
Structure and conformations of the bovine ATP synthase by ...

voiuma 10 Number 161982 Nucleic Acids Research
NMR evidence for the existence of two native conformations of 5S RNA
MJ.Kime and P.B.Moore
Department of Chemistry, Yale University, P.O. Box 6666, New Haven, CT 06511, USA
Received 14 May 1982; Revised and Accepted 6 July 1982
SUMMARYHfiR spectra of the non-exchangeable protons 1n 5S RNA from E.- co l l
show the existence of two d i s t i n c t conformers of the molecule which meetthe operational d e f i n i t i o n of "A form" or native 5S RNA. Both are eas i lydistinguished spectroscopically from denatured, "B form" 5S RNA. Theconditions which Interconvert the two A form conformers strongly suggestthat the t rans i t ion between them gives r ise to the low temperature opt icalmelting t rans i t ion f i r s t reported 1n 5S RNA by Kao and Crothers ( 1 ) .
INTRODUCTION
5S RNA 1s the smallest of the three RNAs found 1n procaryotic r1bo-
somes. In E_. co l l 1 t 1s an RNA of about 40,000 molecular weight consisting
of 120 bases. While many a c t i v i t i e s of the 50S subunit pers is t a t measur-
able levels 1n par t ic les lacking 5S RNA, a l l are markedly stimulated by
I t s presence ( 2 ) . However, beyond I t s contr ibution to the structural
I n t e g r i t y of the 50S subunit, I t s role 1n protein synthesis remains
obscure ( f o r review see reference 3 ) .
I t has been known for many years that 5S RNA can be obtained 1n two
sta tes , the A and B forms ( 4 ) . The two forms are separable chromatogra-
ph ica l l y , and d i f f e r both 1n t h e i r electrophoret ic mobi l i t i es (5) and 1n
t h e i r a f f i n i t y for proteins L5, LI8 and L25, the three found associated
with 5S RNA 1n the ribosome (see (6) and references t h e r e i n ) . Because A
form 5S RNA par t ic ipates 1n ribosome reconsti tut ion while B form does not
( 4 ) , and A form 1s also the one favored thermodynamically under physio-
logical conditions ( 7 ) , I t Is believed to correspond to the s ta te of 5S
RNA In the In tac t ribosome.
In the course of a series of experiments on protein Interact ions
with 5S RNA we have obtained NMR data which Indicate tha t there ex ist two
readi ly distinguishable conformations of the molecule which meet the
operational d e f i n i t i o n of "A form". The conditions required for e l i c i t i n g
C IRL Pros Limited, Oxford, England. 49730306-1048/82/1016-4973S 2.00/0
by guest on September 16, 2016
http://nar.oxfordjournals.org/D
ownloaded from

Nucleic Acids Research
these two forms and interconverting them strongly suggest that they
correspond to the high and low temperature states of 5S RNA Ident i f ied
opt ica l ly by Kao and Crothers (1). The t rans i t ion between these two
states produces a detectable hyperchromic ef fect 1n 5S RNA preparations
at near physiological temperatures and 1on1c conditions (1). The NMR
data indicate that a substantial difference 1n the arrangement of bases
must ex is t between these two forms, consistent with the optical obser-
vations. Both forms bind protein L25.
MATERIALS AND METHODS
5S RNA. Ribosomes were prepared both from £ . co l l MRE 600 and from
£. co l i PR13 by methods described elsewhere (8). RNA was obtained by
phenol extract ion of Intact 70S particles or of L1Cl-urea precipitates
derived from 50S subunits. 5S RNA was puri f ied from the result ing rRNA
mixtures by the method of Kao & Crothers (1) , except that the buffer used
for Sephacryl chromatography was 0.15M NaCl, 1$ methanol, 0.1M sodium
acetate, pH 5.0. Purity was assessed by acrylamide gel electrophoresis.
5S preparations used 1n th is work contained no detectable traces of
other RNA species, and were ent i rely in the A configurat ion. B form 5S
RNA was prepared by urea denaturation followed by gel f i l t r a t i o n as
described by Aubert, Scott, Reynier and Monier (4) . Samples were stored
at -80° C 1n Sephacryl buffer, and appeared to be stable Indef in i te ly .
