
Vitamin E inhibits CD36 scavenger receptor expression in hypercholesterolemic rabbits

In vivo effects of the controlled NO donor/scavenger
ruthenium cyclam complexes on blood pressure$
Fabiana G. Marcondesa, Alessander A. Ferrob, Adriana Souza-Torsonia,Marie Sumitania, Michael J. Clarkec, Douglas W. Francod,
Elia Tfounib,*, Marta H. Kriegera,*,1
aDepartamento de Fisiologia e Biofısica, Instituto de Biologia UNICAMP. Campus Universitario Zeferino Vaz,
13083-970, Barao Geraldo, Campinas-SP, BrazilbDepartamento de Quımica, Faculdade de Filosofia, Ciencias e Letras de Ribeirao Preto,
Universidade de Sao Paulo. Av. dos Bandeirantes, 3900. 14040-901- Ribeirao Preto, SP, BrazilcMerkert Chemistry Center, Boston College, Chestnut Hill, MA 02467, USA
dInstituto de Quımica de Sao Carlos Universidade de Sao Paulo. Av.Dr. Carlos Botelho,
1465 CEP 13560-970, Sao Carlos, SP, Brazil
Received 19 June 2001; accepted 6 December 2001
Abstract
Ruthenium(II/III) complexes able to bind and release NO. were tested in vivo, in conscious Wistar
rats instrumented for continuous blood pressure (BP) measurement and administration of in bolus
injections (5 to 100 nmol/Kg i.v.) of trans-[RuIICl(NO+)(cyclam)](PF6)2 (cyclam-NO) or sodium
nitroprusside (SNP). For normotensive rats, cyclam-NO produced a sustained 10% BP reduction of
basal MAP during 7 ± 0.4 to 11 ± 0.3 min. In acute hypertensive rats, cyclam-NO produced BP
reduction 3-fold larger than in normotensive rats and similar to that of SNP (maximal effect: 41 ± 1.3
vs. 45 ± 2.2 mmHg, respectively). However, the duration of the effect of cyclam-NO was 13 to 21-fold
longer than that of SNP. The hypotensive effect of cyclam-NO was fully blocked in presence of
continuous infusion of a NO. scavenger, carboxy-PTIO (6 mmol/Kg/min), or of the inhibitor of cGMP
0024-3205/02/$ – see front matter D 2002 Elsevier Science Inc. All rights reserved.
PII: S0024 -3205 (02 )01528 -X
$ Previous results of this work were presented at the First International Conference. Biology, Chemistry and
Therapeutic Applications of Nitric Oxide. San Francisco, California, USA, June 2000. Nitric Oxide: Biology and
Chemistry, 2000, 4, 181-182.
* Corresponding authors. Current Address: Departamento de Ciencias Exatas e Tecnologicas- UESC, Rodovia
Ilhues-Itabuna, Km 16, 45650-000, Ilhues/BA. Tel.: +55-16-602-3748, +55-16-602-3877; fax: +55-16-633-8151.
E-mail addresses: [email protected] (E. Tfouni), [email protected] (M.H. Krieger).1 Tel.: +55-19-3788-6189; fax: +55-192893124.
Life Sciences 70 (2002) 2735-2752

activation, methylene blue (83 nmol/Kg/min), or of the cyclam-NO precursor, trans-[RuCl(tfms)(cy-
clam)](tfms) (cyclam-tfms) (500 mmol/Kg/min). The long lasting BP reduction of cyclam-NO can be
interpreted in terms of a slower rate of NO. release (k-NO = 2.2 � 10�3 s�1 at 35 �C) followingchemical reduction (E00 = 0.10 V vs NHE). In summary, cyclam-NO showed an hypotensive effect
around 20 times longer than SNP in either normotensive or hypertensive rats, which was completely
inhibited by methylene blue or carboxy-PTIO. Continuous infusion of cyclam-tfms completely
blocked the hypotensive effect of cyclam-NO by scavenging the NO. released by the reduced cyclam-
NO. D 2002 Elsevier Science Inc. All rights reserved.
Keywords: NO donor; NO scavenger; Nitric oxide; Cyclam; Ruthenium nitrosyl; Hypotensive effect; Nitroprusside
Introduction
Nitric oxide (nitrogen monoxide, NO), a lipid-soluble and unstable gas [1], is one of the
most fascinating substances in biological chemistry. The NO molecule plays a significant role
in a variety of physiological and pathological processes in different cells and tissues, such as
neurotransmission, blood pressure (BP) control, inhibition of platelet aggregation and im-
munological responses [2,3]. Several of the biological actions of NO are attributed to its
stimulation of soluble guanylate cyclase that acts on GTP to produce cGMP, leading to
vascular relaxation [4]. Therefore, the NO-dependent relaxation of vascular smooth muscle
cell (VSMC) may be regulated locally. This may represent the simplest, essential mechanism
for adaptation of the cardiovascular system in maintaining normal blood pressure [5]. The
importance of the balance between the release, uptake, and degradation of NO for the main-
tenance of normal blood pressure is evident i) in that administration of the NO synthase
inhibitors, such as L-NAME (NG-nitro-L-arginine methyl ester), markedly increase blood
pressure in animals [5,6]; and ii) when occurs decrease of NO bioavailability in the vascular
function resulting from oxidative inactivation by excessive production of the superoxide
anion [7,8].
While the importance of investigating drugs, which act as NO donors is well recognized
[9,10], its unstable nature and short half-life, 0.1 to 6 seconds [11], present challenges from a
pharmacological perspective. Furthermore, the mechanism and extent of dissociation from
NO donors is not fully understood, and such compounds are sometimes unstable with respect
to temperature and light [12]. As a consequence, there is considerable interest in kinetically
and thermodynamically stable compounds that can, with minimal toxic effects, release or
scavenge NO in vivo.
A compound that acts as a NO carrier and could be activated to release nitric oxide in a
controlled manner, in the order of minutes, would be of great relevance. The vasodilating
effects of nitrovasodilators, such as organic nitrites (e.g. nitroglycerine, NTG) and
inorganic sodium nitroprusside (SNP) are mediated by generation of NO [13]. Although
SNP and NTG are widely used in the treatment of various cardiovascular disorders and
F.G. Marcondes et al. / Life Sciences 70 (2002) 2735–27522736

