
in the mermithid nematode
7
JOURNAL OF INVERTEBRATE PATHOLOGY 32, 348-354 (1978) Catenaria anguihlae in the Mermithid Nematode Romanomermis culicivorax A. M. STIRLING AND E. G. PLATZER Department of Nematology, University of California. Riverside, California 92521 Received Feburary 28, 1978 Disease epizootics in laboratory cultures of the mermithid nematode Romanomermis culicivorax were caused by a chytridiomycetous fungal parasite, Catenaria anguillulae. Techniques used for mass rearing the nematode provided ideal conditions for dispersal of zoospores and multiplication of the fungus; up to 90% of the postparasites of R. culicivorax were parasitized. The fungus was isolated and grown on yeast extract-soluble starch agar, and the effects of different temperature and p H regimes on the fungus were investigated as possible methods for prevention or control. A temperature suitable for mass rearing the nematodes but unsuitable for the fungus could not be found, as C. angui/lulae had a broad temperature range. Adjustment of pH or a short period of chilling (0°C) were more promising control measures. At acid pH (<5), zoospore motility and germination and infection of postparasites were suppressed. The disease was controlled by rearing nematodes in water adjusted to a pH of 4.5. KEY WORDS: Catenaria anguillulae; Romanomermis culicivorax; fungal epizootics; pH effects; zoospore motility; zoospore germination. INTRODUCTION MATERIALS AND METHODS Recently, a chytridiomycetous fungal parasite, Catenaria anguillulae, reached epizootic proportions in laboratory cultures of the mermithid nematode Romanomermis culicivorax. In many cultures, up to 90% of the postparasites were infected, and, as a result, the potential yields of preparasites were reduced considerably. Studies were initiated to identify the fungus, to ascertain its life history, and to develop control measures. Previous attempts to control the disease in R. culicivorax with cyclohexi- mide, amphotericin B, griseofulvin, hydroxyquinoline, ethoxy-ethylmercury chloride, or copper sulfate were unsuccess- ful (Platzer and Brown, 1976). Therefore, the following abiotic factors were studied in an effort to develop cultural methods for controlling the disease: (i) the response of C. anguillulae to temperature; (ii) the effect of chilling on C. anguillulae; and (iii) the effect ofpH on zoospores of C. anguillulae. Nematode R. culicivorux was reared in mass cultures using the procedures of Petersen and Willis (1972). Autogenous Culex pipens was employed as the host (Brown and Platzer, 1977). The mosquito larvae were fed a mix- ture of finely ground Purina rabbit chow (3 parts) and brewer’s yeast (1 part). Fungus C. anguillulae was isolated by rinsing in- fected nematodes in sterile water and streak- ing discharged zoospores onto plates of YPSS agar medium (yeast extract, 4 g; K2HPOI, 1 g; MgSO,.7H,O, 0.5 g; soluble starch, 20 g; water, 1 liter; agar, 16 g) modi- fied from Emerson (1941). Colonies from single spores were maintained on YPSS at 27 to 30°C and one isolate was selected for further study. 0022-201 l/78/0323-0348$01.00/O Copyright 0 1978 by Academic Ress. Inc. AU rights of reproduction in any form reserved. 348
-
Upload
ucriverside -
Category
Documents
-
view
0 -
download
0
Transcript of in the mermithid nematode

JOURNAL OF INVERTEBRATE PATHOLOGY 32, 348-354 (1978)
Catenaria anguihlae in the Mermithid Nematode Romanomermis culicivorax
A. M. STIRLING AND E. G. PLATZER
Department of Nematology, University of California. Riverside, California 92521
Received Feburary 28, 1978
Disease epizootics in laboratory cultures of the mermithid nematode Romanomermis culicivorax were caused by a chytridiomycetous fungal parasite, Catenaria anguillulae. Techniques used for mass rearing the nematode provided ideal conditions for dispersal of zoospores and multiplication of the fungus; up to 90% of the postparasites of R. culicivorax were parasitized. The fungus was isolated and grown on yeast extract-soluble starch agar, and the effects of different temperature and p H regimes on the fungus were investigated as possible methods for prevention or control. A temperature suitable for mass rearing the nematodes but unsuitable for the fungus could not be found, as C. angui/lulae had a broad temperature range. Adjustment of pH or a short period of chilling (0°C) were more promising control measures. At acid pH (<5), zoospore motility and germination and infection of postparasites were suppressed. The disease was controlled by rearing nematodes in water adjusted to a pH of 4.5.
