
The effect of vitamin A on flatfish development and skeletogenesis: A review
16
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of The effect of vitamin A on flatfish development and skeletogenesis: A review

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Review
The effect of vitamin A on flatfish development and skeletogenesis: A review
Ignacio Fernández ⁎, Enric GisbertIRTA, Centre de Sant Carles de la Ràpita (IRTA-SCR), Unitat de Cultius Experimentals, Crta. del Poble Nou s/n, 43540-Sant Carles de la Ràpita, Spain
a b s t r a c ta r t i c l e i n f o
Article history:Received 7 October 2009Received in revised form 13 November 2010Accepted 19 November 2010Available online 26 November 2010
Keywords:DeformitiesFlatfishLarvaeJuvenilesBroodstockSkeletonMetamorphosisVitamin A
Flatfish can provide an excellent model to study developmental alterations of cartilage and bone tissues inresponse to vitamin A (VA), and therefore, with respect to the underlying morphogenesis, cell differentiationand proliferation processes, and gene expression regulation. Bone and cartilage development andhomeostasis at different life stages (embryonic, larval, juvenile or adult stages) are primarily regulated byretinoic acid, the active metabolite of VA. Since fish are not able to synthesize VA, they have to obtain it fromthe diet. Thus, dietary VA imbalance maymodify VA homeostasis, resulting in a disruption of gene expression,and cell proliferation and differentiation of skeletal tissues (bone and cartilage), leading to the appearance ofskeletal deformities. These skeletal deformities are one of the most important factors that affect productionefficiency and quality of flatfish aquaculture. However, different flatfish species present differentdevelopmental timing in skeletogenesis and consequently, different species may present different VArequirements. In the present work, special attention has been made to different experimental approaches toevaluate the effects of this morphogenetic nutrient on early morphogenesis (bath exposure vs. dietary dose–response nutritional essays), as well as the different retinoid forms in which VA was administered to larvae.We review the current state of knowledge on the effects of dietary VA content in flatfish development with anemphasis on the skeletal deformities induced by dietary VA imbalances.
© 2010 Elsevier B.V. All rights reserved.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 342. Flatfish skeletal development and metamorphosis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 353. Vitamin A metabolism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 364. Dietary VA effects on broodstock spawning performance and offspring quality . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 395. Dietary VA effects on fish larval development . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39
5.1. Larval bath exposure essays . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 395.2. Dietary dose–response essays in larvae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41
5.2.1. Studies with VA-enriched live preys . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 415.2.2. Developmental effects on flatfish larvae fed VA-enriched live preys . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42
6. Effects of dietary VA on juvenile development using compound diets . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 447. Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46
1. Introduction
Since 1970, interest on flatfish aquaculture has grown as theoverexploitation of natural resources and environmental degradationhave caused depletion of fisheries of several of these species, such asJapaneseflounder (Paralichthys olivaceus), summerflounder (P. dentatus),
turbot (Psetta maxima) and Atlantic halibut (Hippoglossus hippoglossus)(Bengtson, 1999). However, within marine fish aquaculture, worldflatfish farming is considered as a minor activity. In 2008, it onlyrepresented around 10% of the total production of marine fish species(1,511,087 tonnes; FAO, 2010). However, driven by the high marketprices in many Asian and European countries, where they are a culturalsource of food, flatfish production has increased from 26,310 to148,807 tonnes between 2000 and 2008, which represents an increaseof 565% in production and 50% in value (FAO, 2010).
Aquaculture 315 (2011) 34–48
⁎ Corresponding author. Tel.: +34 977745427; fax: +34 977443138.E-mail address: [email protected] (I. Fernández).
0044-8486/$ – see front matter © 2010 Elsevier B.V. All rights reserved.doi:10.1016/j.aquaculture.2010.11.025
Contents lists available at ScienceDirect
Aquaculture
j ourna l homepage: www.e lsev ie r.com/ locate /aqua-on l ine

Author's personal copy
During the last decade, flatfish production has been hampered by anarray of biological and physiological difficulties (Bengtson, 1999) suchas: pigmentary disorders (Bolker and Hill, 2000; Gavaia et al., 2002;Hamre et al., 2007); high mortality rate during the weaning feedingphase (Zarza et al., 2003); low quality and discontinuous spawning(Howell et al., 2009); and/or high incidence of skeletal deformities (e.g.Takeuchi et al., 1998; Gavaia et al., 2002; Fernández et al., 2009, amongothers). Skeletal deformities reduce the economic efficiency of the fishproduction process (Divanach et al., 1996). Hatcheries that providejuveniles grade outmalformed fishwhile, those farms that grow out thefish to market size have either to downgrade the product to a lowervalue or discard the deformed fish entirely (Michie, 2001). The extent oflosses at either point is different, dependent on the species and thehusbandry practices followed. In whatever circumstance, these lossesare substantial, in terms of both productivity and profitability. Hatcheryand nursery productions are mostly affected, since the majority ofskeletal structures develop during the larval and juvenile stages, inwhich organogenesis and morphogenesis take place. The developmentof skeletal disorders has been linked to a poorly understood relationshipbetween environmental and biotic factors (reviewed in Lall and Lewis-McCrea, 2007). Among these, larval nutrition atfirst feeding is oneof thekey parameters affecting skeletogenesis during early development.Several works have shown that suboptimal levels and different forms ofnutrients supplied in the diet are responsible for the appearance ofskeletal deformities (Cahu et al., 2003; Lall and Lewis-McCrea, 2007).Both environmental and nutritional requirements closely correlate tothe larval developmental stage and the skeleton ontogeny and anatomyof each fish species. Consequently, understanding the environmentalneeds and nutritional requirements of fish larvae is essential forimproving husbandry and rearing practices, and reducing the incidenceof skeletal disorders and their associated economical costs. However,while mammalian nutritional effects on bone development andremodelling have been widely studied, in fish these types of studiesare limited and depend on the nutrient and fish species underconsideration. Research on compound diet composition for marinefinfish larvae and enriching emulsions for live preys has revealed thatseveral nutrients, particularly minerals, vitamins, and lipids are criticalfor normal fish morphogenesis and skeletogenesis (Lall and Lewis-McCrea, 2007).
Retinoids, also known as Vitamin A (VA), are a group ofmorphogenetic nutrients that include all compounds that possessthe same biological activity as retinol, and since fish are not able tosynthesize VA, they have to obtain it from the diet at the optimumlevel, but also in the proper chemical form. In general, retinoids andtheir precursors (carotenoids) are absorbed from the diet by means ofa complex metabolism, which includes a high number of transportproteins and enzymes, that at the same time, tightly control retinoidabsorption, transport, accumulation and conversion to active forms.There are two primary active forms: (i) retinal, used as thechromophore of rhodopsin in the eye (Pepe, 1999); and (ii) retinoicacid (RA), the main active metabolite of VA that represents the othernon-visual functions of VA (Ross et al., 2000). RA binds to nuclear RAreceptors (retinoic acid receptors and retinoid X receptors, RARs andRXRs, respectively) and through them, regulates gene expression,cellular differentiation and proliferation processes (Ross et al., 2000),determining body pattern, growth, and the development of skeletalstructures, as well as being an important element in other developingsystems such as the nervous and immune systems (Fig. 1).
The impact of dietary VA on flatfish development and aquaculturewill depend on the following factors: the chemical form and dose ofVA in the diet, the duration of this supply, the fish developmentalstage, and the particular ontogeny of the species. Therefore, dietaryVA requirements need to be fine-tuned for each flatfish species,although they share similar skeletal structures and a commondevelopmental process from a symmetrical to an asymmetricalskeleton and shape (Watanabe et al., 2008). To date, different
approaches have been used to understand how VA affects differentaspects of aquaculture production: from adult fish (reproductiveperformance) to larvae and juveniles; although special emphasis hasbeen placed on flatfish skeletogenesis and pigmentation disorders,with larvae at first feeding being the developmental stage moststudied. This article reviews all the research conducted studyingdietary VA effects on skeletogenesis of different flatfish species.
2. Flatfish skeletal development and metamorphosis
Generally, marine fish larvae hatch much earlier than othervertebrates, and their development continues several days/monthsafterwards, representing a good biological model for studying theprocess of skeletogenesis (Haga et al., 2002a). The formation of theskeleton in Teleosts is quite different in their spatiotemporalsequences from those of higher vertebrates. In particular, fish larvaeundergo morphogenesis and skeletogenesis during their earlyontogeny, developing all the organs and systems that will lead thelarva into a juvenile, which in the case of flatfishes becomes morecomplicated due to their metamorphosis and bottom settlement. Athorough description of the metamorphosis process and its endocrineregulation is outside the scope of this review, but readers can consultrecent studies from Geffen et al. (2007), Power et al. (2008) andKlaren et al. (2008).
In brief, metamorphosis is a post-embryonic process, whichinvolves profound morphological, biochemical and physiologicaltransformations in order to adapt to a new habitat. In flatfish, it isan endocrine driven process in which thyroid hormones play a centralrole (Power et al., 2008), where fishes have a dramatic transformationfrom a bilateral symmetrical to an asymmetrical anatomy. Studies inzebrafish (Danio rerio) demonstrate that the nodal-lefty-pitx2 (NLP)pathway directs organ sidedness by promoting the expression ofthese genes on the left side of their embryonic primordia that inflounders occur irrespective of whether the species exhibits sinistralor dextral asymmetry (Hashimoto et al., 2007). Pitx2 is the final left–right determinant of the NLP pathway (Suzuki et al., 2009), as loss ofpitx2 re-expression induces randomization of eye-sidedness, mani-festing as normal, reversed or bilateral symmetry, with the lateralityof the structural asymmetry of the habenula entirely inverted inreversed flounders compared with normal ones. Perhaps the moststriking developmental feature in flatfishes (presumably evolved asan adaptation to their benthic existence) is the eye-sidedness. Themigration of one eye to the opposite side of the head requiresextensive rearrangement and remodelling of tissues, which occur atthe level of individual morphological elements, rather than as a globaldistortion of the entire chondrocranium (Martinez and Bolker, 2003).During metamorphosis, the fish skeleton experiences the followingmajor morphological changes: (i) a 90° rotation of the cranial andvisceral skeleton; (ii) eye migration to the ocular upper side resultingin neurocranium and splachnocranium reorganization; (iii) develop-ment of the pseudomesial bone during eye migration (Okada et al.,2003; Sæle et al., 2006a); and (iv) remodelling of the pectoral fin(Schreiber, 2006). The timing of this process varies among flatfishspecies and seems to depend on the different water temperatures atwhich they live. Osse and Van den Boogaart (1997) proposed twogeneral patterns of metamorphosis: (a) plaice-like, where metamor-phosis occurs at larger larval sizes (as in European plaice Pleuronectesplatessa and Atlantic halibut Hippoglossus hippoglossus); and (b) sole-like, in which metamorphosis takes place at smaller larval sizesand has a short duration (as in Senegalese sole Solea senegalensis).For example, while metamorphosis in Atlantic halibut starts at80–90 days post hatch (dph) or 1035 °C day and finishes 45 daysafter (at 1552 °C day), in Senegalese sole this begins at 10 dph (170 °Cday) and finishes approximately 10 days later (340 °C day), depend-ing on the rearing temperature (Fernández et al., 2009; Gavaia et al.,2002). Furthermore, even flatfish species within the same genera
35I. Fernández, E. Gisbert / Aquaculture 315 (2011) 34–48

Author's personal copy
present different developmental timing: e.g. summer flounder(Bengtson, 1999) starts metamorphosis at 35 dph (647 °C day),while Japanese flounder starts at 12–14 dph (around 210 °C day).Moreover, metamorphosis also seems to be larval size dependent(Geffen et al., 2007). Size of spawned eggs is different among flatfishspecies; bigger eggs produce bigger larvae at hatching, which undergolonger metamorphosis (e.g. Atlantic halibut, Sæle et al., 2004).
In addition to metamorphosis, larvae go through another criticaldevelopmental process to reach the juvenile stage, called settlement.During this process, larvae move from a pelagic to a benthicenvironment, which involves behavioural and ecological changesthat allow the fish to adapt to its final environment. Settlement canoccur during or after metamorphosis, depending on the flatfishspecies considered (Geffen et al., 2007). During development,settlement might happen even when no eye migration has takenplace, suggesting that it is not an adaptive response to eye migration,but rather the result of changing vestibular responses to thyroidhormones (Schreiber, 2006). During settlement, juveniles alsodevelop their normal pigmentation pattern, with no pigmentationon the blind side (Bolker and Hill, 2000). However, even whensuccessful metamorphosis (eyemigration) and settlement occurs, fishmight still show an impaired pigmentation pattern, such as albinismor hyperpigmentation (Haga et al., 2002b; Schreiber, 2006; Martinezet al., 2007). Several works have shown that several factors couldaffect flatfish pigmentation (Hamre et al., 2007), such as larvalnutrition (Estévez and Kanazawa, 1995; Hamre et al., 2005, 2007;Villalta et al., 2005).
