
Regulatory interdependence of myeloid transcription factors ...
Upload
independentCategory
view
2download
0
IL-12 and type I IFN response of neonatal myeloid DCto human CMV infection
Joelle Renneson1, Binita Dutta1, Stanislas Goriely1, Benedicte Danis1,
Sandra Lecomte1, Jean-Franc-ois Laes2, Zsuzsanna Tabi3,
Michel Goldman1 and Arnaud Marchant1
1 Institute for Medical Immunology, Universite Libre de Bruxelles, Charleroi, Belgium2 DNAVision SA, Charleroi, Belgium3 Department of Oncology and Palliative Medicine, Velindre Hospital, Whitchurch, Cardiff, UK
Following congenital human CMV (HCMV) infection, 15–20% of infected newborns develop
severe health problems whereas infection in immunocompetent adults rarely causes
illness. The immaturity of neonatal antigen presenting cells could play a pivotal role in
this susceptibility. Neonatal myeloid DC were shown to be deficient in IFN-b and IL-12
synthesis in response to TLR triggering. We studied the response of cord and adult blood-
derived myeloid DC to HCMV infection. Neonatal and adult DC were equally susceptible to
in vitro HCMV infection. Among immunomodulatory cytokines, IL-12, IFN-b and IFN-k1
were produced at lower levels by neonatal as compared with adult DC. In contrast,
neonatal and adult DC produced similar levels of IFN-a and IFN-inducible genes. Micro-
array analysis indicated that among the more than thousand genes up- or down-regulated
by HCMV infection of myeloid DC, 88 were differently regulated between adult and
neonatal DC. We conclude that neonatal and adult DC trigger a partly different response to
HCMV infection. The deficient IL-12 and mature IFN-a production by neonatal DC exposed
to HCMV are likely to influence the quality of the T lymphocyte response to HCMV
infection in early life.
Key words: Cytokines . DC . IFN . Infectious diseases . Neonate immunity
Supporting Information available online
Introduction
Neonates are highly susceptible to viral infection, as a result of the
immaturity of their immune system [1]. Deficiencies of both innate
and adaptive immune responses contribute to the impaired
neonatal host defense [2, 3]. Human CMV (HCMV) is the most
common cause of congenital infection, affecting between 5 and 20
newborns per thousand births worldwide [4]. Although primary
infection in immunocompetent adults rarely causes serious illness,
HCMV is responsible of severe health problems (deafness,
psychomotor retardation and death) in 10–20% of infected
neonates [5, 6]. Following primary infection, newborns are able
to develop a mature and functional CD81 T-cell response to HCMV
[7]. In contrast, anti-HCMV CD41 T-lymphocyte response is
defective in early life [8, 9]. This defect could be involved in the
prolonged viruria observed in young children [10].
DC play a pivotal role in the generation of antiviral response
and many viruses have evolved to interfere with DC functions
[11]. Myeloid cells are important targets for HCMV infection and
replication. The permissiveness of cells is related to their state of
differentiation, monocytes are non-permissive whereas differ-
entiated macrophages and immature DC support productiveCorrespondence: Dr. Arnaud Marchante-mail: [email protected]
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
Eur. J. Immunol. 2009. 39: 2789–2799 DOI 10.1002/eji.200939414 Immunity to infection 2789
infection [5]. Cytokines and chemokines are critical for the host
to mount an effective antiviral immune response. Two essential
immunomodulatory cytokines can drive the differentiation of
T cells: IL-12 and type I IFN. IL-12 has a central role in the
generation of Th-1 responses [12], acting as a potent inducer of
IFN-g from T cells [13, 14]. Type I IFN (IFN-a/b) are a family of
cytokines specialized in coordinating immunity to viruses, both
directly and indirectly [15]. The cross-linking of type I IFN
receptor stimulates the expression of hundreds of IFN-stimulated
genes (ISG) promoting an antiviral state [16]. IFN-a/b induces a
program that limits viral spreading and activates cytotoxic
response against infected cells. Type III IFN (IFN-l or IL-28/29)
are novel antiviral cytokines that display very similar activities to
type I IFN [17], although they act through a distinct receptor
complex [18, 19]. Several studies have shown that the production
of IL-12 p70 and IFN-b by neonatal monocyte-derived DC
(moDC) stimulated with the TLR-4 ligand LPS is lower than that
of adult moDC [20, 21]. The defective production of IL-12 p70
was shown to involve a lower transcription of the IL-12 p35
subunit [21]. The capacity of neonatal DC to produce T-cell
immunoregulatory cytokines in response to viral infection has not
been characterized. The immaturity of neonatal DC could play an
important role in the susceptibility of young infant to HCMV and
other viral infections [22].
The aim of this study was to compare the immunomodulatory
cytokine response of cord and adult blood-derived DC to infection
with a clinical isolate of HCMV (TB40/E strain). We observed that
infected neonatal moDC produced lower concentrations of IL-12,
IFN-b and IFN-l1 as compared with adult moDC. In contrast,
neonatal and adult moDC produce high and similar concentra-
tions of IFN-a and express similar levels of IFN-stimulated/
inducible genes (ISG).