KEY WORDS: Catenaria anguillulae; Romanomermis culicivorax; fungal epizootics; pH effects; zoospore motility; zoospore germination.
INTRODUCTION MATERIALS AND METHODS
Recently, a chytridiomycetous fungal parasite, Catenaria anguillulae, reached epizootic proportions in laboratory cultures of the mermithid nematode Romanomermis culicivorax. In many cultures, up to 90% of the postparasites were infected, and, as a result, the potential yields of preparasites were reduced considerably. Studies were initiated to identify the fungus, to ascertain its life history, and to develop control measures. Previous attempts to control the disease in R. culicivorax with cyclohexi- mide, amphotericin B, griseofulvin, hydroxyquinoline, ethoxy-ethylmercury chloride, or copper sulfate were unsuccess- ful (Platzer and Brown, 1976). Therefore, the following abiotic factors were studied in an effort to develop cultural methods for controlling the disease: (i) the response of C. anguillulae to temperature; (ii) the effect of chilling on C. anguillulae; and (iii) the effect ofpH on zoospores of C. anguillulae.
Nematode
R. culicivorux was reared in mass cultures using the procedures of Petersen and Willis (1972). Autogenous Culex pipens was employed as the host (Brown and Platzer, 1977). The mosquito larvae were fed a mix- ture of finely ground Purina rabbit chow (3 parts) and brewer’s yeast (1 part).
Fungus
C. anguillulae was isolated by rinsing in- fected nematodes in sterile water and streak- ing discharged zoospores onto plates of YPSS agar medium (yeast extract, 4 g; K2HPOI, 1 g; MgSO,.7H,O, 0.5 g; soluble starch, 20 g; water, 1 liter; agar, 16 g) modi- fied from Emerson (1941). Colonies from single spores were maintained on YPSS at 27 to 30°C and one isolate was selected for further study.
0022-201 l/78/0323-0348$01.00/O Copyright 0 1978 by Academic Ress. Inc. AU rights of reproduction in any form reserved.
348

CATENARIA IN ROMANOMERIS 349
Temperature
The radial growth rate of C. anguillulae was assessed at 9”, 15”, 18”, 21”, 24”, 27”, 30”, 33”, 36”, 39”, and 42°C. Three-millimeter diameter plugs were transferred to YPSS plates and colony diameter was measured 9 days later. Zoospore production was as- sessed by growing five replicate cultures for 4 days at each of these temperatures, flood- ing the plates with glass-distilled water for 30 min, and estimating zoospore discharge by counting aliquots in a hemocytometer. The area of each colony was calculated from measurements of colony diameter.
Chilling
Suspensions of zoospores were obtained by flooding cultures of C. anguillulae on YPSS with sterile glass-distilled water. The numbers of germinated zoospores in the suspensions were estimated by killing zoo- spores with 10% formalin (Held, 1972) and counting an aliquot in a hemocytometer. Aliquots (0.5 ml) of the zoospore suspen- sions were added to 0.5 ml of sterile glass- distilled water in an ice bath at 0°C and maintained therein for 0,0.25,0.5, 1, 2, and 4 hr. Following each cold treatment (six replicates) a portion of each suspension was smeared on the surface of YPSS plates, which were incubated at 24°C for 20 hr. Zoospore viability was assessed by counting germinated and nongerminated spores. The remainder of each suspension was returned to room temperature, and 1 hr after treat- ment the number of moving zoospores in samples of 100 spores was counted.
Effect of pH on Zoospore Motility and Germination
Experiment 1. A zoospore suspension of C. anguillulae was prepared as described above, and 0.2-ml aliquots were added to six replicate test tubes containing 0.4 ml of 40 mM Tris-maleate buffer (pH 7.6, 7.3, 6.3), 12.5 mM succinate buffer (pH 5.0,4.2, 3.8), or glass-distilled water (pH 6.8). Zoo-
spore motility was estimated 1 hr later and after 24 hr the number of germinated zoo- spores was counted.
