
Clean public areas, facilities and equipment D1.HHK.CL3.02 Trainee Manual

Increased cell growth and tumorigenicity in human prostate LNCaP cells byoverexpression to cyclin D1
Yian Chen, Luis A Martinez, Margaret LaCava, Lezlee Coghlan and Claudio J Conti
The University of Texas MD Anderson Cancer Center, Science Park-Research Division, Smithville, Texas 78957, USA
Deregulated expression of cyclin D1 has been found inseveral types of human tumors. In order to investigatefactors involved in human prostate cancer progression,we studied the e�ects of cyclin D1 overexpression onhuman prostate cancer cell proliferation and tumorigeni-city by transfecting LNCaP cells with a retroviral vectorcontaining human cyclin D1 cDNA. When compared tothe parental and control-vector transfected LNCaP cells,these cyclin D1-transfected cells had more cells in S-phase and lower growth factor requirements. Further-more, these cells grew more in androgen-free medium.We also detected higher levels of Rb phosphorylation andE2F-1 protein levels in LNCaP/cyclin D1 cells than thatin the parental and vector control cells in medium with orwithout androgen. Cyclin D1 transfected clones formedtumors more rapidly than control and parental cells.These tumors were refractory to the androgen-ablationtreatment by castration, whereas tumors from parentaland vector-control LNCaP cells regressed within 4 weeksafter castration. These results suggest that overexpres-sion of cyclin D1 changes the growth properties,increases tumorigenicity and decreases the requirementfor androgen stimulation in LNCaP cells both in vitroand in vivo.
Keywords: prostate cancer cell lines; androgen; cyclinD1; gene transfection; cell proliferation; tumorigenicity
Introduction
Cyclin D1 is a 36-Kda protein that belongs to a familyof cell-cycle regulators called cyclins (Xiong et al.,1991, 1992a). It functions in the G1 phase of cell cycleand activates the kinase activities of G1 cyclin-dependent kinases (for review, see Sherr, 1993, 1994).The active cyclin/cdk complexes phosphorylate pro-teins which are important in cell-cycle control. Oneputative substrate of these cyclin D1-activated kinasesis the retinoblastoma protein (Rb). The hypopho-sphorylated form of Rb prevents cells from enteringthe S phase. When Rb is hyperphosphorylated bykinases (e.g., cdks), it can no longer prevent the cellsfrom entering S phase (for review, see Weinberg, 1991).The activities of cyclin/cdk can be inhibited by severalnegative cell-cycle regulatory proteins (for review, seeSherr and Roberts, 1995).Overexpression of cyclin D1 shortens the G1 phase,
decreases cell size and increases tumorigenicity in rat
embryo ®broblasts (Jiang et al., 1993a; Quelle et al.,1993; Resnitzky et al., 1994). Induction of cyclin D1 inthe human breast cancer cell line, T-47D, also increasesthe rate of transition from G1 to S phase and theinduced cyclin D1 is su�cient to allow cells tocomplete the cell cycle even if they are arrested inearly G1 phase after growth-factor deprivation(Musgrove et al., 1994). Microinjection of antibodiesor antisense plasmids into cells in mid-G1 inhibitscyclin D1 and prevents the cells from entering S phase(Baldin et al., 1993; Quelle et al., 1993; Lukas et al.,1994a,b; Tam et al., 1994; Zhou et al., 1995). Thisindicates that cyclin D1 limits the rate of progressionthrough G1 phase. Deregulated expression of cyclin D1has been found in several types of tumors. In someparathyroid adenomas, cyclin D1 expression is elevatedby a translocation that juxtaposes the cyclin D1parathyroid hormone genes at 11p15 (Rosenberg etal., 1991). Ampli®cation of the human 11q13 regionand elevated cyclin D1 transcript levels have beenreported in a subset of head and neck squamous cellcarcinomas, esophagus, breast, bladder and livercarcinomas, and B-cell lymphomas. (Lammie et al.,1991; Buckley et al., 1993; Jiang et al., 1992, 1993b;Schuuring et al., 1992; Zhang et al., 1993). Thus, cyclinD1 is probably involved in tumor development.In androgen-induced rat prostate cell proliferation,
cyclin D1 is greatly induced when prostate cells passfrom G0 into early G1 phase. The changes in the cyclinD1 levels correlate with the proliferation rate afterandrogen stimulation in castrated rats (Furuya et al.,1995; Chen et al., 1996). This suggests that cyclin D1plays an important role in prostate cell proliferation.We also examined the expression of cyclin D1 in threehuman prostate cancer cell lines (PC3, DU145 andLNCaP). The growth of PC3 and DU145 cells isandrogen independent, and that of LNCaP is androgendependent. We found that cyclin D1 was constitutivelyexpressed in PC3 and DU145 cells grown with orwithout serum but cyclin D1 expression was signifi-cantly lower in LNCaP cells grown without serum(Chen et al., 1995). So far it is still not clear whetherthis constitutive high expression of cyclin D1 inandrogen-independent cells is related to their non-responsiveness to androgen ablation in prostate cancer.To further understand the role of cyclin D1 in the
progression of prostate cancer, we overexpressedcyclin D1 in the LNCaP cell line by using a retroviralvector containing human cyclin D1 cDNA (Jiang etal., 1993a). The LNCaP cell line was derived from alymph node of a patient with metastatic stage Dprostate cancer (Horoszewicz et al., 1980, 1983).LNCaP cells are well di�erentiated and have thecharacteristics of prostate cells; they produce prostate-speci®c antigen (Montgomery et al., 1992), prostate-
Correspondence: CJ ContiThis work was supported by American Cancer Society CN143Received 30 April 1995; revised 10 November 1997; accepted 10November 1997
Oncogene (1998) 16, 1913 ± 1920 1998 Stockton Press All rights reserved 0950 ± 9232/98 $12.00
http://www.stockton-press.co.uk/onc
speci®c membrane antigen (Israeli et al., 1994) andandrogen receptor (Veldscholte et al., 1990). Althoughthere are mutations in androgen receptor of LNCaPcells, these cells are androgen responsive in cell culture(Sonnenschein et al., 1989) and androgen dependentwhen they form tumors in male nude mice(Horoszewicz et al., 1980, 1983; Lim et al., 1993).LNCaP cells can become more tumorigenic when theyare mixed with matrigel (Pretlow et al., 1991; Lim etal., 1993) or bone ®broblasts (Wu et al., 1994) andsome tumors become androgen independent in nudemice (Wu et al., 1994). Due to these characteristics,we chose this cell line to study the relationshipbetween cyclin D1 overexpression and androgenindependence in the progression of prostate cancercells. In doing so, we examined the e�ects of androgenon the expression of cyclin D1 in LNCaP cells. Weinvestigated the e�ects of cyclin D1 overexpression onLNCaP cell-cycle progression, cell proliferation,androgen responsiveness and involvement of theother cell-cycle regulators in cell culture. Finally, westudied the tumorigenicity of various LNCaP deriva-tives and their response to androgen ablation in malenude mice.
