
Genetic, cytological and morphological differentiation within the Balkan-Carpathian Sesleria rigida...
15
458 TAXON 62 (3) • June 2013: 458–472 Kuzmanović & al. • Differentiation of Sesleria rigida sensu Fl. Eur. 458 Version of Record (identical to print version). INTRODUCTION The Balkan Peninsula and the adjacent Carpathians are centres of European biodiversity (Turrill, 1929; Davis & al., 1994; Kryštufek & Reed, 2004). In the past ten years several phylogeographic studies have explored diversification patterns on the Balkan Peninsula (e.g., Park & al., 2006; Liber & al., 2008; Stefanović & al., 2008; Kučera & al., 2010; Surina & al., 2011) and in the Carpathians (e.g., Mráz & al., 2007; Puşcaş & al., 2008; Ronikier & al., 2008). However, only a few (e.g., Frajman & Oxelman, 2007; Csergö & al., 2009) have exam- ined the biogeographic connections between these two areas, previously acknowledged on the basis of chorological patterns (e.g., Meusel & al., 1965; Ronikier, 2011). A major force in plant evolution is polyploidy (e.g., Wendel, 2000; De Bodt & al., 2005; Soltis & al., 2009; Van de Peer & al., 2009; Jiao & al., 2011), which fosters adaptation to new ecological niches and confers reproductive isolation, ultimately leading to speciation (Otto & Whitton, 2000; Comai, 2005; but see Mayrose & al., 2011). As much as 15% of angiosperm spe- ciation events are associated with an increase in ploidy (Wood & al., 2009). Whereas allopolyploids usually differ conspicu- ously from their diploid progenitors in their genomic constitu- tion and morphology, autopolyploids that arise from the crosses within or between populations of a single species (Ramsey & Schemske, 1998) are often genetically and morphologically similar to their parents (Levin, 1983, 2002; Parisod & al., 2010). Recent molecular studies, which often made use of methodo- logical advances in flow cytometry such as the availability of protocols optimised for the use of dried plant material (Suda & Trávníček, 2006a, b), indicate a much higher frequency of autopolyploidy than previously assumed (Husband & Sabara, 2004; Kron & al., 2007; Kolář & al., 2009; Bardy & al., 2010). Despite the widely recognised importance of polyploidisation for plant diversification, very little is known about its contri- bution to the high diversity on the Balkan Peninsula. The genus Sesleria Scop. (Pooideae, Seslerieae) comprises ca. 28 mostly European species (Deyl, 1980), with a centre of diversity on the Balkan Peninsula. It is taxonomically intricate which may relate to the high incidence of polyploidy in the genus, as the majority of taxa are tetra- and/or octoploid (2n = 4x = 28, 2n = 8x = 56, e.g., Strgar, 1979; Lysak & Doležel, 1998; Petrova, 2000). One of the polyploid species is S. rigida Heuff. ex Rchb. sensu Fl. Eur. (Deyl, 1980; for simplicity termed “S. rigida s.l.” in the following). So far only tetraploids were reported for this taxon (Ujhelyi, 1959, 1960; Deyl, 1980; Petrova, 2000), even though Deyl (1946) assumed that octoploids are present in some morphologically deviating populations. Genetic, cytological and morphological differentiation within the Balkan-Carpathian Sesleria rigida sensu Fl. Eur. (Poaceae): A taxonomically intricate tetraploid-octoploid complex Nevena Kuzmanović, 1 Petronela Comanescu,2 Božo Frajman, 3 Maja Lazarević, 1 Ovidiu Paun, 4 Peter Schönswetter3 & Dmitar Lakušić1 1 Institute of Botany and Botanical Garden, Faculty of Biology, University of Belgrade, Takovska 43, 11000 Belgrade, Serbia 2 Botanical Garden “Dimitrie Brandza”, Sos. Cotroceni 32, 060101 Bucharest, Romania 3 Institute of Botany, University of Innsbruck, Sternwartestraße 15, 6020 Innsbruck, Austria 4 Department of Systematic and Evolutionary Botany, University of Vienna, Rennweg 14, 1030 Vienna, Austria Authors for correspondence: Nevena Kuzmanović, [email protected]; Peter Schönswetter, [email protected] Abstract Reconstruction of relationships among populations of the morphologically polymorphic and taxonomically intricate Sesleria rigida sensu Fl. Eur. based on Amplified Fragment Length Polymorphisms (AFLPs) revealed four clearly differentiated genetic groups that did only partly follow recent taxonomic concepts, but were strictly allopatric. While some of the previ- ously described taxa constitute distinct genetic entities, others have no taxonomic value. Synthesizing our AFLP data with ploidy-level information obtained from all genetically investigated individuals as well as with chromosome counts revealed that tetraploid individuals prevail, while octoploids occur only within S. filifolia. Lack of AFLP divergence between tetra- and octoploids suggests an autopolyploid origin of the latter. The genetic differentiation pattern was reflected by morphological differentiation, allowing for a taxonomic revision of the constituents of S. rigida sensu Fl. Eur. resulting in recognition of the four species S. achtarovii, S. filifolia, S. rigida, and S. serbica. Keywords AFLP; autopolyploidy; flow cytometry; morphometrics; Sesleria rigida; taxonomy Supplementary Material The Electronic Supplement (Fig. S1) is available in the Supplementary Data section of the online version of this article (http://www.ingentaconnect.com/content/iapt/tax). Received: 12 Nov. 2012; revision received: 11 Mar. 2013; accepted: 27 Apr. 2013
Transcript of Genetic, cytological and morphological differentiation within the Balkan-Carpathian Sesleria rigida...

458
TAXON 62 (3) • June 2013: 458–472Kuzmanović & al. • Differentiation of Sesleria rigida sensu Fl. Eur.
458 Version of Record (identical to print version).
IntroductIon
The Balkan Peninsula and the adjacent Carpathians are centres of European biodiversity (Turrill, 1929; Davis & al., 1994; Kryštufek & Reed, 2004). In the past ten years several phylogeographic studies have explored diversification patterns on the Balkan Peninsula (e.g., Park & al., 2006; Liber & al., 2008; Stefanović & al., 2008; Kučera & al., 2010; Surina & al., 2011) and in the Carpathians (e.g., Mráz & al., 2007; Puşcaş & al., 2008; Ronikier & al., 2008). However, only a few (e.g., Frajman & Oxelman, 2007; Csergö & al., 2009) have exam-ined the biogeographic connections between these two areas, previously acknowledged on the basis of chorological patterns (e.g., Meusel & al., 1965; Ronikier, 2011).
A major force in plant evolution is polyploidy (e.g., Wendel, 2000; De Bodt & al., 2005; Soltis & al., 2009; Van de Peer & al., 2009; Jiao & al., 2011), which fosters adaptation to new ecological niches and confers reproductive isolation, ultimately leading to speciation (Otto & Whitton, 2000; Comai, 2005; but see Mayrose & al., 2011). As much as 15% of angiosperm spe-ciation events are associated with an increase in ploidy (Wood & al., 2009). Whereas allopolyploids usually differ conspicu-ously from their diploid progenitors in their genomic constitu-tion and morphology, autopolyploids that arise from the crosses
within or between populations of a single species (Ramsey & Schemske, 1998) are often genetically and morphologically similar to their parents (Levin, 1983, 2002; Parisod & al., 2010). Recent molecular studies, which often made use of methodo-logical advances in flow cytometry such as the availability of protocols optimised for the use of dried plant material (Suda & Trávníček, 2006a, b), indicate a much higher frequency of autopolyploidy than previously assumed (Husband & Sabara, 2004; Kron & al., 2007; Kolář & al., 2009; Bardy & al., 2010). Despite the widely recognised importance of polyploidisation for plant diversification, very little is known about its contri-bution to the high diversity on the Balkan Peninsula.
The genus Sesleria Scop. (Pooideae, Seslerieae) comprises ca. 28 mostly European species (Deyl, 1980), with a centre of diversity on the Balkan Peninsula. It is taxonomically intricate which may relate to the high incidence of polyploidy in the genus, as the majority of taxa are tetra- and/or octoploid (2n = 4x = 28, 2n = 8x = 56, e.g., Strgar, 1979; Lysak & Doležel, 1998; Petrova, 2000). One of the polyploid species is S. rigida Heuff. ex Rchb. sensu Fl. Eur. (Deyl, 1980; for simplicity termed “S. rigida s.l.” in the following). So far only tetraploids were reported for this taxon (Ujhelyi, 1959, 1960; Deyl, 1980; Petrova, 2000), even though Deyl (1946) assumed that octoploids are present in some morphologically deviating populations.
Genetic, cytological and morphological differentiation within the Balkan-Carpathian Sesleria rigida sensu Fl. Eur. (Poaceae): A taxonomically intricate tetraploid-octoploid complexNevena Kuzmanović,1 Petronela Comanescu,2 Božo Frajman,3 Maja Lazarević,1 Ovidiu Paun,4 Peter Schönswetter3 & Dmitar Lakušić1
1 Institute of Botany and Botanical Garden, Faculty of Biology, University of Belgrade, Takovska 43, 11000 Belgrade, Serbia2 Botanical Garden “Dimitrie Brandza”, Sos. Cotroceni 32, 060101 Bucharest, Romania3 Institute of Botany, University of Innsbruck, Sternwartestraße 15, 6020 Innsbruck, Austria4 Department of Systematic and Evolutionary Botany, University of Vienna, Rennweg 14, 1030 Vienna, AustriaAuthors for correspondence: Nevena Kuzmanović, [email protected]; Peter Schönswetter, [email protected]
Abstract Reconstruction of relationships among populations of the morphologically polymorphic and taxonomically intricate Sesleria rigida sensu Fl. Eur. based on Amplified Fragment Length Polymorphisms (AFLPs) revealed four clearly differentiated genetic groups that did only partly follow recent taxonomic concepts, but were strictly allopatric. While some of the previ-ously described taxa constitute distinct genetic entities, others have no taxonomic value. Synthesizing our AFLP data with ploidy-level information obtained from all genetically investigated individuals as well as with chromosome counts revealed that tetraploid individuals prevail, while octoploids occur only within S. filifolia. Lack of AFLP divergence between tetra- and octoploids suggests an autopolyploid origin of the latter. The genetic differentiation pattern was reflected by morphological differentiation, allowing for a taxonomic revision of the constituents of S. rigida sensu Fl. Eur. resulting in recognition of the four species S. achtarovii, S. filifolia, S. rigida, and S. serbica.
Keywords AFLP; autopolyploidy; flow cytometry; morphometrics; Sesleria rigida; taxonomy
Supplementary Material The Electronic Supplement (Fig. S1) is available in the Supplementary Data section of the online version of this article (http://www.ingentaconnect.com/content/iapt/tax).
Received: 12 Nov. 2012; revision received: 11 Mar. 2013; accepted: 27 Apr. 2013

459
Kuzmanović & al. • Differentiation of Sesleria rigida sensu Fl. Eur.TAXON 62 (3) • June 2013: 458–472
459Version of Record (identical to print version).
