
Differential requirements for basic amino acids in transcription factor IIIA-5S gene interaction
9
Di¡erential requirements for basic amino acids in transcription factor IIIA-5S gene interaction Jay S. Hanas *, Gerald Koelsch, Rodney Moreland, J.Q. Wickham Department of Biochemistry and Molecular Biology, University of Oklahoma College of Medicine, 940 Stanton Young Blvd., Oklahoma City, OK 73140, USA Received 21 October 1997; accepted 1 April 1998 Abstract Basic amino acids Arg, Lys, and His in the Cys 2 His 2 zinc fingers of transcription factor IIIA (TFIIIA) potentially have important roles in factor binding to the extended internal control region (ICR) of the 5S ribosomal gene. Conserved and non- conserved basic residues in the N-terminal fingers I, II, III and the more C-terminal fingers V and IX were analyzed by site- directed mutagenesis and DNase I protection in order to assess their individual requirement in the DNA-binding mechanism. In the DNA recognition helix of finger II, the conserved Arg at position 62 (N-terminal side of the first zinc-coordinating histidine) was changed to a Leu or Gln. Both the R62L and R62Q mutations inhibited Xenopus TFIIIA-dependent DNase I footprinting along the entire 5S gene ICR. When His-58 (non-conserved basic residue with DNA-binding potential in the same helical region) was changed to a Gln, the mutated protein was able to protect the ICR from DNase I digestion. Therefore, Arg-62 is individually required for TFIIIA binding over the entire ICR whereas His-58 is not. Fingers V and IX have conserved Arg residues in positions identical to Arg-62 in finger II (Arg-154 in finger V and Arg-271 in finger IX). When these residues were changed to Leu and Ile respectively, TFIIIA-dependent DNase I protection was observed along the entire 5S gene ICR. These results indicate differing DNA-binding mechanisms by the N-terminal fingers versus the C-terminal fingers at the level of individual amino acid-nucleotide interactions. In the N-terminal finger I, the conserved Lys at position 11 outside the recognition helix and a conserved hydrophobic Trp at position 28 within the helix were changed to an Ala and Ser respectively. The K11A change inhibited TFIIIA-dependent DNase I protection to a much greater extent than the W28S change. ß 1998 Elsevier Science B.V. All rights reserved. Keywords : Protein-DNA interaction; Zinc ¢nger; 5S ribosomal gene 1. Introduction Transcription factor IIIA (TFIIIA) regulates eu- karyotic 5S ribosomal RNA synthesis and ribosome biogenesis through its interaction with the 5S riboso- mal gene [1]. TFIIIA isolated from Xenopus laevis oocytes was the ¢rst cysteine-rich regulatory protein discovered to contain zinc and require the metal for function [2]. The amino acid sequence of this protein revealed repetitive Cys 2 His 2 zinc ¢nger domains [3,4]. Zinc-containing regulatory proteins are now known to be responsible for a multitude of transcription and signal transduction events in eukaryotic cells [5]. 0167-4781 / 98 / $19.00 ß 1998 Elsevier Science B.V. All rights reserved. PII:S0167-4781(98)00070-0 Abbreviations : DTT, dithiothreitol ; ICR, internal control re- gion ; PAGE, polyacrylamide gel electrophoresis ; PCR, polymer- ase chain reaction ; PMSF, phenylmethylsulfonyl £uoride ; TFIIIA, transcription factor IIIA * Corresponding author. Fax: +1 (405) 2713092; E-mail : [email protected] Biochimica et Biophysica Acta 1398 (1998) 256^264
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Differential requirements for basic amino acids in transcription factor IIIA-5S gene interaction

Di¡erential requirements for basic amino acids in transcription factorIIIA-5S gene interaction
Jay S. Hanas *, Gerald Koelsch, Rodney Moreland, J.Q. WickhamDepartment of Biochemistry and Molecular Biology, University of Oklahoma College of Medicine, 940 Stanton Young Blvd.,
Oklahoma City, OK 73140, USA
Received 21 October 1997; accepted 1 April 1998
Abstract
Basic amino acids Arg, Lys, and His in the Cys2His2 zinc fingers of transcription factor IIIA (TFIIIA) potentially haveimportant roles in factor binding to the extended internal control region (ICR) of the 5S ribosomal gene. Conserved and non-conserved basic residues in the N-terminal fingers I, II, III and the more C-terminal fingers V and IX were analyzed by site-directed mutagenesis and DNase I protection in order to assess their individual requirement in the DNA-binding mechanism.In the DNA recognition helix of finger II, the conserved Arg at position 62 (N-terminal side of the first zinc-coordinatinghistidine) was changed to a Leu or Gln. Both the R62L and R62Q mutations inhibited Xenopus TFIIIA-dependent DNase Ifootprinting along the entire 5S gene ICR. When His-58 (non-conserved basic residue with DNA-binding potential in thesame helical region) was changed to a Gln, the mutated protein was able to protect the ICR from DNase I digestion.Therefore, Arg-62 is individually required for TFIIIA binding over the entire ICR whereas His-58 is not. Fingers V and IXhave conserved Arg residues in positions identical to Arg-62 in finger II (Arg-154 in finger V and Arg-271 in finger IX). Whenthese residues were changed to Leu and Ile respectively, TFIIIA-dependent DNase I protection was observed along the entire5S gene ICR. These results indicate differing DNA-binding mechanisms by the N-terminal fingers versus the C-terminalfingers at the level of individual amino acid-nucleotide interactions. In the N-terminal finger I, the conserved Lys at position11 outside the recognition helix and a conserved hydrophobic Trp at position 28 within the helix were changed to an Ala andSer respectively. The K11A change inhibited TFIIIA-dependent DNase I protection to a much greater extent than the W28Schange. ß 1998 Elsevier Science B.V. All rights reserved.