Results
Induction of cyclin D1 mRNA expression in LNCaPcells by androgen
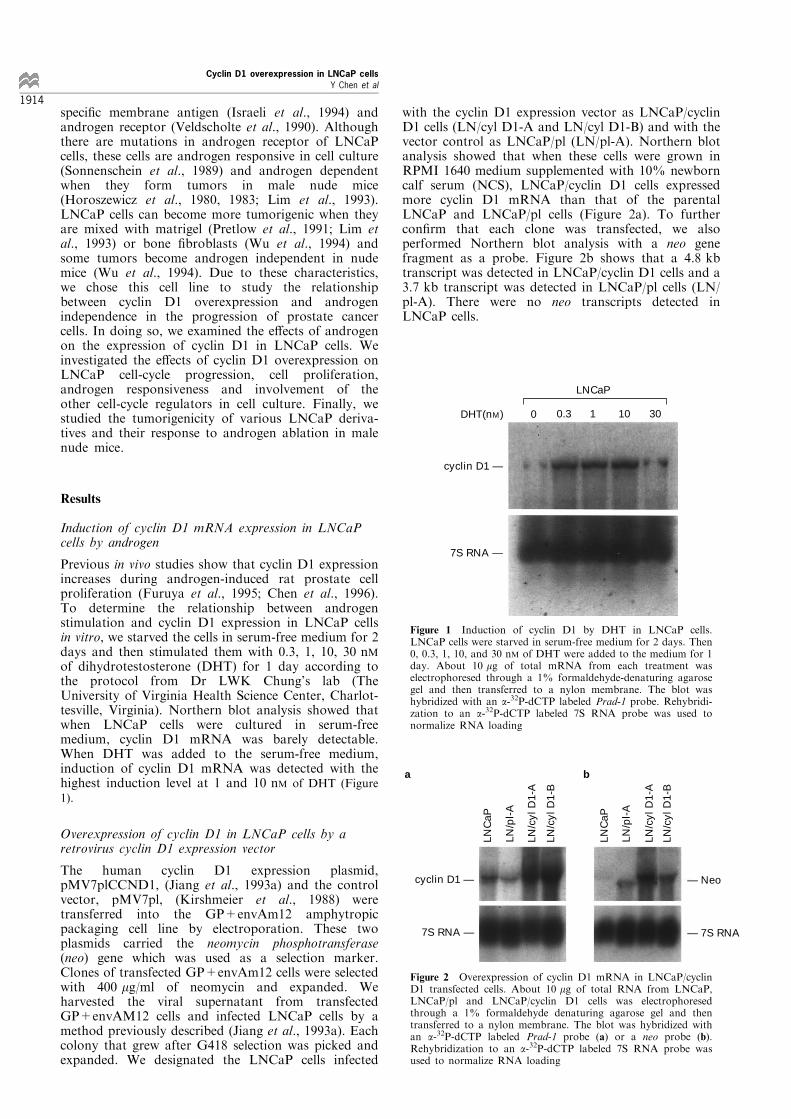
Previous in vivo studies show that cyclin D1 expressionincreases during androgen-induced rat prostate cellproliferation (Furuya et al., 1995; Chen et al., 1996).To determine the relationship between androgenstimulation and cyclin D1 expression in LNCaP cellsin vitro, we starved the cells in serum-free medium for 2days and then stimulated them with 0.3, 1, 10, 30 nMof dihydrotestosterone (DHT) for 1 day according tothe protocol from Dr LWK Chung's lab (TheUniversity of Virginia Health Science Center, Charlot-tesville, Virginia). Northern blot analysis showed thatwhen LNCaP cells were cultured in serum-freemedium, cyclin D1 mRNA was barely detectable.When DHT was added to the serum-free medium,induction of cyclin D1 mRNA was detected with thehighest induction level at 1 and 10 nM of DHT (Figure
1).
Overexpression of cyclin D1 in LNCaP cells by aretrovirus cyclin D1 expression vector
The human cyclin D1 expression plasmid,pMV7plCCND1, (Jiang et al., 1993a) and the controlvector, pMV7pl, (Kirshmeier et al., 1988) weretransferred into the GP+envAm12 amphytropicpackaging cell line by electroporation. These twoplasmids carried the neomycin phosphotransferase(neo) gene which was used as a selection marker.Clones of transfected GP+envAm12 cells were selectedwith 400 mg/ml of neomycin and expanded. Weharvested the viral supernatant from transfectedGP+envAM12 cells and infected LNCaP cells by amethod previously described (Jiang et al., 1993a). Eachcolony that grew after G418 selection was picked andexpanded. We designated the LNCaP cells infected
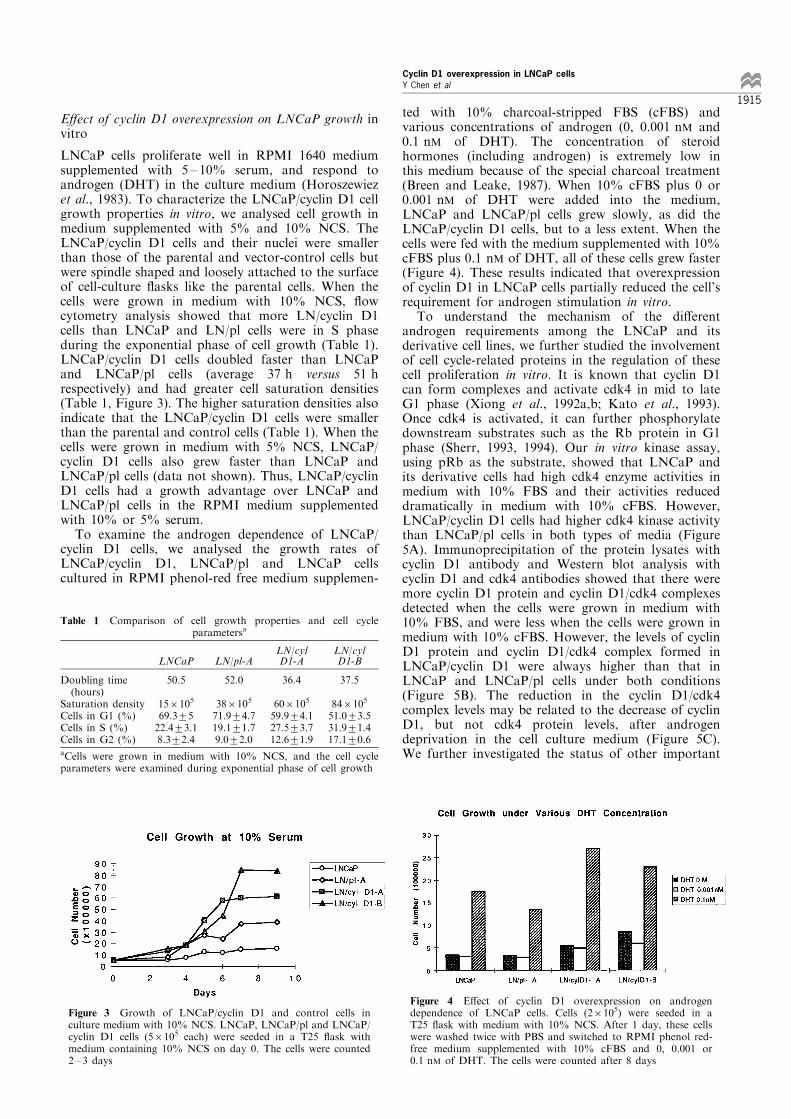
with the cyclin D1 expression vector as LNCaP/cyclinD1 cells (LN/cyl D1-A and LN/cyl D1-B) and with thevector control as LNCaP/pl (LN/pl-A). Northern blotanalysis showed that when these cells were grown inRPMI 1640 medium supplemented with 10% newborncalf serum (NCS), LNCaP/cyclin D1 cells expressedmore cyclin D1 mRNA than that of the parentalLNCaP and LNCaP/pl cells (Figure 2a). To furthercon®rm that each clone was transfected, we alsoperformed Northern blot analysis with a neo genefragment as a probe. Figure 2b shows that a 4.8 kbtranscript was detected in LNCaP/cyclin D1 cells and a3.7 kb transcript was detected in LNCaP/pl cells (LN/pl-A). There were no neo transcripts detected inLNCaP cells.
