
Decomposition of a 3D triangular mesh into quadrangulated patches
Upload
independentCategory
view
1download
0
Original article
Importance of unmanaged wasteland patches for the grey partridgePerdix perdix in suburban habitats
Miroslav Šálek a,*, Pavel Marhoul b, Jan Pintír c, Tomáš Kopecký a, Lukáš Slabý a
a Department of Ecology, Czech Agricultural University, 165 21 Prague 6, Czech Republicb Agency for Nature Conservation and Landscape Protection, Kališnická 4-6, 130 23 Prague 3, Czech Republic
c Department of Forest Conservation, Czech Agricultural University, 165 21 Prague 6, Czech Republic
Received 13 February 2003; accepted 14 October 2003
Abstract
A significant component of breeding habitat of the grey partridge Perdix perdix in Europe includes field boundaries with idle sites andgrassy strips, loss of which is considered as one of the reasons for the decline of grey partridge populations. We associate enduring highdensities of wild grey partridges in Prague, the Czech Republic, with the existence of unmanaged wasteland, herbaceous patches spread outbetween urbanized areas of the capital and its surrounding agricultural landscape. The study combines density estimations, home rangeanalysis, weed seed sampling and plant composition analysis, all with the aim of explaining factors influencing partridges’ habitat preference.Patch size, availability of patches within the surrounding landscape and mean height of plant cover were the most important factors influencingthe numbers of breeding pairs among patches. Numbers of weed seeds collected on the soil surface seems to be of greater importance thanspecific plant composition. Although patches supported high densities (tens of pairs km–2), the neighboring agricultural landscape supportedlow densities (<10 pairs km–2) with an influence of the distance to the nearest patch. April to June home range size varied around 2.9 ha andtended to be smaller for pairs with higher availability of patch habitat. A sufficient availability of temporary patches in the study area enableda persistence of grey partridge populations without any support through hand-reared birds or systematic predator control. A surplus ofoffspring may potentially saturate the less attractive surrounding landscape but only to a reduced extent limited by its carrying capacity.
© 2003 Elsevier SAS. All rights reserved.
Keywords: Food availability; Fragmented landscape; High-density population; Idle habitats; Landscape diversity; Weed seeds
1. Introduction
Grey partridge Perdix perdix (L.) prefers open, low-intensity, mixed farmland comprising small fields boundedby hedges and grassy habitats (Aebischer and Kavanagh,1997) but it occurs commonly also in intensive cereal ecosys-tems in Western Europe (Reitz, 1999; Bro et al., 2003).However, a valuable component of the grey partridge habitatrequirements includes unmanaged or extensively managedsites with a predominantly herbaceous cover such as idlefields, boundaries of managed land or grassy strips withdispersed shrubs or some specific (substitutional) habitatssuch as hedgerows in Britain (Blank et al., 1967; Potts, 1986;Meriggi et al., 1990). Rapid and broad reductions of suchhabitats linked to agriculture intensification that result in a
dramatic decline of European partridge populations maydemonstrate the importance of such habitats for this birdspecies (Potts, 1980; Aebischer and Kavanagh, 1997).
Similarly, agricultural land has started to encroach onmost open country at the expense of the loss of suitablepartridge habitat since the second half of the 1950s in theCzech Republic, Central Europe. This process lead to aprogressive decrease in grey partridge populations, reachinga decline of approximately 95% between 1965 and the end ofthe 1980s (Št{astný et al., 1997). Bouchner and Fišer (1967)reported that extensively grazed wastelands, which wereunequivocally preferred to cornfields as nesting habitat, werevery attractive for the grey partridge during the 1960s. How-ever, wastelands have gradually become rare not only be-cause of agricultural intensification and urbanization but amajority of them have been left ungrazed resulting in theirbeing overgrown by dense shrubs and high trees, which areunsuitable for grey partridges in most cases.
* Corresponding author.E-mail address: [email protected] (M. Šálek).
Acta Oecologica 25 (2004) 23–33
www.elsevier.com/locate/actoec
© 2003 Elsevier SAS. All rights reserved.doi:10.1016/j.actao.2003.10.003

In recent decades, unmanaged land on city outskirts, leftidle for some years prior to its development for suburbs,shopping centers or infrastructure, may thus have played aremarkable role for grey partridge populations. Like non-linear and patchy habitats with herb cover, such land re-sembles former wastelands and forms part of a suburbanlandscape mosaic dominated by fields and various construc-tions. There are some indications that such suburban areasadjacent to large human settlements in the Czech Republicare inhabited by grey partridge populations in higher densi-ties than the remaining majority of the agricultural landscape(Št{astný et al., 1997; Fuchs et al., 2003).
The objectives of this study were to: (a) analyze the spatialpattern of grey partridge numbers within the city of Prague,the largest human settlement of the Czech Republic, in rela-tion to unmanaged wastelands; (b) evaluate the importanceand highlight the best types of wasteland patches for greypartridge populations; (c) emphasize the value of suchpatches for species diversity of suburban landscapes in gen-eral.
2. Study area
The study was carried out on locations at the border of theadministrative territory of Prague, the Czech Republic, be-tween urbanized areas of the city and its surrounding agricul-tural landscape (Fig. 1). This zone of about 165 km2 stretchesout with varying width (but no more than 5 km in the NE–SEdirection) around the entire city and is locally interrupted byimmediately neighboring urbanized areas and forests. It isintersected by a dense infrastructure, numbers of small in-dustrial estates and human settlements (villages). Large lots
of uniform land, formed during the 1950s and 1960s by theprocess of small field unification and by crop assortmentsimplification, recently prevailing in the Czech agriculturallandscape, are regularly managed with a rotation of winterwheat (Triticum, 30%), spring cereal (barley Hordeum,13%), other cereals (13%), rape (Brassica, 12%), corn (Zeamays, 11%), lucerne and clover (Medicago and Trifolium,6%), beet (Beta, 3%) and potato (Solanum, 1.5%) being themost important crops (Data from 2002, Czech StatisticalOffice, Prague).
Non-linear wasteland patches (further referred to as“patches”) have been defined as unmanaged (idle) herba-ceous communities in their early stages of succession (fordominant plant species see Appendix). Patches, physicallyseparated one from another, consisted mainly of land left idleby city planners for future development projects such asshopping centers, roads or houses. Neighboring patches aredefined as those, which are completely separated by con-struction, forests or highways. On the other hand, connectedpatches, joined together even by a short section or by anarrow local road have been pooled to form only one patch.In total, we identified 273 patches totaling up to 840 ha, i.e.an average of 3.1 ha per patch, and representing approxi-mately 5% of the entire zone between the city and the sur-rounding agricultural landscape. This calculation does notinclude the area of the Ruzyne International Airport (north-west of the city) with regularly mowed (thus managed)greenswards also inhabited by numerous long-time localpartridge populations (Št{astný et al., 1997; own unpublisheddata from 2000 to 2002).
