
Regulation of Pol I-Transcribed 45S rDNA and Pol III-Transcribed 5S rDNA in Arabidopsis
Upload
independentCategory
view
0download
0
Nucleic Acids Research
Oranization and expression of 5S rRNA genes in the parasitic nematode, Brugia malayi
Richard M.Ransohoffl'2 John A.Denkert, Adrienne M.Takacst, Gregory J.Hannont and TimothyW.NilsenI *
'Department of Molecular Biology and Microbiology, Case Western Reserve University School ofMedicine and 2Department of Neurology, Cleveland Clinic Foundation, Cleveland, OH 44106, USA
Received November 22, 1988; Revised and Accepted March 30, 1989 EMBL accession no. X14&K
ABSTRACTDNA sequence analysis of genes encoding 5S rRNA in the human parasitic nematode Brugia malayi(B. malayi) indicates a surprising degree of heterogeneity. This variation in coding sequence is notaccompanied by corresponding heterogeneity in flanking regions which are highly conserved. Sixout of eight potential 5S coding regions differed; of these sequence variants, two were abundantin the B. malayi genome. Direct RNA sequence analysis indicated that one of these abundant variantsaccounts .for most if not all of expressed 5S RNA at two stages of development.
INTRODUCTIONCharacterization of 5S ribosomal RNA genes and transcripts in numerous eukaryoticorganisms has produced a wealth of data about 5S gene structure, evolution and expression.In all cases multigene families encode 5S rRNA and these genes are normally organizedin headtotail arrays of repeat units containing 5S rRNA coding regions interspersed withnontranscribed spacer DNA (Reviewed in 1). There is considerable diversity with respectto the detailed organization of 5S genes between organisms including the variable occurrenceof nontranscribed 'pseudogene' sequences (Reviewed in 1).
Typically, 5S coding regions exhibit limited heterogeneity, while a variable degree ofheterogeneity of noncoding spacer DNA is tolerated (2,3, reviewed in 6). For example,in the human parasite Plasmodiafalciparum, the three 5S rRNA genes (the smallest reportedgenomic complement) are identical, while spacer DNA is divergent, as determined bySouthern blot hybridization studies (5). Similarly, in both the newt Notophthalmosviridescens, and in Trypanosoma brucei, sequence comparison of several individual repeatsdemonstrated homogeneity of 5S coding regions (4,7,8). A detailed study of 5S genesin Drosophila melanogaster revealed only three variations in coding sequence in 23 copies(3). In Xenopus laevis, sequence heterogeneity of both 5S-coding and of intergenic regionswithin clusters of 5S genes is limited (2,9) despite the presence of three families ofdevelopmentally-regulated 5S genes.The detailed organization of 5S genes in nematodes has been addressed only in C. elegans
(10). Typical of other eukaryotes, Southern blot hybridization analysis revealed 5S codingregions to be homogeneous (10). Here, we describe the organization and expression of5S genes in the nematode B. malayi. In contrast to C. elegans, 5S coding sequences werefound to be quite heterogeneous in that six out of eight sequenced genes were different.Surprisingly, this sequence heterogeneity was largely confined to coding regions whileintergenic sequences were highly conserved. Two of the 5S coding sequence variants wereabundantly represented in the B. malayi genome. Primer extension analysis of B. malayi
( IRL Press
Volume 17 Number 10 1989
3773

Nucleic Acids Research
RNA with diagnostic oligodeoxynucleotides indicated that, at most, a subset of the potential5S coding regions were expressed. Sequencing of 5S rRNA in both microfilariae and adultworms established that a predominant 5S species was expressed in these two developmentalstages.