Experiment 2. The first experiment was repeated using citrate-phosphate buffer (pH 7.1, 6.1, 5.6, 5.0, 4.5, 4.0), prepared at a concentration 0.25 of that given in Gomori (1955), and glass-distilled water (pH 6.6).
Effect of pH on Infection of R. culicivorax
Experiment 1. Ten 7-day-old R. culici- vorax postparasites were placed in 35-mm Petri dishes containing 2.8 mlof40 mM Tris- maleate buffer (pH 7.9,7.0,6.1, 5.7, 5.3) or 12.5 mM succinate buffer (pH 5.0, 4.7,4.0). Zoospores prepared and counted as de- scribed above were added to eight replicates of each pH treatment, so that each dish contained about 110 x lo3 zoospores/ml. Controls consisted of additional dishes (eight replicates) containing nematodes in glass- distilled water (pH 6.7) and in buffers of pH 7.0, 5.0, and 4.7 to check for latent C. anguillulae infections in the nematodes used for the experiment. After incubation for 66 hr at 24”C, the number of infected nema- todes and final pH of each buffer were determined.
Experiment 2. The first experiment was repeated using citrate-phosphate buffer (pH7.0,6.5,6.0,5.6,5.1,4.6,4.l)andglass- distilled water (pH 6.6) and an inoculum level of 42 x 103 zoospores/ml. Nonexposed nematodes were maintained as checks in all buffers used.
RESULTS
Life History of C. anguillulae in R. culicivorax Cultures
The following observations were made on infected nematodes from the mass-reared system and experimental infections. C. anguillulae was never observed in newly emerged postparasites but after 36 hr at room temperatures (23-24°C) some nema- todes contained maturing sporangia (Fig. 1). Zoospores discharged through exit tubes
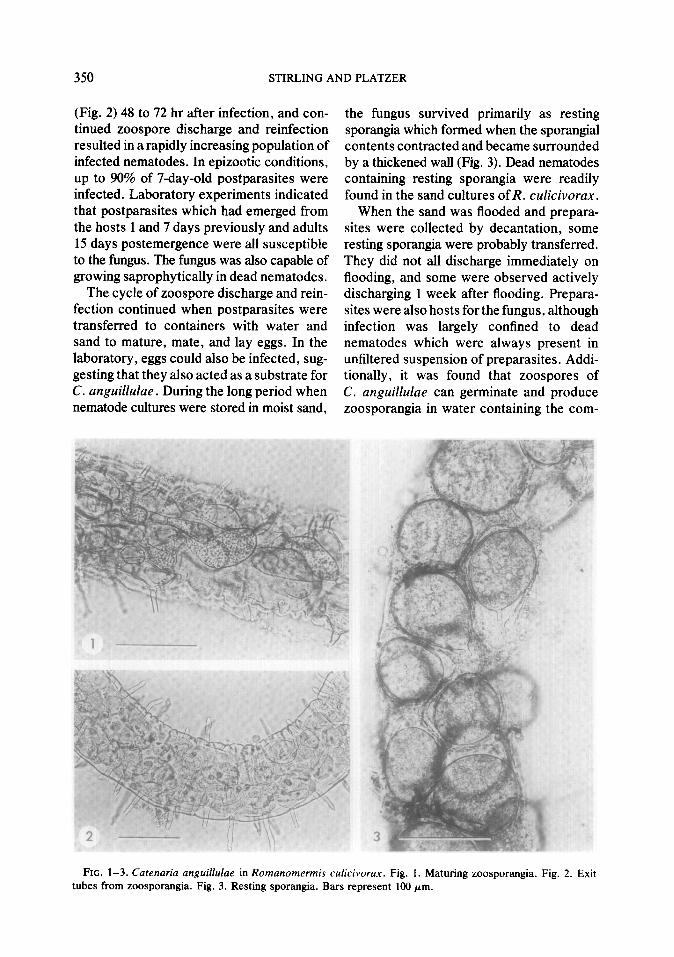
350 STIRLING AND PLATZER
(Fig. 2) 48 to 72 hr after infection, and con- tinued zoospore discharge and reinfection resulted in a rapidly increasing population of infected nematodes. In epizootic conditions, up to 90% of 7-day-old postparasites were infected. Laboratory experiments indicated that postparasites which had emerged from the hosts 1 and 7 days previously and adults 15 days postemergence were all susceptible to the fungus. The fungus was also capable of growing saprophytically in dead nematodes.