Sesleria rigida s.l. belongs to S. sect. Calcariae Deyl, and together with S. tenuifolia Schrad. s.l., S. insularis Sommier and S. taygetea Hayek constitutes the “turma” (= swarm) Rigida (Deyl 1946), whose centre of origin was suggested to be located on the western Balkan Peninsula (Deyl, 1946). Taxa from this species group have narrow, pruinose leaves which are most often convolute or flat. Their thin spikes are composed of large spikelets with elongated glumes and shortly awned lemmas. Sesleria rigida s.l. differs from the other species of Rigida by the absence of reticulate basal leaf sheaths (vs. S. tenuifolia s.l.), less than 13 veins in the tiller leaves and usually hairy leaves (vs. S. insularis) and lack of white bracts subtending the spikes (vs. S. taygetea). It is distributed in Romania, Bulgaria, Greece, Serbia and Bosnia (Deyl, 1946; Tatić, 1976); a doubtful and highly disjunct record from Croatia (Sekulić & al., 1988) could not be confirmed in the course of the present study (Nevena Kuzmanović, field obs.). The species thus spans several moun-tain ranges of the Balkan Peninsula (Dinaric Alps, Balkan Mts. [Stara planina], Rhodope Mts.) as well as the Romanian Car-pathians. It has a wide altitudinal distribution from lowlands to the alpine vegetation belt, growing mostly on carbonates, more rarely on serpentines. Intraspecific morphological and ecological variation of S. rigida s.l. is immense and several intraspecific taxa have been described (e.g., S. rigida var. de-genii Deyl, var. pancicii Deyl, subsp. achtarovii (Deyl) Deyl). Different authors described new species belonging to this group which were later either synonymised with S. rigida (e.g., S. filifolia Hoppe, S. haynaldiana Schur) or considered only as ecotypes, such as S. serbica (Adam.) Ujhelyi from serpentine areas in western Serbia and eastern Bosnia and Herzegovina (Deyl, 1980). Taxonomic value, delimitation and distribution of most of these taxa are not clear and they are mostly consid-ered conspecific with S. rigida (Deyl, 1946, 1980; Tatić, 1976; Diklić & Nikolić, 1986). Deyl (1980) in Flora Europaea rec-ognised only a single species, S. rigida with two subspecies, subsp. rigida and subsp. achtarovii ; all other names were neglected or considered synonyms. In most recently published national Floras Deyl’s (1980) concept with two subspecies is being followed (e.g., Tatić, 1976; Diklić & Nikolić, 1986; Gustavsson, 1991; Delipavlov & al., 2003; Assyov & Petrova, 2006; Ciocarlan, 2009). Some authors (e.g., Sekulić & al., 1988; Stevanović & al., 1995, 2003; Valdés & Scholz, 2011), following Ujhelyi (1959), recognised S. serbica as an independent species, and this was also supported by a recent leaf anatomical study (Kuzmanović & al., 2012). Also S. filifolia is treated at spe-cific rank by some authors (Gergelyi & Beldie, 1972; Assyov & Petrova, 2006; Ciocarlan, 2009), and S. haynaldiana as a subspecies of S. rigida (Gergelyi & Beldie, 1972).
Taking the intricate taxonomy of S. rigida s.l. into consid-eration, our main goal is to evaluate the taxonomic status of its constituents using molecular and morphological characters. The first aim is to reconstruct the relationships among popu-lations based on amplified fragment length polymorphisms (AFLPs; Vos & al., 1995) and to test whether the intraspecific taxa constitute distinct genetic entities. Second, we screen all genetically investigated populations for their ploidy level using flow cytometry combined with several chromosome counts
and synthesize these data with AFLPs in order to test if oc-toploids originated by auto- or allopolyploidy. Third, we search for congruence between the genetic lineages and morpholog-ically distinct groups and characterise well-supported groups with respect to their morphology, chromosome number and distribution. Finally, we provide a taxonomic treatment and an identification key for all taxa investigated.
MaterIals and Methods
Plant material. — As various taxa were described and later considered at different ranks, we assigned the investigated pop-ulations to the rankless entities “rigida“, “degenii“, “pancicii“, “achtarovii“, “filifolia“ and “serbica“ based on their morphol-ogy and distribution (Fig. 1; Appendix 1). Our study includes all described constituents of S. rigida s.l. mentioned in the In-troduction with the exception of S. haynaldiana, for which no type material could be located. Based on the protologue (Schur, 1856), where no locus classicus is specified, the identity of this taxon, which was regarded as conspecific with S. rigida s.str. by Deyl (1946, 1980) and Valdés & Scholz (2011), remains unclear.
Molecular and genome size analyses are based on leaf material of five to ten individuals per population and up to 25 populations per taxon (in total 218 individuals from 45 popu-lations). Leaf material for molecular studies was collected in the field and desiccated in silica gel. For chromosome counts living plants were collected and grown in the Botanical garden “Jevremovac“ in Belgrade. For morphometric analysis plant material of ten to fifteen individuals (453 leaves and 341 stems with spikes) was collected from 21 populations. These analyses were performed on dissected plant organs preserved in 50% ethanol (leaves) or in 1 : 1 glycerol : ethanol solution (stems with spikes). Voucher specimens are deposited at BEOU. Voucher numbers and collecting details are given in Appendix 1.
Chromosome counts. — Chromosome numbers were de-termined for five individuals per population (20 populations in total). Root tips obtained from plants in pots were pre-treated with 0.002 M 8-hydroxyquinoline for 24 h at 4°C. After fixation in cold Carnoy fixative (ethanol/acetic acid 3 : 1) for at least 24–48 h, hydrolysis was performed in 1N HCl for 14 min at 60°C. Staining was performed in Schiff’s reagent as described by Feulgen & Rossenbeck (1924), followed by squashing in a drop of acetic carmine. After freezing at −80°C, cover slips were removed, preparations were dried at least 24 h and then mounted in Euparal. Chromosome plates were observed under a Leica DMLS light microscope (Leica Microsystems, Wetzlar, Germany) and photographs were taken with a Leica DCF 295 camera (Leica Microsystems).
Estimation of relative and absolute genome size. — Flow cytometry (FCM) of 4′,6-diamidino-2-phenylindole (DAPI)-stained nuclei was used to estimate the relative DNA content of five to ten silica gel-dried samples per population following the procedure described in Suda & Trávníček (2006a). After incu-bation for 5 min at room temperature, the relative fluorescence intensity of 3000 particles was recorded using a Partec PA II flow cytometer (Partec, Münster, Germany) equipped with an

460
TAXON 62 (3) • June 2013: 458–472Kuzmanović & al. • Differentiation of Sesleria rigida sensu Fl. Eur.
460 Version of Record (identical to print version).
HBO mercury arc lamp. After initial trials, up to three samples were pooled. If the coefficient of variation (CV) of the G0/G1 peak of a sample exceeded the 5% threshold, the analysis was discarded and the sample re-measured. Pisum sativum ‘Kleine Rheinländerin’ (2C = 8.84 pg, Greilhuber & Ebert, 1994) was selected as primary reference standard (Doležel & al., 1998). To extrapolate the ploidy level of all individuals, the relative genome size was correlated with chromosome numbers de-termined for selected individuals from several populations, representative for the whole genome size range.
FCM of propidium iodide (PI)-stained nuclei was used to estimate the absolute genome size of two individuals each from six populations (Appendix 1). Fresh green leaf tissue (ca.
1 cm²) was processed following the procedure described by Kolář & al. (2012). After incubation for 5 min at room tem-perature, the relative fluorescence intensity of 5000 particles was recorded using a Partec PA II flow cytometer equipped with an HBO mercury arc lamp. Vicia faba ‘Inovec’ (2C = 26.90 pg) served as an internal standard as the peaks of Pisum sativum ‘Kleine Rheinländerin’ partly overlapped with those of tetraploid Sesleria samples.
AFLP analysis. — Total genomic DNA was extracted from ca. 10 mg of dried tissue with the DNeasy 96 plant kit (Qiagen, Hilden, Germany) following the manufacturer’s protocol. AFLP fingerprinting was performed for 45 populations with usually five individuals per population. The AFLP procedure followed
Fig. 1. Sampled populations of Sesleria rigida s.l. and the dis-tantly related S. taygetea used as outgroup in the neighbour-joining analysis presented in Fig. 2. Octoploid populations of “rigida” are marked with a white margin; all other populations exclusively contained tetraploid accessions. Population numbers correspond to Appendix 1, the taxonomic assignment follows Deyl (1946, 1980).
461
Kuzmanović & al. • Differentiation of Sesleria rigida sensu Fl. Eur.TAXON 62 (3) • June 2013: 458–472
461Version of Record (identical to print version).
Vos & al. (1995) with the modifica-tions described in Schönswetter & al. (2009). In addition, 0.25 U of polymer-ase were used in the preselective and selective amplifications (0.4 U for the NED-labelled primer combination). Fifteen selective primer combinations have been initially screened. The three final selective primer combinations for the selective PCR (fluorescent dye in brackets) were EcoRI (6-FAM)-ACT / MseI-CAC, EcoRI (VIC)-ACG / MseI-CAC, and EcoRI (NED)-ACC / MseI-CAC. Purification and visualisation of PCR products were done as described in Rebernig & al. (2010). Twenty-eight samples were used as replicates be-tween PCR batches to test the repro-ducibility of AFLP fingerprinting.
Electropherograms were analysed with Peak Scanner version 1.0 (Applied Biosystems, Foster City, California, U.S.A.) using default peak detection parameters except light peak smooth-ing. The minimum fluorescent thresh-old was set to 50 relative fluorescence units. Automated binning and scoring was performed using RawGeno v.2.0 (Arrigo & al., 2009), a package for the software R (R Development Core Team, 2010), with the following settings: scor-ing range = 50–500 bp, minimum in-tensity = 50 relative fluorescence units (rfu), minimum bin width = 1 bp, and maximum bin width = 1.5 bp. Fragments with a reproducibility lower than 80% based on sample-replicate comparisons were eliminated. The error rate (Bonin & al., 2004) was calculated as the ra-tio of mismatches (scoring of 0 vs. 1) over phenotypic comparisons in AFLP profiles of replicated individuals. Frag-ments present/absent in only one indi-vidual were removed from the dataset.
A neighbor-joining (NJ) analysis based on the matrix of Nei-Li genetic distances (Nei & Li, 1979) was con-ducted and bootstrapped (1000 pseu-do-replicates) with TREECON v.1.3b (Van de Peer & De Wachter, 1997). The tree was rooted with S. taygetea. A principal coordinate analysis (PCoA) based on a matrix of Jaccard distances among individuals (excluding S. tayge-tea) was calculated using the modules “SimQual”, “Dcenter” and “Eigen” from NTSYS-pc v.2.0 (Rohlf, 1997).