reducing blood pressure during anesthesia, there are limitations to their use, and both
classes of compounds have undesirable side effects [13,14]. Continuous exposure to ni-
troglycerine leads to the development of nitrate resistance and nitrate tolerance. Tolerance
to NTG is a clinically important problem that requires periodic discontinuation to restore
effectiveness [14]. High doses, prolonged use, or use of SNP in patients with compromised
hepatic function, are associated with the accumulation of cyanide and its metabolite thio-
cyanate [13].
Several new metal complexes have been studied recently as NO donors or scavengers,
including nitrosyl ruthenium complexes, nitrite complexes, and related species [15-39],
some of which exhibit low toxicity and are quite robust in aqueous solutions even in the
presence of O2. Coordinated NO, in the nitrosonium (NO+) form, may be released, in vitro,
after activation through irradiation by light at a specific wavelength [25,38-41] or a selective
chemical reduction [42] of the nitrosonium site to the nitrosyl (NO.) form. Recently [33], the
complex trans-[RuCl(NO)(cyclam)](PF6)2 (cyclam-NO) (cyclam = 1,4,8,11-tetraazocyclote-
tradecane) has been isolated and its reactivity studied. The slow loss of NO (k-NO = 6.1 �10�4s�1, at 25 �C, in vitro) from the cyclam-NO complex [33] following a biologically
accessible one-electron reduction (E0 = �0.1 V vs NHE [33]), led us to investigate the
possible hypotensive effect of this complex in biological systems. The cyclam-NO or
SNP complexes were administered in the presence and absence of the NO scavenger
carboxy-PTIO [43], or methylene blue [44], an inhibitor of the synthesis of c-GMP,
and trans-[RuCl(tfms)(cyclam)](tfms) (cyclam-tfms). Since the cyclam-NO complex can be
synthesized by reacting cyclam-tfms with NO [33], the latter complex was tested as a
potential NO scavenger. The biological results are interpreted in light of the chemistry of
these complexes.
Materials and methods
In conducting this research the authors adhered to NIH Animal Care guidelines for use of
animals. All surgical procedures were performed under anesthesia produced by ketamine
(40 mg/Kg i.p., Ketalar, Parke Davis) in conjunction with xylazine (6mg/Kg i.m., Rompum,
Bayer). Adult male Wistar rats (250-300 g) were supplied by Centro de Bioterismo da
Universidade Estadual de Campinas CEMIB-UNICAMP. Cyclam-NO and cyclam-tfms com-
plexes were synthesized as described [33,40].
Arterial pressure and heart rate
Polyethylene cannulae (PE10 connected to PE50) filled with heparinized saline were
implanted into the abdominal aorta (through the femoral artery) for recording blood
pressure and into the iliac vein for administration of drugs or vehicle (pH 7.4 PBS, 0.1 M
phosphate buffer, 0.15 M saline). After catheterization, the rats received a single dose
(40000 U) of penicillin G procaine and were kept in individual home cages under a 12 h
light/dark cycle, receiving standard rat chow and tap water ad libitum. To complete animal
F.G. Marcondes et al. / Life Sciences 70 (2002) 2735–2752 2737

recovery, the drug administration section was performed 24 hours after the catheterization.
Blood pressure was continuously recorded in conscious freely moving unrestrained rats
using a pressure transducer (Gould) and an amplifier attached to the intra-arterial cannula,
and the measurements were acquired in a computer for posterior analysis (DI220 AT-
CODAS data acquisition system, DataQ Instruments Co.). The systolic, diastolic, and
mean arterial pressures (MAP), and the heart rate (HR, peak-to-peak counting) in the basal
condition and during drugs administration were calculated using the Advanced CODAS
software, from which the maximum DMAPs were obtained. The total duration of the
effect refers to the elapsed time from the initial of the BP decrease up to total basal
pressure recovery.
Drug administration
Since SNP is light sensitive, appropriate dilutions of the drug in cold phosphate buffered
saline were prepared immediately before use, protected from light, to prevent its photolysis.
This procedure was standardized to the other complexes, cyclam-NO and cyclam-tfms,
although they are photochemically inert [40], and stable [33,40]. Before the intravenous
administration of cyclam-NO, a control record of blood pressure during approximately
20 minutes was performed until stabilization of pressure and heart rate. Blood pressure
variations after in bolus injection of SNP (MW = 298, Sigma Chemical), cyclam-NO (MW =
802) or cyclam-tfms (MW = 634) were determined in 31 normotensive rats. Equimolar doses
of cyclam-NO, cyclam-tfms and SNP were used. The doses of SNP were chosen following
similar experiments described in the literature [13]. The ranges of doses (5, 10, 25, 30 or
100 nmol/Kg i.v.) of each drug were injected in a randomized sequence protocol. Each rat
received 6-10 injections of PBS or PBS containing SNP, or cyclam-NO, or cyclam-tfms, each
injected in a volume of 0.1 ml. Intervals between administrations of different doses were made
after complete recovery of basal values of blood pressure (3 to 5 min for SNP and � 30 min
for cyclam-NO and cyclam-tfms).
Acute hypertension model
The hypotensive effect (DMAP) of cyclam-NO (10 nmol/Kg i.v. in bolus) was tested in
the presence of continuous infusion with phenylephrine (or the PBS vehicle) made by a
pump (Cole Parmer model HP74900-25). The infusion of 0.11 mg/Kg/min phenylephrine
(L-phenylephrine hydrochloride, Sigma Chemical)) was performed over 1 hour to obtain an
approximately 30% increase in basal blood pressure.
NO scavenger infusion
In another group of 7 rats, the effect of in bolus injection of 5 nmol/Kg of cyclam-NO was
tested during infusion of 0.6 or 6 mmol/Kg/min of carboxy-PTIO ([2-(4-carboxyphenyl)-
4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide], MW 277, Calbiochem). Other 6 rats were
tested in the presence of 83 nmol/Kg/min of methylene blue (C16H18ClN3S, MW 320, Sigma
F.G. Marcondes et al. / Life Sciences 70 (2002) 2735–27522738

Chemical) and, finally, in another group of 5 rats, a 5 nmol/Kg in bolus injection of cyclam-
NO was tested during infusion of 50, 250 or 500 nmol/Kg/min of cyclam-tfms.
Evaluation of the toxicity of cyclam-NO
The cytotoxicity of cyclam-NO was evaluated according to standardized methods. The
assays were carried out on a lung fibroblast cell line derived from Chinese hamsters (V-79)
established in culture. Periodic plating was performed as soon as the cells reached the
standardized confluence density. DMEM essential medium was used for culture, sup-
plemented with 10% bovine fetal serum (BFS), 100 IU/ml penicillin and 100 mg/ml
streptomycin sulfate. The material was incubated at 37 �C in a humid atmosphere containing
5% CO2 in a Forma Scientific incubator. Plating was performed with 3�104 viable cells/ml/
well on a 96-well plate. After incubation at 37 �C for 48 h in DMEM supplemented with
10% serum, the culture was exposed for 24 h to different cyclam-NO concentrations in the
0.1-3 mM concentration range. The nucleic acid content (NAC) was used for the direct
evaluation of the number of viable cells. The numbers of cells in the control and the
complexes samples were estimated from the total NAC according to the method of Cingi
et al. [45] The metabolic activity of these cells was evaluated by a colorimetric method
based on the reduction of the tetrazolium salt MTT, according to the method of Mosmann
et al. [46].
Statistical analysis
All values are expressed as mean ± S.E.M. The MAP values obtained before and after drug
administration were compared by using the STAT SOFT Inc. software Statistic version 5,
using the one way ANOVA followed by the Duncan test. Differences were considered
statistically significant when P < 0.05.
Results
Cytotoxicity assay
Fig. 1 shows the V79 cell viability after treatment with cyclam-NO evaluated by two
different assays, MTT and NAC. These tests show that cyclam-NO is particularly non-toxic,
especially when compared with SNP (IC50 = 0.06 mM, Fig. 1, panel B). The IC50 value for
cyclam-NO was not achieved experimentally up to 3.0 mM (Fig. 1, panel A).
Hypotensive effect in conscious normotensive rats
Fig. 2, panel A, shows the effect of intravenous in bolus injections of different
concentrations of cyclam-NO, SNP or cyclam-tfms on mean arterial pressure (MAP) of
conscious normotensive rats. The vasodilator effect upon administration of cyclam-NO
F.G. Marcondes et al. / Life Sciences 70 (2002) 2735–2752 2739