The cycle of zoospore discharge and rein- fection continued when postparasites were transferred to containers with water and sand to mature, mate, and lay eggs. In the laboratory, eggs could also be infected, sug- gesting that they also acted as a substrate for C. anguillulae. During the long period when nematode cultures were stored in moist sand,
the fungus survived primarily as resting sporangia which formed when the sporangial contents contracted and became surrounded by a thickened wall (Fig. 3). Dead nematodes containing resting sporangia were readily found in the sand cultures of R. culicivorux.
When the sand was flooded and prepara- sites were collected by decantation, some resting sporangia were probably transferred. They did not all discharge immediately on flooding, and some were observed actively discharging 1 week after flooding. Prepara- sites were also hosts for the fungus, although infection was largely confined to dead nematodes which were always present in unfiltered suspension of preparasites. Addi- tionally, it was found that zoospores of C. anguillulae can germinate and produce zoosporangia in water containing the com-
FIG. l-3. Catenaria anguillulae in Romanomermis culicivorax. Fig. 1. Maturing zoosporangia. Fig. 2. Exit tubes from zoosporangia. Fig. 3. Resting sporangia. Bars represent 100 pm.
CATENARIA IN ROMANOMERIS 351
7o r
60 - z 5
50 . z
= - z 40
0 30 -
2 0 & 20 . ”
IO
t OL ’ ’ ’ ’ ’ ’
6 9 12 15 I6 21 24 27 30 33 36 39 42
TEMPERATURE IC)
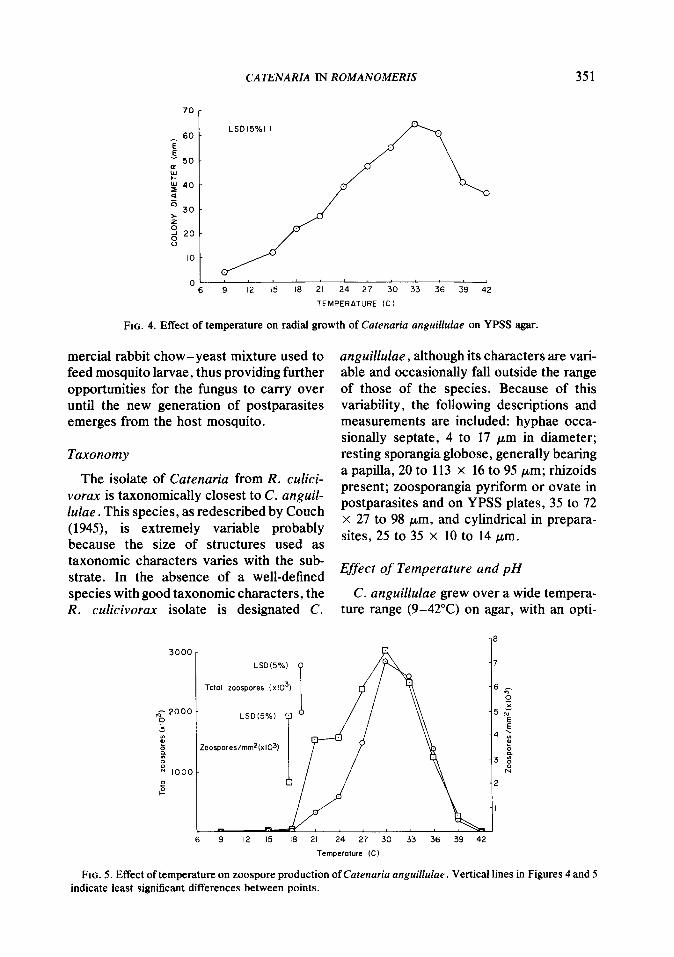
FIG. 4. Effect of temperature on radial growth of Cutennria anguihlae on YPSS agar.
mercial rabbit chow-yeast mixture used to feed mosquito larvae, thus providing further opportunities for the fungus to carry over until the new generation of postparasites emerges from the host mosquito.
