
Improving triploidy induction in the hard clam Mercenaria mercenaria notata: cytological evaluation...
19
ISSN: 1687-4285 Egyptian Journal of Aquatic Research, 2009, 35(3), 385-403 Improving triploidy induction in the hard clam Mercenaria mercenaria notata: cytological evaluation of fertilized eggs *Eman El-Wazzan a , and John Scarpa b a Division of Aquaculture, National Institute of Oceanography and Fisheries, Qaiyet-Bay, El-Anfoushi, Alexandria, 21556, Egypt. b Aquaculture and Stock Enhancement Program, Harbor Branch Oceanographic Institute at Florida Atlantic University, 5600 U.S. 1 North, Fort Pierce, FL, 34946, USA. * corresponding author email: [email protected] Received 30 th September 2009, Accepted 15 th November 2009 Abstract Triploid organisms contain three sets of chromosomes instead of the usual two, thereby rendering them essentially sterile, which may improve growth and stress resistance. Triploidy may be induced by inhibiting extrusion of the first polar body (PBI) or second polar body (PBII) during meiosis using chemicals, such as cytochalasin B (CB). In bivalves, which release large quantities of pre-meiotic eggs, producing a high proportion of triploids relies upon synchrony of meiotic events after fertilization. Therefore, timing of meiotic stages of fertilized eggs of the hard clam Mercenaria mercenaria notata was examined to improve triploid induction yield. Spawned eggs from three females were sampled before insemination and intermittently afterwards up to 90 min post-insemination (PI). Meiotic stages of eggs (avg. 90 eggs/sample) were visualized with the DNA-specific flurochrome Hoechst 33258 and epifluorescence microscopy. The maximum proportion of eggs with PBI and PBII appeared on average at 16 min and 28 min PI, respectively. First cell cleavage (i.e., first mitosis) was greatest on average at 60 min PI. Using these data, triploid induction in hard clams was evaluated in ten trials using cytochalasin B (CB) at 0.67 or 1mg/L egg suspension. CB treatment started at 6.5 to11 min PI with a 10-12 minutes exposure for PBI inhibition and at 15.5 to 21 min with a 13-15 minute exposure for PBII inhibition. The proportions of eggs synchronized at stages just before PBI or PBII extrusion in CB-treated eggs (i.e., potential triploid proportion) were compared to triploid proportions in D-stage larvae as measured by flow-cytometry. Potential triploid proportions from eggs ranged from 0-100%, whereas triploid proportions from larvae ranged from 0- 93%; however, triploid values from both methods were not always similar for each spawn. Parameters that resulted in lower triploid proportions included low CB concentration (0.67 mg/L), pre-fertilization that also severely reduced larval survival (1-2% compared to control), and early or late addition or removal of CB. Depending on environmental conditions, especially temperature that hastens development, induction of triploidy in hard clams using CB should begin 10 minutes PI for PBI inhibition and at 17 min PI for PBII inhibition. CB exposure should not go beyond 25 minutes PI for PBI inhibition or 40 min PI for PBII inhibition. Keywords: Polyploidy, Triploid, Clam, Mercenaria, Bivalve, Cytology 1. Introduction Hard clam culturists in southwest and west Florida, USA report below average survival during the prolonged hot summers (Leslie Sturmer, University of Florida, pers. comm.). It has been suggested that bivalve mortality increases after spring spawning, which decreases tissue glycogen levels (Perdue et al. 1981), and is followed by hot (30 o C) water temperatures, which are associated with lower phytoplankton concentration and oxygen concentrations, as well as large salinity fluctuations from rainfall (Goldman and Ryther 1976, Ohgai et al. 1982, Surge and Lohmann 2002, Weiss et al. 2007). Bivalves that are deprived of energy needed for their maintenance after investment in gametogenesis and spawning are more susceptible to stressors that amplify the already present reproductive cost than those with lower reproductive effort or metabolic needs (Perdue et al. 1981, Eversole 1987, Shpigel et al. 1992, Myrand and Gaudreault 1995). Triploid bivalves may have an energetic advantage over diploids at elevated temperature as triploids are sterile or have severely retarded gametogenesis (e.g., Tabarini 1984, Allen et al. 1986, Lee 1988, Shpigel et al. 1992, Eversole et al. 1996, Utting et al. 1996, Brake et al. 2004, Maldonado- Amparo et al. 2004). Therefore, triploids may be more resistant to summer mortality as they would have higher body mass and more energy reserves (Perdue et al. 1981, Allen et al. 1989, Shpigel et al. 1992, Hand et al. 2004). Successful induction of triploidy in hard clams would provide viable triploids as a first step towards examining their performance in Florida waters, as has been accomplished with other molluscan species (e.g., Baron et al. 1989, Utting and Child 1994, Toro and Sastre 1995). Inducing triploidy in bivalves relies upon EJAR
Transcript of Improving triploidy induction in the hard clam Mercenaria mercenaria notata: cytological evaluation...

ISSN: 1687-4285 Egyptian Journal of Aquatic Research, 2009, 35(3), 385-403
Improving triploidy induction in the hard clam Mercenaria mercenaria notata: cytological evaluation of fertilized eggs
*Eman El-Wazzana, and John Scarpab
a Division of Aquaculture, National Institute of Oceanography and Fisheries, Qaiyet-Bay, El-Anfoushi, Alexandria, 21556, Egypt. b Aquaculture and Stock Enhancement Program, Harbor Branch Oceanographic Institute at Florida Atlantic University, 5600 U.S. 1 North, Fort Pierce, FL, 34946, USA. * corresponding author email: [email protected] Received 30th September 2009, Accepted 15th November 2009 Abstract
Triploid organisms contain three sets of chromosomes instead of the usual two, thereby rendering them essentially
sterile, which may improve growth and stress resistance. Triploidy may be induced by inhibiting extrusion of the first polar body (PBI) or second polar body (PBII) during meiosis using chemicals, such as cytochalasin B (CB). In bivalves, which release large quantities of pre-meiotic eggs, producing a high proportion of triploids relies upon synchrony of meiotic events after fertilization. Therefore, timing of meiotic stages of fertilized eggs of the hard clam Mercenaria mercenaria notata was examined to improve triploid induction yield. Spawned eggs from three females were sampled before insemination and intermittently afterwards up to 90 min post-insemination (PI). Meiotic stages of eggs (avg. 90 eggs/sample) were visualized with the DNA-specific flurochrome Hoechst 33258 and epifluorescence microscopy. The maximum proportion of eggs with PBI and PBII appeared on average at 16 min and 28 min PI, respectively. First cell cleavage (i.e., first mitosis) was greatest on average at 60 min PI. Using these data, triploid induction in hard clams was evaluated in ten trials using cytochalasin B (CB) at 0.67 or 1mg/L egg suspension. CB treatment started at 6.5 to11 min PI with a 10-12 minutes exposure for PBI inhibition and at 15.5 to 21 min with a 13-15 minute exposure for PBII inhibition. The proportions of eggs synchronized at stages just before PBI or PBII extrusion in CB-treated eggs (i.e., potential triploid proportion) were compared to triploid proportions in D-stage larvae as measured by flow-cytometry. Potential triploid proportions from eggs ranged from 0-100%, whereas triploid proportions from larvae ranged from 0-93%; however, triploid values from both methods were not always similar for each spawn. Parameters that resulted in lower triploid proportions included low CB concentration (0.67 mg/L), pre-fertilization that also severely reduced larval survival (1-2% compared to control), and early or late addition or removal of CB. Depending on environmental conditions, especially temperature that hastens development, induction of triploidy in hard clams using CB should begin 10 minutes PI for PBI inhibition and at 17 min PI for PBII inhibition. CB exposure should not go beyond 25 minutes PI for PBI inhibition or 40 min PI for PBII inhibition.
Keywords: Polyploidy, Triploid, Clam, Mercenaria, Bivalve, Cytology
1. Introduction
Hard clam culturists in southwest and west Florida, USA report below average survival during the prolonged hot summers (Leslie Sturmer, University of Florida, pers. comm.). It has been suggested that bivalve mortality increases after spring spawning, which decreases tissue glycogen levels (Perdue et al. 1981), and is followed by hot (30oC) water temperatures, which are associated with lower phytoplankton concentration and oxygen concentrations, as well as large salinity fluctuations from rainfall (Goldman and Ryther 1976, Ohgai et al. 1982, Surge and Lohmann 2002, Weiss et al. 2007). Bivalves that are deprived of energy needed for their maintenance after investment in gametogenesis and spawning are more susceptible to stressors that amplify the already present reproductive cost than those with
lower reproductive effort or metabolic needs (Perdue et al. 1981, Eversole 1987, Shpigel et al. 1992, Myrand and Gaudreault 1995). Triploid bivalves may have an energetic advantage over diploids at elevated temperature as triploids are sterile or have severely retarded gametogenesis (e.g., Tabarini 1984, Allen et al. 1986, Lee 1988, Shpigel et al. 1992, Eversole et al. 1996, Utting et al. 1996, Brake et al. 2004, Maldonado-Amparo et al. 2004). Therefore, triploids may be more resistant to summer mortality as they would have higher body mass and more energy reserves (Perdue et al. 1981, Allen et al. 1989, Shpigel et al. 1992, Hand et al. 2004).
Successful induction of triploidy in hard clams would provide viable triploids as a first step towards examining their performance in Florida waters, as has been accomplished with other molluscan species (e.g., Baron et al. 1989, Utting and Child 1994, Toro and Sastre 1995). Inducing triploidy in bivalves relies upon
EJAR