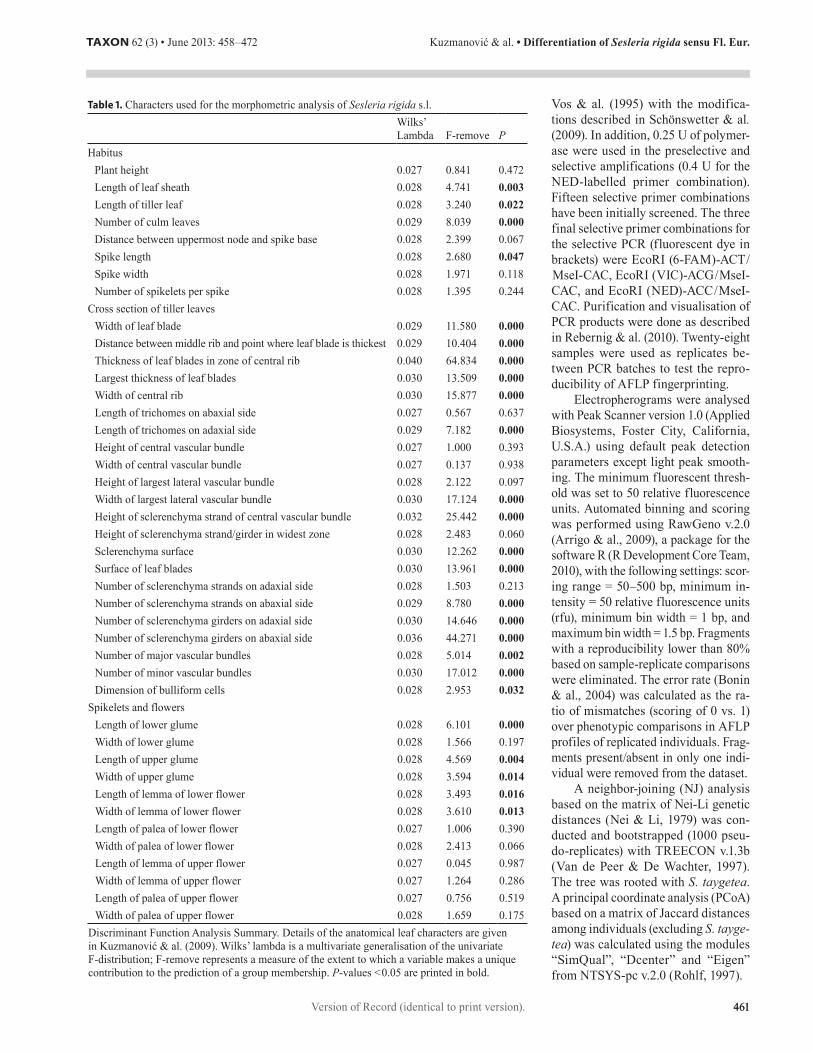
Table 1. Characters used for the morphometric analysis of Sesleria rigida s.l. Wilks’ Lambda F-remove P
Habitus Plant height 0.027 0.841 0.472Length of leaf sheath 0.028 4.741 0.003Length of tiller leaf 0.028 3.240 0.022Number of culm leaves 0.029 8.039 0.000Distance between uppermost node and spike base 0.028 2.399 0.067Spike length 0.028 2.680 0.047Spike width 0.028 1.971 0.118Number of spikelets per spike 0.028 1.395 0.244
Cross section of tiller leaves Width of leaf blade 0.029 11.580 0.000Distance between middle rib and point where leaf blade is thickest 0.029 10.404 0.000Thickness of leaf blades in zone of central rib 0.040 64.834 0.000Largest thickness of leaf blades 0.030 13.509 0.000Width of central rib 0.030 15.877 0.000Length of trichomes on abaxial side 0.027 0.567 0.637Length of trichomes on adaxial side 0.029 7.182 0.000Height of central vascular bundle 0.027 1.000 0.393Width of central vascular bundle 0.027 0.137 0.938Height of largest lateral vascular bundle 0.028 2.122 0.097Width of largest lateral vascular bundle 0.030 17.124 0.000Height of sclerenchyma strand of central vascular bundle 0.032 25.442 0.000Height of sclerenchyma strand/girder in widest zone 0.028 2.483 0.060Sclerenchyma surface 0.030 12.262 0.000Surface of leaf blades 0.030 13.961 0.000Number of sclerenchyma strands on adaxial side 0.028 1.503 0.213Number of sclerenchyma strands on abaxial side 0.029 8.780 0.000Number of sclerenchyma girders on adaxial side 0.030 14.646 0.000Number of sclerenchyma girders on abaxial side 0.036 44.271 0.000Number of major vascular bundles 0.028 5.014 0.002Number of minor vascular bundles 0.030 17.012 0.000Dimension of bulliform cells 0.028 2.953 0.032
Spikelets and flowers Length of lower glume 0.028 6.101 0.000Width of lower glume 0.028 1.566 0.197Length of upper glume 0.028 4.569 0.004Width of upper glume 0.028 3.594 0.014Length of lemma of lower flower 0.028 3.493 0.016Width of lemma of lower flower 0.028 3.610 0.013Length of palea of lower flower 0.027 1.006 0.390Width of palea of lower flower 0.028 2.413 0.066Length of lemma of upper flower 0.027 0.045 0.987Width of lemma of upper flower 0.027 1.264 0.286Length of palea of upper flower 0.027 0.756 0.519Width of palea of upper flower 0.028 1.659 0.175
Discriminant Function Analysis Summary. Details of the anatomical leaf characters are given in Kuzmanović & al. (2009). Wilks’ lambda is a multivariate generalisation of the univariate F-distribution; F-remove represents a measure of the extent to which a variable makes a unique contribution to the prediction of a group membership. P-values < 0.05 are printed in bold.

462
TAXON 62 (3) • June 2013: 458–472Kuzmanović & al. • Differentiation of Sesleria rigida sensu Fl. Eur.
462 Version of Record (identical to print version).
Morphometric analyses. — Analysis of 26 anatomical leaf characters was performed on cross-sections of tiller leaves as described by Kuzmanović & al. (2009). An additional 23 characters of external morphology were analysed following combined and adjusted methods used for Festuca L. (Auquier, 1974; Lakušić, 1999; Foggi & al., 1999, 2006) and Sesleria (Alegro, 2007; Di Pietro, 2007). The measurements were per-formed using Leica Qwin (Leica Microsystem) and Digimizer Image Analysis software (MedCalc Software, Belgium).
Seven invariable characters were excluded. Further charac-ter reduction was envisaged by computing pairwise Spearman correlations and retaining only one out of character pairs with absolute values of correlation coefficients exceeding 0.9, but no
correlations exceeding this threshold were found. The hypoth-esis of morphological separation of four groups of populations recognised by AFLPs was tested using canonical discriminant analysis (CDA) conducted on the resulting data matrix compris-ing 42 characters after standardisation to zero mean and unit variance. Characters included in the CDA are listed in Table 1. The CDA was performed with 4 a priori defined groups × 453 individuals × 42 characters. Discriminant function analysis (Table 1) was done to estimate the contribution of individ-ual characters to overall discrimination. Canonical scores for each case were calculated with the aim to measure distances between individuals, and a scatterplot of canonical scores (see Fig. 5 below) was made to visualise the relationship between a priori defined groups. Statistical analyses were performed using Statistica v.5.1 (StatSoft, 1996).
results
Chromosome numbers and genome size. — Chromosome numbers were determined for 100 individuals from 20 popula-tions (Appendix 1). Two ploidy levels were found, tetraploids with 2n = 4x = 28 in fifteen populations and octoploids with 2n = 8x = 56 in five populations (Fig. 1).
Chromosomes were 2.05–5.99 µm long in tetraploids and 1.75–5.35 µm in octoploids. Several chromosomes in both ploidy levels were bearing satellites. Their number is variable, from two to six in tetraploids and up to ten in octoploids. They are usually situated on large metacentric chromosomes and smaller acrocentric or subtelocentric chromosomes (Fig. 2). In several cases chromatic particles were visible that could be chromosome fragments, detached satellites or B-chromosomes.
Flow cytometry analyses mostly yielded high-resolution histograms. The relative genome size (RGS) ranged from 0.62 to 1.35, whereas the absolute genome size was between 8.58 and 16.90 pg. According to RGS all analyzed individuals were clearly separated into tetraploids (86.4% of the samples) with RGS from 0.62 to 0.76, and octoploids (13.6%) with RGS from 1.29 to 1.35 (Appendix 1; Electr. Suppl.: Fig. S1). Neither odd-ploidy level cytotypes nor mixed-ploidy populations were detected.
AFLP data. — A total of 2580 AFLP fragments were scored in 218 individuals from which high-quality, reproducible AFLP-fingerprints were obtained. A total of 171 and 6 bands were found to be present or absent in only one individual, re-spectively, and were excluded from further analyses. The initial error rate (according to Bonin & al., 2004) before the exclusion of unreliable characters was 3%.
The NJ analysis revealed four clusters with high boot-strap support (BS) between 96% and 100% (Fig. 3). The most
Fig. 2. Mitotic chromosomes of Sesleria rigida s.l. A, population S275 (2n = 28); B, population S468 (2n = 56). — Scale bar = 20 μm.
Fig. 3. Neighbour-joining dendrogram of Sesleria rigida s.l. based on a matrix of Nei-Li distances (Nei & Li 1979) among AFLP phenotypes. The tree is rooted with S. taygetea. Octoploid individuals of “rigida” are shown with thicker branches; all other accessions are tetraploid. Numbers 1 to 4 at the main branches correspond to the main clusters identified by the PCoA (Fig. 4). Bootstrap values (based on 1000 replicates) greater than 50% are given for branches with ≥ 3 terminals. The taxonomic assignment following Deyl (1946, 1980) is given in the middle panel, the new taxonomic concept emerging from the present study is given in the right panel. Symbols correspond to Figs. 1 and 4; symbols at the tips of branches are only given when deviating from the majority assignment in the middle panel. Population numbers given at the tips of branches correspond to Appendix 1.
◄

463
Kuzmanović & al. • Differentiation of Sesleria rigida sensu Fl. Eur.TAXON 62 (3) • June 2013: 458–472
463Version of Record (identical to print version).
“rigida”“pancicii”“filifolia”
464
TAXON 62 (3) • June 2013: 458–472Kuzmanović & al. • Differentiation of Sesleria rigida sensu Fl. Eur.
464 Version of Record (identical to print version).
divergent cluster (BS 100%) corresponds to “serbica“ from ser-pentine areas in western Serbia and eastern Bosnia. A further cluster (BS 100%) contains accessions of “achtarovii“ found in the Rhodopi Mts. (Bulgaria and northeastern Greece). Two well-supported (BS 100) sister clusters include “rigida“ and “degenii“ from the Romanian Carpathians (BS 100%) as well as “rigida“, “filifolia“ and “pancicii“ from eastern and north-eastern Serbia, western and northern Bulgaria and Munţii Ba-natului in Romania (BS 96%).
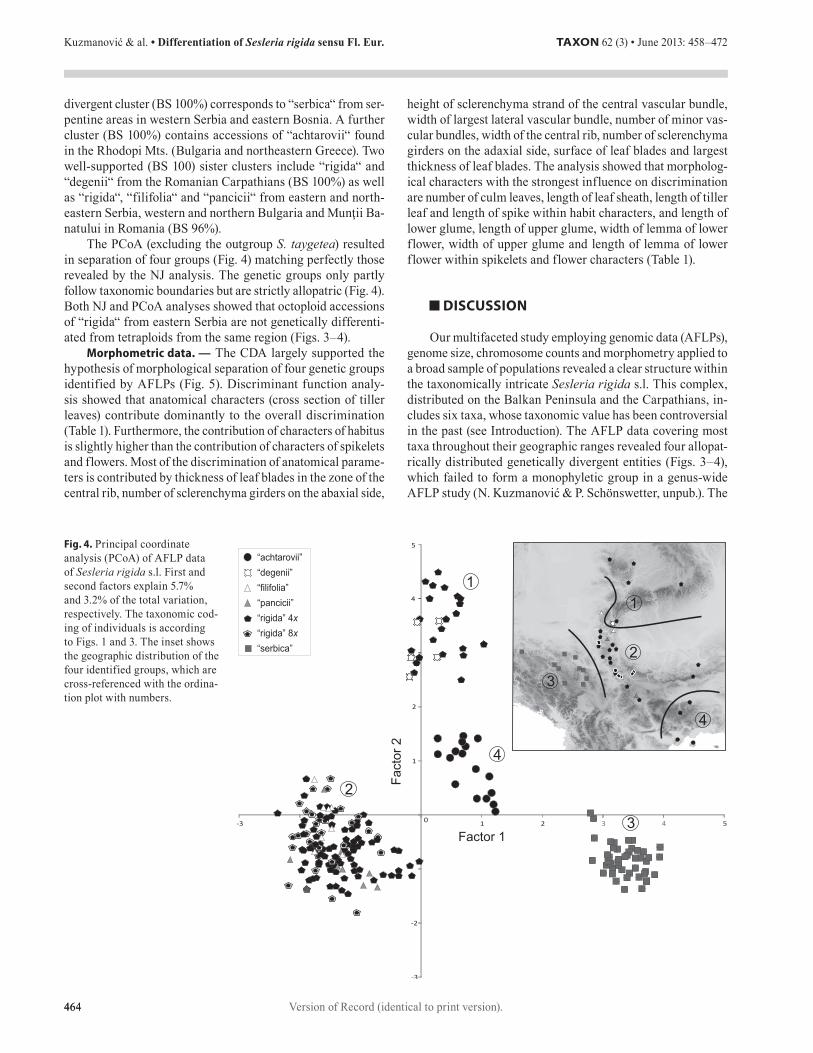
The PCoA (excluding the outgroup S. taygetea) resulted in separation of four groups (Fig. 4) matching perfectly those revealed by the NJ analysis. The genetic groups only partly follow taxonomic boundaries but are strictly allopatric (Fig. 4). Both NJ and PCoA analyses showed that octoploid accessions of “rigida“ from eastern Serbia are not genetically differenti-ated from tetraploids from the same region (Figs. 3–4).