Concentrations of 5S RNA were determined by absorbance at 260 nm assuming
that an 0D26Q of 1.0 corresponds to a 1.18 x 10"6M solution of 5S RNA
(9) , regardless of conformation.
Gel Electrophoresis. RNA and RNA/prote1n complexes were examined on
acrylamide gels. The gel composition was 10% acrylamide, 0.5$ bisacry-
lamide, 0.08% TEMED, 0.05% ammonium persulfate. The buffer used, unless
otherwise noted, was 0.1 M KC1, 5 mM MgCU, 50 mM Tr1s-borate, pH 7.8.
Gels were run at 3V/cm for 16 hours either at 4° C or at room temperature.
Buffer was circulated between reservoirs to s tab i l i ze the pH. RNA was
stained with methylene blue (10) and protein with Coomassie blue (.025%
1n a 50:50 mixture of 7% acetic add and ethanol). The two stains are
specif ic fo r RNA and protein respectively.
Protein L25. The preparation of L25 from 50S subunits has been
described previously (11). The same techniques suf f ice to pur i fy L25
from to ta l 70S protein as wel l , since 1t elutes from carboxymethyl ce l l u -
lose, at pH 5.6 1n urea, separate from a l l other species. Concentrations
4974
by guest on September 16, 2016
http://nar.oxfordjournals.org/D
ownloaded from

Nucleic Acids Research
of L25 were estimated from absorbance at 276 ran. The absorbance of a 13!
solution of L25 at 276 nm 1s 3.7 (Moore, unpublished observations).
WR Spectroscopy. Al l spectra were accumulated on a Bruker WM 500
NMR spectrometer, operating 1n the Fourier transform mode. Typical
spectra required the accumulation of 1000 transients with a cycle time of
about 1.5 sec. Increasing the cycle time to 2.5 sec had no Influence on
the spectra. The residual HOD resonance was not suppressed by preirra-
diat ion. Spectra were obtained on samples held at 25° or 27° C at concen-
trations ranging from 0.5 to 1.0 mM. Dioxane was used as a chemical
sh i f t standard. I ts (single) proton resonance has a chemical sh i f t of
3.741 ppm relat ive to the methyl resonance of 3-(tr1methyls1lyl)propane
sulfonic add .
X-ray Scattering. X-ray solution scattering measurements were done
using an Instrument based on focussing mirror optics and a linear position-
sensitive detector (12). Copper KQ X-rays were used ( X= 1.54 A). Data
were corrected for smearing effects by Indirect transformation (13).
RESULTS
Mg^Dependence of the fMR Spectrum of A form 5S RNA. An aliquot of
5S RNA was dialyzed into 0.1 M KC1, 10 mM Tr1s-HCl, pH 7.6 1n H^ at 4°
C. The sample was concentrated by u l t r a f i l t r a t i o n to bring the RNA
concentration to 0.65 mM. I t was then divided Into two parts and one of
the portions dialyzed into 0.1 M KC1, 10 mM deuterotr1s-HCl, pH 7.6, D20.
The remaining portion was dialyzed Into the same buffer with 2 mM MgCl-.
The spectra given by the two samples at 27° C are shown 1n Figure 1,
traces a and b.