(5.0, 25.0, 30.0, 50.0 and 100 nmol/Kg) on MAP varied from �7 ± 0.4 to �11 ± 0.3 mmHg,
which correspond to an average of ~ 8% decrease of the basal MAP. The highest MAP
reduction, which was produced at 30.0 nmol/Kg, was �11 ± 0.3 mmHg (P < 0.05). When
the doses administered were higher than 30.0 nmol/Kg, the magnitude of the MAP
response did not increase further (Fig. 2, panel A). The hypotensive effect of cyclam-
NO was significantly lower than that produced by equimolar doses of SNP (P < 0.01). The
BP reductions by SNP varied from �12.6 ± 0.8 to �36 ± 1.6 mmHg, and correlated with
dose (P < 0.05). In bolus administration, cyclam-tfms did not result in any significant MAP
variation (Fig. 2, panel A). Fig. 2, panel B, shows the total time duration of the effect on
MAP, produced by the above compounds. The duration of the effect produced by different
doses of SNP varied from 0.5 ± 0.04 to 0.8 ± 0.1 min, almost without dose correlation,
Fig. 1. MTT (dashed lines) and NAC (solid lines) cell viability tests in cultures of fibroblast-like V79 cells. The
percentage of living cells in relations to the control after incubation for 48 h at 37 �C, are plotted versus the
complex concentration (SNP (panel A) or cyclam-NO (panel B)). Data reported as Mean ± S.E.M. of 6 assays.
F.G. Marcondes et al. / Life Sciences 70 (2002) 2735–27522740

whereas those produced by cyclam-NO varied from 6.4 ± 1.4 to 10.6 ± 2.4 min (Fig. 2, panel
B). This duration of cyclam-NO effect was, on average, 13-fold longer than the effect of
SNP, reaching a maximum of 21-fold at 25.0 nmol/Kg (P < 0.05). The latency (time to reach
maximal BP decrease) of the effect for SNP varied from 0.12 ± 0.02 to 0.18 ± 0.03 min,
whereas for cyclam-NO varied from 0.62 ± 0.10 to 1.3 ± 0.18 min.
Hypotensive effect in conscious hypertensive rats
Fig. 3, panel A, shows the BP reduction produced by in bolus injection of different
concentrations of cyclam-NO or SNP in acute hypertensive rats under continuous
infusion of phenylephrine (0.11 mg/Kg/min) and in normotensive rats under continuous
PBS infusion. There was no significant effect of PBS on the BP of normotensive rats.
The phenylephrine infusion resulted in a BP increase from 107 ± 2.7 mmHg to 160 ±
Fig. 2. Effect of intravenous in bolus injections of cyclam-NO, SNP or cyclam-tfms on MAP of conscious
normotensive rats. Panel A: effect of different concentrations of the complexes on MAP. Panel B: duration of the
effect of different complexes concentrations. Data reported as Mean ± S.E.M. of 16 rats.
F.G. Marcondes et al. / Life Sciences 70 (2002) 2735–2752 2741
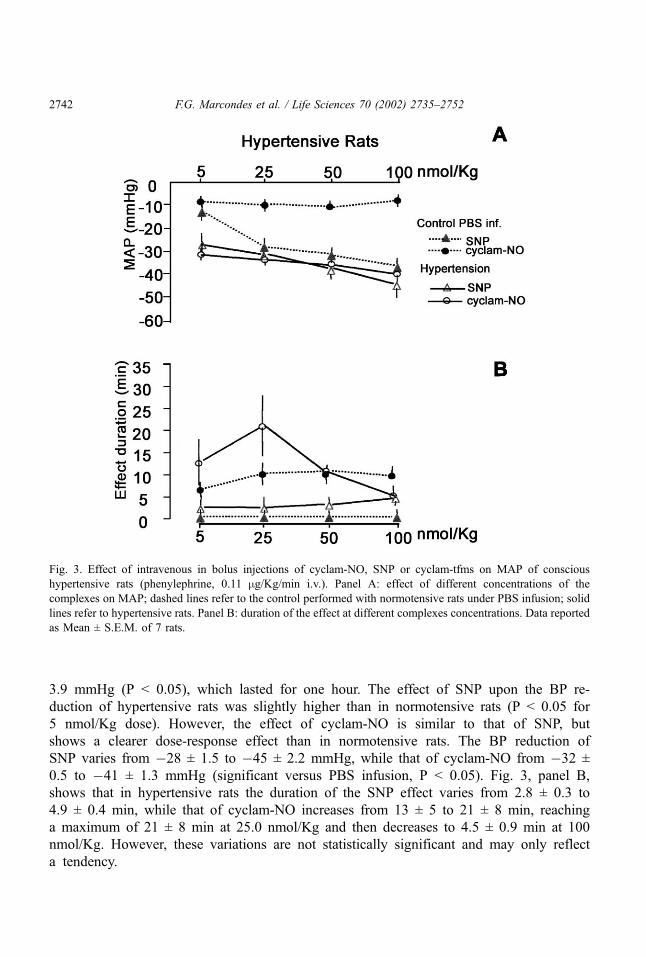
3.9 mmHg (P < 0.05), which lasted for one hour. The effect of SNP upon the BP re-
duction of hypertensive rats was slightly higher than in normotensive rats (P < 0.05 for
5 nmol/Kg dose). However, the effect of cyclam-NO is similar to that of SNP, but
shows a clearer dose-response effect than in normotensive rats. The BP reduction of
SNP varies from �28 ± 1.5 to �45 ± 2.2 mmHg, while that of cyclam-NO from �32 ±
0.5 to �41 ± 1.3 mmHg (significant versus PBS infusion, P < 0.05). Fig. 3, panel B,
shows that in hypertensive rats the duration of the SNP effect varies from 2.8 ± 0.3 to
4.9 ± 0.4 min, while that of cyclam-NO increases from 13 ± 5 to 21 ± 8 min, reaching
a maximum of 21 ± 8 min at 25.0 nmol/Kg and then decreases to 4.5 ± 0.9 min at 100
nmol/Kg. However, these variations are not statistically significant and may only reflect
a tendency.
Fig. 3. Effect of intravenous in bolus injections of cyclam-NO, SNP or cyclam-tfms on MAP of conscious
hypertensive rats (phenylephrine, 0.11 mg/Kg/min i.v.). Panel A: effect of different concentrations of the
complexes on MAP; dashed lines refer to the control performed with normotensive rats under PBS infusion; solid
lines refer to hypertensive rats. Panel B: duration of the effect at different complexes concentrations. Data reported
as Mean ± S.E.M. of 7 rats.
F.G. Marcondes et al. / Life Sciences 70 (2002) 2735–27522742

