
Enhancement of alkalinity tolerance in two cucumber genotypes inoculated with an arbuscular...
11
ORIGINAL PAPER Enhancement of alkalinity tolerance in two cucumber genotypes inoculated with an arbuscular mycorrhizal biofertilizer containing Glomus intraradices Youssef Rouphael & Mariateresa Cardarelli & Elena Di Mattia & Monica Tullio & Elvira Rea & Giuseppe Colla Received: 18 December 2009 / Revised: 2 March 2010 / Accepted: 4 March 2010 # Springer-Verlag 2010 Abstract The aim of the present study was to determine whether arbuscular mycorrhizal (AM) inoculation with a biofertilizer containing clays as granular carriers, leek root pieces and Glomus intraradices spores could improve alkalinity tolerance of two cucumber genotypes, and to study the changes induced by AM at agronomical and physiological level. A greenhouse experiment was carried out to determine yield, growth, fruit quality, net photosyn- thesis (A CO2 ), electrolyte leakage, and mineral composition of two cucumber (Cucumis sativus L.) genotypes (hybrid “Ekron” or open-pollinated variety “Marketmore”) with inoculated and noninoculated arbuscular mycorrhizal bio- fertilizer. Plants were supplied with nutrient solutions at two pH values (6.0 or 8.1). The high pH nutrient solution had the same basic composition with an additional 10 mM NaHCO 3 and 0.5 gl -1 CaCO 3 . The percentage root colonization was higher in “Marketmore” (21.8%) than “Ekron” (12.7%). Total and marketable yield and total biomass were significantly higher by 189%, 213%, and 77%, respectively, with Ekron in comparison to those recorded with Marketmore. The highest crop performance with Ekron in comparison to Marketmore was due to the improved nutritional status (higher N, P, K, Ca, Mg, Fe, Mn, and B), higher leaf area, and net photosynthesis. Increasing the concentration of NaHCO 3 from 0 to 10 mM in the nutrient solution significantly decreased yield, plant growth, A CO2 , N, P, Fe, Cu, Zn, Mn, and B concentration in leaf tissue, whereas the electrolyte leakage increased. The inoculated plants under alkaline conditions had higher total, marketable yield, and total biomass than noninoculated plant. Mycorrhizal cucumber plants grown under alkaline conditions had a higher macronutrient concentration in leaf tissue compared to noninoculated plants. The highest yield and biomass production in inoculated plants seems to be related to the capacity of maintaining higher net A CO2 and to a better nutritional status (high P, K, Mg, Fe, Zn, and Mn and low Na accumulation) in response to bicarbonate stress with respect to -AM plants. Keywords Arbuscular mycorrhizal . Cucumis sativus L. . Genotypes . Nutritional status . Net photosynthesis . pH level Introduction In the Mediterranean basin, alkaline soils are widely represented throughout the cropping regions, particularly those with semiarid climates (Troeh and Thompson 2005). Y. Rouphael Department of Crop Production, Faculty of Agricultural Engineering and Veterinary Medicine, Lebanese University, Dekwaneh-El Maten, Lebanon M. Cardarelli : G. Colla (*) Dipartimento di Geologia e Ingegneria Meccanica, Naturalistica e Idraulica per il Territorio, Università della Tuscia, via S. C. De Lellis snc, 01100 Viterbo, Italy e-mail: [email protected] E. Di Mattia Dipartimento di Agrobiologia e Agrochimica, Università della Tuscia, via S. C. De Lellis snc, 01100 Viterbo, Italy M. Tullio : E. Rea CRA-Centro di ricerca per lo studio delle relazioni tra pianta e suolo, Via della Navicella 2-4, 00184 Roma, Italy Biol Fertil Soils DOI 10.1007/s00374-010-0457-9
-
Upload
xn--2tesisdoctoralhistoriadelcinelaautoficcinenelcine-10f -
Category
Documents
-
view
3 -
download
0
Transcript of Enhancement of alkalinity tolerance in two cucumber genotypes inoculated with an arbuscular...

ORIGINAL PAPER
Enhancement of alkalinity tolerance in two cucumbergenotypes inoculated with an arbuscular mycorrhizalbiofertilizer containing Glomus intraradices
Youssef Rouphael & Mariateresa Cardarelli &Elena Di Mattia & Monica Tullio & Elvira Rea &
Giuseppe Colla
Received: 18 December 2009 /Revised: 2 March 2010 /Accepted: 4 March 2010# Springer-Verlag 2010
Abstract The aim of the present study was to determinewhether arbuscular mycorrhizal (AM) inoculation with abiofertilizer containing clays as granular carriers, leek rootpieces and Glomus intraradices spores could improvealkalinity tolerance of two cucumber genotypes, and tostudy the changes induced by AM at agronomical andphysiological level. A greenhouse experiment was carriedout to determine yield, growth, fruit quality, net photosyn-thesis (ACO2), electrolyte leakage, and mineral compositionof two cucumber (Cucumis sativus L.) genotypes (hybrid“Ekron” or open-pollinated variety “Marketmore”) withinoculated and noninoculated arbuscular mycorrhizal bio-fertilizer. Plants were supplied with nutrient solutions attwo pH values (6.0 or 8.1). The high pH nutrient solutionhad the same basic composition with an additional 10 mM
NaHCO3 and 0.5 gl−1 CaCO3. The percentage rootcolonization was higher in “Marketmore” (21.8%) than“Ekron” (12.7%). Total and marketable yield and totalbiomass were significantly higher by 189%, 213%, and77%, respectively, with Ekron in comparison to thoserecorded with Marketmore. The highest crop performancewith Ekron in comparison to Marketmore was due to theimproved nutritional status (higher N, P, K, Ca, Mg, Fe,Mn, and B), higher leaf area, and net photosynthesis.Increasing the concentration of NaHCO3 from 0 to 10 mMin the nutrient solution significantly decreased yield, plantgrowth, ACO2, N, P, Fe, Cu, Zn, Mn, and B concentration inleaf tissue, whereas the electrolyte leakage increased. Theinoculated plants under alkaline conditions had higher total,marketable yield, and total biomass than noninoculatedplant. Mycorrhizal cucumber plants grown under alkalineconditions had a higher macronutrient concentration in leaftissue compared to noninoculated plants. The highest yieldand biomass production in inoculated plants seems to berelated to the capacity of maintaining higher net ACO2 andto a better nutritional status (high P, K, Mg, Fe, Zn, and Mnand low Na accumulation) in response to bicarbonate stresswith respect to −AM plants.
Keywords Arbuscular mycorrhizal .Cucumis sativus L. .
Genotypes . Nutritional status . Net photosynthesis .
pH level
Introduction
In the Mediterranean basin, alkaline soils are widelyrepresented throughout the cropping regions, particularlythose with semiarid climates (Troeh and Thompson 2005).