Linear habitats (or “lines”) have been defined as perma-nent field edges, grassy strips, ditches or roads flanked byherbs as well as orchard borders, which were also regularlyvisited by partridges (own observations). Temporary (1-year)
Fig. 1. The distribution of monitored patches, regions of agricultural landscapes and position of the Stodulky region, along the Prague City border.
24 M. Šálek et al. / Acta Oecologica 25 (2004) 23–33

rims between fields of different crops, windbreak tree lines orhard edges of buildings or forests where herbaceous cover isstrictly limited, were not included in this category. One linemay be characterized as a section between two opposite sitesof field borders, ditches or roads unless highways providedfor two separated lines.
Predators such as martens Martes spp., fox Vulpes vulpes(L.), marsh harrier Circus aeruginosus (L.), goshawk Accipi-ter gentilis (L.) and sparrowhawk A. nisus (L.) hunting in thestudy area occasionally attacked or killed partridges whilemagpies Pica pica (L.) and hedgehogs Erinaceus spp. pre-dated some grey partridge nests (own unpublished data).
Prague was chosen as a model city for the purposes of thisstudy for the following reasons. First, wild grey partridgepopulations have not been supported by any special manage-ment or stocked by releasing of hand-reared birds from farmsat this location. Second, reduced hunting activities next tohuman settlements exclude any systematic predator control,which may artificially strengthen survival of grey partridges(Potts, 1980).
3. Material and methods
3.1. Data collection
3.1.1. PatchesPatches were mapped using colored aerial orthophoto-
maps (taken in early Summer). They were distinguished assegments with an uneven mosaic of green, rufous (herbs),brown and greyish (bare soil) colors of various hues withsporadic dark-green segments or their clusters (shrubs). Incontrast, field or meadow vegetation cover was uniformlyrich-green with a regular pattern (ruts from machinery or/androws of sown crop). The accuracy of such sorting was con-firmed by checking a number of sites in the field.
Grey partridge pairs were mapped by two to four observ-ers from the second half of April to the beginning of May in2001 and 2002 on a sample of 26 patches of various sizesbetween 2.4 and 16.5 ha (mean area 7.0 ha, 183 ha in total)distributed throughout the study area (Fig. 1). Patches iso-lated from agricultural landscape by buildings or forests wereavoided in this selection. The number of grey partridges(flushed pairs or presence of droppings) in patches was esti-mated through detailed surveys twice a year and the highervalue of these two (expected to be closer to the real pair statusof each patch) was used for analysis.
Habitat description of patches performed in June 2002includes the following parameters: mean height of herb veg-etation, proportion of shrubs and trees and rate of directhuman disturbances. The human disturbance was distin-guished as a subjective category ranging from absence ofdisturbances (category 0) to intense frightening (category 4)by children, cyclists and/or dog walkers depending on thedistance to the nearest human settlement and its size, physi-cal accessibility of the patch and the number of paths cross-
ing the patch found during our visits. The character of plantcommunities was evaluated from a proportional coverage ofplant species in patches or their individual sections in caseswhere they were of markedly different composition. Dis-tances and areas of patches were assessed from maps.
3.1.2. Agricultural landscapeFive regions (Fig. 1) were selected for analysis of par-
tridge spatial distribution outside the patches: Cholupice(south of the city), Uhríneves (southeast), Dubec (east), Vi-nor (northeast) and Horomerice (northwest). The city map(including selected surrounding regions) was lined by a400 × 400 m grid determining 16 ha quadrat areas. This areawas accepted as a compromise between field-work efficiencyand the need to reduce the risk of repeated registration of thesame pair of grey partridges in neighboring quadrats. Pre-breeding and breeding home ranges of 11 radio-tracked greypartridges studied in previous years in Prague were taken intoconsideration; their mean home range sizes varied around3.73 ha with a maximum of 9 ha (Šálek et al., 2002).
Quadrats in which at least 1% of the area (0.16 ha) repre-sented a suitable habitat for the grey partridge (grassy lines,shrubs and small weeded spots) were taken into account forfurther analysis. Therefore, we avoided quadrats in whichforests, buildings, uniformly sowed fields and wetlands aswell as their hard edges without any grassy or weeded habi-tats were exclusively present because they lack fundamentalhabitat requirements for grey partridge. Finally, quadratsmade up of more than 10% (1.6 ha) of small weeded spotswere also disregarded because these spots, when added to-gether may have had an undesirable confounding effect asthey resemble a patch itself. Landscape characteristics wereread from digitalized aerial orthophoto-maps using the GISsoftware ArcView GIS 3.2a (Environmental Systems Re-search Institute, Redlands, California, USA).
All selected quadrats with a total area of 19 km2 werecarefully examined by walking through the areas during Mayand June 2000 and 2001 with the aim of unearthing anyevidence of the presence of grey partridges. Most occur-rences were confirmed by fresh droppings along roads or atfield borders but many pairs were observed directly.
3.1.3. Population density
The Stodulky region (southwest of the city), representingone of the best grey partridge breeding sites in Prague (Šáleket al., 2002), was selected as the area for population densityand food supply analyses. The established study plot had arectangular shape (ca. 2700 × 800 m, 188.6 ha in total)comprised of one large and one small patch with 34.3 ha(18% of the study plot) passing onto a uniform agriculturallandscape of 154.3 ha (82%; Fig. 2). From March to thebeginning of May 2000 and 2001, groups of two to fiveobservers searched this study plot thoroughly while register-ing and recording all pairs and trios (groups of two males
25M. Šálek et al. / Acta Oecologica 25 (2004) 23–33

attempting to mate with one female resulting in one paireventually) onto maps. In total, 13 detailed grey partridgesurveys were organized (six in 2000 and seven in 2001).
3.1.4. Food supply
Weed seeds, as the key item in an adult grey partridge’sdiet, were collected on 60 locations (points) set out over theentire Stodulky study plot. These points were divided amongthe three main habitats (patches: 15 points, grassy strips: 19,fields: 26) representing also various distances to habitatedges as well as to other habitats. The east–west orientationof the site and the presence of patches on both its east andwest ends should control any influence of prevailing westernwinds on seed dispersal.
All seed samples were collected in February (2.2.2000and 18.2.2001) because this represents the period of inten-sive pairing while food is limited after winter and sampling isalso easiest on the vegetation-free soil surface. One small jar(container) with a 3.7 cm radius and 4 cm depth was used forall soil sample extractions. Collected samples were sealed inplastic bags and stored in an icebox (2000) or analyzedimmediately (2001). Seed separation was comprised of fivesteps: 2-minute microwave drying, slow homogenization us-ing a hand eggbeater, gauze filtration and hand-picking ofseeds using tweezers and a magnifying glass. All collectionsand laboratory separation of seeds were performed exclu-sively by one person (T.K.).