Keywords: Protein-DNA interaction; Zinc ¢nger; 5S ribosomal gene
1. Introduction
Transcription factor IIIA (TFIIIA) regulates eu-
karyotic 5S ribosomal RNA synthesis and ribosomebiogenesis through its interaction with the 5S riboso-mal gene [1]. TFIIIA isolated from Xenopus laevisoocytes was the ¢rst cysteine-rich regulatory proteindiscovered to contain zinc and require the metal forfunction [2]. The amino acid sequence of this proteinrevealed repetitive Cys2His2 zinc ¢nger domains [3,4].Zinc-containing regulatory proteins are now knownto be responsible for a multitude of transcription andsignal transduction events in eukaryotic cells [5].
0167-4781 / 98 / $19.00 ß 1998 Elsevier Science B.V. All rights reserved.PII: S 0 1 6 7 - 4 7 8 1 ( 9 8 ) 0 0 0 7 0 - 0
Abbreviations: DTT, dithiothreitol ; ICR, internal control re-gion; PAGE, polyacrylamide gel electrophoresis ; PCR, polymer-ase chain reaction; PMSF, phenylmethylsulfonyl £uoride;TFIIIA, transcription factor IIIA
* Corresponding author. Fax: +1 (405) 2713092;E-mail : [email protected]
BBAEXP 93115 30-6-98
Biochimica et Biophysica Acta 1398 (1998) 256^264

TFIIIA-like zinc ¢nger proteins (those containingCys2His2 metal-binding domains) number over athousand in vertebrates and comprise the largest pro-tein superfamily [5^7]. Individual proteins in thissuperfamily di¡er in ¢nger number as well as nucleo-tide sequence recognition. Vertebrate TFIIIAs con-tain nine contiguous Cys2His2 zinc-binding domainsand a non-¢nger C-terminal tail [8^11]. The zinc ¢n-gers bind to the large internal control region (ICR,nucleotides +43 to +96) of the 5S gene as well as 5SRNA [1,12,13]. Because the structure of TFIIIA isrepresentative of the largest protein superfamily, it isimportant to understand the molecular details ofhow such ¢ngers bind to extended regions of DNA.Finger deletion analysis of Xenopus TFIIIA indicatedthat the nine ¢ngers bind to the 5S gene in threegroups with the N-terminal three ¢ngers interactingwith the C-box of the ICR, the middle group of¢ngers binding the intermediate element (M [14]),and the C-terminal ¢ngers binding the ICR A-box[15,16]. DNA binding by the N-terminal ¢ngers stim-ulates binding of the more C-terminal ¢ngers where-as 5S RNA binding is not mediated by the N-termi-nal ¢ngers but by the middle and C-terminal ¢ngers[15^18].
The N-terminal three ¢ngers of TFIIIA contributesigni¢cantly to the overall interaction energy with the5S gene ICR [19^24]. Remarkably, these ¢ngers ofclosely related Xenopus, Rana, and Bufo frogTFIIIAs share only about 60% sequence identityyet the nucleotide sequences they bind (C-box in 5Sgene) are 100% conserved [9]. Overall, XenopusTFIIIA shares about 55% amino acid identity withhuman TFIIIA, about 40% identity with ¢shTFIIIA, and about 20% identity with yeast TFIIIA[10,11,25,26].
The ¢sh, amphibian, and human TFIIIA zinc ¢n-gers contain a number of conserved and non-con-served basic residues with nucleic acid binding poten-tial [11]. Alignment of the conserved basic residues invertebrae TFIIIAs with conserved guanine bases inthe corresponding species-speci¢c ICRs suggestedhow the N-terminal, middle, and C-terminal groupsof ¢ngers contact the C-box, M element, and A-boxrespectively [11]. With respect to the N-terminalgroup, many of the basic amino acid-guanine con-tacts predicted from this sequence alignment werealso identi¢ed in a NMR solution study of these
¢ngers bound to the 5S gene C-box [27]. This con-served sequence alignment predicted binding sitesgreater than 3 bp for ¢ngers I and III and a novelstructure in ¢nger I where the recognition helix in-teracts with both strands of DNA via a Lys-26 con-tact with guanine 91 on the ICR coding strand, all ofwhich were observed in the NMR structure [27].
Although the NMR structure identi¢ed most ofthe amino acid-nucleotide contacts between TFIIIA¢ngers and the 5S gene C-box, the analysis did notresolve contacts for basic amino acids His-58 and/orHis-59 in the recognition helix of ¢nger II nor exam-ine contacts of the more C-terminal ¢ngers [27]. Inaddition, the NMR structure does not assess the re-quirement for individual contacts in the overall bind-ing mechanism. In the present study, conserved andnon-conserved basic amino acids in Xenopus TFIIIA¢ngers I, II, III, V, and IX are targeted for muta-genesis and DNase I footprint analyses to elucidatetheir requirement in TFIIIA binding to the 5S geneand whether ¢nger-binding mechanisms at the levelof individual contacts are similar between the N-ter-minal and more C-terminal ¢ngers.
