
Role of plant symbionts in nematode management: A review
10
ELSEVIER 0960-8524(95)00137-9 Bioresource Technology 54 (1995) 217-226 © 1996 Elsevier Science Limited Printed in Great Britain. All fights reserved 0960-8524/95 $9.50+0.00 ROLE OF PLANT SYMBIONTS IN NEMATODE MANAGEMENT: A REVIEW Zaki A. Siddiqui* & Irshad Mahmood Section of Plant Pathology and Nematology, Department of Botany, Aligarh Muslim University, Aligarh-202 002, India (Received 25 April 1995; revised version received 2 September 1995; accepted 6 September 1995) Abstract Mycorrhizal fungi increase soil nutrient and water absorption as plant symbionts. Root nodule bacteria, beside fixing atmospheric nitrogen, have the ability to produce antibiotics and phytoalexins, etc. The use of these two symbionts together appears to be more bene- ficial for plant growth than their use individually. The results of most studies indicate that mycorrhizal fungi and root-nodule bacteria generally reduce the severity of nematode diseases of various crops. There are possi- bilities for biological control of nematodes by selecting effective strains of mycorrhizal fungi and root-nodule bacteria, despite some obstacles. Key words: Mycorrhizal fungi, nematode manage- ment, plant symbionts, root-nodule bacteria. INTRODUCTION Lewin (1982) defined symbionts as lodgers who pay rent and parasites as freeloaders who do not. Rh/zo- bium and mycorrhizal fungi are the two prime examples of symbionts. Generally, Rhizobium induce symbiosis only in legume hosts, while mycorrhizal fungi have this association with about 90% of vas- cular and economically important plants. The roles of these symbionts will be discussed in this review in relation to nematode management. Mycorrhizai fungi Albert Bernard Frank in 1885 (Frank, 1885) intro- duced the Greek word 'mycorrhiza', which literally means 'fungus roots'. Mycorrhizal fungi form a sym- biotic relationship with plant roots in a similar fashion to root-nodule bacteria in legumes. The fun- gus takes carbohydrates from the plant and in return supplies the plant with nutrients, hormones, etc. There are four major types of mycorrhizae: (i) ecto- mycorrhiza, (ii) vesicular-arbuscular mycorrhiza (VAM), (iii) ericoid mycorrhiza, (iv) orchidaceous mycorrhiza. Ecto-mycorrhiza are commonly found on the roots of forest trees and form a thick mantle covering the outside of the roots. Inside the roots, * Author to whom correspondence should be addressed. 217 they are present in intercellular spaces and form a characteristic structure called 'harting net'. Ericoid mycorrhizae are associated with heath plants like blueberries, azaleas, Rhododendron, etc. Orchida- ceous mycorrhizal fungi are associated with orchids. Vesicular-arbuscular mycorrhizal (VAM) fungi are found to occur on roots of most of the food and horticultural crops and tropical trees. The presence of vesicles and arbuscules is the diagnostic criterion for identifying VAM fungi in a root. VAM fungi improve plant growth through improved uptake of nutrients, especially phosphorus. They also improve uptake of minor elements like zinc and copper and of water. They also produce plant hormones and increase the activity of nitrogen-fixing organisms. VAM fungi have been observed in 1000 genera of plants, representing some 200 families (Bagyaraj, 1991). There are at least 300 000 receptive hosts in the world flora. According to Gerdemann (1975), it is easier to list plant families that do not form VAM than to list those that do. All the VAM fungi are grouped in a single family, the Endogonaceae. The family is considered to be in an order of its own, the Endogonales, tentatively placed in the class Zygomy- cetes in the subdivision Zygomycotina (Benjamin, 1979). The seven genera, Endogone, Glomus, Scler- ocystis, Entrophospora, Acaulospora, Scutellospora and Gigaspora, presently constitute Endogonaceae. Of these only Endogone form zygospores, while others probably lack sexual reproduction. Species of Endogone do not form VAM associations. The other six genera form VAM associations. Root-nodulating bacteria Symbiotic nitrogen fixation is not confined to the legumes only, some non-legume angiosperms also form root-nodules when invaded by soil actinomy- cetes (Callaham et aL, 1978). Allen and Allen (1947) showed that 11.8% of Mimosoideae, 3.3% of Cae- salpinioideae and 84-9% of Papilionoideae had nodules. However, another estimate showed that the nodulation status of legumes ranges from 25 to 30% for Caesalpinioideae, 60-70% for Mimosoideae and 90-95% for Papilionoideae (Lin & Burton, 1982). The ability of legumes and non-legumes to draw
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Role of plant symbionts in nematode management: A review

ELSEVIER 0 9 6 0 - 8 5 2 4 ( 9 5 ) 0 0 1 3 7 - 9
Bioresource Technology 54 (1995) 217-226 © 1996 Elsevier Science Limited
Printed in Great Britain. All fights reserved 0960-8524/95 $9.50 +0.00
ROLE OF PLANT SYMBIONTS IN NEMATODE MANAGEMENT: A REVIEW
Zaki A. Siddiqui* & Irshad Mahmood
Section of Plant Pathology and Nematology, Department of Botany, Aligarh Muslim University, Aligarh-202 002, India
(Received 25 April 1995; revised version received 2 September 1995; accepted 6 September 1995)
Abstract Mycorrhizal fungi increase soil nutrient and water absorption as plant symbionts. Root nodule bacteria, beside fixing atmospheric nitrogen, have the ability to produce antibiotics and phytoalexins, etc. The use of these two symbionts together appears to be more bene- ficial for plant growth than their use individually. The results of most studies indicate that mycorrhizal fungi and root-nodule bacteria generally reduce the severity of nematode diseases of various crops. There are possi- bilities for biological control of nematodes by selecting effective strains of mycorrhizal fungi and root-nodule bacteria, despite some obstacles.
Key words: Mycorrhizal fungi, nematode manage- ment, plant symbionts, root-nodule bacteria.
INTRODUCTION
Lewin (1982) defined symbionts as lodgers who pay rent and parasites as freeloaders who do not. Rh/zo- bium and mycorrhizal fungi are the two prime examples of symbionts. Generally, Rhizobium induce symbiosis only in legume hosts, while mycorrhizal fungi have this association with about 90% of vas- cular and economically important plants. The roles of these symbionts will be discussed in this review in relation to nematode management.
Mycorrhizai fungi Albert Bernard Frank in 1885 (Frank, 1885) intro- duced the Greek word 'mycorrhiza', which literally means 'fungus roots'. Mycorrhizal fungi form a sym- biotic relationship with plant roots in a similar fashion to root-nodule bacteria in legumes. The fun- gus takes carbohydrates from the plant and in return supplies the plant with nutrients, hormones, etc. There are four major types of mycorrhizae: (i) ecto- mycorrhiza, (ii) vesicular-arbuscular mycorrhiza (VAM), (iii) ericoid mycorrhiza, (iv) orchidaceous mycorrhiza. Ecto-mycorrhiza are commonly found on the roots of forest trees and form a thick mantle covering the outside of the roots. Inside the roots,
* Author to whom correspondence should be addressed. 217
they are present in intercellular spaces and form a characteristic structure called 'harting net'. Ericoid mycorrhizae are associated with heath plants like blueberries, azaleas, Rhododendron, etc. Orchida- ceous mycorrhizal fungi are associated with orchids.
Vesicular-arbuscular mycorrhizal (VAM) fungi are found to occur on roots of most of the food and horticultural crops and tropical trees. The presence of vesicles and arbuscules is the diagnostic criterion for identifying VAM fungi in a root. VAM fungi improve plant growth through improved uptake of nutrients, especially phosphorus. They also improve uptake of minor elements like zinc and copper and of water. They also produce plant hormones and increase the activity of nitrogen-fixing organisms. VAM fungi have been observed in 1000 genera of plants, representing some 200 families (Bagyaraj, 1991). There are at least 300 000 receptive hosts in the world flora. According to Gerdemann (1975), it is easier to list plant families that do not form VAM than to list those that do. All the VAM fungi are grouped in a single family, the Endogonaceae. The family is considered to be in an order of its own, the Endogonales, tentatively placed in the class Zygomy- cetes in the subdivision Zygomycotina (Benjamin, 1979). The seven genera, Endogone, Glomus, Scler- ocystis, Entrophospora, Acaulospora, Scutellospora and Gigaspora, presently constitute Endogonaceae. Of these only Endogone form zygospores, while others probably lack sexual reproduction. Species of Endogone do not form VAM associations. The other six genera form VAM associations.
