliken, G. A., Stroup, W. W. and Wolfinger,R. D. 1996. SAS Systems for Mixed Models. – SASInstitute, SAS Campus drive, Cary, North Caroline, USA.
Magrath, R. D. 1991. Nestling weight and juvenile survival inthe blackbird, Turdus merula. – J. Anim. Ecol. 60: 335–351.
Nilsson, J.-A� . 2000. Time-dependent reproductive decisions inthe blue tit. – Oikos 88: 351–361.
Norman, F. I., Du Guesclin, P. B. and Dann, P. 1992. The1986 wreck of little penguins Eudyptula minor in westernVictoria. – Emu 91: 369–376.
Olsson, O. 1997. Effects of food availability on fledging condi-tion and post-fledging survival in king penguin chicks. –Polar Biol. 18: 161–165.
Oro, D. and Pradel, R. 2000. Determinants of local recruit-ment in a growing colony of Audouin’s gull. – J. Anim.Ecol. 69: 119–132.
Perriman, L. and Steen, H. 2000. Blue penguins Eudyptulaminor nest distribution and breeding success on the Otagopeninsula, 1992 to 1998. – New Zealand J. Zool. 27:269–275.
Perriman, L., Houston, D., Steen, H. and Johannesen, E.2000. Climate fluctuation effects on breeding of blue pen-guins Eudyptula minor. – New Zealand J. Zool. 27: 261–267.
Prevot-Julliard, A.-C., Pradel, R., Julliard, R., Grosbois, V.and Lebreton, J.-D. 2001. Hatching date influences age atfirst reproduction in the black-headed gull. – Oecologia127: 62–68.
Reilly, P. N. and Cullen, J. M. 1979. The little penguinEudyptula minor in Victoria, I: Mortality of adults. – Emu79: 97–102.
Reilly, P. N. and Cullen, J. M. 1981. The little penguinEudyptula minor in Victoria, II: Breeding. – Emu 81: 1–19.
Reilly, P. N. and Cullen, J. M. 1982. The little penguinEudyptula minor in Victoria, III: Dispersal of chicks andsurvival after banding. – Emu 82: 137–142.
Renner, M. and Davies, L. 1999. Sexing little penguins Eudyp-tula minor from Cook Strait, New Zealand using discrimi-nant function analysis. – Emu 99: 74–79.
Ringsby, T. H., Sæther, B. E. and Solberg, E. J. 1998. Factorsaffecting juvenile survival in house sparrow Passer domesti-cus. – J. Avian Biol. 29: 241–247.
Rogers, T., Eldershaw, G. and Walraven, E. 1995. Reproduc-tive success of little penguins, Eudyptula minor, on LionIsland, New South Wales. – Wildlife Res. 22: 709–715.
Sagar, P. M. and Horning, D.S. Jr. 1997. Mass-related sur-vival of fledging sooty shearwaters Puffinus griseus at TheSnares, New Zealand. – Ibis 140: 329–331.
Sandercock, B. K., Lank, D. B. and Cooke, F. 1999. Seasonaldecline in the fecundity of arctic-breeding sandpipers: dif-ferent tactics in two species with an invariant clutch size. –J. Avian Biol. 30: 460–468.
Stahel, J. R. and Gales, R. 1987. Little penguin: Fairy pen-guins in Australia. – New South Wales Univ. Press,Sydney.
Stearns, S. C. 1992. The evolution of life histories. – OxfordUniversity Press, Oxford, UK.
Sydeman, W. J. and Eddy, J. O. 1995. Repeatability in layingdate and its relationship to individual quality for commonmurres. – Condor 97: 1048–1052.
JOURNAL OF AVIAN BIOLOGY 34:2 (2003) 209

van der Jeugd, H. P. and Larsson, K. 1998. Pre-breedingsurvival of barnacle geese Branta leucopsis in relation tofledging characteristics. – J. Anim. Ecol. 67: 953–966.
Viallefont, A., Cooch, E. G. and Cooke, F. 1995. Estimationof trade-offs with capture-recapture models: a case studyon the lesser snow geese. – J. Appl. Stat. 22: 847–862.
White, G. C. and Burnham, K. P. 1999. Program MARK:survival estimation from populations of marked animals. –Bird Study 46: 120–138.
Wiktander, U., Olsson, O. and Nilsson, S. G. 2001. Annualand seasonal reproductive trends in the lesser spottedwoodpecker Dendrocopos minor. – Ibis 143: 72–82.
Williams, T. D. 1995. The penguins, Spheniscidae. – OxfordUniversity Press, Oxford, Oxford, UK.
(Recei�ed 15 February 2002, re�ised 24 October 2002, accepted31 October 2002.)
210 JOURNAL OF AVIAN BIOLOGY 34:2 (2003)
Copyright © 2022 FDOKUMEN




















