
The Molecular and Endocrine Basis of Flatfish Metamorphosis
17
Reviews in Fisheries Science, 16(S1):93–109, 2008 Copyright C Taylor and Francis Group, LLC ISSN: 1064-1262 print DOI: 10.1080/10641260802325377 The Molecular and Endocrine Basis of Flatfish Metamorphosis DEBORAH M. POWER, 1 INGIBJ ¨ ORG E. EINARSD ´ OTTIR, 2 KARIN PITTMAN, 3 GLEN E. SWEENEY, 4 JON HILDAHL, 2 MARCO A. CAMPINHO, 1 NADIA SILVA, 1 ØYSTEIN SÆLE, 3 MALYKA GALAY-BURGOS, 4 HEIDD ´ IS SM ´ ARAD ´ OTTIR, 5 and BJ ¨ ORN THRANDUR BJ ¨ ORNSSON 2 1 CCMAR, FERN, Universidade do Algarve, Campus de Gambelas, Faro, Portugal 2 Fish Endocrinology Laboratory, Department of Zoology/Zoophysiology, University of G¨ oteborg, G¨ oteborg, Sweden 3 Department of Biology, University of Bergen, High Technology Centre, Bergen, Norway 4 School of Biosciences, Cardiff University, Cardiff, United Kingdom 5 Fiskey ehf. Hjalteyri, Akureyri, Iceland A significant component of aquaculture is the production of good quality larvae, and, in the case of flatfish, this is tied up with the change from a symmetric larva to an asymmetric juvenile. Despite the pioneering work carried out on the metamorphosis of the Japanese flounder (Paralichthys olivaceus) and summer flounder (Paralichthys dentatus), the underlying molecular basis of flatfish metamorphosis is still relatively poorly characterized. It is a thyroid hormone (TH) driven process, and the role of other hormones in the regulation of the process along with the interplay of abiotic factors are still relatively poorly characterized as is the extent of tissue and organ remodeling, which underlie the profound structural and functional modifications that accompany the larval/juvenile transition. The isolation of genes for hormones, receptors, binding proteins, and other accessory factors has provided powerful tools with which to pursue this question. The application of molecular methodologies such as candidate gene approaches and microarray analysis coupled to functional genomics has started to contribute to understanding the complexity of tissue and organ modifications that accompany flatfish metamorphosis. A better understanding of the biology of normal metamorphosis is essential to identify factors contributing to abnormal metamorphosis. Keywords Atlantic halibut, endocrine control, hormones, metamorphosis, molecular tools, phenotype and genotype, thyroid hormones INTRODUCTION With the exception of metamorphosis, the process by which the symmetrical larvae becomes an asymmetric juvenile, the challenges and problems associated with flatfish (Division Teleostei, Order Pleuronectiformes) culture are common to the culture of round fish. Abnormal development during metamor- phosis is a frequent problem in aquaculture of flatfish (Gartner, 1986) and European species such as Atlantic halibut, Hippoglos- sus hippoglossus (Bolker and Hill 2000; Pittman et al., 1998; Solbakken et al., 2002), sole, Solea senegalensis (Dinis et al., Address correspondence to Deborah M. Power, CCMAR, FERN, Univer- sidade do Algarve, Campus de Gambelas, 8005-139 Faro, Portugal. E-mail: [email protected] 1999), and turbot, Scophthalmus maximus (Ellis et al., 1997) are no exception. The high incidence of abnormal juveniles limits the cost-effectiveness of their production and thus represents a serious industrial bottleneck. If the problems that cause abnor- mal metamorphosis can be alleviated, juvenile production rates can be significantly increased in existing facilities with only marginally higher running costs. FLATFISH AQUACULTURE AND GENETICS: A EUROPEAN PERSPECTIVE Production of Atlantic species of flatfish is dominated by turbot and Atlantic halibut with an annual yield of over 6,000 and 1,000 tons, respectively. Other flounders and sole amount to 93 Downloaded By: [Göteborg University Library] At: 10:49 16 September 2008
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of The Molecular and Endocrine Basis of Flatfish Metamorphosis

Reviews in Fisheries Science, 16(S1):93–109, 2008Copyright C©© Taylor and Francis Group, LLCISSN: 1064-1262 printDOI: 10.1080/10641260802325377
The Molecular and Endocrine Basisof Flatfish Metamorphosis
DEBORAH M. POWER,1 INGIBJORG E. EINARSDOTTIR,2 KARIN PITTMAN,3
GLEN E. SWEENEY,4 JON HILDAHL,2 MARCO A. CAMPINHO,1
NADIA SILVA,1 ØYSTEIN SÆLE,3 MALYKA GALAY-BURGOS,4
HEIDDIS SMARADOTTIR,5 and BJORN THRANDUR BJORNSSON2
1CCMAR, FERN, Universidade do Algarve, Campus de Gambelas, Faro, Portugal2Fish Endocrinology Laboratory, Department of Zoology/Zoophysiology, University of Goteborg, Goteborg, Sweden3Department of Biology, University of Bergen, High Technology Centre, Bergen, Norway4School of Biosciences, Cardiff University, Cardiff, United Kingdom5Fiskey ehf. Hjalteyri, Akureyri, Iceland
A significant component of aquaculture is the production of good quality larvae, and, in the case of flatfish, this is tied up withthe change from a symmetric larva to an asymmetric juvenile. Despite the pioneering work carried out on the metamorphosisof the Japanese flounder (Paralichthys olivaceus) and summer flounder (Paralichthys dentatus), the underlying molecularbasis of flatfish metamorphosis is still relatively poorly characterized. It is a thyroid hormone (TH) driven process, andthe role of other hormones in the regulation of the process along with the interplay of abiotic factors are still relativelypoorly characterized as is the extent of tissue and organ remodeling, which underlie the profound structural and functionalmodifications that accompany the larval/juvenile transition. The isolation of genes for hormones, receptors, binding proteins,and other accessory factors has provided powerful tools with which to pursue this question. The application of molecularmethodologies such as candidate gene approaches and microarray analysis coupled to functional genomics has startedto contribute to understanding the complexity of tissue and organ modifications that accompany flatfish metamorphosis.A better understanding of the biology of normal metamorphosis is essential to identify factors contributing to abnormalmetamorphosis.
Keywords Atlantic halibut, endocrine control, hormones, metamorphosis, molecular tools, phenotype and genotype,thyroid hormones
INTRODUCTION
With the exception of metamorphosis, the process by whichthe symmetrical larvae becomes an asymmetric juvenile, thechallenges and problems associated with flatfish (DivisionTeleostei, Order Pleuronectiformes) culture are common to theculture of round fish. Abnormal development during metamor-phosis is a frequent problem in aquaculture of flatfish (Gartner,1986) and European species such as Atlantic halibut, Hippoglos-sus hippoglossus (Bolker and Hill 2000; Pittman et al., 1998;Solbakken et al., 2002), sole, Solea senegalensis (Dinis et al.,
Address correspondence to Deborah M. Power, CCMAR, FERN, Univer-sidade do Algarve, Campus de Gambelas, 8005-139 Faro, Portugal. E-mail:[email protected]
1999), and turbot, Scophthalmus maximus (Ellis et al., 1997) areno exception. The high incidence of abnormal juveniles limitsthe cost-effectiveness of their production and thus represents aserious industrial bottleneck. If the problems that cause abnor-mal metamorphosis can be alleviated, juvenile production ratescan be significantly increased in existing facilities with onlymarginally higher running costs.
FLATFISH AQUACULTURE AND GENETICS:A EUROPEAN PERSPECTIVE
Production of Atlantic species of flatfish is dominated byturbot and Atlantic halibut with an annual yield of over 6,000and 1,000 tons, respectively. Other flounders and sole amount to
93
Downloaded By: [Göteborg University Library] At: 10:49 16 September 2008
94 D. M. POWER ET AL.
Ton
s x
1000
2001 2002 2003 2004 2005
AsiaEurope
2
4
6
8
1020406080120
140
Atla
ntic
hal
ibut
Lefte
ye fl
ound
er
Rig
htey
e flo
unde
r
Tot
ol A
sia
Turb
ot
Tota
l E
urop
e
Bas
tard
hal
ibut
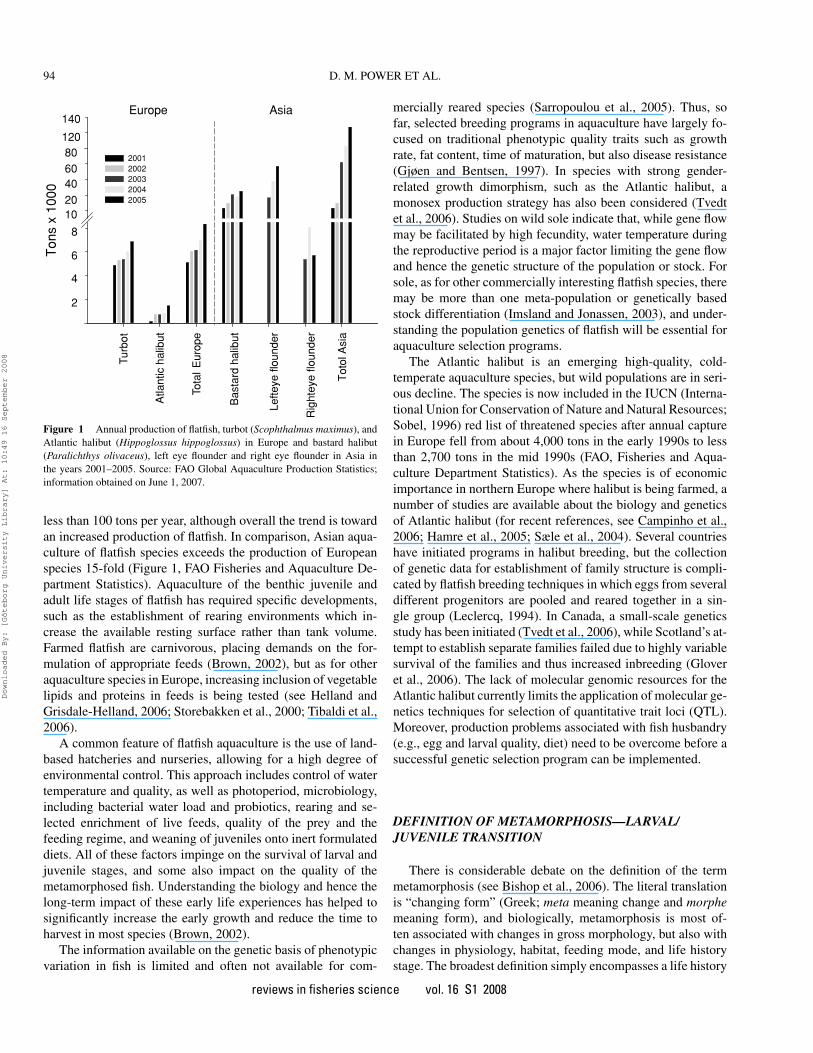
Figure 1 Annual production of flatfish, turbot (Scophthalmus maximus), andAtlantic halibut (Hippoglossus hippoglossus) in Europe and bastard halibut(Paralichthys olivaceus), left eye flounder and right eye flounder in Asia inthe years 2001–2005. Source: FAO Global Aquaculture Production Statistics;information obtained on June 1, 2007.
less than 100 tons per year, although overall the trend is towardan increased production of flatfish. In comparison, Asian aqua-culture of flatfish species exceeds the production of Europeanspecies 15-fold (Figure 1, FAO Fisheries and Aquaculture De-partment Statistics). Aquaculture of the benthic juvenile andadult life stages of flatfish has required specific developments,such as the establishment of rearing environments which in-crease the available resting surface rather than tank volume.Farmed flatfish are carnivorous, placing demands on the for-mulation of appropriate feeds (Brown, 2002), but as for otheraquaculture species in Europe, increasing inclusion of vegetablelipids and proteins in feeds is being tested (see Helland andGrisdale-Helland, 2006; Storebakken et al., 2000; Tibaldi et al.,2006).
A common feature of flatfish aquaculture is the use of land-based hatcheries and nurseries, allowing for a high degree ofenvironmental control. This approach includes control of watertemperature and quality, as well as photoperiod, microbiology,including bacterial water load and probiotics, rearing and se-lected enrichment of live feeds, quality of the prey and thefeeding regime, and weaning of juveniles onto inert formulateddiets. All of these factors impinge on the survival of larval andjuvenile stages, and some also impact on the quality of themetamorphosed fish. Understanding the biology and hence thelong-term impact of these early life experiences has helped tosignificantly increase the early growth and reduce the time toharvest in most species (Brown, 2002).
The information available on the genetic basis of phenotypicvariation in fish is limited and often not available for com-
mercially reared species (Sarropoulou et al., 2005). Thus, sofar, selected breeding programs in aquaculture have largely fo-cused on traditional phenotypic quality traits such as growthrate, fat content, time of maturation, but also disease resistance(Gjøen and Bentsen, 1997). In species with strong gender-related growth dimorphism, such as the Atlantic halibut, amonosex production strategy has also been considered (Tvedtet al., 2006). Studies on wild sole indicate that, while gene flowmay be facilitated by high fecundity, water temperature duringthe reproductive period is a major factor limiting the gene flowand hence the genetic structure of the population or stock. Forsole, as for other commercially interesting flatfish species, theremay be more than one meta-population or genetically basedstock differentiation (Imsland and Jonassen, 2003), and under-standing the population genetics of flatfish will be essential foraquaculture selection programs.
The Atlantic halibut is an emerging high-quality, cold-temperate aquaculture species, but wild populations are in seri-ous decline. The species is now included in the IUCN (Interna-tional Union for Conservation of Nature and Natural Resources;Sobel, 1996) red list of threatened species after annual capturein Europe fell from about 4,000 tons in the early 1990s to lessthan 2,700 tons in the mid 1990s (FAO, Fisheries and Aqua-culture Department Statistics). As the species is of economicimportance in northern Europe where halibut is being farmed, anumber of studies are available about the biology and geneticsof Atlantic halibut (for recent references, see Campinho et al.,2006; Hamre et al., 2005; Sæle et al., 2004). Several countrieshave initiated programs in halibut breeding, but the collectionof genetic data for establishment of family structure is compli-cated by flatfish breeding techniques in which eggs from severaldifferent progenitors are pooled and reared together in a sin-gle group (Leclercq, 1994). In Canada, a small-scale geneticsstudy has been initiated (Tvedt et al., 2006), while Scotland’s at-tempt to establish separate families failed due to highly variablesurvival of the families and thus increased inbreeding (Gloveret al., 2006). The lack of molecular genomic resources for theAtlantic halibut currently limits the application of molecular ge-netics techniques for selection of quantitative trait loci (QTL).Moreover, production problems associated with fish husbandry(e.g., egg and larval quality, diet) need to be overcome before asuccessful genetic selection program can be implemented.
DEFINITION OF METAMORPHOSIS—LARVAL/JUVENILE TRANSITION
There is considerable debate on the definition of the termmetamorphosis (see Bishop et al., 2006). The literal translationis “changing form” (Greek; meta meaning change and morphemeaning form), and biologically, metamorphosis is most of-ten associated with changes in gross morphology, but also withchanges in physiology, habitat, feeding mode, and life historystage. The broadest definition simply encompasses a life history
reviews in fisheries science vol. 16 S1 2008
Downloaded By: [Göteborg University Library] At: 10:49 16 September 2008