Results
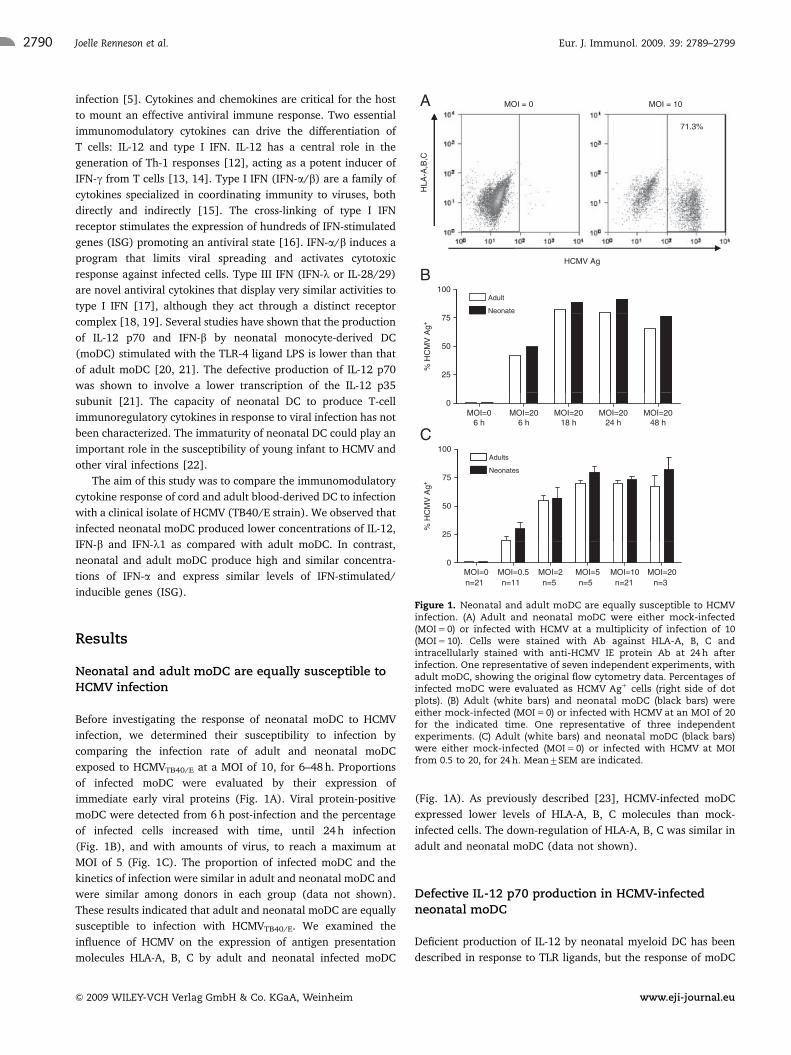
Neonatal and adult moDC are equally susceptible toHCMV infection
Before investigating the response of neonatal moDC to HCMV
infection, we determined their susceptibility to infection by
comparing the infection rate of adult and neonatal moDC
exposed to HCMVTB40/E at a MOI of 10, for 6–48 h. Proportions
of infected moDC were evaluated by their expression of
immediate early viral proteins (Fig. 1A). Viral protein-positive
moDC were detected from 6 h post-infection and the percentage
of infected cells increased with time, until 24 h infection
(Fig. 1B), and with amounts of virus, to reach a maximum at
MOI of 5 (Fig. 1C). The proportion of infected moDC and the
kinetics of infection were similar in adult and neonatal moDC and
were similar among donors in each group (data not shown).
These results indicated that adult and neonatal moDC are equally
susceptible to infection with HCMVTB40/E. We examined the
influence of HCMV on the expression of antigen presentation
molecules HLA-A, B, C by adult and neonatal infected moDC
(Fig. 1A). As previously described [23], HCMV-infected moDC
expressed lower levels of HLA-A, B, C molecules than mock-
infected cells. The down-regulation of HLA-A, B, C was similar in
adult and neonatal moDC (data not shown).
Defective IL-12 p70 production in HCMV-infectedneonatal moDC
Deficient production of IL-12 by neonatal myeloid DC has been
described in response to TLR ligands, but the response of moDC
71.3%
MOI = 0
HLA
-A,B
,C
100Adult
HCMV Ag
25
50
75Neonate
% H
CM
V A
g+%
HC
MV
Ag+
100
MOI=06 h
MOI=206 h
MOI=2018 h
MOI=2024 h
MOI=2048 h
0
25
50
75
Adults
Neonates
0MOI=0 MOI=0.5 MOI=2 MOI=5 MOI=10 MOI=20n=21 n=11 n=5 n=5 n=21 n=3
MOI = 10A
B
C
Figure 1. Neonatal and adult moDC are equally susceptible to HCMVinfection. (A) Adult and neonatal moDC were either mock-infected(MOI 5 0) or infected with HCMV at a multiplicity of infection of 10(MOI 5 10). Cells were stained with Ab against HLA-A, B, C andintracellularly stained with anti-HCMV IE protein Ab at 24 h afterinfection. One representative of seven independent experiments, withadult moDC, showing the original flow cytometry data. Percentages ofinfected moDC were evaluated as HCMV Ag1 cells (right side of dotplots). (B) Adult (white bars) and neonatal moDC (black bars) wereeither mock-infected (MOI 5 0) or infected with HCMV at an MOI of 20for the indicated time. One representative of three independentexperiments. (C) Adult (white bars) and neonatal moDC (black bars)were either mock-infected (MOI 5 0) or infected with HCMV at MOIfrom 0.5 to 20, for 24 h. Mean7SEM are indicated.