One of the most important processes during flatfish larvaldevelopment is skeletogenesis. During this process, the skeletalstructures that support the body develop and transform through boneremodelling from a bilateral symmetry to a fully asymmetrical form(Sæle et al., 2006a,b; Schreiber, 2006), allowing correct eye migrationand leading to normal fish settlement and pigmentation. Similarly tometamorphosis, skeletogenesis also varies among flatfish species(Gavaia et al., 2002; Martinez and Bolker, 2003; Lewis-McCrea et al.,2004; Sæle et al., 2004; Lewis-McCrea and Lall, 2006). Some of thesedifferences are related to the number of structures forming the axialskeleton (e.g. 3–5hypurals, 50–45vertebral bodies) and/or the timingofskeleton formation and ossification. Compared to other marine species,such as European sea bass (Dicentrarchus labrax) and gilthead seabream
(Sparus aurata), with less than 30% and between 15 and 50% ofdeformed fishes (Villeneuve et al., 2005; Boglione et al., 2001,respectively), farmed flatfish shows a higher incidence of skeletaldeformities. In some flatfish species, deformities might reach between40 and 80% of the fry production (e.g. Senegalese sole; Gavaia et al.,2002; Fernández et al., 2009). Between flatfish and other marine fishspecies there exist differences in the sensitivity to nutritional imbal-ances, to xenobiotic factors or abiotic rearing conditions that couldexplain the high percentage of deformed flatfish (Fernández et al.,2009). Additionally, the impact on a fish's viability due to a particulartype of skeletal deformity might be different depending on the fishspecies (round or flatfish). Mechanical stress during swimming mightincrease the severity of some deformities in some marine fish species,leading to a lower survival rate, whereas lower swimming activity inbottom-settled flatfish does not make deformities more severe,rendering a higher survival rate (Schreiber, 2006).
Summarizing, the correct occurrence in time and developmentalstage of metamorphosis (eye migration) and skeletogenesis willimprove the efficiency of juvenile flatfish production, enhancing thequality and market value of the fish (Michie, 2001). Several workshave provided evidence that different factors could affect the normaldevelopment of fish larvae and particularly, skeletogenesis andmetamorphosis in flatfishes (Bolker and Hill, 2000; Solbakken andPittman, 2004; Harboe et al., 2009). Among these factors, nutrition atfirst feeding stage has been identified as a key factor influencinglarvae development. During the last decade, some studies have shownthat balanced nutrition at first feeding plays a critical role in thenormal occurrence of skeletogenesis and metamorphosis (Cahu et al.,2003; Sæle et al., 2003; Hamre et al., 2005; Lall and Lewis-McCrea,2007). As flatfish larvae present different developmental stage at firstfeeding and afterwards different skeletogenesis, fine-tuning of eachnutrient chemical form, dietary dose, and the larvae developmentalstage at which nutrients are offered becomes extremely necessary toreach the proper skeletal development for each flatfish species.
3. Vitamin A metabolism
Vitamin A (VA) is a morphogenetic nutrient that includes allcompounds that perform the same biological activity as retinol. It iswell known that retinoic acid, the main VA active metabolite, plays a
Fig. 1. Retinoid schematic pathways controlling cell proliferation and differentiation, which regulate the development of different vertebrate tissues and systems. Solid arrowsdenote direct regulation of retinoids, whereas dashed arrows indicate possible interactions between different tissues and systems with bone development.
36 I. Fernández, E. Gisbert / Aquaculture 315 (2011) 34–48

Author's personal copy
key role in morphogenesis and cell proliferation and differentiation,determining growth, body patterning, and the development of thenervous system, limbs and skeletal development during earlyvertebrate development (Ross et al., 2000). Most studies of VAmetabolism have been carried out in higher vertebrates, and only afew works have focused on VA metabolism in fish (Miki et al., 1984;Irie and Seki, 2002; Moren et al., 2002, 2004b; Lubzens et al., 2003;Palace and Werner, 2006; Alsop et al., 2008). In addition, furtherresearch must be done for improving the quantification methods ofdifferent retinoids in feeds and animal tissues. Detection limits for RAreported by Moren et al. (2005) might be considered as quite high;thus, considering that this compound is the most active VAmetaboliteand is found in animal tissues at very low levels, decreasing itsdetection limits would improve the knowledge of the effects of thisretinoid.
It is generally accepted that retinoid metabolism is fundamentallysimilar among all vertebrate classes (Duester, 2000) and is summarizedin Fig. 2. In this sense, two different dietary sources of VA compoundscould be considered, vegetable (carotenoids) and animal (retinyl esters)compounds. There are approximately 600 different carotenoids knownto exist, but only about 50 of these have provitamin A activity (Olson,1989). On one hand, β-carotene (the most potent retinol precursor)undergoes central cleavage to produce retinal, which in turns forms acomplex with cellular retinol binding protein II (CRBP-II). This complexprotects retinal from oxidation and allows its reduction into retinol bythe action of retinal reductase (Kakkad and Ong, 1988). On the other
hand, retinyl esters produce retinol byhydrolysis in the intestinal lumen(Napoli, 1996). Interestingly, a number of works have pointed outdifferences between fish and mammalians in their efficiency to convertcarotenoids into VA compounds (Rønnestad et al., 1998a; Ørnsrud et al.,2002; Tanumihardjo, 2002).Moren et al. (2002) reported that Atlantichalibut juveniles were able to convert retinyl acetate, β-carotene,astaxanthin and cantaxanthin included in dry diets into retinol.However, their conversion rates were different, with retinyl acetateand β-carotene reaching significantly higher whole fish levels thancantaxanthin and astaxanthin.Moreover, Takeuchi et al. (1995) showedthat larval Japanese flounder was not able to metabolize high dietarycontent of β-carotene.
Retinol binds to CRBP or CRBP-II forming a substrate for lecithin:retinol acyltransferase (LRAT), which expression is regulated by dietaryVA and exogenous RA and esterifies retinol to retinyl esters (Zolfaghariand Ross, 2002). Then, these retinyl esters (retinyl palmitate and retinylstearate) are incorporated into chylomicrons and secreted into thelymph. In the liver, the chylomicrons are incorporated via endocytosisinto the hepatocytes, where retinyl esters are stored until they arehydrolyzed and released as free retinol. Although the most commonform of VA accumulation in tissues is retinol, some freshwater fishes,such as Carassius auratus and Channa argus deposit retinol anddehydroretinol (Hata and Hata, 1973). Retinol binding protein (RBP)binds the free retinol and transfers it to stellate cells, where it is re-esterified into retinyl esters and stored in cytoplasmic lipid droplets.Mobilization of retinol through plasma from stellate cells is also done
Fig. 2. General vitamin A metabolic reactions, showing vitamin compounds, transport proteins and their cel/tissue localization. Abbreviations: ADH, alcohol dehydrogenase; ALDH,aldehide dehydrogenase; CRABP, cellular retinoic acid binding protein; CRBP, cellular retinol binding protein; RA, retinoic acid; SDR, short chain alcohol dehydrogenase/reductase;P450, Cytochrome P450.
37I. Fernández, E. Gisbert / Aquaculture 315 (2011) 34–48

Author's personal copy
byRBP,which in thefishplasmadoes not seem to forma super-complexwith transthyretin (TTR; Zanotti et al., 2008), one of the proteins thattransport thyroid hormones. In addition to RBP, retinol may be alsotransported by albumin (N'soukpoé-Kossi et al., 2007). Furthermore,while low-density lipoproteins, high-density lipoproteins, and veryhigh-density lipoproteins transport carotenoids (mainly astaxanthins),vitellogenin transports retinal to oocytes (Lubzens et al., 2003).
Once retinol is incorporated into the target cells, it is transformedto retinal and used as the chromophore of rhodopsin in the eye (Pepe,1999) or as a source of retinoids for the future embryogenesis in fisheggs (Irie and Seki, 2002). Additionally, retinol can be stored by re-esterification or undergoing metabolic activation, producing RA(Napoli, 2000). A complex metabolic pathway involving both RAsynthesis and RA catabolism tightly regulates RA intracellularphysiological level (Duester, 1996; Duester et al., 2003). During itsmetabolic activation, retinol dehydrogenase reduces retinol to retinal,which in turn is converted to RA by retinal dehydrogenase. Finally,free cytoplasmic RAmay either be degraded by Cyp26 enzymes into 4-OH-RA, 4-oxo-RA or 18-OH-RA; bound to the cellular retinoic acidbinding protein (CRABP-I), which protects it from degradation byCyp26 enzymes; or transported to the nucleolus to activate theligand-dependent transcription factors RAR and RXR (Lefebvre et al.,2005; Germain et al., 2006a,b). Interestingly, while all-trans-RAactivate specifically RAR isoforms (α, β, and γ), 9-cis-RA could bindand activate both kinds of retinoid receptors (RARs and RXRs);however, showing higher affinity for RXRs (Germain et al., 2006a,b).Surprisingly, a recent work demonstrated that the products of RAdegradation (4-oxo-RA, 4-OH-RA and 5,6-epoxy-RA) by Cyp enzymescould also regulate gene transcription and were able to fully rescuethe vitamin A-deficiency (Reijntjes et al., 2005). Transcription factors
RAR and RXR appear in every vertebrate tissue examined, includingthe kidney, liver, exocrine pancreas, thyroid, bone, cartilage, brain,eyes, gill, ovary, testis, blood, heart, and white muscle of fish (Alsopet al., 2001; Villeneuve et al., 2004) and regulate the expression of awide variety of genes (Bastien and Rochette-Egly, 2004; Arima et al.,2005). Gene expression regulation by RA takes place via dimerformation (Fig. 3) (Germain et al., 2006a,b), such as homodimers likeRXR/RXR or heterodimers like RAR/RXR, RXR/THR, RXR/VDR andothers, that bind to specific DNA sequences of the promoter region ofseveral genes, named retinoic acid response elements (RARE; Balmerand Blomhoff, 2002, 2005), activating or inhibiting their transcription(Germain et al., 2006a,b). Through this molecular pathway, RA plays akey role in morphogenesis and cell differentiation and proliferationprocesses (Ross et al., 2000), determining the development of bonestructures that compose the fish skeleton.
The retinoid content in Senegalese sole larvae presented twodifferent accumulation patterns for retinyl palmitate and retinol(Fernández et al., 2009). In agreement with Moren et al. (2004b),retinol content in larvae decreased, whereas retinyl palmitate contentincreased with time. Furthermore, it has been shown that Japaneseflounder larvae could excrete rapidly dietary atRA, decreasing itscontent around 50% within 18 h after feeding, as well as theisomerisation capacity of RA by flatfish larvae, as a detoxifyingmechanism, leading to the appearance of 13-cis-RA that cannot bindto RAR/RXR (Haga et al., 2004). Therefore, VA metabolism of flatfishhas its own particularities that should be considered in nutritionalapproaches to study the effects of VA on skeletogenesis.
In conclusion, VA metabolism is a highly complex process thatincludes different sources of VA, involves the participation of differentenzymes and transport proteins, and requires specific storage tissues.
Fig. 3. Retinoic acid molecular pathways that may be involved in the regulation of gene expression. While 9-cis-retinoic acid could ligand activate retinoic acid receptors (RAR) orretinoid X receptors (RXR), while all-trans-retinoic acid binds to RAR. RAR and XRX presented three subclases (α, β, and γ). That regulation can be done by homodimers (RAR-RAR orRXR-RXR) or heterodimers (RXR-VDR, RXR-THR, RXR-PPARγ, RXR-RAR) which binds to DNA sequences specifically and activate their gene transcription. HUFA, highly unsaturatedfatty acids RA, retinoic acid; TH, thyroid hormone; THR, thyroid hormone receptor; VD, vitamin D; VDR, vitamin D receptor.
38 I. Fernández, E. Gisbert / Aquaculture 315 (2011) 34–48

Author's personal copy
Complexity increases because dietary VA requirements vary with thefish developmental stage. Additionally, larval stages do not present afully developed digestive system; therefore, dietary VA requirementsand VA compound preferences of larval stages might be different tothose of juveniles (Moren et al., 2002). Further research is needed tostudy the metabolic pathways of retinoids in relation to theirhomeostasis, including their absorption, storage and mobilizationduring fish larval ontogeny.
4. Dietary VA effects on broodstock spawning performance andoffspring quality
Regarding the effects of dietary VA content in sexual maturation andreproduction performance, a high number of works have studied theeffects of dietary VA on mammalian metabolism, accumulation andtransport of retinoids to the gonads, gonad development, reproduction,andquantity andquality of the offspring (reviewed inClagett-DameandDeLuca, 2002). However, limited work has been performed on theeffects of dietaryVAonfish reproduction (Lavens et al., 1999; Palace andWerner, 2006; Alsop et al., 2008; Sawanboochun et al., 2008), and evenfewer have focused on flatfish reproduction and egg retinoid content(Furuita et al., 2001, 2003). Different forms of VA (retinol, retinyl esters,and carotenoids) have been detected in eggs from several freshwater(Costaridis et al., 1996; Lubzens et al., 2003) andmarinefish species (Irieand Seki, 2002). In particular, flatfish eggs contain primarily retinal andsmall amounts of retinol (Rønnestad et al., 1998a; Irie and Seki, 2002).
Furuita et al. (2001) evaluated the effects of different levels ofdietary VA (retinyl palmitate) on Japanese flounder spawningperformance and egg quality. Authors showed that the broodstockfed a control diet (CD) with 110,000 IU VA kg−1 had a slightly longerspawning period and higher number of total eggs produced than thatfed a supplemented VA diet (SD; 3,370,000 IU VA kg−1). However, thepercentage of buoyant eggs and the hatching rate of the SD groupwere significantly higher than in the CD group. Fecundity values(number of eggs produced per kg of female per day) of the SDbroodstock were significantly higher than those of the CD group. Asexpected, the VA content of the liver and ovaries in females reflectedthe dietary VA content; being retinol and retinal the two forms ofretinoids found in ovaries. Surprisingly, the retinol content in SD eggswas only 4 times the content in CD eggs and twice the total VA eggcontent, suggesting the control of retinoid transport from liver toovaries. Furthermore, while retinol was the main retinoid found in SDovaries, retinal was the primary form in CD ovaries. Moreover, thetotal VA content in eggs from wild fish (13–28 IU g−1) wasintermediate between the CD and the SD group. Interestingly, theauthors did not find significant differences on the percentage ofabnormal larvae and larval starvation tolerance between both groups.The lack of significant differences in the percentage of deformedlarvae between SD and CD broodstocks could be due to the fact thatthe authors did not rear hatched larvae long enough to allow thedevelopment of skeletal deformities.