THE MOLECULAR AND ENDOCRINE BASIS OF FLATFISH METAMORPHOSIS 95
transition between a vegetative and sexually reproductive stageand is synonymous with morphogenesis, and includes all mul-ticellular organisms (e.g., fungi, plants and animals).
Most commonly, however, the term metamorphosis is usedto refer to the major morphological, physiological, and eco-logical changes in holometabolous insects, marine crustaceans,amphibians, and certain fish species, most notably flatfish, eels,and lampreys (Youson, 2004). A distinguishing feature in thesegroups is the degree of morphological change, and that larvaland juvenile forms are distinct. Metamorphic morphogenesis isrecognized to be hormonally driven, but there is some debateconcerning the timing and rate of metamorphosis, and thereforewhat changes are actually hormonally regulated (Heyland et al.,2006; Hodin 2006).
Development of fish from a fertilized egg to a larva and thena juvenile is a continuous process, which generally leads to achange in form and function and is characterized by phases ofrapid and slower change (see Balon 1999; Blaxter 1988). Con-siderable debate surrounds the notion of what constitutes meta-morphosis in fish, and it is frequently associated with functionalchanges such as those occurring during the larval to juveniletransition, although the degree of change may be variable. Hodin(2006) suggests that the defining aspect of metamorphosis is“discontinuous” change in morphology. Youson (1988, 2004)goes further, stating that all fish with a larval period experiencemetamorphosis to become a juvenile in contrast to direct devel-opment where the post-embryonic fish resembles the juvenile oradult form, but is smaller and sexually immature. In the preced-ing definition, metamorphosis is considered a phase in the larvalperiod, and it is sub-divided into pre-metamorphosis, metamor-phosis, climax, and post-metamorphosis and is not consideredto be a common developmental strategy. This distinction elim-inates in fish some transitory forms that do not experience apronounced climax event and excludes some juvenile transi-tions such as the salmon parr-smolt transition, which involves amajor change in habitat and inevitably a number of functionalmodifications.
The present review will focus on the dramatic remodeling ofthe pleuronectiform (flatfish) larva. Metamorphosis in flatfish ishighly sensitive to intrinsic and extrinsic signals, leading to atightly regulated climax event required for successful migrationof one eye so that both eyes are on the same side of the head.There is concomitant pigmentation of the ocular side of the fishand the transition from a pelagic larva to a benthic juvenile.The sidedness of flatfish is species specific (Policansky, 1982).Thus, for Atlantic halibut, the left eye migrates to the right side,whereas for the Japanese flounder (Paralichthys olivaceus, here-after referred to as the Japanese founder), the right eye migratesto the left side. Associated with the obvious changes in externalmorphology during metamorphosis, larvae also undergo otherdramatic changes that drive the maturation of organs and alsoshape the morphology to that of the adult animal.
The abnormalities reported in flatfish culture are surprisinglycommon and generally become evident at metamorphosis. Themost common abnormalities include lack of eye migration, bone
deformities, and pigment abnormalities. The main difficulty inidentifying causative factors of these abnormalities is the lackof information on the developmental and physiological changesthat underlie “normal metamorphosis” in fish. This scarcity ofdata is due to several factors, but, unquestionably, the absenceof molecular tools for important aquaculture species and dif-ficulties in obtaining standardized samples are of major sig-nificance. Moreover, as metamorphosis is a cascade of eventswith tissue-specific modifications and timings, the small size oflarvae represents a significant difficulty in studying this process.
Early development of Atlantic halibut is better correlatedwith size than with age (Sæle et al., 2004), although it still re-mains to be established if this is true for other flatfish. In a tankcontaining larvae derived from a single egg batch and thus ofthe same age, some individuals will go through metamorphicclimax and settling earlier than others. Nonetheless, samplingschemes are often based on the day of hatching, as opposed tomorphological criteria, complicating comparisons between andwithin experiments. It is, therefore, important to establish well-defined developmental staging schemes upon which samplingseries can be based. Complex analysis of Atlantic halibut larvae,from first feeding through to settlement, has enabled a robustdefinition of stages (Figure 2). The stages are defined using ex-ternal morphology, skeletal development, degree of ossification,and asymmetry of cranial bones. In turn, these are well corre-lated with myotome height (Sæle et al., 2004), which is readilydetermined and has proven efficient for sample standardizationas individuals can be rapidly separated into the principal devel-opmental stages: pre-metamorphosis, metamorphosis, climax,and post-metamorphosis (Figure 2).
First feeding through metamorphosis to settlement in Atlantichalibut has been defined as stages 5–10 (Saele et al., 2004), whilethe preceding stages are classified as stages 1–4 (Pittman et al.,1990). As the present review is focused on metamorphosis andnot early development, a consideration of morphology linkedto stage will only be given for stages 5–10 (Figure 2). More-over, as a detailed description of the staging scheme and itsvalidation is given in Saele et al. (2004), the present consid-eration will be brief. Atlantic halibut larvae of stages 5–7 arebilaterally symmetrical, transparent, but with pigmented eyes.There is an increase in body pigmentation and depth from stage5–7, and in the latter stage, the fins become defined and cra-nial ossification is notable. In normal Atlantic halibut larvae,stage 8 marks the start of the transition to asymmetry, which ismost evident in the frontal bone of the cranium, which starts tocurve in the direction of the future ocular side. At this stage,the larvae are relatively large but remain pelagic and the finsare all evident. This is the first stage at which larvae with anabnormal developmental trajectory, which may lead to failedmetamorphosis, become evident. In Atlantic halibut undergoingnormal metamorphosis, by stage 9 eye migration is advanced,and it reaches the midline giving the larvae an asymmetric ap-pearance, the body depth is greatly increased, and a distinctiveskin pigment pattern emerges, and the larvae rest occasionallyon the substrate. The last stage, 10, corresponds to the juvenile,
reviews in fisheries science vol. 16 S1 2008
Downloaded By: [Göteborg University Library] At: 10:49 16 September 2008

96 D. M. POWER ET AL.
Figure 2 Schematic drawing of the key developmental stages of the marineflatfish, Atlantic halibut (Hippoglossus hippoglossus). The larval period in fishis characterized by the presence of embryonic organs such as the yolk sac,the development of transitional structures such as the finfold, and by a thinthread-like body shape. Five main stages (5–10) of Atlantic halibut develop-ment through metamorphosis were identified. Stage 5 is first-feeding larvae,bilaterally symmetrical, and transparent except for the eyes, which are fully pig-mented and there is a primordial finfold; stage 6 is larvae that resembles stage 5but with increased body depth; in stage 7 larvae, the body depth increases in thebilaterally symmetrical larvae, and the pelvic fins develop and the primordialfin is in transition to the dorsal, ventral, and caudal fins; stage 8 larvae haveinitiated metamorphosis, the left eye starts to migrate, the body depth increases,and pigmentation become evident on the right side; stage 9 corresponds to theclimax of metamorphosis, eye migration is advanced, and the body is clearlyasymmetrical and pigmentation is intense and covers most of the right side ofthe body; stage 10 corresponds to the juvenile, the right side is pigmented andthe left side is white. The scale bar corresponds to 1 cm.
in which eye migration is complete and both eyes are on the oc-ular side of the fish, pigmentation is complete, and the craniumis a similar shape to that of the adult. The juveniles are dem-ersal and are settled on the substrate the majority of the time.The staging scheme developed for Atlantic halibut representeda fundamental step in studying specific ontogeny of organ andtissue systems, and the underlying molecular changes duringkey developmental stages in metamorphosis.
ENDOCRINE REGULATION OF FLATFISHMETAMORPHOSIS
Gross changes in external morphology occurring during hal-ibut metamorphosis (Moren et al., 2004, 2006; Sæle et al.,2006a, 2006b; Solbakken et al., 1999, 2002) are underlain bymolecular and biochemical processes that are directly or indi-rectly regulated by endocrine factors.
Thyroid Hormones
Thyroid hormones (THs), both thyroxine (T4) and tri-iodothyronine (T3), appear to be the most important endocrine
regulators of metamorphosis in flatfish and other teleost fish(Campinho et al., 2006, 2007b; de Jesus et al., 1990, 1998; Inuiand Miwa, 1985; Leatherland, 1994; Llewellyn et al., 1998,1999; Miwa and Inui, 1987a, 1987b, 1991; Miwa et al., 1988,1992; Power et al., 2001; Schreiber and Specker, 1998, 1999,2000; Soffientino and Specker, 2003; Szisch et al., 2005; Ya-mano, 2005; Yamano and Miwa, 1998; Yamano et al., 1991a,1991b, 1994b). The role of THs in fish metamorphosis has beenstudied for decades (Eales, 1979), and their effect in driving flat-fish metamorphosis is undisputable (de Jesus et al., 1991; Inuiand Miwa, 1985; Miwa et al., 1988; Tanaka et al., 1995). THlevels peak at metamorphic climax during the larvae-to-juveniletransition in the well-studied flatfish models, the Japanese floun-der (Inui and Miwa, 1985; Miwa et al., 1988; Yamano, 2005),and the summer flounder (Schreiber and Specker, 1998), andalso during metamorphosis of Atlantic halibut (Einarsdottiret al., 2006).
Thyroid Hormone Receptors
THs are presumed to mediate their effects by interactingwith thyroid hormone receptors (TRs), members of the nuclearreceptor superfamily, which positively and negatively regulateTH-target gene expression through binding to specific thyroidresponse elements (TREs) (Oetting and Yen, 2007; Zhang andLazar, 2000). TRs have been cloned in a number of differentflatfish, and two key isoforms, encoded by different genes, havebeen identified: TRα and TRβ; in some species, additional genesencoding further isoforms are present, e.g., TRαA and TRαB(Galay-Burgos et al., 2008; Llewellyn et al., 1999; Power et al.,2001; Yamano et al., 1994a; Yamano and Inui, 1995; Yamanoand Miwa, 1998). Relatively little is known about the precisemolecular mechanisms and pathways fish TRs influence. Thisis due in part to a lack of information about TH target genesand the expression of deiodinases during teleost development(Thisse et al., 2003).
Studies of receptor tissue distribution during fish metamor-phosis are infrequent, partly because of the difficulty in cloningTRs and also their generally low tissue abundance. In the At-lantic halibut, both TRα and TRβ expression tends to peakat metamorphosis (stage 9) and declines thereafter, althoughthe increase is only significant for TRβ (Galay-Burgos et al.,2008). The TR expression profiles described in turbot (Scophtal-mus maximus) and Japanese flounder are approximately similarto that of the Atlantic halibut, with a peak of TRβ and TRα
around the metamorphic climax, although the exact pattern ofTR expression prior to, during, and after metamorphosis ap-pears to be species specific (Marchand et al., 2004; Yamanoand Miwa, 1998). The differential expression of TRα and TRβ
during flatfish metamorphosis may explain how THs can haveapparently opposing effects in the tissue maturation that accom-panies metamorphosis. It will be of importance in the future toestablish sensitive methods for TR tissue localization in order toestablish timing of expression and the isoforms that are involved
reviews in fisheries science vol. 16 S1 2008
Downloaded By: [Göteborg University Library] At: 10:49 16 September 2008