MATERIALS AND METHODSB. malayi nucleic acids.B. malayi microfilariae were obtained by peritoneal lavage of infected jirds, as described(11). Adult parasites were obtained from TRS Laboratories (Athens, GA). RNA wasprepared by the guanidinium hot phenol method, as described (1 1). Total cellular DNAwas prepared by lysis in guanidinium thiocyanate buffer followed by phenol extraction.After initial ethanol precipitation, DNA was digested with RNase A and proteinase K beforereprecipitation, as described (12).Sequence analysis.DNA fragments were subcloned into pBS M13+ (Stratagene) as described in the text.Plasmid DNA was prepared as described and both strands were sequenced by thedideoxynucleotide chain termination method (13,14).RNA sequence was determined by primer extension analysis in the presence of
dideoxynucleotides (15).Primer extension analysis.Oligodeoxynucleotide primers complementary to variant sequences present in cloned 5Sgenes of B. malayi were terminally labelled, using [F32P]ATP and T4 polynucleotidekinase. Labelled oligodeoxynucleotides as indicated in figure legends were annealed withB. malani RNA at 550 C in 125 mM KCI, 10 mM Tris, pH 8.3. Extensions were performedas described (16) with the exception that the reactions were carried out at 550 C. Reactionproducts were fractionated on 7 M urea/5 % polyacrylamide gels.
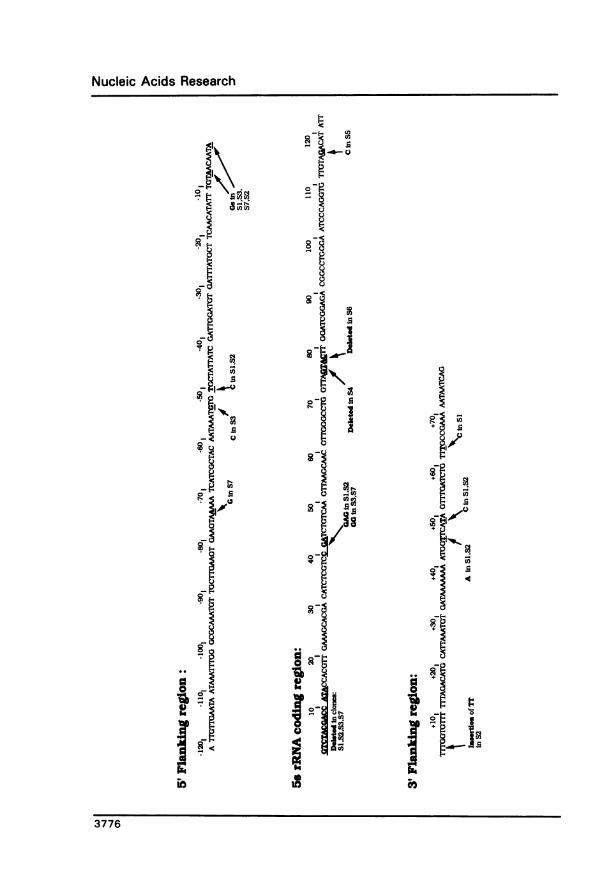
RESULTSNucleotide sequence analysis of subclones containing SS rRNA sequences.We recently reported the isolation and characterization of a genomic clone (designatedclone 8), containing a cluster of B. malayi 5S rRNA genes (16). Clone 8 was initiallyselected by hybridization to two synthetic oligodeoxynucleotides, one complementary toa 22-base spliced leader found on a subset of B. malayi mRNAs and one complementaryto C. elegans 5S rRNA (10).Sequence analysis of three subclones of genomic clone 8 indicated that potential 5S coding
regions were interspersed with the 22-nucleotide spliced leader sequence (Figure 1) (16).During the course of these studies, it became clear that 5S coding regions contained withinthese subclones varied considerably. To investigate this heterogeneity further, we preparedsubclones containing complete 5S rRNA coding regions by subcloning a population of385-bp Xmn I/Pvu II fragments derived from clone 8.Sequence analysis of eight subclones revealed six unique variants of 5S rRNA coding
sequence (Figure 2). Two major patterns of sequence variation were observed:1. S'deleted clones. Four clones, designated S1, S2, S3 and S7, exhibited deletions of
thirteen base pairs (bp) at or near the 5' end of 5S rRNA coding sequence. This interpretationwas based on alignment with putative 5S coding regions and flanking sequences in theremaining four B. malayi 5S genes. Because of sequence redundancy in this region, itwas not possible to differentiate whether this deletion comprised bp + 1-+ 13 or -7- +6.