Furuita et al. (2003) also evaluated the reproductive performanceof Japanese flounder broodstock fed a non-VA-supplemented diet(NSD) and a VA-supplemented (SD) diet, containing 1155 and25,350 IU VA kg−1 (0.77 and 16.9 mg retinyl acetate kg−1), respec-tively. The percentage of buoyant eggs and normal larvae was higherin the SD fish group; however, the hatching egg rate and larvalsurvival were the same in both groups. Surprisingly and contrary tothe previous study of the same authors (Furuita et al., 2001), the VAcontent in SD ovaries was lower than in NSD ovaries. These differentresults could be mainly because the SD group continued to spawn incontrast to the NSD group that ceased feeding and spawning, resultingin a change in VA body content between both groups.
Results from both studies (Furuita et al., 2001, 2003) suggest thathigh dietary VA levels (3,370,000 IU kg−1) increase egg VA contentand spawning performance, but do not affect egg and larval quality in
Japanese flounder, as has similarly been found in studies with rats(Biesalski et al., 1996). RA concentration in eggs might not be highenough to exceed the binding capacity of CRABP, which wouldprevent the ectopic activation of RARs, resulting in an abnormalpattern of gene expression; or embryos could not present the propermetabolic enzymes to convert the high retinol content of their yolksac into other retinoids. In this sense, parallel studies revealed thatJapanese flounder embryos treated with disufiram (a chemical thatinhibits the conversion of retinal to RA) did not present skeletaldeformities (Suzuki et al., 2000), which seems to indicate that duringthe embryonic development up to first feeding, animals could notconvert retinol into retinal and then retinal into RA. In addition, theaforementioned studies confirmed that: (i) in agreement withprevious works, broodstock diet directly affects fish reproductionand egg quality; (ii) fish have a considerable capacity to regulate VAmetabolism and transport from liver to eggs; and (iii) flatfish eggsaccumulate retinol and not just retinal, as reported by Irie and Seki(2002). Furthermore, we propose retinyl palmitate or acetate as theVA form to be supplied in broodstock diets as fishes at thatdevelopmental phase present a fully-developed digestive systemable to convert those retinoids to the needed active retinal and RAforms, but we cannot recommend the optimum level of dietary VA forflatfish reproduction. At most, the optimal dietary VA requirements toimprove reproduction performance of flatfish broodstock shouldexceed 1155 IU kg−1 (Furuita et al., 2003); since this level decreasedfeed intake and stopped spawning in Japanese flounder. However,none of the diets with high VA content showed any benefits on larvalquality at early stages, although a VA content of 25,350 IU kg−1
(Furuita et al., 2003) improved the percentage of normal larvae ascompared to a deficient VA diet (1155 IU kg−1). Further work remainsto be done to determine the optimum VA dietary content for flatfishbroodstock; a requirement that may be specific for each flatfishspecies.
5. Dietary VA effects on fish larval development
In order to study the role of VA and its active metabolite (RA) aswell as the effects of a VA-imbalanced diet on larval flatfishskeletogenesis, several authors have used two different experimentalapproaches: (i) bath exposure of larvae in different concentrationsand isomers of RA, and (ii) feeding of larvae with diets containingdifferent VA levels.
5.1. Larval bath exposure essays
As skeletal deformities are often detected under culture conditionsin fingerlings, knowing the mechanisms that induce them at early lifestages (embryo and larvae) will aid to develop strategies forpreventing their appearance. Furthermore, since cartilage compo-nents in pharyngeal arches, neurocranium, and pectoral fin developfrom embryonic to early larval stage (Bisbal and Bengtson, 1995),maternal nutrients stored in eggs could affect the development ofthose skeletal components. However, modifying the VA content in theegg is not easy as the broodstock are able to control its transport fromthe liver to the ovaries. Furthermore, high retinol content in eggs didnot induce abnormal skeletal development in larvae (Furuita et al.,2001). To further this line of investigation, different experimentalapproaches have been developed to determine the effects of an excessor deficiency of VA compounds in fish eggs. Although feedingstrategies provide quite realistic knowledge about the effects ofdietary VA imbalances on flatfish skeletogenesis (Dedi et al., 1995;Takeuchi et al., 1998; Furuita et al., 2001, 2003; Haga et al., 2002b;Sæle et al., 2003; Tarui et al., 2006; Fernández et al., 2009), they onlygive us information about the effects after first feeding. However,organogenesis and morphogenesis are processes that begin beforefish mouth opening. Therefore, studies with larvae exposed to
39I. Fernández, E. Gisbert / Aquaculture 315 (2011) 34–48

Author's personal copy
different concentrations of RA isomers (all-trans-RA and 9-cis-RA), aswell as to different agonists of retinoic acid and retinoid X receptors,were used to highlight the importance of RA in the development oflarvae before mouth opening. This type of experimental approachshowed how RA controls the development of several skeletaldeformities (pharyngeal arches, pectoral fin, jaw and vertebralbodies) and the underlying molecular pathways involved in theappearance of skeletal deformities at early stages of development(Suzuki et al., 1998, 1999, 2000).
During early development, it has been shown that different genes,such as Hoxd-4 and sonic hedgehog (shh), are associated with thedevelopment of pharyngeal and pectoral fin skeleton in Japaneseflounder (Suzuki et al., 1998, 1999, 2000). Suzuki et al. (1998)demonstrated that Hoxd-4 was expressed between 21-somite (36 hafter fertilization; haf) and prim-5 stages (60 haf) in the central nervoussystem, from rhombomere 7 to the spinal cord, and in the posteriorpharyngeal region, from gill arch 2; in addition, Hoxd-4 was alsoexpressed in the pectoral fin buds. Furthermore, expression of Hoxd-4appeared to be restricted to cartilage precursor cells of gill arches 2–5.These facts suggest that during the formation of the pharyngeal arches,Hoxd-4 functions in the specification of regional identity between gillarches 1 and 2. Interestingly, when embryos in the early shield stagewere exposed to RA concentrations above 10−7 M during one hour, theanterior border of Hoxd-4 expression shifted anteriorly in a dose-dependent manner, affecting the identity of gill arches (Suzuki et al.,1998). In addition, when the same stage embryos were exposed to10−5 M RA, brain segmentation and eye formation were disturbed.Given that Hoxd-4 presented its own RA-responsive upstream element(RARE), the former authors suggested that Japanese flounder exposureto RA affects gill early development directly through the regulation ofgene transcription mediated by RAR/RXR heterodimers.
In addition, Suzuki et al. (1999) reported that Japanese flounderembryos at prim-5 stage showed shh gene expression in themandibular, hyoid and gill primordials. After 40 h, shh domainsexpanded at the posterior edge of the endoderm of each mandibular,hyoid, and gill primordium, reaching maximal gene expression inpectoral fin buds. When prim-5 stage embryos were exposed todifferent RA concentrations (0.5 and 2.5×10−7 M) during 40 h, bothshh and Hoxd-4 expression were depressed, as well as the expansionof the shh expression domainswas reduced. These reductions (in geneexpression and expression domains) were correlated with thedevelopment of malformations in the pharyngeal cartilages as theirgrowth direction was shifted posteriorly. Furthermore, the size ofpharyngeal arches diminished, and the malformations were moresevere when embryos were exposed to 2.5×10−7 M RA, showing thatthe trabecular cartilage typical of those skeletal structures was absentin the treated embryos. As a consequence of the induced malforma-tions in the brain, survival rate of embryos decreased (from 80% to 2%)when RA concentration increased.
In two additional studies, Haga et al. (2002a, 2003) suggested thatRA induces skeletal deformities through the RAR/RXR pathway duringlater larval stages. Firstly, Haga et al. (2002a) showed that Japaneseflounder larvae exposed to different RA isomers (25 nM of atRA, 9-cis-RA and 13-cis-RA) from 6 to 9 dph, presented several types of skeletaldeformities in the jaws, vertebral bodies, and caudal fin complex.These authors found differences in the incidence and severity ofskeletal disorders depending on the RA isomer administered, andconcluded that atRA activated the expression of RAR, inhibitingvitamin D receptor (VDR) expression; whereas 9-cis-RA activated RARand RXR, and 13-cis-RA did not affect either RAR or VDR geneexpression. In addition, pitx2 gene expression was evaluated, showingthat its expression was inhibited in the lower jaw by atRA and 9cRA,increasing lower jaw deformity; however, no different frequency ofright and left body asymmetry was found in metamorphosed RA-treated flounder, as only loss of pitx2 re-expression could inducerandomization of eye-sidedness (Suzuki et al., 2009). In their second
study, Haga et al. (2003) exposed Japanese flounder larvae between6 and 9 dph to two synthetic retinoid compounds: 25 nM Am80(a retinoic acid receptor α/β-selective retinoid) and 25 nM metho-prene acid (a retinoid X receptor-selective retinoid; MA). The Am80treatment increased the RAR gene expression in the lower jaw, whilethe AM treatment did not activate it. Whereas, Am80 and AM did notchange the RXR gene expression, both treatments specificallyinhibited the expression of the patched gene in the lower jaw. Patchedgene is a receptor of the hedgehog family expressed in chondrocytesand osteoblasts (Nakamura et al., 1997). These results showed thatthe RAR signal pathway is the major factor involved in the appearanceof skeletal deformities related to dietary VA imbalance in Japaneseflounder. These findings are in agreement with those results reportedin RAR and RXR mice mutants. While RAR mice mutants died as aconsequence of the severe skeletal deformities induced, RXR mutantsdid not show these severe skeletal deformities (Weston et al., 2002).
In addition to the regulation of gene expression induced by RA,several studies have highlighted the development of VA-inducedskeletal deformities in flatfishes underlying thosemolecular pathways(Suzuki et al., 2000; Haga et al., 2002a, 2003; Suzuki et al., 2003;Martinez et al., 2007). In brief, these studies found that RA inducedskeletal deformities in the pharyngeal arches and pectoral finsdepending on the RA concentration and the larval stage ofdevelopmental at which exogenous RA was administered. Gastrulat-ing embryos at shield stage treated with RA showed severemalformations in their pharyngeal skeletons (e.g. absence of Merckel'scartilage and fusion of cartilage components in the mandibular andhyoid arches), while treatment with RA after hatching, induced adisorganized growth in direction of the pharyngeal cartilages (Suzukiet al., 2000). In contrast, when exposing larvae from 6 to 9 dph todifferent RA isomers deformities were induced in the lower jaw,vertebral bodies and caudal fin complex (Haga et al., 2002a). Growthretardation of the dentary bone was evident in the lower jaw, whilemajor vertebral abnormalities were the hypertrophy and fusion ofcentrums, and the increase of the mean number of vertebraecomposing the axial skeleton. In addition, partial or total absencesof bone structures in the caudal fin complex were reported. Thefollowing results from the aforementioned study showed that atRAexerted a larger biological activity in contrast to the other tested RAisomers due to the incidence and severity of skeletal deformitiesinduced. Those anomalies might be summarized as (i) higherincidence of lower jaw deformities in larvae treated with atRA;(ii) higher percentage of larvae lacking caudal fin complex whenexposed to atRA than those that were exposed to 9 and 13-cis-RA;(iii) a significantly higher percentage of atRA-exposed larvaepresented supranumerary vertebrae; and (iv) larvae exposed toatRA presented the shortest length, followed by 9-cis-RA and 13-cis-RA treated larvae. According to the authors, growth differences amongRA isomer treatments might be a consequence of, on the one hand,skeletal deformities induced in the feeding apparatus, whichinterfered with feeding and, on the other hand, the absence of thecaudal fin, which obliged larvae to make a greater effort duringswimming activity. Furthermore, when Japanese flounder larvae wereexposed from 6 to 9 dph to RAR/RXR selective agonists, RAR selectiveagonist (Am80) treatment caused upper and lower jaw deformities in100% of larvae (depressed growth of maxilla, premaxilla and dentary),while RXR selective agonist (AM) induced skeletal deformities only inlower jaw, although the differences with the control group were notsignificant (Haga et al., 2003). Moreover, exogenous RA (25 nM)between 13 and 16 days post fertilization specifically inhibits themesenchymal condensation prior to the proximal radial formationtogether with the down-regulation of shh and patched expression,resulting in the loss of proximal radials in Japanese flounder (Suzukiet al., 2003). Thus, it was indicated that differentiation of theprecursor cells of radials and lepidotrichia begins in the proximalpart of the fin fold and that the initial mesenchymal condensation
40 I. Fernández, E. Gisbert / Aquaculture 315 (2011) 34–48

Author's personal copy
prior to the proximal radial formation is highly susceptible to theeffects of RA. Lepidotrichia formation does not occur where proximalradials are absent, indicating that lepidotrichia differentiationrequires interaction with the radial cartilages.