THE MOLECULAR AND ENDOCRINE BASIS OF FLATFISH METAMORPHOSIS 97
in specific TH-driven tissue maturation during metamorphosis(e.g., muscle, blood, skin, intestine, fin formation).
Deiodinases
The conversion of T4 into the biologically active T3 and sub-sequent inactivation of these THs occurs through the action of afamily of selenocysteine protein enzymes known as deiodinases(Bianco et al., 2002; Kohrle, 2000). Deiodination reactions oc-cur by outer ring deiodination (ORD) in the 5′-position of thephenolic ring or by inner ring deiodination (IRD) of the 5-position on the tyrosyl ring of T4. ORD of T4 generates T3 inperipheral tissues, and IRD of T4 yields the inactive metabolitereverse T3(rT3). Metabolism of T3 by IRD and rT3 by ORD re-sults in the metabolite T2 (Bianco et al., 2002; Kohrle, 2000). Infish, even though three deiodinases have been cloned and adulttissue distribution characterized, no studies exist of these keyenzymes at any early life stage, apart from the limited analysis ofzebrafish (Brachydanio rerio) D1 and D2 (Thisse et al., 2003).In flatfish, the lack of information on deiodinases is striking.During Atlantic halibut metamorphosis, when T3 levels rise,deiodinase D3 expression decreases and D2 and TR (especiallyTRβ) expression increases concomitantly and peaks at climaxwhen major organ and tissue transformations occur (Campinhoet al., 2005b; Galay-Burgos et al., 2008). Clearly, a better un-derstanding of TR and the deiodinases in flatfish will be crucialto understand the molecular mechanisms THs regulate duringmetamorphosis.
The morphological changes during metamorphosis arematched by a number of internal changes in tissues and organs,which prepare the developing fish for its changing nutritional,metabolic, locomotive, and environmental demands. Althoughthere are a number of studies documenting the tissue-specificmorphological changes that occur during metamorphosis offlatfish, relatively little molecular information is availableabout thyroid responsive genes and the genetic regulatorynetworks that determine the larval to juvenile (metamorphic)development.
TH Effect on Stomach
A surge in TH levels is correlated with stomach developmentin Japanese flounder and summer flounder (Huang et al., 1998;Miwa et al., 1992). In these flatfish, the gastric gland startsdeveloping during pre-metamorphosis, although pepsinogen-like immunoreactivity is detected only after metamorphosis.Furthermore, TH treatment results in accelerated gut develop-ment and the presence of pepsinogen-like immunoreactivity,whereas thiourea delays stomach development and prevents anypepsinogen-like immunoreactivity in the organ (Miwa et al.,1992). In Atlantic halibut, gastric gland development is alsoapparent prior to metamorphosis and pepsin mRNA is ex-pressed in low abundance in the developing gastric gland. Inpost-metamorphic halibut, gastric glands are numerous and well
developed, and there is a highly significant increase in pepsintranscript expression (Power et al., unpublished observations).That the gastric glands first appear prior to the peak in THs,which occurs at stage 9 Atlantic halibut (metamorphic climax),suggests the role of THs on the developing gastric glands of thisspecies may be more complex than that of Japanese and summerflounder.
TH Effects on Muscle Development
Muscle development and growth are of key interest in aqua-culture. Although axial musculature in teleosts appears mor-phologically far simpler than in terrestrial vertebrates, at themolecular level it is complex. Metamorphosis in flatfish is ac-companied by changes in isoform expression of several musclegenes. During Japanese flounder metamorphosis, troponin T(TnT) and DTNB (5,5′-dithio-bis-nitrobenzonic acid) myosinlight chain protein isoform expression changes (Yamano et al.,1991a, 1994b). Moreover, both thiourea and TH treatmentseverely affect troponin T and DTNB myosin light chain proteinisoform expression, emphasizing the role of THs in this process(Yamano et al., 1994a, b).
In Atlantic halibut, THs also influence muscle gene ex-pression during metamorphosis, and there is a change in theexpression of fast TnT gene expression, which is accompa-nied by an alteration in their spatial distribution as musclemorphology changes. Interestingly, despite the change from asymmetrical larva to an asymmetrical juvenile, the distribu-tion of both slow and fast transcripts of TnT in muscle re-mains symmetrical in relation to the left-right axis (Campinhoet al., 2007a). THs also appear to regulate TnT expression dur-ing the larval-juvenile transition in roundfish (Campinho et al.,2006), although the regulation appears to differ among teleosts,and this probably relates to differing functional requirements(Campinho et al., 2005a, 2005c, 2006; Hsiao et al., 2003).
THs Effects on Blood
A further important change that occurs during metamor-phosis is associated with the oxygen-carrying capacity ofhemoglobin in red blood cells. Hemoglobin is a tetramer, whichin almost all vertebrates consists of two identical α-globin sub-units and two identical β-globin subunits (Hardison 1998). Inorder to cope with the different availability of, and requirementsfor, oxygen during development, different hemoglobin tetramersare produced and erythrocyte morphology changes.
In the Atlantic halibut, a shift from a larval to an adult pat-tern of globin isoforms accompanies metamorphosis. However,it has not been possible to identify changes in erythrocyte mor-phology, as circulating erythrocytes are infrequent until devel-opmental stage 9 (metamorphic climax) (Pittman, unpublishedobservations). Expression of larval globin genes becomes de-tectable at stage 9, and transformation to the adult pattern of gene
reviews in fisheries science vol. 16 S1 2008
Downloaded By: [Göteborg University Library] At: 10:49 16 September 2008

98 D. M. POWER ET AL.
expression occurs at an undetermined time after completion ofmetamorphosis (Sweeney, unpublished observations). In con-trast, in pre-metamorphic Japanese flounder larvae, large rounderythrocytes with small round nuclei exist as larva enter meta-morphosis. This cell population is then substituted by ellipticalerythrocytes so that, at the end of metamorphosis, only adult-type erythrocytes are detected. TH treatment can induce thismorphological change, suggesting that THs may suppress larvalerythropoiesis and stimulate adult erythropoiesis; no informa-tion about globin gene expression exists (Miwa and Inui, 1991).
The above examples demonstrate the importance of THs inthe maturation of only a few of the many tissue systems influ-enced by these hormones and which are modified during meta-morphosis. However, although a number of studies describingtissue-specific maturation and some of the genes involved dur-ing flatfish metamorphosis exist, data clearly demonstrating therole of different elements of the thyroid axis are still scarce. Therelative scarcity of information about TRs, deiodinases, andresponsive genes is probably related to their low tissue abun-dance, precise temporal expression, and tissue-specific distribu-tion. These problems are further compounded by the small sizeof the larvae being studied. Moreover, the complex interactionsbetween different elements of the thyroid axis make determin-ing changes and understanding their biological significance amajor challenge.
Relatively few studies exist about the role of other hor-mones during fish metamorphosis even though in a number ofteleosts endocrine glands appear relatively early in development.For example, the pituitary gland, ultimobranchials, endocrinepancreas, interrenals (adrenocorticol homologue), Stannius cor-puscles and pineal, are already well developed in larvae andprobably interact directly or indirectly with the TH axis to reg-ulate different aspects of metamorphosis. In anurans, severalhormones, including pituitary hormones, have been shown toinfluence metamorphosis (Denver et al., 2002), and it remainsto be established if this is also the case in teleosts.
Pituitary Morphology/Histology
In Atlantic halibut, as in other vertebrates, the pituitary issituated ventrally and close to the hypothalamus (Einarsdottiret al., 2006; Weltzien et al., 2003) and consists of a neurohy-pophysis (NH), which is dorsal to and interdigitates with theadenohypophysis (AH), as reported in other teleost fish (Balland Baker, 1969; Norris, 1997; Yadav, 1995). At the end of theyolk sack stage, the pituitary is already present, and the NH is adistinct structure at the start of first feeding (Einarsdottir et al.,2006). The AH is divided into a well-distinguishable rostral parsdistalis, proximal pars distalis, and pars intermedia (Figure 3a).
Growth hormone (GH) producing somatotrophs, prolactin(PRL) producing lactotrophs, TSH-producing thyrotrophs(Figure 3; Power et al., unpublished results), somatolactin(SL) producing somatolatotrophs (Einarsdottir et al., 2006), aswell as adenocorticotropic hormone (ACTH)- and melanocyte-stimulating hormone (MSH)-producing cells are first observed
Figure 3 Sagittal sections of Atlantic halibut (Hippoglossus hippoglossus)larvae at first feeding (stage 5, a and b) and metamorphosis (stage 8, c and d).Sections have been stained with hematoxylin and eosin (a and c) or subject to insitu hybridization using a digoxygenin-labeled riboprobe specific for TSHβ (band d). The outline of the pituitary is highlighted to facilitate its identification,and the rostral pars distalis (RPD), proximal pars distalis (PPD), and neurohy-pophysis (NH) are indicated in the sections stained with hematoxylin and eosin(a and c). In situ TSHβ digoxygenin-labeled riboprobe. A few weakly labeledthyrotrophs (arrow) are evident in stage 5 halibut (b) in the PPD, although bystage 8 the thyrotrophs in the PPD are more numerous and contain abundantmRNA (d). The scale bar corresponds to 20 µm.
using both in situ hybridization and immunohistochemisty(IHC) around the time of yolk sack absorption (Power et al.,unpublished results). As development progresses through tometamorphosis, there is a concomitant increase in the relativenumbers of all endocrine cells, suggesting the hormones theyproduce may play a role in early developmental stages of At-lantic halibut but also during metamorphosis, although their ex-act function remains to be established (Einarsdottir et al., 2006).In order for hormones to function, they must interact withspecific receptors, the presence or absence of which will de-termine tissue responsiveness. Clearly, the identification ofhormone receptors and their localization is an important stepin identifying potential sites of action and function duringdevelopment.
Pituitary Hormone Actions in Larval Development
The GH-IGF-I System
GH is the principal growth-promoting hormone in teleostfish, stimulating the growth of cartilage, bone, and muscle, andregulating food conversion and appetite. GH also plays an im-portant role in osmoregulation, reproduction, and immune func-tion in teleosts (reviewed in Bjornsson, 1997; Bjornsson et al.,2002; Perez-Sanchez, 2000; Peter and Marchant, 1995). GHmay act directly on target tissue or by regulating the hepatic
reviews in fisheries science vol. 16 S1 2008
Downloaded By: [Göteborg University Library] At: 10:49 16 September 2008