Y. RouphaelDepartment of Crop Production, Faculty of AgriculturalEngineering and Veterinary Medicine, Lebanese University,Dekwaneh-El Maten, Lebanon
M. Cardarelli :G. Colla (*)Dipartimento di Geologia e Ingegneria Meccanica,Naturalistica e Idraulica per il Territorio, Università della Tuscia,via S. C. De Lellis snc,01100 Viterbo, Italye-mail: [email protected]
E. Di MattiaDipartimento di Agrobiologia e Agrochimica,Università della Tuscia,via S. C. De Lellis snc,01100 Viterbo, Italy
M. Tullio : E. ReaCRA-Centro di ricerca per lo studio dellerelazioni tra pianta e suolo,Via della Navicella 2-4,00184 Roma, Italy
Biol Fertil SoilsDOI 10.1007/s00374-010-0457-9

Alkaline soils are generally characterized by low bioavail-ability of plant nutrients, high concentrations of CaCO3 andsoil solution HCO3
−, high pH, and almost no exchangeableH+ (Marschner 1995; Misra and Tyler 1999). Bicarbonateions interfere with the uptake of macroelements, inparticular P, K, and Mg (Pissaloux et al. 1995). For instancein alkaline soils, P is largely unavailable to plants due to theformation of insoluble phosphate (e.g., Ca–P and Mg–P).Moreover, the concentration of HCO3
− interacts stronglywith the availability of several micro ions, especially Fe2+,and it is often considered to be the primary factorresponsible for chlorosis of plants on calcareous soils(Mengel 1994; Nikolic and Kastori 2000) leading to seriousyield and quality losses (Valdez-Aguilar and Reed 2006,2007, 2008). Reduction in iron availability is due to theincapacity of sensitive plants to acquire and to transportiron towards shoots (Reynier 1997). Iron deficiency reflectsupon the physiology and biochemistry of the whole plant,as iron is an important cofactor of many enzymes, includingthose involved in the biosynthetic pathway of chlorophylls(Marschner 1995). Thus, under iron deficiency conditions,the reduction in leaf iron concentration is often accompa-nied by a marked reduction of chlorophyll levels(Gogorcena et al. 2004; Molassiotis et al. 2006), by asignificant, although less intense, decrease in the chloro-phyll fluorescence (Morales et al. 1991; Nedunchezhian etal. 1997), and by a reduction in photosynthesis (Marschner1995; Molassiotis et al. 2006).
One way to avoid or reduce losses in production causedby alkalinity in high yielding genotypes would be toinoculate them with arbuscular mycorrhizal fungi capableof reducing the detrimental effect of external pH on cropperformance. Arbuscular mycorrhizal fungi are the mostwidespread root fungal symbionts and are associated withthe vast majority of higher plants (Sensoy et al. 2007).Arbuscular mycorrhizal fungi have been shown to improvesoil structure (Miller and Jastrow 2000), enhance plantnutrient acquisition (P, N, Zn, Cu, and Fe; Clark and Zeto2000), overcome the detrimental effect of salinity (Colla etal. 2008), improve drought tolerance (Bryla and Duniway1997; Sanchez-Blanco et al. 2004), suppresses root knotnematode (Zhang et al. 2008), and alleviate cultural andenvironmental stresses (Jeffries et al. 2003) through greatereffective root area and penetration of substrate (s) andactivation and excretion of various enzymes by infectedarbuscular mycorrhizal fungi roots and/or hyphae(Marschner 1995; Smith and Read 1997). However, thebenefits of arbuscular mycorrhizal fungi inoculation dependon the specific host–fungus combinations and also on thetypes of the inoculums used. Enhancing the tolerance ofplants to alkalinity when inoculated with AM fungi hasbeen studied and proposed until now for ornamental cropproduction such as vinca and roses (Cartmill et al. 2007,
2008), whereas no published data is available on the effectof arbuscular mycorrhizal fungi inoculation under high pHconditions on the agronomical and physiological responsesof an important vegetable crop widely grown in theMediterranean region, such as cucumber.
Differences in mycorrhizal responsiveness betweendifferent crops and between different genotypes within thesame crop have also been demonstrated (Declerck et al.1995; Parke and Kaeppler 2000; Linderman and Davis2004). In wheat, growth of modern cultivars tended tobenefit less from mycorrhizal inoculation than oldercultivars (Hetrick and Wilson 1992). Similarly, improvedsoybean cultivars showed a lower growth response tocolonization than older cultivars (Khalil et al. 1994). Noconsistent trend has been observed among corn cultivars(Toth et al. 1990; Khalil et al. 1994). Cultivated tomatoesand oats, however, are more responsive to mycorrhizalcolonization than closely related wild species (Koide et al.1988; Byrla and Koide 1990).
We hypothesize that arbuscular mycorrhizal inoculationwith a biofertilizer containing Glomus intraradices wouldgive an advantage to overcome alkalinity problems. Toverify this hypothesis, two cucumber genotypes (hybrid oropen-pollinated variety) were inoculated and their yield,growth, fruit quality, net photosynthesis, electrolyte leak-age, and mineral composition were compared with those ofnoninoculated plants when both were grown in sand cultureat two pH values in the nutrient solution (6.0 or 8.1).
Materials and methods
Plant material, growth conditions, treatments,and AM inoculation
The experiment was conducted in 2008 growing season in a400-m2 polyethylene greenhouse situated on the Experi-mental Farm of Tuscia University, Central Italy (42°25′N,12°08′E). Plants were grown under natural light conditions.The greenhouse was maintained at daily temperaturesbetween 18°C and 33°C and day/night relative humiditiesof 55%/85%, respectively.
The experimental plan was based on two pH levels ofthe nutrient solution (6.0 or 8.1), two mycorrhizal treat-ments (with or without AM) and two cucumber genotypes(hybrid or open pollinated variety). Treatments werearranged in a randomized complete block design with fourreplicates. Each experimental unit consisted of six plants.Cucumis sativus L. hybrid Ekron (Enza Zaden, Verona,Italy) and the open-pollinated variety Marketmore (SocietàAgricola Italiana Sementi, Cesena, Italy) were used.
Seeds were germinated in a 2:1 (v/v) mixture of BRILL-Type 3 propagation substrate (Gebr. Brill Substrate GmbH
Biol Fertil Soils

& Co. KG, Georgsdorf, Germany) and quartziferous sandon September 3, 2008. Prior to sowing, half of the potswere inoculated with G. intraradices (Aegis Argilla,Italpollina S.p.A., Rivoli Veronese, Italy) by mixing 1 m3
of the substrate with 15 l of inoculum mixture containing95% of clay minerals as granular carriers, root fragments,and spores (100 spores ml−1). The calcined clay (particulesize average 5 mm) is an attapulgite used as substrate forpropagation of AM fungi (Plenchette et al. 1996). Thecommercial inoculum was originally cultured in leek roots(Allium porrum L.) as reported by Calvet et al. (2001).However, because of the lack of information concerning themolecular characterization of Glomus contained in thebiofertilizer, it may be possible that with genetic recombi-nation and anastomoses compatibility between isolates ofAM, this specie might not be G. intraradices but a cohortof related species (Stockinger et al. 2009).
The seedlings were transplanted 21 days (September 24)after sowing, at the two true-leaf stages into pots (d 30 cm,h 30 cm) containing 17.7 l of quartziferous sand mixed with5 kg m−3 of a slow release fertilizer (Nitrophoska Top 15%N, 4.4% P, 12.4% K, 1.2% Mg, 4.8% S, 0.02% B, and0.01% Zn—BASF S.P.A., Milano, Italy). Pots weredisposed in double rows on the greenhouse floor. Plantrows were 1.4 m apart, and the space between plants withina row was 0.4 m. The distance between the centers ofdouble rows was 2.0 m, resulting in a plant density of 2.5plants per square meter. Plants were grown as verticalcordons and trained to the “umbrella” system where alllaterals branches were pruned back as they developed untilthe plant reached the overhead support wire. There, theterminal bud was removed above the support cable and twolateral branches were trained over the cable and allowed togrow back downward (Hickman 1998).
Nutrient solution management
The basic nutrient solution in this experiment was amodified Hoagland and Arnon formulation. All chemicalsused were of analytical grade, and composition of thenutrient solution was: 7.0 mM N–NO3
−, 1.0 mM S, 0.7 mMP, 2.5 mM K, 2.9 mM Ca, 0.7 mM Mg, 20 µM Fe, 9 µMMn, 0.3 µM Cu, 1.6 µM Zn, 20 µM B, and 0.3 µM Mo.Nutrient solution was applied starting from October 6(14 days after transplanting). The pH of the nutrientsolution was 6.0. The high pH treatment had the samenutrient composition plus 10 mM NaHCO3 and 0.5 gl−1
CaCO3. The sodium bicarbonate and CaCO3 were added tothe basic nutrient solution to mimic the effects of alkalinity.The pH of the bicarbonate treatment was 8.1. The alkalinetreatment was begun 7 days after transplanting.
