Cytological analysis of infection process and the first defence responses induced in winter rye (...
8
Cytological analysis of infection process and the first defence responses induced in winter rye (Secale cereale L.) seedlings inoculated with Microdochium nivale Iwona Anna _ Zur a, * , Ewa Dubas a , Ewa Pociecha b , Franciszek Dubert a , Irena Kolasi nska c , Agnieszka P1a _ zek b a Institute of Plant Physiology, Polish Academy of Sciences, Niezapominajek 21, 30-239 Kraków, Poland b Agricultural University, Faculty of Agriculture and Economics, Department of Plant Physiology Al. Mickiewicza 21, 31-120 Kraków, Poland c Plant Breeding and Acclimatization Institute at Radzikow, Department of Plant Breeding and Genetics 05-870 Blonie, Poland article info Article history: Accepted 28 September 2011 Keywords: Callose Defence response Microdochium nivale (Fr) Samuels & Hallett Pathogenesis Pink snow mould Phenolic Secale cereale (L.) abstract Pink snow mould caused by Microdochium nivale can lead to serious yield loss in many winter cereals and turf grasses. The study examined the infection mechanism in four inbred lines and one cultivar of rye (Secale cereale L.) with different sensitivities to this pathogenic fungus. Fungus development was analyzed at the early stages of the infection, up to the 13th day after inoculation, in hardened and non- hardened seedlings, using light/fluorescence microscopy. The cytological analysis was supplemented with a spectrophotometrical assay of the phenolics. Significantly higher phenolic content was revealed in the leaves of the lines more resistant to M. nivale, as compared to those which are more sensitive. Ó 2011 Elsevier Ltd. All rights reserved. 1. Introduction Owing to its considerable winter hardiness and low environ- mental demands, rye (Secale cereale L.) is a cereal which played a major role in the feeding of European populations throughout the Middle Ages [1]. Today, it is cultivated on 11 mln ha worldwide and annual production amounts to approximately 17.2 millions tons. Its high frost tolerance means that it is frequently grown under conditions where other cereals fail. The winter survival of rye is most seriously endangered by its low resistance to psychrophilic fungi, which can attack when the temperature at the snow-plant interface remains at around 0 C. This group of pathogens contains more than a dozen species that commonly occur in high latitude regions [2]. In Poland and other cold or temperate regions of the northern hemisphere, the most dangerous of these is the pathogenic fungus Microdochium nivale (Fr) Samuels & Hallett. The pink snow mould disease caused by this fungus can inflict damages leading to very serious yield losses and a decrease in yield quality [3,4]. It attacks not only rye, but also many other economically important winter cereals and turf grasses [2,5]. The harmful effects of fungicides on the ecosystem and human health have led to a strong interest in resistant genotype production. In spite of the intensive work being carried out, no report of cultivars highly resistant to snow moulds has been published. One of the possible reasons for this is the fact that neither the mechanism of infection nor the physiological back- ground for snow mould resistance has been recognized. The process of infection induced by M. nivale mycelium, which is the main source of M. nivale infection in winter field conditions, was studied earlier on triticale [6]. Single infection hyphae pene- trating into the host tissues via stomata were visualized in crowns, leaf sheaths and leaves of inoculated seedlings. For the first time, the authors reported the presence of haustoria-like structures produced by M. nivale in host tissues. It has been observed that resistance to snow mould increases with plant size and age [7e10]. Nevertheless, the most crucial factor for the expression of snow mould resistance is the hardening of plants at a low temperature [7,11]. Although this relationship has been reported on numerous occasions, an understanding of the precise nature of this effect is still lacking [9]. This positive influ- ence of low temperature on M. nivale resistance can be explained by the ‘cross-tolerance’ process, where exposure to a stress factor induces a wide spectrum of defence reactions that better prepare the plant to face subsequent stress in the near future [12]. The same positive effect of low temperature on plant resistance has also been observed for other fungal diseases such as rust [13] and powdery mildew [14], which suggests a more non-specific form of plant response. Phenylalanine ammonia-lyase (PAL) acti- vation and a change in phenolic content are common elements in the mechanisms of pathogen resistance acquisition [15]. Phenolic synthesis is initiated very rapidly in response to many stress factors, * Corresponding author. Tel.: þ48 12 425 33 01; fax: þ48 12 425 18 44. E-mail address: [email protected] (I.A. _ Zur). Contents lists available at SciVerse ScienceDirect Physiological and Molecular Plant Pathology journal homepage: www.elsevier.com/locate/pmpp 0885-5765/$ e see front matter Ó 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.pmpp.2011.09.007 Physiological and Molecular Plant Pathology 76 (2011) 189e196
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Cytological analysis of infection process and the first defence responses induced in winter rye (...

at SciVerse ScienceDirect
Physiological and Molecular Plant Pathology 76 (2011) 189e196
Contents lists available
Physiological and Molecular Plant Pathology
journal homepage: www.elsevier .com/locate/pmpp
Cytological analysis of infection process and the first defence responses inducedin winter rye (Secale cereale L.) seedlings inoculated with Microdochium nivale
Iwona Anna _Zur a,*, Ewa Dubas a, Ewa Pociecha b, Franciszek Dubert a, Irena Kolasi�nska c, Agnieszka P1a _zek b
a Institute of Plant Physiology, Polish Academy of Sciences, Niezapominajek 21, 30-239 Kraków, PolandbAgricultural University, Faculty of Agriculture and Economics, Department of Plant Physiology Al. Mickiewicza 21, 31-120 Kraków, Polandc Plant Breeding and Acclimatization Institute at Radzikow, Department of Plant Breeding and Genetics 05-870 Błonie, Poland
a r t i c l e i n f o
Article history:Accepted 28 September 2011
Keywords:CalloseDefence responseMicrodochium nivale (Fr) Samuels & HallettPathogenesisPink snow mouldPhenolicSecale cereale (L.)
* Corresponding author. Tel.: þ48 12 425 33 01; faE-mail address: [email protected] (I.A. _Zur).