THE MOLECULAR AND ENDOCRINE BASIS OF FLATFISH METAMORPHOSIS 99
production of insulin-like growth factor I (IGF-I), which stim-ulates tissue and skeletal growth (see Bjornsson et al., 2002;Kopchick and Andry, 2000). GH receptor (GHR) mRNA is ex-pressed in most fish tissues and organs (Calduch-Giner et al.,2001; Fukada et al., 2004; Kajimura et al., 2004; Lee et al.,2001; Nakao et al., 2004; Tse et al., 2003; Very et al., 2005),further underlining the pluripotency of GH.
Although GH is present in the pituitary gland of flatfish larvaeand juveniles (Einarsdottir et al., 2006, 2007), their small sizehas so far precluded any studies of GH secretion or plasma GHlevels during larval stages. To elucidate the role of GH in larvaldevelopment and metamorphosis, GHR, as well as IGF-I and itsreceptor (IGF-IR), need to be considered.
Nucleotide sequences have been generated for multipleforms of the Atlantic halibut GHR and IGF-IR. In common withseveral other teleost species, Atlantic halibut has two full-lengthGHR (type I and type II hhGHRfl) and two IGF-IR isoforms(hhIGF-IR1 and hhIGF-IR2; Hildahl et al., 2007a, 2007b).Additionally, Atlantic halibut has two forms of a truncated GHR(hhGHRtr1 and hhGHRtr2; Hildahl et al., 2007b), one similarto GHRtr in Japanese flounder and turbot and an alternativelyspliced form with a 132-bp extracellular insert correspondingto intron six in Japanese flounder (Calduch-Giner et al., 2001;Nakao et al., 2004).
Gene cloning and sequencing has paved the way for molec-ular analysis of the GH-IGF-I system during Atlantic halibutmetamorphosis (Hildahl et al., 2007a, 2007b). In situ hybridiza-tion and immunohistochemistry show ubiquitous expression ofGHR mRNA and protein from early pre-metamorphosis (stage5) through to post-metamorphosis (stage 10; Hildahl et al.,2008), likely reflecting its multi-functional role in tissue growthand development. The way in which the GH-IGFI pathway is in-volved in metamorphosis clearly merits further attention as bothGHR and IGFI transcripts and protein are associated with tissuesof the cranium, which undergo remodeling during metamorpho-sis (Hildahl et al., 2008). Type I hhGHRfl, hhGHRtr1, hhIGF-IR1, and hhIGF-IR2 are differentially expressed during meta-morphosis with all but IGF-IR1 decreasing during metamorphicclimax and post-metamorphosis. This general decrease in recep-tor gene expression suggests decreased tissue sensitivity to GHand IGF-I during metamorphosis, and may explain decreasedgrowth rates during metamorphosis (Einarsdottir et al., 2007).
Individuals undergoing abnormal metamorphosis have de-creased GHR transcript levels as well as decreased IGF-I tissuecontent (Hildahl et al., 2007a, 2007b). The reduced receptorgene expression in abnormal individuals further supports thenotion that that expression of these genes could be importantfor metamorphic success. The specific function of the GH andIGF-I system during metamorphosis and if its role is direct orindirect remains to be clarified.
Other Pituitary Hormone Systems
The presence of TSH in halibut larval thyrotrophs is indicatedby results from immunohistochemistry and in situ hybridization,
using a sea bream-specific TSHβ riboprobe. The thyrotrophs,localized in the RPD of the adenohypophysis (Einarsdottir et al.,2006; Power et al., unpublished results), are scarce early aftercommencement of first feeding, but became more abundant priorto and at the climax of metamorphosis (Figure 3). The distinctincrease of thyrotrophs before and at metamorphic climax isconcurrent with an increase in whole body T4 levels (Camp-inho et al., 2007b). After the metamorphic climax, the TSHβinsitu hybridization signal intensity decreases (Power et al., un-published results), although whole body T4 levels remain high(Campinho et al., 2007b). The reason for the apparent discrep-ancy in thyrotroph occurrence and tissue content of T4 after themetamorphic climax remains to be established but may reflectthe maturation of the TH axis to meet the modified physiolog-ical requirements of the juvenile/adult stage and may be theresults of a range of different factors, including the change indynamics of hormone turnover (e.g., reduced tissue deiodinaseactivity), changes in the abundance of thyroid hormone bind-ing proteins, modifications in feeding activity and growth, andthe increased muscle mass and associated increase in the largereservoir of THs in peripheral tissue. The physiological conse-quences of high T4 levels after the metamorphic climax remainto be studied.
In fish, adenocorticotropic hormone (ACTH) released fromthe pituitary regulates cortisol secretion from interrenal tissue.Cortisol levels are elevated in response to stressors, and cor-tisol has catabolic effects, affects hydromineral balance, andreduces immune competence in fish (Wendelaar Bonga, 1997).The POMC gene, which codes for a common precursor of MSH,ACTH, and corticotrophs, are present in the pituitary at all de-velopmental stages of halibut larvae (stages 5–10). Moreover,cortisol is detectable in halibut tissue extracts from stage 5 lar-vae onwards, which indicates that the hypothalamic-pituitary-interrenal axis is active early in development (Power, unpub-lished observations). High cortisol levels occur at metamorphicclimax of the Japanese flounder, and cortisol appears to en-hance some TH actions (de Jesus et al., 1990, 1991, 1993),although there are conflicting opinions about whether it hasa permissive or inhibitory role during metamorphosis (Tanakaet al., 1995). Recent studies in the summer flounder indicatethe hypothalamus-pituitary-interrenal (HPI) axis is active be-fore the onset of metamorphosis, and that cortisol is necessaryfor larval seawater tolerance (Veillette et al., 2007a, 2007b).The detection of a functional HPI axis prior to metamorphosisin the Summer flounder raises a number of intriguing possibil-ities about its interaction with the TH axis before, during, andafter metamorphosis.
PRL is a versatile hormone, influencing growth, develop-ment and metabolism, stimulation of endocrine glands, behav-ior, reproduction, and immune function in different vertebrateclasses (Bole-Feysot et al., 1998; Forsyth and Wallis, 2002;Power, 2005). In freshwater and euryhaline teleosts, includingsome flatfishes, a major function appears to be osmoregulation(Gutt, 1985; McCormick, 2001; Wada et al., 2004). In am-phibians, there are conflicting opinions about the role of PRL in
reviews in fisheries science vol. 16 S1 2008
Downloaded By: [Göteborg University Library] At: 10:49 16 September 2008

100 D. M. POWER ET AL.
metamorphosis; it has been considered to be a juvenile hormone.However, transgenic Xenopus laevis over-expressing PRL andnon-transgenic siblings reach metamorphic climax at the sametime, although tail resorption fails to occur in the former animals(Huang and Brown, 2000). There is little evidence so far aboutthe role of PRL in fish metamorphosis, although the presence ofPRL in the pituitary gland of stage 5 halibut larvae and its in-crease in abundance in subsequent stages raises questions aboutits possible function during flatfish metamorphosis (Einarsdottiret al., 2006).
SL, only found in teleost fish, was first discovered in theflounder pituitary (Ono et al., 1990), and it appears to have di-verse actions (Company et al., 2001; Fukamachi et al., 2004,2006; Kakizawa et al., 1993, 1995, 1996, 1997; Nguyen et al.,2006; Olivereau and Rand-Weaver, 1994; Planas et al., 1992;Rand-Weaver et al., 1992, 1993; Rand-Weaver and Swanson,1993; Renfro, 1997; Zhu and Thomas, 1997), including reg-ulation of adiposity in several fish species (Company et al.,2001) and skin pigmentation in red drum (Sciaenops ocellatus)(Zhu and Thomas, 1997), zebrafish (Nguyen et al., 2006), andmedaka. It remains to be established if SL has a role in theasymmetric pigmentation of halibut juveniles and regulation offat reserves in metamorphosing halibut.
Other Endocrine Hormone Systems
As mentioned previously a number of endocrine glands, suchas the Stannius corpuscles, ultimobranchial gland, and pancreasappear early in larval development. However, there is a dearth ofstudies about their development during metamorphosis and therole of their endocrine secretions in this process. It seems likely,however, taking into consideration the physiological modifica-tions that accompany metamorphosis and the associated reduc-tion in feeding in some flatfish species, that other endocrineaxes, particularly those involved in digestion, energy balance,and ion regulation will be important. However, it remains to beestablished how other endocrine systems are modified duringmetamorphosis and if this is a direct effect of THs or a conse-quence of passage to a new developmental status, e.g., juvenile.For example, the development of the endocrine pancreas is in-timately linked to the development of the endocrine cells ofthe stomach and intestine (Youson and Al-Mahrouki, 1999). Inthe Xenopus, in which a number of studies of metamorphosis-related changes in the gastrointestinal (GI) tract exist (Prettyet al., 1995; Schreiber et al., 2005; Torihashi, 1990), no stud-ies of metamorphosis-associated changes in the pancreas andenteric endocrine cells exist. In fish, there are few studies ofmetamorphosis-associated maturation of the GI tract. There isclearly considerable scope for future studies directed at explor-ing how the enteropancreatic endocrine system is modified orinvolved in metamorphosis-associated modifications of the GItract.
ABNORMAL METAMORPHOSIS
Abnormal metamorphosis may involve one or more tissuesystems, be of greater or lesser severity, and may be visible ex-ternally as incomplete eye migration, deformity of bones, andabnormal pigmentation (Figure 4). Associated with modified ex-ternal morphology, there may be delayed maturation of a numberof systems, such as the skeleton, muscle, and circulatory systems(authors’ observations). Abnormal pigmentation in flatfish cov-ers a broad range of different conditions ranging from albinism(on the ocular side), ambicoloration (pigmentation on ocularand abocular sides), and mosaicism (for references, see Bolkerand Hill, 2000; Tagawa and Aritaki, 2005). Such abnormalitiesare commercially important, although their relative impact maydepend on the way in which flatfish are commercialized, e.g.,intact fish or fillets. One characteristic skeletal abnormality inflatfish is the failure of the eye to migrate (Okada et al., 2001),although spinal abnormalities are also frequent, and both defor-mities are commercially important as they influence growth anddevelopment and are often associated with reduced immunity.
Eye migration can be arrested at various “steps” in the process(Okada et al., 2001, 2003; Sæle et al., 2003, 2006b). The asym-metry of the Atlantic halibut head is generally restricted to theregion around the eyes, even after metamorphosis is completed,and so does not involve the entire cranium or neurocranium.In Atlantic halibut, the first “migration-related asymmetry” ofcranial bone structures is the lateral part of the future blind sideof the ethmoid cartilage, where a sub-dermal condensation offibroblasts begins to migrate in the caudal direction, ventral tothe eye (about stage 7; Sæle et al., 2006a, 2006b). Abnormal hal-ibut at stage 8 exhibit abnormal remodeling of the frontals withbilaterally equal activity of the osteoclasts. In abnormal stage 9fish (climax metamorphosis), the sinistral eye has not migrated,the frontal bones are symmetrical, and there is equal lateral os-teoclastic activity in the frontal processes (Figure 5). There is asignificant difference in osteoclast activity in remodeling boneof abnormal and normal halibut larvae at stage 9, with remod-eling normally more active on the frontal near the migratingeye.
In juvenile Atlantic halibut lacking eye migration (arrestedmetamorphosis), the dermal bone, also called the prefrontal,does not develop. The two abnormal paraethmoids developsymmetrically as two plate-like structures curving anteriorly,whereas normal elongate fused paraethmoids curve at their pos-terior (Sæle et al., 2006b). The abocular side retrorbital vesiclesare largest in volume only after the completion of normal eyemigration (Sæle et al., 2006b).
Pigmentation in fish is, in general, dependent on bilater-ally symmetric embryonic chromatoblasts, which either dif-ferentiate into larval pigment cells or remain undifferentiated.A number of other factors, such as environment, genetics,nutrition, and development of appropriate nervous innerva-tion, are also important in pigmentation (Dickey–Collas, 1993;Gronas et al., 1993; Houde, 1971; Tabata, 1991), but so farmost studies have focused on chromatoblast development.
reviews in fisheries science vol. 16 S1 2008
Downloaded By: [Göteborg University Library] At: 10:49 16 September 2008

THE MOLECULAR AND ENDOCRINE BASIS OF FLATFISH METAMORPHOSIS 101
Figure 4 Some characteristic metamorphosis associated abnormalities encountered in Atlantic halibut (Hippoglossus hippoglossus). (a) Normal stage 10 halibutwith both eyes on the upper pigmented side of the body; (b) An abnormal stage 10 albino halibut in which eye migration has failed to occur completely andthe cranium is deformed; (c) A stage 10 halibut with partial pigmentation and incomplete eye migration; (d) A stage 10 halibut with modified pigmentation, anopercular deficiency, and modified cranium; (e) A pelagic stage 10 halibut with a modified dorsal fin and incomplete eye migration; (f) An albino stage 9 halibutwhich has completed metamorphosis. The arrowhead highlights specific deformations, the closed arrow indicates the migrating eye, and the open arrow indicatespigment abnormalities. The scale bar corresponds to 1 cm.
Asymmetric adult flatfish pigmentation becomes evident duringmetamorphosis. In Japanese flounder, the pattern of develop-ment is accompanied by changes in skin histology, enzymaticactivity, and pigment cell distribution (Seikai et al., 1987). Onthe ocular side, the degeneration of larval melanophores andthe rise of adult type melanophores occur slowly and may re-sult either from the migration of differentiated cells or fromthe differentiation of extant precursors (Bolker and Hill, 2000).Most stem cells on the blind side appear to undergo cytolysis(Matsumoto and Seikai, 1992; Seikai, 1992; Seikai et al., 1987,1993) and not apoptosis.
The molecular and physiological processes underlyingabnormality in juvenile flatfish probably have their origin inearlier larval stages, long before it is manifested in the exter-nal morphology. It is, therefore, important to identify molecularmarkers in pre-metamorphosis larvae that differentiate and arediagnostic of, normal and abnormal metamorphosis. The currentdilemma, therefore, is that separation of individuals into nor-mally and abnormally developing fish is based on macroscopicmorphology, which only allows identification of abnormal indi-viduals at pro-metamorphosis. It is essential to establish a meansby which to separate normal and abnormally developing larvaebefore morphological differences become evident.
A suite of molecular methodologies and tools are now beingdeveloped for a number of different flatfish (express sequencetags, microarrays, specific antisera, stage-specific gene tran-scripts, and proteins, etc.), and these may provide the means for
early identification of larvae undergoing normal and abnormaldevelopment. Several studies have demonstrated the causalrelationship between TH modifications and abnormal metamor-phosis (Okada et al., 2005; Tagawa and Aritaki, 2005), and thegeneration of tools for species-specific assessment of the THaxis will provide important clues about regulation of metamor-phosis. However, considerably more effort will be required toelucidate and understand the molecular basis of normal meta-morphosis in order to understand why and how this process goeswrong during aquaculture.
MOLECULAR METHODS AND TOOLS APPLIEDTO METAMORPHOSIS
Associated with the change in external phenotype, there isalso remodeling of multiple organ systems during metamor-phosis. The process must be underpinned by differential geneexpression which, given the extensive transformations that oc-cur, might include the induction or repression of hundreds, if notthousands of genes. However, knowledge of differential geneexpression remains disappointingly limited, with most of themetamorphosis-associated genes identified to date being clonedas a result of a candidate-gene approach. This approach includescloning of flatfish homologues of genes identified as beingdifferentially expressed during amphibian metamorphosis (for
reviews in fisheries science vol. 16 S1 2008
Downloaded By: [Göteborg University Library] At: 10:49 16 September 2008