Hypotensive effect in presence of a scavenger or GMPc blockage in normotensive
conscious rats
Fig. 4 shows the BP reduction of in bolus administration of cyclam-NO or SNP under
infusion of PBS, 83 nmol/Kg/min methylene blue, 6 � 102 or 6 � 103 nmol/Kg/min carboxy-
PTIO. Compared to the effect in PBS, the hypotensive effect of SNP was decreased 16 ± 5%,
18 ± 4% and 23 ± 4 % under continuous infusion of methylene blue, and 6 � 102 and 6 �103 nmol/Kg/min carboxy-PTIO, respectively. Notably, a larger decrease in effect is observed
for cyclam-NO. Compared to the effect in PBS, the hypotensive effect of cyclam-NO
decreased 47 ± 7 % and 80 ± 11 % for 6 � 102 and 6 � 103 nmol/Kg/min carboxy-PTIO,
respectively. Methylene blue completely blocks the BP reduction of cyclam-NO.
Hypotensive effect of cyclam-NO in the presence of cyclam-tfms in normotensive
conscious rats
Fig. 5 shows the BP reduction of in bolus administration of 5 nmol/Kg cyclam-NO under
continuous infusion of PBS, 50, 250 or 500 nmol/Kg/min of cyclam-tfms in normotensive
rats. The cyclam-tfms infusion per se (1 ml/h) does not change the BP of normotensive rats.
A marked decrease of the hypotensive effect of cyclam-NO occurs with increasing
Fig. 4. Influence of methylene blue endovenous infusion (83 nmol/Kg/min i.v.) and of carboxy-PTIO infusion (6
� 102 or 6 � 103 nmol/Kg/min i.v.) on the hypotensive effect produced by 5 nmol/Kg i.v. in bolus injection of
cyclam-NO or SNP in normotensive conscious rats. Data reported as Mean ± S.E.M. of 7 rats for methylene blue
and 6 rats for carboxy-PTIO (P < 0.05 compared to PBS group).
F.G. Marcondes et al. / Life Sciences 70 (2002) 2735–2752 2743

concentration of cyclam-tfms. Compared to PBS infusion, the hypotensive effect was
reduced to 41 ± 12 % and 66 ± 14 % with 50 and 250 nmol/Kg/min cyclam-tfms, respec-
tively. Remarkably, the hypotensive effect of cyclam-NO was practically completely blocked
(97 ± 12 %) by a concentration of cyclam-tfms (500 nmol/Kg/min) a 100 times higher than
that of cyclam-NO.
Discussion
The results described in this paper refer to the BP reduction effect of cyclam-NO on
normotensive and hypertensive rats and can be interpreted at least partly on the chemical
properties of the complexes involved [33,40], as shown below.
Hypotensive effects in normotensive and hypertensive rats
Cyclam-NO is water soluble, quite stable, and in solution ionizes to yield trans-
[RuIICl(NO+)(cyclam)]2+, which can be reduced to trans-[RuIICl(cyclam)(NO.)]+ (Eq. 1):
trans�½RuIIClðNO2
ÞðcyclamÞ�2þ þ e� ��!k1 trans�½RuIIClðcyclamÞðNO*Þ�þ ð1Þ
Fig. 5. Influence of PBS, 50, 250 or 500 nmol/Kg/min cyclam-tfms endovenous infusion on the hypotensive effect
produced by 5 nmol/Kg i.v. in bolus injection of cyclam-NO in normotensive conscious rats. Data reported as
Mean ± S.E.M. of 8 rats.(P < 0.05 for comparison versus PBS group).
F.G. Marcondes et al. / Life Sciences 70 (2002) 2735–27522744

The formation of trans-[RuIICl(cyclam)(NO.)]+ is followed by the rapid loss of chloride by
first-order kinetics [33], with k = 1.5 s�1, forming trans-[RuII(H2O)(cyclam)(NO.)]+ (Eq. 2):
trans�½RuIIClðcyclamÞðNO*Þ�þ þ H2O
!k2 trans�½RuIIðH2 OÞðcyclamÞðNO*Þ�2þ þ Cl� ð2Þ
This chloride loss is suppressed by 0.1 M chloride, and both, trans-[RuII(H2O)(cyclam)
(NO.)]+ and trans-[RuIICl(cyclam)(NO.)]+ release NO. (Eqs. 3 and 4) with specific rate
constants of 6.2 � 10�4 s�1 at 25 �C, and 2.2 � 10�3 s�1 at 35 �C, which are independent of
chloride concentration up to 0.25 M [33,40]:
trans�½RuIIðH2OÞðcyclamÞðNO*Þ�2þ þ H2O
!k3 trans�½RuIIðH2OÞ2ðcyclamÞ�2þ þ NO* ð3Þ
trans�½RuIIClðcyclamÞðNO*Þ�þ þ H2O
! trans�½RuIIClðH2OÞðcyclamÞ�þ þ NO* ð4Þ
This slow rate of release allowed the observation of the EPR spectra of the coordinated NO.
in trans-[RuII(H2O)(cyclam)(NO.)]+ [34].
The reduction potential of trans-[RuIICl(NO+)(cyclam)]2+, of -0.10 V vs NHE [33] is
accessible to glutathione [47], flavins, and other single electron reductants that may ultimately
be reduced by NADH (�0.32 V vs NHE) [48]. Considering that in blood, chloride loss is
suppressed, then, the hypotensive effects observed in conscious rats are consistent with the
release of NO. from trans-[RuIICl(cyclam)(NO.)]+, following the reduction of trans-
[RuIICl(NO+)(cyclam)]2+.
The hypotensive effect of cyclam-NO is smaller in normotensive rats (Fig. 2), when
compared to the effect produced by SNP. For hypertensive rats, the effects of cyclam-NO and
SNP are similar (Fig. 3). However, for both, normotensive and hypertensive rats, the duration
of the effects of cyclam-NO are longer than those of SNP. This difference is consistent with the
differences in the rates of release of NO. between SNP and cyclam-NO. The specific rate
constant for the release of NO. from SNP through reduction activation is not accessible in the
literature. However, it is well known that this rate is fast, and it is consistent with effecting a BP
decrease similar to that of trans-[RuII(NO+)(NH3)4(P(OEt)3)]3+ [49,50], which releases NO.
three to four orders of magnitude faster (k = 0.97 s�1 at 25 �C and 5.0 s�1 at 37 �C [49,50])
than cyclam-NO (2.2 � 10�3 s�1 at 35 �C [40]). Both, SNP and trans-[RuII(NO+)(NH3)4(P(OEt)3)]
3+are reported to show a similarly short-lived BP reduction [49,50]. Fig. 2 shows the
effect of SNP on BP compared to that of cyclam-NO. The slower rate of release of NO. from
trans-[RuIICl(cyclam)(NO.)]+, compared with these other two complexes, is consistent with a
longer duration of the hypotensive effect of cyclam-NO. It should be noted that the longer
time-course of the hypotensive effect of the cyclam-NO complex might overcome some
possible regulatory blood pressure mechanisms, such as the baroreflex [6].
F.G. Marcondes et al. / Life Sciences 70 (2002) 2735–2752 2745

