
In vitro osteoblastic differentiation of human mesenchymal stem cells and human dental pulp stem...
15
In vitro osteoblastic differentiation of human bone marrow cells in the presence of metal ions S. Morais, 1 N. Dias, 1 J. P. Sousa, 1 M. H. Fernandes, 2 G. S. Carvalho 3 1 INEB-Instituto de Engenharia Biome ´dica, Departamento de Engenharia Qui ´mica, Faculdade de Engenharia da Universidade do Porto, Prac ¸a do Coronel Pacheco, 1, 4050 Porto, Portugal 2 Faculdade de Medicina Denta ´ria da Universidade do Porto, Rua Dr Manuel Pereira da Silva, 4200 Porto, Portugal 3 IEC, Universidade do Minho, Largo do Pac ¸o, 4719 Braga codex, Portugal Received 30 April 1997; accepted 27 July 1998 Abstract: For periods up to 21 days human bone marrow was cultured in control conditions that favor the prolifera- tion and differentiation of osteoblastic cells. The effect of AISI 316L corrosion products and the corresponding major separate metal ions (Fe, Cr, and Ni) were studied in three different phases of the culture period in order to investigate the effects of metal ions in cell populations representative of osteoblastic cells in different stages of differentiation. Tox- icity consequences of the presence of metal ions in bone marrow cultures were evaluated by biochemical parameters (enzymatic reduction of MTT, alkaline phosphatase activity, and total protein content), histochemical assays (identifica- tion of ALP-positive cells and Ca and phosphates deposits), and observation of the cultures by light and scanning elec- tron microscopy. Culture media were analyzed for total and ionized Ca and P and also for metal ions (Fe, Cr, and Ni). The presence of AISI 316L corrosion products and Ni salt in bone marrow cultures during the first and second weeks of culture significantly disturbs the normal behavior of these cultures, interfering in the lag phase and exponential phase of cell growth and ALP expression. However, the presence of these species during the third week of culture, when ex- pression of osteoblastic functions occurs (mineralization process), did not result in any detectable effect. Fe salt also disturbs the behavior of bone marrow cell cultures when present during the lag phase and proliferation phase, and a somewhat compromised response between the normal pat- tern (control cultures) and intense inhibition (AISI 316L cor- rosion products and Ni salt-added cultures) was observed. Fe did not affect the progression of the mineralization phase. Osteogenic cultures exposed to Cr salt (Cr 3+ ) presented a pattern similar to the controls, indicating that this element does not interfere, in the concentration studied, in the osteo- blastic differentiation of bone marrow cells. Quantification of metal ions in the culture media showed that Cr (originat- ed from AISI 316L corrosion products but from not Cr 3+ salt) and Ni (originated from AISI 316L corrosion products and Ni salt) appear to be retained by the bone marrow cultures. © 1999 John Wiley & Sons, Inc. J Biomed Mater Res, 44, 176–190, 1999. Key words: human osteoblast-like cells; differentiation; metal ions INTRODUCTION The principal applications of AISI 316L stainless steel in the biomedical field are bone and joint replace- ment, spinal instrumentation, and fracture fixation. 1 In the vast majority of implant applications, AISI 316L performs its intended function (due to superior corro- sion resistance and good mechanical properties 1 ) without any important adverse physiological effects, and, consequently, it has achieved a large degree of success. 1 However, clinical experience 2 has revealed that in the presence of a severe and prolonged corro- sion process provoked by the aggressive human body environment, inhibition of normal metabolism, 3,4 toxic reactions, 5,6 and alteration of tissue synthesis and structure can occur. Furthermore, although all the ma- jor alloying elements of AISI 316L (Fe, Cr, and Ni) are essential in the human diet, when their levels increase beyond physiological limits, the precise biochemical balance is disturbed, with consequent toxicity. 7 Bone repair and remodeling are carried out by os- teoblasts and osteoblasts. Resorption of old bone is accomplished by osteoclasts, and the subsequent for- mation of new bone is done by osteoblasts. 8 These two processes are highly coupled to each other and are No benefit of any kind will be received either directly or indirectly by the authors. Correspondence to: M.H. Fernandes Contract grant sponsor: JNICT; Contract grant number; PBIC/C/CTM/1954/95 Contract grant sponsor: European Commission; Contract grant number: Brite Euram project BE 7928 © 1999 John Wiley & Sons, Inc. CCC 0021-9304/99/020176-15
Transcript of In vitro osteoblastic differentiation of human mesenchymal stem cells and human dental pulp stem...

In vitro osteoblastic differentiation of human bone marrowcells in the presence of metal ions
S. Morais,1 N. Dias,1 J. P. Sousa,1 M. H. Fernandes,2 G. S. Carvalho3
1INEB-Instituto de Engenharia Biomedica, Departamento de Engenharia Quimica, Faculdade de Engenharia daUniversidade do Porto, Praca do Coronel Pacheco, 1, 4050 Porto, Portugal2Faculdade de Medicina Dentaria da Universidade do Porto, Rua Dr Manuel Pereira da Silva, 4200 Porto, Portugal3IEC, Universidade do Minho, Largo do Paco, 4719 Braga codex, Portugal
Received 30 April 1997; accepted 27 July 1998
Abstract: For periods up to 21 days human bone marrowwas cultured in control conditions that favor the prolifera-tion and differentiation of osteoblastic cells. The effect ofAISI 316L corrosion products and the corresponding majorseparate metal ions (Fe, Cr, and Ni) were studied in threedifferent phases of the culture period in order to investigatethe effects of metal ions in cell populations representative ofosteoblastic cells in different stages of differentiation. Tox-icity consequences of the presence of metal ions in bonemarrow cultures were evaluated by biochemical parameters(enzymatic reduction of MTT, alkaline phosphatase activity,and total protein content), histochemical assays (identifica-tion of ALP-positive cells and Ca and phosphates deposits),and observation of the cultures by light and scanning elec-tron microscopy. Culture media were analyzed for total andionized Ca and P and also for metal ions (Fe, Cr, and Ni).The presence of AISI 316L corrosion products and Ni salt inbone marrow cultures during the first and second weeks ofculture significantly disturbs the normal behavior of thesecultures, interfering in the lag phase and exponential phaseof cell growth and ALP expression. However, the presenceof these species during the third week of culture, when ex-
pression of osteoblastic functions occurs (mineralizationprocess), did not result in any detectable effect. Fe salt alsodisturbs the behavior of bone marrow cell cultures whenpresent during the lag phase and proliferation phase, and asomewhat compromised response between the normal pat-tern (control cultures) and intense inhibition (AISI 316L cor-rosion products and Ni salt-added cultures) was observed.Fe did not affect the progression of the mineralization phase.Osteogenic cultures exposed to Cr salt (Cr3+) presented apattern similar to the controls, indicating that this elementdoes not interfere, in the concentration studied, in the osteo-blastic differentiation of bone marrow cells. Quantificationof metal ions in the culture media showed that Cr (originat-ed from AISI 316L corrosion products but from not Cr3+ salt)and Ni (originated from AISI 316L corrosion products andNi salt) appear to be retained by the bone marrow cultures.© 1999 John Wiley & Sons, Inc. J Biomed Mater Res, 44,176–190, 1999.
Key words: human osteoblast-like cells; differentiation;metal ions
INTRODUCTION
The principal applications of AISI 316L stainlesssteel in the biomedical field are bone and joint replace-ment, spinal instrumentation, and fracture fixation.1
In the vast majority of implant applications, AISI 316Lperforms its intended function (due to superior corro-sion resistance and good mechanical properties1)
without any important adverse physiological effects,and, consequently, it has achieved a large degree ofsuccess.1 However, clinical experience2 has revealedthat in the presence of a severe and prolonged corro-sion process provoked by the aggressive human bodyenvironment, inhibition of normal metabolism,3,4 toxicreactions,5,6 and alteration of tissue synthesis andstructure can occur. Furthermore, although all the ma-jor alloying elements of AISI 316L (Fe, Cr, and Ni) areessential in the human diet, when their levels increasebeyond physiological limits, the precise biochemicalbalance is disturbed, with consequent toxicity.7
Bone repair and remodeling are carried out by os-teoblasts and osteoblasts. Resorption of old bone isaccomplished by osteoclasts, and the subsequent for-mation of new bone is done by osteoblasts.8 These twoprocesses are highly coupled to each other and are
No benefit of any kind will be received either directly orindirectly by the authors.