3.1.5. Home ranges
Grey partridges captured in Stodulky region during Janu-ary–March 2002 were fitted with radio-transmitters (6.5 g,TW-4, Biotrack Ltd., UK) used in a previous study (Šálek etal., 2002). Beginning on the day of catching, radio-taggedbirds were monitored daily or every second day and some-
times twice a day (e.g. in the morning and afternoon). Thesepartridges were located by triangulation or direct observationand their positions were recorded onto maps including notesabout habitat and individual status.
3.2. Data analysis
3.2.1. Patches
We used the higher value of the two obtained as ourestimate of the number of grey partridge pairs in a patch(2001 or 2002). The differences in pair numbers withinparticular patches between the years 2001 and 2002 weretested using Spearman’s rank correlation coefficient (rs) andWilcoxon matched-pairs test. Numbers of pairs in individualpatches (explained variable) was fitted using a generalizedlinear model (GLIM) with Poisson’s distribution (referred toas GLIMpoiss) in which patch area, distance to the nearestmonitored patch (as a control for the potential effect ofneighboring patches), total area of patches available within a1 km radius, level of direct human disturbances, mean heightof herb vegetation and proportion of shrubs and trees repre-sented the potential explanatory factors. The resulting modelis based on stepwise selection of predictors, and on minimiz-ing theAkaike Information Criterion (AIC) statistic. TheAICwas estimated using a Cp statistic from the intermediateresults of the IRLS algorithm. The calculations were per-formed with S-Plus for Windows (S-Plus, 1999). Such anapproach was applied also in other GLIMs in this study.
The relationship between pair numbers and compositionof plant communities in patches was examined separatelyusing CANOCO (ter Braak, 1996) software. The reversecanonical correspondence analysis (CCA) with pair numbersas the environmental variable explaining patch plant compo-sition was applied. Patch area and available patch area withina 1 km radius significantly affecting pair numbers (see Sec-
Fig. 2. Map of the Stodulky study plot (SW Prague) showing borders of the monitored patches and adjacent agricultural landscape. Symbols (points andtriangles) represent grey partridge pairs and singles observed in the area during one counting (21stApril 2000) and document an enormous difference in densitiesbetween these two habitats. No trios occurred on this date of counting.
26 M. Šálek et al. / Acta Oecologica 25 (2004) 23–33

tion 4) were established as covariables. The significance ofthis relationship was evaluated by a Monte-Carlo permuta-tion test.
3.2.2. Agricultural landscape
The presence of grey partridges in individual quadrats(present in both years, present in 1 year or absent) was fittedin GLIMs with a binomial distribution (GLIMbinom). Region,total area of small weeded spots, total length of lines, propor-tion of constructions, proportion of forests and distance ofthe quadrat from the nearest patch were included as factors.For the purpose of highlighting the difference between quad-rats which were occupied in both years and quadrats occu-pied only in any 1 year, we omitted those in which at least onehabitat parameter changed from 2000 to 2001.
3.2.3. Food supply
Variability in seed abundance was analyzed using col-lected seed samples using the GLIMpoiss in which year,habitat and distances from the edge or from other habitatsrepresented explanatory factors.
3.2.4. Home ranges
From the total number of 37 radio-tracked grey partridges,only 13 pairs (12 females each with a tagged mate and onemale with an untagged female) provided sufficient data fromthe entire period defined for this analysis. In the CzechRepublic, this time period from 10th April to 30th June 2002should include egg formation, nest building, egg laying,incubation and rearing of small chicks (Hudec and Cerný,1977). All records of incubating females (i.e. repeatedrecords of nest positions) were excluded from the data set aswell as repeated (dependent) fixes obtained in short (fewhours) intervals within which the birds shifted no more thansome tens of meters. Also, outliers (i.e. 5% of the mostdistant locations representing one or two fixes in individualsets) were omitted to reduce the possibility that the pairswere already outside their home range following accidentalflushing by dogs or people immediately before our visit. Infact, such flushing was a common phenomenon in the Sto-dulky study plot. Disturbed birds often flew far away fromtheir stands to which they, however, returned after a shortwhile.
Home range sizes were calculated by the fixed kernelmethod using 95% area level and automatic least squarescross-validation (LSCV). We preferred this particularmethod from the convex polygons method especially be-cause of the limited (<50) number of locations for each pair(Seaman and Powell, 1996; Seaman et al., 1999). We com-bined the use of the Animal Movement software (Hooge andEichenlaub, 1997) and GIS ArcView in this analysis. Thehome range sizes (area) were fitted in a GLIM with gammadistribution (GLIMgamma) in which numbers of locations forindividual pairs were controlled.
4. Results
4.1. Patches
In 2001, we monitored 26 patches with a 1.83 km2 totalarea of which 21 (81%) were occupied by 68 grey partridgepairs. The mean population density per patch (inclusive ofunoccupied patches) varied widely around a mean of36.1(±S.D. = 41.4) pairs km–2, or 3.4 ha (±S.D. = 1.8) perpair (Fig. 3). Four patches were destroyed during the periodfrom 2001 to 2002 (2× building, 2× restored field work) sothat only 22 remaining patches were available for monitoringin 2002 of which 16 (73%) were occupied. We recorded59 pairs within the 1.61 km2 area giving a density of 37.1(±43.7) pairs km–2, or 3.1 (±2.5) ha per pair. Pair numbersobserved in 2002 were highly correlated with those from2001 (rs = 0.77, n = 22, P < 0.001) and no statisticallysignificant population change towards decrease or increasewas detected (change = –6.3%, Wilcoxon matched-pairs test,Z = –0.47, n = 22, P = 0.638).
Pair numbers in patches varied between 0 and 17 and werepositively correlated with patch area (correlation strengthr = 0.72, GLIMpoiss, F1,20 = 38.73, P < 0.0001) as well as withthe total area of other patches available within a 1 km radius(correlation strength r = 0.26, F1,19 = 6.07, P = 0.024). Thepatch area and the availability of other patches were notcorrelated (P = 0.698) but were in interaction (F1,19 = 34.21,P < 0.0001) indicating different trends in smaller and largerpatches: the effect of total area of other patches was signifi-cant only around patches larger than the mean (7.6 ha;P = 0.035) but not around smaller patches (P = 0.173).Vegetation height was found to be a significant factor with aunimodal pattern (F1,20 = 5.53, P = 0.030; Fig. 4). Distance tothe nearest monitored patch, proportion of shrubs and treesand human disturbances showed no significant influence onpair numbers within patches (all P > 0.2). The three signifi-cant factors mentioned above explained 71.8% of the totaldeviance. Note that the alternative model arising from themean number of pairs (2001 and 2002) provides similarresults and explains 67.7% of the total deviance (not pre-sented here).
Fig. 3. Frequency distribution of grey partridge densities within the patchesin 2001 and 2002 (total n = 48).