Root-nodulating bacteria Symbiotic nitrogen fixation is not confined to the legumes only, some non-legume angiosperms also form root-nodules when invaded by soil actinomy- cetes (Callaham et aL, 1978). Allen and Allen (1947) showed that 11.8% of Mimosoideae, 3.3% of Cae- salpinioideae and 84-9% of Papilionoideae had nodules. However, another estimate showed that the nodulation status of legumes ranges from 25 to 30% for Caesalpinioideae, 60-70% for Mimosoideae and 90-95% for Papilionoideae (Lin & Burton, 1982). The ability of legumes and non-legumes to draw

218 Z. A. Siddiqui, I. Mahmood
atmospheric nitrogen is dependent on the presence of soil microorganisms of particular groups; Rhizo- biacea (Jordan, 1984) and Frankia (Lechevalier & Lechevalier, 1989). The rhizobiaceae includes two genera of root-nodule bacteria; Rhizobium Frank 1889 and Bradyrhizobium Jordan 1982. Members of the genus Rhizobium produce fast growth on yeast extract-mannitol agar and acid reaction in mineral salt-mannitol medium. Members of the genus Brady- rhizobium produce slow growth and alkaline reaction in these media. Rhizobia are aerobic rods and non- spore-forming. Frankia Brunchorst 1886 also readily infect suitable host plants, giving rise to nitrogen- fixing nodules on the roots (Callaham et al., 1978). Legumes stimulate rhizobia much more than other rhizosphere microorganisms (Nutman, 1965). Rhizo- bial nodulation is a complex symbiotic process between host plant and rhizobia. The plant provides an energy source and ecological niche for the bacteria and the bacteria provide a source of fixed nitrogen for the plant (Vance & Johnson, 1981).
INTERACTION OF MYCORRHIZAL FUNGI AND NEMATODES
Mycorrhizal fungi and plant parasitic nematodes are commonly found inhabiting the rhizosphere and colonizing the roots of their host plants. These two groups of microorganisms exert opposite effects on plant growth (Hussey & Roncadori, 1982; Schenck, 1981). A large number of reviews and chapters have appeared on the interaction of nematode and mycorrhizal fungi (Bagyaraj, 1984; Jalali & Jalali, 1991; Khan, 1993; Paulitz & Linderman, 1991). As a result of interactions, in general, the severity of nematode diseases is reduced in mycorrhizal plants (Table 1). However, in some instances root coloniza- tion by mycorrhizal fungi had no effect on nematode reproduction (Table 2), while it was even found to be stimulated in some cases (Table 3).
Reasons for reduction of nematode reproduction by mycorrhizal fungi Some of the possible reasons of reduced nematode reproduction caused by mycorrhizal fungi are as fol- lows.
Nematode parasitism by VAM fungi In some studies, parasitism of nematodes eggs with VAM fungus has been demonstrated, but the level of parasitism was not considered sufficient to nega- tively affect nematode activities (Francl & Dropkin, 1985). VAM fungal chlamydospores have been reported to occupy seeds and dead insects in soil (Rabatin & Rhodes, 1982; Taber, 1982) and have limited saprophytic capabilities (Harley & Smith, 1983). It seems likely that the VAM fungi colonize only stressed or weakened nematode eggs. The nematode parasitism by VAM fungi is opportunistic
and depends on carbon nutrition from autotrophic symbionts, rather than being representative of a true host-parasite relationship.
Changes in root morphology The colonization by mycorrhizal fungi as host-symbi- ont results in morphological changes, leading to formation of an ecto-, ericoid-, orchidaceous- or vesicular-arbuscular-mycorrhizal relationship. In the case of ectomycorrhizae, the plant root morphology changes dramatically from the non-mycorrhizal state, resulting in loss of root hairs, the development of a thick fungal mantle surrounding the epidermis and cortex and the development of extensive root branching. The increased root branching and hyphal network greatly increase the absorptive surface area (Linderman, 1988). The development of endomycor- rhizae results in loss of root hairs, but no external hyphal mantle forms, except for the relatively sparse external hyphae that occur at the rhizoplane and extend out into the surrounding soil. Roots offer structural support to the plant, function in absorp- tion of water and mineral nutrients, are the site of production of growth regulaters, are a site of starch storage and provide a nutrient supply for a wide range of microorganisms (Curl & Truelove, 1986; Rovira, 1985). Change in root morphology will ulti- mately affect their responses to other organisms. Dehne (1982) found that mycorrhizal fungi reduced disease response to plant pathogens, due to some morphological changes in the plant. Mycorrhizal plants also have a strong vascular system, which imparts greater mechanical strength to diminish the effects of pathogens (Schonbeck, 1979).
Histopathological changes Histopathological studies on galls caused by the nematode Meloidogyne ineognita showed that galls in mycorrhizal plants had fewer of the giant cells and syncytia, which are needed for the development of nematode larvae, than did non-mycorrhizal plants. Nematodes in mycorrhizal plants were smaller and took a longer time to develop into adults (Suresh, 1980). Smaller syncytia and fewer giant cells have been reported to confer resistance against nema- todes on the host plant (Trudgill & Parrott, 1969; Fassuliotis, 1970).
Physiological and biochemical changes The physiological changes caused by mycorrhizal fungi in the host plant generally reduce the severity of nematode diseases (Dehne, 1982). Phenolic com- pounds have been shown to be formed after mycorrhizal colonization (Sylvia & Sinclair, 1983) and have been thought to play a role in disease resistance (Goodman et al., 1967). Production of phytoalexin was greater on mycorrhizal roots than on non-mycorrhizal roots (Morandi, 1987) and phytoalexin is believed to play a major role in the host defenee system against pathogens (Kaplan et

Plant symbiotics in nematode control: a review 219
al., 1980). An increase in lignin and phenols in mycorrhizal plants was observed and was associated with reduced nematode reproduction (Singh et al.,
1990; Umesh et al., 1988). Increased phenylalanine and serine in tomato roots due to inoculation with VAM fungi has been observed (Suresh, 1980). These
Table 1. Mycorrhizal fungi had adverse effect on nematode reproduction
Host plant Nematode Mycorrhizal ~ngi Effect of interaction on nematodes Reference
Tobacco Meloidogyne incognita Endogone mosseae
Tobacco, M. incognita M. hapla E. mosseae oat, tomato
Tomato M. incognita Glomus mosseae