Correspondence to: M.H. FernandesContract grant sponsor: JNICT; Contract grant number;
PBIC/C/CTM/1954/95Contract grant sponsor: European Commission; Contract
grant number: Brite Euram project BE 7928
© 1999 John Wiley & Sons, Inc. CCC 0021-9304/99/020176-15

responsible for the renewal of the skeleton whilemaintaining its anatomic and structural integrity.9 Un-der normal physiologic circumstances, bone repairand remodeling proceed in highly regulated cycles.9
However, the presence of corrosion products may af-fect the systemic and local factors that regulate osteo-blastic development and their function of promotingosteolysis surrounding metal implants. Also, oneneeds to bear in mind that osteoblasts occur in a con-tinuum of many different stages of differentiation andthat functional behavior and also response to changesin the local conditions (i.e., presence of drugs) dependon the stage of differentiation in the osteoblastic lin-eage.10,11
In this study, human osteoblastic cells derived frombone marrow were cultured in control conditions forperiods up to 21 days and the effect of metal ionsstudied in three different phases of the culture period.Metal-treated bone marrow cells were cultured for 7days (in the presence of metal ions), 14 days (exposedto metal ions during the second week of culture), and21 days (exposed to metal ions during the third weekof culture). All cells, except those used as controls,were exposed for seven days to AISI 316L corrosionproducts and to the corresponding separate majormetal ions in the same concentration in differentphases of the culture period in order to investigate theeffects of metal ions in cell populations representativeof osteoblastic cells in different stages of differentia-tion. The toxicity consequences of the presence ofmetal ions in human bone marrow cultures wereevaluated by biochemical parameters [enzymatic re-duction of MTT, alkaline phosphatase (ALP) activity,and total protein content], histochemical assays (iden-tification of ALP-positive cells and Ca and phosphatesdeposits), and observation of the cultures by light mi-croscopy and scanning electron microscopy (SEM)coupled with X-ray microanalysis. The culture mediawere analyzed for total and ionized Ca and P and alsofor metal ions (Fe, Cr, and Ni). Ca and metal contentwere determined using a selective electrode12 and amercury microelectrode,13–16 respectively, showingthat electrochemical methods provide a very inexpen-sive and reliable alternative to the traditional methodof atomic absorption spectrometry (AAS), combininga nondestructive character with high sensitivity andaccuracy.
MATERIALS AND METHODS
Metallic slurries
Type AISI 316L stainless steel (Fe 63.9%, Cr 18.0%, Ni12.5%, Mo 2.8%, Si 1.2%, Mn 1.6%, and C 0.025%, weight forweight) was anodically dissolved in Hank’s balanced salt
solution (HBSS), which simulates the composition of physi-ological fluids. The resulting concentrations of the majormetal ions in the stock AISI 316L slurry were determined byatomic absorption spectrometry (AAS): 1.02 × 10−2 mol/LFe, 2.67 × 10−3 mol/L Cr, and 1.95 × 10−3 mol/L Ni (5.2Fe:1.4 Cr:1.0 Ni). Salt solutions containing 8.95 × 10−3 mol/LFe (FeCl3?6H2O), 2.35 × 10−3 mol/L Cr (CrCl3?6H2O), or 1.72× 10−3 mol/L Ni (NiNO3) were prepared separately inHBSS.
Cell culture
Osteogenic cells were obtained from the bone marrow ofhealthy 35 year old man who was undergoing orthopediccorrective surgery. Bone marrow was cultured in a-MEMsupplemented with 10% fetal bovine serum, 2.5 mg/mL fun-gizone, and 50 mg/mL gentamicin, 10−8 mol L−1 dexameth-asone, 2.52 × 10−4 mol L−1 ascorbic acid, and 10−2 mol L−1
b-glycerophosphate17 (control medium). Human bone mar-row cells were cultured at 37°C in a humidified atmospherecontaining 5% CO2 and 95% air. Medium containing non-adherent cells was removed 24 h later, and passages weredone after tripsinization of adherent cells in exponentialgrowth (7 days after the beginning of the incubation). Cellsof the second passage were seeded at a concentration of 104
cells/cm2 in: (a) 35 mm diameter tissue culture dishes forhistochemical assays and SEM observation, (b) 96-well cul-ture plates for cell viability MTT assay, ALP activity assay,and total protein content, and (c) 60 mm in diameter tissueculture dishes for Ca, P and metal quantification in the cul-ture medium.
Bone marrow cells were cultured for periods up to 21 daysin control conditions and in the presence of AISI 316L cor-rosion products (SS) or their corresponding separate metalions. Metal-treated cultures were exposed to (SS) −8.95 ×10−6 mol/L Fe + 2.35 × 10−6 mol/L Cr + 1.72 × 10−6 mol/LNi (obtained by dilution of the stock 316L slurry) and Fe, Cr,and Ni salt solutions, which were added separately to otherculture dishes at equivalent concentrations, that is, 8.95 ×10−6 mol/L, 2.35 × 10−6 mol/L, and 1.72 × 10−6 mol/L Ni,respectively. The effects of metal ions were studied in dif-ferent stages of the culture period. Cells were cultured for 7,14, and 21 days in four experimental situations. In the firstassay, cells were cultured for 7 days and metallic elementswere added 24 h after tripsinization (to allow for cell adhe-sion; day 0); cells were tested at day 0 (before metallic ex-position) and at 1 and 7 days later (after metallic exposition).In the second assay, cells were cultured for 14 days. Thesecells were grown in control medium during the first 6 daysand then exposed to metallic species between days 7 and 14.Cells were tested at days 0 and 7 (before metallic exposition)and at day 14 (after metallic exposition). In the third assay,cells were cultured for 21 days. These cells were grown incontrol medium during the first 13 days and were exposedto the corrosion products and metal salts between days 14and 21, cells were tested at days 0, 7, 14 (before introductionof slurries) and 21 (after metallic exposition). The commonfeature of the three assays is that cells were exposed to me-tallic slurries (metal-treated cultures) or control medium
177IN VITRO OSTEOBLASTIC DIFFERENTIATION

(cultures used as controls) during 7 days without renewal ofmedium. Finally, in the last assay, cells were fed with controlmedium for 21 days and tested at days 0, 7, 14, and 21. In allexperiments, the medium was changed twice a week exceptduring the incubation periods in which the effects of themetallic species were studied. Also, every time the mediumwas changed, its content in total and ionized Ca and P, Fe,Cr, and Ni was determined. For control quantification ex-periments, 60 mm tissue culture dishes containing culturemedium with and without corrosion products and metalions added were incubated, treated, and analyzed as werethe cell culture plates (in the different experimental situa-tions studied).
Biochemical assays
MTT assay
MTT assay18 [reduction of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide to a purple formazanproduct] was used to estimate cell viability and prolifera-tion. Cells (8 wells for each situation) were incubated with0.5 mg/mL of MTT in the last 4 h of the culture periodtested. The media were then decanted, formazan salts weredissolved with 100 mL of dimethylsulphoxide, and the ab-sorbance was determined at 600 nm in a ELISA reader.
Alkaline phosphatase activity
ALP19 was assayed based on the hydrolysis of p-nitrophenylphosphate (used as substrate) into p-nitrophenolat 37°C, pH 10.3. After removal of culture medium, the tis-sue was washed twice with PBS (phosphate-buffered solu-tion), and 100 mL of deionized and distilled water with 0.1%Triton X-100 was added to each well. Then 80 mL of sub-strate (a 25 mmol L−1 solution) were added to 100 mL of eachsample and the mixture was incubated at 37°C for 30 min.The reaction was stopped by the addition of 20 mL of NaOH5 mol L−1. Optical density was determined at 405 nm on anELISA reader and compared with the values of a serie ofp-nitrophenol standards.