Similarly, Martinez et al. (2007) studied the effects of exposure toRA (0–20 nM) and 9-cis-RA (10 nM) on summer flounder pigmenta-tion and skeletogenesis from stages C to E. Although these authorsexposed summer flounder larvae to RA for a longer period (10 days)than the previous studies, they reported similar results to thoseobtained on Japanese flounder larvae. Martinez et al. (2007) not onlyidentified the different skeletal deformities induced by RA treatment,but they also provided a classification of these deformities from slightto severe based on the reduction of larvae viability induced by them.In contrast to previous works (Haga et al., 2002a), deformities did notaffect survival rate (85% mean value) or the percentage of fish thatcompleted metamorphosis (85% mean value). These differencesbetween both studies might be explained by differential flatfish-species sensitivity to RA exposure and/or the different larvaldevelopmental stages used in both works. Similar to Haga et al.(2002a), Martinez et al. (2007) found differences in the frequency ofhyperpigmented and deformed fish between the control and RA-treated groups. When premetamorphic summer flounder larvae wereexposed to RA, the incidence of hyperpigmented fish was higher thanin control fish, although no correlation was found between thefrequency of hyperpigmented fish and the concentration of RA.Regarding the skeletal deformities induced by RA treatment, bothMartinez et al. (2007) and Haga et al. (2002a) found that the rate of finand vertebral malformations increased with the RA concentration;however, no dose–response effect was found in the incidence of jawdeformities (Martinez et al., 2007). In addition, the incidence andseverity of skeletal defects in jaws and pectoral fins were inverselycorrelated with larval size, confirming previous results reported byHaga et al. (2003). In addition, atRA was associated with asignificantly higher frequency of pectoral and vertebral defects, ascompared to 9-cis-RA.
Dissolved RA in water also exerts teratogenic effects on the centralnervous system of flounder embryos, as reported in zebrafish (Zhanget al., 1996) and Xenopus sp. (Durston et al., 1989). However, theseeffects depend on the concentration and isomer of RA, the species offlatfish considered, and the developmental stage used for experimen-tation. These results indicate that RA induces skeletal deformitiesthrough the regulation of the expression of different target genes bythe RAR/RXR pathway, and this gene expression regulation couldcontrol cell proliferation and differentiation, although RA treatmentwas not able to modify eye-sidedness. Therefore, skeletal deformitiesmight be a consequence of two facts: (i) RA regulation of severalgenes that code for extracellular matrix proteins and then, theirprotein synthesis that could alter the properties and characteristics ofthe resulting bone; and/or (ii) the RA regulation of those genes thatregulate cell proliferation and differentiation processes, increasing ordecreasing the number of chondrocytes and/or osteoblasts thatcompose such skeletal structures. Consequently, it is necessary tostudy how RA affects different genes involved in the development ofthe bone tissue, as well as the downstream relationship betweenthose genes.
5.2. Dietary dose–response essays in larvae
Undoubtedly, RA bath exposure studies have greatly contributedto important insights into the molecular pathways leading to skeletaldeformities. However, flatfish aquaculture production requires prac-tical information, and in this regard, several studies have contributedto determine the optimum level of VA compounds for feeding larvae.Some works have evaluated the effects of dietary VA content inmarine fish skeletogenesis using compound feeds (Villeneuve et al.,2004, 2005, 2006; Mazurais et al., 2009). However, flatfish larvae, as
with most marine fish cultured species, only accept live prey from theonset of exogenous feeding. Consequently, studies on nutritionalrequirements and the effects of nutrients on early fish developmentwere conducted using enriched live preys, such as rotifers andArtemia. In general, the nutrient content of live prey reflects that of theenrichment emulsion; but, as Monroig et al. (2006) indicated, manyfactors may affect the enrichment efficiency of the emulsions (i.e., liveprey density, dissolved oxygen, aeration intensity and/or hydrody-namics in the enrichment containers), and consequently, the nutrientcontent in the live prey. Another source of variability might comefrom differences in the strains of Artemia used (Takeuchi et al., 1995).Furthermore, nutritional studies become even more complicated inflatfish larvae because of their species-specific developmental timing,and therefore, their different feeding strategies. For example, summerflounder larval feeding sequences imply supplying rotifers from 3 to20 dph (Bengtson, 1999), while Senegalese sole larvae are only fedrotifers from 3 to 10 dph, or even could be fed without rotifers(Villalta and Estévez, 2005).
5.2.1. Studies with VA-enriched live preysRotifers, as well as Artemia, are able to metabolize different VA
compounds modifying their content (Takeuchi et al., 1998; Haga et al.,2004, 2006; Giménez et al., 2007; Tarui et al., 2006; Fernández et al.,2009), but they have a limited capacity to accumulate them (Giménezet al., 2007). In this sense, Haga et al. (2004) reported quickaccumulation of atRA in Artemia nauplii enriched with 10 mg l−1
atRA in the enriching medium, with a peak of atRA content afterenrichment. These findings suggest that Artemia nauplii may excreteand re-uptake RA metabolites. However, when both live preys wereenriched with the same quantity of VA compound, rotifers accumu-lated a higher quantity of retinoids than Artemia (Haga et al., 2006;Giménez et al., 2007). Furthermore, total retinoid levels in VA-enriched rotifers and Artemia are significantly correlated with theconcentration of total VA contained in the enriching emulsions.However, they did not accumulate in a dose-dependent manner asreported by Giménez et al. (2007). From these results, it can bededuced that both live preys have limited capacity to hydrolyzeretinyl acetate and accumulate VA as retinyl palmitate (the mainretinoid form of VA storage in animals). Rotifers accumulated higherlevels of retinyl palmitate (31 times), retinol (7 times) and retinylacetate than Artemia, even when rotifers were enriched with a loweramount of retinyl acetate in the enriching emulsion and the durationof enrichment was shorter (Giménez et al., 2007). Although theformer authors found trace levels of RA in both types of live preys,they did not find retinal in them. This was in opposition to resultsreported byHaga et al. (2006), even though VA levelswere higher in theenriching emulsion used by Giménez et al. (2007). Two hypothesescould account for such differences between these experiments: (i), VAenrichment of live prey was done under different light conditions in thestudy conducted by Giménez et al. (2007), leading to the degradation ofretinal formed in rotifers and Artemia, and/or (ii) both live preys mightnot be able to metabolize retinyl acetate at a high enough rate toincrease the metabolic pathway from retinyl palmitate to retinal(Takeuchi et al., 1998).
Artemia enriched with different VA compounds (retinol, retinylpalmitate, retinyl acetate, and RA) in the same quantity (10,000 IU l−1)accumulated and metabolized them at different rates (Takeuchi et al.,1998). Total VA content in Artemia was higher when it was enrichedwith retinyl acetate thanwhen it was enrichedwith the other retinoids.RA only represented 19% of the total VA content in Artemia enrichedwith retinyl acetate, whereas RA representedmore than 50% in Artemiaenrichedwith theother retinoids, showing thatArtemiahada lowabilityto metabolize the synthetic VA form (retinyl acetate). In contrast to theresults obtained by Giménez et al. (2007) and Fernández et al. (2009),the RA content in VA-enriched Artemia found by Takeuchi et al. (1998)wasmuch higher than in the aforementioned studies. These differences
41I. Fernández, E. Gisbert / Aquaculture 315 (2011) 34–48

Author's personal copy
might be due to differences in the amount of administered VA, live preyenrichment procedures and/or different Artemia strains used by theauthors in their respective studies. In this sense, it should be noted thatVA and its derivatives are unstable during light irradiation (Murayamaet al., 1997), and therefore this might affect the VA content in biologicalsamples. Haga et al. (2006) found that the retinoid content in rotifersenrichedwith retinyl palmitatewashigherwhen enrichment took placein darkness (b1 lx) rather than under bright conditions (2000 lx).Retinol andRA contents increasedover a 24 hperiod in rotifers enrichedin darkness, and the retinyl palmitate content showed two peaks at 3and 18 h. However, in rotifers enriched under light, retinol and RAcontents decreased 12 h onwards, and the retinyl palmitate contentshowed only one peak at 3 h. Conversely, retinyl palmitate and retinolcontents in Artemia were always higher when nauplii were enrichedunder light conditions, although the dynamics of retinoid content wassimilar under either light or darkness conditions. Rotifers enrichedwitha commercial emulsion in darkness had higher retinal content,suggesting that retinal should be protected from light degradation incommercial emulsions.
The different pattern of retinoid accumulation between both livepreys might be due to species-specific physiological differences intheir developmental stages and filtration rates between adult rotifersand Artemia metanauplii, and/or differences in their respectiveenrichment protocols, in particular, the duration of enrichment(Giménez et al., 2007). All these results should be carefully consideredwhen designing VA nutritional studies with live prey at first feeding,especially because of the difficulties to maintain constant total VAlevels during the transition from the rotifer to the Artemia feedingphase. Paying attention to the RA content in live feeds should be apriority, because RA is up to 40 times more active than retinyl acetate(Ross et al., 2000), and live preys show peaks of RA content at 6 and18 h post enrichment (Haga et al., 2006). From these results, it mightnot be recommended to feed fish larvae with RA-enriched Artemia,mainly after 6 and 18 h post enrichment in order to avoid theunsuitable toxic effects of VA.
5.2.2. Developmental effects on flatfish larvae fed VA-enriched live preysDifferent studies have shown the effects of different levels of VA on
flatfish larval skeletogenesis and the incidence of skeletal disorders.Dedi et al. (1995) fed Japanese flounder larvae from 10 to 40 dph withincreasing levels of retinyl palmitate in enriched Tien-tsin Artemia andfound that for normal skeletogenesis, larvae must be fed with lessthan 50,000 IU kg−1, which was 20 times below the amountsuggested to be included in the diet in order to reduce pigmentationdisorders (Takeuchi et al., 1995). Furthermore, fish larvae fed Artemiaenriched with retinyl palmitate (N4 mg l−1 of enrichment) showedreduced body weight, total length, and higher incidence of skeletaldeformities (mainly in the vertebral column). Authors suggested thatvertebral deformities (fusion and compression) resulted fromaccelerated formation and growth of vertebral bodies, which mainlyoccurred during chondrocyte proliferation and differentiation. How-ever, it has been shown that Japanese flounder, as other teleost fish,undergoes intermembranous ossification in vertebral centra (Hall,2005); thus, compressed and fused vertebrae might be a consequenceof disruption in notochord segmentation as it has been recentlydemonstrated in zebrafish by Haga et al. (2009). Contrarily toprevious results found in flounder embryos exposed to high RAconcentrations by bath exposure (Suzuki et al., 1999), larvae fromTakeuchi et al. (1995) did not present differences in survival rateamong the different dietary regimes tested. Those discrepancies maybe due to the high dietary VA content at 10 dph, which might notinduce severe skeletal deformities that could reduce the viability oflarvae, whereas in embryos that undergo important skeletal structuredevelopment, induced skeletal deformities could critically reducelarvae viability, such as those deformities in the pharyngeal arches(Suzuki et al., 2000) or in the operculum complex. In more developed
larger stages these have been reported as severe deformities thataffect larvae viability (Koumoundouros et al., 1997).
Takeuchi et al. (1998) showed that different VA compounds couldinduce skeletal deformities when they were included in the diet athigh doses (100,000 IU kg−1), although their morphogenetic effectvaried according to the retinoid compound considered, confirmingbath exposure studies (Haga et al., 2002a; Martinez et al. (2007).Larvae fed with retinyl acetate enriched Artemia from 20 to 26 dphgrew better than those fed with RA enriched Artemia, retinylpalmitate, retinal or retinol. This result suggests that the dietarycontent of each retinoid is more important than the overall amount ofVA included in the diet. According to the results presented by Dediet al. (1995), larvae fed Artemia enriched with high levels of VA anddifferent retinoid compounds did not show differences in theirsurvival rate, whichmight be due to the fact that VAwas administeredduring a short and non-critical developmental postmetamorphiclarval stage (20–26 dph).
In addition to those studies examining the effects on flatfishskeletogenesis of the different VA compounds included in live feeds,two other works have shown the different effects of diets enrichedwith VA compounds when those are offered to larvae at differentdevelopmental stages (Haga et al., 2002b; Tarui et al., 2006). Hagaet al. (2002b) demonstrated that feeding larvae with RA-enriched liveprey (12,690 and 1589 μg g−1 DW in rotifer and Artemia, respectively)at different developmental stages (A–B, C–D, E–F, G–H and I; forstaging see Minami, 1982) resulted in different typologies of skeletaldeformities and pigmentation rates. Thus, larvae fed during A–B(premetamorphic) stages showed higher pigmentation abnormalities(albinism) and higher number of vertebrae. Furthermore, no evidentcorrelation was found between the timing of atRA administration andfish survival rate. However, larvae fed both RA-enriched rotifers andRA-enriched Artemia during the G–H (postmetamorphic) stagespresented the lowest survival. These results could be explained bythe incidence, severity, and typology of skeletal deformities inducedin each larval group. In particular, the process of skeletogenesis, whichtakes place during the rotifer-feeding phase, is more sensitive todietary VA imbalances than those taking place at later stages ofdevelopment, resulting in a high incidence and severity of skeletaldeformities (lower jaw deformity, caudal fin complex loss, etc.). Theformer deformities have been reported to be a cause of early larvalmortality in fish (Suzuki et al., 2000; Koumoundouros et al., 2002). Incontrast, lower survival rates at later stages of development (G–Hstages), might be a result of nutritional VA imbalances of a longerduration, which would increase the morphogenetic effects of VA onskeletogenesis (high deformity frequencies). In addition, the skeletaldeformities induced during A–B stages caused these larvae to beshorter as compared to larvae from the other groups. This fact wouldalso explain why these larvae had to be fed with Artemia after 40 dphsince their jaw deformities hampered their ability to capture and bitepellets, while their counterparts were weaned into dry diets.
