
anti-ulcerative effect of aqueous extract of cucumis - baze ...

Increased numbers of immature plasma cells in peripheral bloodspecifically overexpress chemokine receptor CXCR3 and CXCR4 inpatients with ulcerative colitiscei_4290 215..224
S. Hosomi,* N. Oshitani,*N. Kamata,* M. Sogawa,*
H. Okazaki,* T. Tanigawa,*
H. Yamagami,* K. Watanabe,*
K. Tominaga,* T. Watanabe,*
Y. Fujiwara,* K. Maeda,† K. Hirakawa†
and T. Arakawa*Departments of *Gastroenterology, and †Surgical
Oncology, Osaka City University Graduate School
of Medicine, Osaka, Japan
Summary
Ulcerative colitis (UC) is a chronic inflammatory bowel disease featuringinfiltration by plasma cells producing immunoglobulins. We have reportedpreviously the specific and significant proliferation of immature plasma cellsin the inflamed colonic and pouch mucosa of UC patients. The aim of thisstudy was to characterize peripheral blood immature plasma cells and themigration mechanisms of such immature plasma cells to inflamed sites in UC.The characteristics of peripheral blood immature plasma cells and chemokinereceptor expression were examined by flow cytometry. Expression of mucosalchemokine was quantified using real-time reverse transcription–polymerasechain reaction and immunohistochemistry. The number of peripheral bloodimmature plasma cells was significantly higher in patients with active UC andactive Crohn’s disease (CD) than in healthy controls. The proportion ofimmature plasma cells was correlated positively with clinical activities of UCand CD. Many peripheral blood immature plasma cells were positive forCXCR3, CXCR4, CCR9 and CCR10. Expression of CXCR3 and CXCR4 in UCpatients was significantly higher than in controls. CXCL9, CXCL10 andCXCL11 mRNA levels in colonic mucosa of inflamed IBD were higher than incontrols. Immunofluorescence study also showed abundant CXCR3-positiveimmature plasma cells in the inflamed colonic mucosa of UC. Increasednumbers of immature plasma cells may migrate towards inflammatory sites ofUC via the CXCR3 axis, and may participate in UC pathogenesis.
Keywords: chemokine, chemokine receptor, inflammatory bowel disease,
plasma cell, ulcerative colitis
Accepted for publication 11 October 2010
Correspondence: S. Hosomi, Department of
Gastroenterology, Osaka City University
Graduate School of Medicine, 1-4-3,
Asahi-machi, Abeno-ku, Osaka 545-8585, Japan.
E-mail: [email protected]
Introduction
Ulcerative colitis (UC) and Crohn’s disease (CD) arechronic, idiopathic and relapsing forms of inflammatorybowel disease (IBD) of unknown aetiology. A dysregulatedinflammatory immune response has been suggested in thepathogenesis of IBD. Basal plasmacytosis, a dense infiltrationof plasma cells in the lower one-third of the mucosa, is aclassical pathological finding in UC [1]. Activation ofhumoral immunity based on T helper type 2 (Th2) cell-shifted immunity has been implicated in UC pathogenesis,whereas Th1-shifted activation of cellular immunity has animportant role in CD pathogenesis [2]. In UC, laminapropria lymphocytes contain activated B cell subsets [3], andlocal immunoglobulin (Ig) G overproduction shifts to thehighly complement-activating IgG1 subclass [4]. Hibi et al.[5,6] reported that lymphocytes isolated from the inflamed
mucosa and peripheral blood of patients with UC secretedautoantibodies against colonic epithelial cells. Generation ofautoantibodies to tropomyosin-5 [7,8], expressed by theintestinal epithelium, and perinuclear anti-neutrophil anti-bodies (p-ANCA) [9,10] are also a characteristic humoralimmune response in patients with UC. However, littleis known about the mechanisms and processes of B cellactivation in the pathogenesis of inflammatory boweldisease.
We have reported the specific and significant proliferationof immature plasma cells in the inflamed colonic [11] andpouch [12] mucosa of patients with UC compared withhealthy controls and patients with CD. Although theseimmature plasma cells may have important roles in abnor-mal humoral immune responses, including autoantibodyproduction, the mechanisms of migration of immatureplasma cells to inflamed sites of UC have not been clarified.
Clinical and Experimental Immunology ORIGINAL ARTICLE doi:10.1111/j.1365-2249.2010.04290.x
215© 2010 The AuthorsClinical and Experimental Immunology © 2010 British Society for Immunology, Clinical and Experimental Immunology, 163: 215–224

Chemoattractant cytokines (chemokines), together withtissue-specific adhesion molecules, have a central role in themigration of antibody-secreting cells to target tissue [13]. Inparticular, the chemokines/chemokine receptors axis is themost important regulator of leucocyte trafficking in infec-tion or inflammation, by switching the expression of thechemokine receptor profile on these leucocytes during acti-vation and differentiation [14,15]. The association betweeninappropriate activation of the chemokine network andseveral diseases has been investigated by many researchers[16]. In IBD, high expression of chemokine receptors on Tcell lineages has been shown [17,18], however, little has beenreported on the expression of chemokine receptors on B cellpopulations (including plasma cells).
The aim of this study was to characterize peripheral bloodimmature plasma cells and the migration mechanisms ofsuch immature plasma cells to inflamed sites in IBD.
Materials and methods
This study was approved by the Ethics Committee of OsakaCity University. Informed consent was obtained from allpatients and healthy controls.
Patients and samples
The diagnosis of IBD was based on clinical, endoscopic andhistopathological findings. The disease activity of UCpatients was determined according to the Mayo Clinic score[19], whereas the disease activity of CD patients was deter-mined according to the Crohn’s Disease Activity Index(CDAI) [20]. Clinical remission was defined as a total MayoClinic score of 2 or fewer points in patients with UC, and atotal CDAI <150 points in patients with CD.
Peripheral blood samples were obtained from 18 patientswith UC of median age 39 years (25th–75th percentiles32–53) and Mayo Clinic score 5 (2–7), 16 patients with CDaged 31 years (25–36·5) and CDAI 183·4 (129·2, 249·7) and13 healthy controls aged 33 years (31–37). The location ofCD involvement was ileal in two patients, ileocolonic in 13patients and colonic in one patient. UC extent was pancolitisin 12 patients and left-sided colitis in six patients. Forchemokine receptor expression assays, data were collectedfrom 11 patients with UC, 13 patients with CD and 10healthy controls.
Mucosal biopsy samples were obtained from inflamed andnon-inflamed areas of the rectosigmoid colon from 35patients with UC (18 inflamed and 17 non-inflamed speci-mens) and 28 patients with CD (13 inflamed and 15 non-inflamed specimens) during colonoscopy. The location ofCD involvement was ileocolonic in 10 patients and colonic in18 patients. UC extent was pancolitis in 19 patients, left-sided colitis in 12 patients and proctitis in four patients. Forcomparison, control biopsy specimens were taken from thenormal rectosigmoid mucosa of 17 patients who underwent
colonoscopy for various reasons (e.g. cancer screening).Endoscopic activity was determined using an endoscopicscore of 0–2 (0, normal; 1, light erythema or granularity; 2,granularity, friability and bleeding, with or without ulcer-ations) according to Froslie’s criteria [21], with a score of 0or 1 regarded as non-inflamed.