3774

Nucleic Acids Research
8K >' '
8C~i jL*_s
8D~ 'LIi5f Ddeted
Figure 1. Interspersion of SS rRNA genes with genes encoding a spliced leader sequence in B. malayi.The 13.5 kb genomic insert of lambda recombinant clone 8 (16) was digested with restriction endonuclease ScaI which recognizes a sequence within the SS rRNA coding region, and several fragments which hybridized tothe 22-nucleotide sequence of the spliced leader were subcloned into pBS M13+. The nucleotide sequence ofboth strands was determined by a combination of chemical degradation and chain termination methods (14, 25).Portions of potential 5S rRNA coding sequences are indicated by filled arrows, with arrowheads showingtranscriptional orientation. In 5S coding regions designated S4 and S6, the Sca I recognition site is elimiinatedby different 2-base deletions (see Figure 2). The 5S rRNA potential coding sequence designated 5'-deleted containsa 13base deletion (see Figure 2) which eliminates an Acc I recognition site. Arrows designated L correspondto potentially transcribed regions containing the 22-nucleotide spliced exon and are drawn to reflect the 109 nucleotideRNA transcript (16). The position of the 22-nucleotide leader within this region is indicated by a bar above thearrow. A: Acc I; C: Cla I; H: Hpa I; P: Pvu II; S: Sca I; X: Xmn I.
While it was possible that insertion of 13 bp in B. malayi 5S rRNA sequences could alsogive rise to the sequence differences which we attribute to deletion within these genes,this interpretation seems unlikely, since the putative inserted sequence matches 5S rRNAof C. elegans at 11/13 positions, while the 5S rRNA predicted to pre!date such an insertionmatches C. elegans 5S rRNA at only 7/13 positions (10).
In addition to this 5' deletion, these four clones contained base substitutions at positions+40-+42. In clones S3 and S7, two of three bp at +40-+42 were altered, and in clonesSi and S2, all three bp were altered (Figure 2). Clones Si and S2 were identical in SScoding regions, but these clones could be distinguished by a point mutation within 3' flankingsequences. Furthermore, the potential RNA polymerase mI termination signal in clone S2contained seven thymidine residues, while in all other clones it was five residues (17) (Figure2). Clones S3 and S7 were also identical in coding sequences, but could be differentiatedby two different single base substitutions in 5' flanking regions (Figure 2).
2. 5'intact clones. Four clones, S4, S5, S6 and S8 contained neither of the sequencevariations described above. Clone S5 contained a single base substitution at +116, bycomparison with all other clones (Figure 2). In two clones, S4 and S6, different two bpdeletions were present, each of which abolished a recognition site for the restrictionendonuclease Sca I present at positions +74-+79. In clone S4, a recognition site forthe restriction endonuclease Hpa I was generated by the deletion of two residues at bases+75-+76 (Figure 2).In contrast to the heterogeneity of potential 5S rRNA coding regions, flanking sequences
were relatively conserved. Whereas the four 5'-intact clones (S4, S5, S6 and S8) all haddifferent coding sequences, their 5' and 3' flanking sequences were identical. With respectto the 5'-deleted clones, Si and 52 had three scattered base substitutions in common withinthe first 121 bases of the 5'-flanking region. Clones S3 and S7 also displayed limitedheterogeneity within 5'-flanking regions (Figure 2).
3775
Nucleic Acids Research
2
I
_o
(_
si
$i
_0 t
8_ §
*a
}r-C
1 _
_ r
+ S
I
+
1S
+3a
+ 4
* CSRSX - o
X o s85~~~vg~~~~~~~.