Eman El-Wazzan, and John Scarpa
Egyptian Journal of Aquatic Research, 2009, 35(3), 385-403 ISSN: 1687-4285
386 exposing fertilized eggs to agents that inhibit extrusion of either the first polar boy (PBI) or second polar body (PBII), which normally release the extra chromosomal sets during meiosis (Chaiton and Allen 1985, Beaumont and Fairbrother 1991, Scarpa et al. 1994b, Eudeline et al. 2000). The dynamics of polar body release are important for successful induction of triploidy by targeting the correct meiotic stage.
Compared to oysters, there have only been a few studies that examined triploid hard clams (Hidu et al. 1988, Buzzi 1990, Eversole et al. 1996, Yang et al. 2001). Only Buzzi (1990) and Eversole et al. (1996) reported time of addition of the polar body inhibiting agent cytochalasin B (CB) to fertilized eggs; 5 and 10 min post-insemination with an exposure time of 20 min. By knowing the timing of extrusion of each polar body, polar body inhibiting agent application can be done at the appropriate time before the start and removed after the end of this time range.
The aim of the present study was to evaluate and customize triploidy induction in hard clams by: 1) describing timing of meiotic maturation in fertilized eggs of hard clams; 2) attempting triploidy induction at the times calculated in objective one; and 3) evaluating the success of triploidy induction trials. 2. Materials and Methods
Fertilized eggs from commercial spawns of the hard clam Mercenaria mercenaria notata were examined to describe the timing of meiotic and mitotic stages. These data were then used to attempt triploid induction in hard clams. The resulting triploid proportions in larvae, as measured by flow-cytometry, were compared to cytological data of eggs at the time of induction to evaluate the success or failure of triploidy induction in ten trials. 2.1 Timing of meiotic and mitotic stages of fertilized eggs 2.1.1. Spawning and fertilization.
Broodstock Mercenaria mercenaria notata were obtained from a commercial hatchery (HB Clams, Inc., Ft. Pierce, Florida). Eggs from three female clams spawned by thermal induction (i.e., alternating cycles of exposure to chilled (24ºC) and warmed (30–31ºC) seawater) were used. Spawned eggs were suspended in 300 mL seawater (27.5–28oC, 31 ppt) in separate containers. The egg suspension from each female was split into two treatments: unfertilized and fertilized. The unfertilized treatment was used as control to check for pre–fertilization that might occur during mass spawning. Fertilized treatments were inseminated with 3 mL sperm suspension from mass spawned males at 30 min post-spawning.
For cytological observations, eggs were sampled just before insemination (T0 samples). Eight min after insemination, eggs were rinsed from sperm by sieving them on 20 μm mesh screen and washing with seawater (29oC, 31 ppt). Fertilized eggs were then re-suspended in 600 mL seawater. Controls (unfertilized eggs) from the 3 females were also made up to 600 mL with seawater. Samples of 4.5 mL were withdrawn from each control and fertilized treatment for each of the three females at different times post-insemination. Samples were taken at 10, 13, 16, 19, 22, 25, 28, 31, 35, 40, 50, 60, 70, 80 and 90 min post-insemination. Samples were fixed by adding 0.5 mL 10% (v/v) buffered formalin and stored at 4oC until cytological observation. 2.1.2. Cytology
Chromosomal material of eggs was visualized with the DNA–specific fluorochrome Hoechst 33258 and epifluorescence microscopy. Fixed eggs were observed on wet mount using the method of Scarpa and Allen (1992), except that the DAPI solution was replaced by 10 μM Hoechst 33258 solution and chitosan-coated slides were prepared using 2% v/v acetic acid. An average of 90 eggs was observed in each sample using an Olympus BH2 microscope equipped with the UV filter set. Photomicrographs were taken with a Kodak DC290 Zoom digital camera.
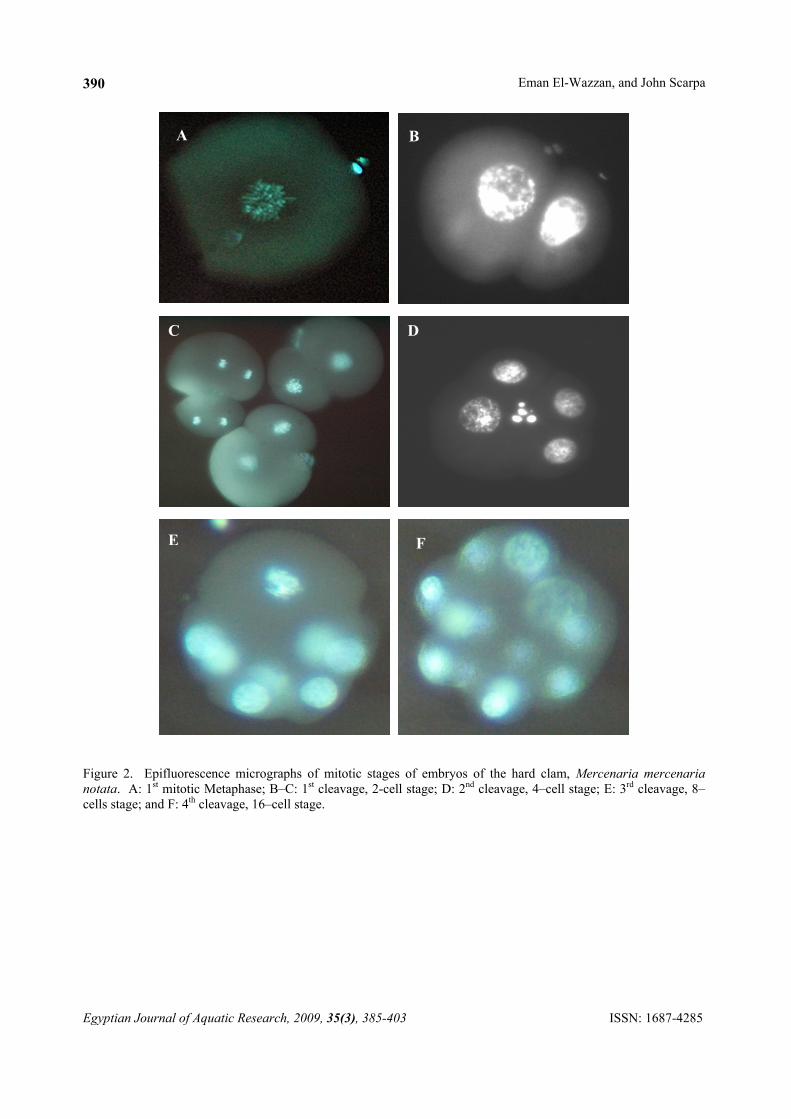
Stages scored were germinal vesicle (GV), prophase I (Pro I), metaphase I (Met I), anaphase I (Ana I), telophase I (Tel I), and polar body I (PBI). The anaphase/telophase I stage was considered as the marker for the extrusion of the first polar body (PBI). The same abbreviations were used to score the second meiotic division, but with addition of II instead of I (Figure 1). In the second meiotic division, anaphase/telophase II stage was considered as the marker for the extrusion of the second polar body (PBII). Mitotic events (Figure 2) were described as 1st cleavage, which is the 2–cell stage, 2nd cleavage (4–cell stage), 3rd cleavage (8–cell stage) and finally 4th cleavage (16–cell stage). The percentages of different stages were calculated for each time interval. 2.2. Induction of triploidy
Triploid induction was attempted with eggs from clams spawned by thermal shock en masse on a spawning table or individually in separate containers.
2.2.1. Clams induced to spawn en masse
Clams in a commercial facility were induced to spawn by thermal cycling. Clams that started spawning were transferred directly into separate small containers filled with clean seawater to resume spawning. Eggs from 6 females were mixed, sperm added (3 mL from mass spawned males), and divided into three treatments