102 D. M. POWER ET AL.
Figure 5 Osteoclast activity as indicated by tartrate resistant acid phosphatase (TRAP) activity in the ocular side of the frontal bones (black) and abocular sideof the frontal bones (white). N = 9 (a), N = 10 (b). (a) Stage 9 normal (left panel) and abnormal (right panel) Atlantic halibut (Hippoglossus hippoglossus) larvae(a), and normal (left panel) and abnormal (right panel) juveniles (b) (taken from Sæle et al., 2006a). No significant difference in TRAP activity is detected in theocular versus abocular frontal bone of normal and abnormal stage 9 halibut. However, the total TRAP activity in the frontal bone (the sum of ocular and abocular)is significantly higher (p < 0.05) in normal stage 9 Atlantic halibut compared to those undergoing abnormal metamorphosis. In normal juvenile Atlantic halibut,the TRAP activity in the ocular frontal bone is significantly higher (p < 0.05) than the abocular frontal bone (b, left panel). No difference in TRAP activity in theocular (black) and abocular (white) frontal bone is present in abnormal halibut juveniles (b, right panel).
references, see Denver et al., 2002; Schreiber and Brown, 2003;Schreiber et al., 2005) and of genes encoding proteins previouslyidentified as being involved in flatfish metamorphosis.
Subtractive Hybridization
Suppression subtractive hybridization has been employedin a number of studies of gene expression in flatfish. These
have mostly involved identification of genes responding toimmune stimuli or to xenobiotics. However, at least onemetamorphosis-induced gene in Japanese flounder, encoding thesplicing factor SFRS3, has been cloned through the use of sub-tractive hybridization (Bao et al., 2005). Subtractive hybridiza-tion has also been used to isolate candidate metamorphosis-associated genes from Atlantic halibut (our unpublished data),and some of the resulting clones represent genes known to
reviews in fisheries science vol. 16 S1 2008
Downloaded By: [Göteborg University Library] At: 10:49 16 September 2008

THE MOLECULAR AND ENDOCRINE BASIS OF FLATFISH METAMORPHOSIS 103
be differentially expressed during metamorphosis (e.g., tro-ponins; Campinho et al., 2007a). However, these efforts arelikely to be only scratching the surface, and there are some se-rious limitations in using differential cloning techniques whenthe starting material is RNA from whole pre-metamorphic andpost-metamorphic individuals. First, many tissues (e.g., pitu-itary gland) will constitute only a small fraction of the massof a larva so that the mRNAs they contain will make a cor-respondingly small contribution to total larval RNA. Genesthat are differentially expressed in such tissues may, there-fore, be under-represented in subtractive libraries due to therelative scarcity of their mRNAs. Second, many genes are ex-pressed in multiple tissues, and it is likely that some genesmay be induced or repressed in one tissue during metamor-phosis while having an unchanged or even an opposite ex-pression elsewhere in the larva. Such genes will, therefore,be difficult to clone by subtractive hybridization and othersimilar techniques.
Microdissection
Microdissection techniques, followed by RNA amplification,may circumvent these pitfalls. For example, laser capture mi-crodissection permits isolation of cells from specific micro-scopic regions of tissue. The cDNA libraries produced fromsuch cells will represent the true pattern of gene expressionin the context of the tissue or organism. Also, treatment ofpre-metamorphic larvae with thyroid hormones may producestronger induction (or repression) of gene expression than oc-curs during natural metamorphosis, thus facilitating the use ofdifferential cloning techniques to isolate such genes.
There is clearly a need for approaches to be applied for theidentification and characterization of metamorphosis-associatedgenes that require no prior knowledge or assumptions. A numberof methods could be used for this, the most powerful of whichis microarray analysis. Such an assay system has recently beenestablished and applied to the study of halibut metamorphosisduring the EU project named ARRDE.
Microarray Analysis
Microarray analysis represents an attractive approach as, inaddition to being a tool for gene discovery, it can be used tosimultaneously quantify the expression of thousands of genes.Microarrays may be generated largely or wholly from clonesobtained from subtractive hybridization or similar techniques.However, for the majority of microarrays, most or all of thespotted DNAs are derived from randomly selected clones (analternative, especially where genome sequence is available, is touse long oligonucleotides, typically of 50–70 nucleotides). Con-sequently, a considerable resource of cloned genes from the or-ganism of interest is normally required. A completely sequencedgenome is ideal as, in principle, microarrays can be prepared in
which all genes from the organism are represented. Currently,however, most of the clones used in microarray constructionare deposited in sequence databases as expressed sequence tags(ESTs) and do not cover the whole transcriptome.
The development of flatfish microarrays is still at an earlystage, although, for at least some species, considerable progressis being made. A European flounder (Platichthys flesus) microar-ray has been prepared and validated for ecotoxicology (Sheaderet al., 2006; Williams et al., 2003). Most of the clones used inthis array were differentially expressed mRNAs isolated fromthe liver of xenobiotic-exposed animals by suppressive, sub-tractive hybridization and known stress genes isolated by PCRwith degenerate primers. Array experiments using cadmium-treated flounder identified 27 up-regulated genes, some of whichwere potential novel biomarkers of cadmium exposure. A cDNAmicroarray of 871 unique cDNA clones, prepared from theleukocytes of Japanese flounder, has been successfully usedin several immunological studies (Byon et al., 2005). Althoughit would be interesting to use these microarrays in studies ofmetamorphosis, they may be of limited utility, as there may bemany metamorphosis-related genes that are expressed in tissuesother than those from which the microarrayed clones have beenobtained.
The authors of the present manuscript have developed a smallAtlantic halibut array, which includes approximately 300 iden-tified genes and a number of unidentified genes. These wereprepared from a combination of clones randomly selected froma larval cDNA library, from subtractive libraries, and candi-date genes, namely those encoding iodothyroinine deiodinases,thyroid hormone receptors, keratin A1, and PitX2 (a geneimplicated in the development of left-right asymmetry—seeHashimoto et al., 2002). Expression profiling with this microar-ray has identified some candidate metamorphosis-associatedgenes though the use of RNA from whole, as opposed to mi-crodissected, larvae is likely to have reduced both the sensitiv-ity and selectivity of the study. The genes encoding β-globin,myosin light chain, and trypsinogen were all identified as beingup-regulated at metamorphosis, consistent with the fact thatmetamorphosis is known to involve changes to muscle, GItract, and the hematopoetic system. Interestingly, three genesimplicated in vision, encoding retinol dehydrogenase, gamma-crystallin, and transducin alpha subunit were also identifiedas being up-regulated during metamorphosis, consistent withknown changes in retinal morphology during halibut metamor-phosis (Kvenseth et al., 1996). However, it should be stressedthat these data are preliminary and await confirmation by tech-niques such as real-time PCR.
In addition to the microarrays already in existence, for atleast some flatfish species, the resources required for the con-struction of more comprehensive higher density arrays are accu-mulating steadily. Analysis of the sequences deposited at Gen-Bank shows Japanese flounder to have the highest number ofannotated gene sequences, while Atlantic halibut has the mostESTs, the majority originating from the Canadian/Spanish Pleu-rogene project (Douglas et al., 2007). At the other end of the
reviews in fisheries science vol. 16 S1 2008
Downloaded By: [Göteborg University Library] At: 10:49 16 September 2008

104 D. M. POWER ET AL.
Table 1 Annotated cDNA and expressed sequence tags(ESTs) for flatfish present in the NCBI database(http://www.ncbi.nlm.nih.gov/sites/entrez) in June 2007
Species Annotated ESTs
Paralichthys dentatus 83 0Paralichthys olivaceus 1,711 9,679Scophthalmus maximus 902 24Platichthys flesus 367 8,392Solea senegalensis 405 1Microstomus kitt(Lemon sole) 3 8Pleuronectes platessa 255 373Hippoglossus hippoglossus 262 17,768Pthalmus rhombus (Brill) 26 0
spectrum, there are only 11 hits on the entire sequence databasefor the lemon sole (Microstomus kitt) (Table 1). It should berealized, however, that where large EST collections are avail-able, many genes will be represented multiple times. Thus, thenumber of individual genes cloned is considerably less thanthe number of ESTs, although the halibut EST collection mayhave a higher proportion of unique sequences, as many of theclones represented have been obtained from “normalized” li-braries. Furthermore, EST collections may under-represent keyregulatory genes such as those encoding transcription factors orendocrine/paracrine signaling molecules and their receptors, asthese tend to encode relatively scarce mRNAs. As an illustra-tion, Blast searches (http://www.ncbi.nlm.nih.gov/Blast) werecarried out to identify members of the sox family of transcrip-tion factors present within the halibut ESTs that were present inGenBank. The sox transcription factors are an important groupof proteins involved in developmental processes ranging fromneural differentiation to sex determination (Galay-Burgos et al.,2004). There are 24 sox genes in the Fugu genome, with theavailable evidence suggesting a similar number in the genomeof other fish species. However, the Blast search revealed onlythree sox genes among the 17,768 halibut ESTs, and this issymptomatic of the limited EST resources currently available forflatfish compared with other important aquaculture species. Forexample, there are over 430,000 ESTs available from Atlanticsalmon, providing the information required for the productionof high density microarrays for this species.
In summary, it will be some time before the majority of thegenes of any flatfish species are represented on a single mi-croarray. Until then, progress is likely to be made by producingarrays mainly based on analysis of ESTs from diverse tissuesand supplemented with candidate genes obtained by subtractivehybridization and by targeting cloning of potential regulatorygenes, such as those encoding transcription factors or involvedin endocrine and paracrine signaling. Microarray-based studiesof metamorphosis would be enhanced by obtaining RNA fromindividual microdissected tissues and then carrying out amplifi-cation reactions to generate sufficient material for screening.
CONCLUSIONS
Flatfish species form a major focus of the diversificationof European marine aquaculture industry. However, productionhas been severely hampered by biological problems in larvalrearing, which affect juvenile quality and production quantity;these problems represent an important bottleneck for industry.Metamorphosis of flatfish, the process by which a symmetricallarva becomes an asymmetric juvenile, is a TH-driven process,although other endocrine and abiotic factors also influence thisprocess. Evaluation of external morphology is a simple and di-rect method to determine when metamorphosis is occurring, butit cannot predict the success of metamorphosis. Moreover, nu-merous critical events also occur during metamorphosis, whichaffect subsequent juvenile quality, and cannot be determined byexamination of the external larval phenotype.
The external manifestation of abnormal metamorphosis iscommon to all commercially important flatfish cultures, al-though it is possible that the origin of abnormalities may differamong species. In a recent European project (ARRDE), sig-nificant advances have been made in understanding the biol-ogy of early life stages of Atlantic halibut, as a model flatfishspecies, that utilize two major approaches of functional ge-nomics. The candidate gene approach, where specific geneshave been cloned, their expression quantified and localized, andin some instances correlated to localization and/or tissue levelsof the gene products (hormones), has been successful in elu-cidating many aspects of thyroid and pituitary endocrinology.This approach has also confirmed that tissues such as skin, mus-cle, and blood undergo transformations that include inductionand/or repression of gene expression, and which provide molec-ular markers for evaluating the progression of metamorphosis.The microarray-based discovery approach has also proven tobe an important research tool, promising to reveal the gene ba-sis of novel regulatory mechanisms. Both approaches have thushelped demonstrate that marked differences between normal andabnormal flatfish exist at the morphological, physiological, andmolecular levels. However, further elucidation of the complexchanges occurring in different organ systems during metamor-phosis is still needed to reach full understanding of the basis ofboth normal and abnormal development.
ACKNOWLEDGEMENTS
The present invited review was prepared in the context ofthe European Specific Support Action, AQUAFUNC (EU-SSA-022685) with financial support from the Commission of theEuropean Communities. We thank the anonymous reviewerswhose comments and suggestions have contributed to improvethe manuscript. Some of the work reported was carried out in anumber of projects: “Arrested Development: The Molecular andEndocrine Basis of Flatfish Metamorphosis” (ARRDE; Q5RS-2002-01192) with financial support from the Commission of theEuropean Communities; the Swedish Council for Agricultural
reviews in fisheries science vol. 16 S1 2008
Downloaded By: [Göteborg University Library] At: 10:49 16 September 2008