Eur. J. Immunol. 2009. 39: 2789–2799Joelle Renneson et al.2790
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

to viral infection has not been reported. We measured the
secretion of IL-12 p70 and p40 in supernatants 24 h after HCMV
infection of adult and neonatal moDC, at two different MOI (0.5
or 10 plaque forming unit (PFU)/cell). We observed a significant
production of those cytokines following HCMV infection of moDC
(adult mean: 12.15 pg/mL for MOI 0.5, 85.23 pg/mL for MOI 10)
(Fig. 2A and B). Neonatal moDC infected with HCMV at an MOI
of 10 produced significantly lower concentrations of IL-12 p70
than adult moDC. As observed in response to TLR ligation, we
observed at the mRNA level that neonatal moDC infected with
HCMV produced lower levels of IL-12 p35 subunit mRNA and
similar levels of IL-12 p40 subunit mRNA as compared with adult
moDC. These results show that the deficient IL-12 p70 production
by neonatal myeloid DC activated by TLR ligation is also observed
in response to a complex viral infection.
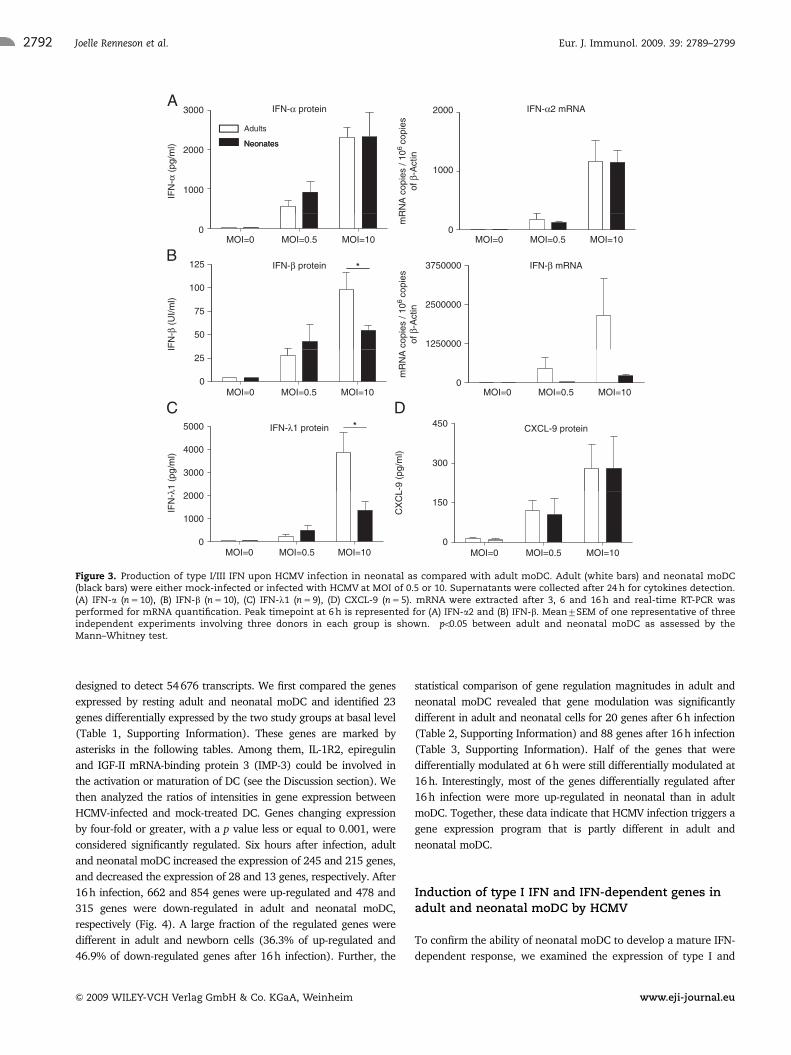
Mature IFN-a and defective IFN-b and IFN-k1production by HCMV-infected neonatal moDC
We then analyzed the production of type I/III IFN that are essential
cytokines for antiviral responses. Previous studies have shown that
neonatal moDC stimulated by TLR ligands produce lower concen-
trations of IFN-b than adult moDC [20]. We observed that HCMV
infection induced the production of high concentrations of IFN-a by
adult moDC even at a low MOI (Fig. 3A). Similar concentrations
were produced by neonatal and adult moDC. Quantitative RT-PCR
confirmed that the IFN-a2 gene was similarly expressed upon
infection by neonatal and adult moDC. IFN-b and IFN-l1 proteins
were also detected in infected adult moDC supernatants, with
higher type I IFN concentrations being induced by increasing MOI
(Fig. 3B and C). In contrast, neonatal moDC infected with HCMV at
an MOI of 10 showed lower production of the two early IFN as
compared with adult cells. The low IFN-b production was associated
with a low accumulation of IFN-b mRNA in neonatal moDC. We
then measured the production of CXCL-9, a typical IFN-inducible
chemokine. As shown in Fig. 3D, the induction of CXCL-9 secretion
was similar in adult and neonatal infected moDC. These experi-
ments indicate that although neonatal moDC produce lower
concentrations of IFN-b and IFN-l1 than adult cells, they develop
mature IFN-a and IFN-dependent responses to HCMV infection.