Nutrient solution was pumped from independent tanksthrough a drip-irrigation system, with one emitter per plant
at a flow rate of 2 lh−1. Irrigation scheduling was performedusing electronic low-tension tensiometers (LT-Irrometer;Riverside, CA, USA) controlled by the substrate matricpotential (Norrie et al. 1994). In each treatment, fourtensiometers were installed and located in different pots toprovide representative readings of the moisture tension.Tensiometers were connected to an electronic programmerthat controlled the beginning (−5 kPa) and the end (−1 kPa)of irrigation, which correspond to the high and low tensionset points for the major part of the substrates (Kiehl et al.1992). Five to 15 fertigations were applied per day, each of1–3 min duration. The duration of irrigations was increasedso as to drain 35% of the nutrient solution from the pots.
Yield and biomass measurements
Fresh weight of marketable fruit and the number of fruitswere recorded three times per week on six plants per plotfrom November 4 to December 9. Fruit were harvestedwhen they reached marketable size, and fruit that weredeformed were considered unmarketable. At final harvest(December 9, 76 days after transplanting), six plants perplot were separated into stems, leaves, and roots. Rootswere rinsed with water to separate them from sand particlesand subsamples were analyzed to evaluate the AM fungiroot colonization. All plant tissues were dried in a forced airoven at 80°C for 72 h for biomass determination. Shootbiomass was the sum of leaves and stems. Root to shootratio was calculated by dividing root dry weight by the sumof leaf and stem dry weights. Leaf area (LA) was measuredwith an electronic area meter (Delta-T Devices Ltd, Cam-bridge, UK). Specific leaf area (SLA) was calculated bydividing leaf area by leaf dry weight of each plant.
Fruit quality analysis
Ten representative fruits of each plot were analyzed for qualityparameters. The liquid extract obtained from liquefying andfiltering the mesocarp of each fruit was analyzed for the totalsoluble solids (TSS) content by an Atago N1 refractometer(Atago Co. Ltd., Japan) and expressed as ºBrix at 20°C.Acidity was determined by potentiometric titration with 0.1MNaOH up to pH8.1 using 10 ml of juice. Results wereexpressed as percentage citric acid in the juice. The pH of thefruit juice was measured with a pH meter (HI-9023; HannaInstruments, Padova, Italy). Fruits were dried in a forced airoven at 80°C for 72 h and weighed to determine the fruit drymatter (DM).
AM fungi root colonization
At the end of the experiment (December 9, 76 days aftertransplanting), the root colonization by arbuscular mycor-
Biol Fertil Soils

rhizal fungi was determined on the same plants sampled forshoot and root measurements. Root samples were clearedwith 10% KOH, stained with 0.05% trypan blue inlactophenol as described by Phillips and Hayman (1970),and microscopically examined for AMF colonization bydetermining percentage of root segments containingarbuscules + vesicles using a gridline intercept method(Giovannetti and Mosse 1980).
Net photosynthesis measurement and electrolyte leakagedetermination
At the end of the experiment, the net assimilation of CO2
(ACO2, µmol CO2m−2s−1) was determined with a portable
photosynthesis system (LI-6200; LI-COR Inc., Lincoln,NE, USA). This measurement was made on the most recentfully expanded leaves, using six replicate leaves pertreatment. The LI-6200 was equipped with a well-stirred2.5×10−5m3 leaf chamber with constant-area inserts (1.2×10−3m2) and fitted with a variable intensity red source(Model QB1205LI-670, Quantum Devices Inc. Barneveld,WI, USA; Tennessen et al. 1994). Leaf temperature withinthe chamber was 30±2°C, vapor pressure differencebetween the leaf and air was 2.6±0.3°C, and CO2
concentration was 365±10 μl l−1. Net photosynthesismeasurement was made between 0900 and 1200 hours.
Electrolyte leakage (EL) was determined as described byLutts et al. (1995). Six randomly chosen plants pertreatment (four mature leaves per plant) were taken andcut into 1-cm segments. Leaf samples were washed threetimes with distilled water to remove surface contaminationand then placed in individual stoppered vials containing10 ml of distilled water. The samples were incubated atroom temperature (25°C) on a shaker (100 rpm) for 24 h.Electrical conductivity of the bathing solution (EC1) wasread after incubation. The same samples were then placedin an autoclave at 120°C for 20 min and a second readingof the EC (EC2) was made after cooling the solution toroom temperature. The EL was calculated as EC1/EC2 andexpressed as percentage.
Mineral analysis
Dried plant tissues (leaf and fruits) were ground in a Wileymill to pass through a 20-mesh screen and 0.5 g of the driedtissues was analyzed for the contents of the following: N, P,K, Ca, Mg, Na, Fe, Cu, Zn, Mn, and B. The Nconcentration of leaf and fruit tissues was determined aftermineralization with sulfuric acid by “Regular Kjeldahlmethod” (Bremner 1965) whereas P, K, Ca, Mg, Na, Fe,Cu, Zn, Mn, and B concentrations were determined by dryashing at 400°C for 24 h, dissolving the ash in HNO3 1:20v/v and assaying the solution obtained by an inductively
coupled plasma emission spectrophotometer (ICP Iris;Thermo Optek, Milano, Italy; Isaac and Johnson 1998).
Statistical analysis
All data were statistically analyzed by analysis of varianceusing the SPSS software package (SPSS 10 for Windows,2001). Duncan’s multiple range test was performed atP=0.05 on each of the significant variables measured.
Results
AM root colonization
The percentage root colonization at the end of the trial wassignificantly affected by genotype (G; P<0.001), mycorrhizaltreatment (M; P<0.001), pH (P<0.01), G×M (P<0.001),and pH × M (P<0.01) interactions. No AM fungi coloniza-tion was recorded in roots of control plants while ininoculated plants, the percentage of AM infection dependedon pH value of the different nutrient solution and on the plantgenotype. Irrespective of nutrient solution pH (plant geno-type × mycorrhizal interaction), the percentage root coloni-zation was higher in Marketmore genotype (21.8%) thanEkron genotype (12.7%). Moreover, when average over bothgenotypes (pH × mycorrhizal interaction), increasing the pHof nutrient solution from 6.0 to 8.1 decreased the coloniza-tion of cucumber roots (19.9% and 14.6%, respectively).
Fruit yield components
Total and marketable cucumber production and fruitnumber were significantly affected by genotype, mycor-rhizal treatment, and pH while the unmarketable yield wassignificantly influenced by the genotype and mycorrhizaltreatment, but not by pH, with no significant plantgenotype × mycorrhizal × pH interaction (Table 1). Totaland marketable yield decreased by 9% and 7%, respec-tively, in response to an increase of pH in the nutrientsolution, whereas the total and marketable yield increasedby 10% and 21%, respectively, when plants wereinoculated with live G. intraradices (Table 1). Irrespectiveof mycorrhizal and pH treatment the total and marketableyield were significantly higher by 189% and 213%,respectively, in the hybrid Ekron (average 2.20 and1.91 kg per plant, respectively) in comparison to thoserecorded with the open-pollinated variety Marketmore(average 0.76 and 0.61 kg per plant, respectively). Thelowest marketable yield observed with Marketmore incomparison to Ekron and with plants grown at high pHlevel in comparison to low pH level in the nutrientsolution was due to a reduction in the number of fruit
Biol Fertil Soils

per plant and not to the fruit mean weight, while thelowest marketable yield observed with noninoculated incomparison to inoculated plants was mainly attributed to areduction in both fruit weight and number of fruits perplant (Table 1).