Histological methods
Light microscopy
The presence of ALP-positive cells and phosphate or Cadeposits were visualized by histochemical staining. The cul-tures were fixed with 1.5% glutaraldehyde in 0.14 mol L−1
sodium cacodylate buffer and rinsed with distilled water.
ALP staining
Fixed cultures were incubated for 1 h in the dark with amixture, prepared in Tris buffer, pH 10, containing 2 mg/
mL of Na-a-naphtyl phosphate (reacting substrate) and 2mg/mL of fast blue RR salt; the incubation was stopped byrinsing the samples with water. ALP-positive cells stainedbrown.
Phosphate staining
Phosphate deposits were assessed by the von Kossa tech-nique,20 that is, the fixed cultures were covered with a 1%silver nitrate solution and kept for 1 h under UV light. Afterrinsing, a 5% sodium thiosulphate solution was added for 2min and cultures were washed again. Phosphate depositsstained black.
Calcium staining
The fixed cultures were covered with a 1% S alizarin so-dium sulfonate solution (0.028% in NH4OH), pH 6.4, for 2min and then rinsed with water and acid ethanol (ethanol,0.01% HCl).20 Calcium deposits stained red.
SEM and X-ray microanalysis
Cultures were fixed as for light microscopy and dehy-drated in a graded ethanol series. Critical point drying of thesamples was followed by carbon sputtering. The prepara-tions were examined in a JEOL JSM-35C microscope coupledwith TRATOR TN-2000 system at 15 keV.
Ion quantification
Ionized and total calcium
The free and total Ca contents in cell culture media wereevaluated by potentiometric means with the procedure de-scribed elsewhere.12 The potential was monitored using acalcium-selective electrode and an Ag/AgCl reference elec-trode, and the values of potential were recorded when thereadings stabilized within ± 0.10 mV for 5 min. Calibrationcurves were prepared with standards of a composition simi-lar to the culture medium twofold and tenfold diluted.These dilutions were made because it is under these condi-tions that free (a twofold dilution) and total (a tenfold dilu-tion) Ca was quantified in osteoblast-like cell culture me-dium. The ionic strength of the standard Ca solutions as wellas of the samples was adjusted and maintained (during thedilution of the samples) at ∼0.17 mol/L, which correspondsto the value of the ionic strength of the culture medium, inorder to keep the equilibrium of the system unchangeable.12
Ionized and total phosphorus
The inorganic (Pi) and total P were determined colorimet-rically using the Fiske and Subbarow procedure.21 For Pi
178 MORAIS ET AL.

quantification, the culture medium was treated with trichlo-roacetic acid to precipitate protein and lipid-bound phos-phates. The supernatant fluid was reacted with ammoniummolybdate in an acid solution to form phosphomolybdate. Amixture of sodium bisulfite, sodium sulfite, and 1-amino-2-naphtol-4-sulfonic acid then was added to reduce the phos-phomolybdate to form a phosphomolybdenum blue com-plex. The intensity of the color was measured at 660 nm in aShimadzu UV-VIS spectrometer. For total P quantification,samples of cell culture medium first were digested in a mi-crowave oven (CEM Model MDS-2000) with nitric acid (su-prapur 65%) in order to destroy organic matter. Then thesolutions obtained were treated as for Pi quantification (ob-viously the trichloroacetic acid step was not performed). Allvalues were calculated from standard curves read at thesame wave length.
Iron, chromium, and nickel
The quantification of these elements was performed byelectrochemical means, after digestion of medium samples,using a mercury film microelectrode, an Ag/AgCl referenceelectrode, and a cylindrical glassy carbon as counter elec-trode.13–16 The technique applied, adsorptive strippingvoltammetry, was based on measurement of the current as-sociated with the reduction of the metal–ligand complex ad-sorbed at the surface of the prepared mercury film micro-electrode. For Fe, Cr, and Ni analysis in osteoblast-like cellculture medium13–16 the organic ligand selected and opti-mized values for detection limit, pH, preconcentration po-tential, and time were, respectively, catechol, detection limit1.93 × 10−8 mol/L, ∼8.0, −1.80 V, and 30 s for Fe; diethylene-triaminepenta-acetic acid, detection limit 2.80 × 10−10 mol/L,∼5.9, −1.00 V, and 10 s for Cr; and dimethylglioxime, detec-tion limit 7.70 × 10−9 mol/L, ∼9.2, −0.70 V, and 10 s for Ni.
For biochemical data each point represents the mean ±standard deviation of at least six replicates, and for Ca andP and metal ions quantification, data presented correspondto the mean ± standard deviation of at least three indepen-dent analyses, each being an individual value obtained byperforming three replicates. The differences observed be-tween groups were considered significant for p values lowerthan 0.05 when analyzed statistically using analysis of vari-ance (ANOVA).
RESULTS
Human bone marrow stromal cells were grown inconditions known to favor the formation of osteoblast-rich cell cultures17,22–26 and in the presence of SS (8.95× 10−6 mol/L Fe + 2.35 × 10−6 mol/L Cr + 1.72 × 10−6
mol/L Ni), Fe salt (8.95 × 10−6 mol/L), Cr salt (2.35 ×10−6 mol/L) and Ni salt (1.72 × 10−6 mol/L). The aimof this work was to investigate the influence of metalelements, commonly present in orthopedic implants,in the osteoblastic differentiation of human bone mar-row cells. For this purpose, osteoblast-like cells were
cultured for periods up to 21 days in control condi-tions and exposed to metal ions in different phases ofthe culture period (i.e., during the first, second, andthird week of incubation). In another experiment,bone cells were cultured in control conditions for 21days. Control and metal treated cultures were evalu-ated by biochemical and histochemical assays and ob-served by SEM. Culture media were analyzed for totaland ionized Ca and P, and also Fe, Cr and Ni ions.
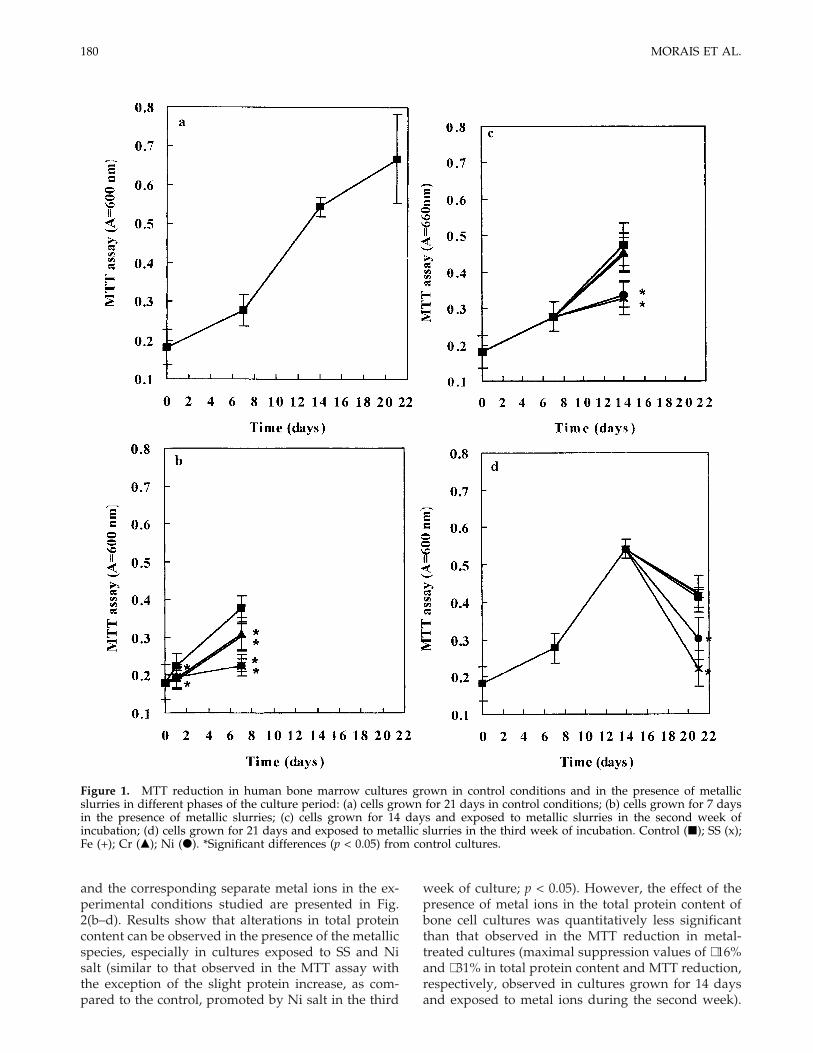
MTT reduction
The MTT test, where tetrazolium salts are trans-formed by active enzymes into intracellular formazandeposits, measures cell activation; only viable cells un-damaged by biomaterials or any other irritant can ex-press this activity.27
Proliferation of cells grown in control conditions for21 days [Fig. 1(a)] increased smoothly in the firstweek, suggesting that during this period the cells wereadapting to the culture conditions and that some ofthem probably died (lag phase). After that, cell growthwas exponential, and a tendency for a stationaryphase was observed by the third week of culture.