On the other hand, Tarui et al. (2006) reported that Japaneseflounder larvae fed Artemia, enriched with over 5 mg l−1 retinylpalmitate during F–G stage (27–31 dph) presented increased hyper-melanosis on the blind side, shorter total length and poor growth.However, the incidence of skeletal deformities (vertebral fusion) onlyincreased when larvae were fed Artemia enriched with 10 mg l−1
retinyl palmitate. These pigmentation and skeletal disorders wereprobably induced by the RA rather than the retinyl ester included inthe emulsion. In addition, no significant differences in survival werefound among feeding regimes, which suggests that the dietary VAcontent tested at F–G stages in Japanese flounder larvae did not eitherproduce toxic effects or induce severe skeletal deformities that couldaffect survival.
InAtlantic halibut, researchhas focussed on comparing the process ofskeletogenesis andmetamorphosis (eyemigration andpigmentation) inlarvae fed with two different live preys, copepods and Artemia (Sæle
42 I. Fernández, E. Gisbert / Aquaculture 315 (2011) 34–48

Author's personal copy
et al., 2003). The former authors showed that larvae fed copepodspresented accelerated eye migration, and a higher percentage of larvaewith completed eye migration as compared to those fed Artemiaenriched with DHA SelcoTM. Furthermore, fish fed natural zooplanktonpresented more mineralized bone than those fed Artemia. However, nodifferences in growth and survival rate were found between larvae fedthose two live preys. These authors suggested that differences inmetamorphosis and skeletogenesis success could be related to thedifferent VA contents in both live preys. In a related study, Moren et al.(2005) quantified the VA content in both live feeds and foundthat Artemia contained 466 IU kg−1, while copepods had only66 IU kg−1 (7 times less). In addition, calanoid copepods contain highlevels of carotenoids (specifically lutein and astaxanthin) and only traceamounts of retinol and retinal, while DHA Selco enriched Artemiapresented cantaxanthin as the major carotenoid (Rønnestad et al.,1998a,b). Atlantic halibut larvae fed natural zooplankton showed ahigher VA content than those fed enriched Artemia (Moren et al., 2001),which could suggest that pro-VA compounds from copepods might bemore efficiently converted to VA than those found in Artemia enrichedwith commercial emulsions. However, it should be noted that both livepreys didnot only presentdifferences in retinoid levels and composition,but they also differ in the levels of many other nutrients, such as iodine,which could influence flatfish metamorphosis and skeletogenesis(Hamre et al., 2002; Van der Meeren et al., 2008). As Sæle et al. (2003)pointed out, differences in the developmental performance of Atlantichalibut larvae fed both live preys might be a consequence of a balancedcontent in VA, vitamin D, and normal thyroid hormone metabolism.These results suggest thatwheneverpossible liveprey, emulsions shouldbe enriched to a nutrient compositionmore similar to that found inwildcaught copepods.
Regarding Senegalese sole, the effects of different levels of VA onlarval skeletogenesis have been recently investigated. Fernández et al.(2009) evaluated the effects of dietary VA imbalance on larvalperformance and skeletogenesis during the Artemia feeding phase. Inthat study, Artemia metanauplii (EG, INVE) were enriched withgraded levels of retinyl palmitate added to a commercial emulsion(Easy Selco™). As previously reported in Japanese flounder (Hagaet al., 2002b), the effects of dietary VA imbalance in Senegalese solelarval performance depended on the developmental stage at whichVAwas supplied. High levels of VA (203,000 IU kg−1) offered to larvaeduring the Artemia feeding phase (6–37 dph) induced a low growthperformance in length, but no significant differences were found inweight and survival rate among larvae fed different VA doses(Fernández et al., 2009). As suggested by Suzuki et al. (2000), dietaryVA content in live prey provided from 6 dphmight not be high enoughto result in toxicity or induce severe skeletal deformities that couldaffect larvae viability or growth performance. This hypothesis issupported by the fact that neither jaw deformities nor retardation ofthe development of digestive system was found in any dietary VAregimen tested, in contrast to other results reported in other marinefish species (Villeneuve et al., 2005; Fernández et al., 2008).
Other recent relevant results dealing with Senegalese sole werethat dietary VA levels affected the development of the thyroid gland(Fernández et al., 2009). Thus, larvae fed higher dietary VA contentpresented lower numbers of developed follicles; however, increasingthe VA dose induced an increase in their size and in the immunore-active staining of T3 and T4 hormones. The effects of dietary VA levelon thyroid development could be a consequence of retinoid andthyroid hormone interaction at the molecular, protein or cellularlevels. In the first case, RA acts as a ligand activator of RAR and RXR,and then the latter could form heterodimers with THR thatconsequently could activate or inhibit the transcription of specifictarget genes (Germain et al., 2006a,b). Secondly, transthyretin as wellas albumin and thyroxine-binding globulin, is known for transportingthyroid hormones. The transport of retinol in plasma is carried out byretinol-binding protein that needs to form a complex with transthy-
retin in mammals (Ingenbleek and Young, 1994), although thisprocess has not been reported in fishes (Zanotti et al., 2008).However, interaction between retinoids and thyroid hormones atthe protein level could be through albumin, which also transportsretinol (N'soukpoé-Kossi et al., 2007). Moreover, it has been shownthat RA down-regulates albumin gene expression (Masaki et al.,2006), which further supports the idea that VA homeostasis couldaffect thyroid hormone transport. Finally, interaction could be at acellular level, as RA regulates cell proliferation and differentiationprocesses (Ross et al., 2000), RA levels might affect the developmentof thyroid follicles. In addition, these results concerning thyroidhormones could explain the higher frequency of fish with acceleratedeye migration found in larvae fed VA supplemented diets. However,no differences in eye migration were found at the end of the trial,indicating that the VA doses tested did not arrest eye migration. Sincethyroid hormones also regulate skeletal development and bonehomeostasis (Wexler and Sharretts, 2007), they could have exertedsome additional influence in the skeletal deformities induced by VA(Fernández et al., 2009).
Regarding the specific effects on Senegalese sole skeletogenesis,we found that feeding larvae with high VA doses during the Artemiafeeding phase led to an increase of deformed fish and the percentageof fish showing more than one deformity, which was correlated to anincrease in the dietary VA content (Fernández et al., 2009). However,no jaw deformities were observed in any larval group, suggesting thatthe dietary VA imbalance was applied at a non-critical developmentalstage for jaw skeletogenesis (most skeletal elements of the jaws wereossifying before feeding larvae with Artemia), and/or the tested VAdietary contents in Artemia were not high enough to affect jawdevelopment. However, larvae fed the highest VA content in Artemiashowed cranial deformities related to the opercular complex that wassignificantly correlated to deformed prehaemal vertebrae (Fernándezet al., 2009). In addition, it is noteworthy that only extremely highdietary VA contents (enriched rotifers with more than 10 times VAcontent in the commercial emulsion) given at premetamorphosis
Fig. 4. Dietary treatments resultedwhen rotifers andArtemiawere enrichedwith differentexperimental emulsions and at different developmental stages to Senegalese sole larvae(pre-, pro-, and post-metamorphosis). Different enriching emulsion resulted from addingretinylpalmitate to control emulsion, EasySelcoTM(ES; 15,000,000 IU kg−1), to increase itsvitaminA content up to10(ES+VA×10) and50(ES+VA×50) times. Control larvaewerefed with rotifers and Artemia enriched with commercial emulsion (Easy Selco; ES), whilethe other dietary treatments resulted from feeding larvae with: enriched rotifers withES+VA×10 or ES+VA×50 (T1 and T2, respectively) and ES enriched Artemia; enrichedrotifers with ES and enriched Artemiawith ES+VA×10 or ES+VA×50 from10 to 20 dphand enriched Artemiawith ES afterwards (T3 and T4, respectively); and enriched rotiferswith ES and enriched Artemiawith ES from 10 to 20 dph and afterwards enriched ArtemiawithES+VA×10orES+VA×50 from21 to 37 dph (T5andT6, respectively). Allfishwerefed with compound feeds from 33 dph to the end of the trial (55 dph).
43I. Fernández, E. Gisbert / Aquaculture 315 (2011) 34–48
Author's personal copy
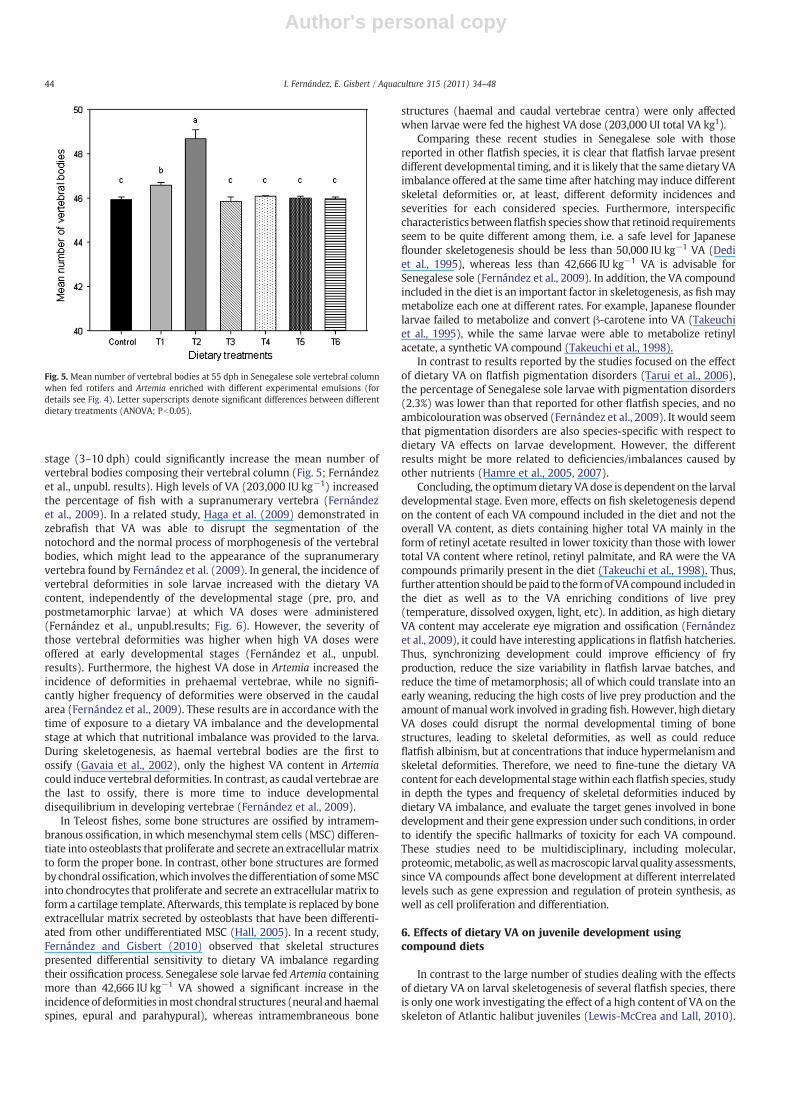
stage (3–10 dph) could significantly increase the mean number ofvertebral bodies composing their vertebral column (Fig. 5; Fernándezet al., unpubl. results). High levels of VA (203,000 IU kg−1) increasedthe percentage of fish with a supranumerary vertebra (Fernándezet al., 2009). In a related study, Haga et al. (2009) demonstrated inzebrafish that VA was able to disrupt the segmentation of thenotochord and the normal process of morphogenesis of the vertebralbodies, which might lead to the appearance of the supranumeraryvertebra found by Fernández et al. (2009). In general, the incidence ofvertebral deformities in sole larvae increased with the dietary VAcontent, independently of the developmental stage (pre, pro, andpostmetamorphic larvae) at which VA doses were administered(Fernández et al., unpubl.results; Fig. 6). However, the severity ofthose vertebral deformities was higher when high VA doses wereoffered at early developmental stages (Fernández et al., unpubl.results). Furthermore, the highest VA dose in Artemia increased theincidence of deformities in prehaemal vertebrae, while no signifi-cantly higher frequency of deformities were observed in the caudalarea (Fernández et al., 2009). These results are in accordance with thetime of exposure to a dietary VA imbalance and the developmentalstage at which that nutritional imbalance was provided to the larva.During skeletogenesis, as haemal vertebral bodies are the first toossify (Gavaia et al., 2002), only the highest VA content in Artemiacould induce vertebral deformities. In contrast, as caudal vertebrae arethe last to ossify, there is more time to induce developmentaldisequilibrium in developing vertebrae (Fernández et al., 2009).
In Teleost fishes, some bone structures are ossified by intramem-branous ossification, in which mesenchymal stem cells (MSC) differen-tiate into osteoblasts that proliferate and secrete an extracellular matrixto form the proper bone. In contrast, other bone structures are formedby chondral ossification,which involves thedifferentiation of someMSCinto chondrocytes that proliferate and secrete an extracellularmatrix toform a cartilage template. Afterwards, this template is replaced by boneextracellular matrix secreted by osteoblasts that have been differenti-ated from other undifferentiated MSC (Hall, 2005). In a recent study,Fernández and Gisbert (2010) observed that skeletal structurespresented differential sensitivity to dietary VA imbalance regardingtheir ossification process. Senegalese sole larvae fed Artemia containingmore than 42,666 IU kg−1 VA showed a significant increase in theincidence of deformities inmost chondral structures (neural andhaemalspines, epural and parahypural), whereas intramembraneous bone
structures (haemal and caudal vertebrae centra) were only affectedwhen larvae were fed the highest VA dose (203,000 UI total VA kg1).