3776
_U
4;:
_2
j/IIc° 0
Nucleic Acids Research
A B1 2 3 23 4
| w ^ ^ 630{550500
* -110
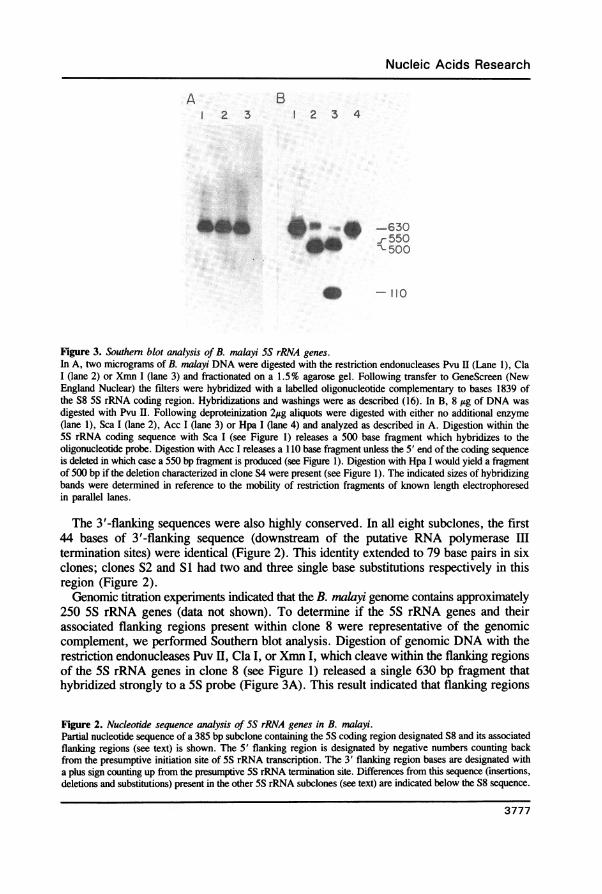
Figure 3. Southern blot analysis of B. malayi 5S rRNA genes.In A, two micrograms of B. malayi DNA were digested with the restriction endonucleases Pvu II (Lane 1), ClaI (lane 2) or Xmn I (lane 3) and fractionated on a 1.5% agarose gel. Following transfer to GeneScreen (NewEngland Nuclear) the filters were hybridized with a labelled oligonucleotide complementary to bases 1839 ofthe S8 5S rRNA coding region. Hybridizations and washings were as described (16). In B, 8 jig of DNA wasdigested with Pvu II. Following deproteinization 2ug aliquots were digested with either no additional enzyme(lane 1), Sca I (lane 2), Acc I (lane 3) or Hpa I (lane 4) and analyzed as described in A. Digestion within the5S rRNA coding sequence with Sca I (see Figure 1) releases a 500 base fragment which hybridizes to theoligonucleotide probe. Digestion with Acc I releases a 110 base fragment unless the 5' end of the coding sequenceis deleted in which case a 550 bp fragment is produced (see Figure 1). Digestion with Hpa I would yield a fragmentof 500 bp if the deletion characterized in clone S4 were present (see Figure 1). The indicated sizes of hybridizingbands were determined in reference to the mobility of restriction fragments of known length electrophoresedin parallel lanes.
The 3'-flanking sequences were also highly conserved. In all eight subclones, the first44 bases of 3'-flanking sequence (downstream of the putative RNA polymerase IIItermination sites) were identical (Figure 2). This identity extended to 79 base pairs in sixclones; clones S2 and SI had two and three single base substitutions respectively in thisregion (Figure 2).Genomic titration experiments indicated that the B. malayi genome contains approximately
250 5S rRNA genes (data not shown). To determine if the 5S rRNA genes and theirassociated flanking regions present within clone 8 were representative of the genomiccomplement, we performed Southern blot analysis. Digestion of genomic DNA with therestriction endonucleases Puv HI, Cla I, or Xmn I, which cleave within the flanking regionsof the 5S rRNA genes in clone 8 (see Figure 1) released a single 630 bp fragment thathybridized strongly to a 5S probe (Figure 3A). This result indicated that flanking regions
Figure 2. Nucleotide sequence analysis of 5S rRNA genes in B. malayi.Partial nucleotide sequence of a 385 bp subclone containing the 5S coding region designated S8 and its associatedflanking regions (see text) is shown. The 5' flanking region is designated by negative numbers counting backfrom the presumptive initiation site of 5S rRNA transcription. The 3' flanking region bases are designated witha plus sign counting up from the presumptive 5S rRNA termination site. Differences from this sequence (insertions,deletions and substitutions) present in the other 5S rRNA subclones (see text) are indicated below the S8 sequence.
3777
Nucleic Acids Research
..... ...