Figure 1(b) shows results concerning the reductionof MTT by human bone marrow cells grown for 7 daysin control conditions and exposed to metal ions. Asignificant decrease (p < 0.05) in cell proliferation wasobserved in metal-treated cultures, especially evidentin the presence of SS and Ni ions, suggesting that thepresence of these species disturbs the initial stages ofthe culture, namely the adaptation of bone cells toculture conditions. In cultures grown for 14 days andexposed to metal ions during the second week of cul-ture [Fig. 1(c)], SS and Ni ions cause a deleteriouseffect in cell viability/proliferation (p < 0.05), as mea-sured by the MTT assay, relative to cells grown incontrol conditions during the 14 days of incubation.These metal species also promoted a decrease in MTTreduction (p < 0.05) in cultures grown for 21 days andexposed to metal ions during the third week of incu-bation [Fig. 1(d)]. Fe and Cr ions interfered with thenormal growth of bone cells in culture only whenadded in the first week of incubation.
Total protein content
In cultures grown in control conditions for 21 days,total protein increased with the incubation time in away similar to cell proliferation, remaining approxi-mately constant during the third week of culture [Fig.2(a)]. Total protein content in cultures grown for 7, 14,and 21 days in control conditions and exposed to SS
179IN VITRO OSTEOBLASTIC DIFFERENTIATION
and the corresponding separate metal ions in the ex-perimental conditions studied are presented in Fig.2(b–d). Results show that alterations in total proteincontent can be observed in the presence of the metallicspecies, especially in cultures exposed to SS and Nisalt (similar to that observed in the MTT assay withthe exception of the slight protein increase, as com-pared to the control, promoted by Ni salt in the third
week of culture; p < 0.05). However, the effect of thepresence of metal ions in the total protein content ofbone cell cultures was quantitatively less significantthan that observed in the MTT reduction in metal-treated cultures (maximal suppression values of ∼16%and ∼31% in total protein content and MTT reduction,respectively, observed in cultures grown for 14 daysand exposed to metal ions during the second week).
Figure 1. MTT reduction in human bone marrow cultures grown in control conditions and in the presence of metallicslurries in different phases of the culture period: (a) cells grown for 21 days in control conditions; (b) cells grown for 7 daysin the presence of metallic slurries; (c) cells grown for 14 days and exposed to metallic slurries in the second week ofincubation; (d) cells grown for 21 days and exposed to metallic slurries in the third week of incubation. Control (j); SS (x);Fe (+); Cr (m); Ni (d). *Significant differences (p < 0.05) from control cultures.
180 MORAIS ET AL.

Although these two parameters (MTT reduction andtotal protein content) are a measure of cell prolifera-tion, the information provided is somewhat different.Protein measured in these cultures includes intracel-lular and extracellular protein, and the latter contrib-utes significantly to total protein content in bone cellcultures. This probably explains the less pronouncedeffect of metal ions on total protein content in cellcultures.
Alkaline phosphatase activity
The identification and characterization of osteo-blast-like cell cultures are based on the assays of spe-
cific metabolites. For this purpose, the activity of ALP,a membrane enzyme routinely used in in vitro experi-ments as a relative marker of osteoblastic differentia-tion,23,28 was determined and the results are shown inFigure 3.
ALP levels of bone marrow cells grown in controlconditions for 21 days were relatively low in the firstweek of culture, rose sharply in the second week tosome peak level of response, and dropped by the thirdweek of incubation [Fig. 3(a)].
In cultures grown for 7 days, only the presence of SSand Ni ions resulted in a significant reduction in ALPactivity (p < 0.05, 77 and 58%, respectively). In culturesgrown for 14 days, the presence of SS in the second
Figure 2. Total protein content (mg/cm2) in human bone marrow cultures grown in control conditions and in the presenceof metallic slurries in different phases of the culture period: (a–d) same as in legend of Figure 1. Control (j); SS (x); Fe (+);Cr (m); Ni (d). *Significant differences (p < 0.05) from control cultures.
181IN VITRO OSTEOBLASTIC DIFFERENTIATION

week of incubation (phase characterized by an expo-nential increase in ALP activity) resulted in a signifi-cant decrease in ALP levels (p < 0.05, 75% reduction);cultures exposed to Fe and Ni ions also presentedlower levels of ALP in the second week of cultures ascompared to those found in control cultures (p < 0.05,a reduction of 31 and 33%, respectively), and Cr ionscaused no detectable effect in the activity of enzyme.In cultures maintained for 21 days, the addition ofmetal species during the third week of incubation didnot affect significantly the behavior of bone marrowcultures although in the presence of SS and Ni ions asmall decrease in ALP activity was observed.
Results relative to the activity of ALP expressed innmol/min/mg protein (results not shown) in controland metal-treated cultures, which give informationabout the mean ALP content per total amount of pro-tein (intracellular + extracellular), presented profilessimilar to those obtained for ALP expressed in nmol/min/cm2.
The results presented in Figure 3 show that SS andNi salt cause a deleterious effect to ALP activity inbone marrow cultures; Cr salt had practically no effecton the activity of this enzyme, and Fe salt caused asmall decrease when present in the second week ofculture.
Figure 3. ALP activity (nmol/min/cm2) in human bone marrow cultures grown in control conditions and in the presenceof metallic slurries in different phases of the culture period: (a–d) same as in legend of Figure 1. Control (j); SS (x); Fe (+);Cr (m); Ni (d). *Significant differences (p < 0.05) from control cultures.
182 MORAIS ET AL.

Quantification of total and ionized Ca and P, andFe, Ni, and Cr in the culture media
Media from cultures grown in control conditionsand in the presence of metallic slurries were collectedthroughout the incubation period and analyzed for thequantification of total and ionized Ca and P and alsofor Fe, Cr, and Ni. Quantification of these species incontrol and metal-ions-exposed media incubated inthe absence of bone marrow cells also was performed.Levels measured of total and ionized Ca and P andmetal ions were not cumulative as the culture mediumwas replaced totally once or twice (during the incuba-tion periods in which the effect of the metallic specieswere studied) a week; values shown reflect changesoccurring in intervals of 3–4 or 7 days throughout theculture period. Results are presented in Table I, Figure4, and Table II, respectively.
Total and ionized Ca and P
In the absence of bone cells, levels of total and ion-ized Ca and P in control and metal-treated culturemedia were similar throughout the incubation peri-ods. Total Ca concentration ranged from a minimumof 2.39 ± 0.030 mmol/L to a maximum of 2.61 ± 0.13mmol/L, and most of it was ionized (1.54 ± 0.027mmol/L to 1.90 ± 0.054 mmol/L); concentration oftotal P ranged from 10.8 ± 0.099 mmol/L to 11.1 ± 0.20mmol/L, and the content of ionized P was signifi-cantly lower (1.28 ± 0.0040 mmol/L to 2.03 ± 0.010mmol/L).