Comparing these recent studies in Senegalese sole with thosereported in other flatfish species, it is clear that flatfish larvae presentdifferent developmental timing, and it is likely that the same dietary VAimbalance offered at the same time after hatching may induce differentskeletal deformities or, at least, different deformity incidences andseverities for each considered species. Furthermore, interspecificcharacteristics betweenflatfish species show that retinoid requirementsseem to be quite different among them, i.e. a safe level for Japaneseflounder skeletogenesis should be less than 50,000 IU kg−1 VA (Dediet al., 1995), whereas less than 42,666 IU kg−1 VA is advisable forSenegalese sole (Fernández et al., 2009). In addition, the VA compoundincluded in the diet is an important factor in skeletogenesis, as fishmaymetabolize each one at different rates. For example, Japanese flounderlarvae failed to metabolize and convert β-carotene into VA (Takeuchiet al., 1995), while the same larvae were able to metabolize retinylacetate, a synthetic VA compound (Takeuchi et al., 1998).
In contrast to results reported by the studies focused on the effectof dietary VA on flatfish pigmentation disorders (Tarui et al., 2006),the percentage of Senegalese sole larvae with pigmentation disorders(2.3%) was lower than that reported for other flatfish species, and noambicolourationwas observed (Fernández et al., 2009). It would seemthat pigmentation disorders are also species-specific with respect todietary VA effects on larvae development. However, the differentresults might be more related to deficiencies/imbalances caused byother nutrients (Hamre et al., 2005, 2007).
Concluding, the optimumdietary VA dose is dependent on the larvaldevelopmental stage. Even more, effects on fish skeletogenesis dependon the content of each VA compound included in the diet and not theoverall VA content, as diets containing higher total VA mainly in theform of retinyl acetate resulted in lower toxicity than those with lowertotal VA content where retinol, retinyl palmitate, and RA were the VAcompounds primarily present in the diet (Takeuchi et al., 1998). Thus,further attention shouldbepaid to the formofVA compound included inthe diet as well as to the VA enriching conditions of live prey(temperature, dissolved oxygen, light, etc). In addition, as high dietaryVA content may accelerate eye migration and ossification (Fernándezet al., 2009), it could have interesting applications in flatfish hatcheries.Thus, synchronizing development could improve efficiency of fryproduction, reduce the size variability in flatfish larvae batches, andreduce the time of metamorphosis; all of which could translate into anearly weaning, reducing the high costs of live prey production and theamount of manualwork involved in grading fish. However, high dietaryVA doses could disrupt the normal developmental timing of bonestructures, leading to skeletal deformities, as well as could reduceflatfish albinism, but at concentrations that induce hypermelanism andskeletal deformities. Therefore, we need to fine-tune the dietary VAcontent for each developmental stagewithin each flatfish species, studyin depth the types and frequency of skeletal deformities induced bydietary VA imbalance, and evaluate the target genes involved in bonedevelopment and their gene expression under such conditions, in orderto identify the specific hallmarks of toxicity for each VA compound.These studies need to be multidisciplinary, including molecular,proteomic,metabolic, aswell asmacroscopic larval quality assessments,since VA compounds affect bone development at different interrelatedlevels such as gene expression and regulation of protein synthesis, aswell as cell proliferation and differentiation.
6. Effects of dietary VA on juvenile development usingcompound diets
In contrast to the large number of studies dealing with the effectsof dietary VA on larval skeletogenesis of several flatfish species, thereis only one work investigating the effect of a high content of VA on theskeleton of Atlantic halibut juveniles (Lewis-McCrea and Lall, 2010).
Fig. 5. Mean number of vertebral bodies at 55 dph in Senegalese sole vertebral columnwhen fed rotifers and Artemia enriched with different experimental emulsions (fordetails see Fig. 4). Letter superscripts denote significant differences between differentdietary treatments (ANOVA; Pb0.05).
44 I. Fernández, E. Gisbert / Aquaculture 315 (2011) 34–48

Author's personal copy
In that study, fish (4.6 g of mean wet weight) were fed two differentcompound diets containing different levels of VA (6649 and52,837 IU kg−1, retinyl acetate was the VA form added to increasethe VA content in the diet) during 14 weeks. Authors found nodifferences between fish groups in several physiological and mor-phological parameters, such as the feed conversion ratio, weight gain,condition factor, hepatosomatic index, liver lipid content, liverα-tocopherol, vertebral ash, phosphorus content, and survival rate.However, a significant increase in retinol liver content was observed,reaching 11.4 μg of retinol g−1 in fish fed the highest VA contentcompared to 5.6 μg of retinol g-1 in control fish. Despite the fact thatthe VA content in the supplemented VA diet was eight times higher
than in the control diet, liver VA content in fish fed a high VA diet wasonly twice than in control fish. With respect to fatty acid compositionof juvenile liver, differences were found in 20:5n−3 (eicosapentae-noic acid, EPA) and n-3 total composition. Fish fed with a high VA dietpresented a mean reduction in fatty acid composition of 14% in EPAand 8.2% in total n−3 fatty acid series with respect to control fish.Although the author did not discuss that result regarding the effect ofdietary VA content, it should be noted that gene expression andprotein levels of cellular retinol-binding protein type II could beaffected by dietary fat (Goda et al., 1994), and in addition, EPA is theprecursor of prostaglandins, thromboxanes, and leucotrienes. Pros-taglandines are potent regulators of bone formation and bone
Fig. 6. Frequency of abnormalities (%) in the axial skeleton of Senegalese sole at 55 dph when fed rotifers and Artemia enriched with different experimental emulsions (for detailssee Fig. 4).
45I. Fernández, E. Gisbert / Aquaculture 315 (2011) 34–48

Author's personal copy
resorption that can have both stimulatory and inhibitory effects(Raisz, 1995; Poulsen et al., 2007). Therefore, in addition to the directeffect of dietary VA content on skeletogenesis, different liver EPAcontents could also indirectly affect bone development.
Regarding skeletal deformities detected and their frequency, nosignificant differences were found in the mean number of vertebraecomposing the vertebral column. This fact may be due to the numberof vertebrae being already established prior to the beginning of thefeeding trial. Although, scoliosis, fused, and compressed vertebraewere not observed in control fish, they were found in fish fed a highVA diet, where 60% of fish presented scoliosis, mainly at the anteriorregion of the vertebral column. These results suggest that a safe levelfor Atlantic halibut skeletal development should be less than52,837 IU kg-1. However, Moren et al. (2004a) and Hernández et al.(2005) found that the optimum dietary VA content for juvenilegrowth should be around 8000–9000 IU kg−1 for Atlantic halibut andJapanese flounder, respectively, being these values different than theoptimum level found for other round fish species (Hernández et al.,2003; Mohamed et al., 2003). However, it should be noted that thoseworks, Moren et al. (2004a) and Hernández et al. (2005), did notevaluate the impact of VA in the skeleton, and theywere only focussedon growth performance. Therefore, the optimum dietary VA level forproper skeletogenesis during the juvenile stage in flatfishes is still tobe determined, although results from Lewis-McCrea and Lall (2010)suggest that it might be less than 52,837 IU kg−1.
7. Conclusions
Of the considerable work already completed towards the under-standing of the effects of dietary VA content on flatfish skeletogenesisduring larval development, many were focused on Japanese flounder.The different nutritional approaches applied (bath exposure versusdietary dose-response trials), diets (compound feed versus enrichedlive preys), and chemical forms of VA tested, among other factors,makes comparing the results from those studies difficult. All theseworks highlight the need to study the dietary VA imbalance for eachflatfish species. However, some conclusions from all the dietary VAstudies can be summarized: (i) VA imbalances have important effectson flatfish skeletogenesis, and these effects depend on the species(ontogenic development and physiology), developmental stage(larva, juvenile or broodstock), VA chemical form used to fulfil dietaryrequirements, as well as all the abiotic and biotic circumstances thattake place during fish rearing (temperature, flow currents, photope-riod, etc.); (ii) broodstock VA requirements are still unknown for eachflatfish species; (iii) larvae from different flatfish species havedifferent requirements for optimum dietary VA content: less than44,666 and 50,000 IU kg−1 for Senegalese sole (Fernández et al.,2009) and Japanese flounder larvae (Dedi et al., 1995), respectively;and (iv) dietary VA effects on juvenile skeletogenesis seems to belower than during larval development and the optimumones could bearound 8000–9,000 IU kg−1 with regard to growth performance(Moren et al., 2004a; Hernández et al., 2005), levels that are higherthan those suggested for salmonids (2500 IU kg−1; NRC 1993); but atleast it should be lower than 52,837 IU kg−1 (Lewis-McCrea and Lall,2010) from the perspective of skeletogenesis.
Finally, during all developmental fish stages, authors recom-mended the use of two retinoid compounds as the better to fulfill thenutritional VA requirements of fishes; however, different reasons leadus to propose those as the better ones. On the one hand, during earlylarval development retinyl palmitate and retinyl acetate are recom-mended, although both could present a limited rate of its conversionto retinol, retinal and RA. Retinyl palmitate is more stable and lesstoxic than retinol, retinal and RA, and a source of lipids (retynilesters), while retinyl acetate has the advantage that is wellaccumulated and less metabolized by live preys (Takeuchi et al.,1998; Giménez et al., 2007). On the other hand, both retinoid
compounds are further recommended to be included in diets as themain source of VA for juveniles and broodstocks, as fishes in thesedevelopmental stages have a fully developed liver with the necessaryenzymes to tightly convert those VA compounds to retinol, retinal,and finally RA.
Acknowledgements
Authors are greatly indebted to the technical staff of IRTA-SantCarles research centre for their excellent technical assistance duringlarvae trials with Senegalese sole, especially to M. Monllaó, J. Canoura,S. Molas, M. Matas and M. Pimentel. In addition, thanks go to N. Gras,M. Sastre, and Dr. A. Estévez for their assistance in biochemistryanalysis; Dr. F. Hontoria from Torre de la Sal Aquaculture Institute forVA quantification in diets and larvae; Dr. Carmen Sarasquete and Dr. J.B. Ortiz-Delgado for thyroid hormones analyses; and Dr. Santosh Lallfor sharing knowledge that greatly enriched this review. I.F. wants tothank the special help given by C. Reverté in recompiling bibliographyand C. Santes for their English review of the manuscript. Funding wasprovided to EG by the Ministry of Education and Culture (MEC) of theSpanish Government (projects AGL2005-02478; AGL2008-03897-CA-01/ACU, and HF2008-0079). I.F was supported by a predoctoral MECfellowship (MEC, Spain).
References
Alsop, D., Brown, S., Van der Kraak, G., 2001. Development of a retinoic acid receptor-binding assay with rainbow trout tissue: characterization of retinoic acid binding,receptor tissue distribution, and developmental changes. Gen. Comp. Endocrinol.123, 254–267.
Alsop, D., Matsumoto, J., Brown, S., van Der Kraak, G., 2008. Retinoid requirements inthe reproduction of zebrafish. Gen. Comp. Endocrinol. 156, 51–62.
Arima, K., Shiotsugu, J., Niu, R., Khandpur, R., Martinez, M., Shin, Y., Koide, T., Cho, K.,Kitayama, A., Ueno, N., Chandraratna, R., Blumberg, B., 2005. Global analysis of RAR-responsive genes in the Xenopus neurula using cDNA microarrays. Dev. Dyn. 234,414–431.
Balmer, J.E., Blomhoff, R., 2002. Gene expression regulation by retinoic acid. J. Lipid Res.43, 1773–1808.
Balmer, J.E., Blomhoff, R., 2005. A robust characterization of retinoic acid responseelements based on a comparison of sites in three species. J. Steroid Biochem. Mol.Biol. 96, 347–354.
Bastien, J., Rochette-Egly, C., 2004. Nuclear retinoid receptors and the transcription ofretinoid-target genes. Gene 328, 1–16.
Bengtson, D.A., 1999. Aquaculture of summer flounder (Paralichthys dentatus): status ofknowledge, current research and future research priorities. Aquaculture 176,39–49.
Biesalski, H.K., Hemmes, C., Hanafy, M.E., Weiser, H., Zshaebitz, H., Stofft, E., 1996. Longterm administration of high dose vitamin A to rat does not cause fatalmalformations: macroscopic, skeletal and physicochemical findings. J. Nutr. 126,973–983.
Bisbal, G., Bengtson, D.A., 1995. Description of the starving condition in summerflounder, Paralichthys dentatus, early life stages. Fish. Bull. 93, 217–230.
Boglione, C., Gagliardi, F., Scardi, M., Cataudella, S., 2001. Skeletal descriptors andquality assessment in larvae and post-larvae of wild-caught and hatchery-rearedgilthead sea bream (Sparus aurata L. 1758). Aquaculture 192, 1–22.
Bolker, J.A., Hill, C.R., 2000. Pigmentation development in hatchery-reared flatfishes. J.Fish Biol. 56, 1029–1052.
Cahu, C., Zambonino Infante, J.L., Takeuchi, T., 2003. Nutritional components affectingskeletal development in fish larvae. Aquaculture 227, 1–4.
Clagett-Dame, M., DeLuca, H.F., 2002. The role of vitamin A in mammalian reproductionand embryonic development. Annu. Rev. Nutr. 22, 347–381.
Costaridis, P., Horton, C., Zeitilinger, J., Holder, N., Maden, M., 1996. Endogenousretinoids in the zebrafish embryo and adult. Dev. Dyn. 205, 41–51.
Dedi, J., Takeuchi, T., Seikai, T., Watanabe, T., 1995. Hypervitaminosis and safe levels ofvitamin A for larval Japanese flounder (Paralichthys olivaceus) fed Artemia nauplii.Aquaculture 133, 135–146.