0885-5765/$ e see front matter � 2011 Elsevier Ltd.doi:10.1016/j.pmpp.2011.09.007
a b s t r a c t
Pink snowmould caused byMicrodochium nivale can lead to serious yield loss in many winter cereals andturf grasses. The study examined the infection mechanism in four inbred lines and one cultivar of rye(Secale cereale L.) with different sensitivities to this pathogenic fungus. Fungus development wasanalyzed at the early stages of the infection, up to the 13th day after inoculation, in hardened and non-hardened seedlings, using light/fluorescence microscopy. The cytological analysis was supplementedwith a spectrophotometrical assay of the phenolics. Significantly higher phenolic content was revealed inthe leaves of the lines more resistant to M. nivale, as compared to those which are more sensitive.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Owing to its considerable winter hardiness and low environ-mental demands, rye (Secale cereale L.) is a cereal which playeda major role in the feeding of European populations throughout theMiddle Ages [1]. Today, it is cultivated on 11 mln ha worldwide andannual production amounts to approximately 17.2 millions tons. Itshigh frost tolerance means that it is frequently grown underconditions where other cereals fail.
The winter survival of rye is most seriously endangered by itslow resistance to psychrophilic fungi, which can attack when thetemperature at the snow-plant interface remains at around 0 �C.This group of pathogens contains more than a dozen species thatcommonly occur in high latitude regions [2]. In Poland and othercold or temperate regions of the northern hemisphere, the mostdangerous of these is the pathogenic fungus Microdochium nivale(Fr) Samuels & Hallett. The pink snowmould disease caused by thisfungus can inflict damages leading to very serious yield losses anda decrease in yield quality [3,4]. It attacks not only rye, but alsomany other economically important winter cereals and turf grasses[2,5]. The harmful effects of fungicides on the ecosystem andhuman health have led to a strong interest in resistant genotypeproduction. In spite of the intensive work being carried out, noreport of cultivars highly resistant to snow moulds has been
x: þ48 12 425 18 44.
All rights reserved.
published. One of the possible reasons for this is the fact thatneither the mechanism of infection nor the physiological back-ground for snow mould resistance has been recognized.
The process of infection induced byM. nivalemycelium, which isthe main source of M. nivale infection in winter field conditions,was studied earlier on triticale [6]. Single infection hyphae pene-trating into the host tissues via stomata were visualized in crowns,leaf sheaths and leaves of inoculated seedlings. For the first time,the authors reported the presence of haustoria-like structuresproduced by M. nivale in host tissues.
It has been observed that resistance to snow mould increaseswith plant size and age [7e10]. Nevertheless, the most crucialfactor for the expression of snowmould resistance is the hardeningof plants at a low temperature [7,11]. Although this relationship hasbeen reported on numerous occasions, an understanding of theprecise nature of this effect is still lacking [9]. This positive influ-ence of low temperature onM. nivale resistance can be explained bythe ‘cross-tolerance’ process, where exposure to a stress factorinduces a wide spectrum of defence reactions that better preparethe plant to face subsequent stress in the near future [12].
The same positive effect of low temperature on plant resistancehas also been observed for other fungal diseases such as rust [13]and powdery mildew [14], which suggests a more non-specificform of plant response. Phenylalanine ammonia-lyase (PAL) acti-vation and a change in phenolic content are common elements inthe mechanisms of pathogen resistance acquisition [15]. Phenolicsynthesis is initiated very rapidly in response tomany stress factors,

IwonaA. _Zur et al. / Physiological and Molecular Plant Pathology 76 (2011) 189e196190
such as high light intensity, UV irradiation, drought, salinity, cold,starvation, wounding, pathogen attack and elicitation [15,16]. Theproducts of phenylpropanoid biosynthesis are mainly utilized ascell wall strengthening compounds, but they also have directantimicrobial activity and can be used as signaling molecules [15].
This study is the first part of a project aimed at the identificationof the physiological factors important for resistance to pink snowmould in winter rye. It examined the early response, namely thefirst two weeks after plant inoculation with fungal mycelium, infour inbred lines and one cultivar of rye (S. cereale L.) with differentsensitivities to M. nivale. In order to explain the mechanism of lowtemperature effect, the progress of the infection process wasanalyzed in hardened and non-hardened seedlings, using light andfluorescence microscopy. The hypothesis that phenolic metabolismcan constitute a part of the defence system against M. nivale wasverified by monitoring the changes in phenolic accumulation afterplant pathogen inoculation.
2. Material and methods
The study was performed in two independent experiments.
2.1. Plant material and growth conditions
The seeds of cultivar ‘Stach’ and four inbred lines (2, 4, 22, 23) ofwinter rye (Secale cereale L.) were obtained from the Institute ofPlant Breeding and Acclimatization in Radzików (Poland). Accord-ing to the breeders’ classification, both cultivar ‘Stach’ and line Nos.2 and 4 were more resistant toM. nivale infection, in comparison tohighly sensitive line Nos. 22 and 23. The seeds were sown in potscontaining a mixture of soil and sand (3/1; v/v) and cultured ina growth chamber, at 18 �C, at a 10/14 h (day/night) photoperiodwith a light intensity of 250 mmolm�2 s�1 photosynthetic photonflux density (PPFD), for a period of six weeks. The plants were thenpre-hardened for two weeks at 12 �C, under the light regimedescribed above. Subsequently, the plants were cold-acclimated forthree weeks at 2 �C, at a 8/16 h (day/night) photoperiod, witha light intensity of 150 mmolm�2 s�1 PPFD.
2.2. Microdochium nivale inoculation
M. nivale mycelium, an isolate of No. 38z/5a/01, collected fromrye in 2001, was kindly provided by Professor Maria Pro�nczuk fromthe Institute of Plant Breeding and Acclimatization, Radzikow,Poland. The myceliumwas grown on Potato Dextrose Agar medium(PDA, SigmaeAldrich) for 7 days, at 20 �C, in darkness. The agar andmycelium were next transferred to Erlenmeyer flasks, with 3 discsof 2 cm in diameter per flask; the flasks contained 200 cm3 ofsterile soil, peat and sand mixture (2/2/1; v/v/v) and 5% w/w ofground wheat seeds. They were then cultured for 14 days at thesame light/temperature conditions.