2. Materials and methods
2.1. Site-directed mutagenesis of X. laevis TFIIIAcDNA
Arg-62, Arg-154, and Arg-271 mutations wereconstructed using the mutagenesis method of Eck-stein and coworkers [28] with reagents provided byAmersham. 5P-Phosphorylated oligonucleotides (20-mers) with the appropriate codon changes were an-nealed to M13mp18 single-stranded DNA containingthe EcoRI-SphI cDNA fragment coding for the N-terminal half or C-terminal half of X. laevis TFIIIA.A primer-extension reaction was performed withKlenow fragment, dNTPs, and [K-S]dCTP followedby ligation with T4 DNA ligase. Double-strandedcircles were nicked with NciI and the wild-typestrand was partially digested with exonuclease III.Double-stranded DNA was resynthesized withDNA polymerase I and dNTPs, religated, and elec-troporated into Escherichia coli TG1 cells. Trans-formed phage DNAs were screened for the mutationsby dideoxy sequencing [29]. Once a mutant clone was
BBAEXP 93115 30-6-98
J.S. Hanas et al. / Biochimica et Biophysica Acta 1398 (1998) 256^264 257

identi¢ed, the entire EcoRI-SphI non-coding strandwas sequenced to con¢rm the lack of spurious muta-tions. The K11A, W28S, H58Q, K92L, and vF94changes were performed by PCR mutagenesis.cDNA coding for the N-terminal portion of TFIIIAwas ampli¢ed from plasmid pT7-TFIIIA which con-tains the 1.5 kb EcoRI cDNA fragment coding forintact Xenopus TFIIIA (kind gift from R. Roeder).PCR was performed with Taq polymerase using anupstream primer complementary to the polylinkerand a downstream primer containing the desired mu-tation. This mutant fragment was gel puri¢ed, heatdenatured, and annealed with heat-denatured, 1.5 kbEcoRI cDNA fragment. Prior to heat denaturation,the 3P ends of the EcoRI fragment were ¢lled in withdideoxy ATP using reverse transcriptase. A fragmentwas ampli¢ed from this annealing reaction using theoriginal upstream primer and a new downstream pri-mer which anneals to sequences coding for the C-terminal tail of TFIIIA. This PCR product was di-gested with EcoRI and XhoI, gel puri¢ed, and clonedinto Phagescript (Stratagene). The 600 bp EcoRI-XhoI insert in recombinant phage DNA was se-quenced initially to identify mutations and then com-pletely to check for the absence of other mutations.
2.2. Expression of wild-type and mutant TFIIIAcDNA in E. coli
Phage DNA inserts were excised by EcoRI-SphI,SphI-HindIII, or EcoRI-XhoI digestion and gel puri-¢ed. Mutant fragments were ligated with the comple-menting wild-type fragment (coding for the N-termi-nal or C-terminal half of TFIIIA) into pT7-7expression vector [30]. The pT7-TFIIIA plasmid,which contains a T7 bacteriophage RNA polymerasepromoter and ribosome binding site upstream of theEcoRI site, was transformed into E. coli K38 cellscontaining plasmid GP1-2 [31]. This plasmid con-tains the gene coding for T7 RNA polymerase underthe control of the VPL promoter, the gene coding forthe temperature-sensitive VCI857 repressor, and thekanamycin-resistance gene. K38 colonies containingboth plasmids were selected on agar plates contain-ing ampicillin and kanamycin. 500 ml colony cultureswere grown with selection overnight at 40³C. Cellswere harvested at 4³C by centrifugation, washed with20 mM Tris-HCl, pH 7.6, 50 mM KCl, 1.5 mM
MgCl2, and pellets were resuspended in 5 ml of bu¡-er A (20 mM Tris-HCl, pH 7.6, 320 mM KCl,1.5 mM MgCl2, 0.2 mM PMSF, 0.4 mM DTT,0.01 mM ZnCl2). Cell suspensions were sonicatedwith a microtip in an ice-bath using a Sonics andMaterials sonicator. After removal of cell debris bylow-speed centrifugation, the samples were fraction-ated by cation-exchange chromatography on BIO-REX 70 (Pharmacia) as described previously [16].Column fractions eluting at 0.75 M KCl in bu¡erA (minus PMSF) were assayed and quanti¢ed forTFIIIA by SDS-PAGE/immunoblotting with rabbitanti-TFIIIA serum and known amounts of puri¢edXenopus TFIIIA [16].
2.3. Isolation of Xenopus 7S RNP/TFIIIA andDNase I footprinting
TFIIIA-containing 7S ribonucleoprotein particleswere isolated from ovarian tissue of immatureX. laevis frogs (4^5 cm, Nasco, Fort Atkinson,WI). Ovarian tissue (from 20 frogs) was homogen-ized in 50 mM Tris-HCl, pH 7.5, 50 mM KCl, 5 mMMgCl2, 0.4 mM DTT, and 0.2 mM PMSF and cen-trifuged at low speed to remove debris. The super-natant containing 7S particles composed of TFIIIAand oocyte 5S RNA was centrifuged at 4³C through15^30% glycerol gradients in a SW41 rotor at 34 000rpm for 22 h. Gradient fractions were assayed forabsorbance at 280 nm and the 7S peak fractionswere pooled. 7S particles were puri¢ed from thesefractions by DEAE cellulose chromatography as de-scribed previously [32]. Protein amounts were deter-mined by the dye-binding method using bovine se-rum albumin as a standard [33]. Oocyte 5S RNAbound to TFIIIA was removed by digesting 7S par-ticles with RNase A (20 Wg/ml) in bu¡er A (minusPMSF) for 30 min at room temperature. For DNaseI footprint analysis, a 303 bp Xenopus borealis so-matic 5S RNA gene-containing template was end-labeled on the coding strand using reverse transcrip-tase and [K-32P]dATP as described previously [16].The end-labeled fragment was puri¢ed by PAGEand its speci¢c activity was determined by absorb-ance at 260 nm and Cerenkov counting. Footprintingassays with native TFIIIA isolated from Xenopusoocytes, wild-type and mutant TFIIIA expressedand isolated from E. coli were performed in 20 Wl
BBAEXP 93115 30-6-98
J.S. Hanas et al. / Biochimica et Biophysica Acta 1398 (1998) 256^264258

reaction volumes containing 20 mM Tris-HCl, pH7.6, 70 mM NH4Cl, 7 mM MgCl2, 0.4 mM DTT,and 0.1% nonidet P-40. TFIIIA and DNA concen-trations in the footprint reactions are indicated in the¢gure legends and reaction conditions were the sameas those described previously [16]. Mutant TFIIIAsdecreased footprint binding saturation (as deter-mined previously [32]) by less than 10% (W28S,H58Q, R154L, R27I) or greater than 90% (K11A,R62L, R62Q, K92L, vF94).