Gene expression analysis of HCMV-infected adultand neonatal moDC
In order to further compare the response of adult and neonatal
moDC to HCMV infection, we analyzed their transcriptome by
oligonucleotide microarrays. Adult and neonatal moDC were either
mock-infected or infected with HCMV at an MOI of 10 for 6 or 16 h
and total RNA was harvested. The time points were chosen to
include an early and a late time period in the first phase of HCMV
life cycle associated with the expression of immediate-early genes
(Fig. 1 and [24]). RNA samples were hybridized to microarrays
IL-12 p70 protein125
IL-12 p35 mRNA30000
50
75
100
∗∗
10000
20000
Adults
Neonates
MOI=0 MOI=0.5 MOI=100
25IL-1
2 p7
0 (p
g/m
l)
MOI=0
0
mR
NA
cop
ies
/ 106
copi
es o
fβ-
Act
in
mR
NA
cop
ies
/ 106
copi
es o
fβ-
Act
in
IL-12 p40 protein
1000
1500IL-12 p40 mRNA
8000
10000
MOI=0 MOI=0.5 MOI=100
500
IL-1
2 p4
0 (p
g/m
l)
MOI=0 MOI=0.5 MOI=100
2000
4000
6000
MOI=0.5 MOI=10
A
B
Figure 2. IL-12 production in HCMV-infected neonatal and adult moDC. Adult (white bars) and neonatal moDC (black bars) were either mock-infected or infected with HCMV at MOI of 0.5 or 10. Supernatants were collected after 24 h for cytokines detection. (A) IL-12 p70 (n 5 7 adults and 9neonates), (B) IL-12 p40 (n 5 10 adults and 8 neonates). mRNA were extracted after 3, 6 and 16 h and real-time RT-PCR was performed for mRNAquantification. Peak timepoint at 6 h is represented for IL-12 p35 (A) and IL-12 p40 (B). Mean7SEM of one representative of three independentexperiments involving three donors in each group is shown. ��po0.01 between adult and neonatal moDC as assessed by the Mann–Whitney test.
Eur. J. Immunol. 2009. 39: 2789–2799 Immunity to infection 2791
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
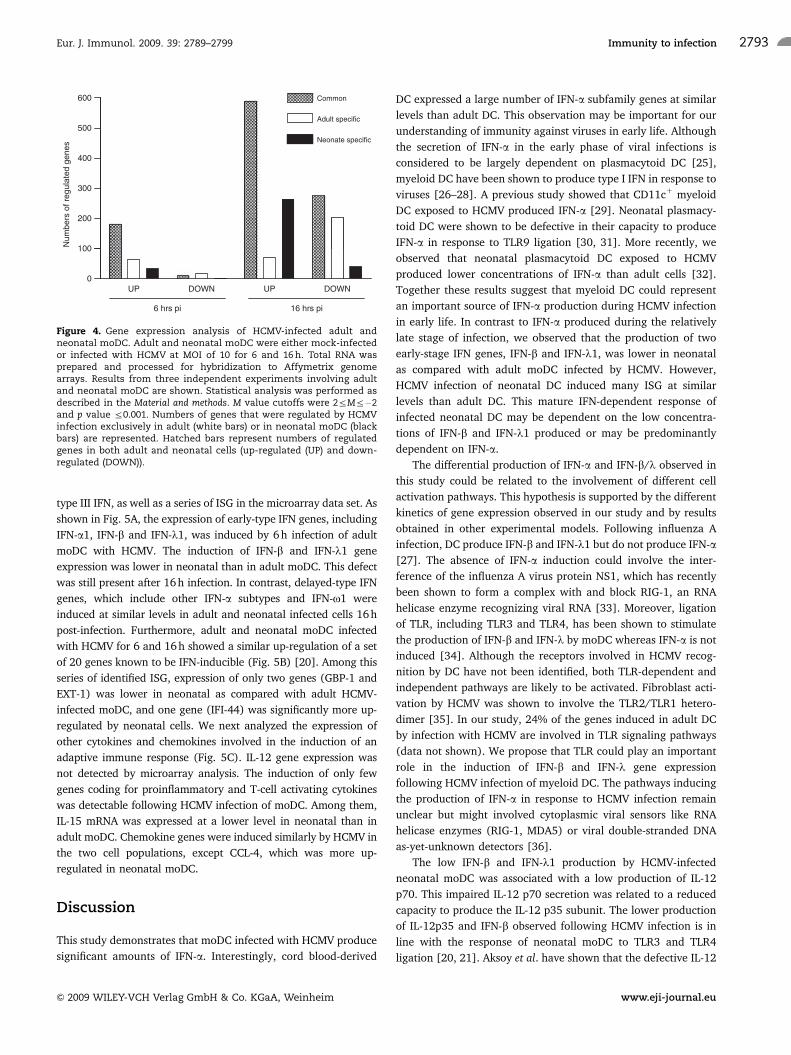
designed to detect 54 676 transcripts. We first compared the genes
expressed by resting adult and neonatal moDC and identified 23
genes differentially expressed by the two study groups at basal level
(Table 1, Supporting Information). These genes are marked by
asterisks in the following tables. Among them, IL-1R2, epiregulin
and IGF-II mRNA-binding protein 3 (IMP-3) could be involved in
the activation or maturation of DC (see the Discussion section). We
then analyzed the ratios of intensities in gene expression between
HCMV-infected and mock-treated DC. Genes changing expression
by four-fold or greater, with a p value less or equal to 0.001, were
considered significantly regulated. Six hours after infection, adult