Improving triploidy induction in the hard clam
ISSN: 1687-4285 Egyptian Journal of Aquatic Research, 2009, 35(3), 385-403
387
(control, PBI and PBII). Sperm were rinsed from eggs 5 min after insemination for all treatments. Cytochalasin B (1.0 mg CB, Scarpa et al. 1994b) was dissolved in dimethyl sulphoxide (DMSO) as a carrier solution and added to fertilized eggs suspended in 1 L of seawater (final concentration of 1.0 mg CB/ 1L of 0.1% DMSO in seawater) at 10 min for PBI treatment and at 20 min for PBII treatment. Eggs were exposed to CB for 10.5 and 15 min for PBI and PBII treatments, respectively. Egg were caught on a 20 μm mesh screen and rinsed with seawater (2 L) to remove CB. Eggs in PBI and PBII treatments were re-suspended in seawater with 0.1% DMSO for removing residual CB for 15 min. Eggs in the control treatment were re-suspended in seawater with 0.1% DMSO at 25 min post–insemination as a control to show the possible effect of DMSO on eggs. Eggs were caught on a 20 μm mesh screen and rinsed with seawater (4 L) to remove residual DMSO at 40 min post–insemination for PBI and at 50 min post–insemination for both PBII and control treatments. Eggs were sampled and fixed for epifluorescence microscopy at time intervals that covered the treatments (pre-insemination =T0, 10 min, 20 min and 55 min post–insemination). Eggs were then placed in 500 L tanks with filtered (1 µm) seawater and reared as described by Hadley et al. (1997). After 24 hours, D–stage larvae in each tank were enumerated to compare survival of treated groups with the control (untreated) group. 2.2.2. Clams induced to spawn in separate containers
To avoid pre–fertilization of eggs, clams were placed in separate containers and spawning attempted by thermal cycling. As spawning could not be induced by thermal cycling alone, irradiated sperm were utilized.
Twenty broodstock clams were kept dry in a refrigerator (4oC) overnight before being transferred to separate 4–L beakers containing 1–2 L of seawater (31oC, 30 ppt). Beakers were kept in a water bath on the spawning table. Thermal induction of spawning was tried three times by removing clams from the warm seawater, exposing them to air for 15 min, and then returning them back to the beakers with new warm seawater and leaving them undisturbed for 1 hour. No clams spawned; therefore, induction of spawning was tried by using sperm killed by UV-irradiation (Scarpa 2002). The irradiation of sperm was conducted in a hood fitted with a single germicidal UV bulb (General Electric G30T8, 30W, Cleveland, Ohio). A thin layer of sperm suspension (~1mm) in a glass tray (25 x 38 cm) was exposed to approximately 480 µW.cm–2.s–1 energy provided by the UV lamp for 3 min. Eggs were sampled and observed visually under the microscope for sperm viability (i.e., motility) and re–exposed to UV for 5 more min to ensure death of all sperm. UV treatment stopped when all sperm appeared to be non–motile. Fifteen mL of the irradiated sperm suspension
was added near the siphon of each individual clam. About 30 min later, two males and two females spawned. Oocytes from the two females were mixed, suspended in 1L of seawater and inseminated by a mixture of sperm from the two spawned males. Sperm were rinsed from eggs 5 min after insemination as described in section 2.2.1. Fertilized eggs were then divided into three treatments (control, PBI and PBII). Chemical treatment was carried out using the same procedures used in the first trial. Eggs were sampled and fixed for epifluorescence microscopy at time intervals that covered the treatments; pre-insemination =T0, at time of CB addition (10 and 20 min) and at 55 min post- insemination.
After rinsing eggs from CB and DMSO as described in section 2.2.1, eggs were placed in 250 L of seawater in 500L tanks with filtered seawater (31oC, 29 ppt) and reared as described by Hadley et al. (1997).
2.3. Triploid induction trials
To evaluate success or failure of triploidy induction in hard clams, fertilized eggs and embryos from ten trials were examined. Hard clam eggs from two to six females were inseminated by sperm from 1 to 3 males (Table 3 A & 3B). Conditions for the ten trials are summarized in Tables (4 A & 4B). Briefly, fertilized eggs were exposed to cytochalasin B (CB) starting 6.5 to 11 min post-insemination to inhibit PBI or 15.5 to 21 min post-insemination to inhibit PBII. CB concentrations ranged from 0.67 mg/mL to 1.05 mg/mL. Initial start times used were suggested by data obtained in section 2.1. Eggs were exposed to CB for 10–12 min to inhibit PBI and for 13–15 min to inhibit PBII. For each attempt, eggs were sampled and fixed, as mentioned above in section 2.1, just before insemination (=T0 sample) to check for pre–fertilization and at the time of CB application. An average of 76 (range 53–122) eggs/sample was scored for meiotic stages as described in section 2.1.2.
Two–day or older larvae were sampled and analyzed for ploidy by flow cytometry as described below. Triploid proportions detected by flow-cytometry were compared to cytological data; i.e. putative triploid proportions as indicated by proportion of eggs at the appropriate stage for inhibition.
2.3.1. Assessment of triploidy Two–day to four–day old larvae were sampled by first catching them on sieves with 20–37 µm openings, depending on larval size, and resuspending in 1-5L of seawater. Three replicate 1.0 mL larval samples were transferred into the compartment of counting slide (Sedgewick–Rafter) and larvae counted with the aid of a compound microscope to determine larvae density. Approximately 1.0 x 104 larvae from each group were concentrated into approximately 0.5 mL suspension and placed in 1.0 mL DAPI/DMSO freezing solution

Eman El-Wazzan, and John Scarpa
Egyptian Journal of Aquatic Research, 2009, 35(3), 385-403 ISSN: 1687-4285
388 (Allen and Bushek 1992) and stored at –80oC until analyzed by flow cytometry to determine the proportions of triploid larvae (Allen and Bushek 1992). 3. Results 3. 1. Timing of meiotic and mitotic events
Observations of developing eggs in three replicate females indicated that at the time of insemination (T0) a significant number of eggs were pre–fertilized and had passed Met I (unfertilized eggs should be arrested at this stage). Therefore, eggs that progressed to any stage after Metaphase I in the control treatment (i.e., unfertilized eggs) were considered to be pre–fertilized and were subtracted from the fertilized samples at the same time interval to correct for the effect of pre–fertilization. Fertilized clams eggs progressed through meiotic maturation (Figures 1 & 2). Polar body I (PBI) was visible at 13 min (Figures 3). The maximum proportion of eggs exhibiting PBI was observed at 16 min. Polar body II (PBII) started to appear at 16 min with maximum proportion at 28 min (Figures 3). Eggs started to enter mitosis at 35 min and 1st cleavage was observed to start at 40 min with a maximum of 1st cleavage (2-cell stage) at 60 min post-insemination (Figures 3). The time frames of polar body formation were utilized as starting points for induction of triploidy in the present study. 3.2. Triploid induction trials 3.2.1. Clams spawned on spawning table
Observations of eggs by epifluorescence microscopy indicated that at the time eggs were inseminated (T0), a significant percentage of eggs has passed Met I (total of 72.4% ≥ Ana I, Table 1). At the time of treatment (10 min post-insemination) to inhibit PBI a substantial portion of eggs (91.2%) had passed through meiosis II and entered mitosis (Table 1, Figures. 4B & 4C). A similar observation was made at 20 min post-insemination; 97.1% of eggs had passed the PBII stage in the PBII treatment group (Table 1). Deformed embryos with inhibited cleavage were observed in both treatment groups at 55 min post-insemination (Figure. 4). Twenty four hours after insemination the control group contained 813000 D-stage larvae, whereas only 11000 and 18000 D-stage larvae were in the PBI and PBII groups, respectively. Survival was 1.4% and 2.2% of control in PBI and PBII treatments, respectively. 3.2.2. Clams spawned in separate containers
Induction of spawning in separate containers by thermal cycles was not successful after two consecutive attempts. Thirty minutes after adding irradiated sperm
to the containers, spawning occurred by two females and two males out of total 20 clams treated. One hr after spawning, no signs of fertilization (e.g., PB or cleavage) were observed by light microscopy. Observation of sampled eggs by epifluorescence microscopy indicated that all eggs were either in Pro I (76.3%) or Met I (23.7%) stage of development (Table 2). Before the addition of live sperm (T0) there was no pre–fertilization of eggs (i.e. no sperm were attached to or had entered the eggs, Table 2). However, a significant proportion (46–77%) of unfertilized eggs were found in all samples at all time intervals in all groups (Table 2).
At the time of treatment to inhibit PBI (10 min post–insemination), 46% of fertilized eggs had passed metaphase I and approached the formation of the first polar body (i.e., were at the targeted stages for PBI inhibition), but no eggs exhibited the first polar body at this time (Table 2). At the time of treatment to inhibit PBII (20 min post–insemination), it was found that only 10% of fertilized eggs were at the targeted stages (i.e., Ana II and Tel II, 3.3% and 6.7%, respectively; Table 2), although a total of 40% of the fertilized eggs had passed the PBI stage (Table 2). Additionally, at this time 26.7% of fertilized eggs progressed in meiosis beyond Met I stage (i.e., were activated), but had not formed the first polar body (i.e., were at Ana I/Tel I stages; Table 2). At 55 min post–insemination, almost all the eggs in the three groups passed the second meiotic division and the embryos were at different mitotic stages (Table 2). In addition to embryos with normal 1st cleavage, there were embryos with abnormal nuclei or dispersed chromosomal material (Figures. 5C & 5G). Furthermore, counting the number of polar bodies in the 2–cell stage embryos of the PBII treatment revealed that triploidy was not induced in all eggs. Both polar bodies were present in 30% of eggs (i.e. triploidy had not been induced in these eggs, Figure 5A–C), one polar body was present in 15% of eggs (i.e., putative triploid embryos, Figure 5D–E) and no polar bodies were present in 55% of eggs (i.e., putative pentaploid embryos, Figure 5F–G). In contrast, all embryos of the control group had both polar bodies. 3.3. Cytological evaluation of triploidy induction When cytological data were compared to flow–cytometry data, several factors were noted that affected triploid induction success. 3.3.1. Evaluation of fertilization
High pre–fertilization occurred in Trials 1 and 3 (85% and 55 % fertilized at T0; Table 3A). At the time of treatment to inhibit PBI (10 min post–insemination), 0% and 50% of eggs were at the targeted stages (Pre–PBI) in the two trials, respectively (Table 3A).