Biomass production and partitioning
Similar to the fruit yield, leaves, fruit, and total dry weightwere significantly affected by plant genotype, mycorrhiza,
and pH, with no interaction of these three variables,whereas the stem, roots dry weight, and the harvest indexwere only influenced by genotype (Table 2). The leaves,fruits, and total dry weight were 17%, 13%, and 20%higher with plants grown at the low pH value (average50.0, 51.0, and 130.8 g per plant, respectively) than plantsgrown at the high pH value (average 42.6, 45.1, and109.3 g per plant, respectively), while the effect of themycorrhizal was more pronounced with an increase of24%, 17%, and 22%, respectively, of inoculated (average
Genotype AM pH Yield (kg per plant) Marketable fruit
T M NM Mean weight (g) Number (no. per plant)
Ekron −AM 6.0 2.25 1.81 0.44 238.0 7.5
8.1 1.92 1.66 0.26 242.4 6.8
+AM 6.0 2.40 2.12 0.28 257.2 8.3
8.1 2.24 2.06 0.18 263.1 7.8
Marketmore −AM 6.0 0.76 0.59 0.17 240.3 2.4
8.1 0.71 0.50 0.21 233.3 2.1
+AM 6.0 0.80 0.71 0.09 262.5 2.7
8.1 0.78 0.65 0.13 254.2 2.5
Significance
Genotype (G) *** *** * NS ***
Mycorrhizae (M) * ** * * *
pH * * NS NS *
G × M NS NS NS NS NS
G × pH NS NS NS NS NS
M × pH NS NS NS NS NS
G × M × pH NS NS NS NS NS
Table 1 Effects of genotype,arbuscular mycorrhizal (AM)inoculation and solution pHvalue on total (T), marketable(M), unmarketable (NM),marketable fruit mean weight,and number of cucumber plants
Values are the means of fourreplicate samples
+AM mycorrhizal plants, −AMnonmycorrhizal plants,NS nonsignificant
*P<0.05; **P<0.01; ***P<0.001 (significant)
Genotype AM pH Dry weight (g per plant) HI
Leaves Stems Fruits Roots Total
Ekron −AM 6.0 48.1 16.3 64.3 18.2 143.6 0.44
8.1 46.1 16.3 56.9 15.5 128.1 0.42
+AM 6.0 66.5 18.4 78.8 19.9 183.7 0.43
8.1 56.6 18.6 72.1 11.2 158.5 0.45
Marketmore −AM 6.0 41.5 15.0 29.6 8.2 93.6 0.31
8.1 29.7 14.4 26.2 5.4 67.6 0.35
+AM 6.0 43.8 15.2 31.2 8.1 102.4 0.32
8.1 38.1 14.3 25.3 5.5 83.1 0.30
Significance
Genotype (G) *** * *** ** *** ***
Mycorrhizae (M) ** NS * NS ** NS
pH * NS * NS ** NS
G × M NS NS NS NS NS NS
G × pH NS NS NS NS NS NS
M × pH NS NS NS NS NS NS
G × M × pH NS NS NS NS NS NS
Table 2 Effects of genotype,arbuscular mycorrhizal (AM)inoculation, and solution pHvalue on biomass production,partitioning, and harvest index(HI) of cucumber plants
Values are the means of fourreplicate samples
+AM mycorrhizal plants, −AMnonmycorrhizal plants, NS non-significant
*P<0.05; **P<0.01; ***P<0.001 (significant)
Biol Fertil Soils

51.2, 51.8, and 131.9 g per plant, respectively) thannoninoculated cucumber plants (average 41.3, 44.2, and108.2 g per plant, respectively; Table 2). Finally, the leaves,stems, fruits, total dry weight, and the harvest index weresignificantly higher by 42%, 18%, 142%, 77%, and 34%,respectively, in Ekron hybrid (average 54.3, 17.4, 68.0,153.5 g per plant, and 0.43, respectively) than in openpollinated variety Marketmore (average 38.3, 14.7, 28.1,86.7 g per plant, and 0.32, respectively).
Fruit quality
No significant difference among treatments was ob-served for the fruit acidity concentration (average0.081%). The fruit DM, the TSS contents, and the fruitpH were influenced by the genotype and were signifi-cantly higher by 12%, 8%, and 3%, respectively, in theopen-pollinated variety Marketmore (average 3.64%,3.50°brix, 5.61, respectively) than those recorded ofthe hybrid Ekron (average 3.26%, 3.23°brix, 5.44,respectively, Table 3). The fruit DM and TSS contentswere significantly affected by the pH level in the nutrientsolution. Increasing the concentration of NaHCO3 from 0to 10 mM in the nutrient solution significantly increasedthe DM and TSS contents by 4.4 (average 3.37% to3.52%) and 5.5% (3.27 to 3.45°brix), respectively(Table 3). The DM was significantly affected by mycor-rhizal treatment. Mycorrhizal colonization enhanced thefruit DM of cucumber fruits, with the highest valuesrecorded on inoculated (average 3.5%) compared tononinoculated plants (average 3.3%, Table 3).
Leaf area, electrolyte leakage, and net photosynthetic rate
Except the SLA, which was significantly affected bygenotype and mycorrhizal treatment, but not by the pH,the final LA and the net ACO2 were significantly influencedby genotype, mycorrhizal treatment, and pH, with no plantgenotype × mycorrhizal × pH interaction (Table 4).Increasing the concentration of NaHCO3 from 0 to10 mM in the nutrient solution decreased the LA andACO2 by 12% and 14%, respectively. Moreover, the finalLA and ACO2 increased by 12% and 7%, respectively,whereas the SLA decreased by 12% when plants wereinoculated with G. intraradices (Table 4). The final LA,SLA, and ACO2 were significantly higher by 34%, 18%,and 10%, respectively, in the hybrid Ekron (average1.05 m2 per plant, 20.8 m2kg−1, 16.3 µmol CO2m
−2s−1,respectively) than in the open-pollinated variety Market-more (average 0.79 m2 per plant, 17.6 m2kg−1, 14.8 µmolCO2m
−2s−1, respectively). Finally, the EL was significantly(P<0.05) affected by the pH level of the nutrient solution(data not shown). Electrolyte leakage increased by increas-ing the concentration of NaHCO3 from 0 to 10 mM, withthe highest values recorded on plants grown at higher pHvalue (average 29.8%) than plants grown at low pH value(average 19.9%).