Morphometric data. — The CDA largely supported the hypothesis of morphological separation of four genetic groups identified by AFLPs (Fig. 5). Discriminant function analy-sis showed that anatomical characters (cross section of tiller leaves) contribute dominantly to the overall discrimination (Table 1). Furthermore, the contribution of characters of habitus is slightly higher than the contribution of characters of spikelets and flowers. Most of the discrimination of anatomical parame-ters is contributed by thickness of leaf blades in the zone of the central rib, number of sclerenchyma girders on the abaxial side,
height of sclerenchyma strand of the central vascular bundle, width of largest lateral vascular bundle, number of minor vas-cular bundles, width of the central rib, number of sclerenchyma girders on the adaxial side, surface of leaf blades and largest thickness of leaf blades. The analysis showed that morpholog-ical characters with the strongest influence on discrimination are number of culm leaves, length of leaf sheath, length of tiller leaf and length of spike within habit characters, and length of lower glume, length of upper glume, width of lemma of lower flower, width of upper glume and length of lemma of lower flower within spikelets and flower characters (Table 1).
dIscussIon
Our multifaceted study employing genomic data (AFLPs), genome size, chromosome counts and morphometry applied to a broad sample of populations revealed a clear structure within the taxonomically intricate Sesleria rigida s.l. This complex, distributed on the Balkan Peninsula and the Carpathians, in-cludes six taxa, whose taxonomic value has been controversial in the past (see Introduction). The AFLP data covering most taxa throughout their geographic ranges revealed four allopat-rically distributed genetically divergent entities (Figs. 3–4), which failed to form a monophyletic group in a genus-wide AFLP study (N. Kuzmanović & P. Schönswetter, unpub.). The
4
.00
1
2
5
1 2 3 4 5
-2
-3
-3 0
Factor 1
Fact
or 2
2
4
4
2
1
3
3
1“rigida” 4x“rigida” 8x
“degenii”“achtarovii”
“serbica”
“pancicii”“filifolia”
Fig. 4. Principal coordinate analysis (PCoA) of AFLP data of Sesleria rigida s.l. First and second factors explain 5.7% and 3.2% of the total variation, respectively. The taxonomic cod-ing of individuals is according to Figs. 1 and 3. The inset shows the geographic distribution of the four identified groups, which are cross-referenced with the ordina-tion plot with numbers.

465
Kuzmanović & al. • Differentiation of Sesleria rigida sensu Fl. Eur.TAXON 62 (3) • June 2013: 458–472
465Version of Record (identical to print version).
first (cluster 1, Fig. 3; PCoA group 1, Fig. 4) includes popula-tions of S. rigida (“rigida” and “degenii”) from the Romanian Carpathians, the second (cluster 2; PCoA group 2) comprises populations of S. filifolia (“filifolia” and “pancicii”) from the Munţii Banatului in Romania and the Balkan Mts. in eastern and northeastern Serbia and western and northern Bulgaria, the third (cluster 3; PCoA group 3) consists of populations of S. serbica from serpentine areas in eastern Bosnia and western
Serbia and the last (cluster 4; PCoA group 4) contains popu-lations of S. achtarovii from the Rhodopi Mts. in Bulgaria, northeastern Greece and the island Thassos. Similar results were obtained by Comanescu (2011), albeit based on a less comprehensive sample, and with the main aim of resolving the taxonomic status of S. filifolia in Romania.
The canonical discriminant analysis of the morphometric dataset (Fig. 5) showed that the genetically defined groups
Fig. 5. Canonical discriminant analysis (CDA) of Sesleria rigida s.l. based on 453 individuals and 42 morpho-anatomical charac-ters. The four a priori defined groups are the genetic groups identified in Figs. 3 and 4. In (A) the labelling is in accordance with the genetic groups, whereas in (B) the labelling follows Deyl (1946, 1980). The informal enti-ties “degenii” and “pancicii” were not included as no material was available for morphometric study.
Root 1
2 tooR
-6 -4 -2 0 2 4 6-6
-4
-2
0
2
4
6
“rigida” 4x“rigida” 8x
“achtarovii”
“serbica”
“filifolia”
B
Root 1
2 tooR
-6 -4 -2 0 2 4 6-6
-4
-2
0
2
4
6
A Sesleria rigidaSesleria serbica
Sesleria filifoliaSesleria achtarovii

466
TAXON 62 (3) • June 2013: 458–472Kuzmanović & al. • Differentiation of Sesleria rigida sensu Fl. Eur.
466 Version of Record (identical to print version).
are also morphologically distinct. Four taxa can thus be rec-ognised: S. achtarovii, S. filifolia, S. rigida and S. serbica, which are presented below in the Taxonomic Treatment. As the populations originally assigned to “degenii” and “pancicii” were neither genetically nor morphologically differentiated, we synonymise S. rigida var. degenii and S. rigida var. pancicii with S. rigida s.str. and S. filifolia, respectively. All four species in the revised circumscription are allopatrically distributed and their distribution ranges are placed within different phytoge-ographic regions and provinces (Meusel & Jäger, 1992; Jäger & Welk 2003). Whereas S. rigida belongs to the Carpathian, the other three species belong to the different provinces of the sub-Mediterranean region. Sesleria filifolia is restricted to the Moesian (i.e., Balkan), S. serbica to the Illyrian and S. achtarovii to the Macedonian-Thracian province, with one isolated population on the island of Thassos in the Aegean province of the Macaronesian-Mediterranean region.
The geographically isolated (Fig. 1) S. achtarovii and S. serbica are also genetically most distant (Fig. 3), whereas S. filifolia and S. rigida with distribution areas separated by minimally 20 km are also genetically closer and form a highly supported clade. Their distribution areas come into close con-tact in the Iron Gates region along the Danube in Serbia and Romania. This area has traditionally been considered a likely migration corridor between the Carpathians and the Dinaric Mts., and this is supported also by phylogeographic studies (e.g., Frajman & Oxelman, 2007; Ronikier, 2011). Our results support this hypothesis, but it is not clear whether both taxa came here into secondary contact after extending their distri-bution from two isolated refugia, or if the Carpathian S. rigida originated from the Balkan S. filifolia in the course of a north-eastern range expansion. The Danube river valley was obvi-ously no impassable barrier for the constituents of S. rigida s.l. as two populations (S317, S372) of S. filifolia were sampled north of the Danube in Romania. The distribution of S. filifolia, extending from southeastern and eastern Serbia to the central Stara planina in Bulgaria but missing from the adjacent Rho-dopi Mts. (Fig. 1) is seen also in the regional distributions of several other plant species such as Aconitum burnatii Gáyer, Allium victorialis L., Anthemis carpatica Waldst. & Kit. ex Willd., Campanula transsilvanica Schur ex Andrae, Carex brevicollis DC., Centaurea napulifera Rochel, Dryas octo-petala L.and Jacobaea abrotanifolia (L.) Moench (Senecio abrotanifolius L.) (Assyov & Petrova, 2006). On the other hand, the Rhodopi Mts. share some taxa with adjacent mountains in Greece (e.g., Haberlea rhodopensis Friv., Petkovia orphanidea (Boiss.) Stef., Saxifraga ferdinandi-coburgi Kellerer & Sünd., Saxifraga stribrnyi (Velen.) Podp., Scabiosa rhodopensis Stoj. & Stef., Polygala rhodopea Janch.), a distribution pattern seen also in S. achtarovii.
Ecologically, S. serbica shows the highest specificity. It is an obligate serpentinophyte that inhabits submontane and mon-tane rocky grasslands (Festuco-Brometea) from the thermophil-ous oak to the cryophilous spruce belt and subalpine pastures (Festuco-Seslerietea). At the same time, this species dominates the understory of specific pine forests (Seslerio-Pinetum nigrae and Seslerio-Pinetum sylvestris community-type) and is an
important component of temperate heaths (Erico-Seslerietum community-type; Lakušić & al., 2005). The other three spe-cies are strictly calcicole and grow in rock crevices (Asple-nietea trichomanis), rocky grasslands (Festuco-Brometea) and subalpine and alpine pastures (Festuco-Seslerietea or Daphno-Festucetea) between 100 and 2140 m a.s.l., and also in beech forests (Seslerio rigidae-Fagetum of the class Quer-co-Fagetea) and Scots pine forests (Seslerio rigidae-Pinetum sylvestris of the class Erico-Pinetea) (Lakušić & al., 2005; Kar-akiev & Tzonev, 2011; Comanescu, 2011).
Previously, only tetraploids were known in S. rigida s.l. (Ujhelyi, 1959, 1960; Deyl, 1980; Petrova, 2000). Our genome size measurements and chromosome counts (Appendix 1; Figs. 2–3) have revealed some octoploid (2n = 56) populations of S. fili folia, mostly from southeastern and eastern Serbia, whereas tetraploids are evenly distributed throughout the studied area in all analysed taxa. The octoploid populations of S. filifolia are genetically indistinguishable from neighbour-ing tetraploid populations. This is supported by the neighbour joining analysis (Fig. 3) as well as by Bayesian cluster analyses conducted with STUCTURE v.2.3 (Pritchard & al., 2000) under different parameterisations (with and without admixture, allele frequencies correlated or not) suggesting a single unstructured gene pool as most likely solution (Peter Schönswetter, unpub.). The lack of genetic structure within S. filifolia strongly suggests a recent autopolyploid origin of the octoploid cytotype. Based on the disjunct distribution of octoploids we hypothesize that they have independently originated at least twice. Autopoly-ploidy is much more common in plants then traditionally as-sumed. Initially considered an evolutionary dead-end (Clausen & al., 1945; Stebbins, 1971), it was later shown that genome multiplication per se may represent an evolutionary advantage, e.g., by enforcing successful range expansion and ecological radiation as demonstrated in different natural autopolyploids (Manton, 1937; Soltis & al., 2007; Parisod & al., 2010).
No cytotype mixtures were found even when expanding our ploidy level screening to ten individuals per population in contact zones of the two cytotypes (Appendix 1). In the same line, geographically close octoploid and tetraploid pop-ulations are ecologically and altitudinally differentiated. For instance, although the horizontal distance between tetraploids from the Gornjak gorge (population S234) and octoploids from Mt. Vukan (population S466) is less than 2 km, tetraploids occur exclusively in rock crevices (Asplenietea trichomanes) in a gorge at ca. 200 m a.s.l., whereas octoploids inhabit rocky grasslands (Festuco-Brometea) on wind-exposed ridges at ca. 700 m a.s.l. Tolerance of plants with higher ploidy level to hab-itats with stronger competition (Hülber & al., 2009) is also re-flected in their overall size. Already Deyl (1946) hypothesised that plants from the eastern Serbian Mt. Basara that are larger in all parts have a higher ploidy level. Herbarium vouchers sampled in populations from which cytological information is available confirmed that octoploids are generally more robust than tetraploids. Particularly in leaves of octoploid individuals all size characters are larger than in tetraploids (Kuzmanović & al., 2009). The absence of any hexaploids, despite screen-ing a large number of individuals, suggests that octoploids are

467
Kuzmanović & al. • Differentiation of Sesleria rigida sensu Fl. Eur.TAXON 62 (3) • June 2013: 458–472
467Version of Record (identical to print version).
reproductively isolated from tetraploids, even when they occur in geographic proximity. The observed ecological differen-tiation may play a role in the apparent isolation (e.g., Lysak & Doležel, 1998; Petrova, 2000).