Tomato M. incognita M. javanica Glomus fasciculatum
Citrus Tylenchulus Glomus mosseae semipenetrans
Soybean M. incognita Glomus macrocarpum
Tomato Rotylenchulus Glomus fasciculatum reniformis
Cotton M. incognita Glomus etunicatum G. mosseae
Peach M. incognita Gigaspora n'uugarita Cotton M. incognita Glomus fasciculatum
Cotton M. incognita Glomus intraradices
Cotton M. incognita
Alfalfa M. hapla
Tomato, M. hapla white clover
Bean M. javanica
Tamarillo M. incognita
Cowpea Heterodera cajani
Tomato M. javanica
Pine Pratylencus penetrans Chickpea M. javanica
Cowpea Heterodera cajani
Banana Radopholus similis
G. intraradices
Mixed inoculum
Mixed inoculum
Glomus etunicatum
Mixed inoculum
Glomus fasciculatum
Glomus fasciculatum
Ectomycorrhizae Glomus manihotis Gigaspora margarita Gigaspora gigantea
Glomus fasciculatum G. epigaeus
Glomus fasciculatura
Citrus Radopholus citrophilus Glomus intraradices Soybean M. incognita Gigaspora margarita Pepper M. incognita Glomus fasciculatum
G. etunicatum
Black pepper M. incognita
Soybean Heterodera glycines
Tomato M. javanica Pigeonpea Heterodera cajani Pigeonpea H. cajani
Glomus mosseae Acaulospora laevis Glomu~ fasciculatum Gigaspora margarita Mixed inoculum
Glomus fasciculatum Glomus fasciculatum Gigaspora margarita
Had antagonistic effect on nematode population Fewer nematode larvae develop into adults in plants pre-inoculated with E. mosseae Nematode development was reduced by mycorrhizal colonization Reduced number and size of root-knot galls on mycorrhizal plants Disease symptoms were less severe on mycorrhizal plants Reduced number of galls per gram root caused by nematodes Adversally affected nematode reproduction Nematode eggs per gram root was lower on mycorrhizal plants Suppressed nematode reproduction Sixty or more spores of VAM fungi had adverse effect on nematode reproduction Adverse effect on nematode reproduction at both intervals (planting and after 28 days) Nematode population was reduced after 28 days of inoculation Reduced nematode number and adult development Number of J2 and adults per gram of root were always lower in mycorrhizal treatments Severity of nematode disease is reduced on mycorrhizal plants Suppress reproduction and development of nematodes Adverse effect on nematode reproduction Reduced nematode reproduction in plants pre-inoculated with VAM Penetration was inhibited Reproduction was differentially suppressed, most pronounced with G. manihotis, less with G. margarita and slightly with G. gigantea Prior inoculation of VAM fungi adversely affected nematode penetration more than simultaneous inoculation Nematodes were lower on mycorrhizal plants when applied simultaneously for 7 days after mycorrhizal inoculation Reduced nematode reproduction Suppressed nematode reproduction Significantly reduce galling and nematode population when pre-inoculated with VAM fungi Individually all the VAM fungi reduced nematode reproduction but the highest reduction wa s caused by A. laevis Increase tolerance to nematode in greenhouse experiment and outdoor microplots Nematode population was inhibited Reduced nematode multiplication Adverse effect on nematode reproduction
Baltruschat et al. (1973)
Sikora & Schonbeck (1975)
Sikora (1978)
Bagyaraj et al. (1979)
O'Bannon et al. (1979)
Kellam & Schenck (1980)
Sitaramaiah & Sikora (1982)
Hussey & Roncadori (1982)
Strobel et al. (1982) Saleh & Sikora (1984)
Smith et al. (1986a)
Smith et al. (1986b)
Grandison & Cooper (1986)
Cooper & Grandison (1986)
Zombolin & Oliveira (1986)
Cooper & Grandison (1987)
Jain & Sethi (1987)
Morandi (1987)
Marks et al. (1987) Diederichs (1987)
Jain&Sethi(1988)
UmeshetaL (1988)
Smith & Kaplan (1988) Carling et al. (1989) Sivaprasad et al. (1990)
Anandaraj et al. (1990)
Tylka et al. (1991)
Sundarababu et al. (1993) Siddiqui & Mahmood (1995a) Siddiqui & Mahmood (1995b)
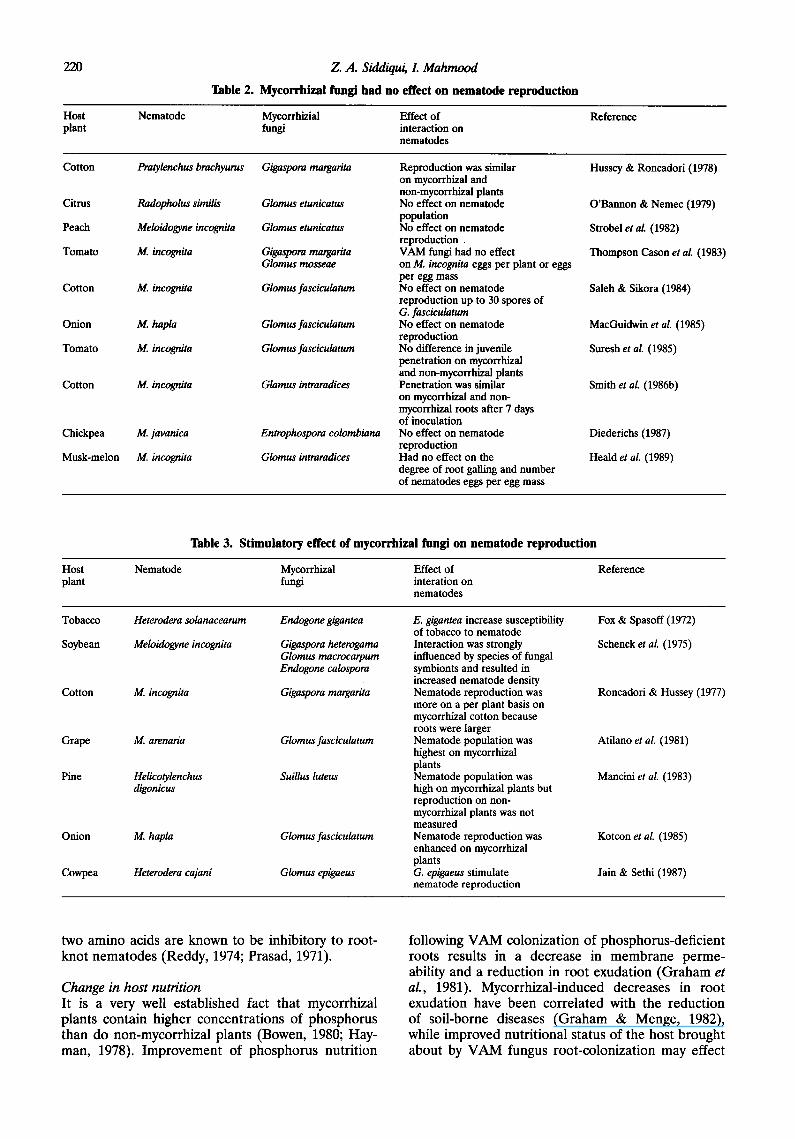
220 Z. A. Siddiqui, L Mahmood
Table 2. Mycorrhizal fungi had no effect on nematode reproduction
Host Nematode Myeorrhizial Effect of plant fungi interaction on
nematodes
Reference
Cotton Pratylenchus brachyurus Gigaspora margarita
Citrus Radopholus similis Glomus etunicatus
Peach Meloidogyne incognita Glomus etunicatus
Tomato M. incognita Gigaspora margarita Glomus masseae
Cotton M. incognita Glomus fasciculatum
Onion M. hapla Glomus fasciculatum
Tomato M. incognita Glomus fasciculatum
Cotton M. incognita Glamus intraradices
Chickpea M. javanica Entrophospora colombiana
Musk-melon M. incognita Glomus intraradices
Reproduction was similar on mycorrhizal and non-myeorrhizal plants No effect on nematode population No effect on nematode reproduction VAM fungi had no effect on M. incognita eggs per plant or eggs per egg mass No effect on nematode reproduction up to 30 spores of G. fasciculatum No effect on nematode reproduction No difference in juvenile penetration on mycorrhizal and non-mycorrhizal plants Penetration was similar on mycorrhizal and non- mycorrhizal roots after 7 days of inoculation No effect on nematode reproduction Had no effect on the degree of root galling and number of nematodes eggs per egg mass
Hussey & Roncadori (1978)
O'Bannon & Nemec (1979)
Strobel et al. (1982)
Thompson Cason et al. (1983)
Saleh & Sikora (1984)
MacOuidwin et al. (1985)
Suresh et al. (1985)
Smith et al. (1986b)
Diederichs (1987)
Heald et al. (1989)
Table 3. Stimulatory effect of mycorrhizal fungi on nematode reproduction
Host Nematode Mycorrhizal Effect of plant fungi interation on
nematodes
Reference
Tobacco Heterodera solanacearum Endogone gigantea
Soybean Meloidogyne incognita Gigaspora heterogama Glomus macrocarpum Endogone calospora
Cotton M. incognita Gigaspora margarita
Grape M. arenaria Glomus fasciculatum
Pine Helicotylenchus Suillus luteus digonicus
Onion M. hapla Glomus fasciculatum
Cowpea Heterodera cajani Glomus epigaeus
E. gigantea increase susceptibility of tobacco to nematode Interaction was strongly influenced by species of fungal symbionts and resulted in increased nematode density Nematode reproduction was more on a per plant basis on mycorrhizal cotton because roots were larger Nematode population was highest on mycorrhizal plants Nematode population was high on mycorrhizal plants but reproduction on non- mycorrhizal plants was not measured Nematode reproduction was enhanced on mycorrhizal plants G. epigaeus stimulate nematode reproduction
Fox & Spasoff (1972)
Schenck et aL (1975)
Roncadori & Hussey (1977)
Atilano et al. (1981)
Mancini et al. (1983)
Kotcon et al. (1985)
Jain & Sethi (1987)
two amino acids are known to be inhibitory to root- knot nematodes (Reddy, 1974; Prasad, 1971).