27M. Šálek et al. / Acta Oecologica 25 (2004) 23–33

In 60 samples of plant communities collected on22 patches, 168 plant species in total were recorded fromwhich only two (Elytrigia repens (L.) Desv. and Calama-grostis epigejos (L.) Roth) covered 48.4% of the total patcharea (Appendix). No relationship was found between pairnumbers and the specific composition of a plant communitywithin a patch (CCA: Monte-Carlo test, 499 permutations,F = 0.95, P = 0.504). Centroids of plant taxa recorded in thepast as partridge food supply in the Czech Republic (accord-ing to Sekera, 1956 and Janda, 1964) showed either lower fit(<10%; Centaurea, Geranium, Rumex, Sanguisorba, Stel-laria) or occupied central positions of the ordination space(Cerastium, Myosotis, Rubus, Vicia, Viola) indicating thatthey are widely distributed throughout communities ratherthan bounded with specific types of vegetation.
4.2. Agricultural landscape
The occurrence of grey partridges within quadrats in 2000and 2002 was found to be positively related primarily to thedistance of the nearest patch (correlation strength r = 0.35,GLIMbinom, F1,94 = 11.65, P < 0.001; Fig. 5) and in a lesserextent negatively to the proportion of present forests (corre-lation strength r = 0.23, F1,95 = 6.98, P < 0.010; Fig. 6). Theomission of all other factors (region, total area of smallweeded spots, total length of lines and proportion of con-structions) did not change the model significantly(P = 0.291). The two statistically significant factors (thedistance of the nearest patch and proportion of present for-ests) were not correlated (P = 0.248) and their interaction wasstrong (F1,101 = 21.10, P < 0.0001): the effect of forests wasnot confirmed close to a patch (<750 m; P = 0.137) unlike inmore distant quadrats (>750 m) where a higher forest propor-tion negatively correlated with the presence of grey par-tridges (P = 0.044).
The proportional occupancy of quadrats was strongly re-lated to the distance of the nearest patch. In 2000, occupancydecreased from 61% within the closest patches to 19% indistant locations ≥1200 m (v3
2 = 10.4, P = 0.016) while in
2001 it was 57% and 17% (v32 = 14.6, P = 0.002; Fig. 5)
respectively.The total area of small weeded spots was the most impor-
tant factor explaining the difference between the combined2-year and any 1-year presence of grey partridges in quadrats(GLIMbinom, F1,101 = 5.49, P = 0.023) while omission of allother factors did not affect the model substantially(P = 0.217; Fig. 7).
4.3. Stodulky region: population density
The density of pairs and trios between March and Mayvaried between 46.6 and 110.7 km–2 within studied patchesand was only 2.6–8.4 km–2 in the surrounding agriculturallandscape. After pooling the data from both years, the meandensity was 72.6 km–2 (±18.7, median 67.0 km–2) in a patch
0
20
40
60
80
100
< 50 51-65 66-80 > 80
Mean height of vegetation (cm)
Pai
rs 1
00 h
a-1
(4)
(5)
(8)
(5)
Fig. 4. Population density of grey partridges in patches in relation to meanheight of vegetation cover in a patch. The higher value of the 2 years (2001or 2002) obtained for each patch was used (n = 22). The standard errors areindicated. Numbers of patches are in parentheses.
Fig. 5. Proportional occupancy of 400 × 400 m quadrats by grey partridgesin relation to the distance of the nearest patch in 2000 (n = 116) and 2001(n = 119). The differences between years are not significant (GLMbinom:F1,233 = 3.099, P = 0.080).
Fig. 6. Proportional occupancy of quadrats by grey partridges in relation tothe proportion of forest. The decline of occupancy within afforested quadratswas significant only in quadrats positioned >750 m far of the nearest patch(see Section 4.2). Numbers of quadrats are in parentheses.
28 M. Šálek et al. / Acta Oecologica 25 (2004) 23–33
but 5.3 km–2 (±1.6, median 4.5 km–2) in agricultural land-scape. In order to obtain the actual nesting density and tominimize the negative effect of population numbers decreas-ing throughout the course of the season, data obtained onlybetween 20th April and 5th May were taken into account forfurther comparisons. A significant difference between thepatch (56.1 ± 6.9, median 56.8 pairs km–2 or 1.6–2.1 ha perpair) and the adjacent agricultural landscape (5.8 ± 1.7,median 5.8 pairs km–2 or 12.9–25.7 ha per pair) was found(Mann–Whitney test, U = 16, n1 =n2 = 4, P = 0.021; Fig. 8).
4.4. Stodulky region: food supply
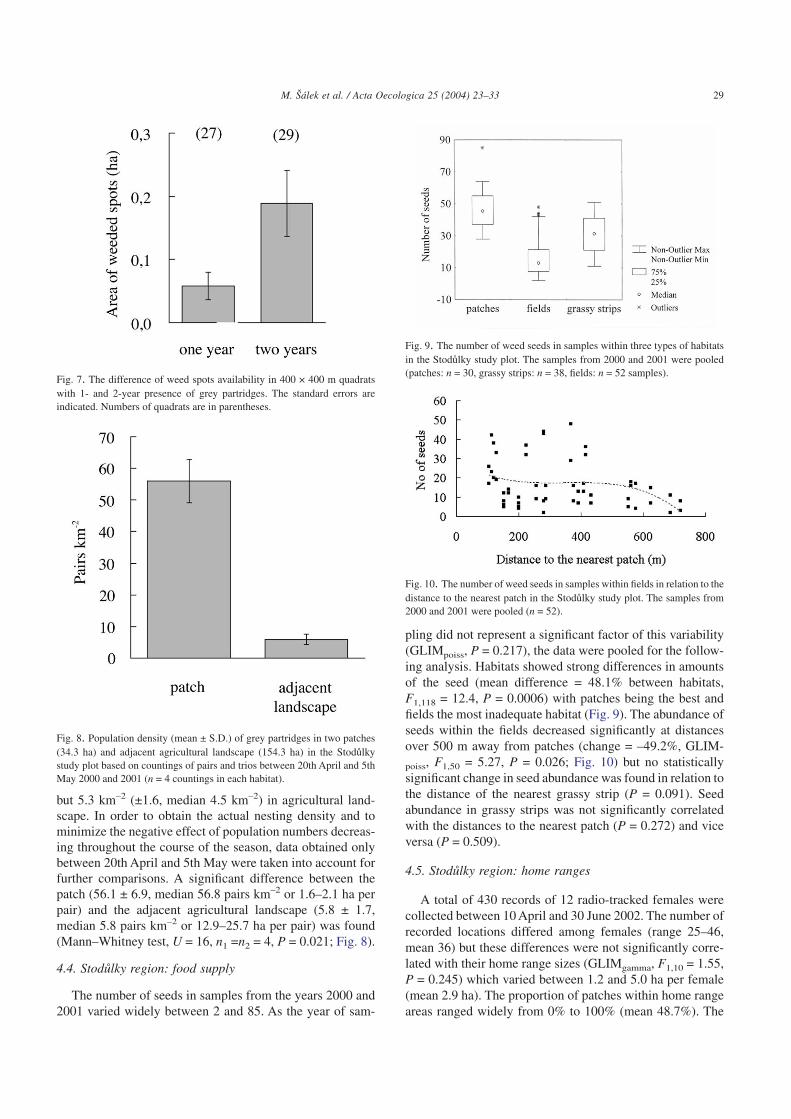
The number of seeds in samples from the years 2000 and2001 varied widely between 2 and 85. As the year of sam-
pling did not represent a significant factor of this variability(GLIMpoiss, P = 0.217), the data were pooled for the follow-ing analysis. Habitats showed strong differences in amountsof the seed (mean difference = 48.1% between habitats,F1,118 = 12.4, P = 0.0006) with patches being the best andfields the most inadequate habitat (Fig. 9). The abundance ofseeds within the fields decreased significantly at distancesover 500 m away from patches (change = –49.2%, GLIM-
poiss, F1,50 = 5.27, P = 0.026; Fig. 10) but no statisticallysignificant change in seed abundance was found in relation tothe distance of the nearest grassy strip (P = 0.091). Seedabundance in grassy strips was not significantly correlatedwith the distances to the nearest patch (P = 0.272) and viceversa (P = 0.509).