0 0.3 1 10 30
cyclin D1 —
7S RNA —
LNCaP
DHT(nM)
Figure 1 Induction of cyclin D1 by DHT in LNCaP cells.LNCaP cells were starved in serum-free medium for 2 days. Then0, 0.3, 1, 10, and 30 nM of DHT were added to the medium for 1day. About 10 mg of total mRNA from each treatment waselectrophoresed through a 1% formaldehyde-denaturing agarosegel and then transferred to a nylon membrane. The blot washybridized with an a-32P-dCTP labeled Prad-1 probe. Rehybridi-zation to an a-32P-dCTP labeled 7S RNA probe was used tonormalize RNA loading
LNC
aP
LNC
aP
LN/p
l-A
LN/p
l-A
LN/c
yl D
1-A
LN/c
yl D
1-A
LN/c
yl D
1-B
LN/c
yl D
1-B
cyclin D1 —
7S RNA —
— Neo
— 7S RNA
a b
Figure 2 Overexpression of cyclin D1 mRNA in LNCaP/cyclinD1 transfected cells. About 10 mg of total RNA from LNCaP,LNCaP/pl and LNCaP/cyclin D1 cells was electrophoresedthrough a 1% formaldehyde denaturing agarose gel and thentransferred to a nylon membrane. The blot was hybridized withan a-32P-dCTP labeled Prad-1 probe (a) or a neo probe (b).Rehybridization to an a-32P-dCTP labeled 7S RNA probe wasused to normalize RNA loading
Cyclin D1 overexpression in LNCaP cellsY Chen et al
1914
E�ect of cyclin D1 overexpression on LNCaP growth invitro
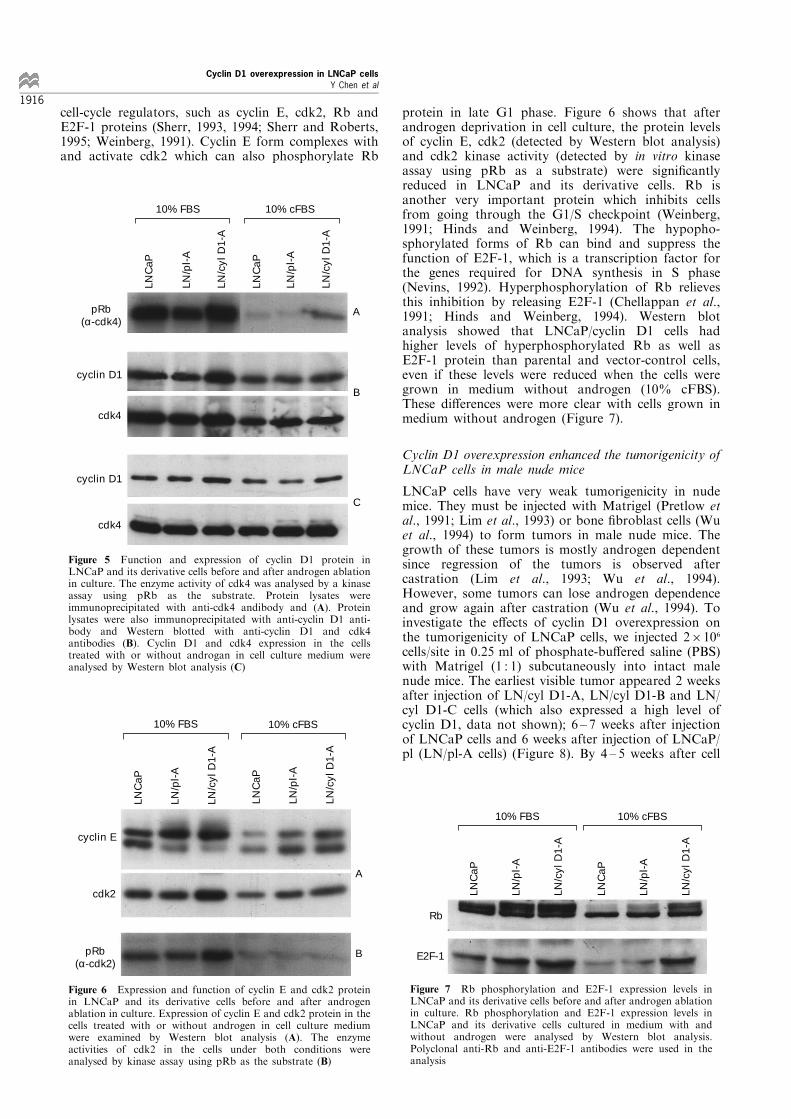
LNCaP cells proliferate well in RPMI 1640 mediumsupplemented with 5 ± 10% serum, and respond toandrogen (DHT) in the culture medium (Horoszewiezet al., 1983). To characterize the LNCaP/cyclin D1 cellgrowth properties in vitro, we analysed cell growth inmedium supplemented with 5% and 10% NCS. TheLNCaP/cyclin D1 cells and their nuclei were smallerthan those of the parental and vector-control cells butwere spindle shaped and loosely attached to the surfaceof cell-culture ¯asks like the parental cells. When thecells were grown in medium with 10% NCS, ¯owcytometry analysis showed that more LN/cyclin D1cells than LNCaP and LN/pl cells were in S phaseduring the exponential phase of cell growth (Table 1).LNCaP/cyclin D1 cells doubled faster than LNCaPand LNCaP/pl cells (average 37 h versus 51 hrespectively) and had greater cell saturation densities(Table 1, Figure 3). The higher saturation densities alsoindicate that the LNCaP/cyclin D1 cells were smallerthan the parental and control cells (Table 1). When thecells were grown in medium with 5% NCS, LNCaP/cyclin D1 cells also grew faster than LNCaP andLNCaP/pl cells (data not shown). Thus, LNCaP/cyclinD1 cells had a growth advantage over LNCaP andLNCaP/pl cells in the RPMI medium supplementedwith 10% or 5% serum.To examine the androgen dependence of LNCaP/
cyclin D1 cells, we analysed the growth rates ofLNCaP/cyclin D1, LNCaP/pl and LNCaP cellscultured in RPMI phenol-red free medium supplemen-
ted with 10% charcoal-stripped FBS (cFBS) andvarious concentrations of androgen (0, 0.001 nM and0.1 nM of DHT). The concentration of steroidhormones (including androgen) is extremely low inthis medium because of the special charcoal treatment(Breen and Leake, 1987). When 10% cFBS plus 0 or0.001 nM of DHT were added into the medium,LNCaP and LNCaP/pl cells grew slowly, as did theLNCaP/cyclin D1 cells, but to a less extent. When thecells were fed with the medium supplemented with 10%cFBS plus 0.1 nM of DHT, all of these cells grew faster(Figure 4). These results indicated that overexpressionof cyclin D1 in LNCaP cells partially reduced the cell'srequirement for androgen stimulation in vitro.To understand the mechanism of the di�erent
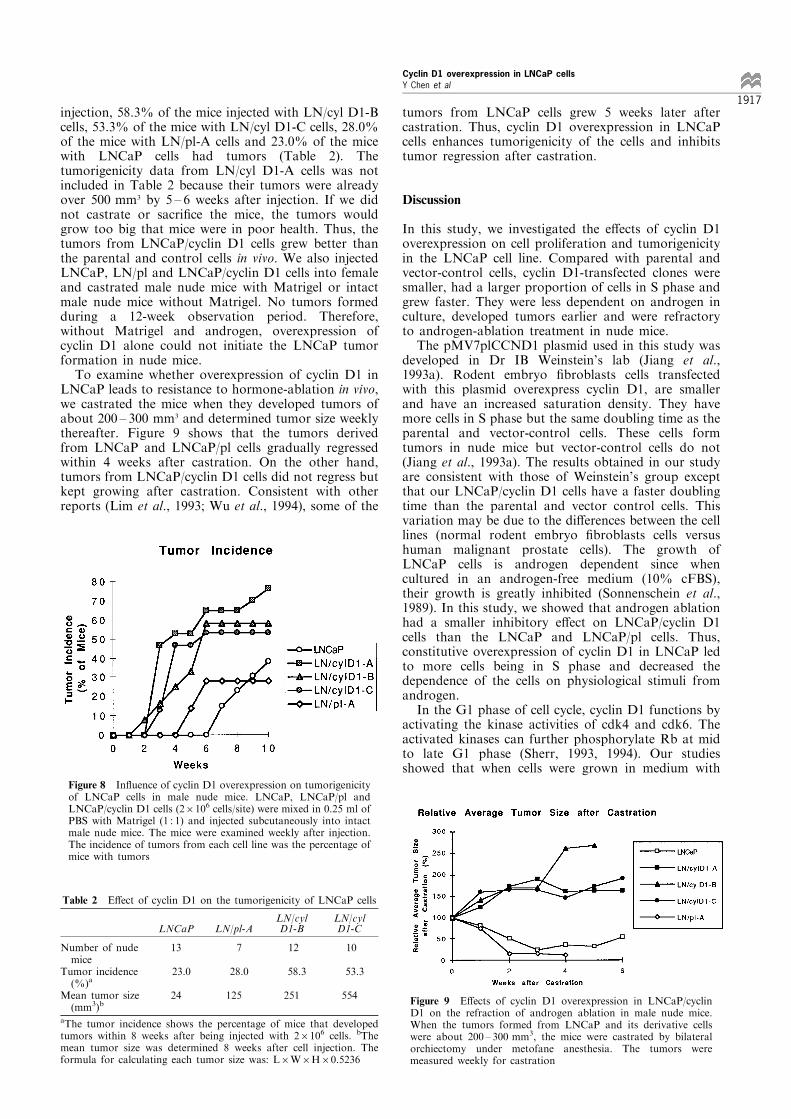