and neonatal moDC increased the expression of 245 and 215 genes,
and decreased the expression of 28 and 13 genes, respectively. After
16 h infection, 662 and 854 genes were up-regulated and 478 and
315 genes were down-regulated in adult and neonatal moDC,
respectively (Fig. 4). A large fraction of the regulated genes were
different in adult and newborn cells (36.3% of up-regulated and
46.9% of down-regulated genes after 16 h infection). Further, the
statistical comparison of gene regulation magnitudes in adult and
neonatal moDC revealed that gene modulation was significantly
different in adult and neonatal cells for 20 genes after 6 h infection
(Table 2, Supporting Information) and 88 genes after 16 h infection
(Table 3, Supporting Information). Half of the genes that were
differentially modulated at 6 h were still differentially modulated at
16 h. Interestingly, most of the genes differentially regulated after
16 h infection were more up-regulated in neonatal than in adult
moDC. Together, these data indicate that HCMV infection triggers a
gene expression program that is partly different in adult and
neonatal moDC.
Induction of type I IFN and IFN-dependent genes inadult and neonatal moDC by HCMV
To confirm the ability of neonatal moDC to develop a mature IFN-
dependent response, we examined the expression of type I and
IFN-α protein3000 IFN-α2 mRNA2000
Adults
Neonates
1000
2000
IFN
-α (
pg/m
l)1000
mR
NA
cop
ies
/ 106
copi
esof
β-A
ctin
Neonates
mR
NA
cop
ies
/ 106
copi
esof
β-A
ctin
MOI=0 MOI=0.5 MOI=100
IFN-β protein125 * IFN-β mRNA
0MOI=0
50
75
100
1250000
2500000
3750000
MOI=0 MOI=0.5 MOI=100
25
IFN
-β (
UI/m
l)
IFN-λ1 protein
0
CXCL-9 protein
3000
4000
5000 *
300
450
MOI=0 MOI=0.5 MOI=100
1000
2000
IFN
-λ1
(pg/
ml)
MOI=0 MOI=0.5 MOI=10
MOI=0 MOI=0.5 MOI=10
MOI=0.5 MOI=10
0
150
CX
CL-
9 (p
g/m
l)
A
B
C D
Figure 3. Production of type I/III IFN upon HCMV infection in neonatal as compared with adult moDC. Adult (white bars) and neonatal moDC(black bars) were either mock-infected or infected with HCMV at MOI of 0.5 or 10. Supernatants were collected after 24 h for cytokines detection.(A) IFN-a (n 5 10), (B) IFN-b (n 5 10), (C) IFN-l1 (n 5 9), (D) CXCL-9 (n 5 5). mRNA were extracted after 3, 6 and 16 h and real-time RT-PCR wasperformed for mRNA quantification. Peak timepoint at 6 h is represented for (A) IFN-a2 and (B) IFN-b. Mean7SEM of one representative of threeindependent experiments involving three donors in each group is shown. �po0.05 between adult and neonatal moDC as assessed by theMann–Whitney test.