3. Results and discussion
3.1. The highly conserved Arg-62 residue in TFIIIA isrequired for binding the 5S gene
Arg-62 in ¢nger II (Fig. 1) is the most conservednon-zinc coordinating amino acid in yeast, ¢sh, am-phibian, and human TFIIIA [8^11,25,26]. BothNMR and sequence conservation alignment analysesdemonstrated this residue interacts with G85 in the5S gene ICR [11,27]. To assess the requirement forArg-62 in TFIIIA-5S gene interactions, site-directedmutagenesis was used to change this residue to aLeu. Leu was chosen because it will not hydrogenbond to a nucleotide base nor will it disrupt the
DNA recognition helix. Signi¢cantly, TFIIIA withthe R62L change is able to a¡ord less than 10%DNase I protection (nucleotides +43 to +96, Fig.2A, lanes 6 and 7) a¡orded by the native proteinpuri¢ed from frogs or recombinant wild-type proteinexpressed in and puri¢ed from E. coli (lanes 2^5).The DNase I protection assay is a stringent and spe-ci¢c binding assay since the footprint depends upona high a¤nity binding event that excludes DNase Ifrom the template during the binding reaction [34].Under these footprinting conditions (large molar ex-cess of DNA-binding protein to DNA template), lossof DNase I protection was previously shown to in-dicate about a 10-fold increase in apparent Kd for thebinding protein [34]. For TFIIIA which binds the 5Sgene with an apparent Kd of about 1039 M [32],these assay conditions most likely discriminate onlythose mutations which substantially increase the Kd,probably greater than 5-fold. The loss of DNase Iprotection by R62L TFIIIA over the entire 5S geneICR is consistent with previous observations thatindicated binding of the N-terminal ¢ngers enhancedbinding by the more C-terminal ¢ngers [15,16]. Aprevious study using ¢lter binding examined the in-teraction of Arg-62 with the 5S gene indirectly bymutating this residue within the context of mutatinga group of surrounding residues in ¢nger II, which
Fig. 1. Amino acid changes in X. laevis TFIIIA. TFIIIA primary sequence (single-letter code [40]) is folded into nine ¢nger domains(I^IX) and a non-¢nger C-terminal tail. The ICR of the Xenopus 5S RNA gene with the C- and A-boxes and the intermediate (M) el-ements are exhibited above the zinc ¢nger domains. The dashed, downward lines at the 3P and 5P ends of the ICR demarcate the re-gion of TFIIIA-dependent DNase I protection. Numbered amino acids in the large circles have been changed in this study from therespective residues positioned next to the small dark circles. ** indicates F94 deletion.
BBAEXP 93115 30-6-98
J.S. Hanas et al. / Biochimica et Biophysica Acta 1398 (1998) 256^264 259

together inhibited DNA binding in the absence of anArg-62 change [35]. The extreme conservation andcentral importance of Arg-62 in TFIIIA functionwarrants its individual biochemical analysis in thepresent study.
As an added test of speci¢city, Arg-62 was alsochanged to a Gln, an amino acid (unlike Leu) capa-ble of hydrogen bonding to nucleotide bases. As ex-hibited in Fig. 2B (lanes 4 and 5), the R62Q changealso strongly inhibited the interaction of TFIIIAwith the 5S gene as evidenced by the s 90% loss ofDNase I protection. These results demonstrate thecritical nature of contacts between individual resi-dues in the N-terminal ¢ngers of TFIIIA and nucleo-tide bases in the C-box. The conserved Lys-92 inamphibian and human ¢nger III (not present in yeastor ¢sh TFIIIA) also was changed to a Leu and theresultant TFIIIA was assayed for its ability to pro-tect the 5S gene ICR from DNase I digestion. Likethe R62L mutant, this leucine modi¢cation led togreater than 90% loss of DNAse I protection of theICR (Fig. 2B, lanes 6 and 7). A K92A mutationanalyzed by ¢lter binding caused about a 6-fold in-crease in Kd [35]. The s 90% loss of DNase I pro-tection observed for the K92L mutant TFIIIA inFig. 2B is consistent with such an a¤nity reduction.NMR indicated K92 interacts with G81 and/or G82[27].
3.2. Conserved arginines in TFIIIA ¢ngers V and IXare not individually required for 5S geneinteraction
Two other highly conserved basic residues in ver-tebrate TFIIIAs, Arg-154 and Arg-271, are locatedin the recognition helices of ¢ngers V and IX inidentical positions to Arg-62 (see Fig. 1). The con-servation alignment method predicted these basic res-idues interact with guanine 70 (in M element) andguanine 48 (in A-box) respectively on the ICR [11].R154 and R271 may also be individually required forTFIIIA interaction along the entire ICR like Arg-62if binding mechanisms are the same for all ¢ngers.To test this hypothesis, these residues were changedto Leu and Ile respectively and assayed by DNase Ifootprinting. This assay has the ability to analyzebinding along the entire 5S gene ICR which can yieldmechanistic information about the polydactyl struc-ture of TFIIIA. For example, a previous study dem-onstrated that individual mutations in the zinc bind-ing residues in the C-terminal ¢ngers of TFIIIAcould inhibit DNase protection by C-terminal ¢ngersbut not by the N-terminal ¢ngers on the C-box re-
Fig. 2. Autoradiogram of the in£uence of Arg-62 mutations onthe interaction of Xenopus TFIIIA with the 5S RNA gene. Site-directed mutagenesis, TFIIIA isolation and expression, TFIIIA-dependent DNase I protection reactions, and autoradiographywere performed as described in Section 2. Nucleotide numbersindicate the DNase I protection boundaries on the codingstrand of the 5S gene; letters and brackets indicate the posi-tions of the C- and A-boxes as well as the intermediate element(M). (A) DNase I protection reactions in the presence of 20nM or 30 nM native TFIIIA from frogs (lanes 2, 3), wild-typeTFIIIA from E. coli (lanes 4, 5), and Arg62-Leu TFIIIA (lanes6, 7). (B) Protection reactions in the presence of 20 or 30 nMwild-type TFIIIA (lanes 2, 3), Arg62-Gln TFIIIA (lanes 4, 5),and Lys92-Leu TFIIIA (lanes 6, 7). All DNase I reactions con-tained 2 nM end-labeled 5S gene and reactions in lanes 1 con-tained no TFIIIA.