THE MOLECULAR AND ENDOCRINE BASIS OF FLATFISH METAMORPHOSIS 105
and Forestry Research (FORMAS) and Plurianual funding toCCMAR from the Portuguese Science and Technology Council.
REFERENCES
Ball, J. N., and B. I. Baker. The pituitary gland: Anatomy and histo-physiology, pp. 1–205. In: Fish Physiology, Vol II (W. S. Hoar andD. J. Randall, Eds.). New York: Academic Press (1969).
Balon, E. K. Alternative ways to become a juvenile or a definitivephenotype (and on some persisting linguistic offenses). Environ.Biol. Fish., 56: 17–38 (1999).
Bao, B., G. Yang, Z. Liu, S. Li, Z. Wang, and D. Ren. Isolation of SFRS3gene and its differential expression during metamorphosis involvingeye migration of Japanese flounder Paralichthys olivaceus. Biochim.Biophys. Acta, 1725: 64–70 (2005).
Bianco, A. C., D. Salvatore, B. Gereben, M. J. Berry, and P. R. Larsen.Biochemistry, cellular and molecular biology, and physiologicalroles of the iodothyronine selenodeiodinases. Endocr. Rev. 23: 38–89 (2002).
Bishop, C. D., D. F. Erezyilmaz, T. Flatt, C. D. Georgiou, M. G.Hadfield, A. Heyland, J. Hodin, M. W. Jacobs, S. A. Maslakova,A. Pires, A. M. Reitzel, S. Santagata, K. Tanaka, and J. H. You-son. What is metamorphosis? Integr. Comp. Biol., 46: 655–661(2006).
Bjornsson, B. Th. The biology of salmon growth hormone: From day-light to dominance. Fish Physiol. Biochem., 17: 9–24 (1997).
Bjornsson, B. Th., V. Johansson, S. Benedet, I. E. Einarsdottir, J.Hildahl, T. Agustsson, and E. Jonsson. Growth hormone endocrinol-ogy of salmonids: Regulatory mechanisms and mode of action. FishPhysiol. Biochem., 27: 227–242 (2002).
Blaxter, J. H. S. Pattern and variety in development, pp. 1–58. In: FishPhysiology: The Physiology of Developing Fish, Part A: Eggs andLarvae, Vol. XI. (W. S. Hoar and D. J. Randall, Eds.). San Diego,CA: Academic Press (1988).
Bole-Feysot, C., V. Goffin, M. Edery, N. Binart, and P. A. Kelly. Pro-lactin (PRL) and its receptor: Actions, signal transduction pathways,and phenotypes observed in PRL receptor knockout mice. Endocr.Rev., 19: 225–268 (1998).
Bolker, J. A., and C. R. Hill. Pigmentation development in hatchery-reared flatfishes. J. Fish Biol., 56: 1029–1052 (2000).
Brown, N. Flatfish farming systems in the Atlantic region. Rev. Fish.Sci., 10: 403–419 (2002).
Byon, J. Y., T. Ohira, I. Hirono, and T. Aoki. Use of a cDNA microarrayto study immunity against viral hemorrhagic septicemia (VHS) inJapanese flounder (Paralichthys olivaceus) following DNA vaccina-tion. Fish Shellfish Immunol., 18: 135–147 (2005).
Calduch-Giner, J. A., H. Duval, F. Chesnel, G. Boeuf, J. Perez-Sanchez,and D. Boujard. Fish growth hormone receptor: Molecular char-acterization of two membrane-anchored forms. Endocrinol., 142:3269–3273 (2001).
Campinho, M. A., D. M. Power, and G. E. Sweeney. Isolation andidentification of two slow troponin T genes in the Sparus aurata:In silico comparative genomic analysis with Fugu rubripes. Arch.Anim. Breed., 48: 77–78 (2005a).
Campinho, M. A., D. M. Power, and G. E. Sweeney. Deiodinase ex-pression during halibut metamorphosis: Quantitative PCR analysis.Paper presented at the 7th International Marine Biotechnology Con-ference, St. John’s, Newfoundland, and Labrador, Canada (2005b).
Campinho, M. A., D. M. Power, and G. E. Sweeney. Identification andanalysis of teleost slow muscle troponin T (sTnT) and intronlessTnT genes. Gene, 361: 67–79 (2005c).
Campinho, M. A., N. Silva, M. A. Nowell, L. Llewellyn, G. E. Sweeney,and D. M. Power. Troponin T isoform expression is modulatedduring Atlantic Halibut metamorphosis. BioMed Central—Dev., 7:71 (2007a).
Campinho, M. A., N. Silva, G. E. Sweeney, and D. M. Power. Molec-ular, cellular, and histological changes in skin from a larval to anadult phenotype during bony fish metamorphosis. Cell Tissue Res.,327: 267–284 (2007b).
Campinho, M. A., G. E. Sweeney, and D. M. Power. Regulation oftroponin T expression during muscle development in sea breamSparus auratus Linnaeus: The potential role of thyroid hormones. J.Exp. Biol., 209: 4751–4767 (2006).
Company, R., A. Astola, C. Pendon, M. M. Valdivia, and J. Perez-Sanchez. Somatotropic regulation of fish growth and adiposity:Growth hormone (GH) and somatolactin (SL) relationship. Comp.Biochem. Physiol. C: Pharmacol. Toxicol. Endocrinol., 130: 435–445 (2001).
de Jesus, E. G., T. Hirano, and Y. Inui. Changes in cortisol and thyroidhormone concentrations during early development and metamor-phosis in the Japanese flounder, Paralichthys olivaceus. Gen. Comp.Endocrinol., 82: 369–376 (1991).
de Jesus, E. G., T. Hirano, and Y. Inui. Flounder metamorphosis: Itsregulation by various hormones. Fish Physiol. Biochem., 11: 323–328 (1993).
de Jesus, E. G., Y. Inui, and T. Hirano. Cortisol enhances the stimulatingaction of thyroid hormones on dorsal fin-ray resorption of flounderlarvae in vitro. Gen. Comp. Endocrinol., 79: 167–173 (1990).
de Jesus, E. G., J. D. Toledo, and M. S. Simpas. Thyroid hormonespromote early metamorphosis in grouper (Epinephelus coioides)larvae. Gen. Comp. Endocrinol., 112: 10–16 (1998).
Denver, R. J., G. C. Boorse, and K. A. Glennemeier. Endocrinologyof complex life cycles: Amphibians, pp. 469–513. In: Hormones,Brain, and Behavior, Vol. 2 (D. Pfaff, A. Arnold, A. Etgen, S.Fahrbach, R. Moss, R. Rubin, Eds). San Diego, CA: AcademicPress (2002).
Dickey-Collas, M. The occurrence of juvenile pigmentation abnor-malities in plaice (Pleuronectes platessa) larvae fed on enrichedand unenriched Artemia salina naupill. J. Fish Biol., 42: 787–795(1993).
Dinis, M. T., L. Ribeiro, F. Soares, and C. Sarasquete. A review on thecultivation potential of Solea senegalensis in Spain and in Portugal.Aquaculture, 176: 27–38 (1999).
Douglas, S. E., L. C. Knickle, J. Kimball, and M. E. Reith. Comprehen-sive EST analysis of Atlantic halibut (Hippoglossus hippoglossus),a commercially relevant aquaculture species. BMC Genom., 8: 144(2007).
Eales, J. G. Thyroid function in cyclostomes and fishes, pp. 341–436.In: Hormones and Evolution, Vol. 1. (E. Barrington, Ed.). New York:Academic Press (1979).
Einarsdottir, I. E., L. Anjos, J. Hildahl, B. Th. Bjornsson, and D. M.Power. Isolation of Atlantic halibut pituitary hormones by contin-uous elution electrophoresis followed by fingerprint identification,and assessment of growth hormone content during larval develop-ment. Gen. Comp. Endocrinol., 150: 355–363 (2007).
Einarsdottir, I. E., N. Silva, D. M. Power, H. Smaradottir, and B. Th.Bjornsson. Thyroid and pituitary gland development from hatching
reviews in fisheries science vol. 16 S1 2008
Downloaded By: [Göteborg University Library] At: 10:49 16 September 2008