Surgical samples obtained from 16 patients with UC(eight inflamed and eight non-inflamed specimens) and 15patients with CD (eight inflamed and seven non-inflamedspecimens). Macroscopically normal mucosa was obtainedfrom seven patients who underwent surgery for colorectalcancer and was used as control samples.
Flow cytometric analysis
Human peripheral blood mononuclear cells (PBMCs) wereisolated by Ficoll-Paque Plus (Amersham Biosciences AB,Uppsala, Sweden) density centrifugation and washed inphosphate-buffered saline (PBS). To analyse the frequency ofimmature plasma cells and expression of the chemokinereceptor, PBMCs were stained with fluorescein isothiocyan-ate (FITC)-conjugated mouse anti-human-CD20 (DakoCy-tomation, Glostrup, Denmark), allophycocyanin (APC)-conjugated mouse anti-human-CD19 (IOTest, Immunotech,Marseille, France), phycoerythrin (PE)-conjugated mouseanti-human-CD138 (DakoCytomation), PE-conjugatedmouse anti-human-CD38 (DakoCytomation), PE-conjugated mouse anti-human-integrin a5/CD49e (R&DSystems, Incorporated, Minneapolis, MN, USA), PE-conjugated mouse anti-human-CXCR3 (R&D Systems,Incorporated), PE-conjugated mouse anti-human-CXCR4(R&D Systems), PE-conjugated mouse anti-human-CXCR5(R&D Systems), PE-conjugated mouse anti-human-CXCR6(R&D Systems), PE-conjugated mouse anti-human-CCR9(R&D Systems) and PE-conjugated rat anti-human-CCR10(R&D Systems) in the dark at room temperature for 30 min.Isotype- and species- matched Ig was the negative control.
After washing in PBS, cells were analysed on a LSR II flowcytometer (BD Biosciences, San Jose CA, USA) using FACS-Diva software.
In-vitro culture of CD19+ B cells
PBMCs isolated from healthy volunteers (as described above)were washed twice in PBS, and CD19+ B cells purified viapositive selection on magnetic affinity cell sorting (MACS)columns (MACS CD19 MicroBeads human; Miltenyi Biotec,Auburn, CA, USA), using specific mouse anti-human CD19specific microbeads. The purity of CD19+ B cells was tested byflow cytometry and found to be >95%. For the analysis of cellproliferation, CD19+ B cells were stained with 5(6)-carboxyfluorescein diacetate-N-hydroxysuccinimide ester(CFSE) (Dojin, Kumamoto, Japan). CD19+ B cells werewashed twice in PBS and cells resuspended in PBS (1 ¥ 106
cells/ml) containing CFSE at a final concentration of 2 mM.
S. Hosomi et al.
216 © 2010 The AuthorsClinical and Experimental Immunology © 2010 British Society for Immunology, Clinical and Experimental Immunology, 163: 215–224

Cells were incubated at 37°C for 10 min. After washing inPBS, CFSE-treated cells were cultured in AIM-V (Gibco BRLInvitrogen, Grand Island, NY, USA) with recombinantinterleukin-6 (IL-6) (20 ng/ml) (R&D Systems) for 7 days at37°C in 5% CO2. Cultured cells were stained with APC-conjugated mouse anti-human-CD19 (IOTest, Immunotech)and PE-conjugated mouse anti-human-CD138 (DakoCyto-mation) in the dark at room temperature for 30 min, andanalysed by flow cytometry. For studies of live cells,7-aminoactinomycin D (7-AAD) (Immunotech) was addedbefore flow cytometric analysis.
Intracellular staining for immunoglobulin
PBMCs isolated as described above were incubated withPE-conjugated mouse anti-human-CXCR3 (R&D Systems),PE-conjugated mouse anti-human-CXCR4 (R&D Systems),APC-conjugated mouse anti-human-CD19 (IOTest, Immu-notech) and PE/Cy7-conjugated mouse anti-human-CD20(BioLegend, San Diego, CA, USA). After washing twice withPBS, cells were fixed and permeabilized using BD Cytofix/Cytoperm Fixation/Permeabilization Kit (BD Biosciences),according to the manufacturer’s instructions. Cells werestained subsequently with FITC-conjugated goat anti-human-IgM (KPL), FITC-conjugated anti-human-IgG(H + l) (KPL) and FITC-conjugated anti-human-IgA (a)(KPL, Gaithersburg, MD, USA) in the dark at room tempera-ture for 30 min. Isotype- and species-matched Ig was thenegative control. Cells were analysed by flow cytometry.
Quantification of levels of chemokine mRNA in colonictissues by real-time reverse transcription–polymerasechain reaction (RT–PCR)
Total RNA was extracted and purified from rectosigmoidcolon biopsy samples using the RNAqueous-Micro kit(Ambion Incorporated, Austin, Texas, USA), according tothe manufacturer’s instructions. The probes (Applied Bio-systems Incorporated, Foster City, CA, USA) used for analy-sis were as follows: CXCL9 (Hs00171065_m1), CXCL10(Hs00171042_m1), CXCL11 (Hs00171138_m1), CXCL12(Hs00930455_m1), CCL27 (Hs00171157_m1) and CCL28(Hs00219797_m). RT–PCR analysis was performed using anABI prism 7700 sequence detection system instrument andsoftware (Applied Biosystems). The reaction mixture wasprepared according to the manufacturer’s instructions usingthe SuperScript III Platinum One-Step quantitative RT–PCR(qRT–PCR) Kit (Invitrogen). Thermal cycling conditionswere 50°C for 15 min and 95°C for 2 min, followed by 40cycles of amplification at 95°C for 15 s and 60°C for 30 s.Total RNA was subjected to real-time RT–PCR for measure-ment of target genes, and glyceraldehyde-3-phosphate dehy-drogenase (GAPDH) was used as the internal standard withTaqMan GAPDH control reagents (Applied Biosystems).Each resulting gene amount was divided by GAPDH gene
amount to obtain a normalized value. The ratio of mRNAlevels was normalized to the mean value for control samples.
Immunohistochemistry
Surgical samples were obtained from patients with CD andUC for immunohistological study. A segment (approxi-mately 5 ¥ 5 mm) was excised from surgically resected colonand immediately fixed in periodate/lysine/2% paraformalde-hyde at 4°C for 6 h, and then incubated in a 10%–15%–20%sucrose gradient in PBS at 4°C for 6 h each. Specimens wereembedded in octreotide (OCT) compound (SakuraFinetechnical Company, Tokyo, Japan) in dry ice/acetone,and stored at –80°C until use.