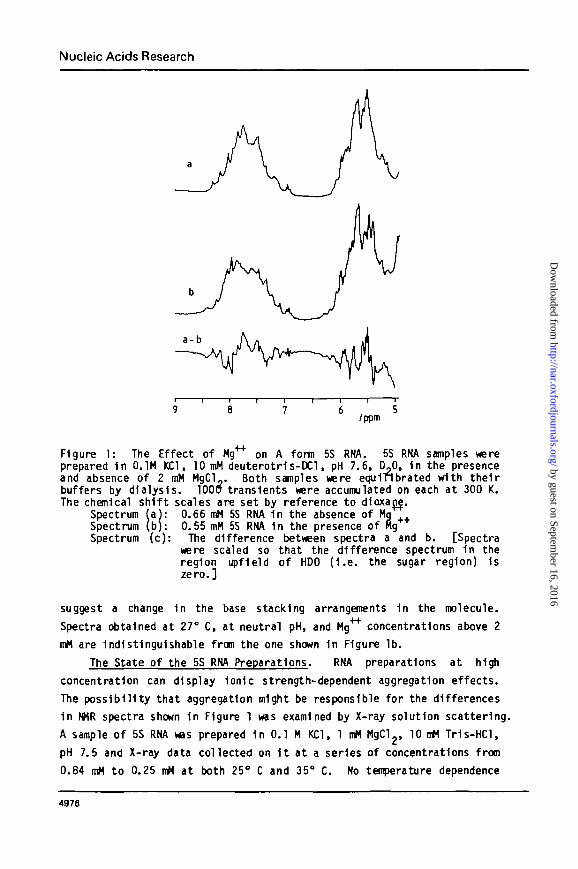
Figure 1 shows the non exchangeable proton spectra of the two samples
downfield of HDO (~4.7 ppm). The cluster of resonances around 5.5 ppm
are those of the ribose I 1 protons and the pyrimidine 5 protons. The
resonances of the remaining nonexchangeable base protons appear 1n the
aromatic region around 7.5 ppm. The two spectra shown 1n Figure 1 d i f fe r
s igni f icant ly 1n both these regions. The th i rd spectrum 1n Figure 1 1si i 1 ^
the low Mg spectrum, (a), less the high Mg spectrum, (b). The ratio
of the Integrated Intensity of the positive (or negative) regions of the
difference spectrum to that of the whole spectrum Indicates that the
environments of at least 15S of the bases 1n the molecule are affected by
the presence of Mg . Since the chemical shifts of these protons are
largely determined by ring current effects, these spectral alterations
4975
by guest on September 16, 2016
http://nar.oxfordjournals.org/D
ownloaded from
Nucleic Acids Research
/ppm
Figure 1: The Effect of Kg** on A form 5S RNA. 5S RNA samples wereprepared 1n 0.1M KC1, 10 mM deuterotr1s-DCl, pH 7.6, D,0, 1n the presenceand absence of 2 mM MgCl_. Both samples were equinbrated with thei rbuffers by dialysis. 100a transients were accumulated on each at 300 K.The chemical sh i f t scales are set by reference to d1oxajj|.
Spectrum (a): 0.66 mM 5S RNA 1n the absence of MqSpectrum (b): 0.55 mM 5S RNA 1n the presence of HgSpectrum (c): The difference between spectra a and b. [Spectra
were scaled so that the difference spectrum 1n theregion upfield of HDO ( I . e . the sugar region) 1szero.]
suggest a change 1n the base stacking arrangements 1n the molecule.
Spectra obtained at 27° C, at neutral pH, and Mg4"*" concentrations above 2
mM are Indistinguishable from the one shown 1n Figure lb .
The State of the 5S RNA Preparations. RNA preparations at high
concentration can display 1on1c strength-dependent aggregation effects.
The possibi l i ty that aggregation might be responsible for the differences
1n NHR spectra shown 1n Figure 1 was examined by X-ray solution scattering.
A sample of 5S RNA was prepared 1n 0.1 M KC1, 1 mM MgClg, 10 mM Tr1s-HCl,
pH 7.5 and X-ray data collected on I t at a series of concentrations from
0.84 mM to 0.25 mM at both 25° C and 35° C. No temperature dependence
4976
by guest on September 16, 2016
http://nar.oxfordjournals.org/D
ownloaded from

Nucleic Acids Research
was noted. The radius of gyration at zero concentration was 34.7 ± 1 Ao
which compares favorably with published values of 36.1 ± 1 A (14) ando
34.5 ± 1.5 A (15). The observed radius f e l l with Increasing RNA concen-t rat ion.
At the same time as these measurements were done data were collectedon a sample of 50S subunits known to be monodisperse by ul t racentr i -fugation. Knowing the chemical compositions of both 50S subunits and 5SRNA, the ratios of their molecular weights can be estimated from theratios of their forward scattering Intensities per unit concentration.The forward scattering of 5S RNA, per unit concentration, extrapolated tozero concentration, Indicated a molecular weight of 44,000 which 1s thevalue expected, 40,000, within the error of this method. Again theconcentration dependence was such that the apparent molecular weight f e l lwith Increasing concentration.