BBAEXP 93115 30-6-98
J.S. Hanas et al. / Biochimica et Biophysica Acta 1398 (1998) 256^264260
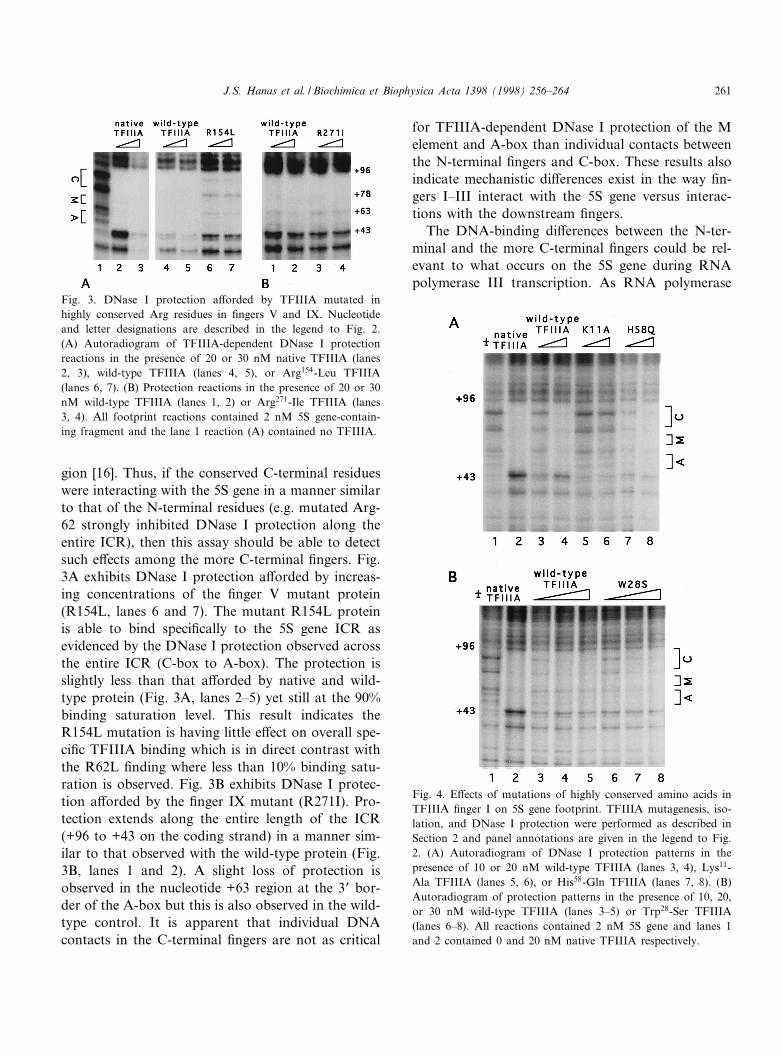
gion [16]. Thus, if the conserved C-terminal residueswere interacting with the 5S gene in a manner similarto that of the N-terminal residues (e.g. mutated Arg-62 strongly inhibited DNase I protection along theentire ICR), then this assay should be able to detectsuch e¡ects among the more C-terminal ¢ngers. Fig.3A exhibits DNase I protection a¡orded by increas-ing concentrations of the ¢nger V mutant protein(R154L, lanes 6 and 7). The mutant R154L proteinis able to bind speci¢cally to the 5S gene ICR asevidenced by the DNase I protection observed acrossthe entire ICR (C-box to A-box). The protection isslightly less than that a¡orded by native and wild-type protein (Fig. 3A, lanes 2^5) yet still at the 90%binding saturation level. This result indicates theR154L mutation is having little e¡ect on overall spe-ci¢c TFIIIA binding which is in direct contrast withthe R62L ¢nding where less than 10% binding satu-ration is observed. Fig. 3B exhibits DNase I protec-tion a¡orded by the ¢nger IX mutant (R271I). Pro-tection extends along the entire length of the ICR(+96 to +43 on the coding strand) in a manner sim-ilar to that observed with the wild-type protein (Fig.3B, lanes 1 and 2). A slight loss of protection isobserved in the nucleotide +63 region at the 3P bor-der of the A-box but this is also observed in the wild-type control. It is apparent that individual DNAcontacts in the C-terminal ¢ngers are not as critical
for TFIIIA-dependent DNase I protection of the Melement and A-box than individual contacts betweenthe N-terminal ¢ngers and C-box. These results alsoindicate mechanistic di¡erences exist in the way ¢n-gers I^III interact with the 5S gene versus interac-tions with the downstream ¢ngers.
The DNA-binding di¡erences between the N-ter-minal and the more C-terminal ¢ngers could be rel-evant to what occurs on the 5S gene during RNApolymerase III transcription. As RNA polymerase
Fig. 4. E¡ects of mutations of highly conserved amino acids inTFIIIA ¢nger I on 5S gene footprint. TFIIIA mutagenesis, iso-lation, and DNase I protection were performed as described inSection 2 and panel annotations are given in the legend to Fig.2. (A) Autoradiogram of DNase I protection patterns in thepresence of 10 or 20 nM wild-type TFIIIA (lanes 3, 4), Lys11-Ala TFIIIA (lanes 5, 6), or His58-Gln TFIIIA (lanes 7, 8). (B)Autoradiogram of protection patterns in the presence of 10, 20,or 30 nM wild-type TFIIIA (lanes 3^5) or Trp28-Ser TFIIIA(lanes 6^8). All reactions contained 2 nM 5S gene and lanes 1and 2 contained 0 and 20 nM native TFIIIA respectively.
Fig. 3. DNase I protection a¡orded by TFIIIA mutated inhighly conserved Arg residues in ¢ngers V and IX. Nucleotideand letter designations are described in the legend to Fig. 2.(A) Autoradiogram of TFIIIA-dependent DNase I protectionreactions in the presence of 20 or 30 nM native TFIIIA (lanes2, 3), wild-type TFIIIA (lanes 4, 5), or Arg154-Leu TFIIIA(lanes 6, 7). (B) Protection reactions in the presence of 20 or 30nM wild-type TFIIIA (lanes 1, 2) or Arg271-Ile TFIIIA (lanes3, 4). All footprint reactions contained 2 nM 5S gene-contain-ing fragment and the lane 1 reaction (A) contained no TFIIIA.