Cryostat sections (thickness, 4 mm) were cut and pre-treated with 5% normal donkey serum with PBS to inhibitnon-specific protein binding. After washing in PBS, sectionswere first reacted with primary antibody at 4°C overnight.Rabbit anti-human-CXCR3 antibody (diluted 1:100 in PBS;GeneTex, Incorporated, San Antonio, TX, USA), mouse anti-human-CD3 antibody (diluted 1:200 in PBS; DakoCytoma-tion), mouse anti-human-CD56 antibody (diluted 1:200 inPBS; DakoCytomation), mouse anti-human-CD68 antibody(diluted 1:200 in PBS; DakoCytomation), mouse anti-human-CD138 antibody (diluted 1:200 in PBS; DakoCyto-mation), goat anti-human-CD19 antibody (diluted 1:50 inPBS; C-20; Santa Cruz Biotechnology, Incorporated, SantaCruz, CA, USA), rabbit anti-human-CD138 antibody(diluted 1:100 in PBS; Thermo Fisher Scientific AnatomicalPathology, CA, USA), mouse anti-human-CXCR3 antibody(diluted 1:50 in PBS; GeneTex, Incorporated), rabbit anti-human-CXCL9/MIG (diluted 1:200 in PBS; H-40; SantaCruz Biotechnology), rabbit anti-human-CXCL10/IP-10(diluted 1:200 in PBS; K-19; Santa Cruz Biotechnology) andrabbit anti-human-CXCL11/I-TAC (diluted 1:200 in PBS;FL-94; Santa Cruz Biotechnology) were the primaryantibodies. Isotype- and species-matched Ig was the negativecontrol. After washing three times in PBS containing 0·05%Tween-20 (PBS-T), sections were incubated for 1 h at roomtemperature with AlexaFluor 488 donkey anti-rabbit IgGantibody (diluted 1:200 in PBS-T; Molecular Probes/Invitrogen, Carlsbad, CA, USA), AlexaFluor 594 donkeyanti-mouse IgG antibody (diluted 1:200 in PBS-T; MolecularProbes/Invitrogen) and AlexaFluor 350 donkey anti-goatIgG antibody (diluted 1:50 in PBS-T; Molecular Probes/Invitrogen). They were then mounted in fluorescent mount-ing medium (DakoCytomation). Images were taken under afluorescence microscope (Olympus BX50, Olympus, Tokyo,Japan) equipped with a colour camera (Olympus ColorChilled 3CCD Camera, M-3204C). Results were obtainedfrom serial sections.
To quantify the numbers of lamina propria CD19+
CD138+ CXCR3+ triple-positive cells, four random high-power fields in each specimen were counted and meancounts were calculated.
Increased immature plasma cells overexpress CXCR3/4 in UC
217© 2010 The AuthorsClinical and Experimental Immunology © 2010 British Society for Immunology, Clinical and Experimental Immunology, 163: 215–224
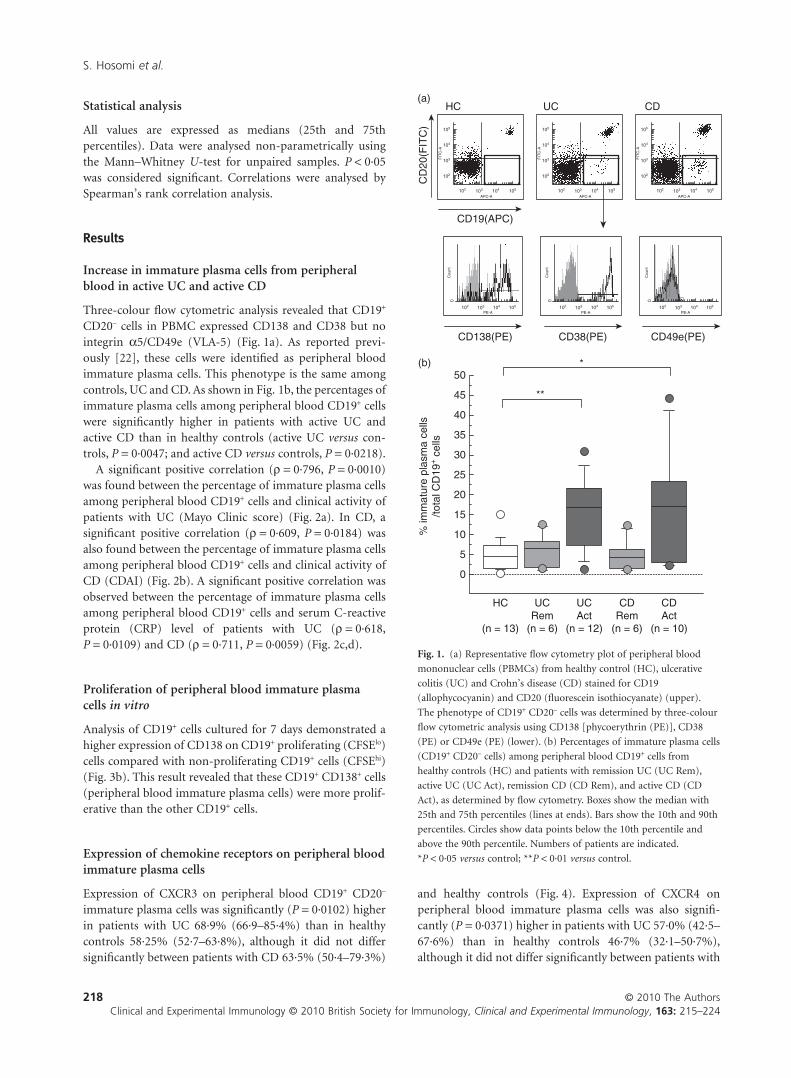
Statistical analysis
All values are expressed as medians (25th and 75thpercentiles). Data were analysed non-parametrically usingthe Mann–Whitney U-test for unpaired samples. P < 0·05was considered significant. Correlations were analysed bySpearman’s rank correlation analysis.
Results
Increase in immature plasma cells from peripheralblood in active UC and active CD
Three-colour flow cytometric analysis revealed that CD19+
CD20– cells in PBMC expressed CD138 and CD38 but nointegrin a5/CD49e (VLA-5) (Fig. 1a). As reported previ-ously [22], these cells were identified as peripheral bloodimmature plasma cells. This phenotype is the same amongcontrols, UC and CD. As shown in Fig. 1b, the percentages ofimmature plasma cells among peripheral blood CD19+ cellswere significantly higher in patients with active UC andactive CD than in healthy controls (active UC versus con-trols, P = 0·0047; and active CD versus controls, P = 0·0218).
A significant positive correlation (r = 0·796, P = 0·0010)was found between the percentage of immature plasma cellsamong peripheral blood CD19+ cells and clinical activity ofpatients with UC (Mayo Clinic score) (Fig. 2a). In CD, asignificant positive correlation (r = 0·609, P = 0·0184) wasalso found between the percentage of immature plasma cellsamong peripheral blood CD19+ cells and clinical activity ofCD (CDAI) (Fig. 2b). A significant positive correlation wasobserved between the percentage of immature plasma cellsamong peripheral blood CD19+ cells and serum C-reactiveprotein (CRP) level of patients with UC (r = 0·618,P = 0·0109) and CD (r = 0·711, P = 0·0059) (Fig. 2c,d).