While chromium and molybdenum nitrosyl complexes have been reported to be efficient
potent vasodilators in vivo and in vitro, with actions compared to that of SNP [15], no
comparisons with cyclam-NO could be made due to the different animal models assayed in
each case. For the chromium and molybdenum complexes anesthetized rats were used,
whereas for cyclam-NO, the rats were conscious. Earlier reports indicated that some Ru
nitrosyl complexes, such as [Ru(NO)Cl5]2� and [Ru(NO)(NH3)5]
3+ relax vascular smooth
muscle, but none were as effective as SNP in relaxing aortic strips and in decreasing
arterial blood pressure in anesthetized cats [51]. The authors also report toxic ip LD50s for
these complexes.
Hypotensive effects in the presence of methylene blue and carboxy-PTIO
Continuous infusion for 30 minutes of methylene blue (83 nmol/Kg/min) alone does not
alter the MAP values of normotensive rats [50], whereas that of carboxy-PTIO (6 � 103
nmol/Kg/min) raises the MAP 15 % [50]. In the presence of methylene blue, a soluble
guanylyl cyclase inhibitor [44,52], the vasodilator effect of SNP is only partly blocked, as
expected [44], whereas that of the cyclam-NO complex is fully blocked, consistent with
NO. release and involvement of the NO/cGMP pathway for in vivo vasorelaxation.
However, methylene blue may act not only by inhibition of the activation of guanylate
cyclase [52], but also through other pathways, such as extra cellular generation of
superoxide anion [53]. Nevertheless, a similar, but less pronounced effect is observed when
carboxy-PTIO, which traps NO. [43], is present during the administration of cyclam-NO.
For SNP, the effect of carboxy-PTIO is only slightly more pronounced than with methylene
blue. For both complexes the effect is larger for greater concentrations of carboxy-PTIO,
although it should be noted that for cyclam-NO the effect of carboxy-PTIO is not as large as
that of methylene blue. This difference could possibly be larger if we take into account that
carboxy-PTIO alone increases the MAP 15 %. Direct comparison of effects is difficult,
since different mechanisms are operative for each drug. Nevertheless, these results are
consistent with carboxy-PTIO acting as a NO. scavenger (the rate constant of reaction of
carboxy-PTIO with NO. is �104 M�1 s�1) [43], and imply NO. release and involvement of
the NO/cGMP pathway for in vivo vasorelaxation. These results strongly suggest that
cyclam-NO delivers NO. in vivo following reduction, and that its hypotensive effect is
primarily dependent on cGMP, in contrast to the other NO-dependent metabolic action of
SNP [54].
Hypotensive effects in the presence of cyclam-tfms
The cyclam-tfms complex is water soluble and undergoes fast aquation in water resulting
in trans-[RuIIICl(H2O)(cyclam)]2+ (Eq. 5).
trans�½RuIIIClðtfmsÞðcyclamÞ�ðtfmsÞ
��!H2O
Hþ trans�½RuIIIClðH2OÞðcyclamÞ�2þ þ 2 tfms ð5Þ
F.G. Marcondes et al. / Life Sciences 70 (2002) 2735–27522746

This last complex, trans-[RuIIICl(H2O)(cyclam)]2+, has a pKa of 3.3 [55], and in water and at
physiological pH is in the hydroxo form, trans-[RuIIICl(OH)(cyclam)]+ (Eq. 6).
trans�½RuIIIClðH2OÞðcyclamÞ�2þ] trans�½RuIIIClðOHÞðcyclamÞ�þ þ Hþ ð6Þ
Then, trans-[RuIIICl(OH)(cyclam)]+ reacts with NO. to form trans-[RuIICl(NO+)(cyclam)]2+
(Eq. 7):
trans� ½RuIIIClðOHÞðcyclamÞ�þ þ NO*
! trans�½RuIIClðNOþÞðcyclamÞ�2þ þ OH� ð7Þ
In fact, the synthesis of the cyclam-NO complex from cyclam-tfms follows Eqs. 4-6 [33,40],
and, thus, cyclam-tfms is the precursor of cyclam-NO. Thus, the blockage of the MAP
decrease effected by the NO. released from the reduction of cyclam-NO, in the presence of
cyclam-tfms (500 times excess) is consistent with cyclam-tfms readily resulting in a NO.
scavenger, trans-[RuIIICl(OH)(cyclam)]+ (Eqs. 5-7). It can be noticed that, for a fixed
cyclam-NO concentration, increasing concentrations of the tfms complex scavenge the
released NO., thereby attenuating the BP lowering effect, and fully blocking the effect with a
500 times excess of cyclam-tfms. In addition, this is also strong evidence that cyclam-NO
decreases the MAP through release of NO.. Other Ru complexes had been proposed and
reported to behave as NO. scavengers, especially polyaminocarboxylates [16-18,27], of
which, [Ru(Hedta)Cl] and [Ru(Hedta)(H2O)] appeared to be efficient NO. scavengers,
resulting in the respective nitrosyl complex [Ru(NO)(Hedta)]+. However, it should be
mentioned that the specific rate constant for the release of NO., after reduction in the edta
complex is 7.3 � 10�3 s�1 at 25 �C [56], one order of magnitude faster than that for the
cyclam-NO complex (6.1 � 10�4 s�1) [33]. Furthermore, it should be noted that the
concentration of cyclam-tfms (500 nmol/Kg/min) to fully block the MAP reduction effect of
cyclam-NO is approximately one order of magnitude smaller than the most efficient
concentration of carboxy-PTIO (6 � 103 nmol/Kg/min).
The cyclam complexes system
After reduction of the cyclam-NO complex and the subsequent reactions (Eqs. 1-4),
trans-[RuIICl(H2O)(cyclam)]+, as the predominant species in blood, and trans-[RuII(H2O)2(-
cyclam)]2+ are formed. These complexes are easily oxidized (E vs NHE = �0.050 V for the
chloro complex [33], and +0.155 V at pH 1 [57], +0.132 V at pH 4.32 [33,40], and ��0.2 V
at pH 7 for the diaqua [55]) by dissolved oxygen (E = +0.815 V at pH 7) or some other
available and accessible oxidizing species present in biological systems, to trans-[RuIIICl
(H2O)(cyclam)]2+ and trans-[RuIII(H2O)2(cyclam)]3+ (pKa = 1.1 [58]), (Eq. 8),
trans�½RuIIðH2OÞ2ðcyclamÞ�2þ ! trans�½RuIIIðH2OÞ2ðcyclamÞ�3þ þ e� ð8Þ
F.G. Marcondes et al. / Life Sciences 70 (2002) 2735–2752 2747

