
Improved flatfish health following remediation of a PAH-contaminated site in Eagle Harbor,...
12
Aquatic Toxicology 88 (2008) 277–288 Contents lists available at ScienceDirect Aquatic Toxicology journal homepage: www.elsevier.com/locate/aquatox Improved flatfish health following remediation of a PAH-contaminated site in Eagle Harbor, Washington Mark S. Myers ∗ , Bernadita F. Anulacion, Barbara L. French, William L. Reichert, Cathy A. Laetz, Jon Buzitis, O. Paul Olson, Sean Sol, Tracy K. Collier Environmental Conservation Division, Northwest Fisheries Science Center, National Marine Fisheries Service/NOAA Fisheries, 2725 Montlake Blvd. E., Seattle, WA 98112, United States article info Article history: Received 10 April 2008 Accepted 2 May 2008 Keywords: English sole Biomarkers of PAH exposure Site remediation Puget Sound Washington Toxicopathic liver lesions abstract Eagle Harbor in Puget Sound, WA became a Superfund site in 1987 due to polycyclic aromatic hydrocarbons (PAHs) released chronically from a nearby creosoting facility. Early studies here (1983–1986) demon- strated up to an ∼80% prevalence of toxicopathic liver lesions, including neoplasms, in resident English sole (Parophrys vetulus). These lesions in English sole are consistently associated with PAH exposure in multiple field studies, and one laboratory study. Later studies (1986–1988) incorporated biomarkers of PAH exposure and effect, including hepatic CYP1A expression and xenobiotic–DNA adducts, and biliary fluorescent aromatic compounds (FACs). Before site remediation, lesion prevalences and other biomarker values in this species from Eagle Harbor were among the highest compared to other sites in Puget Sound and the US Pacific Coast. To sequester PAH-contaminated sediments, in 1993–1994, a primary cap of clean sediment was placed over the most-contaminated 54acres, with a 15-acre secondary cap added from 2000–2002. Lesion prevalences and biomarker values before primary capping were reduced compared to 1983–1986, consistent with facility closure in 1988 and shore-based source controls begun in 1990. Liver lesion risk, hepatic CYP1A activities, and levels of biliary FACs from fish collected immediately after and at regular intervals up to 2 years after primary capping were variable relative to pre-capping. Over the entire monitoring period since primary capping (128 months), but particularly after 3 years, there was a significantly decreasing trend in biliary FACs, hepatic DNA adducts and lesion risk in English sole. In particular, lesion risk has been consistently low (<0.20) compared to primary cap initiation (set at 1.0), from ∼4 years after primary capping through April 2004. These results show that the sediment capping process has been effective in reducing PAH exposure and associated deleterious biological effects in a resident flatfish, and that longer term monitoring of pollutant responses in biological resources, such as resident fish, is needed in order to demonstrate the efficacy of this type of remediation. Published by Elsevier B.V. 1. Introduction Extensive evidence indicates that exposure to polycyclic aro- matic hydrocarbons (PAHs) and their derivatives produces a variety of adverse biological effects in marine fish. While much of this infor- mation is derived from laboratory studies, there is an increasing body of evidence from field studies supporting these relationships between PAH exposure and biological injury. Several of the most definitive field studies in this regard have been done in Puget Sound, Washington (Myers et al., 1987, 1998, 2003; Rhodes et al., 1987; Johnson et al., 1988; Casillas et al., 1991; Landahl and Johnson, 1993) where one of the most heavily PAH-contaminated sites has ∗ Corresponding author. Tel.: +1 206 860 3329; fax: +1 206 860 3335. E-mail address: [email protected] (M.S. Myers). been in Eagle Harbor, on the east side of Bainbridge Island in Puget Sound, WA (Fig. 1). Studies done by Northwest Fisheries Science Center (NWFSC)/National Oceanic and Atmospheric Administra- tion (NOAA) scientists since 1983 in Eagle Harbor showed that resident epibenthic flatfish such as English sole (Parophrys vetulus) there were at increased risk of developing serious toxicopathic liver diseases, including cancer, and of exhibiting impaired reproductive function and altered physiological responses (Malins et al., 1985; Myers et al., 1987; Rhodes et al., 1987; Johnson et al., 1988; Casillas et al., 1991; Stein et al., 1992). In fact, toxicopathic hepatic lesion prevalences (Malins et al., 1985; Myers et al., 1987) and other values for biomarkers of PAH exposure in Eagle Harbor (Stein et al., 1992; Myers et al., 1998) were among the highest recorded in Puget Sound, with up to ∼80% of English sole affected with these liver lesions. Due largely to these findings, as well as the high levels of PAHs in the sediments (up to 120,000 ppb ∑ PAHs), English sole stomach contents (up to 84,000 ppb ∑ PAHs) (Malins et al., 1985) 0166-445X/$ – see front matter. Published by Elsevier B.V. doi:10.1016/j.aquatox.2008.05.005
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Improved flatfish health following remediation of a PAH-contaminated site in Eagle Harbor,...

Aquatic Toxicology 88 (2008) 277–288
Contents lists available at ScienceDirect
Aquatic Toxicology
journa l homepage: www.e lsev ier .com/ locate /aquatox
Improved flatfish health following remediation of a PAH-contaminatedsite in Eagle Harbor, Washington
Mark S. Myers ∗, Bernadita F. Anulacion, Barbara L. French, William L. Reichert,Cathy A. Laetz, Jon Buzitis, O. Paul Olson, Sean Sol, Tracy K. CollierEnvironmental Conservation Division, Northwest Fisheries Science Center, National Marine Fisheries Service/NOAA Fisheries,2725 Montlake Blvd. E., Seattle, WA 98112, United States
a r t i c l e i n f o a b s t r a c t
Article history:Received 10 April 2008
Eagle Harbor in Puget Sound, WA became a Superfund site in 1987 due to polycyclic aromatic hydrocarbonslly frovalenhese
oneinclupound
Eaglo sequr theencesth fac
activ2 yeasince
Accepted 2 May 2008
Keywords:English soleBiomarkers of PAH exposureSite remediationPuget SoundWashingtonToxicopathic liver lesions
(PAHs) released chronicastrated up to an ∼80% presole (Parophrys vetulus). Tmultiple field studies, andPAH exposure and effect,fluorescent aromatic comvalues in this species fromand the US Pacific Coast. Tsediment was placed ove2000–2002. Lesion preval1983–1986, consistent wilesion risk, hepatic CYP1Aat regular intervals up toentire monitoring period
a significantly decreasing trendparticular, lesion risk has beenfrom ∼4 years after primary caprocess has been effective in rresident flatfish, and that longeresident fish, is needed in orde1. Introduction
Extensive evidence indicates that exposure to polycyclic aro-matic hydrocarbons (PAHs) and their derivatives produces a varietyof adverse biological effects in marine fish. While much of this infor-mation is derived from laboratory studies, there is an increasingbody of evidence from field studies supporting these relationshipsbetween PAH exposure and biological injury. Several of the mostdefinitive field studies in this regard have been done in Puget Sound,Washington (Myers et al., 1987, 1998, 2003; Rhodes et al., 1987;Johnson et al., 1988; Casillas et al., 1991; Landahl and Johnson,1993) where one of the most heavily PAH-contaminated sites has
∗ Corresponding author. Tel.: +1 206 860 3329; fax: +1 206 860 3335.E-mail address: [email protected] (M.S. Myers).
0166-445X/$ – see front matter. Published by Elsevier B.V.doi:10.1016/j.aquatox.2008.05.005
m a nearby creosoting facility. Early studies here (1983–1986) demon-ce of toxicopathic liver lesions, including neoplasms, in resident Englishlesions in English sole are consistently associated with PAH exposure inlaboratory study. Later studies (1986–1988) incorporated biomarkers of
ding hepatic CYP1A expression and xenobiotic–DNA adducts, and biliarys (FACs). Before site remediation, lesion prevalences and other biomarker
e Harbor were among the highest compared to other sites in Puget Soundester PAH-contaminated sediments, in 1993–1994, a primary cap of cleanmost-contaminated 54 acres, with a 15-acre secondary cap added fromand biomarker values before primary capping were reduced compared to
ility closure in 1988 and shore-based source controls begun in 1990. Liverities, and levels of biliary FACs from fish collected immediately after andrs after primary capping were variable relative to pre-capping. Over theprimary capping (128 months), but particularly after 3 years, there wasin biliary FACs, hepatic DNA adducts and lesion risk in English sole. In
consistently low (<0.20) compared to primary cap initiation (set at 1.0),pping through April 2004. These results show that the sediment cappingeducing PAH exposure and associated deleterious biological effects in ar term monitoring of pollutant responses in biological resources, such as
r to demonstrate the efficacy of this type of remediation.Published by Elsevier B.V.
been in Eagle Harbor, on the east side of Bainbridge Island in PugetSound, WA (Fig. 1). Studies done by Northwest Fisheries ScienceCenter (NWFSC)/National Oceanic and Atmospheric Administra-tion (NOAA) scientists since 1983 in Eagle Harbor showed thatresident epibenthic flatfish such as English sole (Parophrys vetulus)there were at increased risk of developing serious toxicopathic liverdiseases, including cancer, and of exhibiting impaired reproductivefunction and altered physiological responses (Malins et al., 1985;Myers et al., 1987; Rhodes et al., 1987; Johnson et al., 1988; Casillaset al., 1991; Stein et al., 1992). In fact, toxicopathic hepatic lesionprevalences (Malins et al., 1985; Myers et al., 1987) and other valuesfor biomarkers of PAH exposure in Eagle Harbor (Stein et al., 1992;Myers et al., 1998) were among the highest recorded in Puget Sound,with up to ∼80% of English sole affected with these liver lesions.
Due largely to these findings, as well as the high levels ofPAHs in the sediments (up to 120,000 ppb
∑PAHs), English sole
stomach contents (up to 84,000 ppb∑
PAHs) (Malins et al., 1985)

278 M.S. Myers et al. / Aquatic Toxicology 88 (2008) 277–288
process of hepatocarcinogenesis. These lesions have been strongly
Fig. 1. Map showing location of Eagle Harbor, WA in Puget Sound.
and edible marine biota, Eagle Harbor was designated a Super-fund site in 1987. These high levels of PAH exposure were aresult of long-term and continuing releases of creosote from theonshore facility which had pressure-treated wood pilings with coaltar creosote almost continuously since 1903, referred to as theWyckoff site/facility after the name of the most recent companyoperating the facility. The United States Environmental ProtectionAgency (USEPA) completed a Remedial Investigation/FeasibilityStudy in 1989, and in December 1991 proposed a phased cleanupof the harbor which included capping of PAH-contaminated sedi-ments (http://yosemite.epa.gov/R10/CLEANUP.NSF/sites/Wyckoff).In June 1993, under Superfund Removal authority, the USEPArecommended placement of relatively uncontaminated (<570 ppb∑
PAHs) Snohomish River dredge material over the most contam-inated area of Eagle Harbor by a combined process of release ofdredge material from split-hull barges in the deeper areas of EagleHarbor and wash-off of similar material from regular barges in theshallower regions of the area to be capped. In late September 1993,
the USEPA and US Army Corps of Engineers began placement ofthis sediment cap as a means of (1) controlling transport of con-taminants; (2) isolating contaminants from marine biota; and (3)providing clean habitat for benthic organisms, thus reducing overallrisk of exposure to PAHs contained in the sediments. This primarycapping process was completed in March 1994, resulting in place-ment of an approximately three-foot thick cap of 275,000 cubicyards of clean sandy material and associated wood debris over anoriginal area of approximately 54 acres of the most highly contam-inated subtidal sediments within Eagle Harbor (Fig. 2), includinga PAH “hot spot” documented in 1983 (Malins et al., 1985). Otherremediation actions have included: (1) construction of an onshoregroundwater extraction and treatment system in 1990 and subse-quent intermittent operation of this system up through the present;(2) installation in November 2000–March 2001 of a 2000 feet longsheet pile wall driven 10–100 feet deep along the entire shoreline ofthe former Wyckoff site to curtail continuing subtidal and intertidalseeps of free creosote into Eagle Harbor sediments. These seepshad resulted in multiple areas within the capped area displayinglocally elevated surface high molecular weight PAH concentrationsFig. 2. Map of area of Eagle Harbor in which fish sampling (bottom trawling) oper-ations were conducted, in relation to the past area of high sediment PAHs. Alsoshown are the areas sequentially capped: (1) in September 1993 to March 1994with sediments composed of dredge material from the Snohomish River (polygonalshaded area including the PAH “hot spot”), covering 54 acres, and (2) in November2000–February 2002 as a secondary cap of clean sand, joining the southern edge ofthe original cap with the northern shoreline of the former Wyckoff facility, addinganother 15 acres of area capped with clean sediments.
exceeding both the Washington State Sediment Quality Standardsand Minimum Cleanup Levels, up to ∼72,000 ppb (USEPA/USACOE,1999); and (3) from November 2000–February 2002, extension ofthe original sediment cap from its southern edge to the north-facing shoreline of the former Wyckoff creosoting facility to furthersequester the continuing subtidal and intertidal seeps of free cre-osote. This placement of ∼170,000 cubic yards of clean sand createdanother 15 acres of clean sediment cap over PAH-contaminatedregions of Eagle Harbor (Fig. 2).