androgen requirements among the LNCaP and itsderivative cell lines, we further studied the involvementof cell cycle-related proteins in the regulation of thesecell proliferation in vitro. It is known that cyclin D1can form complexes and activate cdk4 in mid to lateG1 phase (Xiong et al., 1992a,b; Kato et al., 1993).Once cdk4 is activated, it can further phosphorylatedownstream substrates such as the Rb protein in G1phase (Sherr, 1993, 1994). Our in vitro kinase assay,using pRb as the substrate, showed that LNCaP andits derivative cells had high cdk4 enzyme activities inmedium with 10% FBS and their activities reduceddramatically in medium with 10% cFBS. However,LNCaP/cyclin D1 cells had higher cdk4 kinase activitythan LNCaP/pl cells in both types of media (Figure5A). Immunoprecipitation of the protein lysates withcyclin D1 antibody and Western blot analysis withcyclin D1 and cdk4 antibodies showed that there weremore cyclin D1 protein and cyclin D1/cdk4 complexesdetected when the cells were grown in medium with10% FBS, and were less when the cells were grown inmedium with 10% cFBS. However, the levels of cyclinD1 protein and cyclin D1/cdk4 complex formed inLNCaP/cyclin D1 were always higher than that inLNCaP and LNCaP/pl cells under both conditions(Figure 5B). The reduction in the cyclin D1/cdk4complex levels may be related to the decrease of cyclinD1, but not cdk4 protein levels, after androgendeprivation in the cell culture medium (Figure 5C).We further investigated the status of other important
Table 1 Comparison of cell growth properties and cell cycleparametersa
LN/cyl LN/cylLNCaP LN/pl-A D1-A D1-B
Doubling time(hours)
Saturation densityCells in G1 (%)Cells in S (%)Cells in G2 (%)
50.5
156105
69.3+522.4+3.18.3+2.4
52.0
386105
71.9+4.719.1+1.79.0+2.0
36.4
606105
59.9+4.127.5+3.712.6+1.9
37.5
846105
51.0+3.531.9+1.417.1+0.6
aCells were grown in medium with 10% NCS, and the cell cycleparameters were examined during exponential phase of cell growth
Figure 3 Growth of LNCaP/cyclin D1 and control cells inculture medium with 10% NCS. LNCaP, LNCaP/pl and LNCaP/cyclin D1 cells (56105 each) were seeded in a T25 ¯ask withmedium containing 10% NCS on day 0. The cells were counted2 ± 3 days
Figure 4 E�ect of cyclin D1 overexpression on androgendependence of LNCaP cells. Cells (26105) were seeded in aT25 ¯ask with medium with 10% NCS. After 1 day, these cellswere washed twice with PBS and switched to RPMI phenol red-free medium supplemented with 10% cFBS and 0, 0.001 or0.1 nM of DHT. The cells were counted after 8 days
Cyclin D1 overexpression in LNCaP cellsY Chen et al
1915
cell-cycle regulators, such as cyclin E, cdk2, Rb andE2F-1 proteins (Sherr, 1993, 1994; Sherr and Roberts,1995; Weinberg, 1991). Cyclin E form complexes withand activate cdk2 which can also phosphorylate Rb
protein in late G1 phase. Figure 6 shows that afterandrogen deprivation in cell culture, the protein levelsof cyclin E, cdk2 (detected by Western blot analysis)and cdk2 kinase activity (detected by in vitro kinaseassay using pRb as a substrate) were signi®cantlyreduced in LNCaP and its derivative cells. Rb isanother very important protein which inhibits cellsfrom going through the G1/S checkpoint (Weinberg,1991; Hinds and Weinberg, 1994). The hypopho-sphorylated forms of Rb can bind and suppress thefunction of E2F-1, which is a transcription factor forthe genes required for DNA synthesis in S phase(Nevins, 1992). Hyperphosphorylation of Rb relievesthis inhibition by releasing E2F-1 (Chellappan et al.,1991; Hinds and Weinberg, 1994). Western blotanalysis showed that LNCaP/cyclin D1 cells hadhigher levels of hyperphosphorylated Rb as well asE2F-1 protein than parental and vector-control cells,even if these levels were reduced when the cells weregrown in medium without androgen (10% cFBS).These di�erences were more clear with cells grown inmedium without androgen (Figure 7).