Similar to other grass genera (e.g., Festuca, Helictotrichon Besser, Puccinellia Parl., Stipa L.), our results clearly indicate that leaf morphology and especially anatomy play a primary role in morphological discrimination of the constituents of S. rigida s.l. On the basis of leaf shape (convolute vs. flat), presence of indumentum on the adaxial side of leaves (hairy vs. glabrous), type of the subepidermal sclerenchyma bands (continuous vs. interrupted), number of major and minor vas-cular bundles, as well as the number of sclerenchyma girders, it is relatively straightforward to distinguish the four species. Additionally, characters of overall habit, such as shape of tufts (stoloniferous vs. compact) and spikes (lax and interrupted vs. dense and compact), further facilitate their identification.
taxonoMIc treatMent
Sesleria rigida s.l. comprises densely caespitose perennials without reticulate basal leaf sheaths. Leaves are 0.6–2.1 mm wide, plicate, sometimes pruinose above, and have fewer than 13 major vascular bundles. The uppermost stem leaf is 0.2–2.5 cm long. The spike is cylindrical, 3–5 times longer than wide, bluish or whitish, without prominent white bracts subtending the inflorescence. Spikelets are laterally compressed and bear 2–3 florets. Glumes are unequal and membranous with a single vein. The lemma is sparsely pubescent to glabrous between the veins, membranous, five-veined, with usually five aristulate teeth at the apex, the middle tooth being the longest. The palea is as long as or shorter than the lemma and is two-veined.
Based on the cytological, genetic and morphometric results presented here, we recognise four previously described species within the studied group, i.e., S. achtarovii, S. filifolia, S. rigida and S. serbica.
Key to the species of Sesleria rigida s.l.
In the morphological descriptions, value ranges correspond to the mean ± standard deviation, with the minimal and maxi-mal values in brackets. Morphological descriptions are reduced and include only diagnostic morphological characters.
1. Tuft stoloniferous, spike elongated and lax, 4–5 times longer than wide, growing on serpentine ...... S. serbica
1. Tuft compact, spike dense, 3 times longer than wide, grow-ing on carbonate ............................................... 2
2. Leaf transverse section with continuous stratum of sube-pidermal sclerenchyma ............................. S. rigida
2. Leaf transverse section without continuous stratum of sub-epidermal sclerenchyma ..................................... 3
3. Leaves convolute, rarely flat, densely hairy on adaxial side .................................................. S. filifolia
3. Leaves flat, rarely convolute, sparsely hairy on adaxial side ............................................... S. achtarovii
Sesleria achtarovii Deyl in Opera. Bot. Čech. 3: 193. 1946 ≡ S. rigida subsp. achtarovii (Deyl) Deyl in Bot. J. Linn. Soc. 76: 364. 1978 – Lectotype (designated here): [BULGARIA] in saxosis calcareis mt. Rhodope centralis, Kuru-dere ad Stanimaka, 23 Mar 1914, I. Mrkvička s.n. (SOM no. 100252!).Note. – We examined several specimens of original mate-
rial (SOM no. 100252!, SOM no. 4441!, PRC no. PRC519968!, W no. 1892-0009528!), and decided to select the sheet from Kuru-Dere (SOM no. 100252) as lectotype. Of all the speci-mens cited in the protologue it is the most representative, and has several well developed plants.
Description. – Caespitose perennial. Culm (12.7)16.5–33.0(42.3) cm tall. Spike (13)15–20(25) × (4)5–7(8) mm, whit-ish, with (9)12–18(28) spikelets; spikelets on short pedicels, with 2–3 flowers. Tiller leaves (6)11–26(37) cm long, flat or convolute, sparsely hairy on adaxial side; transverse sections of tiller leaves (0.87)0.97–1.31(1.63) × (0.15)0.18–0.23(0.27) mm, with 5–6(7) major vascular bundles and (6)7–10(13) minor vascular bundles, number of sclerenchyma girders varies from (6)8–12(14) on adaxial side of leaves and (4)6–9(12) on abaxial side of leaves, subepidermal sclerenchyma discontinuous.
Chromosome number. – 2n = 4x = 28Ecology. – Rock crevices (Asplenietea trichomanis) and
subalpine and alpine pastures (Daphno-Festucetea) on car-bonate bedrock (limestone, marble); 450–2000 m a.s.l.
Distribution. – Rhodope Mts. in southern Bulgaria and northeastern Greece.
Sesleria filifolia Hoppe in Flora 17: 384. 1834 – Lectotype (designated here): [ROMANIA], in rupibus calcareis ad Krassova [Caraşova], Apr 1831, J. Heuffel s.n. (BP no. 19625!, only the two right-hand plants).
= Sesleria rigida var. pancicii Deyl in Opera. Bot. Čech. 3: 191. 1946 – Lectotype (designated here): SERBIA, auf Kalkfelsen Debeli lug, Majdan pek, Apr 1879, J. Pavlović s.n. (PR no. 161815!).Note on lectotypification of S. filifolia. – On the herbarium
sheet four plants from two different collections are mounted. Only the two right-hand plants clearly from Krassova are se-lected as the lectotype.
Note on lectotypification of S. rigida var. pancicii. – In PR, we found four specimens from Serbia (PR no. 161815!, no. 161821!) and Bulgaria (PR no. 161816!, no. 161817!) that were labelled as Sesleria rigida Heuff. var. pancicii by Deyl. We have excluded the Bulgarian specimens as potential types, as (1) the plant from Zemen collected by Achtaroff (PR no. 161817!) is not cited in “Specimina ut sequuntur vidi” (Deyl 1946: 187–188), which implies that Deyl did not have this material at hand at the time when he described this variety, and (2) the plant from Persenka Balkan in Bulgaria collected by Podpèra (PR no. 161816!) belongs to S. achtarovii (Deyl 1946: 193). Since the specimen from Debeli lug collected by Pavlović (PR no. 161815!) is more representative than the specimen from Niška Banja col-lected by Ilić (PR no. 161821!), we select the former as lectotype.
Description. – Caespitose perennial. Culm (13.1)24.7–42.6(65.3) cm tall. Spike (10)16–23(33) mm × (4)5–8(10) mm,

468
TAXON 62 (3) • June 2013: 458–472Kuzmanović & al. • Differentiation of Sesleria rigida sensu Fl. Eur.
468 Version of Record (identical to print version).
bluish, with (5)11–19(27) spikelets; spikelets on short pedicels, with 2–3 flowers. Tiller leaves (1.3)12.4–25.4(37.5) cm long, convolute, rarely flat, densely hairy on adaxial side, rarely pruinose above; transverse sections of tiller leaf (0.59)0.76–1.13(1.67) mm × (0.15)0.19–0.25(0.33) mm, with (3)4–6(7) major vascular bundles and (3)5–8(11) minor vascular bundles, num-ber of sclerenchyma girders varies from (5)6–9(12) on adaxial side of leaf and (4)6–8(10) on abaxial side of leaf, subepidermal sclerenchyma not continuous.
Chromosome number. – 2n = 4x = 28 and 2n = 8x = 56.Ecology. – Rock crevices (Asplenietea trichomanis), rocky
grasslands (Festuco-Brometea) and subalpine and alpine pastures (Festuco-Seslerietea), on carbonate bedrock; 90–1830 m a.s.l.
Distribution. – Munţii Banatului in Romania and Balkan Mts. in eastern Serbia and western and northern Bulgaria.
Sesleria rigida Heuff. ex Rchb., Fl. Germ. Excurs.: 1403. 1831 – Lectotype (designated here): [ROMANIA], in cacumine montis Domuglett (“3000 ped”), ad Thermas Herculis, Apr [without indication of year], J. Heuffel s.n. (W no. 1889-0204591!).
= Sesleria rigida var. degenii Deyl in Opera. Bot. Čech. 3: 190. 1946 – Lectotype (designated here): [ROMANIA], in saxosis ad cacuminen montis Domugled supra thermas Herkulis, 7 May 1894, A. de Degen s.n. (BP no. 19851!).Note. – The specimen selected as the lectotype of Ses-
leria rigida var. degenii was seen by M. Deyl, and collected prior to the publication of the protologue. Thus we consider it likely represents a part of the original material. Although Deyl’s undated annotation label lists only the name S. rigida, we believe that it was used by him to describe the new variety and simply not reannotated because it was located outside of his home institution (PR). This is supported by the fact that Deyl appears to have only added annotations to specimens at PR after describing new taxa in his monograph.
Description. – Caespitose perennial. Culm (6.9)17.3–35.6(45.8) cm tall. Spike (11)14–23(30) mm × (4)6–8(10) mm, whitish, with (9)13–21(32) spikelets; spikelets on short pedi-cels, with 2–3 flowers. Tiller leaves (2.0)9.3–28.2(39.7) cm long, flat or convolute, pruinose above; transverse sections of tiller leaves (0.72)1.03–1.56(2.09) × (0.15)0.20–0.28(0.34) mm, with (3)5–8(11) major vascular bundles and (4)5–11(21) minor vascular bundles, sclerenchyma girders on adaxial side of leaf (4)8–15(22), on abaxial side (4)8–15(20), with continuous subepidermal sclerenchyma band.
Chromosome number. – 2n = 4x = 28.Ecology. – Rock crevices (Asplenietea trichomanis), rocky
grasslands (Festuco-Brometea), pastures (Festuco-Seslerietea) and forests (Querco-Fagetea, Erico-Pinetea), on carbonate bed-rock; 160–2140 m a.s.l.
Distribution. – Carpathians in Romania.
Sesleria serbica (Adamović) Ujhelyi in Feddes Repert. Spec. Nov. Regni Veg. 62: 64. 1959 ≡ S. rigida var. serbica Ad-amović in Allg. Bot. Z. Syst. 2: 120. 1896 – Lectotype (designated here): [SERBIA], in saxosis ad Gornji Milan-ovac, Apr 1893, L. Adamović s.n. (PRC no. PRC 451931!).
Note. – Ujhelyi (1959), when raising S. rigida var. ser-bica to the species level, incorrectly cited a specimen from BP as the “holotype”: “Bosnien: Gostovic-Gebiet auf Serpentin. Leg. W. Ludwig, Mai 1955” (BP no. 0734231!). However, as he clearly based Sesleria serbica on Adamović’s S. rigida var. serbica, the two names must be taken as homotypic. We ex-amined several specimens of original material (e.g., specimens collected by L. Adamović in the vicinity of Gornji Milanovac in May 1893 [BP no. 593596!, WU no. 0042093!, GZU no. 259577!] and in April 1896 [W no. 1897-0006747!]), and se-lected the specimen from PRC as lectotype. The lectotype is quite representative of the species, and it is very well preserved with several plants on the sheet.
Description. – Stoloniferous perennial. Culm (15.7)23.8–42.6(55.9) cm tall. Spike elongated and lax, (16)19–30(44) mm × (4)5–7(8) mm, bluish; with (7)11–16(20) spikelets. Spikelets on prominent pedicels, with 2–3 flowers. Tiller leaves (4.5)13.4–33.4(47.2) cm long, convolute, hairy on adaxial and abaxial side; transverse sections of tiller leaves (0.60)0.83–1.09(1.42) × (0.18)0.21–0.26(0.29) mm, with (3)4–6(7) major vascular bundles and (3)5–7(12) minor vascular bundles, number of sclerenchyma girders on adaxial side of leaves (6)8–11(14) and on abaxial side of leaves (5)8–11(14), with tendency to form continuous subepidermal sclerenchyma.