Divanach, P., Boglione, C., Menu, B., Koumoundouros, G., Kentouri, M., Cautadella, S.,1996. Abnormalities in finfish mariculture: an overview of the problem, causes andsolutions. Seabass and Seabream Culture: Problems and Prospects. Handbook ofContributions and Short Communications. Verona, Italy, pp. 45–51. October 16–18.
Duester, G., 1996. Involvement of alcohol dehydrogenase, short chain dehydrogenase/reductase, aldehyde dehydrogeanse, and cytochrome p450 in the control ofretinoid signaling by activation of retinoic acid synthesis. Biochemistry 35,12221–12227.
Duester, G., 2000. Families of retinoid dehydrogenases regulating vitamin A function:production of visual pigment and retinoic acid. Eur. J. Biochem. 267, 4315–4324.
46 I. Fernández, E. Gisbert / Aquaculture 315 (2011) 34–48

Author's personal copy
Duester, G., Mic, F.A., Molotkov, A., 2003. Cytosolic retinoid dehydrogenases governubiquitous metabolism of retinol to retinaldehyde followed by tissuespecificmetabolism to retinoic acid. Chem. Biol. Interact. 143–144, 201–210.
Durston, A.J., Timmermans, J.P.M., Hage, W.J., Hendriks, H.F.J., Devries, N.J., Heideveld,M., Nieuwkoop, P.D., 1989. Retinoic acid causes an anteriorposterior transforma-tion in the developing central nervous-system. Nature 340, 140–144.
Estévez, A., Kanazawa, A., 1995. Effect of n-3 PUFA and vitamin A Artemia enrichment onpigmentation success of turbot, Scophthalmusmaximus (L.). Aquacult. Nutr. 1, 159–168.
FAO Fishery and Aquaculture Information and Statistics Service, 2010. Aquacultureproduction. http://www.fao.org/figis/servlet/TabLandArea?tb_ds=Aquaculture&tb_mode=TABLE&tb_act=SELECT&tb_grp=COUNTRY. 2010.
Fernández, I., Gisbert, E., 2010. Senegalese sole bone tissue originated from chondralossification is more sensitive than dermal bone to high vitamin A content inenriched Artemia. J. Appl. Ichthyol. 26, 344–349.
Fernández, I., Hontoria, F., Ortiz-Delgado, J.B., Kotzamanis, Y., Estévez, A., Zambonino-Infante, J.L., Gisbert, E., 2008. Larval performance and skeletal deformities in farmedgilthead sea bream (Sparus aurata) fed with graded levels of Vitamin A enrichedrotifers (Brachionus plicatilis). Aquaculture 283, 102–115.
Fernández, I., Pimentel, M.S., Ortiz-Delgado, J.B., Hontoria, F., Sarasquete, C., Estévez, A.,Zambonino-Infante, J.L., Gisbert, E., 2009. Effect of dietary vitaminAonSenegalese sole(Solea senegalensis) skeletogenesis and larval quality. Aquaculture 295, 250–265.
Furuita, H., Tanaka, H., Takeshi, Y., Shiraishi, M., Takeuchi, T., 2001. Effects of high doseof vitamin A on reproduction and egg quality of Japanese flounder Paralichthysolivaceus. Fish. Sci. 67, 606–613.
Furuita, H., Yamamoto, T., Shima, T., Suzuki, N., Takeuchi, T., 2003. Effect of dietaryarachidonic acid levels on larval and egg quality of the Japanese flounderParalichthys olivaceus. Aquaculture 220, 725–735.
Gavaia, P.J., Dinis, M.T., Cancela, M.L., 2002. Osteological development and abnormal-ities of the vertebral column and caudal skeleton in larval and juvenile stages ofhatchery-reared Senegalese sole (Solea senegalensis). Aquaculture 211, 305–323.
Geffen, A.J., van der Veer, H.W., Nash, R.D.M., 2007. The cost of metamorphosis inflatfishes. J. Sea Res. 58, 35–45.
Germain, P., Chambon, P., Eichele, G., Evans, R.M., Lazar, M.A., Leid, M., de Lera, A.R.,Lotan, R., Mangelsdorf, D.J., Gronemeyer, H., 2006a. International Union ofPharmacology. LX. Retinoic acid receptors. Pharmacol. Rev. 58, 712–725.
Germain, P., Chambon, P., Eichele, G., Evans, R.M., Lazar, M.A., Leid, M., de Lera, A.R.,Lotan, R., Mangelsdorf, D.J., Gronemeyer, H., 2006b. International Union ofPharmacology, LXIII. Retinoid X receptors. Pharmacol. Rev. 58, 760–772.
Giménez, G., Kotzamanis, Y., Hontoria, F., Estévez, A., Gisbert, E., 2007. Modellingretinoid content in live prey: a tool for evaluating the nutritional requirements anddevelopment studies in fish larvae. Aquaculture 267, 76–82.
Goda, T., Yasutake, H., Takase, S., 1994. Dietary-fat regulates cellular retinol-bindingprotein-II gene-expression in rat jejunum. Biochim. Biophys. Acta 1200, 34–40.
Haga, Y., Suzuki, T., Takeuchi, T., 2002a. Retinoic acid isomers produce malformations inpostembryonic development of Japanese flounder Paralichthys olivaceus. Zool. Sci.19, 1105–1112.
Haga, Y., Takeuchi, T., Seikai, T., 2002b. Influence of all-trans retinoic acid on pigmentationand skeletal formation in larval Japanese flounder. Fish. Sci. 68, 560–570.
Haga, Y., Suzuki, T., Kagechika, H., Takeuchi, T., 2003. A retinoic acid receptor-selectiveagonist causes jaw deformity in the Japanese flounder, Paralichthys olivaceus.Aquaculture 221, 381–392.
Haga, Y., Takeuchi, T., Seikai, T., 2004. Changes of retinoid contents in larval Japaneseflounder Paralichthys olivaceus and Artemia nauplii enriched with a large dose ofall-trans retinoic acid. Fish. Sci. 70, 436–444.
Haga, Y., Tarui, F., Ohta, K., Shima, Y., Takeuchi, T., 2006. Effect of light irradiation ondynamics of vitamin A compounds in rotifers and Artemia. Fish. Sci. 72, 1020–1026.
Haga, Y., Dominique, V.J., Du, S.J., 2009. Analyzing notochord segmentation andintervertebral disc formation using the twhh:gfp transgenic zebrafish model.Transgen. Res. 18, 669–683.
Hall, B.K., 2005. Bones and Cartilage: Developmental Skeletal Biology. Academic Press,London.
Hamre, K., Opstad, I., Espe, M., Solbakken, J., Hemre, G.I., Pittman, K., 2002. Nutrientcomposition and metamorphosis success of Atlantic halibut (Hippoglossus hippoglos-sus L.) larvae fed natural zooplancton or Artemia. Aquacult. Nutr. 8, 139–148.
Hamre, K., Moren, M., Solbakken, J., Opstad, I., Pittmann, K., 2005. The impact ofnutrition on metamorphosis in Atlantic halibut (Hippoglossus hippoglossus L.).Aquaculture 250, 555–565.
Hamre, K., Holen, E., Moren, M., 2007. Pigmentation and eye migration in Atlantichalibut (Hippoglossus hippoglossus L.) larvae: new findings and hypotheses.Aquacult. Nutr. 13, 65–80.
Harboe, T., Mangor-Jensen, A., Moren, M., Hamre, K., Rønnestad, I., 2009. Control of lightcondition affects the feeding regime and enables successful eye migration inAtlantic halibut juveniles. Aquaculture 290, 250–255.
Hashimoto, H., Aritaki,M., Uozumi, K., Uji, S., Kurokawa, T., Suzuki, T., 2007. Embryogenesisand expression profiles of charon and nodal-pathway genes in sinistral (Paralichthysolivaceus) and dextral (Verasper variegatus) flounders. Zool. Sci. 24, 137–146.
Hata, M., Hata, M., 1973. Carotenoid pigments in goldfish (Carassius auratus L). 6. Effectof ACTH and phenylthiourea on discoloration and carotenoid metabolism. Comp.Biochem. Physiol. 46, 95–101.
Hernández, H.L.H., Teshima, S., Tanaka, Y., Ishikawa, M., Koshio, S., 2003. Dietaryvitamin A requirements of juvenile red sea bream Chrysophrys major. Program andAbstract Book of the Asian-Pacific Aquaculture 2003. Bangkok, Thailand, September22–25. World Aquaculture Society, Bangkok, Thailand, p. 107.
Hernández, L.H.H., Teshima, S.I., Ishikawa, M., Alam, S., Koshio, S., Tanaka, Y., 2005.Dietary vitamin A requirements of juvenile Japanese flounder Paralichthysolivaceus. Aquacult. Nutr. 11, 3–9.
Howell, B., Conceicão, L., Prickett, R., Cañavate, P., Mañanos, E., 2009. Sole farming:nearly there but not quite? Aquac. Eur. 34, 24–27.
Ingenbleek, Y., Young, V., 1994. Transthyretin (prealbumin) in health and disease:nutritional implications. Annu. Rev. Nutr. 14, 495–533.
Irie, T., Seki, T., 2002. Retinoid composition and retinal localization in the eggs of teleostfishes. Comp. Biochem. Physiol. B 131, 209–219.
Kakkad, B., Ong, D.E., 1988. Reduction of retinaldehyde bound to cellular retinol-binding protein (type II) by microsomes from rat small intestine. J. Biol. Chem. 263,12916–12919.
Klaren, P.H.M., Wunderink, Y.S., Yúfera, M., Mancera, J.M., Flik, G., 2008. The thyroidgland and thyroid hormones in Senegalese sole (Solea senegalensis) during earlydevelopment and metamorphosis. Gen. Comp. Endocrin. 155, 686–694.
Koumoundouros, G., Oran, G., Divanach, P., Stefanakis, S., Kentouri, M., 1997. Theopercular complex deformity in intensive gilthead sea bream (Sparus aurata L.)larviculture. Moment of apparition and description. Aquaculture 156, 165–177.
Koumoundouros, G., Maingot, E., Divanach, P., Kentouri, M., 2002. Kyphosis in rearedsea bass (Dicentrarchus labrax L.): ontogeny and effects on mortality. Aquaculture209, 49–58.
Lall, S.P., Lewis-McCrea, L., 2007. Role of nutrients in skeletal metabolism and pathologyin fish, an overview. Aquaculture 267, 3–19.
Lavens, P., Leregue, E., Jaunet, H., Brunel, A., Dhert, P.H., Sorgeloos, P., 1999. Effect ofdietary essential fatty acids and vitamins on egg quality in turbot broodstocks.Aquacult. Int. 7, 225–240.
Lefebvre, P., Martin, P.J., Flajollet, S., 2005. Transcriptional activities of retinoic acidreceptors. Vitam. Horm. 70, 199–264.
Lewis-McCrea, L.M., Lall, S.P., 2006. Development of the axial skeleton and skeletalabnormalities of Atlantic halibut (Hippoglossus hippoglossus) from first feedingthrough metamorphosis. Aquaculture 257, 124–135.
Lewis-McCrea, L.M., Lall, S.P., 2010. Effects of phosphorus and vitamin C deficiency,vitamin A toxicity, and lipid peroxidation on skeletal abnormalities in Atlantichalibut (Hippoglossus hippoglossus) J. Appl. Ichthyol. 26, 334–343.
Lewis-McCrea, L.M., Lall, S.P., Witten, P.E., 2004. Morphological descriptions of the earlystages of spine and vertebral development in hatchery-reared larval and juvenileAtlantic halibut (Hippoglossus hippoglossus). Aquaculture 241, 47–59.
Lubzens, E., Lissauer, L., Levavi-Sivan, B., Avarre, J.C., Sammar, M., 2003. Carotenoid andretinoid transport to fish oocytes and eggs: what is the role of retinol bindingprotein? Mol. Aspects Med. 24, 441–457.
Martinez, G.M., Bolker, J.A., 2003. Embryonic and larval staging of summer flounder(Paralichthys dentatus). J. Morphol. 255, 162–176.
Martinez, G.M., Baron, M.P., Bolker, J.A., 2007. Skeletal and pigmentation defectsfollowing retinoic acid exposure in larval summer flounder, Paralichthys dentatus. J.World Aquac. Soc. 38, 353–366.
Masaki, T., Matsuuras, T., Ohkawa, K., Miyamura, T., Okazaki, I., Watanabe, T., Suzuki, T.,2006. All-trans retinoic acid down-regulates human albumin gene expressionthrough the induction of C/EBPβ-LIP. Biochem. J. 397, 345–353.
Mazurais, D., Glynatsi, N., Darias, M.J., Christodoulopoulou, S., Cahu, C.L., Zambonino-Infante, J.L., Koumoundouros, G., 2009. Optimal levels of dietary vitamin Afor reduced deformity incidence during development of European sea basslarvae (Dicentrarchus labrax) depend on malformation type. Aquaculture 294,262–270.
Michie, I., 2001. Causes of downgrading in the salmon farmin industry. Fish FarmedQuality. Blackwell Science, Oxford, pp. 129–136.
Miki, W., Yamaguchi, K., Konosu, S., Watanabe, T., 1984. Metabolism of dietarycarotenoids in eggs of red sea bream. Comp. Biochem. Physiol. 77, 665–668.
Minami, T., 1982. Study on the early life-history of flatfishes in Wakasa bay. 6. The earlylife-history of a Paralichthys olivaceus. B. JPN. Soc. Sci. Fish. 48, 1581–1588.