Cyclin D1 overexpression enhanced the tumorigenicity ofLNCaP cells in male nude mice
LNCaP cells have very weak tumorigenicity in nudemice. They must be injected with Matrigel (Pretlow etal., 1991; Lim et al., 1993) or bone ®broblast cells (Wuet al., 1994) to form tumors in male nude mice. Thegrowth of these tumors is mostly androgen dependentsince regression of the tumors is observed aftercastration (Lim et al., 1993; Wu et al., 1994).However, some tumors can lose androgen dependenceand grow again after castration (Wu et al., 1994). Toinvestigate the e�ects of cyclin D1 overexpression onthe tumorigenicity of LNCaP cells, we injected 26106
cells/site in 0.25 ml of phosphate-bu�ered saline (PBS)with Matrigel (1 : 1) subcutaneously into intact malenude mice. The earliest visible tumor appeared 2 weeksafter injection of LN/cyl D1-A, LN/cyl D1-B and LN/cyl D1-C cells (which also expressed a high level ofcyclin D1, data not shown); 6 ± 7 weeks after injectionof LNCaP cells and 6 weeks after injection of LNCaP/pl (LN/pl-A cells) (Figure 8). By 4 ± 5 weeks after cell
LNC
aP
LNC
aP
LN/p
l-A
LN/p
l-A
LN/c
yl D
1-A
LN/c
yl D
1-A
pRb (α-cdk4)
cyclin D1
cyclin D1
cdk4
cdk4
A
B
C
10% FBS 10% cFBS
Figure 5 Function and expression of cyclin D1 protein inLNCaP and its derivative cells before and after androgen ablationin culture. The enzyme activity of cdk4 was analysed by a kinaseassay using pRb as the substrate. Protein lysates wereimmunoprecipitated with anti-cdk4 andibody and (A). Proteinlysates were also immunoprecipitated with anti-cyclin D1 anti-body and Western blotted with anti-cyclin D1 and cdk4antibodies (B). Cyclin D1 and cdk4 expression in the cellstreated with or without androgan in cell culture medium wereanalysed by Western blot analysis (C)
cyclin E
cdk2
pRb (α-cdk2)
LNC
aP
LN/p
l-A
LN/c
yl D
1-A
LNC
aP
LN/p
l-A
LN/c
yl D
1-A
10% FBS 10% cFBS
A
B
Figure 6 Expression and function of cyclin E and cdk2 proteinin LNCaP and its derivative cells before and after androgenablation in culture. Expression of cyclin E and cdk2 protein in thecells treated with or without androgen in cell culture mediumwere examined by Western blot analysis (A). The enzymeactivities of cdk2 in the cells under both conditions wereanalysed by kinase assay using pRb as the substrate (B)
LNC
aP
LN/p
l-A
LN/c
yl D
1-A
LNC
aP
LN/p
l-A
LN/c
yl D
1-A
Rb
E2F-1
10% FBS 10% cFBS
Figure 7 Rb phosphorylation and E2F-1 expression levels inLNCaP and its derivative cells before and after androgen ablationin culture. Rb phosphorylation and E2F-1 expression levels inLNCaP and its derivative cells cultured in medium with andwithout androgen were analysed by Western blot analysis.Polyclonal anti-Rb and anti-E2F-1 antibodies were used in theanalysis
Cyclin D1 overexpression in LNCaP cellsY Chen et al
1916
injection, 58.3% of the mice injected with LN/cyl D1-Bcells, 53.3% of the mice with LN/cyl D1-C cells, 28.0%of the mice with LN/pl-A cells and 23.0% of the micewith LNCaP cells had tumors (Table 2). Thetumorigenicity data from LN/cyl D1-A cells was notincluded in Table 2 because their tumors were alreadyover 500 mm3 by 5 ± 6 weeks after injection. If we didnot castrate or sacri®ce the mice, the tumors wouldgrow too big that mice were in poor health. Thus, thetumors from LNCaP/cyclin D1 cells grew better thanthe parental and control cells in vivo. We also injectedLNCaP, LN/pl and LNCaP/cyclin D1 cells into femaleand castrated male nude mice with Matrigel or intactmale nude mice without Matrigel. No tumors formedduring a 12-week observation period. Therefore,without Matrigel and androgen, overexpression ofcyclin D1 alone could not initiate the LNCaP tumorformation in nude mice.To examine whether overexpression of cyclin D1 in
LNCaP leads to resistance to hormone-ablation in vivo,we castrated the mice when they developed tumors ofabout 200 ± 300 mm3 and determined tumor size weeklythereafter. Figure 9 shows that the tumors derivedfrom LNCaP and LNCaP/pl cells gradually regressedwithin 4 weeks after castration. On the other hand,tumors from LNCaP/cyclin D1 cells did not regress butkept growing after castration. Consistent with otherreports (Lim et al., 1993; Wu et al., 1994), some of the
tumors from LNCaP cells grew 5 weeks later aftercastration. Thus, cyclin D1 overexpression in LNCaPcells enhances tumorigenicity of the cells and inhibitstumor regression after castration.
Discussion
In this study, we investigated the e�ects of cyclin D1overexpression on cell proliferation and tumorigenicityin the LNCaP cell line. Compared with parental andvector-control cells, cyclin D1-transfected clones weresmaller, had a larger proportion of cells in S phase andgrew faster. They were less dependent on androgen inculture, developed tumors earlier and were refractoryto androgen-ablation treatment in nude mice.The pMV7plCCND1 plasmid used in this study was
developed in Dr IB Weinstein's lab (Jiang et al.,1993a). Rodent embryo ®broblasts cells transfectedwith this plasmid overexpress cyclin D1, are smallerand have an increased saturation density. They havemore cells in S phase but the same doubling time as theparental and vector-control cells. These cells formtumors in nude mice but vector-control cells do not(Jiang et al., 1993a). The results obtained in our studyare consistent with those of Weinstein's group exceptthat our LNCaP/cyclin D1 cells have a faster doublingtime than the parental and vector control cells. Thisvariation may be due to the di�erences between the celllines (normal rodent embryo ®broblasts cells versushuman malignant prostate cells). The growth ofLNCaP cells is androgen dependent since whencultured in an androgen-free medium (10% cFBS),their growth is greatly inhibited (Sonnenschein et al.,1989). In this study, we showed that androgen ablationhad a smaller inhibitory e�ect on LNCaP/cyclin D1cells than the LNCaP and LNCaP/pl cells. Thus,constitutive overexpression of cyclin D1 in LNCaP ledto more cells being in S phase and decreased thedependence of the cells on physiological stimuli fromandrogen.In the G1 phase of cell cycle, cyclin D1 functions by
activating the kinase activities of cdk4 and cdk6. Theactivated kinases can further phosphorylate Rb at midto late G1 phase (Sherr, 1993, 1994). Our studiesshowed that when cells were grown in medium with
Figure 8 In¯uence of cyclin D1 overexpression on tumorigenicityof LNCaP cells in male nude mice. LNCaP, LNCaP/pl andLNCaP/cyclin D1 cells (26106 cells/site) were mixed in 0.25 ml ofPBS with Matrigel (1 : 1) and injected subcutaneously into intactmale nude mice. The mice were examined weekly after injection.The incidence of tumors from each cell line was the percentage ofmice with tumors
Table 2 E�ect of cyclin D1 on the tumorigenicity of LNCaP cells
LN/cyl LN/cylLNCaP LN/pl-A D1-B D1-C
Number of nudemice
Tumor incidence(%)a
Mean tumor size(mm3)b
13
23.0
24
7
28.0
125
12
58.3
251
10
53.3
554
aThe tumor incidence shows the percentage of mice that developedtumors within 8 weeks after being injected with 26106 cells. bThemean tumor size was determined 8 weeks after cell injection. Theformula for calculating each tumor size was: L6W6H60.5236