Improving triploidy induction in the hard clam
ISSN: 1687-4285 Egyptian Journal of Aquatic Research, 2009, 35(3), 385-403
389
Figure 1. Epifluorescence micrographs of meiotic stages of eggs of the hard clam, Mercenaria mercenaria notata. A: Oocytes just after spawning (Germinal vesicle, GV); B: Metaphase I of unfertilized egg; C: Metaphase I of fertilized egg (arrow indicates sperm nucleus); D: Early Anaphase I of fertilized egg; E: Late Anaphase I; F: Polar body I extrusion (PBI, arrow); G: Polar Body II extrusion (PBII, arrow); H: Pronuclei; I: Fusion of pronuclei (arrow indicates both polar bodies).
B C
E F
G H I
A
D
Eman El-Wazzan, and John Scarpa
Egyptian Journal of Aquatic Research, 2009, 35(3), 385-403 ISSN: 1687-4285
390
Figure 2. Epifluorescence micrographs of mitotic stages of embryos of the hard clam, Mercenaria mercenaria notata. A: 1st mitotic Metaphase; B–C: 1st cleavage, 2-cell stage; D: 2nd cleavage, 4–cell stage; E: 3rd cleavage, 8–cells stage; and F: 4th cleavage, 16–cell stage.
A B
C D
E F

Improving t
ISSN: 1687
Table 1: Pr in
Treatment
post–insem
Control, 0 m
Control, 10 m
PB I, 10 min
Control, 20 m
PBII, 20 min
Control, 55 m
PBI, 55 min
PBII, 55 min
Figure 3
triploidy indu
7-4285
roportions (%nduced to spaw
t, time
mination GV
min (T0) 0.0
min 0.0
n 0.0
min 0.0
n 0.0
min 0.0
0.0
n 0.0
3: Mean (± SDfirst cleavag
uction in the ha
%) of eggs at eawn by therma
pro I Me
20.7 6.9
0.0 2.2
0.0 2.9
0.0 0.0
0.0 0.0
0.0 0.0
0.0 0.0
0.0 0.0
D, n=3) time pge formation in
ard clam
ach developmal cycles on sp
t I Ana I
9 13.8
2 4.4
9 0.0
0 0.0
0 2.3
0 0.0
0 0.0
0 0.0
post-inseminan eggs of the h
Egyptian
mental stage ofpawning table.
% of eggs at eac
Tel I PB I
3.4 0.0
0.0 0.0
0.0 0.0
0.0 0.0
4.5 0.0
0.0 0.0
0.0 0.0
0.0 0.0
ation of first phard clam Me
Journal of Aq
f the hard clam.
ch developmental
Pro II Met
0.0 31.0
0.0 0.0
0.0 5.9
0.0 0.0
0.0 0.0
0.0 0.0
0.0 0.0
0.0 0.0
polar body (PBrcenaria merc
quatic Researc
m Mercenaria
l stage
II Ana II
0 13.8
0 0.0
9 0.0
0 2.9
0 0.0
0 0.0
0 0.0
0 2.5
BI), second pcenaria notata
ch, 2009, 35(3
mercenaria n
Tel II PB II
0.0 0.0
0.0 0.0
0.0 0.0
0.0 0.0
2.3 0.0
0.0 0.0
0.0 0.0
0.0 0.0
olar body (PBa.
3), 385-403
391
notata
> PB II
10.3
93.3
91.2
97.1
90.9
100.0
100.0
97.5
BII), and

Eman El-Wazzan, and John Scarpa
Egyptian Journal of Aquatic Research, 2009, 35(3), 385-403 ISSN: 1687-4285
392
Figure 4: Epifluorescence micrographs of eggs and embryos of the hard clam, Mercenaria mercenaria notata.
Oocytes and embryos were visualized with DNA–specific flurochrome Hoechst 33258 and epifluorescence microscopy. A: Good synchrony of eggs at anaphase I; B-C: Lack of synchrony (some eggs already entered mitosis, arrows); D–G:
Table 2: Proportions (%) of eggs at each developmental stage in eggs of the hard clam Mercenaria mercenaria
notata induced to spawn by irradiated sperm in separate containers.
Treatment, time post–fertilization
% Number of eggs at each developmental stage
GV pro I Met I Ana I Tel I PB I Pro II Met II Ana II Tel II PB II > PB II
Control, 0 min (T0) 0.0 76.3 23.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 Control, 10 min 0.0 8.1 44.7 32.9 14.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 PB I, 10 min 0.0 7.7 46.2 30.8 15.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 Control, 20 min 0.0 9.4 5.6 8.3 36.1 16.7 10.0 2.8 11.1 0.0 0.0 0.0 PBII, 20 min 0.0 6.7 10.0 6.7 20.0 16.7 13.3 16.7 3.3 6.7 0.0 0.0 Control, 55 min 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 100.0 PBI, 55 min 0.0 5.3 5.3 0.0 5.3 0.0 0.0 0.0 0.0 0.0 0.0 84.2 PBII, 55 min 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 100.0
A B C
D E F
G

Improving t
ISSN: 1687
Figure 5.
At the timeinseminatiotargeted srespectivelyentered mitPBII in PB43% and 67treatment fo3A). Theircytometry treatments, fertilization
A
D
triploidy indu
7-4285
Epifluorescenmercenaria nvisualized wiinhibition occbodies were sperm spread
e of treatmenton), only 2% tages (Pre–Py (Table 3A)tosis at the tim
BI and PBII g7% of eggs haor PBI and PBr triploid propwere 39%
respectivelyn was observ
A
D
uction in the ha
nce micrograpnotata. Embith DNA–speccurred); D–Einhibited (pubetween the t
t to inhibit PBand 30% of
PBII) in Tr). In Trial 1me of treatmegroups, Table ad entered mitBII groups, reportions as mfor PBI an
y (Table 3Aed in Trials
ard clam
phs of embrybryos treated cific flurochro: Only one p
utative pentaptwo blastomer
BII (20 min peggs were at
rials 1 and1, most eggs ent (91% at P
3A). In Triatosis at the timespectively (T
measured by fld 0% for PA & B). P2 and 8 and
B
E
G
Egyptian
yos (55 min pwith CB at
ome Hoechst polar body prploid embryosres.
post–t the
d 3, had
Post–al 3,
me of Table flow-PBII Poor
d 10;
47%(Tabby f(Tab 3.3.2
Ctriplin Tin wused
Journal of Aq
post–insemina20 min pos
33258. A–Cresent (putativs). Arrows sh
%, 44% and 68ble 3A & B). flow-cytometrble 3A & B).
2. Cytochalas
Cytochalasin oidy inductio
Trial 3 and comwhich the lowed. In Trial 5 in
quatic Researc
ation) of the st-inseminatio: Embryos wive triploid emhow chromati
8% of eggs we The triploid
ry were 17%,
sin B concentr
B concentraon. Triploid impletely in Ter CB concenn spite of 100
C
F
ch, 2009, 35(3
hard clams, Mon for 15 miith both polar
mbryo); F-G: in of the UV
ere fertilized rproportions a77%, 26%, r
ration
ation had aninduction fail
Trials 4 and 5 ntration (0.67 % of the eggs
3), 385-403
393
Mercenaria nutes were
r bodies (no Both polar
V–irradiated
respectively as measured respectively
n effect on ed partially (Table 4A) mg/L) was
s being at

Eman El-Wazzan, and John Scarpa
Egyptian Journal of Aquatic Research, 2009, 35(3), 385-403 ISSN: 1687-4285
394 the targeted stages, no triploids were obtained (Table 4A). In contrast, Trials 1 and 2 and 6 through 10 that used 1mg CB/L, produced triploids with different proportions according to other induction conditions (Table 4A & B). 3.3.3. Targeted developmental stages
Comparison of PBI treatment among the ten trials showed that at sampling times (9 to 11 min post–insemination) a large proportion (83%–100%) of eggs were at Pre–PBI in trials 4–7. In Trial 6, 90% of eggs were at the targeted stage (Pre–PBI). Measured triploid percentage was similar to expected (93%, 90%, respectively; Table 3B). In Trial 8, although only 55% of eggs were at the targeted stage (Pre–PBI), actual produced triploid percentage was 77%. In Trial 8, 21% of eggs were also at Pre–PBII (Table 3B). The highest triploid proportion in PBI treatments was obtained in Trial 6 (93%) and next highest was in Trial 8 (77%). Trial 9 had the lowest proportion of eggs at the targeted stage (41%) in the sample collected 6 min post-insemination and yielded 48% triploids (Table 3B) as measured by flow-cytometry. In Trial 7, 85% of eggs were at the Pre–PBI stage at time of treatment, however, only 33% triploids were found in larvae (Table 3B)
For PBII inhibition, CB treatment was suggested to at 15–20 min post-insemination (Figure 3). Start times in the range of 15.5 – 21 min post–insemination were used (Table 3A & B). The lowest triploid proportions obtained were 33% in Trial 6 and 57% in Trial 9 when CB was added at 21 min and at 15.5 min post–insemination, respectively (Table 3B). In Trial 6, at 21 min post–insemination, 44% of eggs were at Pre–PBII, but 56% had passed the targeted stage and entered mitosis (Table 3B). In Trial 9 only 26% of the eggs were at Pre–PBII, but 66% of eggs were at Pre–PBI, which may have increased the proportion of triploids as measured by flow–cytometry (57%, Table 3B). Higher proportions of triploids were noted in larvae from PBII treatments of Trials 7, 8 and 10 than that expected from cytological data (i.e., proportion of eggs at targeted stages; Table 3B).
4. Discussion 4.1. Timing of meiotic and mitotic events
The hard clam Mercenaria mercenaria releases eggs and sperm into the water and fertilization takes place externally (Belding 1931 in Eversole 2001). However, the egg is released pre-meiotic, therefore, meiosis I and meiosis II occur externally during which the two polar bodies are extruded. Triploidy induction in mollusks can be attained by inhibiting either of these two polar bodies (Beaumont and Fairbrother 1991). The timing of meiotic and mitotic events in hard clam
eggs were examined in the present study for use in triploidy induction trials.
In the present study, PBI appeared at 13 min and PBII at 22 min post–insemination (PI). The timing of PBI extrusion was comparable to that observed (15 min PI) by Belding (1931 in Eversole 2001). No data could be obtained regarding PBII extrusion. In dwarf surf clam, Mulinia lateralis, PBI and PBII were formed at 30 and 35 min post–insemination at 20oC (Longo and Scarpa 1991). In a different study, Scarpa et al. (1992) found that 50% of dwarf surf clam eggs extruded PBI at 20 min and PBII at 31 min post–insemination at 21oC. In the blue mussel, Mylillus galloprovincialis, more than 94% of eggs were at Ana I/Met II stages at 20 min post–insemination (Scarpa et al 1994a).
In the present study, first cleavage was first observed at 40 min post–insemination. However, the maximum formation of 2–cell stage was at 60 min post-insemination at 29oC. According to Belding (1931 in Eversole 2001), first cleavage started earlier (30 min PI) than in the present study at a comparable temperature range of 27–30oC. Carriker (2001) reported that eggs of Mercenaria mercenaria at about 22oC arrived to 2–cell stage at 45–50 min PI. In contrast, slower development was observed for the zhikong scallop, Chlamys farreri, at 20oC; first cleavage was seen at 105–120 min PI (Pan et al. 2004). First cleavage was observed in all crosses of the Pacific oyster, Crassostrea gigas, and the Suminoe oyster, C. rivularis, at 90 min PI at temperatures between 23 and 25oC (Scarpa and Allen 1992). These results show variation in early developmental rates among molluscan species as well as for the same species at different conditions. According to the results of this study, induction of triploidy in hard clam, Mercenaria mercenaria, egg should begin at 5–10 min PI for PBI inhibition and 15–20 min PI for PBII inhibition.
Chemical treatment should not last more than 30–40 min PI to avoid inhibiting mitotic events. However, rigorous testing of the chosen timing of induction was still required and is discussed in the following sections. 4.2. Induction of triploidy
Spawning of clams en masse is induced by thermal cycles on spawning tables as a regular procedure in most commercial culture facilities (Hadley et al. 1997). The broodstock are placed in the spawning table, which is filled with seawater of required alternating temperatures. However, these procedures allow passage of water carrying sperm over the whole broodstock resulting in sperm being carried over with females (on shell and internally) into individual containers. This is probably what happened in the first trial of triploidy induction. Therefore, to control fertilization before triploidy induction, spawning and