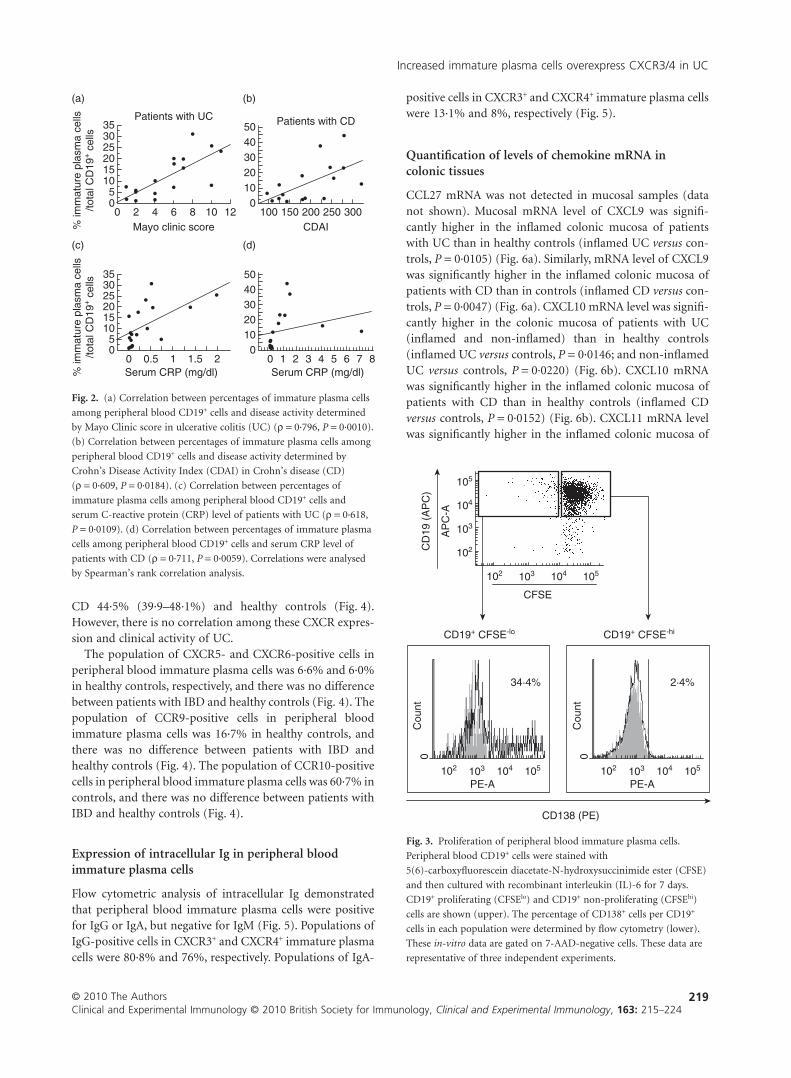
Proliferation of peripheral blood immature plasmacells in vitro
Analysis of CD19+ cells cultured for 7 days demonstrated ahigher expression of CD138 on CD19+ proliferating (CFSElo)cells compared with non-proliferating CD19+ cells (CFSEhi)(Fig. 3b). This result revealed that these CD19+ CD138+ cells(peripheral blood immature plasma cells) were more prolif-erative than the other CD19+ cells.
Expression of chemokine receptors on peripheral bloodimmature plasma cells
Expression of CXCR3 on peripheral blood CD19+ CD20–
immature plasma cells was significantly (P = 0·0102) higherin patients with UC 68·9% (66·9–85·4%) than in healthycontrols 58·25% (52·7–63·8%), although it did not differsignificantly between patients with CD 63·5% (50·4–79·3%)
and healthy controls (Fig. 4). Expression of CXCR4 onperipheral blood immature plasma cells was also signifi-cantly (P = 0·0371) higher in patients with UC 57·0% (42·5–67·6%) than in healthy controls 46·7% (32·1–50·7%),although it did not differ significantly between patients with
HC
CD
20(F
ITC
)
CD19(APC)
APC-A
PE-A
FIT
C-A
Cou
nt
(a)
(b)
UC CD
CD138(PE) CD38(PE) CD49e(PE)
APC-A
FIT
C-A
APC-A
FIT
C-A
PE-A
Cou
nt
PE-A
Cou
nt
102
102
103
103
104
104
105
105
102
0
103 104 105 102
0
103 104 105 102
0
103 104 105
102
102
103
103
104
104
105
105 102
102
103
103
104
104
105
105
0
5
10
HC
(n = 13)
UCRem
(n = 6)
CDRem
(n = 6)
UCAct
(n = 12)
CDAct
(n = 10)
15
20
25
30
35
40
45
50*
**
% im
mat
ure
plas
ma
cells
/tota
l CD
19+ c
ells
Fig. 1. (a) Representative flow cytometry plot of peripheral blood
mononuclear cells (PBMCs) from healthy control (HC), ulcerative
colitis (UC) and Crohn’s disease (CD) stained for CD19
(allophycocyanin) and CD20 (fluorescein isothiocyanate) (upper).
The phenotype of CD19+ CD20– cells was determined by three-colour
flow cytometric analysis using CD138 [phycoerythrin (PE)], CD38
(PE) or CD49e (PE) (lower). (b) Percentages of immature plasma cells
(CD19+ CD20– cells) among peripheral blood CD19+ cells from
healthy controls (HC) and patients with remission UC (UC Rem),
active UC (UC Act), remission CD (CD Rem), and active CD (CD
Act), as determined by flow cytometry. Boxes show the median with
25th and 75th percentiles (lines at ends). Bars show the 10th and 90th
percentiles. Circles show data points below the 10th percentile and
above the 90th percentile. Numbers of patients are indicated.
*P < 0·05 versus control; **P < 0·01 versus control.
S. Hosomi et al.
218 © 2010 The AuthorsClinical and Experimental Immunology © 2010 British Society for Immunology, Clinical and Experimental Immunology, 163: 215–224
CD 44·5% (39·9–48·1%) and healthy controls (Fig. 4).However, there is no correlation among these CXCR expres-sion and clinical activity of UC.
The population of CXCR5- and CXCR6-positive cells inperipheral blood immature plasma cells was 6·6% and 6·0%in healthy controls, respectively, and there was no differencebetween patients with IBD and healthy controls (Fig. 4). Thepopulation of CCR9-positive cells in peripheral bloodimmature plasma cells was 16·7% in healthy controls, andthere was no difference between patients with IBD andhealthy controls (Fig. 4). The population of CCR10-positivecells in peripheral blood immature plasma cells was 60·7% incontrols, and there was no difference between patients withIBD and healthy controls (Fig. 4).
Expression of intracellular Ig in peripheral bloodimmature plasma cells
Flow cytometric analysis of intracellular Ig demonstratedthat peripheral blood immature plasma cells were positivefor IgG or IgA, but negative for IgM (Fig. 5). Populations ofIgG-positive cells in CXCR3+ and CXCR4+ immature plasmacells were 80·8% and 76%, respectively. Populations of IgA-
positive cells in CXCR3+ and CXCR4+ immature plasma cellswere 13·1% and 8%, respectively (Fig. 5).