Ca and P in culture medium originate from Ca andphosphate compounds present in a-MEM (2.4mmol/L and 0.90 mmol/L, respectively) and in fetalbovine serum (∼3.8 mmol/L and ∼4.0 mmol/L, re-spectively). Addition of b-glycerophosphate (10mmol/L) to the incubation medium was responsiblefor the increased value observed for total P concentra-tion as compared to that found for Pi. This ester phos-
phate provides a potential source of P ions requiredfor the mineralization process in bone nodules form-ing cultures (deposition of calcium phosphate).29–31
One can notice that Ca and P content in culture me-dium changed slightly during the time course of theexperiment. This fact may be explained bearing inmind that the culture medium has to be prepared sev-eral times during the assay, and it is very difficult topipette the culture medium uniformly due to the pres-ence of proteins (which bind Ca and P). The possibilityof the incorrect application of the selected methodsmay be discarded since good precision and accuracywere obtained in preliminary comparative studies12
and because other authors30,32 have reported similarvariations in Ca and P content using other techniques.Furthermore, it can be observed that total Ca and Pconcentrations are, in the majority, lower than the ex-pected theoretical values based upon the proportion ofeach in the culture medium (a-MEM + 10% fetal bo-vine serum + 10 mmol/L b-glycerophosphate), that is,∼2.8 mmol/L for Ca and ∼11.3 mmol/L for P, indicat-ing that some precipitation occurs in the stock culturemedium flask and/or in all culture plates during theincubation period.
Culture medium collected from cultures grown incontrol conditions for 21 days (Table I) showed con-stant levels of total Ca and P and also ionized Caduring the first 10 days of incubation although con-centration of Pi increased during the first 2 weeks ofculture (maximal values were 9.30 ± 0.083 mmol/L atday 14), suggesting that most of the b-glycerophosphateadded is utilized as a substrate by ALP, which is pres-ent in high levels in these cultures. However, afterapproximately 2 weeks of incubation, levels of Ca andP started to decrease continuously until the end of theculture. These variations probably were caused by thepronounced reductions verified in the ionized con-tents of these species and reflect, most probably, themineralization process, that is, the deposition of cal-cium phosphate in the cultures. These results seem toagree with those reported in the literature concerningstudies performed in similar culture systems, that is,
TABLE ILevels (Mean ± Standard Deviation) of Total and Ionized Ca and P in the Incubation Medium from Human Bone
Marrow Cultures Grown for 21 Days in Control Conditions
Days
Total Calcium (mmol/L) Ionized Calcium (mmol/L) Total Phosphorus (mmol/L) Ionized Phosphorus (mmol/L)
No Cells Cells No Cells Cells No Cells Cells No Cells Cells
0 2.57 ± 0.015 2.57 ± 0.015 1.90 ± 0.054 1.90 ± 0.054 11.1 ± 0.20 11.1 ± 0.20 1.28 ± 0.004 1.28 ± 0.0043 2.61 ± 0.13 2.50 ± 0.019 1.59 ± 0.034 1.60 ± 0.044 11.2 ± 0.089 11.0 ± 0.14 1.80 ± 0.004 2.17 ± 0.0467 2.51 ± 0.025 2.48 ± 0.065 1.61 ± 0.022 1.56 ± 0.010 10.9 ± 0.077 10.9 ± 0.066 1.79 ± 0.013 2.94 ± 0.071
10 2.45 ± 0.075 2.46 ± 0.005 1.60 ± 0.044 1.62 ± 0.058 10.9 ± 0.15 11.0 ± 0.040 1.74 ± 0.008 3.89 ± 0.03114 2.50 ± 0.042 1.92 ± 0.089 1.61 ± 0.067 1.31 ± 0.010 11.0 ± 0.10 10.6 ± 0.088 1.80 ± 0.015 9.30 ± 0.09017 2.56 ± 0.10 1.00 ± 0.028 1.61 ± 0.020 0.74 ± 0.096 10.9 ± 0.12 10.0 ± 0.11 1.84 ± 0.009 8.10 ± 0.07521 2.52 ± 0.060 0.68 ± 0.018 1.54 ± 0.064 0.45 ± 0.022 11.0 ± 0.085 9.62 ± 0.050 1.81 ± 0.048 7.40 ± 0.15
Levels of these species in the culture medium incubated in the absence of cells are also shown.
183IN VITRO OSTEOBLASTIC DIFFERENTIATION

in cultures forming mineralized bone nodules in theseexperimental conditions.29–31 Results presented inTable I show that nonbiological mineral depositionoccurring in the absence of bone cells seems to beinsignificant when compared with the high consump-tion of Ca and P observed during the biomineraliza-tion that occurred in control cultures from approxi-mately 2 weeks onward.
No significant variations in total Ca and P, and alsoin ionized Ca, were measured in media from controland metal-exposed cultures maintained for 7 days.Nevertheless, the level of Pi increased from 1.28 ±0.0040 mmol/L (at day 0) to 4.39 ± 0.10 mmol/L in thecontrol situation and to 2.56 ± 0.044 mmol/L, 4.73 ±0.18 mmol/L, 4.57 ± 0.089 mmol/L, and 3.46 ± 0.012mmol/L in the presence of SS, Fe, Cr, and Ni slurries,respectively (results not shown).
Results relative to cultures maintained for 14 daysand exposed to metal ions in the second week arepresented in Figure 4(A). As compared to control cul-tures, Cr-treated cultures presented a similar patternof variation in the levels of total and ionized Ca and Pin the culture media; Fe-treated cultures presentedlower levels of Pi in the second week of incubationand also a lower consumption of Ca and P from theculture medium (levels of these species in control andFe-treated cultures were significantly different, p <0.05). In contrast, media from SS- and Ni-treated cul-tures contained as much total Ca and P and ionized Caas media incubated in the absence of cells; Pi levels inthe culture media were markedly lower during thesecond week of incubation (p < 0.05) compared to thecontrol values.
In cultures maintained for 21 days [Fig. 4(B)], theaddition of SS and the corresponding metal salts dur-ing the third week of culture caused no detectableeffect in the behavior of bone marrow cultures relativeto levels of total and ionized Ca and P in the culturemedia.
Fe, Cr, and Ni
In control medium, mean ± standard deviation val-ues of Fe, Cr, and Ni were, respectively, (1.64 ± 0.07) ×10−5 mol/L, (5.63 ± 0.18) × 10−7 mol/L, and (4.96 ±0.22) × 10−7 mol/L; no significant changes in the con-centration of these metal ions were observed duringthe control assays performed in the presence and ab-sence of cells. Results are in agreement with previ-ously reported studies in rat bone marrow osteogeniccultures.33 Addition of SS to the control culture me-dium increased the Fe, Cr, and Ni concentrations to(2.54 ± 0.04) × 10−5 mol/L (∼55%), (2.90 ± 0.07) × 10−6
mol/L (∼ fourfold), and (2.21 ± 0.06) × 10−6 mol/L (∼threefold), respectively. Similarly, addition of metal
salts to the control culture medium rose the metal con-tents to (2.54 ± 0.02) × 10−5 mol/L for Fe, (2.94 ± 0.05)× 10−6 mol/L for Cr, and (2.23 ± 0.07) × 10−6 mol/Lfor Ni.
Table II shows the variations of Fe, Cr, and Ni con-tents measured in SS-treated culture media and also inthe culture media exposed to the metal salts collectedfrom bone marrow cultures maintained for 7, 14, and21 days in the experimental conditions studied. In theabsence of cells, metal contents appeared to be ap-proximately constant in the three situations investi-gated. In contrast, a different pattern occurred in thepresence of osteogenic cells. In SS-treated cultures,substantial reductions in the Cr and Ni concentrationswere verified whereas in the cultures exposed to themetal salts, only the Ni levels were markedly lower.