BBAEXP 93115 30-6-98
J.S. Hanas et al. / Biochimica et Biophysica Acta 1398 (1998) 256^264 261
traverses the 5S gene ICR, it would ¢rst disruptTFIIIA-DNA contacts in the more C-terminal ¢n-gers because this is the direction the polymerase ismoving. The disruption of these individual contactswould still allow TFIIIA to remain bound to the 5Sgene, which still may be necessary for transcriptioncomplex stability. However, once the polymerasemoves forward and disrupts individual contacts inthe N-terminal ¢ngers, the entire TFIIIA proteincould release and possibly bind the nascent 5SRNA transcript. If Arg-154 and Arg-271 are not in-dividually critical for the TFIIIA-5S RNA gene in-teraction, then why are they so highly conserved?Because the more C-terminal ¢ngers of TFIIIAbind 5S ribosomal RNA as well as DNA, these ¢n-gers may have evolved binding mechanisms distinctfrom the N-terminal group to accommodate 5S RNAbinding. Arg-154 and Arg-271 individually couldhave a more critical role in RNA binding thanDNA binding, accounting for their high conserva-tion.
3.3. Non-conserved basic residue His-58 is notrequired for TFIIIA binding.
DNase I protection analyses in Fig. 2 demon-strated the individual requirements for the basic res-idues Arg-62 and Lys-92 in TFIIIA-5S gene interac-tions. His-58 is another basic residue with DNA-binding potential (possibly to a guanine) in thesame recognition helix as Arg-62. This amino acidis not conserved in vertebrate TFIIIAs [11]. NMRanalysis of the TFIIIA-C-box interaction was notable to determine whether His-58 was making abase-speci¢c contact [27]. TFIIIA-5S gene alignmentusing the sequence conservation approach indicatedHis-58 was not involved in a base-speci¢c contact
whereas the conserved His-59 was binding to guanine86 [11]. Therefore, DNase I protection analysis wasperformed on a His-58 mutation in order to deter-mine its necessity in DNA binding. When this resi-
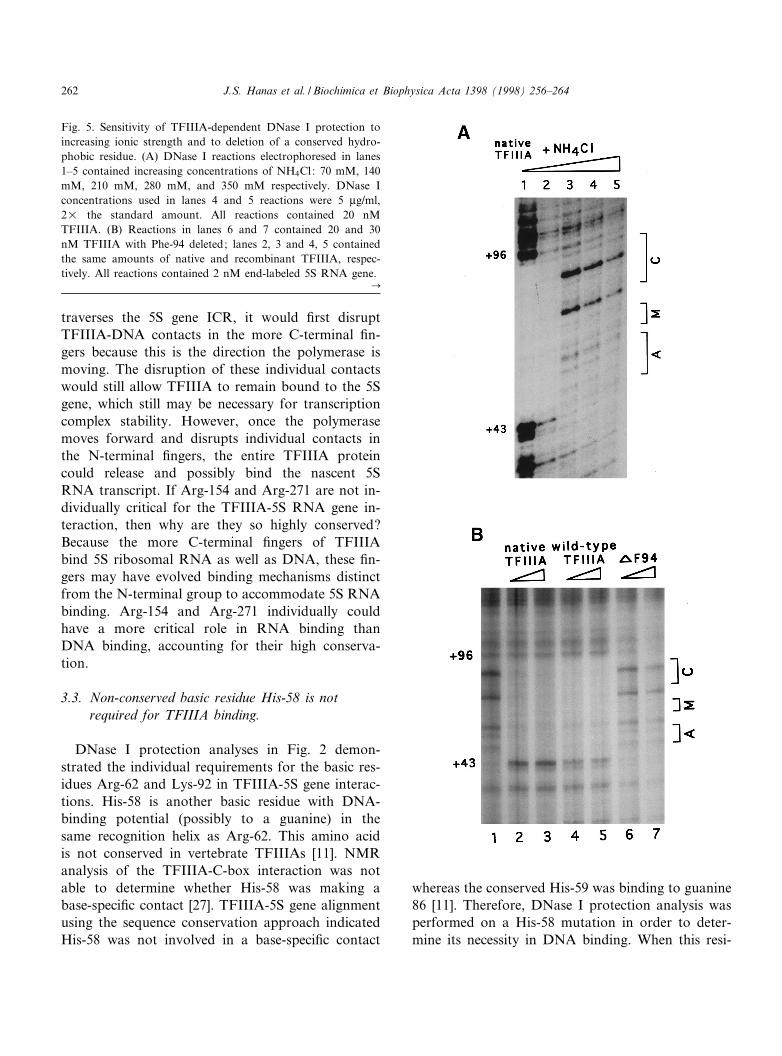
Fig. 5. Sensitivity of TFIIIA-dependent DNase I protection toincreasing ionic strength and to deletion of a conserved hydro-phobic residue. (A) DNase I reactions electrophoresed in lanes1^5 contained increasing concentrations of NH4Cl: 70 mM, 140mM, 210 mM, 280 mM, and 350 mM respectively. DNase Iconcentrations used in lanes 4 and 5 reactions were 5 Wg/ml,2U the standard amount. All reactions contained 20 nMTFIIIA. (B) Reactions in lanes 6 and 7 contained 20 and 30nM TFIIIA with Phe-94 deleted; lanes 2, 3 and 4, 5 containedthe same amounts of native and recombinant TFIIIA, respec-tively. All reactions contained 2 nM end-labeled 5S RNA gene.