Change in host nutrition It is a very well established fact that mycorrhizal plants contain higher concentrations of phosphorus than do non-mycorrhizal plants (Bowen, 1980; Hay- man, 1978). Improvement of phosphorus nutrition
following VAM colonization of phosphorus-deficient roots results in a decrease in membrane perme- ability and a reduction in root exudation (Graham et al., 1981). Mycorrhizal-induced decreases in root exudation have been correlated with the reduction of soil-borne diseases (Graham & Menge, 1982), while improved nutritional status of the host brought about by VAM fungus root-colonization may effect

Plant symbiotics in nematode control: a review 221
quantitative and qualitative changes in root exudates (Linderman, 1985; Reid, 1984).
The severity of nematode damage of cotton was greater on P-fertilized, non-mycorrhizal plants than mycorrhizal plants at a P level deemed adequate to high for cotton (Thompson Cason et al., 1983). This was attributed to zinc deficiency induced by nema- tode infection at high soil-P levels. High levels of P fertilization inhibit zinc uptake (Mengel & Kirkby, 1979) but apparently VAM fungi can alleviate this P-induced zinc deficiency and thus increase host tol- erance to nematode parasitism. VAM fungi have been shown to affect changes induced by environ- mental stress in root growth, root exudation, nutrient absorption and host physiology (Smith, 1988). Many of these factors are related to physiol- ogy and nutrition (Graham et al., 1981; Hayman, 1982). Changes in exudation due to P nutrition alter the chemotaxic attraction of the nematodes to roots and affect exclusion of nematode species that require a hatching stimulus (Baker & Cook, 1982). In general, VAM infection in P-deficient plants affected membrane permeability and exudation pat- terns in a way similar to that caused by P-fertilization in non-mycorrhizal plants.
Reasons for diverse results The diversity of interactions show that each nema- tode-mycorrhizal fungus-plant combination is unique and generalizations regarding such interac- tions are difficult to make. Nematode-mycorrhizal interactions are influenced by the host cultivar, nematode species, fungus species and combination, nematode and fungus initial inoculum densities, soil fertility and sequence of nematode inoculation (Smith, 1987). Most of the evidence is, however, from laboratory, greenhouse and microplot studies. There is a need for extending these studies into field conditions. Moreover, more studies are needed to confirm earlier results. Increased root growth and function, nutritional effects other than that of P, alteration in root exudation, competition for host photosynthates and competition for space and infec- tion site are some reasons which indirectly affect the host-nematode relationship in mycorrhizal plants.
INTERACTION OF ROOT-NODULE BACTERIA AND NEMATODES
The association of rhizobia with legume hosts has a beneficial effect on plant nutrition and growth. In contrast, the plant-nematode relationship has adverse effects on plant growth. The role of plant- parasitic nematodes on rhizobial nodulation and nitrogen fixation of host plants has been reviewed by a number of workers (Huang, 1987; Khan, 1993). As a result of nematode infection, the nodulation and nitrogen fixation has been reported to be suppressed (Hussaini & Seshadri, 1975), or stimulated (Hussey
& Barker, 1976), or remain unaffected (Taha & Raski, 1969). In contrast, the effect of rhizobia on plant parasitic nematodes has been studied in only a few cases (Tables 4 and 5). Inoculation of rhizobia generally had an adverse effect on nematode repro- duction (Table 4), although in a few studies inoculation has been reported to stimulate nema- tode reproduction (Table 5). The role of rhizobia in the control of plant diseases of various leguminous crops has already been discussed (Sawada, 1982) and biological control of plant diseases is now increas- ingly capturing the imagination of plant pathologists (Papavizas & Lumsden, 1980).
Reasons for the reduced nematode reproduction caused by root-nodule bacteria Some of the possible reasons for reduced nematode reproduction are as follows.
Physiological and biochemical changes The root-nodule bacteria which fix atmospheric nitrogen are reported to produce toxic metabolites inhibitory to many plant pathogens (Haque & Gaf- far, 1993) . Rhizobium japonicum secretes rhizobitoxine, which is inhibitory to charcoal root fungus, Macrophomina phaseolina (Chakraborty & Purkayastha, 1984). Chakraborty & Chakraborty (1989) reported an increased level of phytoalexin (4-hydroxy-2,3,9-trimethoxypterocarpan) when pea seeds were bacterized with Rhizobium legumino- sarum prior to inoculation with Fusarium solani f. sp. pisi. This phytoalexin may have an important role in cross-protection against many pathogens. Siddiqui and Mahmood (1994b) observed higher activity of peroxidase, nitrate reductase and catalase in pigeon- pea plants inoculated with Bradyrhizobium japonicum than in plants without B. japonicum. An increase in peroxidase activity due to B. japonicum inoculation indicates increasing resistance of the plant because it catalyzes the polymerization of phe- nolic compounds and forms cross-links between extensin, lignin and feruloylated polysaccharides (Siddiqui & Mahmood, 1994b). An increase in nitrate reductase and catalase may be correlated with the rate of protein synthesis and resistance of the plant to pathogens, respectively (Siddiqui & Mahmood, 1994b). Roslycky (1967) reported pro- duction of an antibiotic bacteriocin by rhizobia bacteria. Some properties of antibiotics of rhizobia bacteria have also been reported by others (Drapeau et al., 1973; Schwinghamer & Belkengren, 1968; Tu, 1978; Tu, 1988). Antibiotics and phytoalexin pro- duced by rhizobia bacteria probably reduce damage from nematodes and other pathogens.
Change in host nutrition Damage to plant growth by nematodes can be less- ened by the application of nitrogen fertilizer (NH4, NOa) (Ross, 1969), indicating that combined nitro- gen can improve growth of diseased plants.

222 Z. A. Siddiqu~ I. Mahmood
Combined nitrogen, such as nitrate, at a high level is a powerful inhibitor of nodulation (Dart, 1977; Pate, 1977) and also has an adverse effect on the develop- ment of nematodes (Barker et al., 1972). Barker and Huisingh (1970) observed necrosis in nodular tissues following invasion by nematodes; this may in part account for reduced nematode development. All this suggests that application of rhizobia bacteria which increase nitrogen content and plant growth can also reduce nematode populations.
Histopathological changes Endo (1964) indicated that nematodes, especially males, often caused plant necrosis and degeneration
of syncytia as the nematodes matured. Endo (1965) found that nematodes induced much necrosis in resistant plants. Some reactions that he observed were very similar to those of nodular tissues where the surrounding tissues, as well as the nematode, died. This type of reaction may partially explain the reduced number of nematodes obtained when nema- todes and Rhizobium were added simultaneously to soybean (Barker et al., 1971).