Both, hardened and non-hardened seedlings were inoculated inaccordance with the ‘cold-chamber’ method [17]. The myceliumwas gentlymixed and spread on the soil in each pot, at a measure of1 g per plant. Each pot was then covered with moistened blottingpaper and black plastic foil to imitate the conditions occurring underthe snow cover and kept for a period of 5 weeks at 2 �C, in darkness.
2.3. Light and fluorescence microscopy
Microscopic observations were performed by means of a NIKONECLIPSE-E 600 fluorescent microscope equipped with Nomarskidifferential interference contrast (DIC). Images were recorded usinga Digital DS-Ri1 camera and processed by the Laboratory ImagingLtd. NIS-Elements AR 2.10 programme.
Five plants from each of three pots for each cultivar/treatmentcombination were studied on the 1st, 5th, 9th and 13th day postinoculation (dpi). For cytological analysis, fragments of living planttissues, namely leaves and leaf sheaths of around 5 mm in length, aswell as cross-sections of rooteshoot junctions, which is to say, thecrowns, and of roots were prepared.
The material was stained for 5 min, in the dark, directly onmicroscopic slides, with drop of fluorescent dyemixture: CalcofluorWhite [18] and Aniline Blue [19] (v/v, 1/1). CalcofluorWhite (18909,stock solution at 1% w/v in H2O; BioChemika) was used at a 0.01%working concentration, and Aniline Blue (415049; stock solution at0.5% w/v in H2O; pH 8.2; SigmaeAldrich) was used at a 0.1%working solution. For visualization of the M. nivale hyphae, theanalysis was carried out under UV light: Exc 330e380 nm, DM400 nm, BA 420 nm.
For improvement of M. nivale hyphae detection, light micros-copy was also used; staining with lactophenol cotton blue (LPCB,Fluka 61335) was preceded by the clearing of the plant tissues.Leaves, leaf sheaths, crowns, and roots cut into pieces of approxi-mately 5 mm in length, were immersed in a clearing solutioncontaining 100% chloral hydrate (w/v) and 90% lactic acid (2/1; v/v),vacuum infiltrated for 1 h and incubated for the next 24 h at roomtemperature. They were then thoroughly washed in tap water,stained with the drop of the dye on a microscopic slide andobserved under a light microscope equipped with a DIC system.
The presence of phenolic compounds and callose was investi-gated on the basis of autofluorescence and fluorescence afteraniline blue staining, respectively. All the samples were analyzedunder a fluorescent microscope in UV light: Exc 330e380 nm, DM400 nm, BA 420 nm.
2.4. Evaluation of regrowth after M. nivale inoculation
At the end of the 5 week-incubation at 1 �C in darkness, theblotting paper and foil were removed. The plants were cut andallowed to grown for 3 days at 12 �C in 100 mmolm�2 s�1 PPFD lightand then for 7 days at 18 �C in 250 mmol�2 s�1 PPFD light.
At the end of this 10-day period, the plant regrowth was eval-uated in the cold-hardened and non-hardened seedlings sepa-rately, using the arbitrary visual rating system (0e5) where0 means a healthy plant with no visible symptoms of infection,1 means a plant without any visible symptoms of infection, but theinner leaves may be brown, 2 means that the plant has survived,but shows damage to less than 50% of its leaves, 3 means that theplant has survived but shows severe damage to 50% of its leaves,4 means that the plant has survived but it is badly damaged, withthe leaves frequently discoloured and curled, and 5 means a deadplant, with no signs of leaf elongation.
The average regrowth was calculated from the ratings describedabove in accordance with the formula:
½ðn� 0Þ þ ðn� 1Þ þ//þ ðn� 5Þ�N�1;
n e the number of plants corresponding to each disease rating(0e5), N e total observations.
The calculation was made in 5 replicates as an average from fivepots, with 25 plants to a pot and each pot¼ one replicate.Decreased values indicate an increase in plant resistance to thepathogen.
2.5. Phenolic estimation
250 mg samples of the third leaf and crown of hardened,infected and hardened, non-infected seedlings were collected onthe 1st, 5th, 9th and 13th dpi and kept at �80 �C.

IwonaA. _Zur et al. / Physiological and Molecular Plant Pathology 76 (2011) 189e196 191
They were then boiled in 1 cm3 of 80% ethanol, homogenized in2 cm3 of 80% ethanol and centrifuged at 2500� g for 20 min. Thesupernatant was mixed with 25% Na2CO3 and FolineCiocalteaureagent [20]. The absorbance of the samples was measured atl¼ 760 nm by means of spectrophotometer Ultrospec 2100 pro(Biosciences Amersham, Sweden). The total phenolic content wascalculated as milligrams of chlorogenic acid per 1 g of fresh mass(FW). The colorimetric method applied permits the estimation ofwater soluble phenolic compounds with an aromatic ring and atleast one hydroxyl group. The analysis was made in 5 replicates(plants).
2.6. Statistical analysis
The effects of the infection on the phenolic content in the leavesand crowns of the rye seedlings were all tested using the F-test(ANOVA/MANOVA). The results obtained were analyzed using
Fig. 1. (1.1) The progress of Microdochium nivale infection on inbred line 2 rye (Secale cerealeof the root-shoot junction. A, B. Dense network of hyphae on the surface of the leaf sheath (leaf initials; cross-section. In the upper left-hand corner of C, a larger magnification of the hyto the endodermis; cross section (dark blue, arrowheads). The small square in F shows a hihaustorium-like structures (white arrowhead) in the metaxylem vessels (black arrowhead100 mm for (the small square in C). Seedlings, 9 days (A and B) and 13 days after inoculation (cleared in lactic acid; Fluorescence microscopy in UV light (BeD). (1.2) The progress of Microinoculated with fungus mycelium. AeF. Cytological analysis of leaves. A. Dense network of hyof the stomata apparatus (st) (dark blue, arrowhead). C. Hyphae tips (dark blue, arrowheapparatus on the surface of epidermis. E. Intercellular hyphae in mesophyl (m) (arrowheads)forming a vesicle (v) within a mesophyl cell (m). Bars¼ 10 mm for (B, D, E), 20 mm for (F); 1Nomarski optics (AeF). The tissues on AeF are all unstained and have been cleared in lacti
Duncan’s multiple range test at p< 0.05. Differences in theregrowth afterM. nivale inoculation were estimated using the non-parametric Friedman test. All the statistical analyses were carriedout using the STATISTICA 9.0 package.