A preparation of 5S RNA 1n 0.1 M NaCl, 0.01M EDTA, pH 5.6 1n D^ wasexamined at a single concentration (0.6 mM) at 25°, 35° and 45° C. Ata l l temperatures both I ts radius of gyration and forward scattering werethe same as those predicted from the data obtained with the Mg"1^ sample,w1 thin error.
These data Indicate that 5S RNA should be monodisperse under theconditions explored In this study and that changes In aggregation stateshould not be contributing to the spectroscopic effects seen.
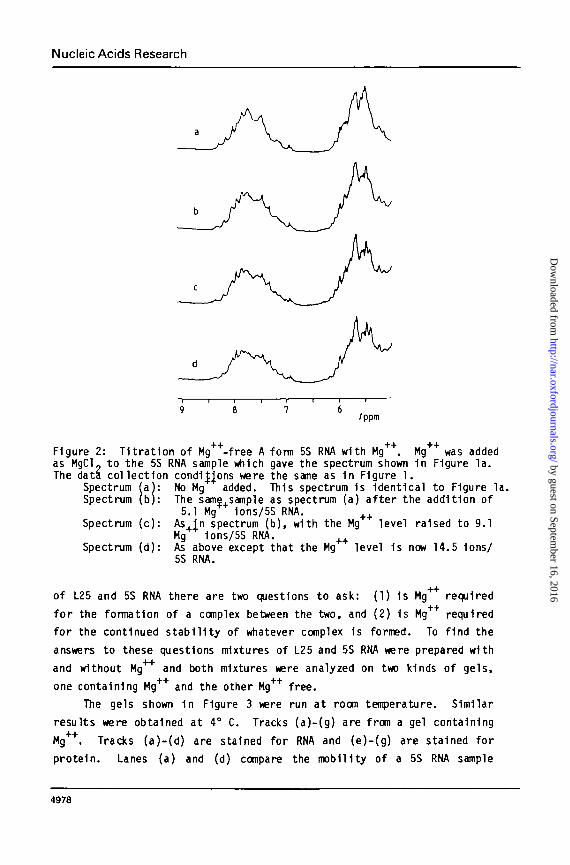
Interconversion of the Two Forms of 5S RNA. MgCl 2 was added tosamples which gave spectra l ike that of Figure la and spectra recorded atseveral ratios of Mg"*"1" Ions per RNA molecule. The results are shown 1nFigure 2. The top trace (a) shows a typical low Mg++ spectrum whichpersists unt i l the rat io of Mg*4 to RNA reaches about 5 Ions/molecule.At 5 1ons/molecule (trace b), small alterations 1n the spectrum becomeapparent which are quite pronounced at 9 Ions/molecule (trace c) , and by14 1ons/molecule (trace d), the conversion 1s complete. [Note that sinceHg 1s being added from the outside, these Hg levels are not equivalentto Mg 1on ac t i v i t i es . ] Thus the conversion of the low Mg form of themolecule to the high Mg form occurs within a relat ively narrow range ofMg44 1on concentration, suggestive of a cooperative transi t ion. Further-more, 1t 1s clear that the molecule 1s capable of making the transit ionat room temperature within a time which 1s short compared to the timerequired to collect NMR data (-1/2 hour).
L25 Binds to Both Forms of 5S RNA. With respect to the Interaction
4977
by guest on September 16, 2016
http://nar.oxfordjournals.org/D
ownloaded from
Nucleic Acids Research
/ppm
Figure 2: T1trat1on of Mg++-free A form 5S RNA with Mg++. Mg++ was addedas MgCl. to the 5S RNA sample which gave the spectrum shown 1n Figure la.The data col lection condi^ons were the same as 1n Figure 1.
Spectrum (a): No Mg added. This spectrum 1s Identical to Figure la.Spectrum (b): The same.sample as spectrum (a) after the addition of
5.1 Mg 1ons/5S RNA. + +Spectrum (c) : As In spectrum (b), with the Mg level raised to 9.1
Mg 1ons/5S RNA. ++Spectrum (d): As above except that the Mg level 1s now 14.5 Ions/
5S RNA.
of L25 and 5S RNA there are two questions to ask: (1) 1s Mg required
fo r the formation of a complex between the two, and (2) 1s Mg++ required
for the continued s tab i l i t y of whatever complex 1s formed. To f ind the
answers to these questions mixtures of L25 and 5S RNA were prepared with
and without Mg"1"1" and both mixtures were analyzed on two kinds of gels,
one containing Mg and the other Mg free.