Mineral composition
The major and trace element concentrations of cucumberleaves and fruits as a function of genotype, mycorrhizaltreatments, and pH are displayed in Tables 5 and 6. The
Genotype AM pH DM (%) TSS (°brix) pH TA (%)
Ekron −AM 6.0 3.15 3.20 5.45 0.082
8.1 3.25 3.25 5.44 0.080
+AM 6.0 3.29 3.23 5.41 0.087
8.1 3.36 3.25 5.46 0.080
Marketmore −AM 6.0 3.42 3.28 5.65 0.075
8.1 3.64 3.63 5.54 0.080
+AM 6.0 3.65 3.40 5.59 0.084
8.1 3.85 3.70 5.66 0.079
Significance
Genotype (G) *** *** *** NS
Mycorrhizae (M) * NS NS NS
pH * * NS NS
G × M NS NS NS NS
G × pH NS NS NS NS
M × pH NS NS NS NS
G × M × pH NS NS NS NS
Table 3 Effects of genotype,arbuscular mycorrhizal (AM)inoculation and solution pHvalue on fruit dry matter (DM),total soluble solids (TSS)contents, fruit juice pH, andtitratable acidity (TA) of fruitpulp of cucumber plants
Values are the means of fourreplicate samples
+AM mycorrhizal plants, −AMnonmycorrhizal plants,NS nonsignificant
*P<0.05; **P<0.01; ***P<0.001 (significant)
Biol Fertil Soils

concentrations of N, P, Fe, Cu, Zn, Mn, and B in leaf tissuewere negatively affected by increasing pH in the nutrientsolution (Table 5). Increasing the concentration of NaHCO3
from 0 to 10 mM in the nutrient solution significantlydecreased the N, P, Fe, Cu, Zn, Mn, and B concentration by8%, 25%, 39%, 6%, 54%, 42%, and 12%, respectively. Theconcentration of P, Mg, Fe, Zn, and Mn in leaves was
significantly higher by 14%, 3%, 14%, 10%, and 11% inleaves of inoculated (average 6.3 and10.3 mg g−1 DW and56.7, 31.8, and 129.1 µg g−1 DW, respectively) than inleaves of noninoculated cucumber (average 5.6 and10.0 mg g−1 DW and 49.9, 29.0, and 116.8 µg g−1 DW,respectively). Moreover, the concentration of N, P, Ca, Fe,Mn, and B was significantly affected by genotype with the
Genotype AM pH LA (m2 per plant) SLA (m2 kg−1) ACO2 (µmol CO2 m−2s−1)
Ekron −AM 6.0 1.07 22.74 17.1
8.1 0.93 21.93 14.1
+AM 6.0 1.15 19.73 17.6
8.1 1.06 18.84 16.3
Marketmore −AM 6.0 0.80 18.15 15.6
8.1 0.66 18.97 13.2
+AM 6.0 0.89 16.63 16.5
8.1 0.79 16.74 13.9
Significance
Genotype (G) *** * *
Mycorrhizae (M) * * *
pH ** NS **
G × M NS NS NS
G × pH NS NS NS
M × pH NS NS NS
G × M × pH NS NS NS
Table 4 Effects of genotype,arbuscular mycorrhizal (AM)inoculation and solution pHvalue on the final leaf area (LA),specific leaf dry weight (SLA),and net assimilation CO2 (ACO2)of cucumber plants
Values are the means of fourreplicate samples
+AM mycorrhizal plants, −AMnonmycorrhizal plants, NS non-significant
*P<0.05; **P<0.01; ***P<0.001 (significant)
Table 5 Effects of genotype, arbuscular mycorrhizal (AM) inoculation, and solution pH value on major and trace elements of cucumber leaves
Genotype AM pH Major elements (mg g−1 DW) Trace elements (µg g−1 DW)
N P K Ca Mg Na Fe Cu Zn Mn B
Ekron −AM 6.0 40.0 6.7 39.9 23.7 10.8 0.5 68.6 12.8 39.5 164.9 63.1
8.1 36.9 5.1 31.9 24.3 10.7 3.7 41.3 11.6 18.3 85.0 52.4
+AM 6.0 42.1 7.3 43.2 24.4 11.6 0.6 76.9 12.7 44.3 177.8 67.3
8.1 38.6 6.1 31.8 24.4 10.6 3.2 48.8 11.8 21.3 100.3 59.8
Marketmore −AM 6.0 39.4 6.4 28.2 21.0 9.3 0.4 56.1 12.3 40.9 136.6 50.4
8.1 35.9 4.0 25.6 21.4 9.2 3.6 33.7 11.9 17.2 80.7 44.3
+AM 6.0 39.5 6.6 33.1 22.1 9.5 0.5 62.9 13.1 42.2 145.0 55.3
8.1 36.7 5.0 28.0 21.1 9.3 3.1 38.2 12.5 19.4 93.3 50.2
Significance
Genotype (G) ** *** *** *** *** NS ** NS NS ** ***
Mycorrhizae (M) NS ** * NS * * * NS * * NS
pH *** *** *** NS NS *** *** ** *** *** **
G × M NS NS NS NS NS NS NS NS NS NS NS
G × pH NS NS ** NS NS NS NS NS NS NS NS
M × pH NS NS NS NS NS ** NS NS NS NS NS
G × M × pH NS NS NS NS NS NS NS NS NS NS NS
Values are the means of four replicate samples
+AM mycorrhizal plants, −AM nonmycorrhizal plants, NS nonsignificant
*P<0.05; **P<0.01; ***P<0.001 (significant)
Biol Fertil Soils
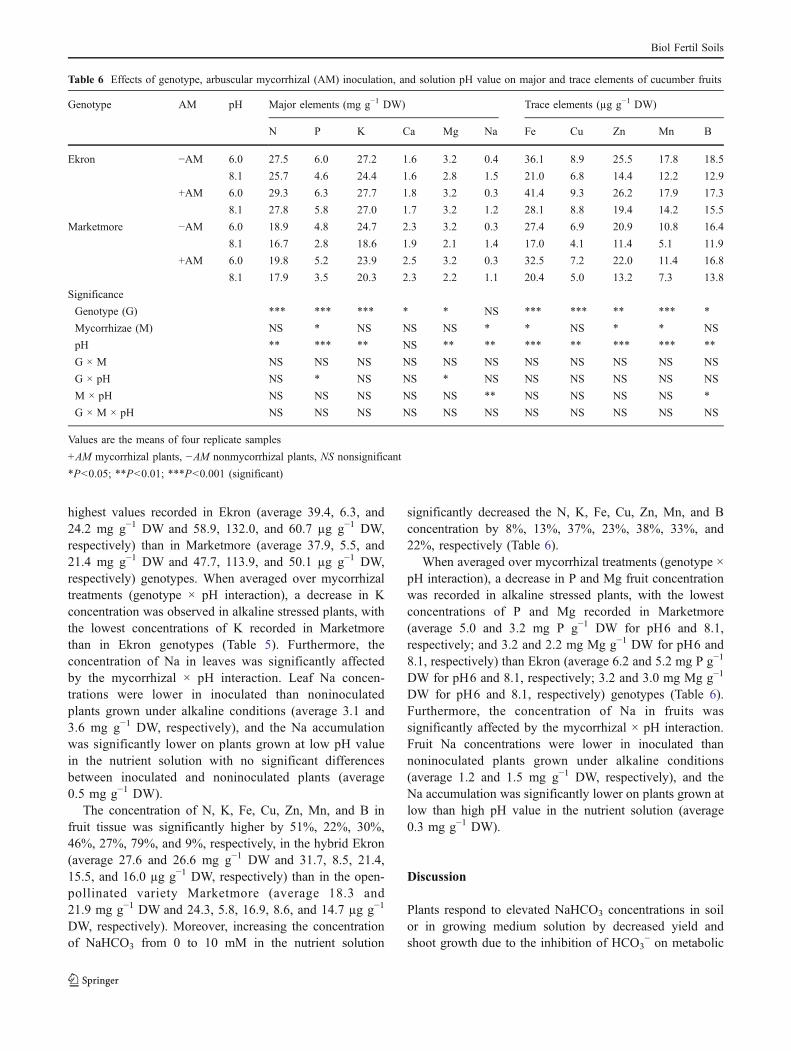
highest values recorded in Ekron (average 39.4, 6.3, and24.2 mg g−1 DW and 58.9, 132.0, and 60.7 µg g−1 DW,respectively) than in Marketmore (average 37.9, 5.5, and21.4 mg g−1 DW and 47.7, 113.9, and 50.1 µg g−1 DW,respectively) genotypes. When averaged over mycorrhizaltreatments (genotype × pH interaction), a decrease in Kconcentration was observed in alkaline stressed plants, withthe lowest concentrations of K recorded in Marketmorethan in Ekron genotypes (Table 5). Furthermore, theconcentration of Na in leaves was significantly affectedby the mycorrhizal × pH interaction. Leaf Na concen-trations were lower in inoculated than noninoculatedplants grown under alkaline conditions (average 3.1 and3.6 mg g−1 DW, respectively), and the Na accumulationwas significantly lower on plants grown at low pH valuein the nutrient solution with no significant differencesbetween inoculated and noninoculated plants (average0.5 mg g−1 DW).
The concentration of N, K, Fe, Cu, Zn, Mn, and B infruit tissue was significantly higher by 51%, 22%, 30%,46%, 27%, 79%, and 9%, respectively, in the hybrid Ekron(average 27.6 and 26.6 mg g−1 DW and 31.7, 8.5, 21.4,15.5, and 16.0 µg g−1 DW, respectively) than in the open-pollinated variety Marketmore (average 18.3 and21.9 mg g−1 DW and 24.3, 5.8, 16.9, 8.6, and 14.7 µg g−1
DW, respectively). Moreover, increasing the concentrationof NaHCO3 from 0 to 10 mM in the nutrient solution
significantly decreased the N, K, Fe, Cu, Zn, Mn, and Bconcentration by 8%, 13%, 37%, 23%, 38%, 33%, and22%, respectively (Table 6).