4.5. Stodulky region: home ranges
A total of 430 records of 12 radio-tracked females werecollected between 10 April and 30 June 2002. The number ofrecorded locations differed among females (range 25–46,mean 36) but these differences were not significantly corre-lated with their home range sizes (GLIMgamma, F1,10 = 1.55,P = 0.245) which varied between 1.2 and 5.0 ha per female(mean 2.9 ha). The proportion of patches within home rangeareas ranged widely from 0% to 100% (mean 48.7%). The
Fig. 7. The difference of weed spots availability in 400 × 400 m quadratswith 1- and 2-year presence of grey partridges. The standard errors areindicated. Numbers of quadrats are in parentheses.
Fig. 8. Population density (mean ± S.D.) of grey partridges in two patches(34.3 ha) and adjacent agricultural landscape (154.3 ha) in the Stodulkystudy plot based on countings of pairs and trios between 20th April and 5thMay 2000 and 2001 (n = 4 countings in each habitat).
Fig. 9. The number of weed seeds in samples within three types of habitatsin the Stodulky study plot. The samples from 2000 and 2001 were pooled(patches: n = 30, grassy strips: n = 38, fields: n = 52 samples).
Fig. 10. The number of weed seeds in samples within fields in relation to thedistance to the nearest patch in the Stodulky study plot. The samples from2000 and 2001 were pooled (n = 52).
29M. Šálek et al. / Acta Oecologica 25 (2004) 23–33
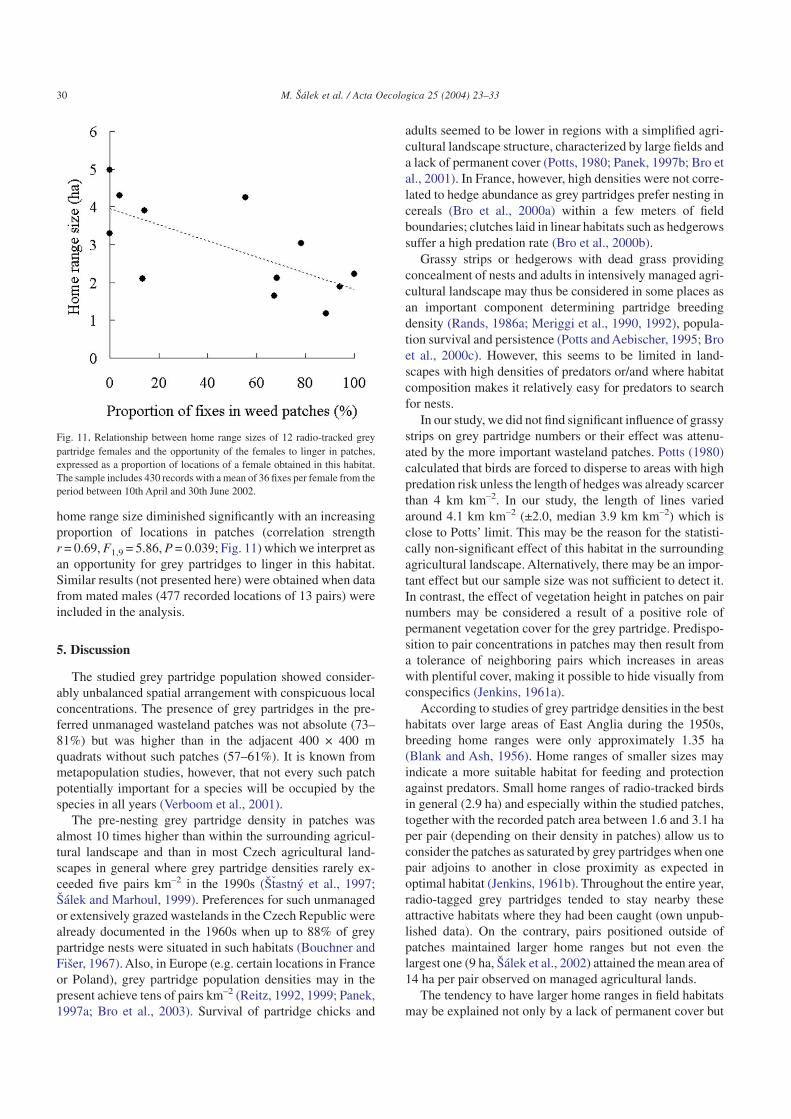
home range size diminished significantly with an increasingproportion of locations in patches (correlation strengthr = 0.69, F1,9 = 5.86, P = 0.039; Fig. 11) which we interpret asan opportunity for grey partridges to linger in this habitat.Similar results (not presented here) were obtained when datafrom mated males (477 recorded locations of 13 pairs) wereincluded in the analysis.
5. Discussion
The studied grey partridge population showed consider-ably unbalanced spatial arrangement with conspicuous localconcentrations. The presence of grey partridges in the pre-ferred unmanaged wasteland patches was not absolute (73–81%) but was higher than in the adjacent 400 × 400 mquadrats without such patches (57–61%). It is known frommetapopulation studies, however, that not every such patchpotentially important for a species will be occupied by thespecies in all years (Verboom et al., 2001).