Taxonomy
The isolate of Catenaria from R. culici- vorax is taxonomically closest to C. anguil- Mae. This species, as redescribed by Couch (1945), is extremely variable probably because the size of structures used as taxonomic characters varies with the sub- strate. In the absence of a well-defined species with good taxonomic characters, the R. culicivorax isolate is designated C.
3000 -
LSD(5%1
Total zoospores (~10~1
anguillulae , although its characters are vari- able and occasionally fall outside the range of those of the species. Because of this variability, the following descriptions and measurements are included: hyphae occa- sionally septate, 4 to 17 pm in diameter; resting sporangia globose, generally bearing a papilla, 20 to 113 x 16 to 95 pm; rhizoids present; zoosporangia pyriform or ovate in postparasites and on YPSS plates, 35 to 72 x 27 to 98 pm, and cylindrical in prepara- sites, 25 to 35 x 10 to 14 pm.
Effect of Temperature and pH
C. anguillulae grew over a wide tempera- ture range (9-42°C) on agar, with an opti-
6 9 12 15 I6 21 24 27 30 33 36 39 42
Temperature (C)
FIG. 5. Effect of temperature on zoospore production of Cutenariu anguillulae . Vertical lines in Figures 4 and 5 indicate least significant differences between points.

352 STIRLING AND PLATZER
mum at 33°C (Fig. 4). Zoospore production was largely restricted to temperatures of 21 to 39”C, with an optimum of 27 to 33°C (Fig. 5).
Chilling zoospores at 0°C for only 15 min was sufficient to significantly reduce their motility (Table 1). However, 4 hr of chilling was required before all zoospores encysted. Chilling did not affect spore viability, as almost all spores germinated on YPSS, re- gardless of the chilling treatment they received.
During the course of the experiments, p H values either remained constant or increased slightly. Zoospores of C. anguillulae showed decreased motility and ability to germinate in acid pH (Table 2) and also decreased ability to infect nematodes (Table 3). The effect was particularly noticeable below pH 5.0. All buffers produced similar results, but differences occurred between distilled water and buffers of similar pH, suggesting that factors other thanpH, e.g., the buffer salts, were also acting on zoospores.
DISCUSSION
C. anguiflulae is the most commonly oc- curring endoparasite of soil nematodes (Barron, 1977) and interest has been shown in its efficacy as a biological control agent
TABLE 1
THE EFFECT OF VARIOUS CHILLING PERIODS ON MOTILITY OF Carenaria anguillulae
ZOOSPORES
Length of Percentage chilling motile
(hr) zoosporesa
0 63.6 k 3.8 A 0.25 37.4 -r- 6.3 B 0.5 32.0 + 2.6 B 1 11.3 f 7.4 c 2 0.8 f 0.4 D 4 0 D
’ Means of six replicates *SE. In each column numbers followed by the same letter are not sig- nificantly different (P > 0.05) by Duncan’s multiple range test.
TABLE 2
EFFECT OF pH ON MOTILITY AND GERMINATION OF Catenaria anguillulae ZOOWORES
PH Mo-
tility”
Number of germinated zoospore
Experiment 1 H,O 6.8 Tris-maleate
buffer 7.6 7.3 6.3
Succinate buffer 5.0 4.2 3.8
Experiment 2 HZ0 6.6 Citrate-phosphate
buffer 7.1 6.1 5.6 5.0 4.5 4.0
+ 3,594 k 1,330 CD
+ + +
-
-
+
27,995 k 5,949 A 27.213 + 4,755 A 7.448 k 1,653 B
6,031 2 2,285 BC 26 k 64 D 78 k 131 D
8,671 t 2,160 C
28,437 k 10,734 B 39,480 + 11,356 A 41,277 2 8,242 A 23,327 k 16,153 B 4,029 + 5,183 C 2,915 t 1,080 C
a Presence (+) or absence (-) of motile zoospores 1 hr after inoculation.
b Means of six replicates *SE. In each experiment, numbers followed by same letter are not significantly different (P > 0.05) by Duncan’s multiple range test.
against plant-parasitic nematodes (Sayre and Keeley, 1%9). The dependence of the fungus on actively swimming zoospores for infec- tion suggests its activity would be greatest in moist environments. During culture of R. culicivorax, conditions are ideal for epizo- otics caused by Catenaria, as large numbers of nematode hosts are present in water, and infective propagules can be easily trans- ferred from one nematode generation to the next.