Quantification of levels of chemokine mRNA incolonic tissues
CCL27 mRNA was not detected in mucosal samples (datanot shown). Mucosal mRNA level of CXCL9 was signifi-cantly higher in the inflamed colonic mucosa of patientswith UC than in healthy controls (inflamed UC versus con-trols, P = 0·0105) (Fig. 6a). Similarly, mRNA level of CXCL9was significantly higher in the inflamed colonic mucosa ofpatients with CD than in controls (inflamed CD versus con-trols, P = 0·0047) (Fig. 6a). CXCL10 mRNA level was signifi-cantly higher in the colonic mucosa of patients with UC(inflamed and non-inflamed) than in healthy controls(inflamed UC versus controls, P = 0·0146; and non-inflamedUC versus controls, P = 0·0220) (Fig. 6b). CXCL10 mRNAwas significantly higher in the inflamed colonic mucosa ofpatients with CD than in healthy controls (inflamed CDversus controls, P = 0·0152) (Fig. 6b). CXCL11 mRNA levelwas significantly higher in the inflamed colonic mucosa of
00 2 4 6 8
Mayo clinic score
Patients with UC
% im
mat
ure
plas
ma
cells
/tota
l CD
19+ c
ells
10 12
252015105
3035
(a)
0100 150 200 250
CDAI
Patients with CD
300
302010
4050
(b)
00 0.5 1 1.5Serum CRP (mg/dl) Serum CRP (mg/dl)%
imm
atur
e pl
asm
a ce
lls/to
tal C
D19
+ c
ells
2
252015105
3035
(c)
0 1 2 3 4 5 6 7 80
302010
4050
(d)
Fig. 2. (a) Correlation between percentages of immature plasma cells
among peripheral blood CD19+ cells and disease activity determined
by Mayo Clinic score in ulcerative colitis (UC) (r = 0·796, P = 0·0010).
(b) Correlation between percentages of immature plasma cells among
peripheral blood CD19+ cells and disease activity determined by
Crohn’s Disease Activity Index (CDAI) in Crohn’s disease (CD)
(r = 0·609, P = 0·0184). (c) Correlation between percentages of
immature plasma cells among peripheral blood CD19+ cells and
serum C-reactive protein (CRP) level of patients with UC (r = 0·618,
P = 0·0109). (d) Correlation between percentages of immature plasma
cells among peripheral blood CD19+ cells and serum CRP level of
patients with CD (r = 0·711, P = 0·0059). Correlations were analysed
by Spearman’s rank correlation analysis.
CD
19 (
AP
C)
AP
C-A
CFSE
CD19+ CFSE-lo CD19+ CFSE-hi
102
102
103
103
104
104
105
105
CD138 (PE)
PE-A
Cou
nt
102
0
103 104 105
34·4%
PE-A
Cou
nt
102
0
103 104 105
2·4%
Fig. 3. Proliferation of peripheral blood immature plasma cells.
Peripheral blood CD19+ cells were stained with
5(6)-carboxyfluorescein diacetate-N-hydroxysuccinimide ester (CFSE)
and then cultured with recombinant interleukin (IL)-6 for 7 days.
CD19+ proliferating (CFSElo) and CD19+ non-proliferating (CFSEhi)
cells are shown (upper). The percentage of CD138+ cells per CD19+
cells in each population were determined by flow cytometry (lower).
These in-vitro data are gated on 7-AAD-negative cells. These data are
representative of three independent experiments.
Increased immature plasma cells overexpress CXCR3/4 in UC
219© 2010 The AuthorsClinical and Experimental Immunology © 2010 British Society for Immunology, Clinical and Experimental Immunology, 163: 215–224

patients with UC than in controls (inflamed UC versus con-trols, P = 0·0248) (Fig. 6c). CXCL11 mRNA was significantlyhigher in the inflamed colonic mucosa of patients with CDthan in controls (inflamed CD versus controls, P = 0·0113)(Fig. 6c). No difference in CXCL12 and CXCL28 mRNAexpression between patients with IBD and controls wasfound (Fig. 6d,e).
Localization of CX chemokine receptor 3-positive cellsand CX chemokine ligands in the colonic mucosa
To identify the type and distribution of immune cells thatexpress the chemokine receptor CXCR3, we conductedimmunofluorescence studies on surgically resected colonictissue of controls and IBD patients. Double- or triple-immunofluorescence experiments on colonic mucosarevealed that CD3+ T cells, CD56+ natural killer (NK) cells,CD19+ cells and CD138+ cells all expressed CXCR3, whileCD68+ macrophages did not. In the ulcer base of UC, mostCXCR3 expression was found on immature plasma cellsdouble-positive for CD19 and CD138 (Fig. 7). In accordwith our previous data [11], numerous CD19+ CD138+
immature plasma cells were observed in the inflamed UCmucosa (Fig. 8a–e). A triple-immunofluorescence studyrevealed that the number of immature plasma cells express-ing CXCR3 was significantly higher in the inflamed colonicmucosa of UC patients than in the colonic mucosa of con-trols (inflamed UC versus controls, P = 0·0026) (Fig. 8c,d,f).
Immunofluorescence study also showed increased expres-sion of CXCL9, 10 and 11 in the inflamed colonic mucosa of
UC and CD patients compared with control colonic mucosa.As reported elsewhere [23] [24], CXCL9, 10 and 11 expres-sion was detected on neutrophils and lamina propria mono-nuclear cells (Fig. 9a–d).
Discussion
Memory B cells activated by antigen in secondary lymphoidtissues leave the lymphoid tissues and migrate through thevessels to target tissues [13]. Activated B cells differentiateinto plasma blasts that express CD19+ CD20– CD138+ inte-grin a5/CD49e– phenotype, and then differentiate intoCD138+ cells within a few days [25]. These CD19+ CD20–
CD138+ CD49e– cells in the peripheral blood are defined asearly (immature) plasma cells because they lack the CD49eexpression characteristic of bone marrow plasma cells[22,26]. These immature plasma cells represent 0·1–0·2% ofthe total PBMCs in healthy donors, but these peripheralblood immature plasma cells increase in the patients with
PE-A
CXCR3
CXCR3 - PE
Cou
nt
1020
5
103
10
15
20
25
104 105
PE-A
CXCR4
CXCR4 - PE
Cou
nt1020
5
103
10
15
20
25
104 105
PE-A
CXCR5
CXCR5 - PE
Cou
nt
1020
5
103
10
15
20
25
104 105
PE-A
CXCR6
CXCR6 - PE
Cou
nt
1020
5
103
10
15
20
25
104 105
PE-A
CCR9
CCR9 - PE
Cou
nt
1020
5
103
10
15
20
25
104 105
PE-A
CCR10
CCR10 - PE
Cou
nt
1020
5
103
10
15
20
25
104 105
HC (n =
10)
UC (n =
11)
CD (n =
13) HCUC CD HC UC CD HC UC CD HC UC CD HCUC CD
0102030405060708090
% C
hem
okin
e re
cept
or c
ells
/Imm
atur
e pl
asm
a ce
lls
* *
Fig. 4. Percentage of chemokine receptor expression (CXCR3,
CXCR4, CXCR5, CXCR6, CCR9 and CCR10) on peripheral blood
immature plasma cells from healthy controls (HC) and patients with
ulcerative colitis (UC) and Crohn’s disease (CD), as determined by
three-colour flow cytometry. Analysis was gated on CD19+ CD20–
cells. Numbers of patients are indicated. *P < 0·05 versus control.