CONCLUSIONS A N D FUTURE PROSPECTS
Mycorrhizal fungi increase soil nutrients and water absorption as plant symbionts: the protection of
Table 4. Inhibitory effect of root-nodule bacteria on nematode reproduction
Host Nematode Nodulating Effect of plant bacterium interaction on
nematode
References
Soybean Heterodera g!ycines Rhizobium japonicum Reduced nematode reproduction
Soybean H. glycines R. japonicum Nodular tissues are unsuitable for nematode development and most nematodes matured as males
Mung Meloidogyne javanica Rhizobium sp. Reduced nematode multiplication
Cowpea M. incognita Rhizobium sp. Reduced nematode reproduction
Mungbean M. incognita Rhizobium sp. Prior inoculation of Rhizobium resulted in reduced nematode multiplication than simultaneous inoculation
Cowpea M. incognita Rhizobium sp. Reduced nematode Rotylenchulus reniformis multiplication
Mungbean M. incognita Rhizobium phaseoli Treatment of R. phaseoli reduce nematode reproduction
Chick pea M. incognita Bradyrhizobium sp. Reduced nematode multiplication
Lentil M. incognita Rhizobium sp. Reduced nematode R. reniformis reproduction
Chickpea M. javanica Bradyrhizobium sp. Reduced nematode R. reniformis reproduction
Pigeonpea Heterodera cajani B. japonicum Reduced nematode reproduction
Pigeonpea H. cajani B. japonicum Caused greater reduction in nematode multiplication in the presence of VAM fungi
Pigeonpea H. cajani B. japonicum Adverse affect on M. incognita nematode reproduction
Barker et al. (1971)
Barker et al. (1972)
Bopaiah et al. (1976)
Sharma & Sethi (1976)
Hussaini & Seshadri (1975)
Khan & Husain (1988)
Tiyagi & Alam (1988)
Siddiqui & Husain (1992)
Fazal et al. (1992)
Siddiqui & Mahmood (1994a)
Siddiqui & Mahmood (1994b)
Siddiqui & Mahmood (1995a)
Siddiqui & Mahmood (1995c)
Table 5. Stimulatory effect of root-nodule bacteria on nematode reproduction
Host Nematode Nodulating Effect of plant bacterium interaction on
nematodes
References
Cowpea Meloidogyne incognita Rhizobium leguminosarum Resulted in greater Ali et al. (1981) nematode multiplication
Cowpea M. incognita Rhizobium sp. Greater nematode Vashney (1982) multiplication
Mungbean, cluster-bean Heterodera cajani Rhizobium sp. Resulted in increased Dalai & Bhatti (1989) nematode reproduction

Plant symbiotics in nematode controk a review 223
roots from nematodes and other pathogens is a secondary role. Schenck (1981) pointed out that mycorrhizae offer an alternative to chemical control and we should pursue their potential as biocontrol agents. The main obstacle is to produce large quan- tities of inoculum because of their obligate nature. Working on this difficulty, Bagyaraj (1992) found Chloris gayana (Rhodes grass) to be the best host, a Perlite:Soilrite mix (1:1 v/v) to be the best substrate, while the pesticides Captan and Furadan added to the pot cultures at half the recommended level checked contaminants with no effect on the mycor- rhizal fungi. Thus the technique to produce a clean mycorrhizal inoculum with high potential in a short time, and which can be supplied to farmers as 'starter cultures', has been developed. Farmers can then multiply it in their own field for inoculating crop plants. Deep furrows can be made in a small area in the field site, the furrows filled with any available straw to partially sterilize the soil, the starter inoculum added to the furrows and the fur- rows sown with sorghum or Rhodes grass. About 2 kg straw may be needed for a 1 m x 1 m plot. After 60 days, the plant tops can be harvested and used for feeding cattle and the top 6 in. of soil, along with the roots (roots chopped into small pieces), can be air-dried and used as a mycorrhizal inoculum for inoculating crop plants. This simple technology can easily be adopted by farmers, in order to raise mycorrhizal inocula and inoculate crop plants. Mycorrhizal fungi may also be used to inoculate the seedlings of certain crops. The seedlings for this pur- pose would be raised in a nursery and then placed in the field. In this way, prospects of mycorrhizal fungi as biocontrol agents can be fully assessed in the field.
The presence of rhizobia in the rhizosphere pre- sumably protects the host roots from pathogens, besides fixing atmospheric nitrogen. The use of rhi- zobia with mycorrhizal fungi is more beneficial for reducing the damage caused by pathogens (Siddiqui & Mahmood, 1995a). The use of these symbionts will reduce the damage without use of chemical pes- ticides, which are costly and have health hazards. Moreover, use of these two symbionts together will increase nitrogen and phosphorus uptakes without the application of any synthetic fertilizer. In this way, the farmers can also save the capital which they invest for fertilizers.
ACKNOWLEDGEMENTS
The senior author thanks the Council of Scientific and Industrial Research, New Delhi, for the award of a Research Associateship. The authors also thank Prof. D. J. Bagyaraj, Head, Department of Agri- cultural Microbiology, University of Agricultural Sciences, G.K.V.K. Campus, Bangalore, India, for his critical suggestions.
REFERENCES
Ali, M. A., Trabulsi, I. Y. & Abd-Elsamea, M. E. (1981). Antagonistic interaction between Meloidogyne incognita and Rhizobium leguminosarum on cowpea. Plant Disease, 65, 432-5.
Allen, D. N. & Allen, E. K. (1947). A survey of nodula- tion among leguminous plants. Proc. Soil Sci. Soc. Am., 12, 203-8.
Anandaraj, M., Ramana, K. V. & Sharrna, Y. R. (1990). Interaction between vesicular-arbuscular mycorrhizal fungi and Meloidogyne incognita in black pepper. Mycor- rhizal symbiosis and plant growth. Proc. Second National Conf. on Mycorrhiza, Banglore, 21-23 November, ed. D. J. Bagyaraj & A. Manjunath, pp. 110-2.
Atilano, R. A., Menge, J. A. & Van Gundy, S. D. (1981). Interaction between Meloidogyne arenaria and Glomus fasciculatus in grape. J. Nematol., 13, 52-7.
Bagyaraj, D. J. (1984). Biological interactions with VA mycorrhizal fungi. In VA Mycorrhiza, ed. C. L. Powell & D. J. Bagyaraj. CRC Press, Boca Raton, FL, pp. 131-53.
Bagyaraj, D. J. (1991). Ecology of vesicular-arbuscular mycorrhizae. In Hand Book of Applied Mycology, Vol. I, ed. D. K. Arora, B. Rai, K. G. Mukerji & G. R. Knud- sen. Marcel Dekker, New York, pp. 3-34.
Bagyaraj, D. J. (1992). Mycorrhizal associations in plants. In Manual of Mycorrhiza Technology, ed. D. J. Bagyaraj & A. Mangunath. University of Agricultural Sciences, Banglore, India.
Bagyaraj, D. J., Manjunath, A. & Reddy, D. D. R. (1979). Interaction of vesicular arbuscular mycorrhiza with root- knot nematodes in tomato. Plant Soil, 51, 397-403.
Baker, K. F. & Cook, R. J. (1982). Biological control of plant pathogens. The American Phytopathological Society, St Paul, MN.
Baltruschat, H., Sikora, R. A. & Schonbeck, F. (1973). Effect of VA Mycorrhiza (Endogone mosseae) on the establishment of Thielaviopsis basicola and Meloidogyne incognita in tobacco. 2nd Int. Cong. Plant Path.
Barker, K. R. & Huisingh, D. (1970). Histological inves- tigations of the antagonistic interaction between Heterodera glycines and Rhizobium japonicum on soy- bean. Phytopathology, 60, 1282-3.
Barker, K. R., Huisingh, D. & Johnston, S. A. (1972). Antagonistic interaction between Heterodera glycines and Rhizobium japonicum on soybean. Phytopathology, 62, 1201-5.
Barker, K. R., Lehman, P. S. & Huisingh, D. (1971). Influence of nitrogen and Rhizobium japonicum on the activity of Heterodera glycines. Nematologica, 17, 377-85.
Benjamin, R. K. (1979). Zygomycetes and their spores. In The Whole Fungus, ed. B. Kendricks. National Museum of Candada, Ottawa, pp. 573-616.
Bopaiah, B. M., Patil, R. B. & Reddy, D. D. R. (1976). Effect of Meloidogyne javanica on nodulation and sym- biotic nitrogen fixation in mung, Vigna radiata. Indian J. Nematol., 6, 124-30.