3. Results
3.1. Microscopic observations
Microscopic analysis permits the precise characterization of thedynamic and progress of the infection process (Fig. 1.1 and 1.2).Generally, the mode of pathogen infection was very similar for allthe rye lines studied; however, the dynamic of fungal growth anddevelopment varied significantly.
In some rye genotypes, namely cv. ‘Stach’ and inbred lines 2 and4, in spite of the lack of visible disease signs on the 1st dpi, singlefine hyphaewere already detectable on the leaf sheaths close to the
L.) seedlings, hardened and inoculated with fungus mycelium. A-G. Cytological analysisdark blue). C, D. Dense network of hyphae (arrowheads) in the crown, at the site of thephae penetration sites (arrowheads). EeF. Hyphae in the cortex cells of the crown, closegher magnification of hyphae penetrating the metaxylem vessel. G. Hyphae producing); cross section. Bars¼ 10 mm for (A, B, E and F), 20 mm for (G); 50 mm for (C and D);CeG). Nomarski optics (A and EeG). The tissues on EeG are all unstained and have beendochium nivale infection on inbred line 2 rye (Secale cereale L.) seedlings, hardened andphae on the surface of the leaf (dark blue). B. The early phase of hyphea (h) penetrationads) penetrating the stomata apparatus. D. Hyphae protruding thorough the stomata; stomata apparatus (st) on the surface of the epidermis. F. A haustorium-like structure00 mm for (A, C). Seedlings, 9 days (A and B) and 13 days after inoculation (C, D, E, F).c acid.

Fig. 1. (continued).
IwonaA. _Zur et al. / Physiological and Molecular Plant Pathology 76 (2011) 189e196192
seedling crowns, at the shoot/root junction. Between the 5th andthe 9th dpi, the M. nivale mycelium grew intensively (Fig. 1.1.AeB).By the 13th dpi, the hyphae had lengthened, branched andprogressively penetrated the crown cortex cells at the site of theleaf initials in the affinity of the endodermis and vascular bundles(Fig. 1.1.CeF). Penetration of hyphae proceeded in the vasculartissue and the production of haustorium-like structures (HLS) inthe metaxylem vessels was observed (Fig. 1.1.G).
Between the 5th and the 9th dpi, fungal mycelium developed onthe leaves growing in contact with infected soil (Fig. 1.2.AeB).Numerous hyphae had entered directly through the stomata(Fig. 1.2.B) into the substomatal chamber and lower layers of themesophyll. On the 13th dpi, a dense hyphal network was found onthe leaf sheaths and leaf epidermis in the line 2 and 4 seedlings.Abundant hyphae had penetrated through the stomata and into theintercellular space. However, hyphal growth in the oppositedirection of was also recorded. The hyphae which first penetratedthe mesophyll grew outwards and protruded through the stomata(Fig. 1.2.CeD). Multiple hyphae were present in the intercellularspaces of the mesophyll (Fig. 1.2.E). In some mesophyll cells, thehyphae swelled and formed haustorium resembling vesicles(Fig. 1.2.F).
In the case of inbred lines 22 and 23, the first signs of theinfectionwere delayed and usually observed on the 5th dpi. Furtherdisease signs were similar, but less severe than in the case of inbredlines 2, 4 and cv. ‘Stach’.
M. nivale infection was accompanied by phenolic accumulationin the cell walls of the leaves and leaf sheath epidermis in almost allthe rye lines studied, with the exception of line 23. It could alreadybe observed on the 1st dpi as a yellow autofluorescence under UVlight. The highest intensity of phenolic autofluorescence wasdetected in the cells nearby the fungal hyphae penetration sites(Fig. 2A). In the leaf sheath epidermis and crown cells of some ryegenotypes: cv. ‘Stach’ and lines 2 and 4, light-green callose fluo-rescence was also detected (Fig. 2B). Callose deposition was
observed in the cell walls of the leaf sheath epidermis, as well as inthe cortex cells of crowns, from the earliest stages of M. nivaleinfection. However, the strongest callose fluorescence was detectedon the 13th dpi.
3.2. Plant regrowth after M. nivale inoculation
Plant resistance to M. nivale was evaluated on the basis ofregrowth ability after 5 week–incubation with fungal mycelium. Inall the rye genotypes studied, the non-hardened seedlings wereinfected so severely that the cut plants display no regrowth ability(Fig. 3). The results obtained for the hardened seedlings are shownin Figs. 3 and 4. The best regrowth in the infected plants was notedin the case of lines 2, 4 and cv. ‘Stach’. A moderate degree of snowmould resistance was demonstrated by line 22, while the leastresistant was line 23.
3.3. Phenolic content
Phenolic content was estimated separately in the leaves andcrowns of rye seedlings which had first been hardened and theninfected with M. nivale and in hardened but non-infected ryeseedlings. The analysis of variance revealed that the phenoliccontent in both plant organs was significantly influenced by plantgenotype and the amount of time which had passed followinginoculation. No effect of fungus infection was detected.
In the leaves of both the control and infected seedlings of all thegenotypes studied, the total phenolic content per mg FW wassignificantly higher as comparedwith its level in the crowns (Fig. 5aand b). Spectophotometrical analysis also revealed that, in theleaves of the more resistant lines, namely cv. ‘Stach’ and lines 2 and4, the average phenolic content ranged from 0.77 mg/g FW to0.92 mg/g FWandwas significantly higher (p� 0.05) in comparisonwith the more susceptible lines 22 and 23, at 0.65e0.67 mg/g FW(Fig. 5a). Moreover, in the case of lines 2 and 4, it increased

Fig. 2. Cytological analysis of the cold hardened seedlings of cv. ‘Stach’ of rye (Secale cereale); host response after inoculation with Microdochium nivale: accumulation of callose andphenolic compounds in infected leave sheaths. A. Autofluorescence of the phenolic compounds (yellow) accumulated in the epidermal cells of the leaf close to the hyphae (blue)penetration site. B. Callose (light-green) in leaf epidermal cells after aniline blue staining. Bars¼ 50 mm for (B); 100 mm for (A). Fluorescence microscopy in UV light (A, B). Non-hardened seedlings, 1 day (A) and 9 days after inoculation (B).