CD
20(P
E-C
y7)
CD19(APC) CXCR3 - PE
CXCR3+
lgG - FITC
CXCR4+
APC-A
PE
-Cy7
-A
102
102
0 0
103
103
104
104
105
105
PE-A
Cou
nt
102 103 104 105
0
FITC-A
60·4%lgG
Cou
nt
102 103 104 1050
FITC-A
80·8%CXCR3-lgG
Cou
nt
102 103 104 1050
FITC-A
76%CXCR4-lgG
Cou
nt
102 103 104 105
lgM - FITC
0%
0
FITC-A
lgM
Cou
nt
102 103 104 105
CXCR4 - PE
PE-A102 103 104 105
lgG - FITC
0
FITC-A
38·9%lgG
Cou
nt
102 103 104 1050
FITC-A
13·1%CXCR3-lgA
Cou
nt
102 103 104 1050
FITC-A
8%CXCR4-lgA
Cou
nt
102 103 104 105
Fig. 5. Immunoglobulin (Ig) type of peripheral blood immature
plasma cells. Intracellular staining for Ig (IgM, IgG and IgA) was
performed in CD19+ CD20– cells by three-colour flow cytometry.
Intracellular staining for Ig (IgG and IgA) was performed in CD19+
CD20– CXCR3+ cells and CD19+ CD20– CXCR4+ cells by four-colour
flow cytometry. These data are representative of three independent
experiments.
S. Hosomi et al.
220 © 2010 The AuthorsClinical and Experimental Immunology © 2010 British Society for Immunology, Clinical and Experimental Immunology, 163: 215–224

active systemic lupus erythematosus (SLE), bacterial septi-caemia and liver cirrhosis [22]. In the present study, becausethree-colour flow cytometric analysis of PBMC revealed thatperipheral blood CD19+ CD20– cells expressed CD138 andCD38 but not CD49e, these peripheral blood CD19+ CD20–
cells could be identified as immature (early) plasma cells.The population of immature plasma cells among peripheralblood CD19+ cells in patients with active UC and active CDwere significantly higher than healthy controls. This is thefirst study to describe increased peripheral blood immatureplasma cells in active UC and CD. There was a significantpositive correlation between the percentage of immatureplasma cells among peripheral blood CD19+ cells and theclinical activity of UC and CD. Alteration in peripheralblood immature plasma cells is therefore considered to be agood parameter to predict clinical activity. The increasednumbers of immature plasma cells in active SLE and bacte-rial septicaemia decrease after treatment [22], therefore anincreased number of peripheral blood immature plasmacells may reflect mobilization of B cells into mature plasmacells.
Earlier, we showed that the immature plasma cells thatinfiltrated into the inflamed colonic mucosa, vermiformappendix and pouchitis mucosa of patients with UC exhib-ited high degrees of labelling for Ki-67 [11,12,27]. Thepresent study revealed that immature plasma cells in periph-eral blood proliferated in vitro in response to IL-6.
Despite an increase in peripheral blood immature plasmacells among patients with UC and CD, infiltration of imma-ture plasma cells in inflamed mucosa is specific for UC [11].These observations may indicate that the processes of cells‘homing’ to the inflamed mucosa differ in UC and CD. Toelucidate the mechanisms of peripheral blood immature
HC (n =
17)
UC –NI (
n =
17)
UC –I (
n =
18)
CD –NI (
n =
15)
CD –I (
n =
13)
0
10
40
(a) CXCL9
30
20
Rel
ativ
e m
RN
A e
xpre
ssio
n ***
HC
UC –NI
CD –NI
UC –I
CD –I
0
10
40
(b) CXCL10
30
20
***
HC
UC –NI
UC –I
CD –NI
CD –I
HC
UC –NI
CD –NI
UC –I
CD –I
HC
UC –NI
UC –I
CD –NI
CD –I
0
40
20
100
(c) CXCL11
80
60
***
0
4
16
CXCL12
12
8
01
32
6
(e)(d) CCL28
54
Fig. 6. Chemokine mRNA levels in colonic tissues of healthy controls
(HC) and patients with non-inflamed ulcerative colitis (UC)
(UC-NI), inflamed UC (UC-I), non-inflamed Crohn’s disease (CD)
(CD-NI) and inflamed CD (CD-I). Real-time reverse
transcription–polymerase chain reaction (RT–PCR) analysis was
performed, and mRNA levels were normalized to the mean value for
control samples. (a) CXCL9 mRNA level. (b) CXCL10 mRNA level.
(c) CXCL11 mRNA level. (d) CXCL12 mRNA level. (e) CCL28 mRNA
level. Numbers of patients are indicated. *P < 0·05 versus control;
**P < 0·01 versus control.
Fig. 7. Expression of CXCR3 (green) on the
lamina propria T cells (CD3, red), natural killer
(NK) cells (CD56, red), macrophages (CD68,
red) and immature plasma cells (CD19, blue;
CD138, red) in active ulcerative colitis (UC).
The specificity of each antibody was confirmed
by negative reactivity of isotype- and
species-matched immunoglobulin.
Increased immature plasma cells overexpress CXCR3/4 in UC
221© 2010 The AuthorsClinical and Experimental Immunology © 2010 British Society for Immunology, Clinical and Experimental Immunology, 163: 215–224

plasma cells homing via chemokine receptors, we analysedchemokine receptor expression on peripheral blood imma-ture plasma cells.
The high expression of CXCR3 on peripheral bloodmemory B cells in rheumatoid arthritis and on B cells incerebrospinal fluid in multiple sclerosis have been reported[28,29]. Several studies have shown that CXCR3 ligandchemokines (CXCL9/MIG, CXCL10/IP10 and CXCL11/ITAC) are up-regulated in inflammatory sites of UC and CD[30–33]. In the present study, flow cytometry experimentsrevealed that many peripheral blood immature plasma cellswere positive for CXCR3, and its expression on peripheralblood immature plasma cells was up-regulated selectively inpatients with UC. Real-time RT–PCR data showed increasedmRNA expression of CXCR3 ligand chemokines (CXCL9, 10and 11) in the inflamed colonic mucosa of patients with UCand CD compared to that in controls. These findings werecorroborated by immunohistochemistry studies that dem-onstrated the following results: (1) increased CXCL9, 10 and11 expression on the inflamed colonic mucosa of UC andCD patients, especially on inflamed UC mucosa, comparedto control mucosa; and (2) an abundance of CXCR3 express-ing CD19+ CD138+ immature plasma cells on the inflamedcolonic mucosa of UC patients compared to controls.However, in CD patients, CXCR3 expression on immatureplasma cells was not high. Specific to UC patients, theup-regulation of both CXCR3 and its ligands may triggerimmature plasma cell migration into inflamed colonicmucosa. These CXCR3-positive peripheral blood immatureplasma cells contained mainly IgG. Our previous study
showed that immature plasma cells in inflamed colonicmucosa of UC contained IgG [11]. This result explains thelocal IgG overproduction and generation of autoantibodiesin patients with UC.