The pre-nesting grey partridge density in patches wasalmost 10 times higher than within the surrounding agricul-tural landscape and than in most Czech agricultural land-scapes in general where grey partridge densities rarely ex-ceeded five pairs km–2 in the 1990s (Št{astný et al., 1997;Šálek and Marhoul, 1999). Preferences for such unmanagedor extensively grazed wastelands in the Czech Republic werealready documented in the 1960s when up to 88% of greypartridge nests were situated in such habitats (Bouchner andFišer, 1967). Also, in Europe (e.g. certain locations in Franceor Poland), grey partridge population densities may in thepresent achieve tens of pairs km–2 (Reitz, 1992, 1999; Panek,1997a; Bro et al., 2003). Survival of partridge chicks and
adults seemed to be lower in regions with a simplified agri-cultural landscape structure, characterized by large fields anda lack of permanent cover (Potts, 1980; Panek, 1997b; Bro etal., 2001). In France, however, high densities were not corre-lated to hedge abundance as grey partridges prefer nesting incereals (Bro et al., 2000a) within a few meters of fieldboundaries; clutches laid in linear habitats such as hedgerowssuffer a high predation rate (Bro et al., 2000b).
Grassy strips or hedgerows with dead grass providingconcealment of nests and adults in intensively managed agri-cultural landscape may thus be considered in some places asan important component determining partridge breedingdensity (Rands, 1986a; Meriggi et al., 1990, 1992), popula-tion survival and persistence (Potts and Aebischer, 1995; Broet al., 2000c). However, this seems to be limited in land-scapes with high densities of predators or/and where habitatcomposition makes it relatively easy for predators to searchfor nests.
In our study, we did not find significant influence of grassystrips on grey partridge numbers or their effect was attenu-ated by the more important wasteland patches. Potts (1980)calculated that birds are forced to disperse to areas with highpredation risk unless the length of hedges was already scarcerthan 4 km km–2. In our study, the length of lines variedaround 4.1 km km–2 (±2.0, median 3.9 km km–2) which isclose to Potts’ limit. This may be the reason for the statisti-cally non-significant effect of this habitat in the surroundingagricultural landscape. Alternatively, there may be an impor-tant effect but our sample size was not sufficient to detect it.In contrast, the effect of vegetation height in patches on pairnumbers may be considered a result of a positive role ofpermanent vegetation cover for the grey partridge. Predispo-sition to pair concentrations in patches may then result froma tolerance of neighboring pairs which increases in areaswith plentiful cover, making it possible to hide visually fromconspecifics (Jenkins, 1961a).
According to studies of grey partridge densities in the besthabitats over large areas of East Anglia during the 1950s,breeding home ranges were only approximately 1.35 ha(Blank and Ash, 1956). Home ranges of smaller sizes mayindicate a more suitable habitat for feeding and protectionagainst predators. Small home ranges of radio-tracked birdsin general (2.9 ha) and especially within the studied patches,together with the recorded patch area between 1.6 and 3.1 haper pair (depending on their density in patches) allow us toconsider the patches as saturated by grey partridges when onepair adjoins to another in close proximity as expected inoptimal habitat (Jenkins, 1961b). Throughout the entire year,radio-tagged grey partridges tended to stay nearby theseattractive habitats where they had been caught (own unpub-lished data). On the contrary, pairs positioned outside ofpatches maintained larger home ranges but not even thelargest one (9 ha, Šálek et al., 2002) attained the mean area of14 ha per pair observed on managed agricultural lands.
The tendency to have larger home ranges in field habitatsmay be explained not only by a lack of permanent cover but
Fig. 11. Relationship between home range sizes of 12 radio-tracked greypartridge females and the opportunity of the females to linger in patches,expressed as a proportion of locations of a female obtained in this habitat.The sample includes 430 records with a mean of 36 fixes per female from theperiod between 10th April and 30th June 2002.
30 M. Šálek et al. / Acta Oecologica 25 (2004) 23–33

also by limited food availability. In spite of the fact that mostof the grey partridge diet represents corn grain (Pulliainen,1984), about one-third of the dry mass of food from latesummer to spring consists of weed seeds (Potts, 1970). Thisfood component was more abundant in patches than in grassylines and fields. As these patches represent vegetation in itsearly stages of plant succession, they are weed-enriched andhave higher seed production while grassy strips are markedlydominated by grasses with lower production of seed (Grime,1979). Only older patches tend to lose weed species as aresult of the expansion of dominant grasses (e.g. C. epigejosand E. repens) which may potentially result in a successiveloss of their attraction for partridges.
In our study, however, we did not detect any preference oravoidance of any type of wasteland patch by grey partridges(e.g. in relation to the proportions of grasses or plant speciesconsidered as a food component). In relation to the greypartridge habitat requirements, food sources and the struc-ture of vegetation cover probably were of similar qualityamong patches over the whole region or, at least, there was nostatistically significant evidence of preferences in oursamples. As noted by Potts (1970), it is the distribution offood, rather than its quality that is important during theestablishment of grey partridge territories in spring. The foodmay vary among regions and years with respect to foodsupply induced by, among others, changes in crop rotationand use of pesticides (Potts, 1970; Pulliainen, 1984). Forexample, dicotyledon weed seeds of the Galeopsis genuswere an important fraction of the grey partridges’ food inFinland (Pulliainen, 1984). Species of the Polygonum genuswere found to be substantial in West Sussex, Britain (Potts,1970). They were not recorded in our patches at all.
Patches unlike fields and narrow grassy strips suffer onlyto a limited extent from the destructive impact of pesticides.Previous studies in France and the UK showed that herbi-cides and insecticides sprayed on crops had a strong effect oninsect abundance (e.g. Serre and Birkan, 1985) and thatpartridge covey size was higher on unsprayed areas (e.g.Rands, 1986b; Sotherton and Robertson, 1990). Thus numer-ous insects may be confined to the patches resulting in highersurvival chances for chicks, a decisive factor affecting greypartridge densities (Rands, 1985; Potts, 1986). Higher num-bers of insects preferred by grey partridge chicks were foundin permanent vegetation than in crops (Southwood andCross, 1969). Moreover, Green’s (1984) study has revealedthat grey partridge broods forage preferably in the edges ofcereal fields where arthropods and weeds are most abundant.
Numbers of pairs present in patches were determinedprimarily by the patch area while the total areas of patchesavailable within a 1 km radius only had this effect whenassociated with a large monitored patch (>7.5 ha). In con-trast, the influences of disturbance rate and vegetation struc-ture were weak with an exception of mean height of herblayer. Groups of larger patches may have a higher return rateof the previous year’s nesters and/or there may be a greatersurvival rate from 1 year to the next based on probability
alone (Helzer and Jelinski, 1999). In a similar manner theavailability of small weed spots may have a positive effect onthe repeated presence of dispersed pairs in subsequent yearsoutside the patches. The presence of grey partridges rapidlydecreased with the distance of the nearest patch and thisradial distribution seemed to be in agreement with a simplereaction–diffusion model of the grey partridge population(e.g. Flather and Bevers, 2002). Only the proportion of for-ests was found to have a negative influence on the presence ofgrey partridges in distances >750 m of the nearest patches. Inother words, small and isolated patches of any plant compo-sition anywhere within the studied agricultural landscapecannot be of the same quality as clusters of large patchessituated nearby, even though they are supported by grassylines or by the absence of human constructions. It still re-mains to be examined, however, whether the patches mayfunction as true sources in a source–sink metapopulationmodel or if they are subsidized by birds from the surroundinglandscape and the population tends towards balanced dis-persal (Diffendorfer, 1998) even though with an uneven spa-tial pattern. Confirming or rejecting such a model will requirecomparisons of natality, mortality and dispersal of popula-tions within and outside the patches.