which at pH � 7 are in the form of trans-[RuIIICl(OH)(cyclam)]+ and trans-[RuIII(OH)
(H2O)(cyclam)]2+ (Eqs. 6 and 9)
trans�½RuIIIðH2OÞ2ðcyclamÞ�3þ] trans�½RuIIIðOHÞðH2OÞðcyclamÞ�2þ þ Hþ ð9Þ
trans-[RuIIICl(OH)(cyclam)]2+ reacts with NO. to form trans-[RuIICl(NO+)(cyclam)]2+
(Eq. 7), and it is reasonable to expect that trans-[RuIII(OH)(H2O)(cyclam)]2+, being similar
to trans-[RuIIICl(H2O)(cyclam)]2+ can react with NO. in a similar manner (Eq. 10),
resulting in trans-[RuII(OH)(NO+)(cyclam)]2+ (pKa of trans-[RuII(NO+)(H2O)(cyclam)]2+ is
3.0 [33,40].
trans� ½RuIIIðOHÞðH OÞðcyclamÞ�2þ þ NO*
! trans� ½RuIIðOHÞðNOþÞðcyclamÞ�2þ þ H2O ð10Þ
In addition to reducing species, oxidizing species are also present in biological systems,
and, thus, conceivably the formed trans-[RuIICl(H2O)(cyclam)]+ and trans-[RuII(H2O)2(cyclam)]2+, resulting from the cyclam-NO administration, can be oxidized to yield the
NO. scavengers trans-[RuIIICl(OH)(cyclam)]+ and trans-[RuIII(OH)(H2O)(cyclam)]2+.
Therefore, since the NO. is released slowly from the reduced cyclam-NO, the resulting
trans-[RuIIICl(OH)(cyclam)]+ and trans-[RuIII(OH)(H2O)(cyclam)]2+ complexes scavenge
the NO., forming trans-[RuIICl(NO+)(cyclam)]2+ and trans-[RuII(OH)(NO+)(cyclam)]2+
(Eqs. 7 and 10) and, thus, attenuating the BP decrease. The NO. scavenging action is
dependent on the trans-[RuIIICl(OH)(cyclam)]+, trans-[RuIII(OH)(H2O)(cyclam)]2+ and NO.
concentrations, and, thus, is more important at higher concentrations. Two factors at least are
involved in these concentrations. They should increase with increasing concentrations of
cyclam-NO and with time due to the slow release of NO. and formation of trans-
[RuIII(OH)(H2O)(cyclam)]2+. There is no full blockage of the hypotensive effect because
the scavenger concentration does not reach the 500 times of cyclam-NO concentration,
needed to fully block the effect.
However, this is not straightforward, as it may seem. Other chemical considerations may
also be involved, and which may contribute to a lack of a dose-response in normotensive rats,
as other chemical equilibria occur that may interact with biological regulatory mechanisms.
For example, while dissociation is suppressed at blood chloride levels, trans-[RuII(OH)
(NO+)(cyclam)]2+ may form inside cells. As this has a low reduction potential (�0.46 V vs
NHE at pH 7 [33,40]) it would be not biologically relevant. Consequently the various
possible ruthenium nitrosyl species may interact differently with available biological
reductants in different compartments. The net result would be dependent on the rate
constants, the nature and concentration of the species involved, and the possible different
biological regulatory mechanisms acting in normotensive and hypertensive rats.
The behavior of the above described cyclam complexes may have important consequences,
since at least conceivably, the pair of complexes cyclam-NO and cyclam-tfms may result in a
NO. buffering system, or lead to the design of one such system.
F.G. Marcondes et al. / Life Sciences 70 (2002) 2735–27522748

Conclusion
Cyclam-NO administration results in BP reduction, especially in hypertensive rats, with a
hypotensive effect 20 times more prolonged than SNP either in normotensive or hypertensive
rats. The hypotensive effect of cyclam-NO is totally inhibited by the unspecific GMPc
inhibitor methylene blue and also by c-PTIO. The continuous infusion of cyclam-tfms is able
to fully block the hypotensive effect of cyclam-NO by scavenging the NO. released from this
last complex after reduction. Finally, the above-mentioned chemical properties of cyclam-
NO, strongly suggest that the long term BP reduction, with cyclam-NO, comes from the slow
release of NO. from this complex after its reduction in vivo. The results are also consistent
with the NO. scavenging by trans-[RuIII(OH)Cl(cyclam)]+, formed from cyclam-tfms. In
addition, the possible attenuation of the BP reduction, from the NO. scavenging by trans-
[RuIII(OH)(H2O)(cyclam)]2+, possibly formed in the subsequent in vivo reactions of cyclam-
NO, would be beneficial in limiting higher concentrations of NO.. The above results may
play an important role in biological applications. The long-term action of cyclam-NO, which
makes it a sort of controlled NO. donor, can be important, under some pathophysiological
conditions, where high concentrations of NO. are harmful, whereas lower and lasting
concentrations are beneficial.
Acknowledgments
The authors thank CAPES, FAPESP, CNPq, FAEP-UNICAMP, NIH GM26390, for grants
and fellowships.
References
1. Ignarro LJ, editor. Nitric Oxide: Biology and Pathology. Orlando: Academic Press, 2000.
2. Whittle BJR. Nitric oxide in physiology and pathology. Hystochem J 1995;27:727-37.
3. Moncada S, Higgs A. The L-arginine: nitric oxide pathway. New Engl J Med 1993;329:2002-12.
4. Rees DD, Palmer RMJ, Moncada S. The role of endothelium-derived nitric oxide in the regulation of blood
pressure. Proc Natl Acad Sci USA 1989;86:3375-78.
5. Moncada S, Palmer RMJ, Higgs A. Nitric oxide: physiology, pathophysiology and pharmacology. Pharmacol
Rev 1991;43:109-42.
6. Umans JG, Levi R. The nitric oxide system in circulatory homeostasis and its possible role in hypertensive
disorders. In: Laragh JH, Brenner BM, editors. Hypertension: Pathology, Diagnosis and Management. 2nd
edn. New York: Raven Press, 1995. pp. 1083-95.
7. Paolocci N, Biondi R, Bettini M, Lee CI, Berlowitz CO, Rossi R, Xia Y, Ambrosio G, Labbate A, Kass DA,
Zweier JL. Oxygen radical-mediated reduction in basal and agonist-evoked NO release in isolated rat heart. J
Mol Cell Cardiol 2001;33(4):671-9.
8. Nakamura T, Igarashi R, Kurashima T, Saito Y, Hoshino J, Sumino H, Sakamoto H, Nagai R. Lecithinized
superoxide dismutase induces vasodilation; evidence of direct contribution of superoxide anions to modulat-
ing vascular tone. Life Sci 1999;64:65-70.
F.G. Marcondes et al. / Life Sciences 70 (2002) 2735–2752 2749