The primary objective of this study was to assess the effi-cacy of these remediation measures in Eagle Harbor in terms ofreducing PAH exposure and improving the health of resident fish.Several biochemical measures of PAH exposure and early toxico-logical response were monitored in the English sole, a residentflatfish species that has been routinely used as a sentinel species inPuget Sound. In addition, we performed histopathological exam-inations of the livers of the same English sole assayed for PAHexposure and early response, and determined the occurrence ofa spectrum of toxicopathic hepatic lesions involved in the stepwise
and consistently associated with PAH exposure in wild English sole(Myers et al., 1987, 2003). Moreover, certain neoplasia-related liverlesions have been induced in sole by injections of a PAH-rich frac-tion extracted from Eagle Harbor sediment (Schiewe et al., 1991).These four measures, or biomarkers, of PAH exposure and effect,are described below.
Because of the high rate of metabolism and depuration ofPAHs by many species of fish, direct measurement of parent com-pounds in tissues is not generally a useful indicator of exposureof fish to PAHs (Varanasi et al., 1989a). Accordingly, methodshave been developed to determine such exposure, based on ourincreased understanding of the mechanisms and pathways of PAHmetabolism in fish.
The first biomarker used in this study is the measurement of flu-orescent aromatic compounds (FACs) in bile by high performanceliquid chromatography (HPLC) with fluorescence detection. Manypolar and fluorescent metabolites of PAHs are primarily excretedvia the hepatobiliary system in fish. This method provides a semi-quantitative means for rapidly assessing recent PAH exposure infish and has the added advantage of being able to be ‘fine-tuned’

Toxico
On 13–14 September 1993, prior to primary sediment cap place-
M.S. Myers et al. / Aquatic
for different classes of PAHs, such as 2–3 ring, 3–4 ring, or 4–5 ringstructures (Krahn et al., 1984, 1986, 1992). This biomarker of short-term PAH exposure responds within hours, is dose-responsive,typically reaches a maximum within 2 weeks in conditions of con-tinuous exposure, and has a half-life of 14–16 days in English sole(Collier and Varanasi, 1991; Anulacion et al., 1995; French et al.,1996).
A second biomarker is the induction of hepatic cytochromeP4501A (CYP1A), which, together with the associated monooxyge-nase activity of aryl hydrocarbon hydroxylase (AHH), can be rapidlyinduced in many fish species after exposure to a wide variety oforganic contaminants, including PAHs (Payne and Penrose, 1975;Payne et al., 1987; Collier and Varanasi, 1991). CYP1A induction rep-resents an early biological effect useful in estimating PAH exposureof fish; however, CYP1A induction is not specific for PAH exposure.The induction of CYP1A is initiated by the interaction of compoundswith the Ah receptor, a step believed to be involved in the subse-quent development of several toxic effects, including reproductivedysfunction, teratogenicity, mutagenicity, and carcinogenicity. Thisis a less specific biomarker of short term PAH exposure that isresponsive within hours of exposure, is dose-responsive, reachesa maximum at 3 days in conditions of continuous exposure, andhas a half-life of 12–28 days in English sole (Anulacion et al., 1995;Collier and Varanasi, 1991).
The third biomarker of PAH exposure we employed is theformation of DNA adducts in liver tissue. Adducts form whenelectrophilic intermediates of high molecular weight PAHs formcovalent bonds with nucleophilic centers within biological macro-molecules, including hepatic DNA. The method for quantifyingDNA adducts is highly sensitive for detecting bulky hydrophobicxenobiotic–DNA adducts (Gupta and Randerath, 1988; Varanasi etal., 1989b; Reichert and French, 1994). This biomarker is considereda longer-term, dose–responsive indicator of PAH exposure, whichcan respond within days of exposure to PAH-contaminated sedi-ments such as those from Eagle Harbor, but also shows persistenceand accumulation of adducts over time, with slow rates of repair(Stein et al., 1995; French et al., 1996) and an estimated half-life of40–100 days (Anulacion et al., 1995). Therefore, it is considered areliable and useful biomarker of cumulative and chronic exposureto PAHs in wild fish.
The fourth method is histopathological examination of liverfrom the same individuals used in the previous assays to documentthe presence/absence of toxicopathic liver lesions involved in hep-atocarcinogenesis in English sole. These lesions are causally related
to exposure to PAHs, and constitute a biomarker of effect of longer-term PAH exposure (Myers et al., 1987, 2003). This biomarkerserves as the primary indicator of English sole health in thisstudy. Lesions such as hepatocellular megalocytosis, nuclear pleo-morphism and hepatocellular regeneration, which occur early inthe histogenesis of liver neoplasia in English sole (Myers et al.,1987) can occur in young-of-the-year (<1 year of age) wild Englishsole from PAH-contaminated sites (Rhodes et al., 1987; Myers etal., 1987, 1998), while the preneoplastic and neoplastic hepaticlesions are rarely detected in English sole less than three yearsof age (Rhodes et al., 1987; Myers et al., 1987, 1998). Such lesionsare therefore considered effective longer-term biomarkers of PAHexposure and indicators of the overall health status of Englishsole.These methods have been employed successfully in determin-ing PAH exposure of fish, especially in English sole. The time– anddose–responsiveness of each of these measures has been deter-mined after exposure to either single PAHs or to complex mixturesof PAHs and other organic contaminants (Collier and Varanasi,1991; Schiewe et al., 1991; Stein et al., 1993, 1994). Moreover, eachof these measures has been extensively field tested, both in Puget
logy 88 (2008) 277–288 279
Sound (Varanasi et al., 1986; Krahn et al., 1986; Myers et al., 1987,1998, 2003; Collier et al., 1992a,b, 1995; Stein et al., 1992) and else-where (Varanasi et al., 1989a,b, 1992; Krahn et al., 1992; Collier etal., 1992a, 1993a,b; Myers et al., 1999). These studies have shownthat the combined use of several indicators is the optimal approach,because each method focuses on a different aspect of PAH toxicoki-netics and provides useful information concerning possible toxiceffects of PAH exposure (Stein et al., 1992; Collier et al., 1993; Myerset al., 1998).
Studies similar to the present one have utilized hepatichistopathology in resident fish species as an assessment tool tomonitor recovery of fish health subsequent to habitat remediationmeasures at other contaminated aquatic or marine sites. For exam-ple, the prevalence of brown bullhead with liver tumors declinedfrom 39% to 10% after closure of a coking plant in 1983 on theseverely PAH-contaminated Black River in Ohio, coincident with adecline in sediment PAH levels at this site of two orders of mag-nitude (Baumann and Harshbarger, 1995). However, subsequentto dredging and resuspension of buried PAH-contaminated sedi-ments in 1990 at this site, neoplasm prevalences in 1992 and 1993were as high or higher than those in the 1980s. By 1994 the tumorprevalence had again declined, especially among 3-year-old fish,which was the first group sampled that were not present duringthe 1990 dredging (Baumann and Harshbarger, 1998). Monitoringof organic contaminant exposure, neoplasm prevalence and that ofthe most commonly occurring toxicopathic hepatic lesion, hydropicvacuolation, in winter flounder from the Boston Harbor regionbetween 1987 and 2003 also demonstrated a parallel reduction incontaminant exposure and the prevalence of hydropic vacuolationfollowing improved effluent quality and relocation of the Bostonsewage outfall into Massachusetts Bay (Moore et al., 2005).
In the present study, biological, histopathological and biochem-ical PAH exposure data collected in September of 1993, prior toinitiation of the sediment capping process, served as the baselinefor comparison with data from samples collected from multiple,and at least annual, time points following sediment cap completion(March 1994) through April of 2004.
2. Materials and methods
2.1. Field collection of English sole
ment, English sole were collected in Eagle Harbor within an areajust west of and adjacent to the cap placement area (Fig. 2). Trawl-ing within the capped area was not permitted in any phase of thisstudy, so as to avoid disturbing or damaging the integrity of thesediment cap. Samples were taken of liver tissue and bile for anal-ysis of biomarkers indicative of exposure to PAHs, as describedearlier. Subsequent samplings of English sole were carried out inMarch 1994, July 1994, September 1994, March 1995, July 1995,October–November 1995, December 1995, October 1996, Decem-ber 1997, May 1998, May 1999, April 2000, May 2001, May 2002,April 2003, and April 2004. A target sample size of at least 15 indi-viduals was established for each of the 17 field samplings and thistarget was achieved in most cases (16 of 17), but at most of theannual samplings in September 1993 and from 1998 on, at least 60English sole were captured and necropsied (Table 1).
All sampling was done either from the R/V HAROLD W.STREETER, a 45 foot NOAA Fisheries research vessel, or by the FVCHASINA, a 58 foot stern trawler/purse seiner operated under con-tract. Fish were collected by otter trawl within the area shown inFig. 2. Trawl duration was between 5 and 15 min, at a speed of1.8–2.2 nautical miles/h.

280 M.S. Myers et al. / Aquatic Toxic
Table 1
Sampling dates and number of English sole samples analyzed for each biomarkerDate(s) Path Adduct CYP1A Bile
September 1993 73 35 39 63March 1994 21 21 20 18July 1994 30 30 26September 1994 37 31 39 34March 1995 32 32 32 31July 1995 19 20 17October–November 1995 21 21 21December 1995 6 6 6October 1996 19 19 13December 1997 26 7May 1998 73 36 45 14May 1999 72 20 72 50April 2000 60 20 45May 2001 60 20 45May 2002 60 20 52April 2003 60 52April 2004 60 20 59
Total 729 255 343 553
Path = liver sample for histopathology; adduct = liver sample for DNA adduct analy-sis; CYP1A = liver sample for CYP1A analysis; Bile = bile sample for FAC analysis.