Histochemical assays and SEM examination ofcell cultures
Human bone marrow cells grown in control condi-tions showed the formation of a nodular structure thatincreases in size with incubation time. Cells associatedwith the nodules presented intense histochemicalstaining for ALP; cultures stained for ALP in the testeddays showed results that were in agreement withthose observed for the biochemical determination ofthe enzyme and are not shown. In 21-day cultures, thethree-dimensional nodules stained intensively for Caand phosphates deposition, as demonstrated by astrong positive reaction in alizarin red and von Kossaassays, respectively; these two assays gave the samekind of information though only the von Kossa assayresults are shown (Fig. 5). The SEM examination of21-days cultures [Fig. 6(a)] shows the presence of nu-merous globular structures, shown by X-ray micro-analysis to contain Ca and P [Fig. 6(b)] in an atomicratio of 1.7 ± 0.06 (n = 10), which is in agreement withthe Ca/P ratio of the hydroxyapatite (HA). Bone mar-row cells grown for 7 days in the presence of Fe and Crions showed behavior similar to that observed in con-trol conditions, but exposition to SS and Ni ions re-sulted in evident signs of deterioration of the cultures,demonstrated by a significative decrease in the num-ber of cells, with the remaining cells presenting alter-ation in cell morphology (cells became bigger andmore spread) and a weak histochemical reaction forALP. Also, in cultures grown for 14 days, the presenceof SS in the second week resulted in a similar delete-rious effect in the behavior of bone marrow cells; nomarked differences were observed in the histochemi-cal reactions and SEM appearance of the culturestreated with Fe and Cr salts compared with controlcultures. In 21-days cultures, the presence of SS andthe corresponding separate metal salts during the
184 MORAIS ET AL.

third week of culture did not result in apparentchanges in the histochemical behavior and SEM ap-pearance compared to cultures grown in control con-ditions (results not shown).
DISCUSSION
In this study we used a combination of biochemicalassays (such as enzymatic reduction of MTT, ALP ac-
tivity, and total protein content) with chemical analy-ses of culture media (with regard to Ca, P, Fe, Cr, andNi concentrations by spectrophotometry and electro-chemistry) and morphological assays. We were able tocharacterize the differential cellular response, depend-ing on the stage of differentiation of the target osteo-blasts, to 0.1% of a stainless steel slurry (8.95 × 10−3
mol/L Fe + 2.35 × 10−3 mol/L Cr + 1.72 × 10−3 mol/LNi) and to its individual major compounds (slurries of
Figure 4. Levels of total and ionized Ca (a, b) and P (c, d) in the culture media from metal-treated cultures. The levels of thesespecies in the culture media incubated in the absence of cells also are shown. (A) Cells grown for 14 days and exposed tometallic slurries in the second week of incubation; (B) cells grown for 21 days and exposed to metallic slurries in the thirdweek of incubation. In the presence of cells: Control (j); SS (x); Fe (l); Cr (m); Ni (d). In the absence of cells (dashed lines):Control (h); SS (✳); Fe (L); Cr (n); Ni (s).
185IN VITRO OSTEOBLASTIC DIFFERENTIATION

Fe, Cr, and Ni salts in the concentration present in theAISI 316L slurry). This concentration of corrosionproducts was selected because our previous studieshad examined the dose and time effects of (a) 0.001%,0.01%, and 0.1% of this AISI 316L slurry and corre-sponding separate major metal ions on rabbit osteo-genic cell proliferation and function,34 and of (b)0.01%, 0.1%, and 1% of another AISI 316L slurry (8.78× 10−3 mol/L Fe + 4.31 × 10−3 mol/L Cr + 2.56 × 10−3
mol/L Ni) in the behavior of osteogenic cells derivedfrom rat bone marrow.33,35 A common feature of bothpreliminary investigations was that 0.001% and 0.01%
of the slurries had a little influence on the osteoblasticbehavior and 1% caused cell death.35 AISI 316L slurryat a concentration of 0.1% appeared to exert some del-eterious effect in the rat culture system.33,35 Bone tis-sue in vivo probably is not exposed to metal ions atconcentrations that cause cell death. However, metalions at sublethal concentrations still can affect the pro-liferation and function of osteoblastic cells and inter-fere with the normal mineral deposition, contributingto implant failure.
The selected biochemical parameters showed thatthe human bone marrow cell cultures grown in control
Figure 4. Continued.
186 MORAIS ET AL.

conditions, that is, in the presence of ascorbic acid,b-glycerophosphate, and dexamethasone, present os-teoblastic features, as demonstrated by the presence ofhigh activity of ALP [Fig. 3(a)] and the formation ofmineralized nodules. These cultures presented a be-havior relative to cell proliferation and osteoblasticdifferentiation similar to that reported in other bone-nodule forming cultures.11,13,17,29,36–38 After a lagphase of approximately 1 week, bone marrow cellsenter a period of active proliferation that is function-ally related to the synthesis of a bone-specific extra-cellular matrix during the second week of incubationand after. In the third week, a tendency for a station-ary phase in cell growth is observed (as a consequenceof the accumulation and maturation of the extracellu-lar matrix36). ALP expression followed a similar pat-tern of variation. The significant increase in the levelsof this enzyme during the second week of incubationsuggest that cells were shifting to a more differenti-ated state.39–41 ALP appears to play a crucial role inthe initiation of matrix mineralization; after that, ex-pression of this enzyme is down regulated.11,13,29,37–38
The mineralization process, an osteoblast-specificfunction, occurred during the third week of culture, asdemonstrated by the strong positive reactions foralizarin red and von Kossa assays, the presence ofnumerous mineral globular structures on SEM obser-vation, and also by a significant consumption of ion-ized Ca and P from the culture medium during thisphase of the culture.
The presence of SS and Ni salt in bone marrow cul-tures during the first and second weeks of culture sig-nificantly disturbs the normal behavior of these cul-tures. These results suggest that these slurries impairthe initial stage of the culture, that is, the lag phase(first week); also, they exert toxic effects during theexponential phase of cell growth and ALP expression(second week), disturbing the proliferation phase thatis associated with the formation of extracellular ma-trix. According to this, cultures treated with these me-tallic species present a weak ability to ionize the b-
glycerophosphate added (probably because of thelower levels of ALP existing in these cultures), and noconsumption of Ca and P from the culture media wasobserved, as compared to control cultures. Resultsalso show that the presence of Ni salt appears to resultin a less pronounced effect compared to that observedin the presence of SS. However, it is interesting to notethat despite their toxicity in these stages of the culture,the presence of SS and Ni salt during the third week ofculture, when expression of osteoblastic functions oc-curs (mineralization process), did not result in anydetectable effect. Fe salt also disturbs the behavior ofbone marrow cell cultures when present during thelag phase and proliferation phase (first and secondweek) and a somewhat compromised response be-tween the normal pattern (control and Cr-treated cul-tures) and intense inhibition (SS- and Ni-salt-addedcultures) is observed. Fe did not affect the progressionof the mineralization phase.
Osteogenic cultures exposed to Cr salt (Cr3+) pre-sented a pattern similar to the controls, indicating thatthis element did not interfere, in the concentration
TABLE IIPercentage of Variation of Fe, Cr, and Ni Contents* in the Media of Cultures Treated With AISI 316L CorrosionProducts and the Corresponding Separate Major Metal Salts in the Absence or in the Presence of Human Bone
Marrow Cells and Relative to the Concentrations That Were Measured After the Addition of AISI 316L CorrosionProducts or the Corresponding Metal Salts to the Culture Medium
Assay†
Fe (%) Cr (%) Ni (%)
AISI 316L-TreatedCultures
Fe Salt-TreatedCultures
AISI 316L-TreatedCultures
Cr Salt-TreatedCultures
AISI 316L-TreatedCultures
Ni Salt-TreatedCultures
No Cells Cells No Cells Cells No Cells Cells No Cells Cells No Cells Cells No Cells Cells
1 −12 −16 −5.5 −8.5 −5.3 −42 −12 −14 −13 −36 −1.1 −532 −14 −12 −2.6 −9.8 +0.92 −62 −17 −14 −2.6 −62 +1.4 −663 −12 −15 −6.8 −1.0 −1.3 −65 −15 −17 −1.2 −50 −1.7 −58
*Increases (+) or decreases (−); maximal relative standard deviations obtained for Fe, Cr, and Ni concentrations were 4.5%,3.5%, and 4.5%, respectively †Key: 1: cells grown for 7 days; 2: cells grown for 14 days and exposed to metal ions during thesecond week; 3: cells grown for 21 days and exposed to metal ions during the third week.