9. Clarke MJ, Gaul JB. Chemistry relevant to the biological effects of nitric oxide and metallonitrosyls. In:
Structure and Bonding, vol. 81. New York: Springer-Verlag, 1993. pp. 147-80.
10. Wink DA, Mitchell JB. Chemical biology of nitric oxide: insights into regulatory, cytotoxic and cytoprotective
mechanisms of nitric oxide. Free Radic Biol Med 1998;25:434-56.
11. Kelm M, Feelisch M, Spahr R, Piper H-M, Noarck E, Schrader J. Quantitative and kinetic characterization of
nitric oxide and EDRF released from cultured endothelial cells. Biochem Biophis Res Commun 1988;
154:236-44.
12. Wink DA, Ford PC. Nitric oxide reactions important to biological systems: a survey of some kinetics in-
vestigations. Methods: A Companion to Methods in Enzymology, vol. 7. San Diego: Academic Press, 1995.
pp. 14-20.
13. Arnold WP, Longnecker DE, Epstein RM. Photodegradation of sodium nitroprussiate: biologic activity and
cyanide release. Anesthesiology 1984;61:254-60.
14. Laursen JB, Mulsch A, Boesgaard S, Mordvintcev P, Trautner S, Gruhn N, Nielsen-Kudsk JE, Busse R,
Aldershvile J. In vivo nitrate tolerance is not associated with reduced bioconversion of nitroglycerin to nitric
oxide. Circulation 1996;94:2241-7.
15. Wang YX, Legzdins P, Poon JS, Pang CY. Vasodilator effects of organotransition-metal nitrosyl complexes,
novel nitric oxide donors. J Cardiovasc Pharmac 2000;35:73-7.
16. Fricker SP. Ruthenium, nitric oxide and disease. A novel inorganic chemistry approach to drug design.
Platinum Metals Rev 1995;39:150-9.
17. Fricker SP, Slade E, Powell NA, Vaughan OJ, Henderson GR, Murrer BA, Megson IL, Bisland SK, Flitney
FW. Ruthenium complexes as nitric oxide scavengers: a potential therapeutic approaches to nitric oxide-
mediated diseases. Br J Pharmacol 1997;122:1441-9.
18. Davies NA, Wilson MT, Slade E, Fricker SP, Murrer BA, Powel NA, Henderrson GR. Kinetics of nitric oxide
scavenging by ruthenium(III) polyaminocarboxylates: novel therapeutic agents for septic shock. Chem Com-
mun 1997;(1):47-8.
19. DeLeo MA, Ford PC. Photoreactions of coordinated nitrite ion. Reversible nitric oxide labilization from the
chromium(III) complex trans-[Cr(cyclam)(ONO)2]+. Coord Chem Rev 2000;208:47-59.
20. Bourassa JL, Ford PC. Flash and continuous photolysis studies of Roussin’s red salt dianion Fe2S2(NO)42� in
solution. Coord Chem Rev 2000;200:887-900.
21. Works CF, Ford PC. Photoreactivity of the ruthenium nitrosyl complex, Ru(salen)(Cl)(NO). Solvent effects on
the back reaction of NO with the Lewis acid Ru-III(salen)(Cl). J Am Chem Soc 2000;122:7592-3.
22. DeLeo MA, Bu XH, Bentow J, Ford PC. The synthesis, characterization and structures of the chromium(III)
dinitrito complexes: trans-[Cr(L)(ONO)2]+(L = 1,4,8,11-tetraazacyclotetradecane or 5,7,7,12,14,14-hexameth-
yl-1,4,8,11-tetraazacyclotetradecane). Inorg Chim Acta 2000;300:944-50.
23. Bourassa J, Lee B, Bernhard S, Schoonover J, Ford PC. Flash photolysis studies of Roussin’s black salt
anion: Fe4S3(NO)7�. Inorg Chem 1999;38:5926.
24. Hoshino M, Laverman L, Ford PC. Nitric oxide complexes of metalloporphyrins: an overview of some
mechanistic studies. Coord Chem Rev 1999;187:75-102.
25. Ford PC, Bourassa J, Miranda K, Lee B, Lorkovic I, Boggs S, Kudo S, Laverman L. Photochemistry of metal
nitrosyl complexes. Delivery of nitric oxide to biological targets. Coord Chem Rev 1998;171:185-202.
26. Laverman LE, Ford PC. Reactions of water soluble iron(II) and cobalt(II) porphyrins with nitric oxide. Im-
plications for the reactivity of NO and biologically relevant metal centers. Chem Commun 1999;18:1843-4.
27. Chen YA, Shepherd RE. [RuII(ttha)(H2O)2]2� is a rapid NO scavenger (ttha6� = triethylenetetraaminehexa-
acetate. J Inorg Biochem 1997;68:183-93.
28. Chen YA, Lin FT, Shepherd RE. 15N NMR and electrochemical studies of [RuII(Hhedta)]� complexes of NO,
NO+, NO2� and NO�. Inorg Chem 1999;38:973-83.
29. Ward MS, Shepherd RE. A reversible NO complex of [FeII(edampda)] and the [NiII(edampda)] analogue
(edampda = N,N0-bis(pyridylmethylenediamina-N,N0-diacetate). Inorg Chim Acta 1999;286:197-206.
30. Chen Y, Sweetland MA, Shepherd RE. A reversible NO complex of Fe(II)-(TIM): An S = 1/2 (FeNO)7
nitrosyl. Inorg Chim Acta 1997;260:163-72.
F.G. Marcondes et al. / Life Sciences 70 (2002) 2735–27522750