2.2. Necropsy procedure
After capture, English sole were placed into holding tanks con-taining flowing seawater and held alive until necropsy. Fish fornecropsy were assigned a unique specimen identification number,measured for total length (mm), and killed by severing the spinalcolumn. Otoliths were collected from each fish for determinationof age by counting of annual growth rings (Chilton and Beamish,1982). The abdominal cavity was opened for sex determination, andany grossly visible liver nodules or tumors were noted. Bile from thegall bladder was collected into an amber vial, and placed on ice untilfrozen in the lab at −20 ◦C. Liver tissue for xenobiotic–DNA adductanalysis (≥250 mg) and CYP1A analysis (≥500 mg) was placed intoseparate plastic cryovials, flash frozen in liquid nitrogen, and storedfrozen in the lab at −80 ◦C prior to analysis. A 2–3-mm thick portionof liver tissue from a standardized region of the liver (central por-tion along the longitudinal axis including the anterior and largestposterior lobe), in addition to any portions showing grossly visiblenodules or alteration in color or texture were collected from eachindividual for histopathological examination and placed immedi-ately into Dietrich’s fixative (Gray, 1954). A transverse section ofthe gonad was also collected and preserved in Dietrich’s fixative
for histological confirmation of the gross sex determination.2.3. Fluorescent aromatic compounds (FACs) in bile
Bile samples were analyzed for metabolites of aromaticcompounds using a high-performance liquid chromatographic pro-cedure with fluorescence detection (Krahn et al., 1984, 1986, 1992),with minor modifications (Collier and Varanasi, 1991). Briefly, the(thawed) bile was injected directly into a high-performance liq-uid chromatograph (HPLC) equipped with a PerkinElmer HC-ODS(reverse-phase) analytical column and two fluorescence detectorsconnected in series, and a gradient (100% water containing 5 �Lacetic acid/L, to 100% methanol). The excitation/emission wave-lengths of one detector were set to 290/335 nm, where metabolitesof naphthalene (NPH) fluoresce. The excitation/emission wave-lengths of the other detector were set to 380/430 nm, where themetabolites of benzo[a]pyrene (BaP) fluoresce. The total integratedarea for each detector was then converted (normalized) to equiva-lents of known concentrations of either NPH or BaP standards. BileFACs data was reported on the basis of mg of biliary protein (Collieret al., 1989).
ology 88 (2008) 277–288
Quality assurance procedures included NPH and BaP calibra-tion standards, a “bile pool” reference material, blank analyses, andreplicate analyses to evaluate HPLC/UV fluorescence performance(Sloan et al., 2006).
2.4. Hepatic hydrophobic xenobiotic–DNA adducts
Analyses of DNA damage were done by the 32P-postlabelingmethod (Reichert and French, 1994). Briefly, these proceduresinvolve: (1) hydrolysis of isolated DNA to normal and adducteddeoxyribonucleotide-3′-monophosphates; (2) nuclease P1 treat-ment of the DNA hydrolysate to remove the normal nucleotides;(3) labeling of the adducts using [�-32P]-ATP; (4) one-dimensionalthin-layer chromatographic separation of the normal nucleotidesfrom the adducts, two-dimensional separation of individual DNAadducts from one another, and one-dimensional separation of indi-vidual DNA bases; and (5) quantitation of adduct levels on thechromatograms by storage phosphor imaging (Reichert et al., 1992;Reichert and French, 1994).
Quality assurance procedures for the 32P-postlabeling assayincluded: (1) the use of salmon testes DNA for measuringthe efficiency of DNA hydrolysis, and as a sample blank andreference standard; (2) the use of 7R,8S,9S,10R-(N2-deoxyguanosyl-3′-phosphate)-7,8,9,10-tetrahydrobenzo(a)pyrene (BaP-dG) as aninternal standard to measure the efficiency of the enzyme-mediated transfer of the 32P-phosphate from ATP to the adducts;(3) the use of the 32P-labeled BaP-dG monophosphate standard as achromatography calibration standard in a separate analysis, ratherthan an internal standard, because of the interference of the stan-dard with the measurement of the unknown adducts; and (4) theuse of 2′-deoxyguanosine-3′-monophosphate to measure the effi-ciency of the enzyme-dependent labeling of the normal nucleotidesby 32P-phosphate from [�32P]ATP.
2.5. Hepatic CYP1A analysis (as AHH activity)
Analyses of hepatic AHH activities were performed by meth-ods as described previously (Collier et al., 1995). Briefly, hepaticmicrosomes were prepared by a slight modification of a previouslydescribed procedure (Collier et al., 1986). The use of glycerol in theresuspension was a modification to the original method, to enhancestability during storage beyond what has been previously reported(Schnell et al., 1980). Protein was measured (Lowry et al., 1951;Collier et al., 1989), using bovine serum albumin as a standard.
AHH activity was assayed by a modification of aradiometricmethod (Van Cantfort et al., 1977; Collier et al., 1986). All enzymeassays were run under conditions in which the reaction rates are inthe linear range for both time and protein. Quality assurance pro-cedures for AHH measurements included duplicate zero-time andboiled enzyme blanks for each set of assays. Each sample was runin triplicate and those samples showing >20% absolute differencebetween duplicates and >10 units (pmoles benzo[a]pyrene metab-olized/mg microsomal protein/min) difference between duplicateswere reanalyzed.
2.6. Histopathology
Tissues preserved in the field (liver and gonad) were dehy-drated, processed using a Shandon Hypercenter XP automatedtissue processor, and embedded in Polyfin (Triangle BiomedicalSciences). Tissue sections (cut to 4–5 �m thickness) were stainedwith Gill’s haematoxylin and eosin-phloxine (Luna, 1968), andexamined by light microscopy by a single, experienced examiner(MS Myers). Lesion classification followed widely accepted crite-ria for the diagnosis of liver lesions in English sole (Myers et al.,

Toxico
M.S. Myers et al. / Aquatic1987) and other fish species (Boorman et al., 1997) including thoseexposed environmentally to creosote in the Elizabeth River, VA(Vogelbein et al., 1990). For reporting and statistical analyses, toxi-copathic hepatic lesions were placed into the following diagnosticcategories: (1) neoplasms (NEO) [hepatocellular adenoma, hepato-cellular carcinoma, cholangioma, cholangiocellular carcinoma, andmixed hepatobiliary carcinoma]; (2) putatively preneoplastic fociof cellular alteration (PRENEO) [eosinophilic focus, clear cell focus,basophilic focus]; (3) non-neoplastic proliferative lesions (PROLIF)[hepatocellular regeneration, biliary hyperplasia or proliferation,“oval cell” proliferation, cholangiofibrosis, and increased mitoticactivity in hepatocellular/biliary epithelial cells]; (4) hepatocellularmegalocytosis or nuclear pleomorphism (HM/NP); and (5) any tox-icopathic liver lesion (TOXLIV), which includes fish affected withany of the lesions within categories 1–4. The microscopic anatomyof these lesions has been previously described in detail in Englishsole and other fish species (Myers et al., 1987; Vogelbein et al., 1990;Boorman et al., 1997; Wolf and Wolfe, 2005).
2.7. Statistical analyses
Statistical analyses of the bile FACs, CYP1A and xenobiotic–DNAadduct data were performed using the Statview 4.01 softwareprogram (Abacus Concepts, Berkeley, CA). All data from measure-ment of biliary FACs, and hepatic CYP1A were log-transformedprior to statistical treatment to reduce heteroscedasticity. The datafor hepatic xenobiotic–DNA adducts were not log-transformedbecause the data were normally distributed. However, for clar-ity of presentation, all figures shown in the results section depictuntransformed values for these respective biomarkers, with theexception of the regression plots of biliary FACs and hepatic CYP1Avs. time post-capping, which are shown with the log-transformeddata. Relationships among these biomarker measures and otherbiological variables such as fish age were determined by analysisfor linear correlation; in all cases age was shown not to be positivelyor negatively correlated with any of these biomarker measures,and was therefore not considered in subsequent analyses (data notshown). Possible differences in biomarker measures between maleand female English sole were tested by two-way analysis of vari-ance, using sex and capture date as factors (Zar, 1984); in all casesthere was no consistently significant difference between values formale and female fish over the sampling periods for any of the abovebiomarker measures, so sexes were combined in subsequent treat-ment of the data (data not shown). The critical level of statistical
significance in all statistical tests was set at p ≤ 0.05.Liver lesion occurrence was statistically related to biologicalrisk factors (fish age and sex), measures of contaminant expo-sure, and time elapsed since initiation of the sediment cappingprocess using stepwise logistic regression techniques, as reportedelsewhere (Johnson et al., 1993; Myers et al., 1994, 1998; Stehr etal., 1997). This type of analysis was done to determine the influ-ence of the PAH-exposure risk factors, as well as the time elapsedsince cap initiation on hepatic disease risk. Logistic regression tech-niques account for the influence of biological risk factors such as ageand sex, and are commonly used on binomial or proportional datain epidemiological and epizootiological studies (Breslow and Day,1980). Logistic models were fitted using the PECAN module of theEGRET statistical package, version III (Statistics and EpidemiologyResearch Corporation, Seattle, WA). Risk factors were consideredsignificant at p ≤ 0.05, with independent analyses performed forall lesion categories. Two types of analyses were performed. Thefirst determined the calculated odds ratio as an estimate of rela-tive risk for lesions in individual fish in relation to the variablesof fish sex and age, and time elapsed since initiation of the sedi-ment capping process in September 1993. In this analysis the odds
logy 88 (2008) 277–288 281
ratio represents the degree of association between a risk factor andlesion occurrence (Fleiss, 1981) as determined from variable coef-ficients of the logistic regression equations (Anderson et al., 1980;Schlesselman, 1982). Odds ratios for lesion categories at a particularsampling time point (months post-cap initiation) were calculatedand interpreted relative to the lesion occurrence at the September1993 time point just prior to the beginning of placement of the pri-mary sediment cap. Decreased probabilities of lesion occurrenceat the respective post-capping sampling points were indicated byodds ratios less than 1.000, and the overall regression could beexpressed in the form of the influence of each successive monthelapsed since cap initiation on the risk of lesion occurrence. Oddsratios for age were interpreted for the influence of each additionalyear in age on the risk of lesion occurrence. A second type of step-wise logistic regression analysis was also used to determine thesignificance of the relationship between hepatic lesion occurrenceand measures for discrete PAH-exposure risk factors (biliary FACs,hepatic CYP1A, xenobiotic–DNA adducts) in individual fish, whilesimultaneously adjusting for the potential influence of fish age andsex.
3. Results
3.1. Field results
Field samplings conducted within the zone adjacent to and westof the capping area in Eagle Harbor at 17 separate time pointsresulted in capture and tissue collection from 729 English sole forhistopathology, 255 for DNA adducts, 343 for CYP1A and 553 forbile FACs (Table 1). In most cases, at least 15 samples for eachbiomarker measurement were obtained at the respective samplingpoints. However, in some cases we were not able to capture suffi-cient numbers of fish to meet this target, and not all fish sampledcontained sufficient bile for analysis (Table 1).
Subsequent to placement of Snohomish River dredge material inthe capped area, we observed a dramatic increase in the proportionof the trawl contents represented by wood chips and small logs. Thischange was presumed to be due to the wood debris present in theSnohomish River dredge material.
3.2. Biomarkers of PAH exposure in English sole
3.2.1. Hepatic CYP1A
Mean hepatic CYP1A activity in English sole captured just priorto initiation of sediment capping was substantially lower than his-torical values determined in 1988 and 1991 (Myers et al., 1998;Collier, previously unpublished data) (Fig. 3a). Activities droppedsomewhat, but not significantly, in the March 1994 sampling (0.5-year post-capping), but rose to levels significantly higher thanpre-capping levels in all subsequent samplings through May of 1999(5.7 years), as compared to the mean activity for the September1993 (time zero) sampling prior to sediment capping (Fig. 3a). Therewas no strongly significant trend toward increasing or decreasingCYP1A activities subsequent to primary capping, as determined bylinear regression (p = 0.1292) (Fig. 3b). Mean activities in samplingssubsequent to March 1994 were typically within the range of val-ues determined for English sole sampled in Eagle Harbor in 1988and 1991. Hepatic CYP1A activities in English sole from Eagle Har-bor in all samplings remained substantially above mean activities(116 pmol/(mg min)) measured in English sole captured at six sep-arate samplings in 1987–1988 from a commonly used reference, orrelatively uncontaminated site in Puget Sound (Polnell Point) (Steinet al., 1992; Collier et al., 1995). Because of the lack of temporal trendin hepatic CYP1A levels subsequent to sediment capping through

282 M.S. Myers et al. / Aquatic Toxicology 88 (2008) 277–288
Fig. 3. (a) CYP1A levels estimated as mean hepatic aryl hydrocarbon hydroxy-lase (AHH) activities (pmol/(mg min)) in English sole sampled from Eagle Harborbetween 9/93 and 5/99. Error bars indicate ± 1 standard error of the mean. Datapoints above the 9/93 sampling point represent values in English sole from EagleHarbor in 1988 and 1991. Also shown are hepatic CYP1A values typical of Englishsole from reference areas in Puget Sound (data from Stein et al., 1992; Collier etal., 1995). (b) Linear regression of log-transformed CYP1A levels estimated as meanhepatic aryl hydrocarbon hydroxylase (AHH) activities (pmol/(mg min)) in Englishsole sampled over time from Eagle Harbor between 9/93 and 5/99, from the samedata as shown in (a). No significant increasing or decreasing trend in CYP1A levelswere shown over this time period (p = 0.1386; Y = 2.59 + 0.016X; R2 = 0.006).