The gels shown 1n Figure 3 were run at room temperature. Similar
results were obtained at 4° C. Tracks (a)-(g) are from a gel containing
Mg"1"1". Tracks (a)-(d) are stained for RNA and (e)-(g) are stained for
protein. Lanes (a) and (d) compare the mobility of a 5S RNA sample
4978
by guest on September 16, 2016
http://nar.oxfordjournals.org/D
ownloaded from

Nucleic Acids Research
b e d e f 9 h i j k I m n
Figure 3: Th^ Protein Binding Properties of 5S RNA 1n the Presence andAbsence of Mg . Acrylamide gels were run to analyze 5S RNA samples andmixtures of 5S RNA and L25. Two gels are shown here: tracks a-g are froma gel run 1n the+Dresence of 5 mM Mg ; tracks h-n are from a gel run 1nthe absence of Mg . The conditions fo r these gels were otherwise as given1n Materials & Methods. The temperature during these runs was about 23° C.The samples 1n each case contained about 0.3 Aprn m of RNA and 7 pg of L25per sample of 40 u l . The 1on1c conditions 1n rne samples ware those of thegel running buffer unless otherwise noted. Samples were kept at 0° C pr io rto loading on the ge ls > + +
Fi rs t gel (5 mM Mg ): Tracks a-d are stained with methylene blue toshow RNA. Tracks e-g are stained with Coomassie blue to show prote in , (a)5S SNA alone, (b) 5S RNA with L25, (c) 5S RNA, previously dialyzed to removeMg , plus L25, (d) 5S RNA, I n i t i a l l y without Mg , no L25, (e) same as (b ) ,( f ) , same as ( c ) , (g) same as (a) .
Second gel (0 Mg++): Tracks h-k are stained with methylene blue.Tracks .1-n are stained with Coomassje blue, (h) 5S plus L25 and Mg++,(1) Mg - f ree 5S plus L25, ( j ) Mg - f ree 5S plus 2 yg L25, (k) 5S RNAwith Mg , no prote in , (1) same as (h ) , (m) same as (1) , (n) same as ( j ) .
++ ++containing Mg with that of a sample which was Mg free at the time 1t
was loaded on the ge l . Lanes (b) and (c) show corresponding samples
containing L25. The reduced mobi l i ty of the RNA 1n (b) and (c) 1s due to
of complex formation as proven by tracks (e) and ( f ) which show the
presence of protein 1n those bands. Lane (g) Is a sample of protein-f ree
5S RNA stained for prote in.
Lanes (h)-(n) 1n Figure 3 repeat the observations of tracks (a)-(g)
4979
by guest on September 16, 2016
http://nar.oxfordjournals.org/D
ownloaded from
Nucleic Acids Research
1n gels which lack Mg 1on. Tracks (h) and (1) show a mixture of 5S
RKA, Mg and a saturating amount of L25 stained for RNA, ( h ) , and
protein, ( 1 ) . Tracks (1) and (DI) are the corresponding experiments 1n the
absence of Mg . Track ( j ) shows Mg free 5S RNA mixed with a less than
saturating quantity of L25 and track (k) 1s 5S RNA with no L25 added,
I n i t i a l l y 1n the presence of Mg++.
Clearly Mg++-free and Mg++ containing samples of A form RNA cannot
be distinguished by gel electrophoresis under these conditions consistent
with the view that 1nterconvers1on between the two forms 1s fast compared
to the time of electrophoresis. Moreover, both forms bind to L25 regard-
less of the presence or absence of Mg In the binding mixture or 1n the
analyzing gel subsequently. Kg"1"1" 1s not required for binding or s t a b i l i -
zation of the complex. This la t te r observation contradicts the finding
of Spierer and Zimmermann (16) that there 1s a Mg++ requirement for this
Interact ion, but 1s 1n accord with other more recent observations (3 ) .
I t 1s possible that differences 1n the binding assay methods used may be
the cause of the disagreement with Spierer and Zimmermann, 1n addition
there are some small differences 1n the 1on1c conditions examined.