106 D. M. POWER ET AL.
through metamorphosis of a teleost flatfish, the Atlantic halibut.Anat. Embryol. (Berl), 211: 47–60 (2006).
Ellis, T., B. R. Howell, and J. Hayes. Morphological differences be-tween wild and hatchery-reared turbot. J. Fish Biol., 50: 1124–1128(1997).
Forsyth, I. A., and M. Wallis. Growth hormone and prolactin molecularand functional evolution. J. Mammary Gland Biol. Neoplasia, 7:291–312 (2002).
Fukada, H., Y. Ozaki, A. L. Pierce, S. Adachi, K. Yamauchi, A. Hara,P. Swanson, and W. W. Dickhoff. Salmon growth hormone receptor:Molecular cloning, ligand specificity, and response to fasting. Gen.Comp. Endocrinol., 139: 61–71 (2004).
Fukamachi, S., M. Sugimoto, H. Mitani, and A. Shima. Somatolactinselectively regulates proliferation and morphogenesis of neural-crestderived pigment cells in medaka. Proc. Natl. Acad. Sci. USA, 101:10661–10666 (2004).
Fukamachi, S., Y. Wakamatsu, and H. Mitani. Medaka double mu-tants for color interfere and leucophore free: Characterization ofthe xanthophore-somatolactin relationship using the leucophore freegene. Dev. Genes Evol., 216: 152–157 (2006).
Galay-Burgos, M., L. Llewellyn, C. C. Mylonas, A. V. M. Canario,S. Zanuy, and G. E. Sweeney. Analysis of the Sox gene familyin the European sea bass (Dicentrarchus labrax). Comp. Biochem.Physiol., 137: 279–284 (2004).
Galay-Burgos, M., D. M. Power, L. Llewellyn, and G. E. Sweeney.Thyroid hormone receptor expression during metamorphosis of At-lantic halibut (Hippoglossus hippoglossus). Mol. Cell. Endocrinol.,281: 56–63 (2008).
Gartner, Jr., J. V. Observation on anomalous conditions is some flatfish(Pisces: Pleuronectiformes), with a new record of partial albinism.Environ. Biol. Fish., 17: 141–152 (1986).
Gjøen, H. M., and H. B. Bentsen. Past, present, and future of geneticimprovement in salmon aquaculture. ICES J. Mar. Sci., 54: 1009–1014 (1997).
Glover, K. A., T. Svasand, I. Olesen, and M. Rye. Atlantic halibut(Hippoglossus hippoglossus). In: Genetic Effects of Domestication,Culture, and Breeding of Fish and Shellfish, and Their Impactson Wild Populations. GENIMPACT Project: Evaluation of GeneticImpact of Aquaculture Activities on Native Populations. June 12–17,Viterbo (D. Crosetti, S. Lapegue, I. Olesen, and T. Svaasand, Eds.).http://genimpact.imr.no/ (2006).
Gronas, W., V. Kiingsheim, Y. Attramadal, and B. T. Walther. Devel-opment of pigmentation in turbot (Scophthalamus maximus) larvaeunder different rearing conditions. Fish Farm. Technol., 129–133(1993).
Gutt, J. The growth of juvenile flounders (Platichthys flesus) at salinitiesof 0, 5, 15, and 35 percentages. Z. Angew. Ichtyol., 1: 17–26 (1985).
Hamre, K., M. Moren, J. Solbakken, I. Opstad, and K. Pittman. The im-pact of nutrition on metamorphosis in Atlantic halibut (Hippoglossushippoglossus L.). Aquaculture, 250: 555–565 (2005).
Hardison, R. Hemoglobins from bacteria to man: Evolution of differentpatterns of gene expression. J. Exp. Biol., 201: 1099–1117 (1998).
Hashimoto, H., A. Mizuta, N. Okada, T. Suzuki, M. Tagawa, K. Tabata,Y. Yokoyama, M. Sakaguchi, M. Tanaka, and H. Toyohara. Isolationand characterization of a Japanese flounder clonal line, reversed,which exhibits reversal of metamorphic left-right asymmetry. Mech.Dev., 111: 17–24 (2002).
Helland, S. J., and B. Grisdale-Helland. Replacement of fish mealwith wheat gluten in diets for Atlantic halibut (Hippoglossus
hippoglossus): Effect on whole-body amino acid concentrations.Aquaculture, 261: 1363–1370 (2006).
Heyland, A., D. A. Price, M. Bodnarova-Buganova, and L. L. Moroz.Thyroid hormone metabolism and peroxidase function in two non-chordate animals. J. Exp. Zool. B: Mol. Dev. Evol., 306: 551–566(2006).
Hildahl, J., M. Galay-Burgos, G. E. Sweeney, I. E. Einarsdottir, andB. Th. Bjornsson.Identification of two isoforms of Atlantic halibutinsulin-like growth factor-I receptor genes and quantitative geneexpression during metamorphosis. Comp. Biochem. Physiol., 147:395–401 (2007a).
Hildahl, J., D. M. Power, B. Th. Bjornsson, and I. E. Einarsdottir.The involvement of the growth hormone—Insulin-like growth factorI system in cranial remodeling during halibut metamorphosis isindicated by tissue- and stage-specific receptor gene expression andthe presence of growth hormone receptor protein. Cell Tiss. Res.,332: 211–225 (2008).
Hildahl, J., G. E. Sweeney, M. Galay-Burgos, I. E. Einarsdottir, and B.Th. Bjornsson. Cloning of Atlantic halibut growth hormone receptorgenes and quantitative gene expression during metamorphosis. Gen.Comp. Endocrinol., 151: 143–152 (2007b).
Hodin, J. Expanding networks: Signaling components in and a hypoth-esis for the evolution of metamorphosis. Integr. Comp. Biol., 46:719–742 (2006).
Houde, E. D. Developmental abnormalities of the flatfish Acbirus lin-eatus reared in the laboratory. Fish. Bull., 69: 537–544 (1971).
Hsiao, C. D., W. Y. Tsai, L. S. Horng, and H. J. Tsai. Molecu-lar structure and developmental expression of the three muscle-type troponin T genes in zebrafish. Dev. Dyn., 227: 266–279(2003).
Huang, H. C., and D. D. Brown. Prolactin is not a juvenile hormonein Xenopus laevis metamorphosis. Proc. Natl. Acad. Sci. USA, 97:195–199 (2000).
Huang, L., A. M. Schreiber, B. Soffientino, D. A. Bengston, and J. L.Specker. Metamorphosis of the summer flounder (Paralichthys den-tatus): Thyroidal status and the timing of gastric gland formation. J.Exp. Zool., 280: 413–429 (1998).
Imsland, A. K., and T. M. Jonassen. Growth and age at first maturity inturbot and halibut reared under different photoperiods. Aquac. Int.,11: 463–475 (2003).
Inui, Y., and S. Miwa. Thyroid hormone induces metamorphosis offlounder larvae. Gen. Comp. Endocrinol., 60: 450–454 (1985).
Kajimura, S., N. Kawaguchi, T. Kaneko, I. Kawazoe, T. Hirano, N.Visitacion, E. G. Grau, and K. Aida. Identification of the growthhormone receptor in an advanced teleost, the tilapia (Oreochromismossambicus), with special reference to its distinct expression pat-tern in the ovary. J. Endocrinol., 181: 65–76 (2004).
Kakizawa, S., A. Ishimatsu, T. Takeda, T. Kaneko, and T. Hirano.Possible involvement of somatolactin in the regulation of plasmabicarbonate for the compensation of acidosis in rainbow trout. J.Exp. Biol., 200: 2675–2683 (1997).
Kakizawa, S., T. Kaneko, S. Hasegawa, and T. Hirano. Activation ofsomatolactin cells in the pituitary of the rainbow trout Oncorhynchusmykiss by low environmental calcium. Gen. Comp. Endocrinol., 91:298–306 (1993).
Kakizawa, S., T. Kaneko, S. Hasegawa, and T. Hirano. Effects of feed-ing, fasting, background adaptation, acute stress, and exhaustive ex-ercise on the plasma somatolactin concentrations in rainbow trout.Gen. Comp. Endocrinol., 98: 137–146 (1995).
reviews in fisheries science vol. 16 S1 2008
Downloaded By: [Göteborg University Library] At: 10:49 16 September 2008

THE MOLECULAR AND ENDOCRINE BASIS OF FLATFISH METAMORPHOSIS 107
Kakizawa, S., T. Kaneko, and T. Hirano. Elevation of plasma so-matolactin concentrations during acidosis in rainbow trout (On-corhynchus mykiss). J. Exp. Biol., 199: 1043–1051 (1996).
Kohrle, J. The selenoenzyme family of deiodinase isozymes controlslocal thyroid hormone availability. Rev. Endocr. Metab. Disord., 1:49–58 (2000).
Kopchick, J. J., and J. M. Andry. Growth hormone (GH), GH receptor,and signal transduction. Mol. Genet. Metab., 71: 293–314 (2000).
Kvenseth, A. M., K. Pittman, and J. V. Helvik. Eye development inAtlantic halibut (Hippoglossus hippoglosus)—Differentiation anddevelopment of the retina from early yolk sac stages through meta-morphosis. Can. J. Fish. Aquat. Sci., 54: 2524–2532 (1996).
Leatherland, J. F. Reflections on the thyroidology of fishes:From molecules to humankind. Guelph Icthyol. Rev., 2: 1–67(1994).
Leclercq, D. Turbot broodstock management: A key point to mid-term progress of the turbot industry, pp. 3–13. In: Turbot Culture:Problems and Prospects (P. Lavens and R. A. M. Remmerswaal,Eds.). Oostende, Belgium: European Aquaculture Society (1994).
Lee, L. T. O., G. Nong, Y. H. Chan, D. L. Y. Tse, and C. H. K.Cheng. Molecular cloning of a teleost growth hormone receptor andits functional interaction with human growth hormone. Gene, 270:121–129 (2001).
Llewellyn, L., M. A. Nowell, V. P. Ramsurn, T. Wigham, G. E. Sweeney,B. Kristjansson, and O. Halldorsson. Molecular cloning and devel-opmental expression of the halibut thyroid hormone receptor-α. J.Fish Biol., 55: 148–155 (1999).
Llewellyn, L., V. P. Ramsurn, G. E. Sweeney, T. Wigham, and D. M.Power. Expression of thyroid hormone receptor during early devel-opment of the sea bream (Sparus aurata). Ann. NY Acad. Sci., 839:610–611 (1998).
Marchand, O., M. Duffraisse, G. Triqueneaux, R. Safi, and V. Laudet.Molecular cloning and developmental expression patterns of thy-roid hormone receptors and T3 target genes in the turbot (Scoph-thalmus maximus) during post-embryonic development. Gen. Comp.Endocrinol., 135: 345–357 (2004).
Matsumoto, J., and T. Seikai. Asymmetric pigmentation and pigmentdisorders in Pleuronectiformes (flounders). Pigment Cell. Res., 2:275–282 (1992).
McCormick, S. D. Endocrine control of osmoregulation in teleost fish.Am. Zool., 41: 781–794 (2001).
Miwa, S., and Y. Inui. Effects of various doses of thyroxine and tri-iodothyronine on the metamorphosis of flounder (Paralichthys oli-vaceus). Gen. Comp. Endocrinol., 67: 356–363 (1987a).
Miwa, S., and Y. Inui. Histological changes in the pituitary-thyroidaxis during spontaneous and artificially-induced metamorphosis oflarvae of the flounder Paralichthys olivaceus. Cell. Tiss. Res., 249:117–123 (1987b).
Miwa, S., and Y. Inui. Thyroid hormone stimulates the shift of ery-throcyte populations during metamorphosis of the flounder. J. Exp.Zool., 259: 222–228 (1991).
Miwa, S., M. Tagawa, Y. Inui, and T. Hirano. Thyroxine surge inmetamorphosing flounder larvae. Gen. Comp. Endocrinol., 70: 158–163 (1988).
Miwa, S., K. Yamano, and Y. Inui. Thyroid hormone stimulates gastricdevelopment in flounder larvae during metamorphosis. J. Exp. Biol.,261: 424–430 (1992).
Moren, M., I. Opstad, and K. Hamre. A comparison of retinol, reti-nal, and retinyl ester concentrations in larvae of Atlantic halibut
(Hippoglossus hippoglossus L.) fed Artemia or zooplankton. Aquac.Nutr., 10: 253–259 (2004).
Moren, M., I. Opstad, T. Van der Meeren, and K. Hamre. Iodine enrich-ment of Artemia and enhanced levels of iodine in Atlantic halibutlarvae (Hippoglossus hippoglossus L.) fed the enriched Artemia.Aquac. Nutr., 12: 97–102 (2006).
Nakao, N., Y. Higashimoto, T. Ohkubo, H. Yoshizato, N. Nakai, K.Nakashima, and M. Tanaka. Characterization of structure and ex-pression of the growth hormone receptor gene of the Japanese floun-der (Paralichtys olivaceus). J. Endocrinol., 182: 157–164 (2004).
Nguyen, N., M. Sugimoto, and Y. Zhu. Production and purificationof recombinant somatolactin beta and its effects on melanosomeaggregation in zebrafish. Gen. Comp. Endocrinol., 145: 182–187(2006).
Norris, D.O. (Ed.). Vertebrate Endocrinology. San Diego, CA: Aca-demic Press (1997).
Oetting, A., and P. M. Yen. New insights into thyroid hormone action.Best Pract. Res. Clin. Endocrinol. Metab., 21: 193–208 (2007).
Okada, N., T. Morita, M. Tanaka, and M. Tagawa. Thyroid hormonedeficiency in abnormal larvae of the Japanese flounder Paralichthysolivaceus. Fish. Sci., 71: 107–114 (2005).
Okada, N., Y. Takagi, T. Seikai, M. Tanaka, and M. Tagawa. Asymmet-rical development of bones and soft tissues during eye migration ofmetamorphosing Japanese flounder (Paralichthys olivaceus). CellTiss. Res., 304: 59–66 (2001).
Okada, N., Y. Takagi, M. Tanaka, and M. Tagawa. Fine structure ofsoft and hard tissues involved in eye migration in metamorphosingJapanese flounder (Paralichthys olivaceus).Anat. Rec. A: Discov.Mol. Cell. Evol. Biol., 273: 663–668 (2003).
Olivereau, M., and M. Rand-Weaver. Immunocytochemical study of thesomatolactin cells in the pituitary of Pacific salmon, Oncorhynchusnerka, and O. keta at some stages of the reproductive cycle. Gen.Comp. Endocrinol., 93: 28–35 (1994).
Ono, M., Y. Takayama, M. Rand-Weaver, S. Sakata, T. Yasunaga, T.Noso, and H. Kawauchi. Complementary DNA cloning of somato-lactin, a pituitary protein related to growth hormone and prolactin.Proc. Natl. Acad. Sci. USA, 87: 4330–4334 (1990).
Perez-Sanchez, J. The involvement of growth hormone in growth regu-lation, energy homeostasis, and immune function in the gilthead seabream (Sparus aurata): A short review. Fish Physiol. Biochem., 22:135–144 (2000).
Peter, R. E., and T. A. Marchant. The endocrinology of growth in carpand related species. Aquaculture, 129: 299–321 (1995).
Pittman, K., A. Jelmart, T. Naess, S. Harboe, and K. Watanabe. Plas-ticity of viable post-metamorphic forms of farmed Atlantic halibutHippoglossus hippoglossus. Aquac. Res., 29: 949–954 (1998).
Pittman, K., A. B. Skiftesvik, and L. Berg. Morphological and behav-ioral development of halibut, (Hippoglossus hippoglossus L.) larvae.J. Fish Biol., 37: 455–472 (1990).
Planas, J., P. Swanson, M. Rand-Weaver, and W. Dickhoff. Somato-lactin stimulates in vitro gonadal steroidogenesis in coho salmon,Oncorhynchus kisutch. Gen. Comp. Endocrinol., 87: 1–5 (1992).
Policansky, D. The asymmetry of flounders. Sci. Am., 246: 96–102(1982).
Power, D. M. Developmental ontogeny of prolactin and its receptor infish. Gen. Comp. Endocrinol., 142: 25–33 (2005).
Power, D. M., L. Llewellyn, M. Faustino, M. A. Nowell, B. Th.Bjornsson, I. E. Einarsdottir, A. V. M. Canario, and G. E. Sweeney.Thyroid hormones in growth and development of fish. Comp.
reviews in fisheries science vol. 16 S1 2008
Downloaded By: [Göteborg University Library] At: 10:49 16 September 2008