May of 1999 and the high sensitivity and lack of PAH-specificityof this biomarker (Stegeman and Hahn, 1994; Collier et al., 1995),analyses for this biomarker were not done past this time point.
3.2.2. Fluorescent aromatic compounds (FACs) in bilePAH metabolites in bile were analyzed at two wavelength pairs,
one estimating exposure to and metabolism of high molecularweight PAHs (benzo[a]pyrene or BaP wavelengths) and the other
for low molecular weight PAHs (naphthalene or NPH wavelengths).However, because the BaP measure is more reflective of exposure tothe high molecular weight PAHs present in coal tar creosote (Malinset al., 1985; Krahn et al., 1987), and because these two measureswere so highly correlated in the present study (0.842) only the FACsdata for compounds fluorescing at BaP wavelengths (FACsBaP) arepresented here.Mean concentrations of biliary FACsBaP at the pre-capping sam-pling point were far below the peak levels determined in EagleHarbor English sole in 1984 (Malins et al., 1985), and lower thanin 1988 (Myers et al., 1998) and slightly lower than those measuredin 1991 (Thom, previously unpublished data) (Fig. 4a). At the firstpost-capping sampling point (March 1994, 0.5-year post-capping),the mean concentration of FACsBaP in bile was significantly lowerthan concentrations measured in sole sampled at the pre-cappingsampling point, but by July 1994 (0.8-year post-capping) levelswere significantly higher than the pre-capping values. In sub-sequent sampling time points through October 1995 (2.1-yearpost-capping), mean concentrations of biliary FACsBaP were highlyvariable, ranging from ∼250 to ∼1300 ng BaP equiv./mg bile pro-tein, suggesting that English sole from Eagle Harbor were exposed
Fig. 4. (a) Mean concentrations (ng BaP equiv./mg bile protein) of fluorescent aro-matic compounds measured at benzo(a)pyrene wavelengths (FACsBaP) in bile ofEnglish sole sampled from Eagle Harbor between 9/93 and 4/04. Error bars indi-cate ± 1 standard error of the mean. Data points above the 9/93 sampling pointrepresent mean values in English sole from Eagle Harbor in 1988 and 1991. Alsoshown are biliary FACsBaP values typical of English sole from reference areas in PugetSound (data from Stein et al., 1992; Collier et al., 1995; Myers et al., 1998). (b) Lin-ear regression of log-transformed FACsBaP levels in bile of English sole sampled overtime from Eagle Harbor between 9/93 and 4/04, from the same data as shown in(a). A highly significant (p < 0.0001; Y = 2.717 − 0.003X; R2 = 0.107) temporal trend ofdecreasing values was shown over this time period extending to April 2004, or 128months after the initiation of sediment capping activities.
to varying concentrations of high molecular weight PAHs for up to2 years following cessation of sediment capping operations. How-ever, subsequent to this sampling point, there was a sharp declinein FACsBaP in bile until December 1997 (4.3-year post-capping),with levels remaining relatively constant through April 2004 (10.7-year post-capping), with only a slight increase since May 1998(4.7-year post-capping). Over the entire time course of the study,there has been a significant decline in FACsBaP in bile subsequent
to the September 1993 sampling as determined by linear regres-sion (p < 0.0001) (Fig. 4b). However, the relatively low mean FACsBaPconcentrations in Eagle Harbor English sole subsequent to theDecember 1997 (4.3-year post-capping) sampling still remainedslightly above the mean concentrations (162 ng BaP equiv./mg bileprotein) measured in English sole during six separate samplings in1987–1988 at the Polnell Point reference site (Myers et al., 1998).3.2.3. Hepatic xenobiotic–DNA adductsMean adduct concentrations at the pre-capping sampling in
September 1993 (70 ± 8 nmol DNA adducts/mol DNA bases) werecomparable to those measured in Eagle Harbor English sole in1992 (WL Reichert, previously unpublished data), but higher thanthose measured in 1988 (Stein et al., 1992) (Fig. 5a). Because ofanalytical cost considerations, hepatic xenobiotic–DNA adductconcentrations were determined only at the March 1994 (0.5-yearpost-capping), September 1994 (1 year), March 1995 (1.5 years),October 1995 (2.1 years), May 1998 (4.7 years), May 1999 (5.7years), April 2000 (6.6 years), May 2001 (7.7 years), May 2002(8.7 years) and April 2004 (10.7 years) samplings (Fig. 5a). Thesedata show a progressive decrease in adduct concentrations over

M.S. Myers et al. / Aquatic Toxicology 88 (2008) 277–288 283
Table 2Sample sizes, mean ages, and prevalences (percent affected) of toxicopathic hepaticlesions in English sole from Eagle Harbor, WA at sampling points between September1993 and April 2004, with comparisons to lesion prevalences in Eagle Harbor from1983–1986 (from data in Myers et al., 1987)
Fig. 5. (a) Mean concentrations (nmol adducts/mol bases) of hydrophobicxenobiotic–DNA adducts in liver of English sole sampled from Eagle Harbor at vari-ous time points between 9/93 and 4/04. Error bars indicate ± 1 standard error of themean. Data points below and to the left of the 9/93 sampling point represent meanvalues in English sole from Eagle Harbor in 1988, and 1992. Also shown are DNAadduct levels typical of English sole from reference areas in Puget Sound (Reichertet al., 1998; French et al., 1996; Reichert, unpublished data). (b) Linear regressionof untransformed levels of bulky hydrophobic xenobiotic–DNA adducts in bile ofEnglish sole sampled over time from Eagle Harbor between 9/93 and 4/04, fromthe same data as shown in (a). A highly significant (p < 0.0001; Y = 54.313 − 0.444X;R2 = 0.322) temporal trend of decreasing values was shown over this time periodextending to April 2004, or 10.7 years (128 months) after the initiation of sedimentcapping activities.
time. Significantly lower hepatic DNA adduct concentrations wereobserved at all of the post-capping sampling points relative topre-capping values (p ≤ 0.05), with the April 2004 values beingindistinguishable from adduct levels in sole from reference areas(Stein et al., 1992; French et al., 1996; WL Reichert, previouslyunpublished data;) such as in Tulalip Bay (7 nmol adducts/molbases) and Useless Bay (4 nmol adducts/mol bases) in 1991. Thisgenerally decreasing trend was confirmed as highly significant by
linear regression (p < 0.0001) (Fig. 5b).3.2.4. Toxicopathic hepatic lesionsHepatic lesion prevalences at the September 1993 pre-capping
sampling were substantially below those in adult English solefrom Eagle Harbor in 1983–1986 (Table 2). This was especiallytrue for hepatic neoplasms, which were detected in only 1% ofthe sole examined, as compared to 26% in the 1983–1986 sam-plings. The lesion most commonly detected in September 1993 (asin 1983–1985) was HM/NP (16%), followed by PROLIF (10%), PRE-NEO (5%), and NEO (1%), with prevalences for all of these categoriessubstantially below the 1983–1986 values. Overall, the prevalencefor the “any toxicopathic lesion” category (TOXLIV) was 25% in theSept. 1993 sampling, as compared to 83% in the 1983–1986 stud-ies. However, this reduction in lesion prevalence could be partiallydue to the relatively low mean age (3.4 years) of fish examined inthe September 1993 sampling, as compared to 5.2 for sole fromthe 1983–1986 studies. Age has been determined to be a strongrisk factor for most of the toxicopathic liver lesion types, espe-cially preneoplastic lesions and neoplasms (Rhodes et al., 1987;Myers et al., 1994, 1998, 2003). Overall, hepatic lesion prevalence
Date(s) N AGE HM/NP PROLIF PRENEO NEO TOXLIV
1983–1986 151 5.2 77 39 36 26 82September 1993 73 3.4 16 10 5 1 25March 1994 21 8.5 52 10 24 5 62July 1994 30 6.6 40 23 20 3 47September 1994 37 4.2 14 3 0 0 14March 1995 32 7.0 6 3 16 3 19July 1995 19 7.1 37 16 21 5 58October–November 1995 21 3.9 29 10 19 10 33December 1995 6 3.5 33 0 0 0 33October 1996 19 5.5 21 5 16 0 32December 1997 26 6.5 0 0 4 12 12May 1998 73 5.2 3 3 3 1 7May 1999 72 5.6 4 0 1 3 8April 2000 60 4.1 0 0 2 2 3May 2001 60 4.7 3 0 3 0 7May 2002 60 5.1 0 2 3 2 3April 2003 60 4.8 10 2 2 7 17April 2004 60 4.7 7 5 3 0 7
HM/NP = hepatocellular megalocytosis/nuclear pleomorphism; PROLIF = nonneo-plastic proliferative lesions; PRENEO = putatively preneoplastic foci of cellular alter-ation; NEO = neoplasms; TOXLIV = any toxicopathic liver lesion.
showed a reduction between 1983 and 1986 and cap initiation, fol-lowed by an approximate 3-year period of highly variable lesionprevalences, and then a sharp decline in TOXLIV prevalences sinceOctober 1996 (3.1-year post-capping) to values consistently below20%, and typically below 10%, through April of 2004.
Mean fish ages among the different sampling periods were quitevariable throughout the study, with individual ages ranging from 1to 17 years, and mean ages for the separate sampling points rangedfrom 3.4 to 8.5 years (Table 2). Consequently, it was not possible toaccurately compare basic lesion prevalences among the samplingperiods. To account and adjust for these age differences and thestrong influence of fish age on risk of toxicopathic liver lesion occur-rence, we utilized stepwise logistic regression to (1) determine theinfluence of fish age on lesion risk, followed by (2) determiningthe influence of the period of elapsed time since capping on riskof hepatic lesion occurrence, or (3) the influence on lesion risk ofchemical risk factors such as the PAH-exposure biomarkers mea-sured. The net effect is to be able to compare fish of equivalentages among the different sampling periods, or in the examination
of chemical risk factors such as the PAH-exposure risk factors. Wealso used logistic regression to assess the influence of sex on thelikelihood of hepatic lesion occurrence. Sex was not a significantrisk factor for any of the hepatic lesion categories, including theinclusive TOXLIV category (data not shown). Therefore, both sexeswere combined in the subsequent logistic regression analyses.In the analysis above, the risk of occurrence of lesions in theTOXLIV category, relative to the pre-capping risk (referred to asrelative risk), was highly variable for approximately 3 years fol-lowing cap initiation. Relative risks ranged from 0.209 at 1.5-yearpost-cap initiation to 1.690 at 1.8-year post-capping (Fig. 6). At the3-year post-capping sampling point the relative risk was 0.793,with relative risks subsequent to this time point dropping to val-ues consistently well below 0.500, and ranging between 0.0597 at8.7-year post-capping to 0.397 at 9.7 years after cap initiation. Infact, all relative risks for TOXLIV occurrence subsequent to the 3-year post-capping sampling point (October 1996) and up throughthe final April 2004 (10.7 years) sampling have been significantlylower (p ≤ 0.05), than the September 1993 baseline value of 1.000.