Bowen, G. D. (1980). Misconceptions, concepts and approaches in rhizosphere biology. In Contemporary Microbial Ecology, ed. D. C. Ellwood, J. N. Hedger, M. J. Lathem, J. M. Lynch & J. H. Slater. Academic Press, London.
Callaham, D., Tredici, P. D. & Torrey, J. G. (1978). Isola- tion and cultivation in vitro of the actinomycete causing root nodulation in Comptonia. Science, 199, 899-902.
Carling, D. E., Roncadori, R. W. & Hussey, R. S. (1989). Interactions of vesicular-arbuscular mycorrhizal fungi, root-knot nematode and phosphorus fertilization on soybean. Plant Disease, 73, 730-3.
Chakraborty, U. & Chakraborty, B. N. (1989). Interaction

224 Z. A. Siddiqui, I. Mahmood
of Rhizobium leguminosarum and Fusarium solani f. sp. pisi on pea affecting disease development and phyto- alexin production. Can J. Bot., 67, 1698-701.
Chakraborty, U. & Purkayastha, R. P. (1984). Role of Rhizobitoxine in protecting soybean roots from Macro- phomina phascolina infection. Can. J. Microbiol., 30, 285 -9.
Cooper, K. M. & Grandison, G. S. (1986). Interaction of vesicular-arbuscular mycorrhizal fungi and root-knot nematode on cultivars of tomato and white dover sus- ceptible to Meloidogyne hapla. Ann. Appl. Biol., 108, 555-65.
Cooper, K. M. & Grandison, G. S. (1987). Effects of vesicular-arbuscular mycorrhizal fungi on infection of tomarillo (Cyphomandra betacea) by meloidogyne incog- nita in fumigated soil. Plant Disease, 71, 1101-6.
Curl, E. A. & Truelove, B. (1986). The Rhizosphere. Springer, New York.
Dalai, M. R. & Bhatti, D. S. (1989). Pathogenicity of Heterodera cajani in mungbean and cluster bean as affected by presence or absence of Rhizobium. Indian J. Nematol., 19, 153-8.
Dart, P. J. (1977). Infection and development of legumi- nous nodules. In A Treatise on Dinitrogen Fixation, Section 3: Biology, ed. R. W. F. Hardy & W. S. Silver. Wiley Interscience, New York, pp. 367-72.
Dehne, H. W. (1982). Interaction between vesicular arbuscular mycorrhizal fungi and plant pathogens. Phy- topathology, 72, 1115-9.
Diederichs, C. (1987). Interaction between five endomy- corrhizal fungi and the root-knot nematode Meloidogyne javanica on chickpea under tropical conditions. Tropical Agriculture, 64, 353-5.
Drapeau, R., Fortin, J. A. & Cagnon, C. (1973). Anti- fungal activity of Rhizobium. Can. J. Bot., 51, 681-2.
Endo, B. Y. (1964). Penetration and development of Het- erodera g~cines in soybean roots and related anatomical changes. Phytopathology, 54, 79-88.
Endo, B. Y. (1965). Histological responses of resistant and susceptible soybean varieties and back cross progeny to entry and development of Heterodera gly- cines. Phytopathology, 55, 375-81.
Fassuliotis, G. (1970). Resistance in Cucumis spp. to root- knot nematode. Meloidogyne incognita acrita. J. Nematol., 2, 174.
Fazal, M., Siddiqui, Z. A. & Imran, M. (1992). Effect of pre-, post- and simultaneous-inoculations of Rhizobium, Rotylenchulus reniformis and Meloidogyne incognita on lentil. Nematol. Medit., 20, 159-61.
Fox, J. A. & Spasoff, L. (1972). Interaction of Heterodera solanacearum and Endogone gigantea on tobacco. J. Nematol., 4, 224-5.
Francl, L. J. & Dropkin, V. H. (1985). Glomus fascicula- tum, a weak pathogen of Heterodera glycines. J. Nematol., 17, 470-5.
Frank, A. B. (1885). Uber di auf werzelsymbiose ber- uhende Ernahrung gewisser Baume durch unterirdische pilze. Ber. Dtsch. Bot. Ges., 3, 128-45.
Gerdemann, J. W. (1975). Vesicular-arbuscular mycor- rhizae. In The Development and Function of Roots, ed. J. G. Torrey & D. T. Clarkson. Academic Press, London, pp. 576-91.
Goodman, R. N., Kiraly, Z. & Zaitlin, M. (1967). The biochemistry and physiology of infections. In Plant DIS- ease. Van Nostrand, Princeton.
Graham, J. H., Leonard, R. T. & Menge, J. A. (1981). Membrane-mediated decreases in root exudation responsible for phosphorus inhibition of vesicular- arbuscular mycorrhiza formation. Plant Physiol., 68, 548-52.
Graham, J. H. & Menge, J. A. (1982). Influence of vesicu- lar-arbuscular mycorrhizae and soil phosphorus on
take-all disease of wheat. Phytopathology, 72, 95-6. Grandison, G. S. & Cooper, K. M. (1986). Interaction of
vesicular-arbuscular mycorrhizae and cultivars of lucerne susceptible and resistant to Meloidogyne hapla. J. Nematol., 18, 141-9.
Haque, S. E. & Gaffar, A. (1993). Use of Rhizobia in the control of root rot diseases of sunflower, okra, soybean and mungbean. Phytopath. Z., 138, 157-63.
Harley, J. L. & Smith, S. E. (1983). Mycorrhizal symbiosis. Academic Press, London.
Hayman, D. S. (1978). Endomycorrhizas. In Interactions Between Non-pathogenic Soil Microorganisms and Plants, ed. Y. R. Dommergues & S. V. Krupa. Elsevier, Amsterdam.
Hayman, D. S. (1982). The physiology of vesicular-arbus- cular endomycorrhizal symbiosis. Can. J. Bot., 6, 944-63.
Heald, C. M., Bruton, B. D. & Davis, R. M. (1989). Influence of Glomus intraradices and soil phosphorus on Meloidogyne incognita Cucumis melo. J. Nematol., 21, 69-73.
Huang, J. S. (1987). Interactions of nematodes with rhizo- bia. In Vistas on Nematology, ed. J. A. Veech & D. W. Dickson. Society of Nematologists, Hyattsville.
Hussaini, S. S. & Seshadri, A. R. (1975). Interrelationship between Meloidogyne incognita and Rhizobium sp. on mung (Phaseolus aureus). Indian J. Nematol., 5, 189-99.
Hussey, R. S. & Barker, K. R. (1976). Influence of nema- todes and light sources on growth and nodulation of soybean. J. Nematol., 8, 48-52.
Hussey, R. S. & Roncadori, R. W. (1978). Interaction of Pratylenchus brachyurus and Gigaspora margarita on cot- ton. J. Nematol., 10, 16-20.
Hussey, R. S. & Roncadori, R. W. (1982). Vasicular- arbuscular mycorrhizae may limit nematode activity and improve plant growth. Plant Disease, 66, 9-14.
Jain, R. K. & Sethi, C. L. (1987). Pathogenicity of Hetero- dera cajani on cowpea as influenced by the presence of VAM fungi, Glomus fasciculatum or G. epigaeus. Indian J. Nematol., 17, 165-70.
Jain, R. K. & Sethi, C. L. (1988). Influence of endomycor- rhizal fungi Glomus fasciculatum and G. epigaeus on penetration and development of Heterodera cajani on cowpea. Indian J. Nematol., 18, 89-93.
Jalali, B. L. & Jalali, I. (1991). Mycorrhiza in plant disease control. In Handbook of Applied Mycology, Vol. I, ed. D. K. Arora, B. Rai, V. G. Mukerji & G. R. Knudsen. Marcel Dekker, New York, pp. 131-54.