108 D. M. POWER ET AL.
Biochem. Physiol. C: Pharmacol. Toxicol. Endocrinol., 130: 447–459 (2001).
Pretty, R., T. Naitoh, and R. J. Wassersug. Metamorphic shortening ofthe alimentary tract in anuran larvae (Rana catesbeiana). Anat. Rec.,242: 417–423 (1995).
Rand-Weaver, M., T. G. Pottinger, and J. P. Sumpter. Plasma soma-tolactin concentrations in salmonid fish are elevated by stress. J.Endocrinol., 138: 509–515 (1993).
Rand-Weaver, M., and P. Swanson. Plasma somatolactin levels in cohosalmon (Oncorhynchus kisutch) during smoltification and sexualmaturation. Fish Physiol. Biochem., 11: 175–182 (1993).
Rand-Weaver, M., P. Swanson, H. Kawauchi, and W. W. Dickhoff. So-matolactin, a novel pituitary protein: Purification and plasma levelsduring reproductive maturation of coho salmon. J. Endocrinol., 133:393–403 (1992).
Renfro, J. L. Hormonal regulation of renal inorganic phosphate trans-port in the winter flounder, Pleuronectes americanus. Fish. Physiol.Biochem., 17: 377–383 (1997).
Sæle, Ø., N. Silva, and K. Pittman. Post-embryonic remodeling of neu-rocranial elements, a comparative study of normal versus abnormaleye migration in the flatfish Atlantic halibut. J. Anat., 209: 31–41(2006a).
Sæle, Ø., H. Smaradottir, and K. Pittman. The twisted story of eyemigration in flatfish. J. Morphol., 267: 730–738 (2006b).
Sæle, Ø., J. S. Solbakken, K. Watanabe, K. Hamre, and K. Pittman. Theeffect of diet on ossification and eye migration in Atlantic halibutlarvae (Hippoglossus hippoglossus L.). Aquaculture, 220: 683–696(2003).
Sæle, Ø., J. S. Solbakken, K. Watanabe, K. Hamre, D. M. Power, andK. Pittman. Staging of Atlantic halibut (Hippoglossus hippoglossusL.) from first feeding through metamorphosis, including cranial os-sification independent of eye migration. Aquaculture, 239: 445–465(2004).
Sarropoulou, E., G. Kotoulas, D. M. Power, and R. Geisler. Geneexpression profiling of gilthead sea bream during early developmentand detection of stress-related genes by the application of cDNAmicroarray technology. Physiol. Genom., 23: 182–191 (2005).
Schreiber, A. M., and D. D. Brown. Tadpole skin dies autonomouslyin response to thyroid hormone at metamorphosis. Proc. Natl. Acad.Sci. USA, 100: 1769–1774 (2003).
Schreiber, A. M., L. Cai, and D. D. Brown. Remodeling of the intestineduring metamorphosis of Xenopus laevis. Proc. Natl. Acad. Sci.USA, 102: 3720–3725 (2005).
Schreiber, A. M., and J. L. Specker. Metamorphosis in the summerflounder (Paralichthys dentatus): Stage-specific developmental re-sponse to altered thyroid status. Gen. Comp. Endocrinol., 111: 156–166 (1998).
Schreiber, A. M., and J. L. Specker. Metamorphosis in the summerflounder, Paralichthys dentatus: Thyroidal status influences salinitytolerance. J. Exp. Zool., 284: 414–424 (1999).
Schreiber, A. M., and J. L. Specker. Metamorphosis in the summerflounder, Paralichthys dentatus: Thyroidal status influences gillmitochondria-rich cells. Gen. Comp. Endocrinol., 117: 238–250(2000).
Seikai, T. Process of pigment cell differentiation in skin on the left andright sides of the Japanese flounder, Paralichthys olivaceus, duringmetamorphosis. Jpn. J. Ichthyol., 39: 85–92 (1992).
Seikai, T., E. Hirose, and J. Matsumoto. Dual appearances of pig-ment cells from in vitro cultured embryonic cells of Japanese
flounder—An implication for a differentiation-associated clock.Pigment Cell Res., 6: 423–431 (1993).
Seikai, T., J. Matsumoto, M. Shimozaki, A. Oikawa, and T. Akiyama.An association of melanophores appearing at metamorphosisas vehicles of asymmetric skin color formation with pigmentanomalies developed under hatchery conditions in the Japaneseflounder, Paralichthys olivaceus. Pigment Cell Res., 1: 143–151(1987).
Sheader, D. L., T. D. Williams, B. P. Lyons, and J. K. Chipman. Ox-idative stress response of European flounder (Platichthys flesus) tocadmium determined by a custom cDNA microarray. Mar. Environ.Res., 62: 33–44 (2006).
Sobel, J. (Ed.). International Union for Conservation of Nature andNatural Resources. http://www.iucnredlist.org/ (1996).
Soffientino, B., and J. L. Specker. Age-dependent changes in the re-sponse of the stomach to thyroidal signaling in developmentallyarrested larval summer flounder. Gen. Comp. Endocrinol., 134: 237–243 (2003).
Solbakken, J. S., M. H. G. Berntssen, B. Norberg, K. Pittman, andK. Hamre. Different iodine and thyroid hormone levels betweenAtlantic halibut larvae fed wild zooplankton or artemia from firstexogenous feeding until post metamorphosis. J. Fish. Biol., 61:1345–1362 (2002).
Solbakken, J. S., B. Norberg, K. Watanabe, and K. Pittman. Thyroxineas a mediator of metamorphosis of Atlantic halibut (Hippoglossushippoglossus L.). Environ. Biol. Fish., 56: 53–65 (1999).
Storebakken, T., S. Refstie, and B. Ruyter (Eds.). Soy products as fatand protein sources in fishfeeds for intensive aquaculture. In J. K.Drackney (ed.), Soy in Animal Nutrition. Fed. An. Sci. (2000).
Szisch, V., N. Papandroulakis, E. Fanouraki, and M. Pavlidis. On-togeny of the thyroid hormones and cortisol in the gilthead seabream, Sparus aurata. Gen. Comp. Endocrinol., 142: 186–192(2005).
Tabata, K. Genetic factors on the albinism of Hirame Paralichthysolivaceus. Suisanzoshoku, 39: 29–35 (1991).
Tagawa, M., and M. Aritaki. Production of symmetrical flatfish bycontrolling the timing of thyroid hormone treatment in spotted hal-ibut Verasper variegatus. Gen. Comp. Endocrinol., 141: 184–189(2005).
Tanaka, M., J. B. Tanangonan, M. Tagawa, E. G. de Jesus, H. Nishida,M. Isaka, R. Kimura, and T. Hirano. Development of the pituitary,thyroid, and interrenal glands and applications of endocrinology tothe improved rearing of marine fish larvae. Aquaculture, 135: 111–126 (1995).
Thisse, C., A. Degrave, G. V. Kryukov, V. N. Gladyshev, S. Obrecht-Pflumio, A. Krol, B. Thisse, and A. Lescure. Spatial and temporalexpression patterns of selenoprotein genes during embryogenesis inzebrafish. Gene. Expr. Patt., 3: 525–532 (2003).
Tibaldi, E., Y. Hakim, Z. Uni, F. Tulli, M. de Francesco, U. Luzzana,and S. Harpaz. Effects of the partial substitution of dietary fish mealby differently processed soybean meals on growth performance,nutrient digestibility and activity of intestinal brush border enzymesin the European sea bass (Dicentrarchus labrax). Aquaculture, 261:182–193 (2006).
Torihashi, S. Morphological changes of the myenteric plexus neurons inthe bullfrog (Rana catesbeiana) duodenum during metamorphosis.J. Comp. Neurol., 302: 54–65 (1990).
Tse, D. L. Y., M. C. L. Tse, C. B. Chan, L. Deng, W. M. Zhang, H. R.Lin, and C. H. K. Cheng. Sea bream growth hormone receptor:
reviews in fisheries science vol. 16 S1 2008
Downloaded By: [Göteborg University Library] At: 10:49 16 September 2008

THE MOLECULAR AND ENDOCRINE BASIS OF FLATFISH METAMORPHOSIS 109
Molecular cloning and functional studies of the full-length cDNAand tissue expression of two alternatively spliced forms. BBA-GeneStruct. Express., 1625: 64–76 (2003).
Tvedt, H. B., T. J. Benfey, D. J. Martin-Robichaud, C. McGowan,and M. Reith. Gynogenesis and sex determination in Atlantichalibut (Hippoglossus hippoglossus). Aquaculture, 252: 573–583(2006).
Very, N. M., J. D. Kittilson, L. A. Norbeck, and M. A. Sheridan. Isola-tion, characterization, and distribution of two cDNAs encoding forgrowth hormone receptor in rainbow trout (Oncorhynchus mykiss).Comp. Biochem. Physiol. B: Biochem. Mol. Biol., 140: 615–628(2005).
Veillette, P. A., M. Merino, N. D. Marcaccio, M. M. Garcia, and J. L.Specker. Cortisol is necessary for seawater tolerance in larvae of amarine teleost the summer flounder. Gen. Comp. Endocrinol., 151:116–121 (2007a).
Veillette, P. A., X. Serrano, M. M. Garcia, and J. L. Specker. Ev-idence for the onset of feedback regulation of cortisol in lar-val summer flounder. Gen. Comp. Endocrinol., 154: 105–110(2007b).
Wada, T., M. Aritaki, and M. Tanaka. Effects of low-salinity on thegrowth and development of spotted halibut Verasper variegatus inthe larva-juvenile transformation period with reference to pituitaryprolactin and gill chloride cells responses. J. Exp. Mar. Biol. Ecol.,308: 113–126 (2004).
Weltzien, F. A., B. Norberg, J. V. Helvik, O. Andersen, P. Swanson,and E. Andersson. Identification and localization of eight distincthormone-producing cell types in the pituitary of male Atlantic hal-ibut (Hippoglossus hippoglossus L.). Comp. Biochem. Physiol. A:Mol. Integr. Physiol., 134: 315–327 (2003).
Wendelaar Bonga, S. E. The stress response in fish. Physiol. Rev., 77:591–652 (1997).
Williams, T. D., K. Gensberg, S. D. Minchin, and J. K. Chipman. ADNA expression array to detect toxic stress response in Europeanflounder (Platichthys flesus). Aquat. Toxicol., 65: 141–157 (2003).
Yadav, B. N. The Pituitary Gland. Fish Endocrinology. Delhi: DayaPublishing House, pp. 24–60 (1995).
Yamano, K. The role of thyroid hormone in fish development withreference to aquaculture. JARQ, 39: 161–168 (2005).
Yamano, K., K. Araki, K. Sekikawa, and Y. Inui. Cloning of thyroidhormone receptor genes expressed in metamorphosing flounder.Dev. Genet., 15: 378–382 (1994a).
Yamano, K., and Y. Inui. cDNA cloning of thyroid hormone receptorβ for the Japanese flounder. Gen. Comp. Endocrinol., 99: 197–203(1995).
Yamano, K., and S. Miwa. Differential gene expression of thyroidhormone receptor α and β in fish development. Gen. Comp.Endocrinol., 109: 75–85 (1998).
Yamano, K., S. Miwa, T. Obinata, and Y. Inui. Thyroid hormoneregulates developmental changes in muscle during floundermetamorphosis. Gen. Comp. Endocrinol., 81: 464–472 (1991a).
Yamano, K., M. Tagawa, E. G. de Jesus, T. Hirano, S. Miwa, and Y.Inui. Changes in whole body concentrations of thyroid hormonesand cortisol in metamorphosing conger eel. J. Comp. Physiol. B,161: 371–375 (1991b).
Yamano, K., H. Takano-Ohmuro, T. Obinata, and Y. Inui. Effectof thyroid hormone on developmental transition of myosin lightchains during flounder metamorphosis. Gen. Comp. Endocrinol.,93: 321–326 (1994b).
Youson, J. H. First metamorphosis pp. 135–198. In: Fish Physiology,Vol. XI, Part B (W. S. Hoar and D. J. Randall, Eds.). New York:Academic Press (1988).
Youson, J. H. The impact of environmental and hormonal cues on theevolution of fish metamorphosis pp. 239–277. In: Environment,Development and Evolution (B. K. Hall, R. D. Pearson, and G. B.Muller, Eds.). Cambridge: MIT Press (2004).
Youson, J. H., and A. A. Al-Mahrouki. Ontogenetic and phylogeneticdevelopment of the endocrine pancreas (islet organ) in fishes. Gen.Comp. Endocrinol., 116: 303–335 (1999).
Zhang, J., and M. A. Lazar. The mechanism of action of thyroidhormones. Ann. Rev. Physiol., 62: 439–466 (2000).
Zhu, Y., and P. Thomas. Effects of somatolactin on melanosomeaggregation in the melanophores of red drum (Sciaenops ocellatus)scales. Gen. Comp. Endocrinol., 105: 127–133 (1997).
reviews in fisheries science vol. 16 S1 2008
Downloaded By: [Göteborg University Library] At: 10:49 16 September 2008




