Relative risks for individual lesion categories showed similartemporal patterns to the inclusive lesion category, TOXLIV. The

284 M.S. Myers et al. / Aquatic Toxicology 88 (2008) 277–288
ivid
ual
Engl
ish
sole
capt
ure
dfr
omEa
gle
Har
bor,
WA
atsa
mp
lin
gp
erio
ds
from
Oct
ober
1996
(36-
mon
thp
ost-
cap
init
iati
on)t
hro
ugh
Ap
ril2
004
(128
-mon
th
May
1999
Ap
ril2
00
0M
ay20
01M
ay20
02A
pri
l20
03A
pri
l20
04
(16)
1983
NEO
(8)
1992
PRE
(13)
1988
PRE
(9)
1993
NEO
,PR
EP-
HR
(11)
1992
NEO
(9)
1995
NP,
PRE
(15)
1984
NO
(6)
1994
NEO
(4)
1997
NP
(4)
1998
PRE
(9)
1994
NP
(7)
1997
HM
/NP
P-H
R(1
3)19
86N
EO(3
)19
98PR
E(8
)19
95N
EO(7
)19
96N
EO(7
)19
97H
M/N
PPR
E,P-
CF
(12)
1987
NP
(3)
1998
HM
/NP
(6)
1997
PRE
(5)
1995
HM
/NP,
P-H
R(8
)19
91H
M/N
P(6
)19
97N
EO,N
P(7
)19
92PR
E(4
)19
99N
P(4
)19
99N
P(3
)20
00
NP,
P-H
R
N=
6(8
%)
N=
2(3
%)
N=
4(7
%)
N=
2(3
%)
N=
10(1
7%)
N=
4(7
%)
llu
lar
nu
clea
rp
leom
orp
his
m;
HM
=h
epat
icm
egal
ocyt
osis
;H
M/N
P=
com
bin
edh
epat
ocel
lula
rn
ucl
ear
ple
omor
ph
ism
and
hep
atic
meg
aloc
ytos
is;
P-P-
CF
=n
onn
eop
last
icp
roli
fera
tive
lesi
ons,
chol
angi
ofibr
osis
;PR
E=
pre
neo
pla
stic
foca
lle
sion
s;N
EO=
neo
pla
sms.
Age
sfo
rin
div
idu
alfi
shar
esh
own
in,1
986,
1992
,etc
.Th
efi
nal
row
inth
ista
ble
rep
rese
nts
the
nu
mbe
r(N
)of
fish
from
the
par
ticu
lar
sam
pli
ng
that
wer
eaf
fect
edw
ith
any
toxi
cop
ath
icliv
er
Fig. 6. Estimated odds ratios or relative risks of toxicopathic hepatic lesion (TOXLIV)occurrence in English sole from Eagle Harbor over the period 1983 through April2004. The risk of toxicopathic lesion occurrence is defined as 1.000 at the start ofsediment capping activities in September 1993. Relative risks at other samplingpoints are determined by stepwise logistic regression, after first accounting for theinfluence of fish age on risk of lesion occurrence, which in this analysis showed asignificantly increased risk of 1.317× for each additional year of age. The relativerisk of lesion occurrence for the period 1983–1985 (∼3.0) is estimated based onmean age and hepatic lesion prevalences for English sole at sampling points overthat period. The decreasing temporal trend line (gray line) is determined by stepwiselogistic regression, and is highly significant (p < 0.001), with risk of lesion occurrencedecreasing by 0.9813 times for each month after initiation of sediment capping.Asterisks indicate that the relative risk for that particular sampling time point issignificantly lower than that of the baseline risk determined for September 1993, bystepwise logistic regression.
most common lesion type, HM/NP, showed highly variable rela-tive risks (0.186–2.835) for the sampling periods through 3-yearpost-cap initiation, followed by a sharp decline past that pointto values consistently below 0.500, and ranging from near-zeroto 0.458 (other data not shown). Relative risks for PROLIF werealso highly variable through the 2-year post-capping sampling(0.254–2.479), and have remained consistently below 0.500 sinceOctober–November 1995, ranging from near-zero to 0.468 (otherdata not shown). Relative risks for PRENEO were also highly variable(near-zero to 3.205) through the 3-year post-capping sampling,followed by a sharp reduction to values consistently below 0.400(0.073–0.397) through April 2004 (other data not shown). Hepaticneoplasms were the least common hepatic lesion type throughoutthe study, and showed a slightly different temporal pattern, withhighly variable relative risks throughout the study (near zero to2.982), with no temporal pattern (other data not shown). This dif-ferent pattern than that shown for the other toxicopathic lesions
is most likely an artifact of the very low prevalence detected inthe baseline pre-capping sampling point (1% in September 1993).For example, in both cases where the relative risk approached 3.0(October–November 1995 and April 2003), the actual prevalenceswere still ≤10%. In addition, the lack of a clear temporal pattern inthe relative risk for neoplasm occurrence may be due to the pre-sumed irreversibility of these lesions (Boorman et al., 1997). Evenin an environment of reduced PAH exposure, if neoplasms werealready present in a cohort or collection of age classes of exposedEnglish sole, their occurrence would not be likely to decrease sub-stantially over the remaining lifetime in that cohort.The improvement in health of English sole as measured by thedecline in occurrence of toxicopathic hepatic lesions since the ini-tiation of sediment capping operations in Eagle Harbor becameeven more apparent when we conducted an analysis of the agesof affected fish and estimated the date of birth (“birthdate”) ofaffected fish, while also taking into account the types of hepaticlesions represented in the individually affected sole (Table 3). In thisanalysis several major assumptions were made. First, we assumedthat English sole captured within Eagle Harbor have resided con-tinuously in Eagle Harbor for their entire lifetime, having settled Ta
ble
3A
ges
(in
year
s),e
stim
ated
year
clas
ses,
and
hep
atic
lesi
ons
affe
ctin
gin
dp
ost-
cap
init
iati
on)
Sam
pli
ng
dat
e
Oct
ober
1996
Dec
embe
r19
97M
ay19
98
(11)
1985
,PR
E(1
7)19
80N
EO(1
6)19
82N
EO,P
RE
(10)
1986
,PR
E(9
)19
86N
EO(1
6)19
82P-
CF
(8)
1988
NP
(6)
1991
NEO
(10)
1988
HM
/NP
(5)
1991
,NP
(6)
1992
HM
/NP
(5)
1991
PRE
(3)
1995
PRE
(4)
1992
HM
/NP,
P-H
R
Tota
lN
=6
(32%
)N
=3
(12%
)N
=5
(7%
)
Abb
revi
atio
ns
for
hep
atic
lesi
onty
pes
are
asfo
llow
s:N
P=
hep
atoc
eH
R=
non
neo
pla
stic
pro
life
rati
vele
sion
s,h
epat
ocel
lula
rre
gen
erat
ion
;p
aren
thes
is;
esti
mat
edye
arcl
asse
sor
year
ofbi
rth
are
show
nas
1985
lesi
on,a
nd
the
over
allp
reva
len
cefo
rth
atu
niq
ue
sam
pli
ng.
Toxicology 88 (2008) 277–288 285
M.S. Myers et al. / Aquaticout after spawning, fertilization and metamorphosis at a nearby,but as yet-undetermined spawning area in the vicinity of EagleHarbor. Available data from acoustic tagging studies in 2003–2005(O’Neill et al., 2005) suggest that a substantial proportion of adultEnglish sole do reside nearly continuously in Eagle Harbor, with theexception of making annual spawning migrations to an as-yet unde-termined spawning area nearby. We also assumed that neoplasmsin the liver of English sole are irreversible lesions, consistent withprinciples of comparative oncology (Robbins et al., 1984; Boormanet al., 1997), and that the time frame for their induction in wildEnglish sole is approximately 3 years (Rhodes et al., 1987; Myers etal., 2003). Likewise, preneoplastic focal lesions are rarely induciblewithin a 1-year time frame, and more commonly within a 2-yearperiod, and HM/NP and hepatocellular regenerative lesions (PRO-LIF) occur both in young-of-the-year wild English sole (Rhodes etal., 1987; Myers et al., 1998). From the combined age, estimateddate of birth and lesion type occurrence data in Table 3, of the42 English sole affected with toxicopathic liver lesions at samplingperiods subsequent to the periods showing highly variable fluctu-ations in hepatic lesion risk (March 1994 through December 1995),a large proportion of these lesions could easily be attributed toPAH exposure in Eagle Harbor prior to the sampling time pointwhere biliary FACsBaP and xenobiotic–DNA adducts levels clearlybegan a steady decline, which was October of 1995 (Figs. 4a and 5a,respectively). For example, of the 22 affected sole captured in thesampling periods between October 1996 and April 2000, 20 indi-viduals (90%) were of year classes where lesion induction can beexplained by PAH exposure prior to October of 1995; in fact all these20 cases could also be explained by PAH exposure prior to Septem-ber 1993. Only two fish, one of which had a “birthdate” of 1994and was affected by a neoplasm, and another “born” in 1995 andaffected by a preneoplastic focal lesion, had lesions that could beexplained by PAH exposure subsequent to September 1993, and thelatter fish had a lesion (preneoplastic focal lesion) whose inductioncould be explained by PAH exposure prior to October 1995. In sam-pling periods after April 2000, a much smaller proportion of the 20sole affected with liver lesions had lesions that could be attributedto exposure prior to either September 1993 or October 1995; onlythree fish fell into this category, with the remaining 17 having lesiontypes and “birthdates” consistent with lesion induction being dueto PAH exposure after October of 1995.
Therefore, of the 42 English sole detected with toxicopathic liverlesions subsequent to December 1995, 23 (55%) of these had lesiontypes and were of year classes consistent with induction by PAH
exposure prior to October of 1995. Overall, the prevalence of Englishsole, sampled between October 1996 and April 2004, with toxi-copathic liver lesions whose induction can be attributed to PAHexposure in Eagle Harbor subsequent to October 1995, was in factonly 3.9%, providing further evidence of the efficacy of sedimentremediation measures in reducing PAH exposure and associatedoccurrence of toxicopathic liver lesions in Eagle Harbor English sole.3.3. Relationship between PAH-exposure biomarkers andtoxicopathic hepatic lesion occurrence
The statistical association between exposure to PAHs and occur-rence of the toxicopathic liver lesions was tested, in individual fish,by stepwise logistic regressions. Fish age was first accounted forin the regression, followed by an assessment of the influence onlesion risk of levels of one of the PAH-exposure biomarkers. Thisallowed us the unique opportunity to assess the influence of eachbiomarker of PAH exposure on the risk of hepatic lesion exposure inindividual fish, as opposed to comparing lesion prevalences in onegroup of fish to mean biomarker values in separate fish that werecaptured at the same sampling.