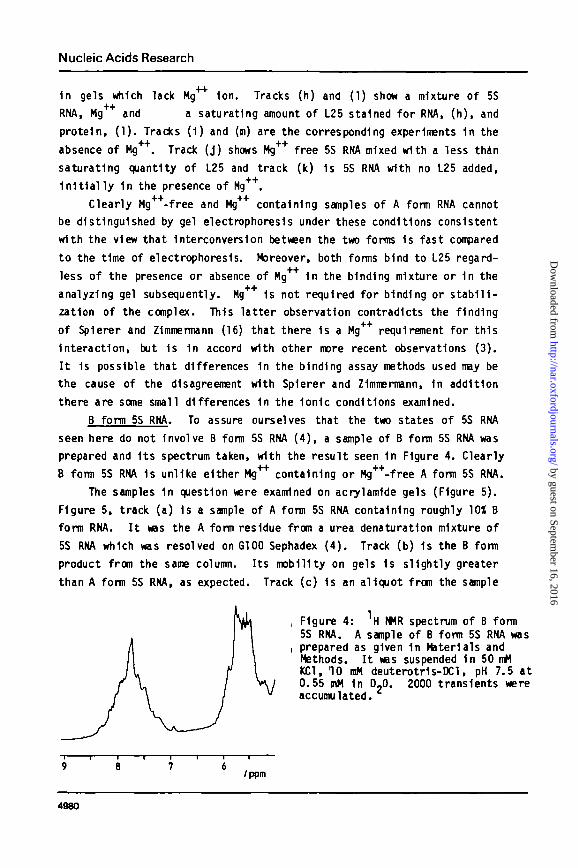
B form 5S RNA. To assure ourselves that the two states of 5S RNA
seen here do not Involve B form 5S RNA ( 4 ) , a sample of B form 5S RNA was
prepared and I t s spectrum taken, with the result seen 1n Figure 4. Clearly
B form 5S RNA 1s unlike either Mg'1"1' containing or Mg^-free A form 5S RNA.
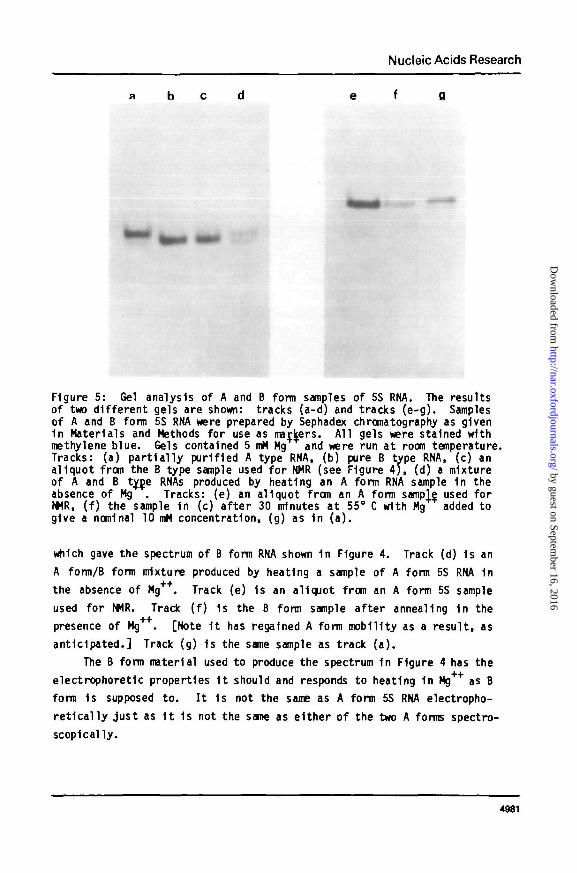
The samples 1n question were examined on acrylamide gels (Figure 5 ) .
Figure 5, track (a) 1s a sample of A form 5S RNA containing roughly 10X B
form RNA. I t was the A form residue from a urea denaturation mixture of
5S RNA which was resolved on G100 Sephadex ( 4 ) . Track (b) 1s the B form
product from the same column. I ts mobility on gels 1s sl ight ly greater
than A form 5S RNA, as expected. Track (c) 1s an aliquot from the sample
, Figure 4: H WR spectrum of B form5S RNA. A sample of B form 5S RNA was
i prepared as given 1n Materials andMethods. I t was suspended 1n 50 mMKC1, 10 mM deuterotr1s-DCl, pH 7.5 at0.55 mM 1n D?0. 2000 transients wereaccumulated.