Jordan, D. C. (1984). Family III. Rhizobiaceae Conn 1938. Bergey's Manual of systematic Bacteriology, Vol. 1, ed. N. R. Krieg & J. G. Holt. Williams and Wilkins, Baltimore, pp. 875-82
Kaplan, D. T., Keen, N. T. & Thomason, I. J. (1980). Association of glyceollin with the incompatible response of soybean roots to Meloidogyne incognita. Physiol. Plant Path., 16, 309-18.
Kellam, M. K. & Schenck, N. C. (1980). Interaction between a vesicular-arbuscular mycorrhizal fungus and root-knot nematode on soybean. Phytopathology, 70, 293-6.
Khan, M. W. (1993). Nematode Interactions. Chapman & Hall, London.
Khan, T. A. & Husain, S. I. (1988). Effect of individual, concomitant and sequential inoculations of Rhizobium, Rotylenchulus reniformis, Meloidogyne incognita and Rhi- zoctonia solani on cowpea plant growth, disease development and nematode multiplication. Indian J. Nematol., 18, 232-8.
Kotcon, J. B., Bird, G. W., Rose, L. M. & Dimoff, K. (1985). Influence of Glomus fasciculatum and Meloido- gyne hapla on Allium cepa in organic soils. J. Nematol., 17, 55-60.
Lechevalier, M. P. & Lechevalier, H. A. (1989). Genus

Plant symbiotics in nematode control: a review 225
Frankia Brunchorst 1886, 174. In Bergey's Manual of Systematic Bacteriology, Vol. 4, ed. S. T. Williams. Wil- liams and Wilkins, Baltimore, pp. 2410-7.
Lewin, R. A. (1982). Symbiosis and parasitism: definitions and evaluations. Bioscience, 32, 254-60.
Lin, G. & Burton, J. C. (1982). Nodulation status of the leguminosae. In Nitrogen Fixation. Vol. 2: Rhizobium, ed. W. J. Broughton. Clarendon Press, Oxford, pp. 1-34.
Linderman, R. G. (1985). Microbial interaction in mycor- rhizosphere. In Proc. 6th N. Am. Conf. on Mycorrhizae, ed. R. Molina, pp. 117-20
Linderman, R. G. (1988). Mycorrhizal interactions with rhizosphere microflora: the mycorrhizosphere effect. Phytopathology, 78, 366-70.
MacGuidwin, A. E., Bird, G. W. & Safir, G. R. (1985). Influence of Glomus fasciculatum on Meloidogyne hapla infecting AUium cepa. J. Nematol., 17, 389-95.
Mancini, G., Cotroneo, A. & Moretti, F. (1983). Response of three pines to parasitism by Helicotylenchus digonicus (Nematoda: Hoplolaimidae). Eur. J. Forest Path., 13, 245 -50.
Marks, G. C., Winoto-Suatmadi, R. & Smith, I. W. (1987). Effects of nematode control on shoot, root and mycorrhizal development of Pinus radiata seedlings growing in a nursery soil infested with Pratylenchus pen- etrans. Australian Forest Res., 17, 1-10.
Mengel, K. & Kirkby, E. A. (1979). Principles of Plant Nutrition. International Potash Institute, Bern, Switzer- land.
Morandi, D. (1987). VA mycorrhizae, nematodes, phos- phorus and phytoalexins on soybean. In Mycorrhizae in the Next Decade, Practical Application and Research Pri- orities. Proceedings of the 7th North American Conf. on Mycorrhizae, ed. D. M. Sylvia, L. L. Hung and J. H. Graham. University of Food and Agricultural Sciences, University of Florida, Gainesville.
Nutman, P. S. (1965). The relation between nodule bacteria and the legume host in the rhizosphere and in the process of infection. In Ecology of Soil-borne Plant Pathogens, Prelude to Biological Control, ed. K. F. Baker & W. C. Snyder. University of California Press, Berke- ley, pp. 231-47.
O'Bannon, J. H., Inserra, R. N., Nemec, S. & Vovlas, N. (1979). The influence of Glomus mosseae on Tylenchulus semipenetrans infected and uninfected Citrus lemon seedling. J. Nematol., 11, 247-50.
O'Banrron, J. H. & Nemec, S. (1979). The response of Citrus lemon seedlings to a symbiont, Glomus etunicatus, and a pathogen, Radopholus similis. J. Nematol., 11, 270-5.
Papavizas, G. C. & Lumsden, R. D. (1980). Biological control of soil borne fungal propagules. Ann. Rev. Phyto- path., 18, 389-413.
Pate, J. S. (1977). Functional biology of dinitrogen fixation by legumes. A Treatise on Dinitrogen Fixation, Section 3: Biology, ed. R. W. F. Hardy & W. S. Silver. John Wiley & Sons, New York, pp. 473-517.
Paulitz, T. C. & Linderman, R. G. (1991). Mycorrhizal interactions with soil organisms. In Handbook of Applied Mycology, Vol. I, ed. D. K. Arora, B. Rai, K. G. Mukerji & G. R. Knudsen. Marcel Dekker, New York, pp. 77-129.
Prasad, K. S. K. (1971). Effects of amino acids and plant growth substances on tomato and its root knot nema- tode Meloidogyne incognita Chitwood. M.Sc. (Agric.) Thesis, University of Agricultural Sciences, Banglore, India.
Rabatin, S. C. & Rhodes, L. H. (1982). Acaulospora bireti- culata inside oribatid mites. Mycologia, 74, 859-61.
Reddy, P. P. (1974). Studies on the action of amino acids to root-knot nematode. Meloidogyne incognita, Ph.D. thesis, University of Agricultural Sciences, Banglore,
India. Reid, C. P. P. (1984). Mycorrhizae: a root-soil interface
in plant nutrition. In Microbial-Plant Interactions, A SA special publication, Vol. 47, ed. R. L. Todd & J. E. Giddens, pp. 29-50.
Roncadori, R. W. & Hussey, R. S. (1977). Interaction of endomycorrhizal fungus Gigaspora margarita and root- knot nematode on cotton. Phytopathology, 67, 1507-11.
Roslycky, E. B. (1967). Bacteriocin production in the rhi- zobia bacteria. Can J. Microbiol., 13, 431.
Ross, J. P. (1969). Effect of Heterodera glycines on yields of non-nodulating soybean grown at various nitrogen levels. J. Nematol., 1, 40-2.
Rovira, A. D. (1985). Manipulation of the rhizosphere microflora to increase plant production. In Reviews of Rural Science, Vol. 6, Biotechnology and Recombinant DNA Technology in the Animal Production Industries, ed. R. A. Leng, J. S. F. Barker, D. B. Adams & K. J. Hutchinson. University of Enland, Armidale, NSW, Australia, pp. 185-97.
Saleh, H. & Sikora, R. A. (1984). Relationship between Glomus fasciculatum root colonization on cotton and its effect on Meloidogyne incognita. Nematologica, 30, 230-7.
Sawada, Y. (1982). Interaction of rhizobial nodulation of alfalfa and root-rot caused by Fusarium oxysporum. Bull. National Grassland Research Inst. Japan, 22, 19-26.
Schenck, N. C. (1981). Can mycorrhizae control root dis- ease? Plant Disease, 65, 230-4.
Schenck, N. C., Kinloch, R. A. & Dickson, D. W. (1975). Interaction of endomycorrhizal fungi and root-knot nematode on soybean. In Endomycorrhizas, ed. F. E. Sanders, B. Mosse & P. B. Tinker. Academic Press, New York, pp. 607-17.
Schonbeck, F. (1979). Endomycorrhiza in Relation to Plant Diseases in Soil-borne Plant Pathogens, ed. B. Schippers & W. Gams. Academic Press, London, pp. 271-80.
Schwinghamer, E. A. & Belkengren, R. D. (1968). Inhibi- tion of Rhizobium by a strain of Rhizobium trifolii; some properties of the antibiotic and of the strain. Arch. Mik- robiol., 64, 130-45.
Sharma, N. K. & Sethi, C. L. (1976). Inter-relationship between M. incognita, H. cajani and Rhizobium spp. on cowpea (Vigna sinensis). Indian J. Nematol., 6, 117-23.