Chromosome number. – 2n = 4x = 28.Ecology. – Rocky grasslands (Festuco-Brometea), forests
(Erico-Pinetea) and subalpine pastures (Festuco-Seslerietea), exclusively on serpentine bedrock; 250–1900 m a.s.l.
Distribution. – Dinaric Mts. of eastern Bosnia and western Serbia.
acKnowledgeMents
This work was financed by the European Commission in the SEE-ERA.NET PLUS framework (project “Evolution, biodiversity and conservation of indigenous plant species of the Balkan Peninsula” to P.S.) and the Ministry of Education and Science of the Republic of Serbia (grant 173030 to D.L.). The Austrian Agency for Interna-tional Cooperation in Education & Research (OeAD; financed by the Austrian Federal Ministry of Science and Research) granted a one-month visit at the Institute of Botany of the University of Innsbruck (ICM-2012-02562) to N.K. Support from the Royal Botanic Gardens Kew and the European SYNTHESYS programme (Project No. GB-TAF-334) granted to P.C. is also acknowledged. We thank M. Magauer, D. Pirkebner and M. Winkler for conducting the laboratory work in an excellent way. We are grateful to P.D. Schlorhaufer and his col-leagues from the Botanical Garden of the University of Innsbruck for successfully cultivating our living collection of Sesleria. We gratefully acknowledge the herbarium curators L. Pignotti (W), W. Till (WU), Z. Barina (BP), L. Somlyay (BP), O. Šida (PRC), J. Štepanek (PR) and S. Bancheva (SOM) for help with searching for type specimens of the investigated taxa. Our colleagues S. Vukojičić, M. Niketić, G. Tomović, M. Plećaš (Belgrade), V. Ranđelović, B. Zlatković, I. Fusi-janović (Niš), D. Uzunov, Ch. Gusev (Sofia) and S. Redžić (Sarajevo) helped with field work and collecting of the plant samples. We are grateful to B. Foggi (Florence) for help with nomenclatural problems.

469
Kuzmanović & al. • Differentiation of Sesleria rigida sensu Fl. Eur.TAXON 62 (3) • June 2013: 458–472
469Version of Record (identical to print version).
Alegro, A. 2007. Sistematika i rasprostranjenost kompleksa Sesleria juncifolia na području Dinarida [Systematics and distribution of Sesleria juncifolia complex in the Dinaric area]. Dissertation, Uni-versity of Zagreb, Croatia. [in Croatian with English summary]
Arrigo, N., Tuszynski, J.W., Ehrich, D., Gerdes, T. & Alvarez, N. 2009. Evaluating the impact of scoring parameters on the structure of intra-specific genetic variation using RawGeno, and R package for automating AFLP scoring. B. M. C. Bioinf. 10: 33.
http://dx.doi.org/10.1186/1471-2105-10-33Assyov, B. & Petrova, A. (eds.) 2006. Conspectus of the Bulgarian
vascular flora: Distribution maps and floristic elements, 3rd, rev. and enl. ed. Sofia: Bulgarian Biodiversity Foundation.
Auquier, P. 1974. Biosystématique, taxonomie et nomenclature du groupe de Festuca ovina L. s.l. (Poaceae) en Belgique et dans quelques régions voisines. Dissertation, University of Liège, Bel-gium.
Bardy, E.K., Albach, C.D., Schneeweiss, M.G., Fischer, A.M. & Schönswetter, P. 2010. Disentangling phylogeography, polyploid evolution and taxonomy of a woodland herb (Veronica chamae-drys group, Plantaginaceae s.l.) in southeastern Europe. Molec. Phylogen. Evol. 57: 771–786.
http://dx.doi.org/10.1016/j.ympev.2010.06.025Bonin, A., Bellemain, E., Eidesen, P.B., Pompanon, F., Brochmann,
C. & Taberlet, P. 2004. How to track and assess genotyping errors in population genetic studies. Molec. Ecol. 13: 3261–3273.
http://dx.doi.org/10.1111/j.1365-294X.2004.02346.xCiocarlan, V. 2009. Flora ilustrată a României: Pteridophyta et Sper-
matophyta. Bucharest: Edit. Ceres.Clausen, J., Keck, D.D. & Hiesey, W.M. 1945. Plant evolution through
amphiploidy and autoploidy, with examples from the Madiinae. Washington, D.C.: Carnegie Institution of Washington.
Comai, L. 2005. The advantages and disadvantages of being polyploid. Nat. Rev. Genet. 6: 836–846.
http://dx.doi.org/10.1038/nrg1711Comanescu, P. 2011. Genul Sesleria în Flora României: Taxonomie,
morfo-anatomie, ecologie, biosistematică [Genus Sesleria in Flora of Romania: Taxonomy, morpho-anatomy, biosystematics]. Dis-sertation, University of Bucharest, Romania. [in Romanian]
Csergö, A.M., Schönswetter, P., Gyöngyvér, M., Deák, T., Boşcaiu, N. & Höhn, M. 2009. Genetic structure of peripheral, island-like populations: A case study of Saponaria bellidifolia Sm. (Caryo-phyllaceae) from the southeastern Carpathians. Pl. Syst. Evol. 278: 33–41. http://dx.doi.org/10.1007/s00606-008-0129-5
Davis, S.D., Heywood, V.H. & Hamilton, A.C. 1994. Centres of plant diversity, vol. 1, Europe, Africa, southwest Asia and the Middle East. Gland: International Union for the Conservation of Nature and Natural Resources.
De Bodt, S., Maere, S. & Van de Peer, Y. 2005. Genome duplication and the origin of angiosperms. Trends Ecol. Evol. 20: 591–597.
http://dx.doi.org/10.1016/j.tree.2005.07.008Delipavlov, D., Cheshmedzhiev, I., Popova, M., Tersiyski, D. &
Kovachev, I. 2003. Key to the Bulgarian plants. Plovdiv: Agrar-ian University Press. [in Bulgarian]
Deyl, M. 1946. Study of the genus Sesleria. Opera Bot. Čech. 3: 1–246.Deyl, M. 1980. Sesleria Scop. Pp. 173–177 in: Tutin, T.G., Heywood,
V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M. & Webb, D.A. (eds.), Flora Europea, vol. 5, Alismataceae to Orchid-aceae. Cambridge: Cambridge University Press.
Finally, the two anonymous reviewers, the subject editor P. Comes, the editor-in-chief J. Kadereit and the nomenclature editors Gerry Moore and James C. Lendemer provided very helpful suggestions to improve previous versions of the manuscript.
lIterature cIted
Di Pietro, R. 2007. Taxonomic features of Sesleria calabrica (Poaceae), a neglected species from southern Italy. Folia Geobot. 42: 289–313.
http://dx.doi.org/10.1007/BF02806468Diklić, N. & Nikolić, V. 1986. Sesleria rigida Heuff. var. serbica
Adamović. P. 248 in: Sarić, M.R. & Diklić, N. (eds.), Flora SR Srbije, vol. 10. Belgrade: Srpska akademija nauka i umetnosti.
Doležel, J., Greilhuber, J., Lucretti, S., Meister, M., Lysák, M.A., Nardi, L. & Obermayer, R. 1998. Plant genome size estimation by flow cytometry: Inter-laboratory comparison. Ann. Bot. (Oxford) 82(Suppl. A): 17–26.
http://dx.doi.org/10.1006/anbo.1998.0730Feulgen, R. & Rossenbeck, H. 1924. Mikroskopisch-chemischer Nach-
weis einer Nukleinsäure vom Typus der Thymonukleinsäure und die darauf beruhende elektive Färbung von Zellkernen in mi kro-skopischen Präparaten. Hoppe-Seyler’s Z. Physiol. Chem. 135: 203–248. http://dx.doi.org/10.1515/bchm2.1924.135.5-6.203
Foggi, B., Rossi, G. & Signorini, M.A. 1999. The Festuca violacea aggregate (Poaceae) in the Alps and Apennines (Central-Southern Europe). Canad. J. Bot. 77: 989–1013.
Foggi, B., Gherardi, M.E., Signorini, M.A., Rossi, G. & Bruschi, P. 2006. Festuca inops De Not. and Festuca gracilior (Hack.) Markgr.-Dannenb. (Poaceae): Are they two different species? Bot. J. Linn. Soc. 151: 239–258.
http://dx.doi.org/10.1111/j.1095-8339.2006.00496.xFrajman, B. & Oxelman, B. 2007. Reticulate phylogenetics and phyto-
geographical structure of Heliosperma (Sileneae, Caryophyllaceae) inferred from chloroplast and nuclear DNA sequences. Molec.Phylogen. Evol. 43: 140–155.
http://dx.doi.org/10.1016/j.ympev.2006.11.003Gergelyi, I. & Beldie, A. 1972. Sesleria. P. 215 in: Savulescu, T. (ed.),
Flora României, vol. 12. Bucharest: Editura Academiei Române.Greilhuber, J. & Ebert, I. 1994. Genome size variation in Pisum sa-
tivum. Genome 37: 646–655. http://dx.doi.org/10.1139/g94-092Gustavsson, L.A. 1991. Sesleria Scop. Pp. 777–783 in: Strid, A. & Kit
Tan (eds.), Mountain flora of Greece, vol. 2. Edinburgh: Edinburgh University Press.
Hülber, K., Sonnleitner, M., Flatscher, R., Berger, A., Dobrovsky, R., Niessner, S., Nigl, T., Schneeweiss, G.M., Kubešová, M., Rauchová, J., Suda, J. & Schönswetter, P. 2009. Ecological seg-regation drives fine scale cytotype distribution of Senecio carnio-licus (Asteraceae) in the Eastern Alps. Preslia 81: 309–319.
Husband, B.C. & Sabara, H.A. 2004. Reproductive isolation between autotetraploids and their diploid progenitors in fireweed Chamer-ion angustifolium (Onagraceae). New Phytol. 161: 703–713.
http://dx.doi.org/10.1046/j.1469-8137.2004.00998.xJäger, E.J. & Welk, E. 2003. Pflanzengeographische Gliederung Euro-
pas. Pp. 79–86 in: Bohn, U., Neuhäusl, R., Gollub, G., Hettwer, C., Neuhäuslová, Z., Raus, T., Schlüter, H. & Weber, H. (eds.), Karte der natürlichen Vegetation Europas = Map of the natural vegetation of Europe. Maßstab / Scale 1 : 2 500 000. Münster: Land-wirtschaftsverlag.
Jiao, Y., Wickett, N.J., Ayyampalayam, S., Chanderbali, A.S., Land-herr, L., Ralph, P.E., Tomsho, L.P., Hu, Y., Liang, H., Soltis, P.S., Soltis, D.E., Clifton, S.W., Schlarbaum, S.E., Schuster, S.C., Ma, H., Leebens-Mack, J. & dePamphilis, C.W. 2011. Ancestral polyploidy in seed plants and angiosperms. Nature 473: 97–100. http://dx.doi.org/10.1038/nature09916
Karakiev, T. & Tzonev, R. 2011. Saxifrago ferdinandi-coburgi-Ses-lerietum actarovii—A new association from the subalpine belt of the Slavianka (Orvilos) Mts. (Bulgaria). Hacquetia 10: 49–71.
http://dx.doi.org/10.2478/v10028-011-0004-8Kolář, F., Štech, M., Trávníček, P., Rauchová, J., Urfus, T., Vít, P.,
Kubešová, M. & Suda, J. 2009. Towards resolving the Knau-tia arvensis agg. (Dipsacaceae) puzzle: Primary and secondary contact zones and ploidy segregation at landscape and microgeo-graphic scales. Ann. Bot. (Oxford) 103: 963–974.
http://dx.doi.org/10.1093/aob/mcp016

470
TAXON 62 (3) • June 2013: 458–472Kuzmanović & al. • Differentiation of Sesleria rigida sensu Fl. Eur.