C
BBAEXP 93115 30-6-98
J.S. Hanas et al. / Biochimica et Biophysica Acta 1398 (1998) 256^264262

due was changed to a Gln as was done with Arg-62,the mutant TFIIIA was still able to a¡ord completeDNase I protection for the 5S gene (Fig. 4A, lanes 7and 8) like the wild-type protein (lane 4). This experi-ment is consistent with His-58 not making a strongcontact with the C-box like Arg-62. In order to pos-sibly obtain a more detailed picture of the interactionof the His58-His59 region of TFIIIA ¢nger II with the5S gene C-box, molecular modeling of the TFIIIAN-terminal ¢ngers was performed using the X-raycrystallography coordinates from Zif268-DNA com-plex [36]. In the model for ¢nger II, the highly con-served His-59 residue is observed hydrogen bondingwith guanine 86 on the non-coding strand (model notshown). Importantly, the adjacent residue His-58which is not conserved in vertebrate TFIIIAs wasnot observed hydrogen bonding to a base. The lackof a His-58-DNA interaction in the sequence conser-vation analysis and TFIIIA molecular model is con-sistent with the negative results of the H58Q muta-genesis experiment (Fig. 4A).
Unlike His-58, Lys-11 is a conserved basic aminolocated on the N-terminal side of the ¢nger ICys....Cys zinc coordination sphere, well away fromthe ¢nger recognition helix (see Fig. 1). This locationsuggests it may not be capable of hydrogen bondingto a base in the major groove of the 5S gene C-box.This Lys was in the abbreviated polypeptide used inthe TFIIIA-C-box NMR study but its structure (aswell as some other Lys residues) was not well re-solved [27]. In the present study, Lys-11 was replacedby an Ala which has no basic charge and the abilityof the mutant TFIIIA to bind the 5S gene was exam-ined (Fig. 4A). Lanes 5 and 6 exhibit the DNase Iproducts in the presence of increasing concentrationsof K11A TFIIIA. TFIIIA-dependent DNase I pro-tection is inhibited relative to wild-type TFIIIA(lanes 3 and 4); higher concentrations of the mutantprotein did not yield further protection (not shown).Previously, a deletion of the ¢nger I region contain-ing Lys-11 (as well the potential zinc-coordinatingCys-15) was found to inhibit TFIIIA-dependentDNase I protection of the 5S gene [19]. Because ofits location, Lys-11 is possibly involved in a contactwith the DNA backbone phosphate similar to thatobserved in the Zif278-DNA complex [36]. Trp-28 isa conserved hydrophobic residue in ¢nger I (presentin ¢sh, amphibian and human TFIIIA). Such resi-
dues may have a role in ¢nger structure stability[36] but recent NMR data indicate a more directrole in DNA binding [27]. When this residue ischanged to a hydrophilic Ser, little e¡ect on 5Sgene binding is observed (Fig. 4B, cf. lanes 6^8,W28S, with lanes 3^5, wild-type TFIIIA). TheW28S structural change may not be large enoughto strongly in£uence 5S gene binding as assayed byDNase I protection.
3.4. Interaction of TFIIIA zinc ¢ngers with the5S gene is highly sensitive to ionic strength
NMR analysis of the TFIIIA-5S gene C-box inter-action demonstrated a large number of contacts (atleast 16) involving amino acids in the N-terminal¢ngers and DNA backbone phosphates [27]. Phos-phate contacts are likely present in the DNA inter-actions of the more C-terminal ¢ngers as well sincesuch contacts were observed over most of the ¢ve¢nger structure of GL1 [37]. Protein-DNA phosphatecontacts are usually sensitive to elevated salt concen-trations and such contacts disrupted by salt concen-trations less than 0.4 M are considered highly sensi-tive to ionic strength [38]. In order to determine thesalt sensitivity of the TFIIIA-5S gene interaction aswell as possibly distinguish non-speci¢c contacts inthe N-terminal ¢ngers from the more C-terminal ¢n-gers, TFIIIA-dependent DNase I protection was an-alyzed under conditions of increasing amounts ofsalt. Lane 1 in Fig. 5A exhibits TFIIIA-dependentDNase I protection of the 5S gene in the presenceof 70 mM NH4Cl, the normal salt concentrationused in this assay; complete protection is observed.Lanes 2^5 exhibit DNase I protection in the presenceof increasing salt concentrations; inhibition ofTFIIIA-dependent protection over the entire 5Sgene ICR (+43 to +96) is observed at NH4Cl con-centrations of 210 mM and above (lanes 3^5) but notat 140 mM (lane 2). Because TFIIIA binding is in-hibited by elevated salt along the entire length of the5S gene ICR (nucleotides +43 to +96, lanes 2^5),ionic contacts between the N-terminal ¢ngers andthe C-box are at least as sensitive or more sensitiveto ionic strength than such contacts in the C-terminal¢ngers.
In a mutagenesis experiment targeting anotherconserved amino acid, inhibition of 5S gene binding
BBAEXP 93115 30-6-98
J.S. Hanas et al. / Biochimica et Biophysica Acta 1398 (1998) 256^264 263

(Fig. 5B, lanes 6 and 7) is observed upon deletion ofa Phe-94 located between the two zinc-coordinatingHis residues of ¢nger III. Most TFIIIA ¢ngers havethree residues between the His...His structure except¢ngers III, VI, and VIII which have four (see Fig. 1).This quartet structure in ¢nger III is highly con-served in vertebrate TFIIIAs (but not yeast TFIIIA)and may have a role in organizing TFIIIA ¢ngersalong DNA as was proposed for other Cys2His2 ¢n-ger proteins [11,39]. A direct DNA-binding role forthis Phe residue was not observed in the NMR struc-ture of ¢nger III bound to DNA but rather a stabi-lizing e¡ect on ¢nger structure was suggested [27].Therefore, the Phe-94 deletion is likely altering afunction dependent upon its conserved hydrophobic-ity and possibly the presence of four residues be-tween the His..His structure and not three. A pre-vious study demonstrated DNA-binding inhibitionwith a F94K change [35].