When averaged over mycorrhizal treatments (genotype ×pH interaction), a decrease in P and Mg fruit concentrationwas recorded in alkaline stressed plants, with the lowestconcentrations of P and Mg recorded in Marketmore(average 5.0 and 3.2 mg P g−1 DW for pH6 and 8.1,respectively; and 3.2 and 2.2 mg Mg g−1 DW for pH6 and8.1, respectively) than Ekron (average 6.2 and 5.2 mg P g−1
DW for pH6 and 8.1, respectively; 3.2 and 3.0 mg Mg g−1
DW for pH6 and 8.1, respectively) genotypes (Table 6).Furthermore, the concentration of Na in fruits wassignificantly affected by the mycorrhizal × pH interaction.Fruit Na concentrations were lower in inoculated thannoninoculated plants grown under alkaline conditions(average 1.2 and 1.5 mg g−1 DW, respectively), and theNa accumulation was significantly lower on plants grown atlow than high pH value in the nutrient solution (average0.3 mg g−1 DW).
Discussion
Plants respond to elevated NaHCO3 concentrations in soilor in growing medium solution by decreased yield andshoot growth due to the inhibition of HCO3
− on metabolic
Table 6 Effects of genotype, arbuscular mycorrhizal (AM) inoculation, and solution pH value on major and trace elements of cucumber fruits
Genotype AM pH Major elements (mg g−1 DW) Trace elements (µg g−1 DW)
N P K Ca Mg Na Fe Cu Zn Mn B
Ekron −AM 6.0 27.5 6.0 27.2 1.6 3.2 0.4 36.1 8.9 25.5 17.8 18.5
8.1 25.7 4.6 24.4 1.6 2.8 1.5 21.0 6.8 14.4 12.2 12.9
+AM 6.0 29.3 6.3 27.7 1.8 3.2 0.3 41.4 9.3 26.2 17.9 17.3
8.1 27.8 5.8 27.0 1.7 3.2 1.2 28.1 8.8 19.4 14.2 15.5
Marketmore −AM 6.0 18.9 4.8 24.7 2.3 3.2 0.3 27.4 6.9 20.9 10.8 16.4
8.1 16.7 2.8 18.6 1.9 2.1 1.4 17.0 4.1 11.4 5.1 11.9
+AM 6.0 19.8 5.2 23.9 2.5 3.2 0.3 32.5 7.2 22.0 11.4 16.8
8.1 17.9 3.5 20.3 2.3 2.2 1.1 20.4 5.0 13.2 7.3 13.8
Significance
Genotype (G) *** *** *** * * NS *** *** ** *** *
Mycorrhizae (M) NS * NS NS NS * * NS * * NS
pH ** *** ** NS ** ** *** ** *** *** **
G × M NS NS NS NS NS NS NS NS NS NS NS
G × pH NS * NS NS * NS NS NS NS NS NS
M × pH NS NS NS NS NS ** NS NS NS NS *
G × M × pH NS NS NS NS NS NS NS NS NS NS NS
Values are the means of four replicate samples
+AM mycorrhizal plants, −AM nonmycorrhizal plants, NS nonsignificant
*P<0.05; **P<0.01; ***P<0.001 (significant)
Biol Fertil Soils

processes and/or impairment of root activity/growth and/ornutrient solubility (Alhendawi et al. 1997; Campbell andNishio 2000; Zribi and Gharsalli 2002; Valdez-Aguilarand Reed 2006, 2007, 2008). Although plant species andcultivars may differ in their tolerance to HCO3
− stress,root physiology and nutrient solubility are affected by thebuffering capacity of HCO3
−, which is related to anincrease in pH. Similarly, in the present study, significantdepression in total and marketable yield and shootbiomass production (Tables 1 and 2) in bicarbonate-treated cucumber plants was observed and that effectvaried as a function of AM inoculation and genotypes.Under alkaline conditions, yield and biomass productionwere higher in inoculated than noninoculated plants.Alkalinity tolerance of cucumber plants inoculated withG. intraradices may be due to the better uptake andtranslocation of mineral elements in particular P, Mg, Fe,Zn, and Mn to the shoot. Arbuscular mycorrhizae canmitigate growth reduction by alkalinity (Cartmill et al.2007, 2008) but the mechanism involved remains unre-solved. Cartmill et al. (2007) concluded that the alkalinetolerance of Rosa multiflora is primarily related to theenhanced nutrient uptake (N, P, K, Ca, Fe, Zn, Al, and B),leaf chlorophyll concentration, low Fe reductase activity,and low soluble alkaline phosphatase activity. Similarly,Cartmill et al. (2008) stated that the tolerance of vincaplants inoculated with a mix (ZAC-19) of Glomus speciesisolate (Glomus albidum, Glomus claroideum, and Glomusdiaphanum) to alkalinity was associated with increased Puptake due to a higher soluble phosphatase activity atmoderate HCO3
− concentrations, with the maintenance ofother leaf nutrients when HCO3
− increased up to 10 mM,and with an increased of antioxidant activity, and thusdetoxifying activity, probably as a result of their enhancedmicronutrient status.
In many plant species, the high alkalinity induces Fedeficiency due to reduced Fe availability in soil or uptake(Alhendawi et al. 1997; Römheld 2000). Bicarbonate canalso precipitate Fe internally making this nutrient lessavailable in roots (Römheld 2000). We have shown that theincrease in the concentration of NaHCO3 from 0 to 10 mMin the nutrient solution significantly decreased the leaf Feconcentration by 39%, but inoculated plants accumulatedon average 14% more Fe than noninoculated plants(Table 5). This suggests that arbuscular mycorrhizalinoculation with a biofertilizer containing G. intraradicesenhance the uptake and translocation of Fe toward theshoot. The higher uptake and accumulation of Fe ininoculated than noninoculated cucumber plants was themain mechanism reducing the detrimental effect of alkalin-ity (Fe deficiency) on yield and plant growth.
Iron deficiency can cause various physiological changesin leaves, as iron is an important cofactor of many enzymes,
and interferes with several aspects of plant biochemistry,including photosynthesis and pigment synthesis (Marschner1995). The changes induced by the inoculation with G.intraradices in the shoot growth were also reflected atphysiological level, as shown by the LA, SLA, and ACO2
values of inoculated plants (Table 4). The highest yield andbiomass production in inoculated plants seems to be relatedto the capacity of maintaining higher net ACO2 in responseto bicarbonate stress with respect to noninoculated plants(Table 4). This indicates that inoculated plant had a higherphotosynthetic area per plant in relative terms, probably asa result of carbon cost necessary to maintain AM fungiassociations (Wright et al. 1998). The inhibitory effect ofroot zone alkalinity on photosynthesis has been observed inmany plant species (Iturbe-Ormaexte et al. 1995; Yakushijiet al. 1998); however, the exact mechanisms involved arenot clear (Molassiotis et al. 2006). In addition to reducednet photosynthetic rates, leaf area decreased in response toan increase of alkalinity in the nutrient solution especiallyin noninoculated plants. Restriction of leaf area may be theresult of the suppressed net photosynthetic rates since thelatter effect reduces the available assimilates for leafgrowth. A reduction in leaf area with excess NaHCO3 hasbeen reported for several greenhouse crops (chrysanthe-mum, mini-rose, vinca, and hibiscus) irrigated with alkalinewater (Valdez-Aguilar 2004).