IwonaA. _Zur et al. / Physiological and Molecular Plant Pathology 76 (2011) 189e196 193
significantly on the 9th and 14th dpi, respectively. This increasewasobserved in both infected and non-infected seedlings. At the sametime, no significant changes in phenolic content were noted in themore susceptible rye genotypes, namely lines 22 and 23. Phenolicquantities in seedling crowns were not correlated with the degreeof rye resistance to M. nivale. The same average phenolic level,namely 0.37 mg/g FW, was found in cv. ‘Stach’ and line no. 22,which are quite different in their level of resistance to M. nivale.This was observed in the case of the most resistant line, 4, and themost susceptible line, 23; the average phenolic level gained0.33 mg/g FW in both. The lowest phenolic content was detected ininfected crowns of line 2, at 0.29 mg/g FW.
In each rye genotype studied, the quantity of phenolicsincreased with the time which had lapsed since the transfer of theseedlings to the conditions imitating snow cover (Fig. 5b).
4. Discussion
For winter cereals, low temperature is the most common stressfactor limiting growth, development and productivity [21].However, as rye has the highest winter hardiness and frost resis-tance among all the cereals, the main influence on its winter
Fig. 3. Regrowth ability of hardened seedlings of four Polish winter rye (Secalecereale L.) inbred lines and cv. ‘Stach’, after inoculation with M. nivale mycelium. Themeans of 5 replicates (25 plants to a pot and each pot¼ one replicate)� SE.
survival is usually the duration of snow cover and the severity ofinfection caused by psychrophilic fungi such as M. nivale.
It has been shown that, although the level of resistance toM. nivale depends mainly on the plant genotype [5], it might bemodified by plant age and condition, and by various environmentalfactors. Temperature is particularly crucial, because cold hardeningis necessary for the expression of resistance to this pathogenicfungus [9,22e24]. The results obtained for non-hardened plants inthis study have clearly confirmed this. The mechanism of thisphenomenon is not fully understood, because low temperatureinduces a wide spectrum of physiological and biochemical changesin the plan, while it might be that only some of them have a directeffect on the pathogenesis process and plant resistance acquisition[25].
The inbred lines of rye used in this study were selected bybreeders as being significantly different in their resistance toM. nivale. The estimation of the resistance level, based on the fielddata was, in general, consistent with the results received in thisexperiment, which was carried out in accordance with the ‘cold-chamber’ method.
Cytological examination of the pathogenesis process is quiteoften reported; however, it is mainly related to infection by fungalspores. This differs considerably from infection induced by fungalmycelium, which is the main source ofM. nivale infection in winterfield conditions [5]. The advanced light and fluorescent microscopytechniques used in this study permitted the precise analyses ofM. nivale infection in the seedlings of four inbred lines and onevariety of rye, with various degrees of resistance to this pathogenicfungus. No matter what the plant genotype, the course of theinfection process was very similar. The first sites of fungus attackwere the leaf sheaths and leaf blades growing in contact withinfected soil. The fungal hyphae then entered the leaf initials in thecrowns and the crown’s cortex cells. Here, the mycelium fungalgrew intensively, the hyphae lengthened, branched and progres-sively penetrated the crown cortex cells and vascular bundles(metaxylem). There, swollen hyphae forming vesicle-like struc-tures resembling haustoria were observed. Similar HLS were alsoobserved earlier in triticale infected by M. nivale [6]. However, it isthis study which has detected hyphal penetration into vasculartissue and the production of HLS in metaxylem vessels for the firsttime. Haustorium is a specialized fungal feeding structure that onlydevelops during plant-pathogen interaction and is formed from

Fig. 4. Regrowth ability of winter rye (Secale cereale L.) seedlings of four Polish inbred lines, namely 2, 4, 22 and 23, and cv. ‘Stach’(St), hardened and non-inoculated (control, C) andhardened and inoculated with M. nivale mycelium (Inc).
IwonaA. _Zur et al. / Physiological and Molecular Plant Pathology 76 (2011) 189e196194
both fungal and host components [26,27]. The hyphae also pene-trated the stomata and grew in the mesophyll, producing HLSbefore entering the vascular tissue via the minor veins in the leaf.This observation may suggest that this pathogenic fungus repre-sents the hemibiotrophic lifestyle [27]. Such an infection method isalso typical for other hemibiotrophic fungi, such as rusts (Uromycesfabae on Vicia fabae) or powdery mildew (Blumeria graminis onHordeum vulgare) [28]. However, a more precise analysis is neces-sary in order to confirm this hypothesis.
It could be assumed that the penetration of the hyphae into themetaxylem vessels permits the rapid spread of the fungus insidehost plant. It also can cause embolism of the water transportsystem, which results in wilting and finally, in plant death. Thissupposition, based on the studies of Fusarium infection on lucerne,was presented by Parry and Pegg [29], who identified F. avenaceumand F. culmorum infection hyphae in plant protoxylem. Fortunately,winter rye belongs to the species in which the hydraulic segmen-tation of the root vessel system from the shoot vessel system had
been found [30]. The separation of both conduit systems takes placeat the root/shoot junction and protects the plant from embolismcaused by unlimited fungal hyphae development. No matter howfast the disease development, as long as the root-shoot junctionand root system are not seriously damaged, the plant still possessesa recovery and regrowth ability.