Improving triploidy induction in the hard clam
ISSN: 1687-4285 Egyptian Journal of Aquatic Research, 2009, 35(3), 385-403
395
Table 3A: Cytological data from different experiments (Trials 1-5) for attempting triploidy induction in the hard clam Mercenaria mercenaria notata. .
Pre-PBI Pre-PBII Post-PBII
Control, T0 85 15 17 45 10PBI, 10 min 85 15 0 6 91 0 **NAPBII, 20 min 88 12 7 2 91 2 **NAControl, T0 0 100 0 0 0PBI, 10 min 23 77 46 0 0 46 poor resol.PBII, 20 min 47 53 27 47 0 47 17%Control, T0 55 45 2 2 96PBI, 10 min 90 10 50 7 43 50 39PBII, 20 min 100 0 3 30 67 30 0Control, T0 5 95 0 0 100PBI, 11 min 99 1 83 2 3 83 0PBII, 20 min 100 0 23 77 0 77 0Control, T0 1 99 0 0 100PBI, 10 min 91 9 100 0 0 100 0PBII, 20 min 100 0 0 100 0 100 0
Triploids (%)Fertilization evaluation Expected
(cytological data)
Measured (flow
cytometry)%
fertilized %
unfertilized
Cytological Data*Targeted Developmental Stages
(% of fertilized eggs)
5
2
3
4
2F:2M
5F:2M
4F:3M
2F:1M
Induction Conditions
1
Females:Males
6F:Mass (3mL)
Treatment, Egg Sampling Time
post-insemination
Trial #
* Synchrony of clam eggs at targeted stages: proportion of eggs at Pre-PBI (Anaphase I/ Telophase I), Pre-PBII (PBI to Telophase II), Pot-PBII (PBII and Mitosis) at time of treatment. Fertilized eggs at Prophase I and Metaphase I stages considered among the total fertilized eggs (add to 100%) when calculating the proportion of each stage as % of total fertilized. ** Not analyzed. Table 3B: Cytological data from different experiments (Trials 6–10) for attempting triploidy induction in the hard clam Mercenaria mercenaria notata.
Pre-PBI Pre-PBII Post-PBIIControl, T0 5 95 0 0 100
PBI, 9.5 min 88 12 90 10 0 90 93PBII, 21 min 96 4 0 44 56 44 33Control, T0 0 100 0 0 0PBI, 9 min 97 3 85 0 0 85 33
PBII, 16.5 min 100 0 28 72 0 72 83Control, T0 6 94 0 0 100PBI, 9 min 44 56 55 21 0 55 77
PBII, 16.5 min 87 13 56 39 5 39 86Control, T0 5 95 0 0 0PBI, 6 min 100 0 41 0 4 41 48
PBII, 15 min 99 1 66 26 8 26 57Control, T0 4 96 0 0 0
PBI, 7.5 min 68 32 69 2 8 69 26PBII, 17 min 96 4 36 48 13 48 69
6
10
7
8
9
2F:1M
2F:1M
2F:1M
2F:1M
3F:2M
Trial #Females:
Males
Treatment, Egg Sampling Time post-
insemination
*Targeted Developmental Stages (% of fertilized eggs)
Fertilization evaluation%
fertilized %
unfertilized
Triploids (%)Expected
(cytological data)
Measured (flow
cytometry)
Induction Conditions Cytological Data
* Synchrony of clam eggs at targeted stages: proportion of eggs at Pre-PBI (Anaphase I/ Telophase I), Pre-PBII (PBI to Telophase II), Pot-PBII (PBII and Mitosis) at time of treatment. Fertilized eggs at Prophase I and Metaphase I stages considered among the total fertilized eggs (add to 100%) when calculating the proportion of each stage as % of total fertilized.

Eman El-Wazzan, and John Scarpa
Egyptian Journal of Aquatic Research, 2009, 35(3), 385-403 ISSN: 1687-4285
396 Table 4 A: Triploidy induction condition and resultant triploids (%) from different experiments (Trials 1-5) for
attempting triploidy induction in the hard clam Mercenaria mercenaria notata.
Start End Expoure
Control, T0
PBI, 10 min 10.50 21.00 10.50 **NAPBII, 20 min 20.00 35.00 15.00 **NAControl, T0
PBI, 10 min 10.50 21.00 10.50 poor resol.PBII, 20 min 20.00 35.00 15.00 17%Control, T0
PBI, 10 min 10.00 22.00 12.00 39PBII, 20 min 20.00 35.00 15.00 0Control, T0
PBI, 11 min 11.00 23.00 12.00 0PBII, 20 min 20.00 35.00 15.00 0Control, T0 0.00PBI, 10 min 10.00 21.00 11.00 0PBII, 20 min 20.00 35.00 15.00 0
29
27‐28
28‐29
27
Triploidy Induction Conditions
1 1.00
5 0.67
2 1.00
3 0.67
4 0.67
31
Trial # T (oC)*CB Conc.
(mg/L)
Timing of CB Application (min) Measured (flow
cytometry)
Salinity (ppt.)
Treatment, Egg Sampling Time
post-insemination
27
29
31
28‐29
27‐28
* The polar body inhibiting agent, cytochalasin B. **Not analyzed.
Table 4B: Triploidy induction condition and resultant triploids (%) from different experiments (Trials 6–10) for attempting triploidy induction in the hard clam Mercenaria mercenaria notata.
Start End Expoure
Control, T0
PBI, 9.5 min 10.00 20.00 10.00 93PBII, 21 min 21.00 35.00 14.00 33Control, T0
PBI, 9 min 9.50 20.00 10.50 33PBII, 16.5 min 17.00 30.00 13.00 83
Control, T0
PBI, 9 min 9.50 20.00 10.50 77PBII, 16.5 min 17.00 32.00 15.00 86
Control, T0
PBI, 6 min 6.50 16.50 10.00 48PBII, 15 min 15.50 30.00 14.50 57Control, T0
PBI, 7.5 min 8.25 19.25 11.00 26PBII, 17 min 18.00 33.00 15.00 69
Measured (flow
cytometry)Salinity (ppt.)
Triploidy Induction Conditions
Trial # T (oC)*CB Conc.
(mg/L)
Timing of CB Application (min)Treatment, Egg
Sampling Time post-insemination
10 1.00
7 1.00
8 1.00
9 1.00
27‐28‐26 26.5
29‐30‐27 26.5‐27
29‐30‐28 27
27
29 27
6 1.0530
* The polar body inhibiting agent, cytochalasin B.