Mohamed, J.S., Sivaram, V., Roy, T.S.C., Maria, S.P., Murudagass, S., Hussain, M.R., 2003.Dietary vitamin A requirements of juvenile greasy grouper (Epinephelus tauvina).Aquaculture 219, 693–701.
Monroig, O., Navarro, J.C., Amat, F., González, P., Hontoria, F., 2006. Effects of nauplialdensity, product concentration and product dosage on the survival of the naupliiand EFA incorporation during Artemia enrichment with liposomes. Aquaculture261, 659–669.
Moren, M., Opstad, I., Pittman, K., Hamre, K., 2001. A comparision of retinol, retinal andretinyl ester concentrations in larvae of Atlantic halibut (Hippoglossus hippoglossusL.) fed Artemia or Zooplankton. In: Hendry, C.I., Van Stappen, G., Wille, M.,Sorgeloos, P. (Eds.), European Aquaculture Society, Special Publication, vol. 30.Oostende, Belgium.
Moren, M., Næss, T., Hamre, K., 2002. Converstion of β-carotene, canthaxanthin andastaxanthin to vitamin A in Atlantic halibut (Hippoglossus hippoglossus L.) juveniles.Fish Physiol. Biochem. 27, 71–80.
Moren, M., Opstad, I., Berntssen, M.H.G., Zambonino Infante, J.L., Hamre, K., 2004a. Anoptimum level of vitamin A supplements for Atlantic halibut (Hippoglossushippoglossus L.) juveniles. Aquaculture 235, 587–599.
Moren, M., Opstad, I., Hamre, K., 2004b. A comparision of retinol, retinal and retinylester concentrations in larvae of Atlantic halibut (Hippoglossus hippoglossus L.) fedArtemia or zooplankton. Aquacult. Nutr. 10, 253–259.
Moren, M., Gundersen, T.E., Hamre, K., 2005. Quantitative and qualitative analysis ofretinoids in Artemia and copepods by HPLC and diode array detection. Aquaculture246, 359–365.
Murayama, A., Suzuki, T., Matsui, M., 1997. Photoisomerization of retinoic acids inethanol under room light: a warning for cell biological study of geometrical isomersof retinoids. J. Nutr. Sci. Vitaminol. 43, 167–176.
N'soukpoé-Kossi, C.N., Sedaghat-Herati, R., Ragi, C., Hotchandani, S., Tajmir-Riahi, H.A.,2007. Retinol and retinoic acid bind human serum albumin: stability and structuralfeatures. Int. J. Biol. Macromol. 40, 484–490.
47I. Fernández, E. Gisbert / Aquaculture 315 (2011) 34–48

Author's personal copy
Nakamura, T., Aikawa, T., Iwamoto-Enomoto, M., Iwamoto, M., Higuchi, Y., Maurizio, P.,Kinto, N., Yamaguchi, A., Noji, S., Kurisu, K., Matsuya, T., 1997. Induction ofosteogenic differentiation by hedgehog proteins. Biochim. Biophys. Res. Commun.237, 465–469.
Napoli, J.L., 1996. Biochemical pathways of retinoid transport, metabolism, and signaltransduction. Clin. Immunol. Immunopathol. 80, S52–S62.
Napoli, J.L., 2000. Retinoic acid: Its biosynthesis and metabolism. Prog. Nucleic Acid Res.Mol. Biol. 63, 139–188.
National Research Council, 1993. Nutrient requirements of fish. Committee on AnimalNutrition Board on Agriculture. National Academy Press, Washington, D.C.
Okada, N., Takagi, Y., Tanaka, M., Tagawa, M., 2003. Fine structure of soft and hardtissues involved in eye migration in metamorphosing Japanese flounder (Para-lichthys olivaceus). Anat. Rec. 273A, 663–668.
Olson, J.A., 1989. Provitamin-A function of carotenoids. The conversion of beta-caroteneinto vitamin A. J. Nutr. 119, 105–108.
Ørnsrud, R., Graff, I.E., Høie, S., Totland, G.K., Hemre, G.I., 2002. Hypervitaminosis A infirst-feeding fry of the Atlantic salmon (Salmo salar L.). Aquacult. Nutr. 8, 7–13.
Osse, J.W.M., Van den Boogaart, J.G.M., 1997. Size of flatfish larvae at transformation,functional demands and historical constraints. J. Sea Res. 37, 229–239.
Palace, V.P., Werner, J., 2006. Vitamins A and E in thematernal diet influence egg qualityand early life stage development in fish: a review. Sci. Mar. 70, 41–57.
Pepe, I.M., 1999. Rhodopsin and phototransduction. J. Photochem. Photobiol. B. Biol. 48,1–10.
Poulsen, R.C., Moughan, P.J., Kruger, M.C., 2007. Long-chain polyunsaturated fatty acidsand the regulation of bone metabolism. Exp. Biol. Med. 232, 1275–1288.
Power, D.M., Silva, N., Campinho, M.A., 2008. Metamorphosis. In: Finn, R.N., Kapoor, B.G.(Eds.), Fish Larval Physiology. Science Publishers, New Hampshire, pp. 607–638.
Raisz, L.G., 1995. Physiological and pathological roles of prostaglandins and othereicosanoids in bone metabolism. J. Nutr. 125, S2024–S2027.
Reijntjes, S., Blentic, A., Gale, E., Maden, M., 2005. The control of morphogen signalling:Regulation of the synthesis and catabolism of retinoic acid in the developingembryo. Dev. Biol. 285, 224–237.
Rønnestad, I., Hemre, G.I., Finn, R.N., Lie, Ø., 1998a. Alternate sources and dynamics ofvitamin A and its incorporation into the eyes during the early endotrophic andexotrophic larval stages of Atlantic halibut (Hippoglossus hippoglossus L.). Comp.Biochem. Physiol. 119, 787–793.
Rønnestad, I., Helland, S., Lie, Ø., 1998b. Feeding Artemia to larvae of Atlantic halibut(Hippoglossus hippoglossus L.) results in lower larval vitamin A content comparedwith feeding copepods. Aquaculture 165, 159–164.
Ross, S.A., Caffery, P.J., Draguer, U.C., De Luca, L.M., 2000. Retinoids in embryonaldevelopment. Physiol. Rev. 80, 1021–1054.
Sæle, Ø., Solbakken, J.S., Watanabe, K., Hamre, K., Pittman, K., 2003. The effect of diet onossification and eye migration in Atlantic halibut larvae (Hippoglossus hippoglossusL.). Aquaculture 220, 683–696.
Sæle, Ø., Solbakken, J.S., Watanabe, K., Hamre, K., Power, D., Pittman, K., 2004. Staging ofAtlantic halibut (Hippoglossus hippoglossus L.) from first feeding throughmetamorphosis, including cranial ossification independent of eye migration.Aquaculture 239, 445–465.
Sæle, Ø., Smáradóttir, H., Pittman, K., 2006a. Twisted story of eye migration in flatfish. J.Morph. 267, 730–738.
Sæle, Ø., Silva, N., Pittman, K., 2006b. Post-embryonic remodelling of neurocranialelements: a comparative study of normal versus abnormal eye migration in aflatfish, the Atlantic halibut. J. Anat. 209, 31–41.
Sawanboochun, J., Roy, W.J., Robertson, D.A., Bell, J.G., 2008. The impact of dietarysupplementation with astaxanthin on egg quality in Atlantic cod broodstock(Gadus morhua, L.). Aquaculture 283, 97–101.
Schreiber, A.M., 2006. Asymmetric craniofacial remodeling and lateralized behavior inlarval flatfish. J. Exp. Biol. 209, 610–621.
Solbakken, J.S., Pittman, K., 2004. Photoperiodic modulation of metamorphosis inAtlantic halibut (Hippoglossus hippoglossus L.). Aquaculture 232, 613–625.
Suzuki, T., Oochara, I., Kurokawa, T., 1998. Hoxd-4 expression during pharyngeal archdevelopment in flounder (Paralichthys olivaceus) embryos and effects of retinoicacid on expression. Zool. Sci. 15, 57–67.
Suzuki, T., Oochara, I., Kurokawa, T., 1999. Retinoic acid given at late embryonic stagedepresses sonic hedgehog and Hoxd-4 expression in the pharyngeal area andinduces skeletal malformation in flounder (Paralichthys olivaceus) embryos. Dev.Growth Differ. 41, 143–152.
Suzuki, T., Srivastava, A.S., Kurokawa, T., 2000. Experimental induction of jaw, gill andpectoral fin malformation in Japanese flounder, Paralichthys olivaceus, larvae.Aquaculture 185, 175–187.
Suzuki, T., Haga, Y., Takeuchi, T., Uji, S., Hashimoto, H., Kurokawa, T., 2003.Differentiation of chondrocytes and scleroblasts during dorsal fin skeletogenesisin flounder larvae. Dev. Growth Differ. 45, 435–448.
Suzuki, T., Washio, Y., Aritaki, M., Fujinami, Y., Shimizu, D., Uji, S., Hashimoto, H., 2009.Metamorphic pitx2 expression in the left habenula correlated with lateralization ofeye-sidedness in flounder. Dev. Growth Differ. 51, 797–808.
Takeuchi, T., Dedi, J., Ebisawa, C.,Watanabe, T., Seikai, T.,Hosoya,K., Nakazoe, J.-I., 1995. Theeffect of β-carotene and vitamin A enriched Artemia nauplii on the malformation andcolor abnormality of larval Japanese flounder. Fish. Sci. 61, 141–148.
Takeuchi, T., Dedi, J., Haga, Y., Seikai, T., Watanabe, T., 1998. Effect of vitamin Acompounds on bone deformity in larval Japanese flounder (Paralichthys olivaceus).Aquaculture 169, 155–165.
Tanumihardjo, S.A., 2002. Factors influencing the conversion of carotenoids to retinol:bioavailability to bioconversion to bioefficacy. Int. J. Vit. Nutr. Res. 72, 40–45.
Tarui, F., Haga, Y., Ohta, K., Shima, Y., Takeuchi, T., 2006. Effect of Artemia naupliienriched with vitamin A palmitate on hypermelanosis on the blind side in juvenileJapanese flounder Paralichthys olivaceus. Fish. Sci. 72, 256–262.
Van der Meeren, T., Olsen, R.E., Hamre, K., Fyhn, H.J., 2008. Biochemical composition ofcopepods for evaluation of feed quality in production of juvenile marine fish.Aquaculture 274, 375–397.
Villalta, M., Estévez, A., 2005. Culture of Senegal sole larvae without the need forrotifers. Aquacult. Int. 13, 469–478.
Villalta, M., Estévez, A., Brandsen, M.P., 2005. Arachidonic acid enriched live prey inducesalbinism in Senegal sole (Solea senegalensis) larvae. Aquaculture 245, 193–209.
Villeneuve, L., Gisbert, E., Le Gall, M.M., Cahu, C.L., Zambonino-Infante, J.L., 2004.Expression and localization of some retinoid receptors during European sea bass(Dicentrarchus labrax) larvae development. Aquaculture 242, 537–551.
Villeneuve, L., Gisbert, E., Le Delliou, H., Cahu, C.L., Zambonino-Infante, J.L., 2005. Dietarylevels of all-trans retinol affect retinoid nuclear receptor expression and skeletaldevelopment in European sea bass larvae. Br. J. Nutr. 93, 791–801.
Villeneuve, L.A.N., Gisbert, E., Moriceau, J., Cahu, C.L., Zambonino, J.L., 2006. Intake ofhigh levels of vitamin A and polyunsaturated fatty acids during differentdevelopmental periods modifies the expression of morphogenesis genes inEuropean sea bass (Dicentrarchus labrax). Br. J. Nutr. 95, 677–687.
Watanabe, K., Washio, Y., Fujinami, Y., Aritaki, M., Uji, S., Suzuki, T., 2008. Adult-typepigment cells, which color the ocular sides of flounders at metamorphosis, localizeas precursor cells at the proximal parts of the dorsal and anal fins in early larvae.Dev. Growth Differ. 50, 731–741.
Weston, A.D., Chandraratna, R.A., Torchia, J., Underhill, T.M., 2002. Requirement forRAR-mediated gene repression in skeletal progenitor differentiation. J. Cell Biol.158, 39–51.
Wexler, J.A., Sharretts, J., 2007. Thyroid and bone. Endocrinol. Metab. Clin. N. Am. 36,673–687.
Zanotti, G., Folli, C., Cendron, L., Alfieri, B., Nishida, S.K., Gliubich, F., Pasquato, N., Negro,A., Berni, R., 2008. Structural and mutational analyses of protein-proteininteractions between transthyretin and retinol-binding protein. FEBS J. 275,5841–5854.
Zarza, C., Padrós, F., Estévez, A., Crespo, S., Furones, M.D., 2003. New fish species foraquaculture, old pathological problems: the case of Solea sp. Proc. 11th EuropeanAssoc. Fish Pathologist, St Julians, Malta.
Zhang, Z.Y., Balmer, J.E., Lovlie, A., Fromm, S.H., Blomhoff, R., 1996. Specific teratogeniceffects of different retinoic acidi somers and analogs in the developing anteriorcentral nervous system of zebrafish. Dev. Dyn. 206, 73–86.
Zolfaghari, R., Ross, A.C., 2002. Lecithin:retinol acyltransferase expression is regulatedby dietary vitamin A and exogenous retinoic acid in the lung of adult rats. J. Nutr.132, 1160–1164.
48 I. Fernández, E. Gisbert / Aquaculture 315 (2011) 34–48




