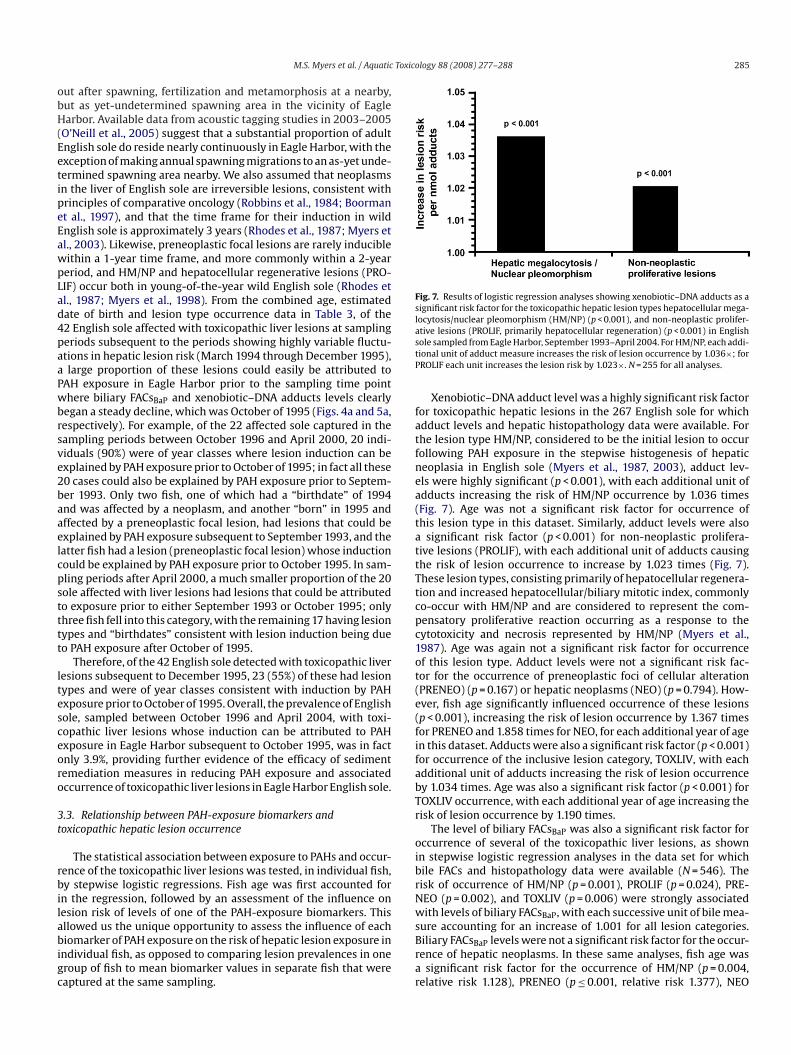
Fig. 7. Results of logistic regression analyses showing xenobiotic–DNA adducts as asignificant risk factor for the toxicopathic hepatic lesion types hepatocellular mega-locytosis/nuclear pleomorphism (HM/NP) (p < 0.001), and non-neoplastic prolifer-ative lesions (PROLIF, primarily hepatocellular regeneration) (p < 0.001) in Englishsole sampled from Eagle Harbor, September 1993–April 2004. For HM/NP, each addi-tional unit of adduct measure increases the risk of lesion occurrence by 1.036×; forPROLIF each unit increases the lesion risk by 1.023×. N = 255 for all analyses.
Xenobiotic–DNA adduct level was a highly significant risk factorfor toxicopathic hepatic lesions in the 267 English sole for whichadduct levels and hepatic histopathology data were available. Forthe lesion type HM/NP, considered to be the initial lesion to occurfollowing PAH exposure in the stepwise histogenesis of hepaticneoplasia in English sole (Myers et al., 1987, 2003), adduct lev-els were highly significant (p < 0.001), with each additional unit ofadducts increasing the risk of HM/NP occurrence by 1.036 times(Fig. 7). Age was not a significant risk factor for occurrence ofthis lesion type in this dataset. Similarly, adduct levels were alsoa significant risk factor (p < 0.001) for non-neoplastic prolifera-tive lesions (PROLIF), with each additional unit of adducts causingthe risk of lesion occurrence to increase by 1.023 times (Fig. 7).These lesion types, consisting primarily of hepatocellular regenera-tion and increased hepatocellular/biliary mitotic index, commonlyco-occur with HM/NP and are considered to represent the com-pensatory proliferative reaction occurring as a response to thecytotoxicity and necrosis represented by HM/NP (Myers et al.,1987). Age was again not a significant risk factor for occurrenceof this lesion type. Adduct levels were not a significant risk fac-tor for the occurrence of preneoplastic foci of cellular alteration(PRENEO) (p = 0.167) or hepatic neoplasms (NEO) (p = 0.794). How-
ever, fish age significantly influenced occurrence of these lesions(p < 0.001), increasing the risk of lesion occurrence by 1.367 timesfor PRENEO and 1.858 times for NEO, for each additional year of agein this dataset. Adducts were also a significant risk factor (p < 0.001)for occurrence of the inclusive lesion category, TOXLIV, with eachadditional unit of adducts increasing the risk of lesion occurrenceby 1.034 times. Age was also a significant risk factor (p < 0.001) forTOXLIV occurrence, with each additional year of age increasing therisk of lesion occurrence by 1.190 times.The level of biliary FACsBaP was also a significant risk factor foroccurrence of several of the toxicopathic liver lesions, as shownin stepwise logistic regression analyses in the data set for whichbile FACs and histopathology data were available (N = 546). Therisk of occurrence of HM/NP (p = 0.001), PROLIF (p = 0.024), PRE-NEO (p = 0.002), and TOXLIV (p = 0.006) were strongly associatedwith levels of biliary FACsBaP, with each successive unit of bile mea-sure accounting for an increase of 1.001 for all lesion categories.Biliary FACsBaP levels were not a significant risk factor for the occur-rence of hepatic neoplasms. In these same analyses, fish age wasa significant risk factor for the occurrence of HM/NP (p = 0.004,relative risk 1.128), PRENEO (p ≤ 0.001, relative risk 1.377), NEO

Toxic
286 M.S. Myers et al. / Aquatic(p ≤ 0.001, relative risk 1.623) and TOXLIV (p ≤ 0.001, relative risk1.245).
Hepatic CYP1A levels, as estimated by AHH activities, were notsignificant risk factors for occurrence of any of the toxicopathichepatic lesion categories in the dataset of 340 English sole forwhich hepatic CYP1A and histopathology data were available. Theseresults strongly suggest that hepatic CYP1A levels are not asso-ciated with hepatic lesion occurrence in individual adult Englishsole, and that hepatic CYP1A induction is not a specific measureof PAH exposure. Moreover, the correlations were extremely poorbetween CYP1A levels and both biliary FACsBaP (−0.025) and hep-atic xenobiotic–DNA adduct (−0.031) levels.
4. Discussion
This investigation clearly demonstrated that the sequentialplacement of two major sediment caps over a large portion of east-ern Eagle Harbor contributed towards an overall reduction in PAHexposure, and a consequent reduction in occurrence of toxicopathichepatic lesions, in English sole captured within Eagle Harbor. How-ever, factors other than exposure associated with the capped areamay have also been important in determining total PAH exposureand associated biological effects in resident English sole. Generally,indicators of short-term exposure to PAHs (e.g., CYP1A inductionand concentrations of FACs in bile) were considerably reduced inEnglish sole from Eagle Harbor even before cap placement began, incomparison to historical values. This indicates that previous sourcecontrol measures enacted between 1988 and September 1993, suchas closure of the facility which pressure-treated wood pilings withcreosote, and control of upland and groundwater PAH contamina-tion via the on-site groundwater extraction and treatment system,likely also contributed to reduced PAH exposure in English soleliving in Eagle Harbor.
Over the first 2 years since capping started in September 1993,the rapidly responsive biomarker of PAH exposure, biliary FACsBaP,showed a highly variable response, increasing at times to levelscomparable to and even higher than those present several yearsprior to sediment capping. However, subsequent to the highestlevels detected at the October–November 1995 sampling (2.1-year post-capping), FACsBaP levels have dropped considerably tolevels consistently below pre-capping values. This reduction hasbeen sufficient to overcome the highly variable response withinthe first 2-year post-capping, and result in an overall, highly sig-nificant decrease from September 1993 to the final sampling in
April 2004. With the exception of the May 2001 sampling (7.7-year post-capping), the biliary FACsBaP levels since December 1997(4.3-year post-capping) have remained no higher than 30–50%of the pre-capping levels. However, a slight increase in levelsbetween 4.3-year post-capping and 10.7-year post-capping, pointsto the necessity of continued monitoring of this PAH exposurebiomarker in Eagle Harbor English sole. This continued monitor-ing is especially advisable considering the intermittent operationof the aging onshore wastewater treatment facility, and the esti-mated existence of almost 1 million gal of remaining free creosotein the soils underneath the former Wyckoff creosoting facility(http://yosemite.epa.gov/R10/CLEANUP.NSF/sites/Wyckoff).Hepatic CYP1A level, as estimated by AHH activity, is also arapidly responsive indicator of PAH exposure. Although levels ofthis biomarker were also reduced at the pre-capping samplingpoint as compared to historical levels in English sole, CYP1A lev-els were highly variable throughout the period in which CYP1Awas measured, and showed no overall temporal trend throughMay of 1999 (5.7-year post-capping). This lack of a downwardtrend in hepatic CYP1A occurred contemporaneously with a sig-nificantly decreasing trend in the other measured biomarkers of
ology 88 (2008) 277–288
PAH exposure and effect, strongly suggesting that hepatic CYP1Ais a nonspecific biomarker in this field monitoring situation. It iswell known that hepatic CYP1A is mediated by multiple chemi-cal groups whose metabolism is mediated via initial binding to theAh receptor (Stegeman and Hahn, 1994), such as PCBs and dioxins.Moreover, the introduction of bark and wood debris with associatednatural PAHs such as retene within the original capping materialmay have contributed to continued CYP1A induction (Nuno et al.,1998). Because of the lack of a relationship between CYP1A levelsand the other biomarkers of PAH exposure and effect, its use as amonitoring tool in Eagle Harbor was discontinued in 1999.
Hepatic DNA adducts have slow repair rates and a longer halflife, and are therefore a longer-term biomarker of PAH expo-sure (Anulacion et al., 1995; Reichert et al., 1998). Levels at thepre-capping sampling point were actually somewhat elevated com-pared to historical values in English sole (1988 and 1992). However,after that sampling point adduct levels have dropped in a highly sig-nificant and continuous trend, to near-reference values in the finalApril 2004 sampling. These findings clearly demonstrate that sed-iment capping has significantly reduced PAH exposure in Englishsole over the longer term, nearly 11 years.
The risk of toxicopathic hepatic liver lesion occurrence in EagleHarbor English sole is another longer-term biomarker of PAH expo-sure and effect (Myers et al., 2003). Lesion occurrence was reducedsubstantially (by 66%) at the pre-capping sampling as comparedto historical risk in English sole adults prior to any remediationmeasures in Eagle Harbor (1983–1986). This also suggests thatshore-based source controls and facility closure ameliorated hep-atic lesion risk. Subsequent to this sampling point, age-adjustedlesion risks were highly variable for up to 3 years, with relativerisks of toxicopathic lesion occurrence (compared to risk at theSeptember 1993 sampling point) ranging from 0.209 at 1.5-yearpost-cap initiation to 1.690 at 1.8-year post-capping. However, afterthe 3-year post-cap initiation sampling point, relative risks of lesionoccurrence for the inclusive lesion category, TOXLIV, and all of theunique lesion categories except NEO, have shown a substantial andconsistent reduction to risks typically between near 0 and 0.5, suchthat there exists a significant decreasing trend in toxicopathic lesionrisk after sediment cap initiation, with a reduction in risk of 0.9813times for each month after capping through April 2004. Overall,this substantial reduction in risk of lesion occurrence has stayedrelatively consistent over the last 8 years of the present study, andlow relative risks have continued for the years 2005, 2006 and 2007(MS Myers, unpublished observations).