Improving triploidy induction in the hard clam
ISSN: 1687-4285 Egyptian Journal of Aquatic Research, 2009, 35(3), 385-403
397
fertilization of clam eggs should take place in separate containers, which was utilized in the later triploidy induction attempts.
Very low survival was observed in CB–treated groups (1.4% and 2.2% in PBI and PBII, respectively) compared to the control group for clams spawned on the spawning table. Detailed cytological observation of the egg stages at the time of treatment indicated that most of the eggs had already started mitotic events at the time of CB treatments. Therefore, CB treatment was blocking early mitosis instead of meiosis, which resulted in abnormal embryo development (karyokinesis without parallel cell division leading to multiple-nuclei embryos) and very high mortality.
Cytochalasin B is an effective triploid inducing agent in producing viable triploids (Beaumont and Fairbrother 1991). However, the effect of CB on larval survival, especially the early larval stages, is conflicting. Eggs of the blue mussel, Mytillus galloprovincialis, treated with 1 mg CB/L showed 0% mortality in D–stage larvae (Scarpa et al. 1994b). In contrast, high mortalities (74%–76%) were observed in the embryos of the European clam (Ruditapes decussates) after using 1 mg CB/L (Gerard et al. 1994). In the soft–shell clam Mya arenaria, Allen et al. (1982) observed altered early developmental events and indication of disruption throughout a four–hour period in 1 mg CB/L treated groups. Similarly, survival to pediveligar larvae was lower for triploids than diploids in the Venerid clam, Tapes dorsatus; even within triploid treatments, survival was highest in groups with lowest triploid proportions (Nell et al. 1995). These data indicate a negative effect of CB on survival of larvae of some species of bivalves, even if used at the correct time for inducing triploidy. The treatment that was used in the present study was not expected to result in such high mortality as the same concentration and similar or shorter exposure time was used compared to other studies. High mortality may be attributed not only to the CB, but to its use during mitotic events that led to deformation of embryos and subsequent death of larvae.
The second trial conducted utilized clams in separate containers to prevent pre-fertilization and synchronize the targeted developmental stages. This would allow for testing the correct timing of chemical treatment for inhibition of the first or second polar bodies. Although repeated temperature cycling normally induces spawning in hard clams (Hadley et al. 1997), repeated attempts did not induce spawning; further stimulation was still necessary. Hadley et al. (1997) suggested that further stimulation with sperm or eggs might be necessary, especially if the clams are not very ripe. However, live sperm would cause pre–fertilization. Therefore, ultra–violet light (UV)–irradiated sperm were tested as a chemical cue to induce spawning without fertilizing the spawned eggs (Scarpa 2002). UV–irradiation had been used as a genetic manipulation technique to inactivate sperm
DNA, but still activate the egg to develop (Scarpa et al. 1994a).
UV–irradiated sperm induced spawning in 2 males and 2 females about 30 min after adding killed sperm without fertilizing the eggs as observed epifluorescence microscopy. This allowed for good egg meiotic synchrony at the time of insemination. However, poor fertilization (46–77%) and abnormal first cleavage were observed in both control and CB–treated treatments. This may be attributed to the use of irradiated-sperm, which may have activated the eggs (i.e., initiating the calcium waves and starting meiosis; Kline and Kline 1992, Bordignon and Smith 1999) and prevented further fertilization with live sperm. This high level of poor fertilization was not observed in any other trial during the present study in which only live sperm were used. However, abnormal mitosis was observed in the eggs that was similar to the abnormalities recorded by Longo and Scarpa (1991) in the dwarf surf clam, Mulinia lateralis, and Li et al. (2000) with the Pacific abalone, Haliotis discus. The chromatin of the UV–irradiated sperm was spread between the blastomeres of the 2-cell stage embryos. It was seen either as an extended tail near the cleavage furrow or as a bridge–like material crossing the cleavage furrow. These observations indicate that the assumed dead UV–irradiated sperm did inseminate some of eggs in both control and CB–treated groups.
In contrast to the observed meiotic synchrony at 10 min post–insemination in eggs treated with irradiated sperm, eggs were not meiotically synchronized at 20 min post-insemination (27% were at Pre–PBI and 47% at Pre–PBII). Therefore, the CB treatment probably interfered with the extrusion of both first polar body and the second polar body. This may be the reason for the absence of both polar bodies in 55% of the embryos of the PBII group, which may have led to the formation of tetraploids, pentaploids or both. Larvae with a ploidy higher than triploid are expected to show low survival. This assumption is based upon the results of Peruzzi and Guo (2002) who found that inhibition of polar body I and II in surf clams produced mainly pentaploids and embryos with ploidy levels higher than triploid developed only to trochophores and died a few days after fertilization. In the present study, larvae in the three groups including the controls, died at 48 h post-insemination when water temperature in larval tanks increased to 32oC. This sudden mortality did not allow further comparisons among the 3 groups regarding their survival and growth.
In clams induced to spawn by UV–irradiated sperm, although better synchrony was observed in eggs, a very low proportion of triploids was produced (17%). This shows that using irradiated sperm decreased the triploid production in hard clams. Therefore, it was not used in further triploidy induction attempts.

Eman El-Wazzan, and John Scarpa
Egyptian Journal of Aquatic Research, 2009, 35(3), 385-403 ISSN: 1687-4285
398 4.3. Cytological evaluation of triploidy induction
The results of the present study indicated that four factors determined the success or failure of triploidy induction in hard clams. These were 1) fertilization (pre–fertilization or poor fertilization), 2) developmental synchrony of eggs, 3) CB concentration, and 4) time of addition and removal of CB.
Pre–fertilization and poor fertilization may affect the total production of triploids by decreasing the number of available eggs at the targeted stage for CB application. Pre–fertilization disturbs synchrony and disrupts triploidy induction especially if the pre–fertilized eggs have entered mitosis by the time of CB treatment. In this case, mitotic cleavage is inhibited producing deformed embryos that are not expected to develop to D–stage larvae (Beaumont and Fairbrother 1991). The harsh effect of CB on survival of larvae was recorded even in cases when it was applied at the correct meiotic stage (Allen et al. 1982, Gerard et al. 1994). Overall survival from eggs to competent larvae will be affected.
It was possible to avoid or reduce pre–fertilization even when using normal hatchery practices (i.e., spawning table), as was done in triploidy induction Trials 3–10. This was accomplished by moving spawning females from the spawning table into clean seawater, keeping them undisturbed until they started to spawn again and then moving them again to another container of clean seawater to complete spawning. Proportions of pre–fertilized eggs in trials in which this strategy was used eggs did not exceed 6%. Furthermore, leaving eggs for 1 hour before attempted fertilization may allow for pre–fertilized eggs to enter mitosis and synchronization of non–fertilized eggs at Met I (Scarpa and Allen 1992). This may remove the pre–fertilized eggs from the total produced population as these embryos will be deformed by inhibited mitotic cleavage and will not develop to larval stage. However, caution should be taken in storage of sperm or using fresh sperm to avoid decreased fertilization rate and increased mortality in the resulting larvae (Eversole 2001). Examining eggs at this time was useful in evaluating the resulting embryos for possible deformation and for counting polar bodies.
The effectiveness of cytochalasin B (CB) for inducing triploidy depended on both CB concentration and the proportion of eggs at the targeted stage at the time of treatment application. No or low triploid proportions were obtained at a CB concentration of 0.67 mg/L even with high proportions (85%–100%) of eggs at the correct stages. In contrast, triploidy was induced in all the trials when CB concentration was 1mg/L. Similar dose-dependent triploidy induction was observed by Gerard et al. (1994) in the European clam, Ruditapes decussates. Examining two CB concentrations, 1mg/L was more successful (95%) than 0.5 mg/L (63%) in producing triploids (Gerard et al. 1994). However, lower CB concentrations were
efficient for inducing triploidy in other bivalve species. For example, 0.25 mg CB/L was optimum in producing triploidy in the oyster Crassostrea gigas (Barber et al. 1992). Similarly, Stepto and Cook (2004) produced 70.9% PBII triploids and 48.4% PBI triploids in the South African abalone, Haliotis midae, using 0.5 mg/L CB. In the present study, a low CB concentration (0.67 mg/L) may not be the only reason behind failure in triploidy induction as it produced 39% in one of the three trials in which this concentration was used. It was not determined why no triploids were obtained in the other two trials.
Time of CB addition also affected triploid induction success. In the present study, maximum triploid inductions were obtained by using 1mg/L CB starting at 10 min post–insemination with a 10 min exposure for inhibiting PBI and at 17 min post–insemination with a 13 min exposure for inhibiting PBII. Low triploid proportions were produced when CB application started earlier than 10 min post–insemination for PBI inhibition (for example, 8.25 min in Trial 10). It seems that the problem was not the start time as 69% of eggs were at Pre–PBI at time of CB application and 26% triploids were measured by flow cytometry. Removing CB at 19.25 min post-insemination may have been too early. It is possible that more eggs were still in the pre-PBI stage at the time of CB removal due to a delay in development. Similarly, in Trial 9, when CB was added at 6.5 min and removed at 16.5 min post–insemination, only 48% triploids were produced. In this trial (Trial 9) at the time of CB addition, only 41% of eggs were ready to extrude PBI. Cytological data of eggs sampled at the time to induce PBII (15 min PI) revealed that 66% of eggs were still at Pre–PBI and only 26% of eggs had passed PBI reflecting delayed development. Therefore, removing CB at 16.5 min post–insemination allowed some of the eggs that did not extrude PBI to resume PBI extrusion. CB exerts its effect by inhibiting the microfilaments of the contractile ring whose formation is necessary for cytokinesis (Copland 1974 in Beaumont and Fairbrother 1991). Removal of CB, will allow these rings to form and cytokinesis to resume, resulting in extrusion of polar body in more eggs.
The highest proportions of triploids were produced in trials when all or most eggs had good synchrony at one or both pre–PB stages at the time of CB application. In these trials the expected proportion of triploids from the cytological data was less than the flow-cytometry data (e.g., PBI treat. in Trial 8 and PBII treat. in Trials 7–10). However, triploids may have occurred from inhibiting either PBI or PBII not just the one targeted by the designated treatment. It was observed that in some trials, although CB treatments were applied while eggs were either at the Pre–PBI or Pre–PBII stage, a low triploid proportion was produced. In contrast, when eggs were simultaneously synchronized at both Pre–PBI and Pre–PBII stages, the proportion of triploids was higher. For example,