*" f*1t'5SRNA..*.....5S RNA
.A.fji 78,
92. nt C58 ni j-
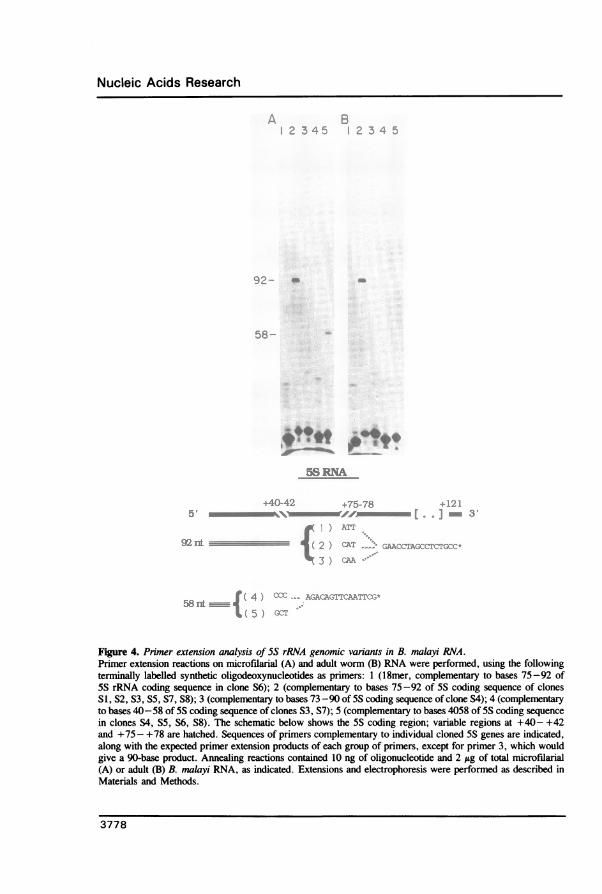
Figure 4. Primer extension analysis of 5S rRNA genomic variants in B. malayi RNA.Primer extension reactions on microfilarial (A) and adult worm (B) RNA were performed, using the followingterminally labelled synthetic oligodeoxynucleotides as primers: 1 (18mer, complementary to bases 75-92 of5S rRNA coding sequence in clone S6); 2 (complementary to bases 75-92 of 5S coding sequence of clonesSI, S2, S3, S5, S7, S8); 3 (complementary to bases 73 -90 of 5S coding sequence of clone S4); 4 (complementaryto bases 40-58 of 5S coding sequence of clones S3, S7); 5 (complementary to bases 4058 of 5S coding sequence
in clones S4, S5, S6, S8). The schematic below shows the 5S coding region; variable regions at +40- +42and +75 - +78 are hatched. Sequences of primers complementary to individual cloned 5S genes are indicated,along with the expected primer extension products of each group of primers, except for primer 3, which wouldgive a 90-base product. Annealing reactions contained 10 ng of oligonucleotide and 2 jg of total microfilarial(A) or adult (B) B. malayi RNA, as indicated. Extensions and electrophoresis were performed as described inMaterials and Methods.
3778
...... " 'l-
Nucleic Acids Research
A BG AUCO
5-a. 5-10- 10-20- * 20-30- 30-
40- 40-
50- Z 50-
60- 60-
4.
YY
70- 70-
4..
S
80- 4. 80-
90- 90-
GAUCO
-4'--a
'w4
-a
._
_
_,
_k
S5/S8 sequence:
5 10 20 30 40 50 60 70 80 90
GTCTACGACCATACCACGTTGAAAGCACGACATCTCGTCCGATCTGTCAAGTTAAGCAACGTTGGGCCTGGTTAGTACTTGGATCGGAGACGG
A: ACGACCAUACCACGUUGAAAGCACGACAUCUCGUCCGAUCUGUCAAGUUAAGCAACGUUGGGCCUGGUUAGUACUUGGAUCGGAGACGG
B: ACGACCAUACCACGUUGAAAGCACGACAUCUCGUCCGAUCUGUCAAGUUAAGCAACGUUGGGCCUGGUUAGUACUUGGAUCGGAGACGG
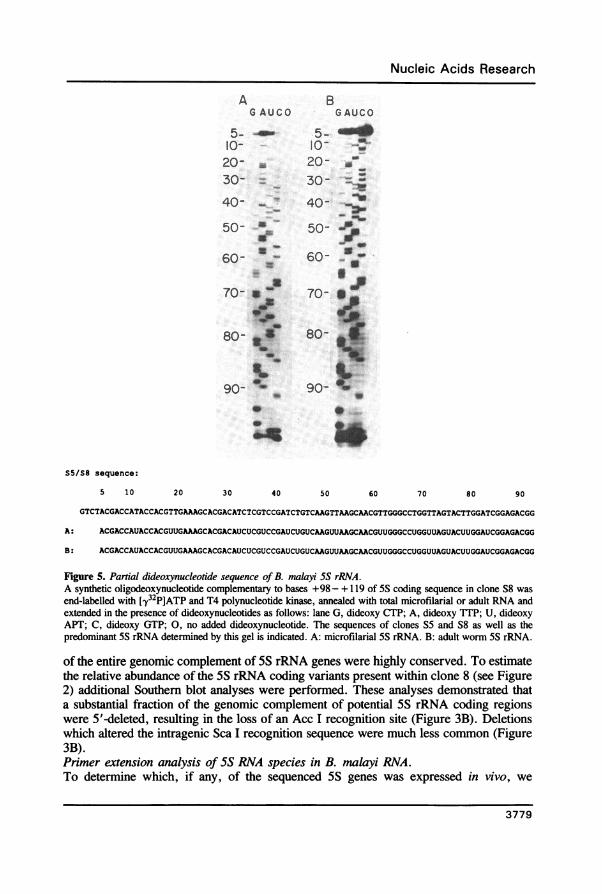
Figure 5. Partial dideoxynucleotide sequence of B. malayi 5S rRNA.A synthetic oligodeoxynucleotide complementary to bases +98- + 119 of 5S coding sequence in clone S8 wasend-labelled with ['y32P]ATP and T4 polynucleotide kinase, annealed with total microfilarial or adult RNA andextended in the presence of dideoxynucleotides as follows: lane G, dideoxy CTP; A, dideoxy TTP; U, dideoxyAPT; C, dideoxy GTP; 0, no added dideoxynucleotide. The sequences of clones S5 and S8 as well as thepredominant 5S rRNA determined by this gel is indicated. A: microfilarial 5S rRNA. B: adult worm 5S rRNA.