The M. nivale infection process was accompanied by the accu-mulation of phenolic compounds and callose, which were seen asstrong autofluoresence or fluorescence, usually around theentrance site of the pathogen. A similar effect was observed byDubas et al. [6,31] during M. nivale infection in triticale and Loliumperenne seedlings. This observation was not confirmed by thespectrophotometric assay of the phenolics carried out under thisstudy, where no effect of fungus infection on the content of thesecompounds was revealed. However, the lines more resistant toM. nivalewere characterized by a significantly higher phenolic levelin the leaves of both the control and the inoculated seedlings, ascompared to more sensitive ones. This could be explained by the
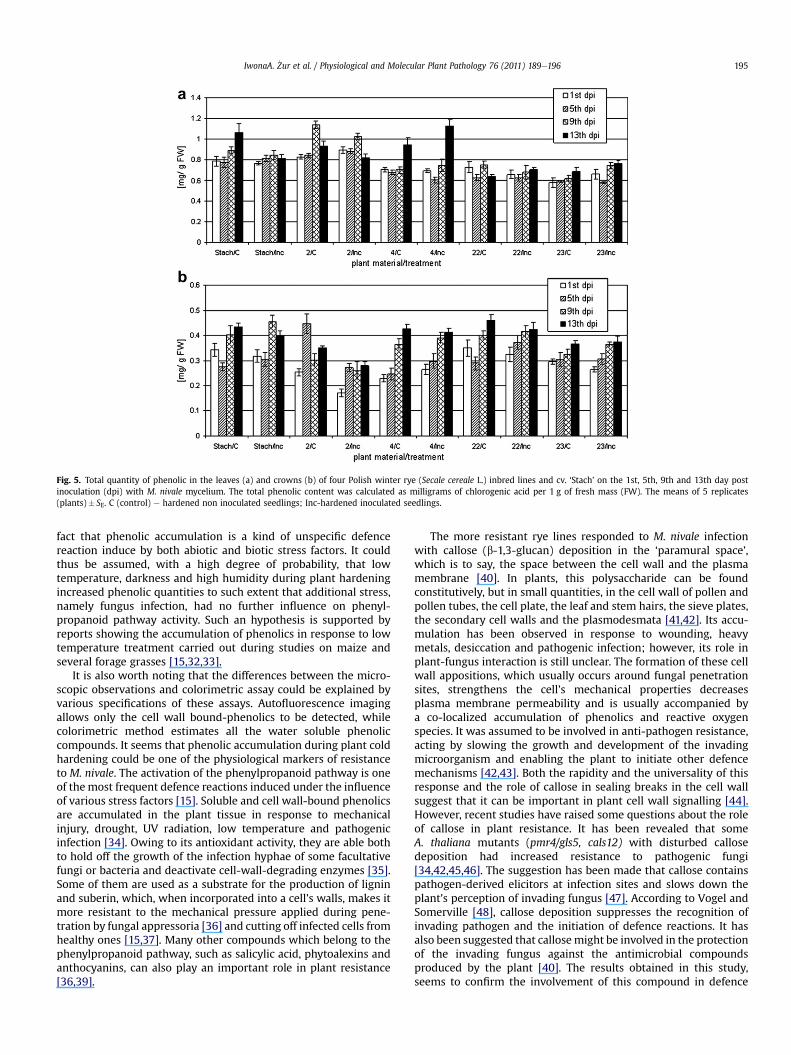
Fig. 5. Total quantity of phenolic in the leaves (a) and crowns (b) of four Polish winter rye (Secale cereale L.) inbred lines and cv. ‘Stach’ on the 1st, 5th, 9th and 13th day postinoculation (dpi) with M. nivale mycelium. The total phenolic content was calculated as milligrams of chlorogenic acid per 1 g of fresh mass (FW). The means of 5 replicates(plants)� SE. C (control) e hardened non inoculated seedlings; Inc-hardened inoculated seedlings.
IwonaA. _Zur et al. / Physiological and Molecular Plant Pathology 76 (2011) 189e196 195
fact that phenolic accumulation is a kind of unspecific defencereaction induce by both abiotic and biotic stress factors. It couldthus be assumed, with a high degree of probability, that lowtemperature, darkness and high humidity during plant hardeningincreased phenolic quantities to such extent that additional stress,namely fungus infection, had no further influence on phenyl-propanoid pathway activity. Such an hypothesis is supported byreports showing the accumulation of phenolics in response to lowtemperature treatment carried out during studies on maize andseveral forage grasses [15,32,33].
It is also worth noting that the differences between the micro-scopic observations and colorimetric assay could be explained byvarious specifications of these assays. Autofluorescence imagingallows only the cell wall bound-phenolics to be detected, whilecolorimetric method estimates all the water soluble phenoliccompounds. It seems that phenolic accumulation during plant coldhardening could be one of the physiological markers of resistanceto M. nivale. The activation of the phenylpropanoid pathway is oneof the most frequent defence reactions induced under the influenceof various stress factors [15]. Soluble and cell wall-bound phenolicsare accumulated in the plant tissue in response to mechanicalinjury, drought, UV radiation, low temperature and pathogenicinfection [34]. Owing to its antioxidant activity, they are able bothto hold off the growth of the infection hyphae of some facultativefungi or bacteria and deactivate cell-wall-degrading enzymes [35].Some of them are used as a substrate for the production of ligninand suberin, which, when incorporated into a cell’s walls, makes itmore resistant to the mechanical pressure applied during pene-tration by fungal appressoria [36] and cutting off infected cells fromhealthy ones [15,37]. Many other compounds which belong to thephenylpropanoid pathway, such as salicylic acid, phytoalexins andanthocyanins, can also play an important role in plant resistance[36,39].