470 Version of Record (identical to print version).
Kolář, F., Lučanová, M., Těšitel, J., Loureiro, J. & Suda, J. 2012. Glycerol-treated nuclear suspensions—An efficient preservation method for flow cytometric analysis of plant samples. Chromosome Res. 20: 303–315. http://dx.doi.org/10.1007/s10577-012-9277-0
Kron, P., Suda, J. & Husband, B.C. 2007. Applications of flow cy-tometry to evolutionary and population biology. Annual Rev. Ecol. Evol. Syst. 38: 847–876.
http://dx.doi.org/10.1146/annurev.ecolsys.38.091206.095504Kryštufek, B. & Reed, J.M. 2004. Pattern and process in Balkan bio-
diversity—An overview. Pp. 1–8 in: Griffiths, H.I., Kryštufek, B. & Reed, J.M. (eds.), Balkan biodiversity. Dordrecht: Kluwer Academic Publishers.
http://dx.doi.org/10.1007/978-1-4020-2854-0_1Kučera, J., Marhold, K. & Lihová, J. 2010. Cardamine maritima
group (Brassicaceae) in the amphi-Adriatic area: A hotspot of species diversity revealed by DNA sequences and morphological variation. Taxon 59: 148–164.
Kuzmanović, N., Šinžar-Sekulić, J. & Lakušić, D. 2009. Leaf anat-omy of the Sesleria rigida Heuffel ex Reichenb. (Poaceae) in Ser-bia. Bot. Serbica 33: 51–67.
Kuzmanović, N., Šinžar-Sekulić, J. & Lakušić, D. 2012. Ecologically determined variation in leaf anatomical traits of Sesleria rigida Heuffel ex Reichenb. (Poaceae) in Serbia – Multivariate morpho-metric evidence. Folia Geobot. 47: 41–57.
http://dx.doi.org/10.1007/s12224-011-9104-yLakušić, D. 1999. Ekološka i morfološka diferencijacija uskolisnih
vijuka (Festuca L. subgen. Festuca) na prostoru Durmitora [Eco-logical and morphological differentiation of narrow-leaved fescue (Festuca L. subgen. Festuca) in the area of Mt. Durmitor]. Dis-sertation, University of Belgrade, Serbia. [in Serbian with English summary]
Lakušić, D., Blaženčić, J., Ranđelović, V., Butorac, B., Vukojičić, S., Zlatković, B., Jovanović, S. & Šinžar-Sekulić, J. 2005. Fito-cenoze Srbije—Baza podataka [Phytocoenosis of Serbia—Data base]. Institute of Botany and Botanical Garden Jevremovac, Fac-ulty of Biology, University of Belgrade. http://habitat.bio.bg.ac.rs/stanista_srbije.htm
Levin, D.A. 1983. Polyploidy and novelty in flowering plants. Amer. Naturalist 122: 1–25. http://dx.doi.org/10.1086/284115
Levin, D.A. 2002. The role of chromosomal change in plant evolution. Oxford: Oxford University Press.
Liber, Z., Kovačić, S., Nikolić, T., Likić, S. & Rusak, G. 2008. Rela-tions between western Balkan endemic Campanula L. (Campanul-aceae) lineages: Evidence from chloroplast DNA. Pl. Biosyst. 142: 40–50. http://dx.doi.org/10.1080/11263500701872283
Lysak, M.A. & Doležel, J. 1998. Estimation of nuclear DNA content in Sesleria (Poaceae). Caryologia 51: 123–132.
http://dx.doi.org/10.1080/00087114.1998.10589127Manton, I. 1937. The problem of Biscutella laevigata L. II. The evi-
dence from meiosis. Ann. Bot. (Oxford) 51: 439–465.Mayrose, I., Zhan, S.H., Rothfels, C.J., Magnuson-Ford, K., Barker,
M.S., Rieseberg, L.H. & Otto, S.P. 2011. Recently formed poly-ploid plants diversify at lower rates. Science 333: 1257.
http://dx.doi.org/10.1126/science.1207205Meusel, H. & Jäger, E.J. 1992. Vergleichende Chorologie der zentra-
leuropäischen Flora, vol. 3. Jena: Fischer.Meusel, H., Jäger, E. & Weinert, E. 1965. Vergleichende Chorologie
der zentraleuropäischen Flora, vol. 1. Jena: Fischer.Mráz, P., Gaudeul, M., Rioux, D., Gielly, L., Choler, P., Taberlet,
P. & Intrabiodiv Consortium 2007. Genetic structure of Hypo-chaeris uniflora (Asteraceae) suggests vicariance in the Carpath-ians and rapid post-glacial colonization of the Alps from an eastern Alpine refugium. J. Biogeogr. 34: 2100–2114.
http://dx.doi.org/10.1111/j.1365-2699.2007.01765.xNei, M. & Li, W.H. 1979. Mathematical model for studying genetic vari-
ation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. U.S.A. 76: 5269–5273. http://dx.doi.org/10.1073/pnas.76.10.5269
Otto, S.P. & Whitton, J. 2000. Polyploid incidence and evolution. Annual Rev. Genet. 34: 401–437.
http://dx.doi.org/10.1146/annurev.genet.34.1.401Parisod, C., Holderegger, R. & Brochmann, C. 2010. Evolutionary
consequences of autopolyploidy. New Phytol. 186: 5–17. http://dx.doi.org/10.1111/j.1469-8137.2009.03142.xPark, J.M., Kovačić, S., Liber, Z., Eddie, W.M.M. & Schneeweiss,
G.M. 2006. Phylogeny and biogeography of isophyllous species of Campanula (Campanulaceae) in the Mediterranean area. Syst. Bot. 31: 862–880. http://dx.doi.org/10.1600/036364406779695924
Petrova, A. 2000. Karyological study of some species of Sesleria (Po aceae) growing in Bulgaria. Bot. Chron. (Patras) 13: 133–140.
Pritchard, K.J., Stephens, M. & Donnelly, P. 2000. Inference of population structure using multilocus genotype data. Genetics 155: 945–959.
Puşcaş, M., Choler, P., Tribsch, A., Gielly, L., Rioux, D., Gaudeul, M. & Taberlet, P. 2008. Post-glacial history of the dominant al-pine sedge Carex curvula in the European Alpine System inferred from nuclear and chloroplast markers. Molec. Ecol. 17: 2417–2429.
http://dx.doi.org/10.1111/j.1365-294X.2008.03751.xR Development Core Team 2010. R: A language and environment
for statistical computing, version 2.13.1. Vienna: R Foundation for Statistical Computing. http://www.r-project.org/foundation/
Ramsey, J. & Schemske, D.W. 1998. Pathways, mechanisms, and rates of polyploid formation in flowering plants. Annual Rev. Ecol. Syst. 29: 467–501. http://dx.doi.org/10.1146/annurev.ecolsys.29.1.467
Rebernig, C.A., Schneeweiss, G.M., Bardy, K.E., Schönswetter, P., Villasenor, J.L., Obermayer, R., Stuessy, T.F. & Weiss-Schnee-weiss, H. 2010. Multiple Pleistocene refugia and Holocene range expansion of an abundant southwestern American desert plant species (Melampodium leucanthum, Asteraceae). Molec. Ecol. 19: 3421–3443. http://dx.doi.org/10.1111/j.1365-294X.2010.04754.x
Rohlf, F.J. 1997. NTSYS-pc: Numerical taxonomy and multivariate analysis system, version 2.0.2. Setauket, New York: Exeter Software.
Ronikier, M. 2011. Biogeography of high-mountain plants in the Car-pathians: An emerging phylogeographical perspective. Taxon 60: 373–389.
Ronikier, M., Cieślak, E. & Korbecka, G. 2008. High genetic dif-ferentiation in the alpine plant Campanula alpina Jacq. (Cam-panulaceae): Evidence for glacial survival in several Carpathian regions and long-term isolation between the Carpathians and the Alps. Molec. Ecol. 17: 1763–1775.
http://dx.doi.org/10.1111/j.1365-294X.2008.03664.xSchönswetter, P., Solstad, H., García, P.E. & Elven, R. 2009. A com-
bined molecular and morphological approach to the taxonomically intricate European mountain plant Papaver alpinum s.l. (Papav-eraceae)—taxa or informal phylogeographical groups? Taxon 58: 1326–1343.
Schur, P.J.F. 1856. Ueber siebenbürgische Seslerien. Verh. K.K. Zool.-Bot. Ges. Wien 6: 191–214.
Sekulić, B., Lovrić, A.Ž. & Rac, M. 1988. Značajna flora i fitocenoze ultramafitnih stijena Banije i odnos prema ostalim balkanskim ofiolitima [Important flora and phytocoenosis of ultramaphitic rocks of Banija and their relationship to other Balkan ophiolits]. Pp. 349–356 in: Slišković, T. (ed.), Zbornik referata naučnog skupa Minerali, stijene, izumrli i živi svijet Bosne i Hercegovine [Pro-ceedings of the Symposium Minerals, rocks, extinct and living world of Bosnia and Herzegovina]. Sarajevo: Zemaljski muzej Bosne i Hercegovine. [in Croatian]
Soltis, D.E., Soltis, P.S., Schemske, D.W, Hancock, J.F., Thomp-son, J.N., Husband, B.C. & Judd, W.S. 2007. Autopolyploidy in angiosperms: Have we grossly underestimated the number of species? Taxon 56: 13–30.
Soltis, D.E., Albert, V.A., Leebens-Mack, J., Bell, C.D., Paterson, A.H., Zheng, C., Sankoff, D., dePamphilis, C.W., Wall, P.K. & Soltis, P.S. 2009. Polyploidy and angiosperm diversification. Amer. J. Bot. 96: 336–348. http://dx.doi.org/10.3732/ajb.0800079

471
Kuzmanović & al. • Differentiation of Sesleria rigida sensu Fl. Eur.TAXON 62 (3) • June 2013: 458–472
471Version of Record (identical to print version).
StatSoft 1996. STATISTICA (data analysis software system), version 5.1. Tulsa: StatSoft Inc. www.statsoft.com
Stebbins, G.L. 1971. Chromosomal evolution in higher plants. London: Edward Arnold.
Stefanović, S., Lakušić, D., Kuzmina, M., Međedović, S., Kit Tan & Stevanović, V. 2008. Molecular phylogeny of Edraianthus (Grassy Bells; Campanulaceae) based on non-coding plastid DNA sequences. Taxon 57: 452–475.
Stevanović, V., Jovanović, S., Lakušić, D. & Niketić, M. 1995. Diverzitet vaskularne flore Jugoslavije sa pregledom vrsta od međunarodnog značaja [Diversity of vascular plants of Yugosla-via, with review of internationally significant species]. Pp. 183–217 in: Stevanović, V. & Vasić, V. (eds.), Biodiverzitet Jugoslavije sa pregledom vrsta od međunarodnog značaja [Biodiversity of Yugo-slavia with an overview of species of global importance]. Belgrade: Ecolibri, Biološki fakultet. [in Serbian]
Stevanović, V., Kit, T. & Iatrou, G. 2003. Distribution of the endemic Balkan flora on serpentine I. – Obligate serpentine endemics. Pl. Syst. Evol. 242: 149–170.
http://dx.doi.org/10.1007/s00606-003-0044-8Strgar, V. 1979. Kromosomska števila treh balkanskih seslerij. Biol.