31. Shepherd RE, Sweetland MA, Junker DE. Ligand field factors in promoting S = 3/2 (FeNO)7 nitrosyls. J Inorg
Biochem 1997;65:1-14.
32. Bezerra CWB, Silva SC, Gambardella MTP, Santos RHA, Plicas LMA, Tfouni E, Franco DW. Water p-donation in trans-tetraammineruthenium(II): effect on coordinated water properties induced by NO trans
ligand. Inorg Chem 1999;38:5660-7.
33. Lang D, Davis JA, Lopes LGF, Ferro AA, Vasconcellos LCG, Prock A, Franco DW, Tfouni E, Wieraszko A,
Clarke MJ. A controlled NO releasing compound: Synthesis, molecular structure, spectroscopy, electrochem-
istry, and chemical reactivity of (R,R,S,S) trans-[RuCl(NO)(cyclam)]2+ (cyclam = 1,4,8,11-tetraazacyclote-
tradecane). Inorg Chem 2000;39:2394-400.
34. McGarvey BR, Ferro AA, Tfouni E, Bezerra CWB, Bagatin I, Franco DW. Detection of the EPR spectrum of
NO in ruthenium(II) complexes. Inorg Chem 2000;39:3577-81.
35. Gorelsky SI, Silva SC, Lever ABP, Franco DW. Electronic spectra of trans-[Ru(NH3)4L(NO)]3+/2+ com-
plexes. Inorg Chim Acta 2000;300:698-708.
36. Da Silva SC, Franco DW. Metastable excited states and electronic structure of [Ru(NH3)5(NO)]3+ and
[Ru(NH3)4(OH)(NO)]2+. Spectrochim Acta A 1999;55:1515-25.
37. Lopes LGF, Gomes MG, Borges SSS, Franco DW. Correlation between the lever parameter and electronic
properties of nitrosyl ruthenium(II) complexes. Austr J Chem 1998;51:865-6.
38. Borges SDS, Davanzo CU, Castellano EE, Z-Schpector J, Silva SC, Franco DW. Ruthenium nitrosyl com-
plexes with N-heterocyclic ligands. Inorg Chem 1998;37:2670-7.
39. Gomes MG, Davanzo CU, Silva SC, Lopes LGF, Santos PS, Franco DW. Cis- and trans-nitrosyltetraammi-
neruthenium(II). Spectral and electrochemical properties and reactivity. J Chem Soc Dalton 1998;601-7.
40. AA Ferro, Sıntese, Caracterizacao e Reatividade Quımica e Fotoquımica de Aminas de Rutenio (trans-
[Ru(NO)(NH3)4L](BF4)3 (L = pz, 4-acpy, imN ou L-hist) e trans-[RuCl(NO)(cyclam)](PF6)2 (cyclam =
1,4,8,11-tetraazaciclotetradecano)). D.Sc.Thesis, Departamento de Quımica, Faculdade de Filosofia, Ciencias
e Letras de Ribeirao Preto, Universidade de Sao Paulo, Ribeirao Preto, SP, Brazil, 2000.
41. Tfouni E. Photochemical reactions of ammineruthenium(II) complexes. Coord Chem Rev 2000;196:281-305.
42. Callahan RW, Meyer TJ. Reversible electron-transfer in ruthenium nitrosyl complexes. Inorg Chem 1977;
16:574-81.
43. Akaike T, Yoshida M, Miyamoto Y, Sato K, Kohno M, Sasamoto K, Miyazaki K, Ueda S, Maeda H.
Antagonist action of imidazolineoxyl N-oxides against endothelium-derived relaxing factor/NO through a
radical reaction. Biochem 1993;32:827-32.
44. Mayer B, Brunner F, Schmidt F. Inhibition of the nitric oxide synthesis by methylene blue. Biochem Phar-
macol 1993;45:367-74.
45. Cingi MR, De Angelis I, Fortunati E, Reggiani E, Bianchi V, Tiozzo R, Zucco F. Choice standardization of
test protocols in cytotoxicology: a multicentre approach. Toxicology In vitro 1991;5:119-25.
46. Mosmann T. Rapid colorimetric assay for cellular grownth and survival: application to proliferation and
citotoxicity assays. J Immun Methods 1983;65:55-63.
47. Frasca DR, Clarke MJ. Alterations in the binding of [Cl(NH3)5RuIII]2+ to DNA by glutathione: reduction,
autoxidation, coordination, and decomposition. J Am Chem Soc 1999;121:8523-32.
48. Clarke MJ, Bitler S, Rennert D, Buchbinder M, Kelman AD. Reduction and Subsequent Binding of Ruthe-
nium Ions Catalyzed by Subcellular Components. J Inorg Biochem 1980;12:79-87.
49. Franco DW, Tfouni E, Krieger MH, Toledo JC, Barros BF. First International Conference. Biology, chemistry,
and therapeutic applications of nitric oxide. San Francisco, CA, 2000, in Nitric Oxide. Biology and Chemistry
2000;4:254.
50. Torsoni AS, Barros BF, Toledo Jr. JC, Haun M, Krieger MH, Tfouni E, Franco DW, Hypotensive properties
and acute toxicity of trans-[Ru(NH3)4P(OEt)3(NO)](PF6)3. A new nitric oxide donor. Nitric Oxide, in press.
51. Kruszyna H, Kruszyna R, Hurst J, Smith RP. Toxicology and pharmacology of some ruthenium compounds:
Vascular smooth muscle relaxation by nitrosyl derivatives of ruthenium and iridium. J Toxic Envir Health
1980;6:757-73.
52. Gruetter CA, Gruetter DY, Lyon JE, Kadowitz JP, Ignarro LJ. Relationship between cyclic guanosine
F.G. Marcondes et al. / Life Sciences 70 (2002) 2735–2752 2751

3:5-monophososphate. Formation and relaxtion of coronary arterial smooth muscle by glyceryl trinitrate,
nitropruuside, nitrite and nitric oxide: effects of methylene blue and methemoglobin. J Pharm Exp Therap
1981;2199:181-6.
53. Wolin MS, Cherry PD, Rodenburg JM, Messina EJ, Kaley G. Methylene blue inhibits vasodilation of skeletal
muscle arterioles to acethylcholine and nitric oxide via the extracellular generation of superoxide anion. J
Pharm Exp Ther 1990;254:872-6.
54. Kowaluk EA, Seth P, Fung H-L. Metabolic activation of sodium nitroprussitee to nitric oxide in vascular
smooth muscle. J Pharm Exp Ther 1992;262:916-22.
55. Ferreira KQ, Tfouni E, unpublished results.
56. Guidorzi APB, Lopes LGF, Franco DW. Reatividade do ligante NO em [RuHedtaNO]2+/+. 20a Reuniao
Annual da SBQ, Pocos de Caldas MG, 1997. Livro de Resumos, vol. 1, Sao Paulo, Brazil, abstract QI-121.
57. Walker DD, Taube H. N-Macrocyclic complexes of ruthenium(II) and ruthenium(III). Inorg Chem 1981;
20:2828-34.
58. Walker DD. Saturated tetraaza macrocyclic ligand complexes of ruthenium(II) and ruthenium(III). PhD
Thesis. Stanford University, Stanford, 1980.
F.G. Marcondes et al. / Life Sciences 70 (2002) 2735–27522752
Copyright © 2022 FDOKUMEN




