Bicarbonate ions may interfere with essential nutrientuptake and transport and thereby can affect nutrientcomposition of plants (Marschner 1995). In the presentstudy, a significant reduction of macro- (N, P, and K) andmicroelement (Fe, Cu, Zn, Mn, and B) contents of leaf andfruit tissue of cucumber was found at high pH value(Tables 5 and 6). Decreased tissue P, K, and Fe concen-trations are typically observed in alkaline conditions(Pissaloux et al. 1995). Moreover, in terms of leaf nutrientconcentration, inoculation of cucumber plants with arbus-cular mycorrhizal fungi increased the P, K, Mg, Fe, Zn, andMn content (Table 5). Similar results have been reported fora number of species (Cartmill et al. 2007, 2008), and theyhave been attributed to a greater effective root area andpenetration of the substrate(s) provided by AM fungi ZAC-19, (composed of G. albidum, G. claroideum, and G.diaphanum) and to the activation and excretion of enzymesby colonized roots and/or hyphae (Marschner 1998; Clarkand Zeto 2000). In the present experiment, the plants weregrown in relatively small containers and probably rootgrowth was not important and thus the beneficial effect ofAM fungi may be due to enhanced nutrient acquisition andtransport to the plant rather than greater effective root area.
Data of this study also showed that Na concentration incucumber leaves was lower in inoculated than noninocu-lated plants grown under alkaline conditions (Table 5). Thismight indicate that Na might be retained in intraradical AM
Biol Fertil Soils

fungal hyphae or was compartmentalized in the root cellvacuoles without translocation to the shoots (Cantrell andLinderman 2001). However, Al-Karaki (2006) suggestedthat lower Na concentrations in inoculated than noninocu-lated tomato plants with Glomus mosseae might beexplained by dilution effects due to growth enhancementby AM fungi colonization.
Finally, we have shown a role of cucumber genotype inmycorrhizal colonization of the plant since the percentage rootcolonization was higher in open-pollinated variety Market-more (21.8%) than hybrid Ekron (12.7%). Our findings agreewith those of Linderman and Davis (2004) for marigolds,whereas growth of modern wheat cultivars tended to benefitless from mycorrhizal inoculation than older cultivars(Hetrick and Wilson 1992). Similarly, improved soybeancultivars showed a lower growth response to colonizationthan older cultivars (Khalil et al. 1994). The highest AM rootcolonization recorded in Marketmore did not improve theyield and biomass production since the total and marketableyield and total biomass were significantly higher by 189%,213%, and 77%, respectively, in the hybrid Ekron than thoserecorded with the open-pollinated variety Marketmore. Theabove findings confirmed the lack of correlation betweenmycorrhiza colonization and the growth response as reportedfor pepper (Sensoy et al. 2007). The highest crop perfor-mance with Ekron in comparison to Marketmore was due tothe improved nutritional status (higher N, P, K, Ca, Mg, Fe,Mn, and B), higher LA, and ACO2, which had promotedcucumber fruit production.
A previous study showed that the incidence of thebiofertilizer used in the present trial on the production costof mycorrhized transplants in 60-hole tray was very low(0.68 eurocent per plantlet, Tullio et al. unpublished data).Taking into consideration that the average price ofcucumber fruits during 2009 in Italy was 52 eurocent perkilogram (www.ismea.it) and that the mycorrhized plantsproduced 0.245 kg per plant more than the nonmycorrhizedplants, the extra profit was 13 eurocents per plant. From aneconomical point of view, the increasing of the cost due tothe application of the AM biofertilizer was largely coveredby the sale of the additional cucumber yield.
In conclusion, arbuscular mycorrhizal inoculation with abiofertilizer containing G. intraradices alleviate the detri-mental effect of alkalinity on growth and productivity byimproving the nutritional status (higher P, K, Mg, Fe, Zn,and Mn and lower Na concentration in leaf tissue) and thecapacity of maintaining higher ACO2 of the cucumberplants. Furthermore, cucumber fruit quality in particularfruit dry matter was improved by mycorrhizal colonizationbut these effects depended on the plant genotype. Finally,future studies in which a “non viable inoculum treatment”is included could clarify if any other component of thebiofertilizer affects the crop performance.
Acknowledgment We are grateful to Emanuele Mastronardi for thehelp during the carrying out of the experiment.
References
Alhendawi RA, Römheld V, Kirkby EA, Marschner H (1997)Influence of increasing bicarbonate concentrations on plantgrowth, organic acid accumulation in roots and iron uptake bybarley, sorghum and maize. J Plant Nutr 20:1731–1753
Al-Karaki GN (2006) Nursery inoculation of tomato with arbuscularmycorrhizal fungi and subsequent performance under irrigationwith saline water. Sci Hortic 109:1–7
Bremner JM (1965) Total nitrogen. In: Methods of Soil Analysis,Black CA, Evans DD, White IL, Ensminger LE, Clark FE(eds) Methods Agronomy Monograph 9. ASA, Madison, WI,pp 1149–1178
Bryla DR, Duniway JM (1997) Effects of mycorrhizal infection ondrought tolerance and recovery in safflower and wheat. Plant Soil197:95–103
Byrla DR, Koide RT (1990) Role of mycorrhizal infection in thegrowth and reproduction of wild vs cultivated plants. II. Eightwild accessions and two cultivars of Lycopresicum esculentumMill. Oecologia 77:537–543
Calvet C, Pinochet J, Hernandez-Dorrego A, Estaun V, Camprubi A(2001) Field microplot performance of the peach-almond hybridGF-677 after inoculation with arbuscular mycorrhizal fungi in areplant soil infested with root-knot nematodes. Mycorrhiza10:295–300
Campbell SA, Nishio JN (2000) Iron deficiency studies of sugar beetusing an improved sodium bicarbonate-buffered hydroponicgrowth system. J Plant Nutr 23:741–757
Cantrell IC, Linderman RG (2001) Preinoculation of lettuce and onionwith VA fungi reduces deleterious effects of soil salinity. PlantSoil 233:269–281
Cartmill AD, Alarcón A, Valdez-Aguilar LA (2007) Arbuscularmycorrhizal fungi enhance tolerance of Rosa multiflora cv.Burr to bicarbonate in irrigation water. J Plant Nutr 30:1517–1540
Cartmill AD, Valdez-Aguilar LA, Bryan DL, Alarcón A (2008)Arbuscular mycorrhizal fungi enhance tolerance of vinca to highalkalinity in irrigation water. Sci Hortic 115:275–284
Clark RB, Zeto SK (2000) Mineral acquisition by arbuscularmycorrhizal plants. J Plant Nutr 23:867–902
Colla G, Rouphael Y, Cardarelli M, Tullio M, Rivera CM, Rea E(2008) Alleviation of salt stress by arbuscular mycorrhizal inzucchini plants grown at low and high phosphorus concentration.Biol Fertil Soils 44:501–509
Declerck S, Plenchette C, Strullu DG (1995) Mycorrhizal dependencyof banana (Musa acuminate, AAA group) cultivar. Plant Soil176:183–187
Giovannetti M, Mosse B (1980) An evaluation of techniques formeasuring vesicular arbuscular mycorrhizal infection in roots.New Phytol 84:489–500
Gogorcena Y, Abadía J, Abadía A (2004) A new technique forscreening iron-efficient genotypes in peach rootstocks: elicitationof root ferric chelate reductase by manipulation of external ironconcentrations. J Plant Nutr 27:1701–1715