C. anguillulae has generally been re- garded as parasitic on nematodes, although Boosalis and Mankau (1965) suggested that it was largely saprophytic on dead or injured nematodes. Although the Romanomermis isolate of the fungus is capable of growing

CATENARIA IN ROMANOMERIS 353
TABLE 3
EFFECT OF pH ON INFECTIVITY OF Catenaria anguillulae IN Romanomermis culicivorax
PH
Number of infected
nematode@
Experiment 1 Tris-maleate buffer
7.9 7.0 6.1 5.7 5.3
Succinate buffer 5.0 4.7 4.0
Experiment 2 Hz0 6.6 Citrate-phosphate buffer
7.0 6.5 6.0 5.6 5.1 4.6 4.1
4.1 2 2.7 B 8.4 T 2.1 A 0.5 f 0.8 C 0.1 + 0.4c 0.1 -r- 0.4 c
0.0 C 0.0 C 0.0 C
6.6 * 1.2 A
2.3 ” 0.7 B 2.8 ‘- 1.5 B 2.5 k 1.7 B 2.1 f 0.8 B 0.8 k 0.7 C 0.0 C 0.0 C
B Mean number infected out of 10 nematodes *SE (eight replicates). In each experiment, numbers followed by the same letter are not significantly different (P > 0.05) by Duncan’s multiple range test.
saprophytically on dead nematodes, it is also a virulent parasite of R. culicivorax.
Another isolate of C. anguillulae from Hemicycliophora arenaria was unable to infect R. culicivorax (unpubl). These obser- vations support the contention of Sayre and Keeley (1969) that C. anguillulae contains different strains which vary in virulence and suggest that the Romanomermis isolate might be a good strain to test as a biological control agent for plant parasitic nematodes.
The response of C. anguillulae to tem- perature was studied because environmental factors such as temperature influence the initiation and development of fungal diseases of other invertebrates (Bell, 1974; Tanada, 1%3). C. anguillulae grew over an extremely wide range of temperatures (9-42°C) but
zoospore production was largely restricted to temperatures of 21 to 39°C. Previous studies on the effects of temperature on R. culicivorax have shown that it most effec- tively infects mosquitoes at temperatures of 21 to 33°C (Brown and Platzer, 1977) and the growth of the parasitic stages is optimal at 20 to 32°C (Hughes and Platzer, 1977). Con- sequently, it is probably not possible to control epizootics by adjusting the tempera- ture at which cultures are maintained. The low temperatures required to limit zoospore production would undoubtedly reduce the fecundity of the nematode. However, temperatures might be lowered after the nematodes have mated and the majority of eggs have developed.
Chilling is known to induce encystment and reduce germination of zoospores of some species of lower fungi. The swimming period of Phytophthora zoospores was shortened by low temperature (SC) (Ho and Hickman, 1967) and chilling (O-l”(Z) pro- moted encystment of zoospores of Blas- tocladiella (Cantino et al., 1969, Truesdell and Cantino, 1971) and ofRosella allomycis (Held, 1972). The response of C. anguillulae was similar, as encystment of zoospores occurred with as little as 15 min of chilling and all zoospores had encysted after 4 h chilling. Catenaria zoospores are attracted to the body orifices of nematodes (Boosalis and Mankau, 1965) perhaps moving direc- tionally along chemical gradients produced by exudates from the host (Barron, 1977). The premature encystment of zoospores fol- lowing chilling would effectively prevent such directional movement and could be expected to reduce ability of zoospores to locate and infect their hosts. However, chilling time required for complete encyst- ment of zoospore suggested that chilling could not be successfully incorporated into the Romanomermis mass-rearing procedure.
The reduced ability of C. anguillulae to infect nematodes in acid pH was also ob- served by Sayre and Keeley (1%9). Infection of R. culicivorax was completely inhibited below pH 5.0, probably because zoospores

354 STIRLING AND PLATZER
lost motility and the ability to germinate at these pH levels. Low pH had no obvious detrimental effect on the nematodes.