/ppm
4980
by guest on September 16, 2016
http://nar.oxfordjournals.org/D
ownloaded from
Nucleic Acids Research
Figure 5: Gel analysis of A and B form samples of 5S RNA. The resultsof two di f ferent gels are shown: tracks (a-d) and tracks (e -g) . Samplesof A and B form 5S RNA were prepared by Sephadex chromatography as given1n Materials and Methods for use as markers. All gels were stained withmethylene blue. Gels contained 5 mM Mg and were run at room temperature.Tracks: (a) pa r t i a l l y purified A type RNA, (b) pure B type RNA, (c) analiquot from the B type sample used for NMR (see Figure 4 ) , (d) a mixtureof A and B ty_D.e RNAs produced by heating an A form RNA sample 1n theabsence of Mg . Tracks: (e) an aliquot from an A form sampVe used forNMR, ( f ) the sample 1n (c) a f te r 30 minutes at 55° C with Mg added togive a nominal 10 mM concentration, (g) as 1n ( a ) .
which gave the spectrum of B form RNA shown 1n Figure 4. Track (d) 1s an
A form/B form mixture produced by heating a sample of A form 5S RNA 1n
the absence of Mg++. Track (e) Is an aliquot from an A form 5S sample
used for NMR. Track ( f ) 1s the B form sample af ter annealing In the
presence of Mg . [Note I t has regained A form mobility as a resul t , as
ant ic ipated. ] Track (g) 1s the same sample as track ( a ) .
The B form material used to produce the spectrum 1n Figure 4 has the
electrophoretic properties I t should and responds to heating 1n Mg++ as B
form 1s supposed to . I t 1s not the same as A form 5S RNA electropho-
r e t i c a l l y jus t as I t 1s not the same as e i ther of the two A forms spectro-
scopically.
4981
by guest on September 16, 2016
http://nar.oxfordjournals.org/D
ownloaded from

Nucleic Acids Research
DISCUSSION
In 1980 Kao and Crothers reported the existence of a low temperature
melting transit ion 1n 5S RNA (1). Its salient features are that 1t
occurs 1n buffers of moderate 1on1c strength and neutral pH, at tempe-
ratures close to room temperature, that 1t 1s fast, having a half time of
milliseconds, and that the high temperature form of the molecule 1s
stablized by Mg++ 1 on.Recent t r i t i um exchange results also point to the
existence of a Mg dependent conformational change 1n 5S RNA (17).
The two states of 5S RNA presented 1n this study behave as would be
expected i f they corresponded to those Identified by Kao and Crothers
(1). The transit ion between them 1s Induced by Mg 1on at room tempe-
rature, and the 1nterconvers1on between them 1s fast . An attempt was
made to Induce the same conformational change by raising the temperature.
However under the conditions tested, the material converted Into the B
form upon heating rather than Into the high Mg form hoped for . Given
the sensi t iv i ty of the Kao-Crothers transit ion to 1on1c conditions 1t 1s
possible that under dif ferent conditions of pH and 1on1c strength, the
appropriate transit ion would be observed.
Several kinds of evidence Indicate that the two A states examined
here have nothing to do with the classical B form of 5S RNA. F i rs t , both
forms have NMR spectra d is t inc t ly dif ferent from that of B form 5S RNA.
Second, neither coelectrophoreses with B form RNA. Third, the rate of
1nterconvers1on between these two forms at room temperature 1s orders of
magnitude faster than the A to B conversion which under the conditions
Investigated here should take days (19). I t 1s not possible k inet ica l ly
that one form could be the A form and the other the B form.
We suggest that the two conformers of A type 5S RNA f i r s t recognized
by Kao & Crothers (1), and Identi f ied here by NMR spectroscopy, be called
"H" and "L". The H form would be that which 1s favored by high tempera-
ture and high Mg concentration, and the L form would be the one which
1s stable at lower temperatures 1n the absence of Mg .