Hetrick BAD, Wilson GWT (1992) Mycorrhizal dependence ofmodern wheat cultivars, landraces, and ancestors. Can J Bot70:2032–2040
Hickman GV (1998) Commercial greenhouse vegetable hand-book. University of California, Division of Agriculture andNatural Resources Communication Services–Publications,Oakland, p 15
Biol Fertil Soils

Isaac RA, Johnson WC Jr (1998) Elemental determination byinductively coupled plasma atomic emission spectrophotometry.In: Kalra YP (ed) Handbook of reference methods for plantanalysis. CRC, Raton, pp 165–170
Iturbe-Ormaexte I, Moran JF, Arrese-Igor C, Gogorcena Y, Klucas RV,Becana M (1995) Activated oxygen and antioxidant defenses iniron deficient pea plants. Plant Cell Environ 18:421–429
Jeffries P, Gianianazzi S, Perotto S, Turnau K, Barea JM (2003) Thecontribution of arbuscular mycorrhizal fungi in sustainable mainte-nance of plant health and soil fertility. Biol Fertil Soils 37:1–16
Khalil S, Loynachan TE, Tabatabai MA (1994) Mycorrhizal depen-dency and nutrient uptake by improved and unimproved corn andsoybean cultivars. Agron J 8:949–958
Kiehl PA, Lieth JH, Burger DW (1992) Growth response ofchrysanthemum to various container medium moisture tensionlevels. J Am Soc Hortic Sci 114:387–392
Koide R, Minguang L, Lewis L, Irby C (1988) Role of mycorrhizalinfection in the growth and reproduction of wild vs. cultivatedplants. I. Wild vs. cultivated oats. Oecologia 77:537–543
Linderman RG, Davis AE (2004) Varied response of marigold(Tagetes spp.) genotypes to inoculation with different arbuscularmycorrhizal fungi. Sci Hortic 99:67–78
Lutts S, Kinet JM, Bouharmont J (1995) Changes in plant response toNaCl during development of rice varieties differing in salinityresistance. J Exp Bot 46:1843–1852
Marschner H (1995) Mineral nutrition of higher plants, IIth edn. Press,London, 889 p
Marschner H (1998) Role of root growth, arbuscular mycorrhiza, androot exudates for the efficiency in nutrient acquisition. FieldCrops Res 56:203–207
Mengel K (1994) Iron availability in plant tissues-iron chlorosis oncalcareous soil. Plant Soil 165:275–283
Miller RM, Jastrow JD (2000) Mycorrhizal fungi influence soilstructure. In: Kapulnik Y, Douds DD Jr (eds) Arbuscularmycorrhizas: physiology and function. Kluwer Academic Publi-cation, Dordrecht, pp 3–18
Misra A, Tyler G (1999) Influence of soil moisture on soil solutionchemistry and concentrations of minerals in the calcicolesPhleum phleoides and Veronica spicata grown on a limestonesoil. Annal Bot 84:401–410
Molassiotis A, Tanou G, Diamantidis G, Patakas A, Therios I (2006)Effects of 4-month Fe deficiency exposure on Fe reductionmechanism, photosynthetic gas exchange, chlorophyll fluores-cence and antioxidant defense in two peach rootstocks differingin Fe deficiency tolerance. J Plant Physiol 163:176–185
Morales F, Abadía A, Abadía J (1991) Chlorophyll fluorescence andphoton yield of oxygen evolution in iron-deficient sugar beet(Beta vulgaris L.) leaves. Plant Physiol 97:886–893
Nedunchezhian N, Morales F, Abadía A, Abadía J (1997) Decline inphotosynthetic electron transport activity and changes in thyla-koid protein pattern in field grown iron deficient peach (Prunuspersica L.). Plant Sci 129:29–38
Nikolic M, Kastori R (2000) Effect of bicarbonate and Fe supply onFe nutrition of grapevine. J Plant Nutr 23:1619–1627
Norrie J, Graham MED, Gosselin A (1994) Potential evapotranspira-tion as a means of predicting irrigation timing in greenhousetomatoes grown in peat bags. J Am Soc Hortic Sci 119:163–168
Parke JL, Kaeppler SW (2000) Effects of genetic differences amongcrop species and cultivars upon the arbuscular mycorrhizalsymbiosis. In: Kapulnik Y, Douds DD Jr (eds) ArbuscularMycorrhizas: Physiology and function. Kluwer Academic Publi-cation, Dordrecht, pp 131–146
Phillips J, Hayman D (1970) Improved procedures for clearing rootsand staining parasitic and vesicular-arbuscular mycorrhizal fungifor rapid assessment of infection. Trans Br Mycol Soc 55:158–161
Pissaloux A, Morarad P, Bertoni G (1995) Alkalinity-bicarbonatecalcium effects on iron chlorosis in white lupine in soillessculture. In: Abadía J (ed) Development in plant and soil science,59. Iron nutrition in soils and plants; Seventh InternationalSymposium on Iron Nutrition and Interactions in Plants,Zaragoza, Spain, June 27–July 2, 1993. Kluwer AcademicPublishers, Dordrecht, pp 127–133
Plenchette C, Declerck S, Diop T, Strullu DG (1996) Infectivity ofmonoaxenic subcultures of the AM fungus Glomus versiformeassociated with Ri-TDNA transformed root. Appl MicrobiolBiotechnol 46:545–548
Reynier A (1997) Manuel de viticulture. Technique et Documentation(Lavoisier). 7ème éd. Bailliere, Paris, p 414
Römheld V (2000) The chlorosis paradox: Fe inactivation as asecondary event in chlorotic leaves of grapevine. J Plant Nutr23:1629–1643
Sanchez Blanco MJ, Ferrández T, Angeles Morales M, Morte A,Alarcón JJ (2004) Variations in water status, gas exchange,and growth in Rosmarinus officinalis infected with Glomusdesrticola under drought conditions. J Plant Physiol 161:675–682
Sensoy S, Demir S, Turkmen O, Erdinc C, Savur OB (2007)Responses of some different pepper (Capsicum annuum L.)genotypes to inoculation with two different arbuscular mycorrhi-zal fungi. Sci Hortic 113:92–95
Smith SE, Read DW (1997) Mycorrhizal symbiosis, 2nd edn.Academic, London
SPSS (2001) SPSS 10 for Windows. Applications Guide. SPSS Inc,Chicago
Stockinger H, Walker C, Schüßler A (2009) ‘Glomus intraradicesDAOM197198’, a model fungus in arbuscular mycorrhizaresearch, is not Glomus intraradices. New Phytol 183:1176–1187
Tennessen DJ, Singsaas EL, Sharkey TD (1994) Light-emitting diodesas a light source of photosynthesis research. Photosyn Res39:85–92
Toth R, Toth D, Starke D, Smith DR (1990) Vesicular-arbuscularmycorrhizal colonization in Zea mays affected by breeding forresistance to fungal pathogens. Can J Bot 68:1039–1044
Troeh FR, Thompson LM (2005) Soils and soil fertility, 6th edn.Blackwell, Ames, IA 50014, p 489
Valdez-Aguilar LA (2004) Effect of alkalinity in irrigation water onselected greenhouse crops. PhD thesis in Horticulture, TexasA&M University, 500014, p 489
Valdez-Aguilar LA, Reed DW (2006) Comparison of growth andalkalinity-induced responses in two cultivars of hibiscus (Hibis-cus rosa-sinensis L.). HortScience 41:1704–1708
Valdez-Aguilar LA, Reed DW (2007) Response of selected green-house ornamental plants to alkalinity in irrigation water. J PlantNutr 30:441–452
Valdez-Aguilar LA, Reed DW (2008) Influence of potassiumsubstitution by rubidium and sodium on growth, ion accumula-tion, and ion partitioning in bean under high alkalinity. J PlantNutr 31:867–883
Wright DP, Scholes JD, Read DJ (1998) Effects of VA mycorrhizalcolonization on photosynthesis and biomass production ofTrifolium repens L. Plant Cell Environ 21:209–216
Yakushiji H, Morinaga K, Nonami H (1998) Sugar accumulationand partitioning in Satsuma mandarin tree tissues and fruitin response to drought stress. J Am Soc Hortic Sci 123:719–726
Zhang L, Zhang J, Christie P (2008) Pre-inoculation with arbuscualrmycorrhizal fungi suppresses root knot nematode (Meloidogyneincognita) on cucumber (Cucumis sativus). Biol Fert Soils45:205–211
Zribi K, Gharsalli M (2002) Effect of bicarbonate on growth and ironnutrition of pea. J Plant Nutr 25:2143–2149
Biol Fertil Soils


