This declining risk of liver lesion occurrence in English soleclosely parallels the reduction in biomarkers of PAH exposure inthe form of biliary FACsBaP and hepatic xenobiotic–DNA adducts, inthe same fish over the same time period. Moreover, the lesion riskfor the categories HM/NP, PROLIF, PRENEO, and TOXLIV was signif-icantly influenced by the PAH-exposure risk factor biliary FACsBaPlevels in a large dataset (N = 546). Xenobiotic–DNA adduct concen-tration was a significant risk factor for the occurrence of HM/NP,PROLIF, and the inclusive TOXLIV category in a smaller dataset ofvalues matched in the same individual fish (N = 267). The exis-tence of these PAH-exposure biomarkers as significant, positiverisk factors for the initial and more commonly occurring lesionsinvolved in the histogenesis of hepatic neoplasia in adult Englishsole (HM/NP, PROLIF, and PRENEO) parallels results in subadultEnglish sole (Myers et al., 1998). Data demonstrating the persis-tence of hepatic DNA adducts for up to 84 days after exposure toDuwamish River sediment extract or benzo[a]pyrene (Stein et al.,1995) and dose- and exposure time–responsiveness of DNA adductsin sole held for up to 5 weeks on PAH-amended sediments (Frenchet al., 1996) suggest that these adducts can represent cumulativechanges resulting from chronic exposure to and damage from geno-

Toxico
M.S. Myers et al. / Aquatictoxic compounds such as PAHs and their reactive intermediates.The strong and significant influence of DNA adduct concentra-tions on risk of occurrence of these earlier-occurring lesions in thislarge and robust dataset of English sole reinforces the importantroles of exposure to bulky aromatic compounds such as 4–5 ringPAHs and consequent formation of molecular lesions in the formof xenobiotic–DNA adducts in the etiology of these lesions. Overall,these findings parallel reductions in hepatic lesion risks and theother biomarkers of PAH exposure and effect. Along with the sig-nificant positive relationships between lesion occurrence and thesemeasures of PAH exposure in individual fish, these data form a lineof evidence that PAH exposure is a major causative factor in theetiology of these hepatic lesions (Myers et al., 2003).
Overall, the temporal trends showing parallel declines in hep-atic lesion risks, biliary FACs and hepatic DNA adducts, especiallysince 3 years after the initiation of capping activities, show that cap-ping of contaminated sediments and prior and continuing onshoresource control measures to reduce entry of creosote into Eagle Har-bor’s sediments have been effective in reducing PAH exposure andthe associated adverse health effect of hepatic lesions in residentEnglish sole.
A major lesson of this important biomonitoring study is thatno trends in decline of the measured PAH-exposure and effectparameters were apparent within an approximate time frame of3 years following the beginning of placement of the primary sedi-ment cap. However, longer term monitoring over at least a 5-yearperiod after sediment remediation was able to clearly demonstratethe efficacy of sediment remediation measures, as well as asso-ciated improvements in the health of a resident epibenthic fishspecies. Site remediation measures conducted just within the EastHarbor Operable Unit of the Wyckoff/Eagle Harbor Superfund sitehave cost over $120 million to date (personal communication, MaryJane Nearman, USEPA Region X). Therefore, it seems prudent toallocate a small fraction of the total remediation costs towards bio-logical monitoring of the health in living marine resources withinthe area remediated, as a relevant assessment of the efficacy ofthose remediation actions.
There were some limitations inherent in the current investi-gation. Because fish are able to move freely over the capped anduncapped areas of Eagle Harbor, and because we did not trawlwithin the capped area so as to not disturb the cap, the project couldnot be designed to precisely delineate reduced exposure specificallyin the capped area only. Our assumption throughout the study wasthat English sole captured in the adjacent trawl area are represen-
tative of English sole that reside in the capped area, and that there isconsiderable movement from the nearby capped areas to the trawlarea. This assumption is supported by recent studies using acous-tic tagging and subsequent tracking of tagged English sole in EagleHarbor, which have unequivocally shown that a large proportion ofadult English sole do in fact reside for long periods in the cappedareas of Eagle Harbor (O’Neill et al., 2005).The current investigation did not begin until after onshore reme-dial activities had been conducted. Additional questions regardingreduction of PAH exposure in the nearshore marine environmentcould likely have been answered had appropriate measurementsbeen made prior to these onshore activities. Nonetheless, the datapresented in this report provide a useful reference for evaluatingthe effectiveness of future efforts to restore essential habitat for liv-ing marine resources. Clearly, there are many areas of Eagle Harborperipheral to the capped area which are still contaminated withPAHs. We recommend that, as remedial actions are planned forthese other units, they be accompanied by biological surveys ofmarine biota as well as biochemical assessment of biota prior to,during, and subsequent to these actions. As agencies take actionsto redress past insults to marine resources resulting from chemical
logy 88 (2008) 277–288 287
contamination, the timely use of a combination of assessment toolswill allow for an informed and objective evaluation of the efficacyof such actions.
Acknowledgements
This study initially represented a cooperative effort betweenUSEPA Region 10 and NOAA’s National Marine Fisheries Service,Northwest Fisheries Science Center, Environmental ConservationDivision (ECD). We thank Ken Marcy, Bruce Duncan, Ellie Hale,and Mary Jane Nearman of EPA Region 10, for partial funding ofthis study in its early phase, and for providing sediment chemistrydata, maps, and photographs from their website. We also thankGarry Mayer of NOAA’s Restoration Center for partial funding of thisstudy in its early phase. Dan Lomax, Paul Plesha, Lyndal Johnson,Maryjean Willis, Herb Sanborn and Bill Mowitt of the NorthwestFisheries Science Center, and Jim Beam, Greg Lippert, Sandie O’Neill,Jim West, and Steve Quinnell of the Washington State Departmentof Fish and Wildlife, and Kurt Dobszinski of the FV CHASINA alsoassisted with boat operation, fish necropsy, and sample collection.We also thank Tom Lee and Jim Peacock of the Northwest FisheriesScience Center for preparation of histological slides and graph-ics assistance, respectively, and Johnathan Sivilay for determiningfish ages from otoliths. Thanks are also due Lyndal Johnson, JohnIncardona, and Nathaniel Scholz of the Northwest Fisheries ScienceCenter for reviewing the manuscript.
References
Anderson, S., Augier, A., Hauck, W.W., Oates, D., Vandaele, W., Weisburg, H.I. (Eds.),1980. Statistical Methods for Comparative Studies. John Wiley and Sons, NewYork.
Anulacion, B.F., Hom, T., Reichert, W.L., Collier, T.K., 1995. Temporal changes in CYP1A,DNA adducts, and contaminant residues in fish removed from contaminatedsites. In: Proceedings of the 2nd SETAC World Congress, Vancouver, BC, Canada,November 9, Abstract book, p. 96.
Baumann, P.C., Harshbarger, J.C., 1995. Decline in liver neoplasms in wild brownbullhead catfish after coking plant closes and environmental PAHs plummet.Environ. Health Perspect. 103, 168–170.
Baumann, P.C., Harshbarger, J.C., 1998. Long term trends in liver neoplasm epizooticsof brown bullhead in the Black River, Ohio. Environ. Monit. Assess. 53, 213–223.
Boorman, G.A., Botts, S., Bunton, T.E., Fournie, J.W., Harshbarger, J.C., Hawkins, W.E.,Hinton, D.E., Jokinen, M.P., Okihiro, M.S., Wolfe, M.J., 1997. Diagnostic criteriafor degenerative, inflammatory, proliferative nonneoplastic and neoplastic liverlesions in medaka (Oryzias latipes): consensus of a National Toxicology ProgramPathology Working Group. Toxicol. Pathol. 25, 202–210.
Breslow, N.E., Day, N.E., 1980. Statistical Methods in Cancer Research, Volume 1. TheAnalysis of Case-control Studies. International Agency for Research on Cancer,Lyon, France.
Casillas, E., Misitano, D., Johnson, L.L., Rhodes, L.D., Collier, T.K., Stein, J.E., McCain,
B.B., Varanasi, U., 1991. Inducibility of spawning and reproductive success offemale English sole (Parophrys vetulus) from urban and nonurban areas of PugetSound, Washington. Mar. Environ. Res. 31, 99–122.Chilton, D.E., Beamish, R.J., 1982. Age determination methods for fishes studied bythe Groundfish Program at the Pacific Biological Station. Can. Spec. Publ. Fish.Aquat. Sci. 60, 1–54.
Collier, T.K., Stein, J.E., Wallace, R.J., Varanasi, U., 1986. Xenobiotic metabolizingenzymes in spawning English sole (Parophrys vetulus) exposed to organic-solvent extracts of marine sediments from contaminated and reference areas.Comp. Biochem. Physiol. C 84, 291–298.
Collier, T.K., MacLeod, W.D. Jr., Krahn, M.M., 1989. Standard analytical proceduresof the NOAA National Analytical Facility: Protein in bile. Supplement to NOAATechnical Memorandum NMFS/F/NWC-102, 3 pp.
Collier, T.K., Varanasi, U., 1991. Hepatic activities of xenobiotic metabolizing enzymesand biliary levels of xenobiotics in English sole (Parophrys vetulus) exposed toenvironmental contaminants. Arch. Environ. Contam. Toxicol. 20, 462–473.
Collier, T.K., Connor, S.D., Eberhart, B-T.L., Anulacion, B.F., Goksøyr, A., Varanasi,U., 1992a. Using cytochrome P450 to monitor the aquatic environment: initialresults from regional and national surveys. Mar. Environ. Res. 34, 195–199.
Collier, T.K., Singh, S.V., Awasthi, Y.C., Varanasi, U., 1992b. Hepatic xenobioticmetabolizing enzymes in two species of benthic fish showing different preva-lences of contaminant-associated liver neoplasms. Toxicol. Appl. Pharmacol. 113,319–324.
Collier, T.K., Stein, J.E., Goksøyr, A., Myers, M.S., Gooch, J.W., Huggett, R.J., Varanasi,U., 1993a. Biomarkers of PAH exposure in oyster toadfish (Opsanis tau) from theElizabeth River, Virginia. Environ. Sci. 2, 161–177.
Collier, T.K., Krahn, M.M., Krone, C.A., Johnson, L.L., Myers, M.S., Chan, S.-L., Varanasi,U., 1993b. Oil exposure and effects in subtidal fish following the EXXON Valdez

Toxic
1999 Environmental Monitoring Report; Wyckoff/Eagle Harbor Superfund Site,
288 M.S. Myers et al. / Aquatic
oil spill. In: Proceedings of the 1993 International Oil Spill Conference. AmericanPetroleum Institute, pp. 301–305.
Collier, T.K., Anulacion, B.F., Stein, J.E., Goksøyr, A., Varanasi, U., 1995. A field evalu-ation of cytochrome P450 1A as a biomarker of contaminant exposure in threespecies of flatfish. Environ. Toxicol. Chem. 14, 143–152.
Fleiss, J.L., 1981. Statistical Methods for Rates and Proportions, 2nd ed. John Wileyand Sons, New York.
French, B.L., Reichert, W.L., Hom, T., Nishimoto, M., Sanborn, H.R., Stein, J.E., 1996.Accumulation and dose-response of hepatic DNA adducts in English sole (Pleure-onectes vetulus) exposed to a gradient of contaminated sediments. Aquat. Toxicol.36, 1–16.
Gray, P., 1954. The Microtomist’s Formulary and Guide. Blakiston, New York.Gupta, R.C., Randerath, K., 1988. Analysis of DNA adducts by 32P-labelling and thin
layer chromatography. In: Freidberg, E.C., Hanawalt, P.H. (Eds.), DNA Repair, vol.3. Marcel Dekker, New York, pp. 399–418.
Johnson, L.L., Stehr, C.M., Olson, O.P., Myers, M.S., Pierce, C.A., McCain, B.B., Varanasi,U., 1993. Chemical contaminants and hepatic lesions in winter flounder (Pleu-ronectes americanus) from the Northeast Coast of the United States. Environ. Sci.Technol. 27, 2759–2771.
Johnson, L.L., Casillas, E., Collier, T.K., McCain, B.B., Varanasi, U., 1988. Contaminanteffects on ovarian development in English sole (Parophrys vetulus) from PugetSound, Washington. Can. J. Fish. Aquat. Sci. 45, 2133–2146.
Krahn, M.M., Myers, M.S., Burrows, D.G., Malins, D.C., 1984. Determination ofmetabolites of xenobiotics in the bile of fish from polluted waterways. Xeno-biotica 14, 633–646.
Krahn, M.M., Rhodes, L.D., Myers, M.S., Moore, L.K., MacLeod Jr., W.D., Malins, D.C.,1986. Associations between metabolites of aromatic compounds in bile andthe occurrence of hepatic lesions in English sole (Parophrys vetulus) from PugetSound, Washington. Arch. Environ. Contam. Toxicol. 15, 61–67.