Eur. J. Immunol. 2009. 39: 2789–2799Joelle Renneson et al.2792
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
type III IFN, as well as a series of ISG in the microarray data set. As
shown in Fig. 5A, the expression of early-type IFN genes, including
IFN-a1, IFN-b and IFN-l1, was induced by 6 h infection of adult
moDC with HCMV. The induction of IFN-b and IFN-l1 gene
expression was lower in neonatal than in adult moDC. This defect
was still present after 16 h infection. In contrast, delayed-type IFN
genes, which include other IFN-a subtypes and IFN-o1 were
induced at similar levels in adult and neonatal infected cells 16 h
post-infection. Furthermore, adult and neonatal moDC infected
with HCMV for 6 and 16 h showed a similar up-regulation of a set
of 20 genes known to be IFN-inducible (Fig. 5B) [20]. Among this
series of identified ISG, expression of only two genes (GBP-1 and
EXT-1) was lower in neonatal as compared with adult HCMV-
infected moDC, and one gene (IFI-44) was significantly more up-
regulated by neonatal cells. We next analyzed the expression of
other cytokines and chemokines involved in the induction of an
adaptive immune response (Fig. 5C). IL-12 gene expression was
not detected by microarray analysis. The induction of only few
genes coding for proinflammatory and T-cell activating cytokines
was detectable following HCMV infection of moDC. Among them,
IL-15 mRNA was expressed at a lower level in neonatal than in
adult moDC. Chemokine genes were induced similarly by HCMV in
the two cell populations, except CCL-4, which was more up-
regulated in neonatal moDC.
Discussion
This study demonstrates that moDC infected with HCMV produce
significant amounts of IFN-a. Interestingly, cord blood-derived
DC expressed a large number of IFN-a subfamily genes at similar
levels than adult DC. This observation may be important for our
understanding of immunity against viruses in early life. Although
the secretion of IFN-a in the early phase of viral infections is
considered to be largely dependent on plasmacytoid DC [25],
myeloid DC have been shown to produce type I IFN in response to
viruses [26–28]. A previous study showed that CD11c1 myeloid
DC exposed to HCMV produced IFN-a [29]. Neonatal plasmacy-
toid DC were shown to be defective in their capacity to produce
IFN-a in response to TLR9 ligation [30, 31]. More recently, we
observed that neonatal plasmacytoid DC exposed to HCMV
produced lower concentrations of IFN-a than adult cells [32].
Together these results suggest that myeloid DC could represent
an important source of IFN-a production during HCMV infection
in early life. In contrast to IFN-a produced during the relatively
late stage of infection, we observed that the production of two
early-stage IFN genes, IFN-b and IFN-l1, was lower in neonatal
as compared with adult moDC infected by HCMV. However,
HCMV infection of neonatal DC induced many ISG at similar
levels than adult DC. This mature IFN-dependent response of
infected neonatal DC may be dependent on the low concentra-
tions of IFN-b and IFN-l1 produced or may be predominantly
dependent on IFN-a.
The differential production of IFN-a and IFN-b/l observed in
this study could be related to the involvement of different cell
activation pathways. This hypothesis is supported by the different
kinetics of gene expression observed in our study and by results
obtained in other experimental models. Following influenza A
infection, DC produce IFN-b and IFN-l1 but do not produce IFN-a[27]. The absence of IFN-a induction could involve the inter-
ference of the influenza A virus protein NS1, which has recently
been shown to form a complex with and block RIG-1, an RNA
helicase enzyme recognizing viral RNA [33]. Moreover, ligation
of TLR, including TLR3 and TLR4, has been shown to stimulate
the production of IFN-b and IFN-l by moDC whereas IFN-a is not
induced [34]. Although the receptors involved in HCMV recog-
nition by DC have not been identified, both TLR-dependent and
independent pathways are likely to be activated. Fibroblast acti-
vation by HCMV was shown to involve the TLR2/TLR1 hetero-
dimer [35]. In our study, 24% of the genes induced in adult DC
by infection with HCMV are involved in TLR signaling pathways
(data not shown). We propose that TLR could play an important
role in the induction of IFN-b and IFN-l gene expression
following HCMV infection of myeloid DC. The pathways inducing
the production of IFN-a in response to HCMV infection remain
unclear but might involved cytoplasmic viral sensors like RNA
helicase enzymes (RIG-1, MDA5) or viral double-stranded DNA
as-yet-unknown detectors [36].
The low IFN-b and IFN-l1 production by HCMV-infected
neonatal moDC was associated with a low production of IL-12
p70. This impaired IL-12 p70 secretion was related to a reduced
capacity to produce the IL-12 p35 subunit. The lower production
of IL-12p35 and IFN-b observed following HCMV infection is in
line with the response of neonatal moDC to TLR3 and TLR4
ligation [20, 21]. Aksoy et al. have shown that the defective IL-12
400
500
600 Common
Adult specific
Neonate specific
100
200
300
UP0
16 hrs pi6 hrs pi
DOWN UP DOWN
Num
bers
of r
egul
ated
gen
es
Figure 4. Gene expression analysis of HCMV-infected adult andneonatal moDC. Adult and neonatal moDC were either mock-infectedor infected with HCMV at MOI of 10 for 6 and 16 h. Total RNA wasprepared and processed for hybridization to Affymetrix genomearrays. Results from three independent experiments involving adultand neonatal moDC are shown. Statistical analysis was performed asdescribed in the Material and methods. M value cutoffs were 2rMr�2and p value r0.001. Numbers of genes that were regulated by HCMVinfection exclusively in adult (white bars) or in neonatal moDC (blackbars) are represented. Hatched bars represent numbers of regulatedgenes in both adult and neonatal cells (up-regulated (UP) and down-regulated (DOWN)).