Improving triploidy induction in the hard clam
ISSN: 1687-4285 Egyptian Journal of Aquatic Research, 2009, 35(3), 385-403
399
treatments used to inhibit PBI formation induced low triploidy levels in all trials that had eggs at only Pre–PBI. The high triploid proportions (93% and 77%, Trials 6 and 8, respectively) occurred in cases that had both stages of Pre–PBI and Pre–PBII present. Similarly, treatments used to inhibit only PBII produced triploids in all cases (33%–86%), but with higher levels (69%–86%) in trials that had both Pre–PBI and Pre–PBII stages present. The highest proportion of triploids (86%) produced among PBII treatments was observed in Trial 8, when a total of 95% of the eggs were at these stages (56% and 39% of fertilized eggs were at Pre–PBI and Pre–PBII, respectively) at the time of CB treatment. These results are in agreement with the observation of Liu et al. (2004) in the blacklip abalone, Haliotis rubra. They found that, targeting PBI produced triploid levels of 71.5% and 57.6% in two different experiments. In contrast, treatments that targeted both Pre–PBI and Pre–PBII resulted in higher triploid proportions of (84.8–89.5% and 86.5–96.5%). However, very low survival (1.8% and 5.4%) was observed in the treatments that targeted overlapped stages in the two experiments (Liu et al 2004).
It was observed that, timing of PBI or PBII extrusion could not be precisely separated when using eggs and sperm from multiple females and males. Overlapping of extrusion of a significant number of both polar bodies occurred as early as 9.5 min post-insemination and continued until 21 min post–insemination. This may be attributed to differential development of eggs from different females and sperm from different males. Therefore, it may be hard to produce pure PBI or PBII triploids of hard clams in a commercial setting. 5. Conclusions
The commercial method of mass spawning clams by thermal shock is not efficient for producing triploids because of high pre–fertilization, even with removal of females from the spawning table and being placed in separate containers to continue spawning. However, triploid induction may be possible with more caution to avoid pre–fertilization or by leaving eggs for 1 hour before attempted fertilization to allow time for all pre–fertilized eggs to develop to mitotic stages.
The recommended practice for triploid induction in clams by spawning and inseminating in separate containers is also not efficient due to the difficulty of spawning in females in the absence of chemical cues (i.e., sperm). Using UV–irradiated sperm as a chemical cue was not an appropriate practice as it affected the rate of fertilization, as well as resulted in chromosomal abnormalities that inhibited triploid induction attempts.
Depending on environmental conditions, especially temperature that hastens development, induction of triploidy using CB application should begin at 5–10 min post–insemination for PBI inhibition with an
optimum start time of 10 min. CB addition should start at 15–20 min for PBII inhibition with an optimum start time of 17 min. CB application time for PBII inhibition may be adjusted by observing eggs under light microscopy and applying CB when approximately 50% of the eggs exhibit PBI as recommended by Allen and Bushek (1992). However, optimum time for exposure to CB still needs evaluation to allow for inhibition of either polar body in all eggs, as well as to avoid inhibition of both polar bodies. Chemical treatment should last until 25 min post-insemination for PBI and 40 min for PBII inhibition. Chemical treatment should not last beyond 40 min post-insemination to avoid inhibiting mitotic events. Acknowledgements
The authors thank Mr. Joe Weissman, Ms. Stacy Plummer and Mr. Federico Prahl of HB Clams, Inc. for providing clam broodstock and assistance with spawning, respectively. We thank Dr. Randal Niedz, Mr. Scott Hyndman, and Mr. Eldridge Wynn of the USDA-Horticultural Research Laboratory in Ft Pierce for access to the Partec PA and to Ms. Sandra Scarpa for assistance with ploidy analysis. EE-W thanks the Link Foundation for her HBOI 2003 Summer Internship. This research was also, supported by grants to JS from the U.S. Department of Agriculture - Agricultural Research Services and Florida Sea Grant. This research was part of EE-W's dissertation research. Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the funding organizations. The views expressed are those of the authors and do not necessarily reflect the view of the funding organizations. This is HBOI-FAU Contribution # 1801.
References
Allen, SK. Jr.; Bushek. D.: 1992, Large scale
production of triploid oysters, Crassostrea virginica (Gmelin), using “stripped” gametes. Aquaculture, 103: 241-251.
Allen, SK. Jr., Downing, S.L.; Chew, K.K.: 1989, Hatchery manual for producing triploid oysters. University of Washington Press, Seattle, Washington, 27pp.
Allen, SK. Jr.; Gagnon, P.S.; Hidu, H.: 1982, Induced triploidy in the soft–shell clam Mya arenaria: cytogenetic and allozymic confirmation. Journal of Heredity, 73: 421-428.
Allen, S.K.Jr., Hidu, H.; Stanley, J.G.: 1986, Abnormal gametogenesis and sex ratio in triploid

Eman El-Wazzan, and John Scarpa
Egyptian Journal of Aquatic Research, 2009, 35(3), 385-403 ISSN: 1687-4285
400 soft-shell clams (Mya arenaria). The Biological Bulletin, 170: 198-210.
Barber, B.J.; Mann, R.; Allen, SK. Jr.: 1992, Optimization of triploid induction for the oyster Crassostrea virginica (Gmelin). Aquaculture 106: 21-6.
Baron, J.; Dieter, S.A.; Bodoy, A.: 1989, Triploidy induction in the black scallop (Chlamys varia L.) and its effect on larval growth and survival. Aquaculture, 77:103-11.
Beaumont AR, Fairbrother JE.: 1991, Ploidy manipulation in molluscan shellfish: a review. Journal of Shellfish Research, 10:1-18.
Belding, D.L.: 1931, The quahog fishery of Massachusetts. Commonw. Mass. Dep. Conserv., Div. Fish. Game, Mar. Ser. 2, 41 p. (cited in Eversole 2001).
Bordignon, V.; Smith, L.C.: 1999, Ultraviolet-irradiated spermatozoa activate oocytes but arrest preimplantation development after fertilization and nuclear transplantation in cattle. Biology Reproduction, 61: 1513-1520.
Brake, J.; Davison, J.; Davis, J.: 2004, Field observations on growth, gametogenesis, and sex ratio of triploid and diploid Mytilus edulis. Aquaculture, 236: 179-91.
Buzzi WR.: 1990, Effects of induced polyploidy on the growth and survival of juvenile hard clams Mercenaria mercenaria. Masters Thesis, College of Charleston, South Carolina. 58 pp.
Carriker MR.: 2001, Embryogenesis and organogenesis of veligers and early juveniles. In: Kraeuter JN and Castagna M, editors. The biology of hard clams. Amsterdam (The Netherlands): Elsevier Science. P 77–115.
Chaiton JA, Allen SK.: 1985, Early detection of triploidy in the larvae of the Pacific oysters Crassostrea gigas by flow cytometry. Aquaculture, 48: 35-43.
Eudeline B, Allen SK Jr., Guo X.: 2000, Delayed meiosis and polar body release in eggs of triploid Pacific oysters, Crassostrea gigas, in relation to tetraploid production. Journal of Experimental Marine Biology and Ecology, 248(2): 151-61.
Eversole, AG.: 1987, Species profiles: life histories and environmental requirements of coastal fishes and invertebrates (South Atlantic ) hard clam. U.S. Fish Wildl. Serv. Biol. Rep. 82 (11.75). U.S. Army Corps of Engineers, TR EL–82–4. 33 pp.
Eversole, AG.: 2001, Reproduction in Mercenaria mercenaria. In: Kraeuter JN and Castagna M, editors. The biology of hard clams. Amsterdam (The Netherlands): Elsevier Science. P 221–60.
Eversole, AG., Kempton CJ, Hadley NH, Buzzi WR.: 1996, Comparison of growth, survival, and reproductive success of diploid and triploid Mercenatia mercenaria. Journal of Shellfish Research, 15: 689-94.
Gerard, A.; Naciri, Y.; Nioret, C.; Ledu, C.; Peignon, JM.; Phelipot, P.: 1994, Induced triploidy in the European clam, Ruditapes decussates (L), and performance of triploid larvae. Aquac Fish Manag 25: 769-79.
Goldman, J.C.; Ryther, JH.: 1976, Temperature-influenced species competition in mass cultures of marine phytoplankton. Biotechnology and Bioengineering,18: 1125-1144.
Hadley, NH.; Manzi, JJ.; Eversole, AG.; Dillon, RT.; Battey, CF.; Peacock, NM.: 1997, A manual for the culture of the hard clam Mercenaria spp. Charleston (SC). Sea Grant Consortium, 135pp.
Hand RE, Nell JA, Thompson PA.: 2004, Studies of triploid oysters in Australia XIII. Performance of diploid and triploid Sydney rock oysters, Saccostrea glomerata (Gould, 1850), progeny from a third generation breeding line. Aquaculture, 233: 93-107.
Hidu, H.; Mason, KM.; Shumway, SE.; Allen, SK.: 1988, Induced triploidy in Mercenaria mercenaria L. Effects on performance in the juveniles [abstract]. Journal of Shellfish Research, 7: 202.
Kline, D.; Kline, JT.: 1992, Repetitive calcium transients and the role of calcium in exocytosis and cell cycle activation in the mouse egg. Developmental Biology, 149: 80-89.
Lee, MM.: 1988, Abnormal gametogenesis in triploid American oysters Crassostrea virginica. Journal of Shellfish Research, 7: 201-202.
Li, Q, Osada, M, Kashihara M, Hirohashi K, Kijima A.: 2000, Cytological studies on artificially induced gynogenesis in the Pacific abalone. Fish Science, 66: 701-707.
Liu, W.; Heasman, M.; Simpson. R.: 2004, Evaluation of cytochalasin B (CB) treatments for triploidy induction in the blacklip abalone, Haliotis rubra (Leach, 1814). Aquaculture Research, 35: 1062-1075.
Longo, FJ.; Scarpa, J.: 1991, Expansion of the sperm nucleus and association of the maternal and paternal genomes in fertilized Mulinia lateralis Eggs. Biological Bulletin, 180: 56-64.
Maldonado-Amparo, R.; Ramirez, JL.; Avila, S.; Ibarra, AM.: 2004, Triploid lion-paw scallop (Nodipecten subnodosus Sowerby); growth, gametogenesis, and gametic cell frequencies when grown at a high food availability site. Aquaculture, 235: 185-205.
Myrand, B.; Gaudreault, J.: 1995, Summer mortality of blue mussels (Mytilus edulis, Linneaus, 1758) in the Magdalin Islands (Southern Gulf of St Lawrence, Canada). Journal of Shellfish Research, 14: 395-404.
Nell, JA.; O’Connor, WA.; Hand, RE.; McAdam, SP.: 1995, Hatchery production of diploid and triploid clams, Tapes dorsatus (Lamarck 1818): A potential new species for aquaculture. Aquaculture, 130: 389-394.