Figure 9 E�ects of cyclin D1 overexpression in LNCaP/cyclinD1 on the refraction of androgen ablation in male nude mice.When the tumors formed from LNCaP and its derivative cellswere about 200 ± 300 mm3, the mice were castrated by bilateralorchiectomy under metofane anesthesia. The tumors weremeasured weekly for castration
Cyclin D1 overexpression in LNCaP cellsY Chen et al
1917

10% FBS, LNCaP/cyclin D1 cells had higher cdk4kinase activity and higher levels of Rb phosphorylationthan LNCaP and LNCaP/pl cells. Thus, these resultswere consistent with the function of cyclin D1previously described. E2F-1 is another importantprotein in cell cycle regulation because it regulatesthe expression of genes required for DNA synthesis inS phase, including dehydrofolate reductase, DNApolymerase a, thymidine kinase and cdc2 (reviewedby Nevins 1992). The transcription of E2F-1 is alsoknown to be activated by itself (Johnson et al., 1994).The function of E2F-1 is regulated by the members ofthe Rb protein. Hypophosphorylated Rb binds to E2F-1 and inhibit E2F-1 from activating gene transcription.Phosphorylation of Rb relieves this inhibition byreleasing E2F-1 which leads to activation of thisprotein and drives cells into S phase (Chellappan etal., 1991; Hinds and Weinberg, 1994; Johnson, 1995).Our studies showed that when cells were grown inmedium with 10% FBS, LNCaP/cyclin D1 cells hadhigher expression levels of E2F-1 than LNCaP andLNCaP/pl cells. This observation is consistent withprevious reports from other labs stating that cyclin D1/cdk4 induces E2F-1 indirectly by phosphorylation ofRb resulting in the release of E2F-1 which thenactivates its own transcription (Schulze et al., 1994;Oswald et al., 1994; Vairo et al., 1995; Johnson, 1995;Shao and Robbins, 1995).Although cyclin D1 is overexpressed in LNCaP/
cyclin D1 cells, cell proliferation was greatly inhibitedwhen these cells were grown in medium supplementedwith 10% cFBS, as that of the LNCaP and vector-control cells, but to a less degree. The mediumsupplemented with 10% cFBS contains most of thegrowth factors and hormones except steroid hormoneswhich includes androgen. Thus, androgen is animportant mitogen for the LNCaP, vector-control aswell as its cyclin D1 overexpressing cells. The kinaseassay and Western blot analysis performed in thisstudy also indicate that androgen plays a role in theregulation of the cell-cycle machinery. The expressionlevels of cyclin D1, cyclin E, cdk2, E2F-1, phosphor-ylation of Rb protein and cyclin D1/cdk4 complexformation, as well as kinase activities of cdk4 and cdk2were reduced in all of these cells after androgendeprivation from the medium. Hence, these changesmay be a result of the androgen deprivation and partof the factors leading to inhibition of cell proliferation.In LNCaP/cyclin D1 cells, there was a less reduction ofcyclin D1, E2F-1 protein expression, Rb phosphoryla-tion levels and cdk4 kinase activity. This is related tothe constitutively higher expression levels of cyclin D1which may lead to less growth inhibition of these cellsin androgen deprived environment.LNCaP cells are not tumorigenic when they are
injected alone into nude mice. When they are injectedwith Matrigel (Pretlow et al., 1991; Lim et al., 1993) orbone ®broblast cells (Wu et al., 1994), they formtumors in male mice. LNCaP cells are androgendependent not only in vitro but also in vivo as theirtumors regress after castration (Lim et al., 1993; Wu etal., 1994). In our study, LNCaP/cyclin D1 cells mixedwith Matrigel formed tumors in male nude mice earlierthan the parental and vector-control cells did. Aftercastration, these tumors did not regress, whereas thetumors from parental and vector-control cells did.
Thus, overexpression of cyclin D1 enhances prostatetumor growth in male nude mice and reduces tumordependence on androgen in vivo. Interestingly, LNCaP/cyclin D1 cells showed higher resistance to androgenablation in vivo than in vitro. One of the reasons maybe that tumor formation and growth in the nude miceare a more complicated process than cell growth invitro, which is a di�erent growth environment for cells.The degree of tumorigenicity is not only determined bythe ability of cell proliferation but also by theinteraction between host and tumor cells, as well asother events, such as angiogenesis and geneticinstability during tumor formation and growth. Thus,the mechanisms that lead to androgen resistance of thetumors formed from LNCaP/cyclin D1 cells are verycomplicated and will be further studied. We also foundthat cyclin D1-overexpressing LNCaP cells, just likeLNCaP cells, could not form tumors in male nude micewithout Matrigel in female nor in castrated male nudemice with Matrigel. These results suggest that over-expression of cyclin D1 alone is not su�cient forinitiating LNCaP tumor formation in nude mice. Otherfactors such as androgen, components in the Matrigel(including extracellular matrix proteins and growthfactors such as ®broblast growth factors, tissueplasminogen activators, etc.) and other biologicalchanges are required to initiate tumor formation.Many studies have shown that resistance to androgen
ablation in advanced prostate cancer may be the resultof genetic alterations and deregulation of geneexpression. Several groups have reported that muta-tions in androgen receptors or reduction in itsexpression may be related to androgen resistance inadvanced prostate cancer (Tilly et al., 1994; Ruizeveldde Winter et al., 1994). Alterations in tumor suppressorgenes, for example, p53 mutation are also associatedwith hormone resistance in advanced prostate cancer(Navone et al., 1993; Voeller et al., 1994; Kallakury etal., 1994). In addition, the Rb gene is mutated in 20 ±50% of advanced prostate cancers, while mutation israre in early tumors (Isaacs et al., 1994). In this study,we observed that androgen has an important role in theregulation of the cell-cycle machinery including induc-tion of cyclin D1 leading to phosphorylation of Rb andrelease of E2F-1 transcription factor in the LNCaPcells. Thus, changes at any points in this pathway maylead to the inactivation of Rb which represents a criticalevent during tumorigenesis of prostate. The results fromour study suggest that overexpression of cyclin D1 inLNCaP cells cooperates with other biological altera-tions in the cells and facilitates the cell growth, tumorformation and resistance to androgen ablation both invitro and in vivo.
Materials and methods
Cell culture
The human prostate cancer cell line LNCaP (AmericanType Culture Collection, Rockville, MD) was cultured inRPMI 1640 medium (GIBCO, Gaithersburg, MD) supple-mented with NCS or FBS (Hyclone, Logan, Utah). Theamphytropic retrovirus packaging cell line, GP+envAm12,a gift from Dr Yacob Ben-David and Dr J Filmus(Sunnybrook Health Science Center, Toronto, Canada),was cultured in Dulbecco's modi®ed Eagle's medium
Cyclin D1 overexpression in LNCaP cellsY Chen et al
1918

(DMEM) with high glucose, and 10% NCS. Both cell lineswere also supplemented with 2 mM of glutamine and 50 mg/ml of gentamicin and incubated in a humidi®ed atmospherewith 5% CO2. To examine the dependence of LNCaP andits derivatives on androgen, we cultured the cells in RPMI1640 phenol red-free medium (Sigma Chemical Co., St.Louis, MO) supplemented with 10% FBS or 10% cFBSwith various concentrations of DHT (Sigma ChemicalCo.). The dextran and activated charcoal used in thepreparation of cFBS were obtained from Sigma ChemicalCo.