Acknowledgements
This work was supported by grants from the Na-tional Institute for General Medical Sciences, theNational Science Foundation, and the OklahomaCenter for the Advancement of Science and Technol-ogy. We thank J. Smith for the excellent technicalassistance.
References
[1] D.R. Engelke, S.-Y. Ng, B.S. Shastry, R.G. Roeder, Cell 19(1980) 717^728.
[2] J.S. Hanas, D.J. Hazuda, D.F. Bogenhagen, F.Y.-H. Wu,C.W. Wu, J. Biol. Chem. 258 (1983) 14120^14125.
[3] A.M. Ginsberg, B.O. King, R.G. Roeder, Cell 39 (1984)479^489.
[4] J. Miller, A.D. McLachlan, A. Klug, EMBO J. 4 (1985)1609^1614.
[5] J.M. Berg, Y. Shi, Science 271 (1996) 1081^1085.[6] J.S. Hanas, C.J. Gaskins, J.F. Smith, M.K. Ogilvie, Prog.
Nucleic Acid Res. Mol. Biol. 43 (1992) 205^239.[7] S. Heniko¡, E.A. Greene, S. Pietrokovski, P. Bork, T.K.
Attwood, L. Hood, Science 278 (1997) 609^614.[8] C.J. Gaskins, J.S. Hanas, Nucleic Acids Res. 18 (1990) 2117^
2123.[9] C.J. Gaskins, J.F. Smith, M.K. Ogilvie, J. Hanas, Gene 120
(1992) 197^206.[10] P.D. Drew, J.W. Nagle, R.D. Canning, K. Ozato, W.E.
Biddison, K.G. Becker, Gene 159 (1995) 215^218.
[11] M.K. Ogilvie, J.S. Hanas, Gene 203 (1997) 103^112.[12] B.M. Honda, R.G. Roeder, Cell 22 (1980) 119^126.[13] H. Pelham, D.D. Brown, Proc. Natl. Acad. Sci. USA 77
(1980) 4170^4174.[14] T. Pieler, J. Hamm, R.G. Roeder, Cell 48 (1987) 91^100.[15] J.S. Hanas, R.M. Littell, C.J. Gaskins, R. Zebrowski,
Nucleic Acids Res. 17 (1989) 9861^9870.[16] J.F. Smith, J. Hawkins, R.E. Leonard, J.S. Hanas, Nucleic
Acids Res. 19 (1991) 6871^6876.[17] O. Theunissen, F. Rudt, U. Guddat, H. Mentzel, T. Pieler,
Cell 71 (1992) 679^690.[18] S.J. McBryant, N. Veldhoen, B. Gedulin, A. Leresche, M.P.
Foster, P.E. Wright, P.J. Romainuk, J.M. Gottesfeld, J. Mol.Biol. 248 (1995) 44^57.
[19] K.E. Vrana, M.E.A. Churchill, T.D. Tulius, D.D. Brown,Mol. Cell. Biol. 8 (1988) 1684^1696.
[20] R.M. Fiser-Littell, A.L. Duke, J.S. Yanchick, J.S. Hanas,J. Biol. Chem. 263 (1988) 1607^1610.
[21] X.B. Liao, K.R. Clemens, L. Tennant, P.E. Wright, J.M.Gottesfeld, J. Mol. Biol. 223 (1992) 857^871.
[22] P.K. Hansen, J.H. Christensen, J. Nyborg, O. Lillelund,H.C. Thogersen, J. Mol. Biol. 233 (1993) 191^202.
[23] K.R. Clemens, X. Liao, V. Wolf, P.E. Wright, J.M. Gottes-feld, Proc. Natl. Acad. Sci. USA 89 (1992) 10822^10826.
[24] J.J. Hayes, K.R. Clemens, Biochemistry 31 (1992) 11600^11605.
[25] J. Archambault, C.A. Milne, K.T. Schappert, B. Baum, J.D.Friesen, J. Segall, J. Biol. Chem. 267 (1992) 3282^3288.
[26] N.A. Woychik, R.A. Young, Proc. Natl. Acad. Sci. USA 89(1992) 3999^4003.
[27] D.S. Wuttke, M.P. Foster, D.A. Case, J.M. Gottesfeld, P.E.Wright, J. Mol. Biol. 273 (1997) 183^206.
[28] J.R. Sayers, W. Schmidt, F. Eckstein, Nucleic Acids Res. 16(1988) 791^802.
[29] F. Sanger, S. Nicklen, A.R. Coulson, Proc. Natl. Acad. Sci.USA 74 (1977) 5463^5467.
[30] S. Tabor, C.C. Richardson, Proc. Natl. Acad. Sci. USA 82(1985) 1074^1078.
[31] M. Russel, P. Model, J. Bacteriol. 159 (1985) 1034^1039.[32] J.S. Hanas, D.F. Bogenhagen, C.-W. Wu, Proc. Natl. Acad.
Sci. USA 80 (1983) 2142^2145.[33] M.M. Bradford, Anal. Biochem. 72 (1976) 248^254.[34] D.J. Galas, A. Schmitz, Nucleic Acids Res. 5 (1978) 3157^
3170.[35] W.-Q. Zang, N. Veldhoen, P.J. Romaniuk, Biochemistry 34
(1995) 15545^15552.[36] N.P. Pavletich, C.O. Pabo, Science 252 (1991) 809^817.[37] N.P. Pavletich, C.O. Pabo, Science 261 (1993) 1701^1707.[38] J.W. Chase, K.R. Williams, Annu. Rev. Biochem. 55 (1986)
103^136.[39] M. Kochoyan, T.F. Havel, D.T. Nguyen, C.E. Dahl, H.T.
Keutmann, M.A. Weiss, Biochemistry 30 (1991) 3371^3386.[40] M.O. Dayho¡, in: Atlas of Protein Sequence and Structure
5, Suppl. 3, National Biomedical Research Foundation,Washington, DC, 1978.
BBAEXP 93115 30-6-98
J.S. Hanas et al. / Biochimica et Biophysica Acta 1398 (1998) 256^264264




