6. Implications for management
Wasteland patches are highly valuable for grey partridgeconservation but only once they begin to be dominated by anoverly high vegetation (grasses, shrubs). If a seed-bank,optimal herb cover and wild partridge populations werepresent, the patches did not require any other specific man-agement or, as r-selective habitats, any protection againstdisturbances such as human activity nearby the settlements.Moreover, they may act as refuges for other taxa (weedplants, insects, birds) inhabiting specific habitats in the earlystage of succession and enhance, at least provisionally, spe-cies diversity within the landscape.
The persistence of sufficiently large and grouped waste-land patches passing into agricultural land may sustain thegrey partridge population at a high density and low dispersalwithout the need of support from reared birds released fromcaptivity. A surplus of offspring may become a potentialsource of a breeding population in the surrounding sub-optimal agricultural landscape. These habitats are, in a certainway, analogous to abandoned fields that are dominated by tallgrasses. The presence of weed seeds (and probably also asufficient amount of insects) may enhance the carrying capac-ity for the grey partridge especially during the winter season,pre-nesting period (and chick rearing) more than linear grassyhabitats or hedgerows but may cause problems with farmers.On the other hand, the patches are highly fragmented and mayact as an ecological trap in which birds generally inhabitingattractive habitats suffer from high predation. Moreover, thesehabitats are unstable because they naturally evolve to shrubvegetation or uniform and continuous covers of bushgrass (C.
31M. Šálek et al. / Acta Oecologica 25 (2004) 23–33

epigejos), which consequently blocks the succession, bothbeing unsuitable to the former bird assemblage. However, asthis habitat is not primarily predestined to wildlife in the citydevelopment scheme, successive build-up or older unsuitablepatches may be adequately replaced by new sets of aban-doned or set-aside fields.
Acknowledgements
We thank plant ecologist Bohumil Mandák for the collec-tion and determination of plant species. We are also gratefulto Petr Šmilauer who provided us with necessary statisticaladvise and Michal Tomášek for assistance in GIS procedures.Hana Rychlíková, Mark Burgman and two anonymous refer-ees reviewed earlier versions of the manuscript and providedvaluable comments. The study was financially supported bythe Grant Agency of the Czech Republic (No 206/02/1392).
Appendix
List of plant species occurring in wasteland communitiesin the studied patches. Species present on at least 50% of thesites are only included. No species with a coverage >1% wasomitted. The species are ordered according to total coverage.
Species Presence(%)
Total coverage(%)
Elymus repens (L.) Desv. 95,45 26,07Calamagrostis epigejos (L.) Roth 100,00 22,29Taraxacum sect. Ruderalia 90,91 6,87Epilobium spp. 72,73 4,97Trifolium pratense L. 81,82 3,27Urtica dioica L. 81,82 2,74Cirsium arvense (L.) Scop. 86,36 2,10Poa pratensis L. 68,18 1,70Dactylis glomerata L. 59,09 1,62Tanacetum vulgare L. 54,55 1,54Tussilago farfara L. 72,73 1,40Solidago gigantea Aiton 63,64 1,19Artemisia vulgaris L. 77,27 1,18Daucus carota L. 77,27 1,00Hypericum perforatum L. 77,27 0,96Achillea millefolium L. 68,18 0,93Tripleurospermum inodorum (L.) Schul 50,00 0,74Cirsium vulgare (Savi) Ten 77,27 0,73Dipsacus fullonum L. 81,82 0,69Potentilla reptans L. 54,55 0,52Solidago canadensis L. 77,27 0,49Rumex crispus L. 50,00 0,45Rumex obtusifolius L. 50,00 0,42Geum urbanum L. 68,18 0,37Plantago lanceolata L. 50,00 0,37Pastinaca sativa L. 59,09 0,37Galium album s.str. Miller 50,00 0,36Cardaria draba (L.) Desv 59,09 0,35Cerastium holosteoides Fries ampl. Hyl. 54,55 0,30Lamium album L. 50,00 0,29Potentilla anserina L. 50,00 0,15
References
Aebischer, N., Kavanagh, B., 1997. Grey partridge. In: Hagemeijer, W.J.M.,Blair, M.J. (Eds.), The EBCC Atlas of European Breeding Birds, theirDistribution and Abundance. T&AD Poyser, London, pp. 212–213.
Blank, T.H., Ash, J.S., 1956. The concept of territory in the partridge (Perdixperdix). Ibis 98, 379–389.
Blank, T.H., Southwood, T.R.E., Cross, D.J., 1967. The ecology of thepartridge: I. Outline of population processes with particular reference tochick mortality and nest density. Journal of Animal Ecology 36, 549–556.
Bouchner, M., Fišer, Z., 1967. Príspevek ke hnízdní bionomii koroptví a keztrátám na hnízdech a vejcích. Lesnický casopis 13, 713–725.
Bro, E., Deldalle, B., Massot, M., Reitz, F., Selmi, S., 2003. Density-dependence of reproductive success in grey partridge populations inFrance, management implications. Wildlife Biology 9, 93–102.
Bro, E., Reitz, F., Clobert, J., 2000a. Nest-site selection of grey partridge(Perdix perdix) on agricultural lands in north-central France. Game andWildlife Science 17, 1–16.
Bro, E., Reitz, F., Clobert, J., Mayot, P., 2000b. Nesting success of greypartridges (Perdix perdix) on agricultural land in north-central France,relation to nesting cover and predator abundance. Game and WildlifeScience 17, 199–218.
Bro, E., Sarrazin, F., Clobert, J., Reitz, F., 2000c. Demography and thedecline of the grey partridge Perdix perdix in France. Journal of AppliedEcology 37, 432–448.
Bro, E., Reitz, F., Clobert, J., Migot, P., Massot, M., 2001. Diagnosing theenvironmental causes of the decline in grey partridge Perdix perdixsurvival in France. Ibis 143, 120–132.
Diffendorfer, J.E., 1998. Testing models of source–sink dynamics and bal-anced dispersal. Oikos 81, 417–433.
Flather, C.H., Bevers, M., 2002. Patchy reaction–diffusion and populationabundance: the relative importance of habitat amount and arrangement.American Naturalist 159, 40–56.