Improving triploidy induction in the hard clam
ISSN: 1687-4285 Egyptian Journal of Aquatic Research, 2009, 35(3), 385-403
401
Ohgai, M.; Takesue, K.; Tsurata, A.: 1982, The effect of heat shock on growth of marine phytoplankton. Bulletin of the Japanese Society of Scientific Fisheries, 48: 1233-1236.
Pan, Y.; Li, Q.; Yu, R.; Wang, R.: 2004, Induction of gynogenetic diploids and cytological studies in the zhikong scallop, Chlamys farreri. Aquatic Living Resources, 17: 201-206.
Perdue, JA.; Beattie, JH.; Chew, KK.: 1981, Some relationships between gametogenic cycle and summer mortality phenomenon in the pacific oysters (Crassostrea gigas) in Washington State. Journal of Shellfish Research, 1: 9-16.
Peruzzi, S.; Guo, XM.: 2002, Tetraploid induction by meiosis inhibition with cytochalasin B in the dwarf surfclam Mulinia lateralis say: Effect of temperature. Journal of Shellfish Research, 21: 677-684.
Scarpa, J.: 2002, General method for the production of developmentally-arrested bivalve trochophore larvae as a potential food for marine fish larvae. Journal of Applied Aquaculture, 12:1-11.
Scarpa, J.; Allen, SK. Jr.: 1992, Comparative kinetics of meiosis in hybrid crosses of Pacific Oyster Crassostrea gigas and Suminoe oyster C. rivularis with the American oyster C. virginica. Journal of Experimental Zoology, 263: 316-322.
Scarpa, J.; Komaru, A.; Wada, KT.: 1994a, Gynogenetic induction in the mussel Mytilus galloprovincialis. Bulletin of National Research Institute of Aquaculture 23: 33–41.
Scarpa, J.; Toro, JE.; Wada, KT.: 1994b, Direct comparison of six methods to induce triploidy in bivalves. Aquaculture, 119: 119-133.
Scarpa, J.; Wada, KT.; Allen, SK. Jr.: 1992, Parthenogenetic development of dwarf surf clam, Mulina lateralis, oocytes treated with polar body suppressing agents. Invertebrate Reproduction and Developmen, 22: 47-56.
Shpigel, M.; Barber, BJ.; Mann, R.: 1992, Effect of elevated temperature, gametogenesis, physiology and biochemical composition in diploid and triploid Pacific Oysters, Crassostrea gigas Thunberg. Journal of Experimental Marine Biology and Ecology, 161: 15-25.
Stepto, NK.; Cook, PA.: 1998, Induction of triploidy in the South African abalone using cytochalasin B. Aquaculture International, 6: 161-9.
Surge, DM.; Lohmann, KC.: 2002, Temporal and spatial differences in salinity and water chemistry in SW Florida estuaries: Effects of human-impacted watersheds. Estuarine Coasts, 25: 393-408.
Tabarini, CL.: 1984, Induced triploidy in the bay scallop, Argopecten irradians, and its effect on growth and gametogenesis. Aquaculture, 42: 151-60.
Toro JE, Sastre HD.: 1995, Induced triploidy in the Chilean blue mussel, Mytilus chilensis (Hupe, 1854), and performance of triploid larvae. Journal of Shellfish Research, 14: 161-4.
Utting, SD.; Child, AR.: 1994, Genetic manipulation of Manila clam (Tapes philippinarum) using cytochalasin B to induce triploidy. Aquaculture, 120: 271-82.
Utting, SD.; Millican, PF.; Laing, I.: 1996, The breeding potential and biochemical composition of triploid Manila clams, Tapes philippinarum Adams and Reeve. Aquaculture Research, 27: 537-580.
Weiss, MB.; Curran, PB.; Peterson, BJ.; Gobler, CJ.: 2007, The influence of plankton composition and water quality on hard clam (Mercenaria mercenaria L.) populations across Long Island's south shore lagoon estuaries (New York, USA). Journal of Experimental Marine Biology and Ecology, 345: 12-25.
Yang, H.; Wang, J.; Guo, X.: 2001, Production and evaluation of meiosis I and II triploids in the hard clam Mercenaria mercenaria. Aquaculture, Book of Abstracts, p.708.

Eman El-Wazzan, and John Scarpa
Egyptian Journal of Aquatic Research, 2009, 35(3), 385-403 ISSN: 1687-4285
402
البطلينوس الصلد من نوع مرسيناريا فيتحسين الحث علي تخليق ثالثي الكروموسومات
تقييم خلوي للبويضات المخصبة: مرسيناريا نوتاتا
ين الحيوانات ثالثية الكروموسومات تحوي ثالثة مجموعات من الكروموسومات بدال من احتوائها علي اثن
لبية ؤثرات الس ة للم ا المناعي ومن . آالعادة ، ولذلك تصبح هذه الحيوانات عقيمة مما قد يحسن نموها ومقاومته
اق الجسم القطبي االول يط انبث ة الكروموسومات بواسطة تثب اي )Polar body I( الممكن أن تستحث ثالثي
(PBI) اني او وزي ) Polar body II, PBII(الجسم القطبي الث ام المي اء االنقس ي (أثن باستخدام ) االختزال
ويعتمد انتاج ثالثية الكروموسومات في ). Cytochalasin B, CB(بعض الكيماويات مثل سيتوآاالزين ب
ى يح وعل د التلق وزي في البويضات بع ام المي المحاريات ذات المصراعين علي تزامن حدوث مراحل االنقس
ع ذلك فقد تم تحدي ك بتجمي ي في بويضات البطلينوس الصلد الملقحة، وذل د توقيتات مراحل االنقسام االختزال
ى يح حت د التلق ة بع ات مختلف د اوق يح وعن ل التلق اث قب ة ان ات البويضات المفرخة من ثالث د ٩٠عين ه بع دقيق
وميه ادة الكروموس ار الم م اختب يح، ث والي ) DNA(التلق ي ح ت ٩٠ف ة باس ل عين ن آ خدام الفحص بويضة م
بعد صبغ البويضات بصبغة مشعة متخصصة لصبغ ) epifluereseence microscopy(الميكروسكوبي
ى تكون للجسم القطبي االول .)Hoechst 33258( ٣٣٢٥٨الكروموسومات وهي مادة هوخست وآان اعل
د اني بع ي الث م القطب ي أول ٢٨، ١٦والجس ر أعل والي ، وظه ي الت د التخصيب عل ة بع ج دقيق ام ( تفل انقس
يح ٦٠للخلية بعد مرور حوالي ) ميتوزي ي . دقيقة من التلق وين ثالث ي تك م الحث عل ائج ، ت ذه النت باستخدام ه
يتوآاالزين ب زات Cytochalasin B, CB)(الكروموسومات في عشرة محاوالت باستخدام مادة الس بترآي
دة تعرض ١١حتي ٦.٥عند توقيتات من لتر من معلق البويضات/مجم ١او ٠.٦٧ يح لم د التلق -١٠دقيقة بع
اق الجسم القطبي االول ١٢ يط انبث ي ١٥.٥من و دقيقة لتثب دة تعرض ٢١ال ة لم يط ١٥-١٣دقيق ة لتثب دقيق
.الجسم القطبي الثاني
تم تقييم النجاح في هذه المحاوالت بمقارنة نسب البويضات عند مراحل ما قبل انبثاق الجسم القطبي االول
ي الكروموسومات (انبثاق الجسم القطبي الثاني او ة لتكون ثالث ي النسبة المتوقع د وقت استخدام ) مما يعن عن
وي اس السريان الخل ات والمقاسة بمقي ي الكروموسومات في اليرق المادة المحفزة، بالنسبة الفعلية لتكون ثالث
)flow cytometry(.
وت %. ٩٢ – ٠والفعلية من % ١٠٠ – ٠من وقد تراوحت نسب ثالثي الكروموسومات المتوقعة د احت وق
يتوآاالزين ب ز الس ة ترآي ي قل ي الكروموسومات عل ون ثالث اءة حث تك ي آف ؤثرة بالسلب عل الظروف الم
ر /مجم ١لتر بالمقارنة ب /مجم ٠.٦٧( ة، اضافة او ) لت ة التجرب ل بداي ادة نسبة البويضات المخصبة قب ، زي
الي نزع مادة السيتوآاالزين ب الم زمن المث أخرا عن ال ذلك . حفزه لتكون ثالثي الكروموسومات مبكرا او مت ل

Improving triploidy induction in the hard clam
ISSN: 1687-4285 Egyptian Journal of Aquatic Research, 2009, 35(3), 385-403
403
يتوآاالزين ادة الس اظهرت النتائج أنه لحث تكوين ثالثي الكروموسومات في البطلينوس الصلد فإن استخدام م
لتثبيط الجسم دقيقة بعد التلقيح ١٧دقائق بعد التلقيح لتثبيط الجسم القطبي االول وعند ١٠ب يجب ان تبدأ عند
د ا بع يتوآاالزين ب لم دي التعرض للس اني وال يجب ان يتع ي الث م ٢٥القطب يط الجس يح لتثب ن التلق ة م دقيق
.دقيقة بعد التلقيح لتثبيط الجسم القطبي الثاني ٤٠القطبي االول أو




