Eur. J. Immunol. 2009. 39: 2789–2799 Immunity to infection 2793
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

p35, IFN-b and ISG synthesis by LPS-stimulated neonatal DC is
associated with decreased IRF (IFN regulatory factor)-3/CBP
(CREB-binding protein) interaction [20]. Another study sugges-
ted that IFN-l1 gene expression could also be dependent on IRF-3
[27]. We suggest that the low IL-12, IFN-b and IFN-l1 gene
expression by HCMV-infected neonatal DC may also involve a
reduced activity of IRF-3, whereas IFN-a gene expression by
HCMV-infected myeloid DC may not be dependent on IRF-3. IRF-
7 is a key regulator of type I IFN during viral infections [37]. IRF-
7 efficiently activates IFN-a and IFN-b gene expression and
represents a positive feedback loop as it is induced by type I IFN
[37]. It will be important to evaluate the role of IRF-7 in the
induction of IFN-a by HCMV-infected adult and neonatal moDC.
The mechanisms involved in the reduced capacity of neonatal
moDC to produce IL-12 and IFN-b in response to LPS stimulation
and HCMV infection remain to be identified. Aksoy et al. detected
similar levels of the signal transduction molecules TRIF, TRAM,
IRF-3, IKKe and TBK1 in immature neonatal and adult moDC by
Western blotting [20]. The comparison of gene expression
profiles of uninfected neonatal and adult moDC did not reveal
differences involving specific pathways but identified several
genes coding for proteins that could be involved in DC matura-
tion and function. Adult moDC expressed higher levels of IL-1R2
that could participate in an autocrine loop amplifying DC acti-
vation. Adult DC also expressed higher levels of epiregulin, a
member of epidermal growth factor superfamily involved in
Type I and III IFNs (6h)60 Type I and III IFNs (16h)300
Adults
10
20
30
40
50
Fol
d ch
ange
10
20
30
40200
250
Fol
d ch
ange
Neonates
1α
IFN
-
2α
IFN
-
4α
IFN
-
5α
IFN
-
7α
IFN
-
8α
IFN
- 10α
IFN
-
13α
IFN
-
14α
IFN
-
16α
IFN
-
1 βIF
N-
1 λIF
N-
1ω
IFN
-
1α
IFN
-
2α
IFN
-
4α
IFN
-
5α
IFN
-
7α
IFN
-
8α
IFN
- 10α
IFN
-
13α
IFN
-
14α
IFN
-
16α
IFN
-
1 βIF
N-
1 λIF
N-
1ω
IFN
-
0
IFN-inducible genes (6h)
0
IFN-inducible genes (16h)
100
200
300
Fol
d ch
ange
200
300
400
500600
750
Fol
d ch
ange
CX
CL-
11C
XC
L-10
CX
CL-
9T
RA
ILIS
G-1
5IS
G-5
4IS
G-5
6IS
G-6
0O
AS
LO
AS
2IF
I-44
MX
-1M
X-2
AP
OB
EC
3AG
BP
-1G
BP
-2IN
DO
EX
T-1
PD
GF
RL
IRF
-7
0
CX
CL-
11C
XC
L-10
CX
CL-
9T
RA
ILIS
G-1
5IS
G-5
4IS
G-5
6IS
G-6
0O
AS
LO
AS
2IF
I-44
MX
-1M
X-2
AP
OB
EC
3AG
BP
-1G
BP
-2IN
DO
EX
T-1
PD
GF
RL
IRF
-7
0
100
Cytokines-chemokines (6h) Cytokines-chemokines (6h)
20
25
30
35
40
45
30
40
50
60
βIL
-1 IL-8
IL-1
5
IL-1
6 αT
NF
-
CC
L-2
CC
L-3
CC
L-4
CC
L-5
CC
L-7
CC
L-8 β
IL-1 IL
-8
IL-1
5
IL-1
6 αT
NF
-
CC
L-2
CC
L-3
CC
L-4
CC
L-5
CC
L-7
CC
L-8
0
5
10
15
Fol
d ch
ange
Fol
d ch
ange
0
10
20
A
B
C
Figure 5. Induction of type I/III IFN, IFN-inducible and cytokine/chemokine genes in adult and neonatal moDC by HCMV. Adult (white bars) andneonatal moDC (black bars) were either mock-infected or infected with HCMV at MOI of 10 for 6 and 16 h. Total RNA was prepared and processedfor hybridization to Affymetrix genome arrays. M value cutoffs were 2rMr�2 and p value r0.001. Fold change in gene expression induced byHCMV infection are represented for (A) type I and III IFN genes, (B) IFN-inducible genes and (C) cytokine-chemokine genes. �po0.05 between adultand neonatal moDC using the statistical methods described in the Materials and methods section.
Eur. J. Immunol. 2009. 39: 2789–2799Joelle Renneson et al.2794
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

proinflammatory cytokine production in mouse macrophages
[38]. Neonatal moDC expressed higher levels of IMP-3. IMP-3 is a
translational activator of IGF-II, a molecule that could be
involved in the maturation of mouse DC [39, 40]. Differences in
IL-1R2 and IMP-3 between neonatal and adult immature moDC
were also observed in an independent study by Aksoy et al. [20].