of the entire genomic complement of 5S rRNA genes were highly conserved. To estimatethe relative abundance of the 5S rRNA coding variants present within clone 8 (see Figure2) additional Southern blot analyses were performed. These analyses demonstrated thata substantial fraction of the genomic complement of potential 5S rRNA coding regionswere 5'-deleted, resulting in the loss of an Acc I recognition site (Figure 3B). Deletionswhich altered the intragenic Sca I recognition sequence were much less common (Figure3B).Primer extension analysis of SS RNA species in B. malayi RNA.To determine which, if any, of the sequenced 5S genes was expressed in vivo, we
3779

Nucleic Acids Research
constructed diagnostic oligodeoxynucleotides for primer extension assays. We first showedthat such assays could distinguish among the predicted transcription products of genescontaining varying sequences at positions +40- +42 and +75 - +78 by performing primerextensions on in vitro transcripts synthesized with T3 or T7 RNA polymerase (data notshown). When similar experiments were performed with B. malayi RNA, no primerextension product was observed using primers designed to detect SS RNAs transcribedfrom the potential coding regions of clones SI, S2 S3, S4, S6 and S7 (Figure 4). Extensionproducts of expected length were observed when primers designed to detect transcriptsfrom clones S5 or S8 were used (Figure 4). No differences between microfilarial and adult5S rRNA were detected (Figure 4).This analysis demonstrated that a restricted set of 5S RNA genes was expressed and
that the expressed genes contained sequences at +40- +42 and +75 - +78 correspondingto S5 and S8.Sequence of the predominant 5S rRNA species in B. malayi RNA.It remained possible that the gene from which 5S rRNA was expressed might differ fromS5 and S8 elsewhere in the coding region. To address this question, primer-extensionsequencing was performed using a synthetic 22-base oligodeoxynucleotide complementaryto bases +98-+ 119 of 5S rRNA. This primer was used since seven of eight sequencedSS variants were identical within this region. While clone S5 had a single base substitutionat + 116, it was also recognized by this primer (see Figure 2). Using the dideoxynucleotidechain termination method, we sequenced 5S RNA from microfilariae and adult worms.The sequence of bases +5 - +93 was unambiguously determined in this analysis, and itcorresponded exactly to the sequence predicted to be present in transcripts of clones S5and S8 (Figure 5). In these base positions, the sequence of 5S rRNA in adult worms andmicrofilariae is identical.