Vestn. 27: 71–74. [in Slovenian with German summary]Suda, J. & Travniček, P. 2006a. Estimation of relative nuclear DNA
content in dehydrated plant tissues by flow cytometry. Unit 7.30, 7.30.1–7.30.14 in: Robinson, J.P., Darzynkiewicz, Z., Dobrucki, J., Hyun, W., Nolan, J., Orfao, A. & Rabinovitch, P. (eds.), Current protocols in cytometry. New York: Wiley.
Suda, J. & Travniček, P. 2006b. Reliable DNA ploidy determination in dehydrated tissues of vascular plants by DAPI flow cytometry–new prospects for plant research. Cytometry 69A: 273–280.
http://dx.doi.org/10.1002/cyto.a.20253Surina, B., Schönswetter, P. & Schneeweiss, G.M. 2011. Quaternary
range dynamics of ecologically divergent species (Edraianthus
serpyllifolius and E. tenuifolius, Campanulaceae) within the Balkan refugium. J. Biogeogr. 38: 1381–1393.
http://dx.doi.org/10.1111/j.1365-2699.2011.02493.xTatić, B. 1976. Sesleria Scop. Pp. 325–330 in: Josifović, M. (ed.), Flora
SR Srbije, vol. 8. Belgrade: Srpska akademija nauka i umetnosti.Turrill, W.B. 1929. The plant-life of the Balkan peninsula: A phytogeo-
graphical study. Oxford: Clarendon Press.Ujhelyi, J. 1959. Species Sesleriae generis novae. Feddes Repert. 62:
59–70. http://dx.doi.org/10.1002/fedr.19590620108Ujhelyi, J. 1960. Weitere zytotaxonomische Beiträge zur Kenntnis der
Gattung Sesleria. Bot. Közlem. 48: 278–280.Valdés, B. & Scholz, H. Last updated January 2011. Sesleria serbica
(Adamović) Ujhelyi. Euro+Med Plantbase – the information re-source for Euro-Mediterranean plant diversity. http://ww2.bgbm .org/EuroPlusMed/PTaxonDetail.asp?NameId=142052&PTRefFk=7100000 (accessed 4. Nov. 2012).
Van de Peer, Y. & De Wachter, R. 1997. Construction of evolution-ary distance trees with TREECON for Windows: Accounting for variation in nucleotide substitution rate among sites. Computer Applic. Biosci. 13: 227–230.
Van de Peer, Y., Fawcett, J.A., Proost, S., Sterck, L. & Vandepoele, K. 2009. The flowering world: A tale of duplications. Trends Pl. Sci. 14: 680–688. http://dx.doi.org/10.1016/j.tplants.2009.09.001
Vos, P., Hogers, R., Bleeker, M., Reijans, M., Van de Lee, T., Hornes, M., Frijters, A., Pot, J., Peleman, J., Kuiper, M. & Zabeau, M. 1995. AFLP: A new technique for DNA fingerprinting. Nucl. Acids Res. 23: 4407–4414. http://dx.doi.org/10.1093/nar/23.21.4407
Wendel, J.F. 2000. Genome evolution in polyploids. Pl. Molec. Biol. 42: 225–249. http://dx.doi.org/10.1023/A:1006392424384
Wood, T.E., Takebayashi, N., Barker, M.S., Mayrose, I., Greenspoon, P.B. & Rieseberg, L.H. 2009. The frequency of polyploid specia-tion in vascular plants. Proc. Natl. Acad. Sci. U.S.A. 106: 13875–13879. http://dx.doi.org/10.1073/pnas.0811575106
Appendix 1. The names in bold represent four species recognized in our study, the rankless entities used in this study (see Materials and Methods) are in roman letters. The columns are as follows: Population code (ID), sampling locality, number of individuals used for relative genome size measurements (NGS), ploidy level, chromosome number (2n), relative and absolute genome size (GS) of the investigated populations of Sesleria rigida s.l., number of individuals used for AFLP analysis (NAFLP), population used for morphometrics (M), number of herbarium voucher stored in the herbarium BEOU (No.), collector information.
Taxon ID Locality NGS
Ploidy level 2n
Relative GS ± SD
Absolute GS NAFLP M No. Collector(s)
S. achtarovii
achtarovii S333 Bulgaria, Asenovgrad, Kuru Dere 5 4x 0.700 ± 0.005 5 x 31961 Vukojičić, S. & al.
achtarovii S335 Bulgaria, Trigrad, Trigradsko ždrelo, near waterfall
5 4x 28 0.678 ± 0.005 5 x 31973 Vukojičić, S. & al.
achtarovii S350 Greece, Makedonia, Pangaeon 5 4x 0.701 ± 0.003 3 x 31559 Lakušić, D., Lakušić, B.
achtarovii S351 Greece, Aegian islands, Tassos, Ipsarion
5 4x 0.699 ± 0.004 4 x 31558 Lakušić, D., Lakušić, B.
S. filifolia
filifolia S317 Romania, Dubova, Mali Kazan 5 4x 28 0.667 ± 0.015 3 x 31515 Kuzmanović, N., Comanescu, P.
filifolia S372 Romania, Carasova 5 4x 0.646 ± 0.007 5 32107 Kuzmanović, N., Plećaš, M.
pancicii S245 Bulgaria, Zemen, Zemenski prolom 10 4x 28 0.662 ± 0.015 5 30442 Lakušić, D. & al.
pancicii S439 Serbia, Debeli Lug, Valja Fundata 5 4x 0.667 ± 0.006 3 33936 Kuzmanović, N.
rigida S225 Serbia, Golubac, Golubac gorge 6 4x 28 0.624 ± 0.007 5 21166 Lakušić, D., Lakušić, B.
rigida S228 Serbia, Malinik, canyon of Lazareva river
10 4x 28 0.659 ± 0.018 5 x 27098 Lakušić, D., Kuzmanović, N.
rigida S230 Serbia, Niš, Jelašnica gorge 10 4x 28 0.649 ± 0.017 5 x 27444 Kuzmanović, N.
rigida S234 Serbia, Gornjak gorge 10 4x 28 0.692 ± 0.001 5 30320 Lakušić, D.
rigida S236 Serbia, Kučevo, Pek gorge 10 4x 0.670 ± 0.012 8.584 5 x 30322 Lakušić, D.

472
TAXON 62 (3) • June 2013: 458–472Kuzmanović & al. • Differentiation of Sesleria rigida sensu Fl. Eur.
472 Version of Record (identical to print version).
S. filifolia (continued)
rigida S237 Serbia, Paraćin, Izvor, gorge of river Grza
10 4x 0.660 ± 0.015 5 30323 Vukojičić, S.
rigida S238 Serbia, Pirot, Rsovci 10 8x 56 1.312 ± 0.008 5 x 30324 Zlatković, B.
rigida S239 Serbia, Bor, Stol 5 4x 28 0.666 ± 0.003 5 30325 Fusijanović, I.
rigida S240 Serbia, Bor, Rgotski kamen 10 4x 0.681 ± 0.009 5 30326 Fusijanović, I.
rigida S241 Serbia, Sićevo gorge, monastery Sv. Petka
10 8x 1.305 ± 0.015 5 30327 Ranđelović, V.
rigida S246 Bulgaria, Mt. Konjevska, Viden 10 4x 0.664 ± 0.007 5 x 30495 Lakušić, D. & al.
rigida S247 Serbia, Vidlič, Visoka stena 10 8x 56 1.325 ± 0.029 5 30497 Lakušić, D. & al.
rigida S250 Serbia, Mt. Suva planina, Trem 10 8x 56 1.292 ± 0.004 5 x 30546 Kuzmanović, N. & al.
rigida S354 Serbia, Mt. Rtanj 5 4x 0.651 ± 0.005 5 32062 Lakušić, D. & al.
rigida S355 Serbia, Mt. Rudina 5 4x 0.651 ± 0.010 4 32003 Ranđelović, V.
rigida S356 Serbia, Soko Banja, Soko Grad 5 4x 0.664 ± 0.006 5 32002 Vukojičić, S., Kuzmanović, N., Jušković, M.
rigida S402 Bulgaria, Veliko Trnovo, Jantra river gorge (Kandiljka hill)
5 4x 0.661 ± 0.023 5 33234 Niketić, M., Tomović, G.
rigida S403 Bulgaria, Mts. Central Stara Planina, Šipčenska Planina (Ispolin peak)
5 4x 0.661 ± 0.009 5 33275 Niketić, M., Tomović, G.
rigida S466 Serbia, Mt. Mali Vukan 10 8x 56 1.321 ± 0.011 16.649 4 34165 Lakušić, D. & al.
rigida S468 Serbia, Mt. Suva planina, Sokolov kamen
10 8x 56 1.345 ± 0.017 16.895 5 34169 Lakušić, D. & al.
S. rigida
degenii S353 Romania, Domogled 5 4x 28 0.682 ± 0.001 5 x 31538 Lakušić, D. & al.
rigida S242 Romania, Deva, Craciunesti 10 4x 28 0.669 ± 0.013 8.872 5 x 30329 Kuzmanovic, N., Comanescu, P.
rigida S243 Romania, Baia Mare, Remeti 10 4x 28 0.668 ± 0.011 5 x 30333 Kuzmanovic, N., Comanescu, P.
rigida S244 Romania, Braşov, Timpa 10 4x 28 0.690 ± 0.010 9.208 4 x 30337 Kuzmanovic, N., Comanescu, P.
rigida S319 Romania, Turda, Cheile Turzii 5 4x 28 0.642 ± 0.010 8.917 3 x 31517 Kuzmanovic, N., Comanescu, P.
rigida S352 Romania, Domogled, Prolaz 5 4x 0.664 ± 0.009 5 31537 Lakušić, D. & al.
S. serbica
serbica S269 Serbia, Zlatibor, Zlatiborsko jezero 10 4x 0.643 ± 0.003 5 28741 Kuzmanović, N.
serbica S272 Serbia, Divčibare, Ljuti krš 10 4x 0.650 ± 0.007 4 x 30308 Kuzmanović, N.
serbica S273 Serbia, Mokra Gora, Šargan 10 4x 0.637 ± 0.010 4 x 30309 Lakušić, D.
serbica S274 Serbia, Ibar valley, Maglić old town 10 4x 0.645 ± 0.011 5 30310 Lakušić, D.
serbica S275 Serbia, Mali Vujan, Banjska vrata 10 4x 28 0.651 ± 0.008 5 30311 Lakušić, D.
serbica S361 Serbia, Mt. Studena, Krčanik 5 4x 0.643 ± 0.002 4 31552 Kuzmanović, N.
serbica S373 Bosnia & Herzegovina, Zavidovići, Gostovićka river (Sv. Pantelej)
5 4x 0.635 ± 0.007 4 32105 Niketić, M., Tomović, G.
serbica S374 Serbia, Rogozna, Izbice-Negotinac (mouth of Seoski potok)
5 4x 0.631 ± 0.004 4 32150 Niketić, M., Tomović, G.
serbica S422 Serbia, Mt. Kopaonik, Treska 5 4x 28 0.666 ± 0.007 5 33674 Kuzmanović, N. & al.
serbica S436 Serbia, Mt. Ozren, Kamarišta 5 4x 0.669 ± 0.004 5 33875 Niketić, M., Tomović, G.
S. taygetea
taygetea S362 Greece, Peloponnisos, Taigetos, Langada canyon
3 4x 0.758 ± 0.000 5 31851 Lakušić, D. & al.
Appendix 1. Continued.
Taxon ID Locality NGS
Ploidy level 2n
Relative GS ± SD
Absolute GS NAFLP M No. Collector(s)




