CXCR4 expression on peripheral blood immature plasmacells was also up-regulated selectively in patients with UC.IgA and IgG antibody-secreting cells (ASCs) with CXCR4expression migrate to the bone marrow and mucosal sites inresponse to CXCL12/SDF1a [13]. However, mRNA expres-sion of the CXCR4 ligand CXCL12/SDF1a in colonicmucosa was not significantly different between IBD andcontrols. These findings indicate that CXCR4 axis may play aless important part in the migration of immature plasmacells to inflamed colonic mucosa in UC than the CXCR3 axis.
Most peripheral blood immature plasma cells were nega-tive for CXCR5 and CXCR6. CXCR5 expression on B cells isdecreased on plasma cells during plasmacytic differentiation[34]. Human bone marrow plasma cells express CXCR6, andthe CXCR6–CXCL16 axis contributes to migration andtissue localization of human plasma cells in bone marrow[35]. These findings indicate that peripheral blood immatureplasma cells are intermediate between B cells and plasmacells.
The CCR9–CCL25/TECK and CCR10–CCL28/MEC axeshave homeostatic roles in mucosal immunity by homing ofantigen-specific IgA ASCs to small intestinal mucosa [36]and epithelial cells in various mucosal tissues [37],respectively. Many peripheral blood immature plasma cellswere negative for CCR9 (16·7%), and no difference wasfound in the expression of CCR9 on peripheral blood imma-
(a)
CD19 CD138 CXCR3 Merged image HE
control
UC-NI
UC-I
CD
(b)
(c)
(d)
(e)
(f)
HC (n =
7)
UC I (n
= 8
)
CD I (n
= 7
)
CD NI (
n =
8)
UC NI (
n =
8)0
102030405060708090/mm2
Num
ber
of C
D19
+ C
D13
8+ C
XC
R3+
trip
le-p
ositi
ve c
ells
**
Fig. 8. Triple immunofluorescent staining for CD19 (blue), CD138 (green) and CXCR3 (red) in the lamina propria of control (a), non-inflamed
ulcerative colitis (UC) (b), inflamed UC (c,d) and inflamed Crohn’s disease (CD) (e). (d) High-magnification images of the boxed areas in (c). The
specificity of each antibody was confirmed by negative reactivity of isotype- and species-matched immunoglobulin. Adjacent sections were stained
with haematoxylin and eosin. This figure is representative of staining performed in four controls and six patients with UC or CD. Scale bar
represents 200 mm. (f) Numbers of CD19, CD138 and CXCR3 triple-positive cells in the lamina propria of healthy controls (HC) and patients with
non-inflamed UC (UC-NI), inflamed UC (UC-I), non-inflamed CD (CD-NI) and inflamed CD (CD-I). Numbers of patients are indicated.
**P < 0·01 versus control.
S. Hosomi et al.
222 © 2010 The AuthorsClinical and Experimental Immunology © 2010 British Society for Immunology, Clinical and Experimental Immunology, 163: 215–224

ture plasma cells between IBD and controls. Many peripheralblood immature plasma cells were positive for CCR10(60·7% positive), but no differences in CCR10 expressionwere found on peripheral blood immature plasma cells or inCCL28 expression in colonic mucosa between patients withIBD and controls. These results suggest that CCR9 andCCR10 may not explain the migration of peripheral bloodimmature plasma cells to inflamed sites in UC specifically.
Hardi et al. [38] reported positive safety data from Phase 1trials of anti-CXCL10 monoclonal antibody (mAb) (MDX-1100) for the treatment of UC and rheumatoid arthritis.Inhibition of the CXCL10/CXCR3 axis may become a noveltherapeutic approach for the treatment of UC due to thereduction of the migration of immature plasma cells. Thepathogenic effects of immature plasma cells have not beenstudied in detail. Further studies are needed to clarify thefunctions of immature plasma cells in the pathogenesis ofUC.
In conclusion, our study demonstrates that peripheralblood immature plasma cells in patients with IBD areelevated with a positive correlation to clinical activity.CXCR3 expression on peripheral blood immature plasma
cells is selectively up-regulated in patients with UC. Theseresults indicate that immature plasma cells may migratetowards inflammatory sites of UC via the CXCR3 axis andparticipate in the pathogenesis of UC.
Disclosure
The authors declare no conflicts of interest.
References
1 Bitton A, Peppercorn MA, Antonioli DA et al. Clinical, biological,
and histologic parameters as predictors of relapse in ulcerative
colitis. Gastroenterology 2001; 120:13–20.
2 Fiocchi C. Inflammatory bowel disease: etiology and pathogenesis.
Gastroenterology 1998; 115:182–205.
3 Yacyshyn BR. Activated CD19+ B cell lamina propria lymphocytes
in ulcerative colitis. Immunol Cell Biol 1993; 71:265–74.
4 Kett K, Rognum TO, Brandtzaeg P. Mucosal subclass distribution
of immunoglobulin G-producing cells is different in ulcerative
colitis and Crohn’s disease of the colon. Gastroenterology 1987;
93:919–24.
5 Hibi T, Ohara M, Toda K et al. In vitro anticolon antibody produc-
Fig. 9. Immunofluorescent staining for CXCL9,
CXCL10 and CXCL11 in the lamina propria of
control (a), non-inflamed ulcerative colitis
(UC) (b), inflamed UC (c) and inflamed
Crohn’s disease (CD) (d). The specificity of
each antibody was confirmed by negative
reactivity of isotype- and species-matched
immunoglobulin. Scale bar represents 200 mm.
(a)
CXCL9 CXCL10 CXCL11
control
UC-I
UC-NI
CD
(b)
(c)
(d)
Increased immature plasma cells overexpress CXCR3/4 in UC
223© 2010 The AuthorsClinical and Experimental Immunology © 2010 British Society for Immunology, Clinical and Experimental Immunology, 163: 215–224

tion by mucosal or peripheral blood lymphocytes from patients
with ulcerative colitis. Gut 1990; 31:1371–6.
6 Hibi T, Aiso S, Ishikawa M et al. Circulating antibodies to the
surface antigens on colon epithelial cells in ulcerative colitis. Clin
Exp Immunol 1983; 54:163–8.
7 Onuma EK, Amenta PS, Ramaswamy K, Lin JJ, Das KM. Autoim-
munity in ulcerative colitis (UC): a predominant colonic mucosal B
cell response against human tropomyosin isoform 5. Clin Exp
Immunol 2000; 121:466–71.
8 Ebert EC, Geng X, Bajpai M, Pan Z, Tatar E, Das KM. Antibody to
tropomyosin isoform 5 and complement induce the lysis of
colonocytes in ulcerative colitis. Am J Gastroenterol 2009;
104:2996–3003.