The results reported above indicated that pH adjustment of R. culicivorax cultures might effect contol of C. anguillulae. The p H change was easily incorporated into the mass-rearing technique, and it is now stand- ard procedure to adjust the water used in the cultures to pH 4.5 with acetic acid (Platzer and Stirling, 1978). This change has been in effect for 6 months; infected nematodes have almost been eliminated and the disease is now of minor importance in our mass- rearing system.
ACKNOWLEDGMENTS
We thank Dr. R. Mankau for his initial identification of &terraria, and Dr. G. Stirling, Dr. B. Federici, and Dr. P. Tsao for reviewing the manuscript.
REFERENCES
BARRON, G. L. 1977. The nematode destroying fungi. “Topics in Mycobiology,” No. 1. Canadian Biologi- cal Publications Ltd., Guelph, Ontario.
BELL, J. V. 1974. Mycoses. In “Insect Diseases” (G. E. Cantwell, ed.), Vol. I, Chap. 4. Dekker, New York.
BOOSALIS, M. G., AND MANKALJ, R. 1965. Parasitism and predation of soil microorganisms. In “Ecology of Soil-Borne Plant Pathogens.” (K. F. Baker and W. C. Snyder, eds.), pp. 374-391. Univ. of California Press, Berkeley.
BROWN, B. J., AND PLATZER, E. G. 1977. The effects of temperature on the infectivity of Romanomermis culicivorax. J. Nematol., 9, 166- 172.
CANTINO, E. C., SUBERKROPP, K. F., AND TRUESDELL, L. C. 1969. Form and function in the zoospore of Blastocladiella emersonii: II, Spheroidal mito- chondria and respiration. Nova Hedwigia Z. Krypto- gamenkde., 18, 148- 158.
COUCH, J. N. 1945. Observations on the genus Catenaria. Mycologia, 37, 163-193.
EMERSON, R. 1941. An experimental study of the life cycles and taxonomy of Allomyces. Lloydia, 4, 77- 144.
GOMORI, G. 1955. Preparation of buffers for use in enzyme studies. In “Methods in Enzymology” (C. P. Colowick and N. 0. Kaplan, eds.), Vol. I, pp. 138- 146, Academic Press, New York.
HELD, A. A. 1972. Fungal zoospores are induced to encyst by treatments known to degrade cytoplasmic microtubules. Arch. Mikrobiol., 85, 209-224.
Ho, H. H., AND HICKMAN, C. J. 1967. Asexual repro- duction and behaviour of zoospores ofPhytophthora megasperma var. sojae. Canad. J. Bot., 45, 1963- 1981.
HUGHES, D. S., AND PLATZER, E. G. 1977. Tempera- ture effects on the parasitic phase ofRomanomermis culicivorax in Culex pipens. J. Nematol., 9, 173-175.
PETERSON, J. J., AND WILLIS, 0. R. 1972. Procedures for the mass rearing of a mermithid parasite of mos- quitoes. Mosquito News, 32, 226-230.
PLATZER, E. G., AND STIRLING, A. M. 1978. Improved rearing conditions for Romanomermis culicivorax. Proc. Calif. Mosquito Cont. Ass., 46, in press.
PLATZER, E. G., AND BROWN, B. J. 1976. Physiological ecology ofReesimermis nielseni. In “Proceedings of the 1st International Colloquium on Invertebrate Pathology and IXth Annual Meeting of the Society for Invertebrate Pathology,” Queen’s University at Kingston, Canada. pp. 263-267.
SAYRE, R. M., AND KEELEY, L. S. 1969. Factors influencing Catenaria anguillulae infections in a free- living and a plant parasitic nematode. Nematologica , 15, 492-502.
TANADA, Y. 1963. Epizootiology of infectious diseases. In “Insect Pathology, an Advanced Treatise” (E. A. Steinhaus, ed.), Vol. 2., Chap. 13. Academic Press, New York.
TRUESDELL, L. C., AND CANTINO, E. C. 1971. The introduction and early events of germination in the zoospore of Blastocladiella emersonii. In “Current Topics in Developmental Biology” (A. A. Moscona, and A. Monroy, eds.), Vol. 6, pp. l-44. Academic Press, New York.




