Siddiqui, Z. A. & Husain, S. I. (1992). Interaction between Meloidogyne incognita race 3, Macrophomina phaseolina and Bradyrhizobium sp. in the root-rot dis- ease complex of chick-pea. Cicer arietinum. Fundam. Appl. Nematol., 15, 491-4.
Siddiqui, Z. A. & Mahmood, I. (1994a). Interactions of Meloidogyne javanica, Rotylenchulus reniformis, Fusarium oxysporum f. sp. ciceri and Bradyrhizobium japonicum on the wilt disease complex of chikpea. Nematol. Medit., 22, 135-40.
Siddiqui, Z. A. & Mahmood, I. (1994b). Effect of Hetero- dera cajani on growth, chlorophyll content and activity of some enzymes in pigeonpea. Nematropica, 24, 103-11.
Siddiqui, Z. A. & Mahmood, I. (1995a). Biological control of Heterodera cajani and Fusarium udum by Bacillus sub- tilis, Bradyrhizobium japonicum and Glomus fasciculatum on pigeonpea. Fundam. Appl. Nematol., 18, 559-66.
Siddiqui, Z. A. & Mahmood, I. (1995b). Some observa- tions on the management of the wilt disease complex of pigeonpea by the treatment with a vesicular arbuscular fungus and biocontrol agents for nematodes. Biores. Technol., 54, 227-30.
Siddiqui, Z. A. & Mahmood, I. (1995c). The effect of inoculations of Heterodera cajani and Meloidogyne incog- nita with Fusarium udum and Bradyrhizobium japonicum on the wilt disease complex of pigeonpea. Indian Phyto- path. (accepted).

226 Z. A. Siddiqui, I. Mahmood
Sikora, R. A. (1978). Einfluss der endotrophen mykor- rhiza (Glomus mosseae) auf das wirt-parasit-verhaltnis Von Meloidogyne incognita in tomaten. Zeitschrift fur Pflanzenkrankheiten und Pflanzenschutz, 85, 197-202.
Sikora, R. A. & Schonbeck, F. (1975). Effect of vesicular- arbuscular mycorrhiza (Endogone mosseae) on the population dynamics of the root-knot nematodes (Mel- oidogyne incognita and M. hapla). 8th International Congress of Plant Protection, 5, 158-66.
Singh, V. P., Singh, R. S. & Sitaramaiah, K. (1990). Mech- anism of resistance of mycorrhizal tomato against root-knot nematode. In Trends in Mycorrhizal Research, Proc. National Conf. on Mycorrhiza, ed. B. L. Jalali & H. Chand. Haryana Agricultural University, Hissar, pp. 96-7.
Sitaramaiah, K. & Sikora, R. A. (1982). Effect of mycor- rhizal fungus Glomus fasciculatus on the host parasite relationship of Rotylenchulus reniformis in tomato. Nematologica, 28, 412-9.
Sivaprasad, P., Jacob, A., Nair, S. K. & George, B. (1990). Influence of VA mycorrhizal colonization on root-knot nematode infestation in Piper nigrum L. In Trends in Mycorrhizal Research, Proc. National Conf. on Mycor- rhiza, ed. B. S. Jalali & H. Chand. Haryana Agricultural University, Hissar, pp. 100-1.
Smith, G. S. (1987). Interaction of nematodes with mycor- rhizal fungi. In Vistas on Nematology, ed. J. A. Veech & D. W. Dickson, pp. 133-43.
Smith, G. S. (1988). The role of phosphorus nutrition in interactions of vesicular-arbuscular mycorrhizal fungi with soil-borne nematodes and fungi. Phytopathology, 78, 371-4.
Smith, G. S., Hussey, R. S. & Roncadori, R. W. (1986a). Penetration and post infection development of Meloido- gyne incognita on cotton as affected by Glomus intraradices and phosphorus. J. Nematol., 18, 429-35.
Smith, G. S., Roncadori, R. W. & Hussey, R. S. (1986b). Interaction of endomycorrhizal fungi, superphosphate and Meloidogyne incognita on cotton in microplot and field studies. J. Nematol., 18, 208-16.
Smith, G. S. & Kaplan, D. T. (1988). Influence of mycor- rhizai fungus, phosphorus and burrowing nematode interactions on growth of rough lemon citrus seedlings. J. Nematol., 20, 539-44.
Strobel, N. E., Hussey, R. S. & Roncadori, R. W. (1982). Interactions of vesicular-arbuscular mycorrhizal fungi, Meloidogyne incognita and soil fertility on peach. Phyto- pathology, 72, 690-4.
Sundarababu, R., Sankaranarayanan, C. & Santhi, A. (1993). Interaction between vesicular-arbuscular mycor- rhiza and Meloidogyne javanica on tomato as influenced by time of inoculation. Indian J. Nematol., 23, 125-7.
Suresh, C. K. (1980). Interaction between vesicular- arbuscular mycorrhiza and root-knot nematode in
tomato. M.Sc. (Agric.) Thesis, University of Agricultural Sciences, Banglore, India.
Suresh, C. K., Bagyaraj, D. J. & Reddy, D. D. R. (1985). Effect of vesicular-arbuscular mycorrhiza on survival, penetration and development of root-knot nematode in tomato. Plant Soil, 87, 305-8.
Sylvia, D. M. & Sinclair, W. A. (1983). Phenolic com- pounds of resistance to fungal pathogens induced in primary roots of Douglas-fir seedlings by the ecotomy- corrhizal fungus Laccaris laccata. Phytopathology, 73, 390-7.
Taber, H. A. (1982). Occurrence of Glomus spores in weed seeds in soil. Myeologia, 74, 515-20.
Taha, A. H. Y. & Raski, D. J. (1969). Interrelationships between root-nodule bacteria, plant parasitic nematodes and their leguminous host. J. Nematol., 1, 201-11.
Thompson Cason, K. M., Hussey, R. S. & Roncadori, R. W. (1983). Interaction of vesicular-arbuscular mycor- rhizal fungi and phosphorus with Meloidogyne incognita on tomato. J. Nematol., 15, 410-7.
Tiyagi, S. A. & Alam, M. M. (1988). Pathogenicity of root-knot on mungbean. Int. Nematol. Network News., 5, 22-4.
Trudgill, D. L. & Parrott, D. M. (1969). The behavior of the population of potato cyst nematode Heterodera ros- tochinensis towards three resistant potato hybrids. Nematologica, 15, 381.
Tu, J. C. (1978). Protection of soybean from severe phyto- phthora root-rot by Rhizobium. Physiol. Plant Path., 12, 233 -40.
Tu, J. C. (1988). Incidence of root rot and over-wintering of alfalfa as influenced by Rhizobia. Phytopath. Z., 97, 233-40.
Tylka, G. L., Hussey, R. S. & Ronadori, R. W. (1991). Interactions of vesicular-arbuscular mycorrhizal fungi, phosphorus and Heterodera glycines on soybean. J. Nematol., 23, 122-3.
Umesh, K. C., Krishnappa, K. & Bagyaraj, D. J. (1988). Interaction of burrowing nematode, Radopholus similis (Cobb, 1893) Thorne 1949, and VA mycorrhiza, Glomus fasciculatum (Thaxt) Gerd & Trappe, in banana (Musa acuminata Colla). Indian J. Nematol., 18, 6-11.
Vance, C. P. & Johnson, L. E. B. (1981). Nodulation: a plant disease perspective. Plant Disease, 65, 118-28.
Vashney, V. P. (1982). Changes in plant growth, nema- tode population and nodule index as a result of inoculation of cowpea, Vigna unguiculata (L.) Walp., with Meloidogyne incognita (Kofold & White) Chitwood and Rhizoctonia solani Kuhn. Ph.D. Thesis, Aligarh Muslim University, Aligarh.
Zombolin, L. & Oliveira, A. A. R. (1986). Interacaco entre Glomus etunieatum and Meloidogyne javanica em feijao (Phaseolus vulgaris L.). Fitopatologia Brasileira, 11, 217.




