Cyclin D1 retroviral expression plasmid transfection
The cyclin D1 expression plasmid pMV7plCCND1 (Jianget al., 1993a) and the control vector pMV7pl (Kirshmeir,1988) were gifts from Dr IB Weinstein (ColumbiaUniversity, New York, NY). The plasmidpMV7plCCND1 contains a 1.1 kb human cyclin D1cDNA and the 5' Moloney murine leukemia virus longterminal repeat (MoMuLV LTR) as the cyclin D1promoter. A neo gene was included as a selection marker(Jiang et al., 1993a). We transfected pMV7plCCND1 orpMV7pl into GP+envAM12 cells by electroporation.Forty-eight hours later, cells were trypsinized and replatedin DMEM with 10% NCS and 400 mg/ml of G418. G418-resistant clones were selected and expanded. To transfectpMV7plCCND1 or pMV7pl into LNCaP cells, weharvested the viral supernatant of the G418-resistantGP+envAm12 cells and infected LNCaP cells by amethod previously described (Jiang et al., 1993a). Forty-eight hours after transfection, 400 mg/ml of G418 wasadded to the RPMI medium with 10% NCS. Individualcolonies were picked and grown in RPMI medium with10% NCS and 100 mg/ml of G418 for further analysis.
Northern blot analysis
Total RNA was isolated from cells by using TRI-reagent(Molecular Research Center, Inc., Cincinnati, OH). Tenmicrograms of total cellular RNA from each sample waselectrophoresed through a 1.5% agarose gel containing 8%formaldehyde. After electrophoresis, the RNA wastransferred to a nylone membrane by the capillarymethod, and then cross-linked to the membrane withultraviolet light. Prad-1 and neo probes were labeled witha-32P-dCTP by using a random primed DNA labeling kit(Boehringer Mannheim Corporation, Indianapolis IN).The Prad-1 probe, a human cyclin D1 probe, was kindlyprovided by Dr A Arnold (Massachusetts GeneralHospital, Boston, MA). The hybridization was performedat 688C with Quickhyb solution (Strategene, La Jolla, CA).The next day, each membrane was washed twice for 15 mineach time at room temperature with 26SSC and 0.1%SDS and once for 30 min at 688C with 0.16SSC and 0.1%SDS and then exposed to Kodak X-AR5 ®lm for 1 day at7708C. Rehybridization with a-32P-dCTP-labeled 7S RNAwas used to normalize the amount of RNA loaded in eachline.
Western blot analysis and Rb kinase assay
Protein was extracted from cells and kept frozen at7808C by methods described previously (Chen, 1995).One hundred micrograms of protein lysate from eachsample was electrophoresed through a 10% SDS-polyacrylamide gel, transferred to a nitrocellulosemembrane by wet transfer and blotted with variousantibodies. Polyclonal anti-cyclin D, cdk4, cyclin E, cdk2,Rb and E2F-1 antibodies were from Santa Cruz
Biotechnology, Inc. (Santa Cruz, CA). A secondaryantibody conjugated with horseradish peroxidase (Amer-sham Corp., Arlington Heights, IL) was used and the®nal detection was performed by using an ECL kit(Amersham Corp.). For the kinase assay, we followed theprotocol from Dr CJ Sherr's Lab (St. Jude Children'sResearch Hospital, Memphis, TN) (Matsushime et al.,1994). The frozen protein lysates were thawed at 48C and500 mg of protein lysates from each sample wereimmunoprecipitated with either cdk4 or cdk2 antibodies(Santa Cruz, CA). pRb, a GST fusion protein producedin E. coli (Santa Cruz, CA), was used as substrate in thekinase assay. The phosphorylation of pRb was labeled byg-32P-ATP. For quanti®cation of cyclin D1/cdk4 com-plexes formed in the cells, protein lysates wereimmunoprecipitated with cyclin D1 antibody andWestern blotted with cdk4 antibodies.
In vitro cell growth analysis
To characterize cell growth in vitro, we performed ¯owcytometric analysis as well as studies of cell growth curvesand cell population doubling time. For the ¯owcytometric analysis, exponentially growing cells inmedium with 10% NCS were trypsinized, washed withcold PBS and ®xed in 75% ethanol. Propidium iodidewith 2 mg/ml RNase was used to stain the cells. DNAcontent and the percentage of cells in various phases ofthe cell cycle were determined by measuring ¯uorescentintensity with a Coulter Elite Flow cytometer. To examinecell growth, 56105 cells were plated in each of three25 mm2 ¯asks in DMEM with 10%, 5% NCS or 26105
cells in RPMI 1640 phenol red-free medium supplementedwith 10% cFBS with 0, 0.001, 0.1 nM of DHT. Cells werecounted every 2 ± 3 days with a hematocytometer. Thecells were fed twice a week.
Tumorigenicity assay in nude mice
Male nude mice were purchased from Frederick CancerResearch and Development Center (Frederick, MD) andkept aseptically in an athymic animal room. The pMV7pl-and pMV7plCCND1-transfected LNCaP cells and parentalLNCaP cells were trypsinized, washed with PBS, resus-pended in PBS and mixed with an equal volume ofMatrigel (Collaborative Biomedical Products/Becton Dick-inson, Bedford, MA). Then 26106 cells in 0.25 ml of themixture were injected subcutaneously into nude mice by amethod previously described (Lim et al., 1993). The miceunderwent bilateral orchiectomy under metofane anesthesia(Pitman-Moore, Washington Crossing, NJ) when thetumors were 200 ± 300 mm3. All the mice were examinedonce a week, and each tumor's length (L), width (W) andthickness (H) were measured. Tumor volume wascalculated by the formula L6W6H60.5236 (Janek etal., 1975).
AcknowledgementsWe would like to thank Dr IB Weinstein for providing thepMV7plCCND and pMV7pl plasmids; Dr Yacob Ben-David and Dr J Filmus for providing GP+envAM12 cells;Dr A Arnold for providing the Prad-1 (cyclin D1) probe;Dr LWK Chung for sharing their protocol of DHTsupplementation in cell culture and Dr CJ Sherr forproviding their protocol for kinase assays. Dale Wiss andRobert B Whalin for their extensive assistance with theanimal surgery and care; Dennis Walker for ¯ow cyto-metric analysis; Judy G Ing for artwork and Dr MaureenGoode for editing the paper.
Cyclin D1 overexpression in LNCaP cellsY Chen et al
1919

References
Baldin V, Lukas J, Marcote MJ, Pagano M and Draetta G.(1993). Genes & Dev., 7, 812 ± 821.
Breen B and Leake RE. (ed). (1987). Steroid Hormones: APractical Approach. IRL Press: London.
Buckley MF, Sweeney KJE, Hamilton JA, Sini RL, ManningDL, Nicholson RI, deFazio A, Watts CKW, MusgroveEA and Sutherland RL. (1993). Oncogene, 8, 2127 ± 2133.
Chellappan SP, Hiebert S, Mudryj M, Horowitz JM andNevins JR. (1991). Cell, 65, 1053 ± 1061.
Chen Y, Navone NM and Conti CJ. (1995). Urol. Oncol., 1,101 ± 108.
Chen Y, Robles AI, Martinez LA, Liu F, Gimenez-Conti IBand Conti CJ. (1996). Cell Growth Di�er., 7, 1571 ± 1578.
Furuya Y, Lin XS, Walsh JC, Nelson WG and Isaacs JT.(1995). Endocrinology, 136, 1898 ± 1906.
Hinds PW and Weinberg RA. (1994). Curr. Opin. Genet.Dev., 4, 135 ± 141.