Figure 5. Histochemical assay (original magnification ×40)for the identification of phosphate deposition in humanbone marrow cultures grown in control medium.
187IN VITRO OSTEOBLASTIC DIFFERENTIATION

studied, in the osteoblastic differentiation of bonemarrow cells, an observation suggested by the lack ofeffect of MTT reduction (except a small reduction inthe first week), ALP activity, and also the pattern ofvariation of Ca and P levels in the incubation medium.In a recent study,42 Cr3+ also was recognized as havinglow cytotoxicity in ROS 17/2.8 rat osteoblast-like cellscompared to other metal ions that can be releasedfrom metallic implants in vivo, probably because of itsmembrane impermeability and biologic inactivity.Previous studies43 have revealed that Cr6+ induces themost cytotoxicity response from bone marrow stromalcells cultured in vitro, as compared to Fe3+ and Ni2+.These apparent contradictory results may be ex-plained by the different oxidation state used since thecytotoxicity of soluble and insoluble hexavalent Crcompounds appears to be 100–1000 times greater thanthat demonstrated by trivalent Cr compounds.44
Results show that metallic slurries do not interferein the normal behavior of bone cells cultures whenthey are present during the culture period where themineralization process occurs (third week). In controlconditions, mineralization appears to begin from 2weeks onward. An enhanced expression of ALP isneeded just before the onset of matrix mineralizationand well may be the triggering event of the calcifica-tion cascade,39,45 providing localized enrichment of Pifor HA crystal nucleation and proliferation; followingthe initiation of mineralization there is a decrease inALP activity. Thus cells are more importantly in-volved in mineral initiation (providing high levels ofALP) while propagation remains primarily physico-chemical45; exposition of the initial crystals formed tothe rich Ca and P culture medium activates crystalproliferation in what is essentially a physicochemicalprocess.45
The pattern of variation for Ni and Cr content in theincubation medium of cultures exposed to SS wassimilar to that observed previously33; decreased con-centrations of the ions were similar throughout thethree incubation periods, showing an almost regularrate of deposition of these metal ions in the cell cul-tures.33 Media of cultures treated with Ni salt pre-sented the same trend of variation. As demonstratedby other authors,44,46–48 uptake of these species fromits complexes by cells via a transport system may oc-cur with consequent interference in the cellular me-tabolism (intracellular metallic deposition). Also, itmay be hypothesized that some metabolites producedby bone cells bind Cr (originated from SS) and Ni(originated from SS or Ni salt), preferentially promot-ing a sort of selective precipitation, in this case anextracellular precipitation. These two pathways, sepa-rately or together, can contribute to the decreased con-tents measured in cell cultures. Chemical precipitationappears not to contribute significantly to these reduc-tions since in the absence of cells only slight variationson the levels of these metal ions were observed, sug-gesting a biologically induced phenomenon. In con-trast to Ni, the Cr salt used (Cr3+) did not reproducethe biological behavior of Cr compounds that origi-nate from the corrosion of AISI 316L; the maximumCr3+ level of variation in the presence of cells was16.6%, which is not significantly different from themaximum decrease (17.1%) observed in the absence ofcells. Merritt et al.46 showed that corrosion productsfrom stainless steel and cobalt–chrome alloy behavesimilarly to the metal salts and that fretting49 of AISI316L in serum results in corrosion products containingCr6+. Cr6+ is able to penetrate the cell membrane butnot trivalent Cr, which is excluded (except at concen-trations of 100 or 1000 times those for the hexavalentform).44,50 Since Cr can exist in a variety of states, thepossibility of several Cr valence states also exists;however, these other valence states will not be nearly
Figure 6. (a) SEM appearance of human bone marrow cul-tures grown for 21 days in control conditions. Bar = 10 mm.(b) Energy dispersive X-ray microanalysis spectrum of thethree-dimensional globular deposits showing the Ca and Ppeaks.
188 MORAIS ET AL.

as toxic as the Cr6+.51 These hypotheses, in combina-tion with previous results obtained by Lucas et al.51
that indicate in vitro induction of cellular ultrastruc-tural damage by Ni, may help to explain the cytotox-icity of SS and also the low levels of Cr found in cul-ture media exposed to SS and osteogenic cells.
Although the primary constituents of AISI 316L arebiologically essential trace elements, implantation of adevice containing this alloy will constitute a long-termin vivo source of reactive trace elements, any of whichmay produce bone sequel when introduced at levelsnot physiologically regulated. Furthermore, metalspecies react in biological systems, and as differentspecies are formed, significant changes in toxicity mayoccur.
CONCLUSIONS
The results of this study emphasize that corrosionproducts have differential effects, depending on thestate of differentiation of the target cells, and also thatour understanding of these processes will be incom-plete until the relative proportions and differentchemical species (of the same element) formed duringimplant degradation are determined and character-ized.
S. M. is grateful to CIENCIA/PRAXIS XXI for her Ph.D.scholarship.
References
1. Williams D, Cahn RW, Bever MB, editor. Concise encyclopediaof medical & dental materials. Oxford, New York, Beijing,Frankfurt, Sao Paulo, Sydney, Tokyo, Toronto: PermagonPress; 1990. p 4–70.
2. Linden JV, Hopler SM Gossling HR, Sunderman Jr FW. 1985.Blood nickel concentrations in patients with stainless-steel hipprotheses. Ann Clin Lab Sci 1985;15:459–464.
3. Weber GW, Reid BL, Nickel toxicity in young growing mice. JAnimal Sci 1969;28:620–623.
4. O’Dell GD, Miller WJ, King WA, Moore SL, Blackmon DM.Nickel toxicity in the young bovine. J Nutrition 1970;100:1447–1453.
5. Louria DB, Joselow MM, Browder AA. The human toxicity ofcertain trace elements. Ann Intern Med 1972;76:307–319.
6. Whanger PD. Effects of dietary nickel on enzyme activities andmineral contents in rats. Toxicol Appl Pharmacol 1973;25:323–331.
7. Woodman JL, Black J, Jimenez S. Organometallic corrosionproducts of 316L SS and HS-21 in serum: In vitro and in vivocomparison in an acute rat model. In Winter GD, Gibbon DF,Plenk Jr H, editors. Biomaterials 1980. Chichester, New York,Brisbone, Toronto, Singapore: John Wiley & Sons, 1982. p 245–253.
8. Parfitt AM. The physiological and clinical significance of bonehistomorphometric data. In: Recker RR, editor. Bone histomor-
phometry: Techniques and interpretation. Boca Raton, Florida:CRC Press; 1983. p 143–223.
9. Jilka RL, Manolagas SC. The cellular and biochemical basis ofbone remodeling. In: Marcus R, editor. Osteoporosis. Boston:Blackwell Scientific Publications; 1994. p 17–48.
10. Owen M. Lineage of osteogenic cells and their relationship tothe stromal system. In: Peck WA, editor. Bone and mineralresearch, Vol. 3. Amsterdam: Elsevier Science Publishers; 1985.p 1–25.
11. Owen TA, Aronow M, Shalhoub V. Progressive developmentof the rat osteoblast phenotype in vitro: Reciprocal relation-ships in expression of genes associated with osteoblast prolif-eration and differentiation during formation of the bone extra-cellular matrix. J Cell Physiol 1990;143:420–430.
12. Morais S, Carvalho GS, Sousa JP. Potentiometric determinationof total and ionized Ca in osteoblast-like cell culture medium.Electroanalysis 1996;8(12):1174–1178.
13. Morais S, Carvalho GS, Sousa JP. Iron determination in osteo-blast-like cell culture medium by adsorptive cathodic strippingvoltammetry with a mercury microelectrode. Electroanalysis1997;9(10):791–795.
14. Morais S, Carvalho GS, Sousa JP. Chromium determination inosteoblast-like cell culture medium by catalytic cathodic strip-ping voltammetry with a mercury microelectrode. J Trace El-ements Med Biol 1998;2:101–108.