DISCUSSIONWe have used a combination ofDNA and RNA sequence analysis to address the organizationand expression of 5S rRNA genes in the human parasitic nematode, B. malayi. An unusualfeature of the 5S genes of B. malayi is the heterogeneity of coding sequences, which contraststo the relative conservation of intergenic sequences. We observed six unique variants of5S coding sequence among eight clones. In 121 bp of 5'-flanking DNA, only scatteredindividual base substitutions were present in four of the eight clones; similarly, the first44 bp of 3'-flanking DNA was entirely conserved. In previous studies of 5S genes innumerous organisms, variation within 5S coding regions was less than or equal toheterogeneity within flanking sequences (210).
It is possible that the overall conservation of sequences flanking 5S coding regions inB. malayi is a consequence of the presence of interspersed genes encoding the 109-basenonadenylated RNA which is the likely donor of a 22-base transspliced leader found onsome mRNAs (16). However, it should be emphasized that 5S subclones described in Figure2 do not contain the 109 bases which comprise the spliced-leader transcript gene.
Similar interspersion of 5S rRNA genes with a gene encoding a transspliced leader se-quence was described in the nematode C. elegans, although in this organism 5S codingsequences appear to be homogeneous (10, 18). Additionally, 5S genes are interspersedwith histone genes in the brine shrimp Artemia (19,20). Although the intergenic sequencesof 5S gene clusters have not been analyzed in detail in Artemia and C. elegans, it willbe of interest to determine if flanking-sequence conservation proves to be characteristicof 5S repeat units where other interspersed genes are present.
3780

Nucleic Acids Research
Several observations indicate that the genomically-abundant 5'-deleted 5S genes are notexpressed in vivo. In primer extension assays, we did not detect transcripts containingvariant sequences at +40- +42, which were invariably associated with the 5'-deleted geneswe characterized. Furthermore, the RNA sequence data argues strongly that 5'-deletedgenes are not expressed. Initiation of transcription within sequences upstream of the pointof divergence between 5'-deleted genes and the predominant expressed species wouldgenerate truncated 5S RNAs or base substitutions at as many as six of the initial thirteenresidues. The RNA sequence analysis could have detected four of these base substitutionsand none were present. Finally, stops suggestive of truncated species were not observed.
It is also apparent that the minority genomic species containing heterogeneity in therecognition sequence for Sca I were not expressed to any appreciable level, since sequence5' of the 2-base deletion present at +75-76 (S4) or +77-78 (S6) would appear out ofregister on the autoradiogram of the sequencing ladder. Additionally, primer extensionproducts predicted by these sequences were not detected.
In view of the conservation of flanking regions in these eight clones, it is possible thatthe failure of 5'-deleted genes to be expressed is attributable to the 5S codingsequencevariation. 5S RNA transcription initiation in Xenopus is directed primarily by an intergenicpromoter termed the internal control region (ICR), which corresponds to positions+50- +97 (Reviewed in 21). It appears possible that efficient transcription of 5S genesin B. malayi requires additional sequence elements, since regions homologous to the ICRare identical in expressed and silent 5S genes. This suggestion is not without precedent,since Morton and Sprague (22) observed that deletion of 5' flanking DNA to -9 in 5SrRNA genes in Bombyx mori silkworms abolished transcription in homologous extracts.Furthermore, we have utilized a cellfree extract derived from eggs of the parasitic nematodeAscaris suum to assess the transcriptional competence of various cloned B. malayi 5S genes.In these experiments, 5'-deleted clones were transcriptionally inert, whereas 5'-intact cloneswere efficiently transcribed (26). Therefore, we think it likely that the unexpressed 5Sgenes in B. malayi are not transcribed, although it is formally possible that their transcriptionresults in the synthesis of an unstable product. The maintenance of multiple copies of inactive5S variant genes (pseudogenes) has previously been described in both Xenopus and D.melanogaster (2,23). It has been speculated that these sequences may function to sequestertranscription factors (24). Conserved intragenic promoter sequences in inactive 5 '-deleted5S genes of B. malayi could provide a similar function.