Fuchs, R., Škopek, J., Formánek, J., Exnerová, A., 2003. Atlas hnízdníhorozšírení ptáku Prahy. Consult, Praha.
Green, R.E., 1984. The feeding ecology and survival of partridge chicks(Alectoris rufa and Perdix perdix) on arable farmland in East Anglia,UK. Journal of Applied Ecology 21, 817–830.
Grime, J.P., 1979. Plant Strategies and Vegetation Processes. Wiley, Chich-ester, New York, Brisbane, Toronto.
Helzer, C.J., Jelinski, D.E., 1999. The relative importance of patch area andperimeter-area ratio to grassland breeding birds. Ecological Applications9, 1448–1458.
Hooge, P.N., Eichenlaub, B., 1997. Animal Movement Extension to Arc-view, ver. 2.0. Alaska Science Center, Biological Science Office, USGeological Survey, Anchorage.
Hudec, K., Cerný, W. (Eds.), 1977. Fauna CSSR, Ptáci II. Academia, Praha.Janda, J., 1964. Potravní nároky mladých koroptví a bažantu. Lesnický
casopis 6, 580–597.Jenkins, D., 1961a. Social behaviour in the partridge (Perdix perdix). Ibis
103a, 155–188.Jenkins, D., 1961b. Population control in protected partridges (Perdix
perdix). Journal of Animal Ecology 30, 235–258.Meriggi, A., Montagna, D., Zacchetti, D., Matteucci, C., Toso, S., 1990.
Population dynamics of the gray partridge in relation to agriculture andweather in northern Italy. In: Church, K.E., Warner, R.E., Brady, S.J.(Eds.), Perdix V: Gray Partridge and Ring-necked Pheasant Workshop.Mankato, Minnesota, pp. 241–256.
Meriggi, A., Saino, N., Montagna, D., Zacchetti, D., 1992. Influence ofhabitat on density and breeding success of grey and red-legged par-tridges. Bolletino di Zoologia 59, 289–295.
Panek, M., 1997a. Density-dependent brood production in the grey partridgePerdix perdix in relation to habitat quality. Bird Study 44, 235–238.
Panek, M., 1997b. The effect of agricultural landscape structure on foodresources and survival of grey partridge Perdix perdix chicks in Poland.Journal of Applied Ecology 34, 787–792.
32 M. Šálek et al. / Acta Oecologica 25 (2004) 23–33

Potts, G.R., 1970. Studies on the changing role of weeds of the genusPolygonum in the diet of the Partridge Perdix perdix L. Journal ofAnimal Ecology 7, 567–576.
Potts, G.R., 1980. The effects of modern agriculture, nest predation andgame management on the population ecology of partridges (Perdixperdix and Alectoris rufa). Advances in Ecological Research 11, 2–79.
Potts, G.R., 1986. The Partridge: Pesticides, Predation and Conservation.Collins, London.
Potts, G.R., Aebischer, N.J., 1995. Population dynamics of the grey par-tridge Perdix perdix 1793–1993: monitoring, modelling and manage-ment. Ibis 137 (supplement), 29–37.
Pulliainen, E., 1984. Changes in the composition of the autumn food ofPerdix perdix in west Finland over 20 years. Journal of Applied Ecology21, 133–139.
Rands, M.R.W., 1985. Pesticide use on cereals and the survival of partridgechicks: a field experiment. Journal of Applied Ecology 22, 49–54.
Rands, M.R.W., 1986a. Effect of hedgerow characteristics on partridgebreeding densities. Journal of Applied Ecology 23, 479–487.
Rands, M.R.W., 1986b. The survival of gamebird (Galliformes) chicks inrelation to pesticide use on cereals. Ibis 128, 57–64.
Reitz, F., 1992. Adult survival and reproductive success in abundant popu-lations of grey partridge (Perdix perdix) in north central France. In:Birkan, M., Potts, G.R., Aebischer, N.J., Dowell, S.D. (Eds.), Perdix VI:First International Symposium on Partridges, Quails and Francolins.Gib. Faune Sauv., 9, pp. 313–324.
Reitz, F., 1999. The status of partridges in north-central France. Proceedingsof Perdix VIII, Third International Symposium on Partridges, Quails andPheasants in the Western Paleartic and Neartic (26–29 October 1998,Sopron, Hungary). Hungarian Small Game Bulletin, 5, pp. 151–163.
Seaman, D.E., Powell, R.A., 1996. An evaluation of the accuracy of kerneldensity estimators for home range analysis. Ecology 77, 2075–2085.
Seaman, D.E., Millspaugh, J.J., Kernohan, B.J., Brundige, G.C.,Raedeke, K.J., Gitzen, R.A., 1999. Effects of sample size on kernel homerange estimates. Journal of Wildlife Management 63, 739–747.
Sekera, J., 1956. Chov koroptví. Státní zemedelské nakladatelství, Praha.Serre, D., Birkan, M., 1985. Incidence de traitement insecticides sur les
ressources alimentaires de poussins de perdrix grise. Gib. Faune Sauv. 4,21–61.
Sotherton, N.W., Robertson, P.A., 1990. Indirect impacts of pesticides on theproduction of wild gamebirds in Britain. In: Church, K.E., Warner, R.E.,Brady, S.J. (Eds.), Perdix V: Gray Partridge and Ring-necked PheasantWorkshop. Kans. Dep. Wildl. and Parks, Emporia, pp. 84–102.
Southwood, T.R.E., Cross, D.J., 1969. The ecology of the partridge. III.Breeding success and the abundance of insects in natural habitats. Jour-nal of Animal Ecology 38, 497–509.
S-Plus, 1999. S-Plus for Windows 2000: Guide to Statistics. Data AnalysisProducts Division, MathSoft, Seattle.
Šálek, M., Marhoul, P., 1999. Sezónní dynamika a príciny ztrát koroptvepolní (Perdix perdix): výsledky scítání a telemetrického sledování vletech 1997–1999. Sylvia 35, 55–67.
Šálek, M., Marhoul, P., Pintír, J., 2002. Spring to autumn home range andhabitat use of a high density population of the grey partridge (Perdixperdix) in Prague, Czech Republic. Folia Zoologica 51, 299–306.
Št{astný, K., Bejcek, V., Hudec, K., 1997. Atlas hnízdního rozšírení ptáku vCeské republice 1985–1989. H&H, Jinocany.
ter Braak, C.J.F., 1996. Unimodal Models to Relate Species to Environment.DLO-Agricultural Mathematics Group, Wageningen.
Verboom, J., Foppen, R., Chardon, P., Opdam, P., Luttikhuizen, P., 2001.Introducing the key patch approach for habitat networks with persistentpopulations: an example for marshland birds. Biological Conservation100, 89–101.
33M. Šálek et al. / Acta Oecologica 25 (2004) 23–33
Copyright © 2022 FDOKUMEN




