15. Morais S, Carvalho GS, Sousa JP. Determination of Ni in os-teoblast-like cell culture medium by adsorptive cathodic strip-ping voltammetry with mercury microelectrodes. Electro-analysis 1997;9(5):422–426.
16. Morais S, Carvalho GS, Sousa JP. Optimization study for nickelquantification in cell culture medium using mercury film mi-croelectrodes. Portugaliae Electrochimica Acta 1996;14:267–272.
17. Maniatopoulos C, Sodek J, Melcher AH. Bone formation invitro by stromal cells obtained from bone marrow of youngadult rats. Cell Tiss Res 1988;254:317–330.
18. Berridge V, Tan AS. Characterization of the cellular reductionof 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bro-mide (MTT): Subcellular localization, substrate dependence,and involvement of mitochondrial electron transport in MTTreduction. Arch Biochem Biophys 1993;303:474–482.
19. Bessey OE Lowry O↗ Breck Mm A method for the rapid deter-mination of alkaline phosphatase with five cubic millileters ofserum. J Biol Chem 1946;164:321–326.
20. McGee–Russell M. Histochemical methods for calcium. J His-tochem Cytochem 1958;6:22–42.
21. Fiske CH, Subbarow Y. The colorimetric determination ofphosphorus. J Biol Chem 1925;66:373–400.
22. Bellows CG, Aubin JE, Heersche JNM. Physiological concen-trations of glucocorticoids stimulate formation of bone nodulesfrom isolated rat calvaria cells in vitro. Endocrinology 1987;121:1985–1992.
23. Aronow MA, Gerstenfeld LC, Owen TA. Factors that promoteprogressive development of the osteoblast phenotype in cul-tured fetal rat calvaria cells. J Cell Phys 1990;143:213–221.
24. Gotoh Y, Hiraiwa K, Narajama M. In vitro mineralization ofosteoblastic cells derived from human bone. Bone and Miner1990;8:239–250.
25. Cheng SL, Yang JW, Riffas L, Zhang SF, Avioli LV. Differen-tiation of human bone marrow osteogenic stromal cells invitro. Induction of the osteoblast phenotype by dexametha-sone. Endocrinology 1994;134:277–286.
26. Gundle R, Joyner CJ, Triffit JT. Human bone tissue formationin diffusion chamber culture in vivo by bone-derived cells andmarrow stromal fibroblastic cells. Bone 1995;16:597–601.
27. Pizzoferrato A, Ciapetti G, Stea S, Cenni E, Arcicla CR, GranchiDC, Savarino L. Cell culture methods for testing biocompat-ibility. Clin Mater 1994;15:173–190.
189IN VITRO OSTEOBLASTIC DIFFERENTIATION

28. Gerstenfeld LC, Chipman SD, Glowacki J, Lian JB. Expressionof differential function by mineralizing cultures of chicken os-teoblasts. Dev Biol 1987;122:46–60.
29. Bellows CG, Aubin JE, Heersche JNM. Initiation and progres-sion of mineralization of bone nodules formed in vitro: Therole of alkaline phosphatase and organic phosphate. Bone andMiner 1991;14:27–40.
30. Bellows CG, Heersche JNM, Aubin JE. Inorganic phosphateadded exogenously or released from b-glycerophosphate ini-tiates mineralization of osteoid nodules in vitro. Bone andMiner 1992;17:15–29.
31. Chung CH, Golub EE, Forbes E, Tokuoka T, Shapiro IM.Mechanisms of action of b-glycerophosphate on bone cell min-eralization. Calcif Tissue Int 1992;51:305–311.
32. Boskey AL, Stiner D, Doty SB, Binderman I, Levoy P. Studies ofmineralization in tissue culture: Optimal conditions for carti-lage calcification. Bone and Miner 1992;16:11–36.
33. Morais S, Sousa JP, Fernandes MH, Carvalho GS, de Bruijn JDvan Blitterswijk CA. Decreased consumption of Ca and P dur-ing in vitro biomineralization and biologically induced depo-sition of Ni and Cr in presence of stainless steel corrosionproducts. J Biomed Mater Res 1998;42:199–212.
34. Morais S, Sousa JP, Fernandes MH, Carvalho GS. In vitrobiomineralization by osteoblast-like cells. 1. Retardation of tis-sue mineralization by metal salts. Biomaterials 1998;19:13–21.
35. Morais S, Sousa JP, Fernandes MH, Carvalho GS, de Bruijn JD,van Blitterswijk CA. Effects of AISI 316L corrosion products inin vitro bone formation. Biomaterials 1998;19:999-1007.
36. Stein GS, Lian JB. Molecular mechanisms mediating develop-mental and hormone-regulated expression of genes in osteo-blasts: An integrated relationship of cell growth and differen-tiation. In: Noda M, editor. Cellular and molecular biology ofbone. Tokyo: Academic Press Inc., 1993. p 47–95.
37. de Bruijn JD, Klein CPAT, de Groot K, van Blitterswijk CA. Theultrastructure of the one-hydroxyapatite interface in vitro. JBiomed Mater Res 1992;26:1365–1382.
38. Stein GS, Lion JB. Molecular mechanisms mediated prolifera-tion–differentiation interrelationships during progressive de-velopment of the osteoblast phenotype: Update 1995. EndocrinRev 1995;4:290–297.
39. Abe Y, Akamine A, Aida Y, Maeda K. Differentiation and min-
eralization in osteogenic precursor cells derived from fetal ratmandibular bone. Calcif Tissue Int 1993;52:365–371.
40. Lomri A, Tran MPV, Mott M. Characterization of endostealosteoblastic cells isolated from mouse caudal vertebrae. Bone1988;9:165–175.
41. McCulloch CAG, Fair CA, Tenebaum HC, Limeback H, Hom-areau R. Clonal distribution of osteoprogenitor cells in cul-tured chick peristoa: Functional relationship to bone forma-tion. Dev Biol 1990;140:352–361.
42. Sun ZL, Wataha JC, Hanks CT. Effects of metal ions on osteo-blast-like cell metabolism and differentiation. J Biomed MaterRes 1997;34:29–37.
43. Puleo DA, Huh WW. Acute toxicity of metal ions in cultures ofosteogenic cells derived from bone marrow stromal cells. JAppl Biomater 1995;6:109–116.
44. Katz SA, Salem H. The toxicology of Cr with respect to itschemical speciation: A review. J Appl Toxicol 1993;13:217–224.
45. Anderson HC, Morris DC. Mineralization. In: Mundy GR,Martin TJ, editors. Handbook of experimental pharmacology.Berlin: Springer–Verlag; 1993.p 267–298.
46. Merritt K, Brown SA. Biological effects of corrosion productsfrom metals. In: Fraker AC, Griffin CD, editors. Corrosion anddegradation of implant materials: Second symposium. ASTMSTP 859. Philadelphia: American Society for Testing and Ma-terials; 1985. p 195–207.
47. Rae T. The toxicity of metals used in orthopaedic prostheses. JBone Joint Surg 1981;63-B:435–440.
48. Webb M, Weinzier SM. Uptake of 63Ni2+ from its complexeswith proteins and other ligands by mouse dermal fibroblasts invitro. Br J Cancer 1972;26:292–298.
49. Merritt K, Wortman RS, Millard M, Brown SA. XPS analysis of316LVM corroded in serum and saline. Biomater Med DevicesArtif Organs 1983;11:115–124.
50. Bianchi V, Toso RD, Debetto P. Mechanisms of chromium tox-icity in mammalian cell cultures. Toxicology 1980;17:219–224.
51. Lucas LC, Bearden LJ, Lemons JE. Ultrastructural examinationof in vitro and in vivo cells exposed to elements from type 316Lstainless steel. In: Fraker AC, Griffin CD, editors. Corrosionand degradation of implant materials: Second symposium.ASTM STP 859. Philadelphia: American Society for Testing andMaterials; 1985. p 208–222.
190 MORAIS ET AL.




