ACKNOWLEDGEMENTSWe thank Drs. R. Davis, P.A. Maroney and D. Setzer for helpful discussions. Thisinvestigation was supported by a grant from the John D. and Catherine T. MacArthurFoundation, a Presidential Young Investigator Award of the NSF (to TWN) and a JuniorFaculty Award of the National Multiple Sclerosis Society (to RMR).
*To whom correspondence should be addressed
REFE:RENCES1. Long, E.O. and Dawid, I.B. (1980). Ann. Rev. Biochem. 49, 727-764.2. Fedoroff, N.V. and Brown, D.D. (1977). Cold Spring Harbor Symp. Quant. Biol. 42, 1195-1200.3. Tschudi, C. and Pirrotta, V. (1980). Nucleic Acids Res. 8, 441-451.4. Kay, B.K. and Gall, J.G. (1981). Nucleic Acids Res. 9, 6457-6469.5. ShippenLentz, D.E. and Vezza, A.C. (1988). Mol. Biochem. Parasitol. 27, 263-274.6. Fedoroff, N.V. (1979). Cell 16, 697-710.
3781

Nucleic Acids Research
7. Cordingley, J.S. (1985). Mol. Biochem. Parasitol. 17,321-330.8. Lenardo, M.J., Dorfman, D.M., Reddy, L.V. and Donelson, J.E. (1985). Gene 35, 131-141.9. Peterson, R.C., Doering, J.L. and Brown, D.D. (1980). Cell 20, 131-141.10. Nelson, D.W. and Honda, B.M. (1985). Gene 38, 245-251.11. Nilsen, T.W., Maroney, P.A., Goodwin, R.G., Perrine, K.G., Denker, J.A., Nanduri, J. and Kazura, J.W.
(1988). Proc. Natl. Acad. Sci. USA 85, 3604-3607.12. Perrine, K.G., Denker, J.A. and Nilsen, T.W. (1988). Mol. Biochem. Parasitol. 30, 97-104.13. Hattori, M. and Sakaki, Y. (1986). Anal. Biochem. 152, 232-238.14. Sanger, F.S., Nicklen, S. and Coulson, A.R. (1977). Proc. Natl. Acad. Sci. USA 74, 5463-5467.15. Davis, R.E., Davis, A.H., Carroll, S., Rajkovic, A. and Rottman, F.M. (1988). Mol. Cell. Biol. 8,
4745-4755.16. Takacs, A.M., Denker, J.A., Perrine, K.G., Maroney, P.A. and Nilsen, T.W. (1988). Proc. Natl. Acad.
Sci. USA 85, 7932-7936.17. Bogenhagen, D.F. and Brown, D.D. (1981). Cell 24, 261-270.18. Krause, M. and Hirsch, D. (1987). Cell 49, 753-761.19. Andres, M.T., Vaughn, J.C., Perry, B.A. and Bagshaw, J.C. (1987). Gene 51, 61-67.20. Bagshaw, J.C., Skinner, H.B., Burn, T.C. and Perry, B.A. (1987). Nucleic Acids Res. 15, 3628.21. Geiduschek, E.P. and TocchiniValentini, G.P. (1988). Ann. Rev. Biochem. 57, 873-914.22. Morton, D.G. and Sprague, K.U. (1984). Proc. Natl. Acad. Sci. USA 81, 5519-5522.23. Sharp, S., Garcia, A., Cooley, L. and Soll, D. (1984). Nucleic Acids Res. 12, 7617-7632.24. Emerson, B.M. and Roeder, R.G. (1984). J. Biol. Chem. 259, 7916-7925.25. Maxam, A. and Gilbert, W. (1980). Methods Enzymol. 65, 449-560.26. Maroney, P.A., Hannon, G.J. and Nilsen, T.W. (1989). Molec. Biochem. Parasit (in press).
3782
This article, submitted on disc, has been automaticallyconverted into this typeset format by the publisher.
Copyright © 2022 FDOKUMEN

















![Pulmonary Inflammation Induced by a Recombinant Brugia malayi [gamma]-glutamyl transpeptidase Homolog: Involvement of Humoral Autoimmune Responses](https://static.fdokumen.com/doc/165x107/631e10e40ff042c6110c2b14/pulmonary-inflammation-induced-by-a-recombinant-brugia-malayi-gamma-glutamyl-transpeptidase.jpg)