Preliminary data on the NMR spectra of complexes of 5S RNA and L25
show that the mode of binding of L25 to the H and L forms 1s the same and
that L25 has no effect on the i r 1nterconvers1on (Moore, unpublished
data). Furthermore, part ial digestion experiments done with RNase A show
that the large RNase resistant fragment Isolatable from H form 5S RNA
(13) can also be Isolated from the L form (Chabot 4 Moore, unpublished
observation). This fragment comprises nucleotides 1-11, 69-87 and 89-120
1n the Intact 5S molecule and Includes the L25 binding s i te (13). Thus
4982
by guest on September 16, 2016
http://nar.oxfordjournals.org/D
ownloaded from

Nucleic Acids Research
1t would appear that the spectral changes observed here must relate
either to 5S tertiary interactions which do not affect L25 binding or
RNAse resistence or be localized 1n the 12-68 loop. Much more work wil l
have to be done to further Identify the part of the molecule where these
changes are taking place.
Since, as Kao and Crothers (1) pointed out, the two states, H and L,
Interconvert under conditions very close to physiological, the possi-
b i l i ty exists that the transition between them 1s related to 5S RNA
function. Thus the characterization of the transition could turn out to
be of more than passing Interest.
ACKNOWLEDGEMENTS
I wish to thank Drs. D.M. Crothers and J.M. Prestegard f o r t h e i r
advice and encouragement. Darcy Fazio and Bet ty Rennie con t r i bu ted
exper t technica l ass is tance . Gregory Dalach and El izabeth Roche con-
t r i b u t e d to the development o f the work. NMR spectroscopy was done a t
the NSF Northeast Regional NMR F a c i l i t y (Grant CHE-791621O) and the a id
of Mr. Peter Demou. This research was supported by a grant from t he NIH
(AI-09167).
REFERENCES
1 . Kao, T.H. & Crothers , D.M. (1980) Proc. Nat. Acad. Sc1. USA TT_,3360-3364.
2. Dohme, F. & Nierhaus, K.H. (1976) Proc. Nat. Acad. USA 73, 2221-2225.3. Douthwaite, S . , G a r r e t t , R.A. Wagner, R. & Feuteun, J . "(T979)
Nucleic Adds Res. 6, 2454-2470.4. Aubert , M., Sco t t , T . F . , Reynier, M. & Monier, R. (1968) Proc. Nat.
Acad. Sc1. USA 6 1 , 292-299.5. P h i l l i p s , G.P. JTr imko, J . L . (1972) Ana ly t . Biochem. 45_, 319-325.6. Chen-Schme1sser, V. & G a r r e t t , R.A. (1977) FEBS Le t te rs 74, 287-291.7. Lecanidou, R. & Richards, E.G. (1975) Eur. J . Biochem. 5?7 127-133.8. Moore, P.B. (1979) Methods I n Enzymol. 59, 639-655.9. Osterberg, R., S jdberg , B. & G a r r e t t , R7A~. (1976) Eur. J . Biochem. £ 8 ,
481-487.10. Zimmerman, R.A. (1979) Methods 1n Enzymol. 59, 551-583.1 1 . K1me, M . J . , R a t c l i f f e , R.G., Moore, P.B. &¥T l l1ams, R.J.P. (1981)
Eur. J . Biochem. 116, 269-276.12. McDonald, R.C., SteTtz , T.A. & Engelman, D.M. (1979) Biochemistry
Jj5, 338-342.13. Moore, P.B. (1980) J . App l . Crys t 1 3 , 168-175.14. Osterberg, R., Sj f lberg, B. & G a r r e t t , R.A. (1976) Eur. J . Biochem.
6j i , 481-487.15. Conners, P.G. & Beeman, W.W. (1972) J . Mol . B1o l . 7 1 , 31-37.16. Sp ierer , P. & Zimmermann, R.A. (1978) B1ocheffl1stry"T_7, 2474-2479.17. Ramstein, J . & Erdmann, V.A. (1981) Nuc. Adds Res. 9 , 4081-4086.18. G a r r e t t , R.A., Douthwaite, S. i N o l l e r , H.F. (1981) T . I . B . S . 6 , 137-139.19. Richards, E.G., Lecanidou, R. & Geroch, M.E. (1973) Eur. J . BTochem.
34, 262-267.
4983
by guest on September 16, 2016
http://nar.oxfordjournals.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