Horoszewicz JS, Leong SS, Chu TM, Wajsman ZL,Friedman M, Papsidero L, Kim U, Chai LS, Kakati S,Arya SK and Sandberg AA. (1980). Prog. Clin. Biol. Res.,37, 115 ± 132.
Horoszewicz JS, Leong SS, Kawinski E, Karr JP, RosenthalH, Chu TM, Mirand EA and Murphy GP. (1983). CancerRes., 43, 1809 ± 1818.
Israeli RS, Powell CT, Corr JG, Fair WR and Heston WDW.(1994). Cancer Res., 54, 1807 ± 1811.
Issacs WB, Bova GS, Morton RA, Bussemakers MJ, BrooksJD and Ewing CM. (1994). Cold Spring Harb. Symp.Quant. Biol., 59, 653 ± 659.
Janek P, Briand P and Hartman NR. (1975). Cancer Res., 35,3698 ± 3704.
Jiang W, Kahn SM, Tomita N, Zhang YJ, Lu SH andWeinstein IB. (1992). Cancer Res., 52, 2980 ± 2983.
Jiang W, Kahn SM, Zhou P, Zhang Y-J, Cacace AM, InfanteAS, Doi S, Santella RM and Weinstein IB. (1993a).Oncogene, 8, 3447 ± 3457.
Jiang W, Zhang Y-J, Kahn SM, Hollstein MC, Santella RM,Lu S-H, Harris CC, Montesano R and Weinstein IB.(1993b). Proc. Natl. Acad. Sci. USA, 90, 9026 ± 9030.
Johnson DG. (1995). Oncogene, 11, 1685 ± 1692.Johnson DG, Ohtani K and Nevins JR. (1994). Genes & Dev.,8, 1514 ± 1525.
Kallakury BV, Figge J, Ross JS, Fisher HA, Figge HL andJennings TA. (1994). Hum. Pathol., 25, 92 ± 97.
Kato J-Y, Matsushime H, Hiebert SW, Ewen ME and SherrCJ. (1993). Genes & Dev., 7, 331 ± 342.
Kirshmeir PT, Housey GM, Johnson MD, Perkins AS andWeinstein IB. (1988). DNA, 7, 219 ± 225.
Lammie GA, Fant IV, Smith R, Schuuring E, Brookes S,Michalides R, Dickson C, Arnold A & Peters G. (1991).Oncogene, 6, 439 ± 644.
Lim DJ, Liu X-L, Sutkowski DM, Braun EJ, Lee C andKozlowski JM. (1993). Prostate, 22, 109 ± 118.
Lukas J, Jadayel D, Bartkova J, Nacheva E, Dyer MJ,Strauss M and Bartek J. (1994a).Oncogene, 9, 2159 ± 2167.
Lukas J, Pagano M, Staskova Z, Draetta G, Bartek J.(1994b). Oncogene, 9, 707 ± 718.
Matsushime H, Quelle DE, Shurtle� SA, Shibuya M, SherrCJ and Kato J-Y. (1994).Mol. Cell. Biol., 14, 2066 ± 2076.
Montgomery BT, Young CY, Bilhartz DL, Andrews PE,Prescott JL, Thompson NF and Tindall DJ. (1992).Prostate, 21, 63 ± 73.
Musgrove EA, Lee CSL, Buckley MF and Sutherland RL.(1994). Proc. Natl. Acad. Sci. USA, 91, 8022 ± 8026.
Navone NM, Troncoso P, Piters LL, Goodrow TL, PalmerJL, Nichols WW, von Eschenbach AC and Conti CJ.(1993). J. Natl. Cancer Inst., 85, 1657 ± 1669.
Nevins JR. (1992). Science, 258, 424 ± 429.Oswald F, Lovec H, MoÈ roÈ y T and Lipp M. (1994). Oncogene,9, 2029 ± 2036.
Pretlow TG, Delmoro CM, Dilley GG, Spadafora CG andPrelow TP. (1991). Cancer Res., 51, 3814 ± 3817.
Quelle DE, Ashmun RA, Shurtle� SA, Kato J-Y, Bar-SagiD, Roussel MF and Sherr CJ. (1993). Genes & Dev., 7,1559 ± 1571.
Resnitzky D, Gossen M, Bujard H and Reed S. (1994). Mol.Cell. Biol., 14, 1669 ± 1679.
Rosenberg CL, Kim HG, Shows TB, Kronenberg HM andArnold A. (1991). Oncogene, 6, 449 ± 453.
Ruizeveld de Winter JA, Jannssen PJ, Sleddens HM,Verleiun-Mooijman MC, Trapman J, Brinkmann AO,Santerse AB, Schroder FH and van der Kwast TH. (1994).Am. J. Pathol., 144, 735 ± 746.
Schulze A, Zerfa K, Spitkovsky D, Henglein B and Jansen-DuÈ rr P. (1994). Oncogene, 9, 3475 ± 3482.
Schuuring E, Verhoeven E, Mooi WJ and Michalides RJ.(1992). Oncogene, 7, 355 ± 361.
Shao A and Robbins PD. (1995). Oncogene, 10, 221 ± 228.Sherr CJ and Roberts JM. (1995). Genes & Dev., 9, 1149 ±1163.
Sherr CJ. (1993). Cell, 73, 1059 ± 1065.Sherr CJ. (1994). Cell, 79, 551 ± 555.Sonnenschein C, Olea N, Pasanen E and Soto AM. (1989).
Cancer Res., 49, 3474 ± 3481.Tam SW, Theodoras AM, Shay JW, Draetta GF and PaganoM. (1994). Oncogene, 9, 2663 ± 2674.
Tilly WD, Lim-Tio SS, Horsfall DJ, Aspinall JO, MarshallVR and Skinner JM. (1994). Cancer Res., 54, 4096 ± 4102.
Vairo G, Livingston DM and Ginsberg D. (1995). Genes &Dev., 9, 869 ± 881.
Veldscholte J, Ris-Stalpers C, Kuiper GG, Jenster G,Berrevoets C, Claassen E, van Rooij HC, Trapman J,Brinkman AO and Mulder E. (1990). Biochem. Biophys.Rev. Commun., 173, 534 ± 540.
Voeller HJ, Sugars LY, Pretlow T and Gelman EP. (1994). J.Urol., 151, 492 ± 495.
Weinberg RA. (1991). Science, 254, 1138 ± 1146.Wu H-C, Hsieh J-T, Gleave ME, Brown NM, Pathak S andChung LW. (1994). Int. J. Cancer, 57, 406 ± 412.
Xiong Y, Connolly T, Futcher B and Beach D. (1991). Cell,65, 691 ± 699.
Xiong Y, Menninger J, Beach D and Ward DC. (1992a).Genomics, 13, 575 ± 584.
Xiong Y, Zhang H and Beach D. (1992b). Cell, 71, 505 ± 514.Zhang Y-J, Jiang W, Chen CJ, Lee CS, Kahn SM, SanntwllaRM and Weinstein IB. (1993). Biochem. Biophys. Res.Commun., 196, 1010 ± 1016.
Zhou P, Jiang W, Zhang Y-J, Kahn SM, Schieren I, SantellaRM and Weinstein IB. (1995). Oncogene, 11, 571 ± 580.
Cyclin D1 overexpression in LNCaP cellsY Chen et al
1920
Copyright © 2022 FDOKUMEN




