9 Duerr RH, Targan SR, Landers CJ, Sutherland LR, Shanahan F.
Anti-neutrophil cytoplasmic antibodies in ulcerative colitis. Com-
parison with other colitides/diarrheal illnesses. Gastroenterology
1991; 100:1590–6.
10 Oudkerk Pool M, Ellerbroek PM, Ridwan BU et al. Serum
antineutrophil cytoplasmic autoantibodies in inflammatory bowel
disease are mainly associated with ulcerative colitis. A correlation
study between perinuclear antineutrophil cytoplasmic autoanti-
bodies and clinical parameters, medical, and surgical treatment.
Gut 1993; 34:46–50.
11 Jinno Y, Ohtani H, Nakamura S et al. Infiltration of CD19+ plasma
cells with frequent labeling of Ki-67 in corticosteroid-resistant
active ulcerative colitis. Virchows Arch 2006; 448:412–21.
12 Hirata N, Oshitani N, Kamata N et al. Proliferation of immature
plasma cells in pouchitis mucosa in patients with ulcerative colitis.
Inflamm Bowel Dis 2008; 14:1084–90.
13 Kunkel EJ, Butcher EC. Plasma-cell homing. Nat Rev 2003; 3:822–9.
14 Moser B, Loetscher P. Lymphocyte traffic control by chemokines.
Nat Immunol 2001; 2:123–8.
15 Bowman EP, Campbell JJ, Soler D et al. Developmental switches in
chemokine response profiles during B cell differentiation and
maturation. J Exp Med 2000; 191:1303–18.
16 Gerard C, Rollins BJ. Chemokines and disease. Nat Immunol 2001;
2:108–15.
17 Yuan YH, ten Hove T, The FO, Slors JF, van Deventer SJ, te Velde
AA. Chemokine receptor CXCR3 expression in inflammatory
bowel disease. Inflamm Bowel Dis 2001; 7:281–6.
18 Mikami S, Nakase H, Yamamoto S et al. Blockade of CXCL12/
CXCR4 axis ameliorates murine experimental colitis. J Pharmacol
Exp Ther 2008; 327:383–92.
19 Schroeder KW, Tremaine WJ, Ilstrup DM. Coated oral
5-aminosalicylic acid therapy for mildly to moderately active ulcer-
ative colitis. A randomized study. N Engl J Med 1987; 317:1625–9.
20 Best WR, Becktel JM, Singleton JW, Kern F, Jr. Development of a
Crohn’s disease activity index. National Cooperative Crohn’s
Disease Study. Gastroenterology 1976; 70:439–44.
21 Froslie KF, Jahnsen J, Moum BA, Vatn MH. Mucosal healing in
inflammatory bowel disease: results from a Norwegian population-
based cohort. Gastroenterology 2007; 133:412–22.
22 Harada Y, Kawano MM, Huang N et al. Identification of early
plasma cells in peripheral blood and their clinical significance. Br J
Haematol 1996; 92:184–91.
23 Noguchi A, Watanabe K, Narumi S et al. The production of
interferon-gamma-inducible protein 10 by granulocytes and
monocytes is associated with ulcerative colitis disease activity.
J Gastroenterol 2007; 42:947–56.
24 Gasperini S, Marchi M, Calzetti F et al. Gene expression and pro-
duction of the monokine induced by IFN-gamma (MIG), IFN-
inducible T cell alpha chemoattractant (I-TAC), and IFN-gamma-
inducible protein-10 (IP-10) chemokines by human neutrophils.
J Immunol 1999; 162:4928–37.
25 Jego G, Robillard N, Puthier D et al. Reactive plasmacytoses are
expansions of plasmablasts retaining the capacity to differentiate
into plasma cells. Blood 1999; 94:701–12.
26 Kawano MM, Mihara K, Huang N, Tsujimoto T, Kuramoto A.
Differentiation of early plasma cells on bone marrow stromal cells
requires interleukin-6 for escaping from apoptosis. Blood 1995;
85:487–94.
27 Kawachiya T, Oshitani N, Jinno Y et al. Significance of increased
proliferation of immature plasma cells in the appendix of patients
with ulcerative colitis. Int J Mol Med 2005; 15:417–23.
28 Henneken M, Dorner T, Burmester GR, Berek C. Differential
expression of chemokine receptors on peripheral blood B cells
from patients with rheumatoid arthritis and systemic lupus
erythematosus. Arthritis Res Ther 2005; 7:R1001–13.
29 Sorensen TL, Roed H, Sellebjerg F. Chemokine receptor expression
on B cells and effect of interferon-beta in multiple sclerosis.
J Neuroimmunol 2002; 122:125–31.
30 Autschbach F, Giese T, Gassler N et al. Cytokine/chemokine
messenger-RNA expression profiles in ulcerative colitis and
Crohn’s disease. Virchows Arch 2002; 441:500–13.
31 Singh UP, Venkataraman C, Singh R, Lillard JW, Jr. CXCR3 axis:
role in inflammatory bowel disease and its therapeutic implication.
Endocr Metab Immune Disord Drug Targets 2007; 7:111–23.
32 Uguccioni M, Gionchetti P, Robbiani DF et al. Increased expression
of IP-10, IL-8, MCP-1, and MCP-3 in ulcerative colitis. Am J Pathol
1999; 155:331–6.
33 Egesten A, Eliasson M, Olin AI et al. The proinflammatory CXC-
chemokines GRO-alpha/CXCL1 and MIG/CXCL9 are concomi-
tantly expressed in ulcerative colitis and decrease during treatment
with topical corticosteroids. Int J Colorectal Dis 2007; 22:1421–7.
34 Hargreaves DC, Hyman PL, Lu TT et al. A coordinated change in
chemokine responsiveness guides plasma cell movements. J Exp
Med 2001; 194:45–56.
35 Nakayama T, Hieshima K, Izawa D, Tatsumi Y, Kanamaru A, Yoshie
O. Cutting edge: profile of chemokine receptor expression on
human plasma cells accounts for their efficient recruitment to
target tissues. J Immunol 2003; 170:1136–40.
36 Pabst O, Ohl L, Wendland M et al. Chemokine receptor CCR9
contributes to the localization of plasma cells to the small intestine.
J Exp Med 2004; 199:411–16.
37 Kunkel EJ, Kim CH, Lazarus NH et al. CCR10 expression is a
common feature of circulating and mucosal epithelial tissue IgA
Ab-secreting cells. J Clin Invest 2003; 111:1001–10.
38 Hardi R, Mayer L, Targan SR, Yellin M, Cardarelli PM, Das KM. A
phase 1 open-label, single-dose, dose-escalation study of Mdx-
1100, a high-affinity, neutralizing, fully human Igg1 kappa anti-
CXCL10 (Ip10) monoclonal antibody, in ulcerative colitis.
Gastroenterology 2008; 134:A99–A100.
S. Hosomi et al.
224 © 2010 The AuthorsClinical and Experimental Immunology © 2010 British Society for Immunology, Clinical and Experimental Immunology, 163: 215–224
Copyright © 2022 FDOKUMEN




