Krahn, M.M., Burrows, D.G., MacLeod Jr., W.D., Malins, D.C., 1987. Determinationof individual metabolites of aromatic compounds in hydrolyzed bile of Englishsole (Parophrys vetulus) from polluted sites in Puget Sound, Washington. Arch.Environ. Contam. Toxicol. 16, 511–522.
Krahn, M.M., Burrows, D.G., Ylitalo, G.M., Brown, D.W., Wigren, C.A., Collier, T.K.,Chan, S.-L., Varanasi, U., 1992. Mass spectrometric determination of metabo-lites of aromatic compounds in bile of fish captured from Prince William Sound,Alaska, after the EXXON Valdez oil spill. Environ. Sci. Technol. 26, 116–126.
Landahl, J.T., Johnson, L.L., 1993. Contaminant exposure and population growth ofEnglish sole in Puget Sound: the need for better early life history data. Am. Fish.Soc. Symp. 14, 117–123.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1951. Protein measurement withthe Folin phenol reagent. J. Biol. Chem. 193, 265–275.
Luna, L.G., 1968. Armed Forces Institute of Pathology: Manual of Histologic StainingMethods, 3rd ed. MacGraw-Hill, New York.
Malins, D.C., Krahn, M.M., Myers, M.S., Rhodes, L.D., Brown, D.W., Krone, C.A., McCain,B.B., Chan, S.-L., 1985. Toxic chemicals in sediments and biota from a creosote-polluted harbor: relationships with hepatic neoplasms and other hepatic lesionsin English sole (Parophrys vetulus). Carcinogenesis 6, 1463–1469.
Moore, M., Lefkovitz, L., Hall, M., Hillman, R., Mitchell, D., Burnett, J., 2005. Reductionin organic contaminant exposure and resultant hepatic hydopic vacuolation inwinter flounder (Pseudopleuronectes americanus) following improved effluentquality and relocation of the Boston sewage outfall into Massachusetts Bay, USA:1987–2003. Mar. Pollution Bull. 50, 156–166.
Myers, M.S., Rhodes, L.D., McCain, B.B., 1987. Pathologic anatomy and patterns ofoccurrence of hepatic neoplasms, putative preneoplastic lesions, and other idio-pathic hepatic conditions in English sole (Parophrys vetulus) from Puget Sound,Washington. J. Natl. Cancer Inst. 78, 333–363.
Myers, M.S., Stehr, C.M., Olson, O.P., Johnson, L.L., McCain, B.B., Chan, S.-L., Varanasi,U., 1994. Relationships between toxicopathic hepatic lesions and exposure tochemical contaminants in English sole (Pleuronectes vetulus), starry flounder(Platichthys stellatus), and white croaker (Genyonemus lineatus) from selectedmarine sites on the Pacific Coast, USA. Environ. Health Perspect. 102, 200–215.
Myers, M.S., Johnson, L.L., Hom, T., Collier, T.K., Stein, J.E., Varanasi, U., 1998. Tox-icopathic hepatic lesions in subadult English sole from Puget Sound, WA;Relationship to other indicators of contaminant exposure. Mar. Environ. Res.45, 47–67.
Myers, M.S., Johnson, L.L., Olson, O.P., Stehr, C.M., Horness, B.H., Collier, T.K., McCain,B.B., 1999. Toxicopathic hepatic lesions as biomarkers of chemical contaminantexposure and effects in marine bottomfish species from the Northeast and PacificCoasts, USA. Mar. Pollution Bull. 37, 92–113.
Myers, M.S., Johnson, L.L., Collier, T.K., 2003. Establishing the causal relationshipbetween polycyclic aromatic hydrocarbon (PAH) exposure and hepatic neo-plasms and neoplasia-related liver lesions in English sole (Pleuronectes vetulus).Hum. Ecol. Risk. Assess. 9, 67–94.
Nuno, M., Fragoso, N.M., Parrott, J.L., Hahn, M.E., Hodson, P.V., 1998. Chronic reteneexposure causes sustained induction of CYP1A activity and protein in rainbowtrout (Oncorhynchus mykiss). Environ. Toxicol. Chem. 17, 2347–2353.
O’Neill, S.M., Myers, M.S., Moser, M.L., Anulacion, B.F., Quinnell, S.R., West, J.E., 2005.Use of acoustic tagging to study home range and migration of English sole(Parophrys vetulus) in Puget Sound: application to management of contaminatedsediments. In: 2005 Puget Sound/Georgia Basin Research Conference 2005, Seat-tle, WA, March 29–31, Abstract Book, p. 91.
Payne, J.F., Penrose, W.R., 1975. Induction of aryl hydrocarbon benzo[a]pyrenehydroxylase in fish by petroleum. Bull. Environ. Contam. Toxicol. 14, 112–116.
ology 88 (2008) 277–288
Payne, J.F., Fancey, L.L., Rahimtula, A.D., Porter, E.L., 1987. Review and perspective onthe use of mixed-function oxygenase enzymes in biological monitoring. Comp.Biochem. Physiol. 86C, 233–245.
Reichert, W.L., Stein, J.E., French, B., Goodwin, P., Varanasi, U., 1992. Storage phosphorimaging technique for detection and quantitation of DNA adducts measured bythe 32P-postlabeling assay. Carcinogenesis 13, 1475–1479.
Reichert, W.L., French, B., 1994. The 32P-Postlabeling protocols for assaying lev-els of hydrophobic DNA adducts in fish. U.S. Dept. Commerce, NOAA TechnicalMemorandum NMFS-NWFSC-14, 89 pp.
Reichert, W.L., Myers, M.S., Peck-Miller, K., French, B., Anulacion, B.F., Collier, T.K.,Stein, J.E., Varanasi, U., 1998. Molecular epizootiology of genotoxic events inmarine fish: Linking contaminant exposure, DNA damage, and tissue-level alter-ations. Mutation Res. 411, 215–225.
Rhodes, L.D., Myers, M.S., Gronlund, W.D., McCain, B.B., 1987. Epizootic characteris-tics of hepatic and renal lesions in English sole (Parophrys vetulus) from PugetSound. J. Fish. Biol. 31 (3), 395–408.
Robbins, S.L., Cotran, R.S., Kumar, V., 1984. The Pathologic Basis of Disease, 3rd ed.W.B Saunders Company, Philadelphia.
Schiewe, M.S., Weber, D.D., Myers, M.S., Jacques, F.J., Reichert, W.L., Krone, C.A.,Malins, D.C., McCain, B.B., Chan, S.-L., Varanasi, U., 1991. Induction of foci ofcellular alteration and other hepatic lesions in English sole (Parophrys vetulus)exposed to an extract of an urban marine sediment. Can. J. Fish. Aquat. Sci. 48,1750–1760.
Schlesselman, J., 1982. Case-control Studies: Design, Conduct and Analysis. OxfordUniversity Press, New York.
Schnell, J.V., Gruger, E.H., Malins, D.C., 1980. Monooxygenase activities of cohosalmon (Oncorhynchus kisutch) liver microsomes using three polycyclic aromatichydrocarbon substrates. Xenobiotica 10, 229–234.
Sloan, C.A., Brown, D.W., Pearce, R.W., Boyer, R.H., Bolton, J.L., Burrows, D.G., Herman,D.P., Krahn, M.M., 2006. Quality assurance plan for analyses of environmen-tal samples for polycyclic aromatic compounds, persistent organic pollutants,fatty acids, stable isotope ratios, lipid classes, and metabolites of polycyclicaromatic compounds. U.S. Dept. Commerce, NOAA Technical MemorandumNMFS-NWFSC-77, 30 pp.
Stegeman, J.J., Hahn, M.E., 1994. Biochemistry and molecular biology of monooxyge-nases: current perspectives on forms, functions, and regulation of cytochromeP450 in aquatic species. In: Malins, D.C., Ostrander, G.K. (Eds.), Aquatic Tox-icology: Molecular, Biochemical and Cellular Perspectives. CRC Press, LewisPublishers, Boca Raton, pp. 87–206.
Stehr, C.M., Myers, M.S., Burrows, D.G., Krahn, M.M., Meador, J.P., McCain, B.B.,Varanasi, U., 1997. Chemical contamination and associated liver diseases in twospecies of fish from San Francisco Bay and Bodega Bay. Ecotoxicology 6, 35–65.
Stein, J.E., Collier, T.K., Reichert, W.L., Casillas, E., Hom, T., Varanasi, U., 1992. Bioindi-cators of contaminant exposure and sublethal effects: studies with benthic fishin Puget Sound, Washington. Environ. Toxicol. Chem. 11, 701–714.
Stein, J.E., Reichert, W.L., French, B., Varanasi, U., 1993. 32P-Postlabeling analysisof DNA adduct formation and persistence in English sole (Pleuronectes vetulus)exposed to benzo[a]pyrene and 7H-dibenzo[c,g]carbazole. Chem-Biol. Interact.88, 55–69.
Stein, J.E., Reichert, W.L., Varanasi, U., 1994. Molecular epizootiology: assessmentof exposure to genotoxic compounds in teleosts. Environ. Health Perspect. 102(Suppl. 12), 19–23.
Stein, J.E., Reichert, W.L., French, B.L., Anulacion, B.F., Varanasi, U., 1995. Toxicoki-netics of xenobiotic-DNA adducts in English sole (Pleuronectes vetulus): studieswith laboratory and environmentally exposed sole. Marine Environ. Res. 39, 373.
United States Environmental Protection Agency, U.S. Army Corps of Engineers, 1999.
East Harbor Operable Unit, Bainbridge Island, Washington. Draft, December 3,1999. Prepared for USEPA, Region 10 and U.S. Army Corps of Engineers, SeattleDistrict, by Striplin Environmental Associates, Inc. Olympia, WA 98502.
Van Cantfort, J., De Graeve, J., Gielen, J.E., 1977. Radioactive assay for aryl hydrocarbonhydroxylase. Improved method and biological importance. Biochem. Biophys.Res. Commun. 79.
Varanasi, U., Collier, T.K., Williams, D.E., Buhler, D.R., 1986. Hepatic cytochrome P-450enzymes and aryl hydrocarbon hydroxylase in English sole (Parophrys vetulus).Biochem. Pharmacol. 35, 2967–2971.
Varanasi, U., Stein, J.E., Nishimoto, M., 1989a. Biotransformation and disposition ofPAH in fish. In: Varanasi, U. (Ed.), Metabolism of Polycylic Aromatic Hydrocar-bons in the Aquatic Environment. CRC Uniscience Series. CRC Press, Inc., BocaRaton, pp. 93–150.
Varanasi, U., Reichert, W.L., Stein, J.E., 1989b. 32P-postlabeling analysis of DNAadducts in liver of wild English sole (Parophrys vetulus) and winter flounder(Pseudopleuronectes americanus). Cancer Res. 49, 1171–1177.
Varanasi, U., Stein, J.E., Johnson, L.L., Collier, T.K., Casillas, E., Myers, M.S., 1992. Evalu-ation of bioindicators of contaminant exposure and effects in coastal ecosystems.In: McKenzie, D.H., Hyatt, D.E., McDonald, V.J. (Eds.), Ecological Indicators, vol.1, Proceedings of International Symposium on Ecological Indicators. ElsevierApplied Science, pp. 461–498.
Vogelbein, W.K., Fournie, J.W., Van Veld, P.A., Huggett, R.J., 1990. Hepatic neoplasmsin the mummichog (Fundulus heteroclitus) from a creosote-contaminated site.Cancer Res. 50, 5978–5986.
Wolf, J.C., Wolfe, M.J., 2005. A brief overview of nonneoplastic hepatic toxicity infish. Toxicol. Pathol. 33, 75–85.
Zar, J.H., 1984. Biostatistical Analysis. Prentice-Hall, Englewood Cliffs, NJ, 620 pp.




