The more resistant rye lines responded to M. nivale infectionwith callose (b-1,3-glucan) deposition in the ‘paramural space’,which is to say, the space between the cell wall and the plasmamembrane [40]. In plants, this polysaccharide can be foundconstitutively, but in small quantities, in the cell wall of pollen andpollen tubes, the cell plate, the leaf and stem hairs, the sieve plates,the secondary cell walls and the plasmodesmata [41,42]. Its accu-mulation has been observed in response to wounding, heavymetals, desiccation and pathogenic infection; however, its role inplant-fungus interaction is still unclear. The formation of these cellwall appositions, which usually occurs around fungal penetrationsites, strengthens the cell’s mechanical properties decreasesplasma membrane permeability and is usually accompanied bya co-localized accumulation of phenolics and reactive oxygenspecies. It was assumed to be involved in anti-pathogen resistance,acting by slowing the growth and development of the invadingmicroorganism and enabling the plant to initiate other defencemechanisms [42,43]. Both the rapidity and the universality of thisresponse and the role of callose in sealing breaks in the cell wallsuggest that it can be important in plant cell wall signalling [44].However, recent studies have raised some questions about the roleof callose in plant resistance. It has been revealed that someA. thaliana mutants (pmr4/gls5, cals12) with disturbed callosedeposition had increased resistance to pathogenic fungi[34,42,45,46]. The suggestion has been made that callose containspathogen-derived elicitors at infection sites and slows down theplant’s perception of invading fungus [47]. According to Vogel andSomerville [48], callose deposition suppresses the recognition ofinvading pathogen and the initiation of defence reactions. It hasalso been suggested that callosemight be involved in the protectionof the invading fungus against the antimicrobial compoundsproduced by the plant [40]. The results obtained in this study,seems to confirm the involvement of this compound in defence

IwonaA. _Zur et al. / Physiological and Molecular Plant Pathology 76 (2011) 189e196196
reactions as all the rye genotypes in which callose deposition wasdetected demonstrated the highest regrowth ability after fungusinfection.
Summarizing, the received results have shed a little light on themechanism of defence induced in rye plants by M. nivale infection.They suggest the important role of phenolics and callose in thehostepathogen interactions. However, more precise analysis of theM. nivale colonization method is necessary. The electron micros-copy should confirm if observed by us vesicles are indeed typical forhaustoria produced by hemibiotrophs.
Acknowledgement
This research was supported by project 592/N-COST/2009/0 ‘Physiological and biochemical indicators of winter rye (Secalecereale L.) resistance to pink snow mould caused by Microdochiumnivale’, under Action COST FA0604 ‘Triticeace genomics for theadvancement of essential European crops’ (TritiGen).
References
[1] Schlegel R. Plant breeding. Updates Version 11.10. New Delhi, Mumbai,Chennai, Kolkata, Bangalore: BOD, Viva Books Private Limited; 2010.
[2] McBeath JH. Snow mold-plant-antagonist interactions: survival of the fittestunder the snow. Plant Health Instructor; 2002. doi:10.1094/PHI-I-2002-1010-01.
[3] Domsch KH, Gams W, Anderson TH. Monographella Petr. 1924. In: Compen-dium of soil fungi, vol. 1. Academic Press; 1980. pp. 428e30.
[4] Smith JD. Snow molds of winter cereals: guide for diagnosis, culture, andpathogenicity. Can J Plant Pathol 1981;3:15e25.
[5] Tronsmo AM, Hsianh T, Okuyama H, Nakajama T. Low temperature diseasescaused by Microdochium nivale. In: Iriki N, Gaudet DA, Tronsmo AM,Matsumoto N, Yoshida M, Nishimune, editors. Low temperature plantemic-robe interactions under snow. Sapporo: Hokkaido National AgriculturalExperimental Station; 2001. p. 75e82.
[6] Dubas E, Go1ebiowska G, _Zur I, Wedzony M. Microdochium nivale (Fr., Samuels& Hallett): cytological analysis of the infection process in triticale (x Tritico-secale Wittm.). Acta Physiol Plant; 2010. doi:10.1007/s11738-010-0576-9.
[7] Ǻrsvoll K. Effects of hardening, plant age, and development in Phleum pra-taense and Festuca pratensis on resistance to snow mould fungi. Sci Rep AgricUniv Norway 1977;56:1e13.
[8] Gaudet DA, Bhalla MK, Clayton GC, Chen THH. Effect of cottony snow mouldand low temperatures on winter wheat survival in central and northernAlberta. Can J Plant Pathol 1989;11:291e6.
[9] Gaudet DA. Progress towards understanding interactions between coldhardiness and snow mold resistance and development of resistant cultivars.Can J Plant Pathol 1994;16:241e6.
[10] Nakajima T, Abe J. Environmental factors affecting expression of resistance topink snow mould caused by Microdochium nivale in winter wheat. Can J Bot1996;74:1783e8.
[11] Tronsmo AM. Resistance to winter stress factors in half-sib families of Dactylisglomerata, tested in a controlled environment. Acta Agric Scand 1993;43:89e96.
[12] Bowler C, Fluhr R. The role of calcium and activated oxygens as signals forcontrolling cross-tolerance. Trends Plant Sci 2000;5:241e6.
[13] Tronsmo AM. Resistance to the rust fungus Puccinia poae-nemoralis in Poapratensis induced by low-temperature hardening. Can J Bot 1984;62:2891e2.
[14] White N, Jenkyn JF. Effects of sowing date and vernalisation on the growth ofwinter barley and its resistance to powdery mildew (Erysiphe graminis f. sp.hordei). Ann Appl Biol 1995;126:269e83.
[15] Dixon RA, Paiva NL. Stress-induced phenylpropanoid metabolism. Plant Cell1995;7:1085e97.
[16] Graham TL, Graham MY. Signaling in soybean phenylpropanoid responses,dissection of primary secondary and conditioning effect of light woundingand elicitor treatments. Plant Physiol 1996;110:1123e33.
[17] Pro�nczuk M, Madej L, Kolasi�nska I. Research for resistance to Microdochiumnivale among inbred lines of rye. Plant Breed Seed Sci 2003;48(2):83e6.
[18] O’Brien TP, McCully ME, editors. The study of plant structure. Principles andselected methods. Australia: Termarcarphi Pty. Ltd. Melbourne Press; 1981.
[19] M1odzianowski F, Wo�zny A. Wyk1ady i �cwiczenia z biologii komórki ro�slinnej.Wydawnictwo Naukowe Uniwersytetu im. A. Mickiewicza, Pozna�n, wyd. III;1990, p. 344.
[20] Singleton VS, Rossi Jr JA. Colorimetry of total phenolics with phosphomo-lybdicphosphotungstic acid reagent. Am J Enol Viticult 1965;16:144e57.
[21] Efremova TT, Laikova LI, Arbuzova VS. Popova OMEffect of the 5R(5A) AlienChromosome Substitution on the Growth Habit and Winter Hardiness ofWheat. Rus J Gen 2004;40(7):810e2.
[22] Tronsmo AM. Predisposing effects of low temperature on resistance to winterstress factors in grasses. Acta Agric Scand 1984;34:210e20.
[23] Ergon A, Klemsdal SS, Tronsmo AM. Interactions between cold hardening ofMicrodochium nivale infection on expression of pathogenesis-related genesin winter wheat. Physiol Mol Plant Pathol 1998;53:301e10.
[24] Hofgaard IS, Browne RA, Mascher F, Golebiowska G. Components of partialdisease resistance in wheat detected in a detached leaf assay inoculated withMicrodochium majus using first, second and third expanding seedling leaves.J Phytopathol 2006;154:204e8.
[25] Guy CL. Cold acclimation and freezing stress tolerance: role of proteinmetabolism. Annu Rev Plant Physiol Plant Mol Biol 1990;41:187e223.
[26] Rumbolz J, Kassemeyer HH, Steinmetz V, Deising HB, Mendgen K, Mathys D,et al. Differentiation of infection structures of the powdery mildew fungusUnicinula necator and adhesion to the host cuticle. Can J Bot 2000;78(3):409e21.
[27] Mendgen KW, Hahn M. Plant infection and the establishment of fungal bio-trophy. Trends Plant Sci 2002;7(8):352e6.
[28] Perfect SE, Green JR. Infection structures of biotrophic and hemibiotrophicfungal plant pathogens. Mol Plant Pathol 2001;2:101e8.
[29] Parry DW, Pegg GF. Surface colonization, penetration and growth of threeFusarium species in lucerne. Trans Br Mycol Soc 1986;85(3):495e500.
[30] Aloni R, Griffith M. Functional xylem anatomy in root-shoot junctions of sixcereal species. Planta 1991;184(1):123e9.
[31] Dubas E, Marzec K, P1a _zek A. Cytological studies on Microdochium nivaleinfection in Lolium perenne (L.) plants. Zeszyty Problemowe Postepów NaukRolniczych PAN 2010;545:389e401.
[32] Pociecha E, P1a _zek A, Janowiak F, Janeczko A, Zwierzykowski Z. Physiologicalbasis for differences in resistance to Microdochium nivale (Fr.) Samuels andHallett in two androgenic Genotypes of festulolium derived from tetraploid F1hybrids of Festuca pratensis � Lolium multiflorum (Festulolium) 2008;156(3):155e63.
[33] Pociecha E, P1a _zek A, Rapacz M, Niemczyk E, Zwierzykowski Z. Photosyntheticactivity and soluble carbohydrate content induced by the cold acclimationeffect frost tolerance and resistance to Microdochium nivale of androgenicFestulolium genotypes. J Agron Crop Sci 2010;196(1):48e54.
[34] Hückelhoven R. Cell wall-associated mechanisms of disease resistance andsusceptibility. Annu Rev Phytopathol 2007;45:101e27.
[35] Samapundo S, De Meulenaer B, Osei-Nimoh D, Lamboni Y, Debevere J,Devlieghere F. Can phenolic compounds be used for the protection of cornfrom fungal invasion and mycotoxin contamination during storage? FoodMicrobiol 2007;24(5):465e73.
[36] Bechinger C, Giebel KF, Schnell M, Leiderer P, Deising HB, Bastmeyer M.Optical measurements of invasive forces exerted by appressoria of a plantpathogenic fungus. Science 1999;285:1896e9.
[37] Peltonen S, Mannonen L, Karjalainen R. Elicitor-induced changes of phenyl-alanine ammonia-lyase activity in barley cell suspension cultures. Plant CellOrg Cult 1997;50:185e93.
[39] Ganesan V, Thomas G. Salicylic acid response in rice: influence ofsalicylic acid on H2O2 accumulation in oxidative stress. Plant Sci 2001;160:1095e106.
[40] Schulze-Lefert P. Knocking on the heaven’s wall: pathogenesis of andresistance to biotrophic fungi at the cell wall. Curr Opin Plant Biol 2004;7:377e83.
[41] Trillas M, Cotxarrera L, Casanova E, Cortadellas N. Ultrastructural changes andlocalization of chitin and callose in compatible and incompatible interactionsbetween carnation callus and Fusarium oxysporum. Physiol Mol Plant Pathol2000;56:107e16.
[42] Dong X, Hong Z,$Chatterjee J, Kim S, Verma DPS. Expression of callose syn-thase genes and its connection with Npr1 signaling pathway during pathogeninfection. Planta 2008;229:87e98.
[43] Brown I, Trethowan J, Kerry M, Mansfield J, Bolwell GP. Localization ofcomponents of the oxidative cross-linking of glycoproteins and of callosesynthesis in papillae formed during the interaction between non-pathogenicstrains of Xanthomonas campestris and French bean mesophyll cells. Plant J1998;15:333e43.
[44] Hématy K, Cherk C, Somerville S. Hostepathogen warfare at the plant cellwall. Curr Opin Plant Biol 2009;12:406e13.
[45] Jacobs AK, Lipka V, Burton RA, Panstruga R, Strizhov N, Schulze-Lefert P, et al.An Arabidopsis callose synthase, GSL5, is required for wound and papillarycallose formation. Plant Cell 2003;15:2503e13.
[46] Nishimura MT, Stein M, Hou BH, Vogel JP, Edwards H, Somerville SC. Loss ofa callose synthase results in salicylic acid dependent disease resistance.Science 2003;301:969e72.
[47] Gomez-Gomez L, Boller T. Flagellin perception: a paradigm for innateimmunity. Trends Plant Sci 2002;7:251e6.
[48] Vogel J, Somerville S. Isolation and characterization of powdery mildewresistant Arabidopsis mutants. Proc Natl Acad Sci USA 2000;97(4):1897e902.