Further studies are required to evaluate the role of these mole-
cules in the response of neonatal and adult DC to HCMV infection
and TLR ligation.
Microarray analysis showed that HCMV infection has a major
impact on the transcriptional activity of neonatal and adult
moDC. Sixteen hours after infection of adult DC, 1140 genes were
found to be up-regulated or down-regulated by at least four-fold.
Multiple pathways are affected by the regulation of those genes,
particularly TLR and cytokine receptor signaling, Jak-STAT
signaling, cell cycle and apoptosis (data not shown). IFN-
dependent response was strongly induced in HCMV-infected DC,
as previously described in infected fibroblasts [41]. Recently,
HCMV has been shown to induce a transcriptional program
similar to IFN-treated cells, suggesting that a majority of genes
are regulated coordinately by viral infection and type I IFN
treatment [42]. Comparing adult and neonatal response to
HCMV, we demonstrated that HCMV infection triggers a partly
different gene expression program in neonatal and adult moDC.
More than a third of the genes regulated by HCMV infection were
different in adult and newborn cells. The 88 genes that were
differentially regulated in adult and neonatal cells after 16 h
infection belong to multiple families and no unique pathway
could be identified (data not shown). However, important
differences were observed in the expression of immunomodula-
tory genes between adult and neonatal moDC. In addition, the
expression of IMP-3, which was expressed more by uninfected
neonatal DC than by adult DC, was significantly up-regulated
following HCMV infection of adult but not neonatal moDC. These
results suggest that IMP-3 may play an important role in the
regulation of the biology of neonatal and adult moDC. Together,
our results indicate that neonatal DC infected with HCMV show
defective production of the immunomodulatory cytokines
IL-12p70, IFN-b and IFN-l1 but are able to produce adult-like
concentrations of IFN-a and IFN-dependent genes. These defects
could be involved in the differential induction of CD81 and CD41
T-cell responses observed following congenital HCMV infection
[7, 8]. IL-12 is an essential cytokine for development of effector
functions and cytotoxic responses. Viruses have been shown to
induce IL-12 p70 release by DC, including influenza [43] and
dengue virus [28]. IFN-g production by antigen-specific CD41
T cells requires IL-12 and STAT-4 activation, whereas CD81
T cells can produce IFN-g independently of IL-12 [44]. In addi-
tion, both IL-12 and type I IFN signaling were shown to be
required for IFN-g production by antigen-specific CD41 T cells in
Listeria monocytogenes infection [45]. The defective IL-12
production by HCMV-infected neonatal moDC observed in our
study could therefore be involved in the deficient CD41 T-cell
responses in congenitally infected newborns. Defective CD41
T-cell responses were also observed following infection of infants
with HIV or HSV [46]. These defective responses are associated
with a poorer control of viral replication and more severe
outcome. To our knowledge, the responses of neonatal moDC to
infection with HSV and HIV have not been studied. In order to
increase our understanding of the pathogenesis of viral infections
in early life, it will be important to extend the study we have
conducted with HCMV to other pathogens causing severe infec-
tions in young infants. On the other hand, type I IFN contribute to
cytotoxic CD81 T-cell response by diverse mechanisms, indirect
action through induction of DC maturation [47] but also more
direct action [48]. In mice infected with lymphocytic chor-
iomeningitis virus, the expansion and the differentiation of CD81
T cells as well as their production of IFN-g was shown to be
dependent on endogenous IFN-a/b but not on IL-12 [49, 50]. The
preferential production of IFN-a by HCMV-infected neonatal
moDC could therefore play an important role in the functional
CD81 T-cell responses to this virus during fetal life. The TB40/E
strain that was used in the study was selected because it has the
property of inducing a full replicative cycle in DC. Indeed, human
DC are not permissive to conventional laboratory strains of
HCMV, such as the AD169 strain. Genetic variability of HCMV
strains, in particular of clinical isolates, could influence the
response of DC. However, the fact that the defect in IL-12 and
IFN-b production is also observed following activation of
neonatal moDC with LPS suggests that it is not specific to the
HCMV strain used in the study.
In conclusion, our study shows that neonatal moDC respond
differently to HCMV infection than adult moDC by producing
lower levels of specific immunomodulatory cytokines. These
findings have important implications for our understanding of the
pathogenesis of viral infections in early life and for the develop-
ment of vaccines protecting young infant from intracellular
pathogens.
Materials and methods
Generation and culture of moDC
Cord blood was obtained from normal full-term deliveries at the
obstetric departments of CHU Tivoli, La Louviere, CHU, Charleroi
and Clinique Notre Dame, Charleroi, Belgium. Fresh adult blood
was obtained from healthy laboratory staff volunteers. HCMV
seropositive and seronegative donors were included. This study
was approved by the institutional scientific and ethics commit-
tees. Mononuclear cells were isolated by centrifugation over
Lymphoprep density gradient (Nycomed, Oslo, Norway) and
monocytes were purified by positive selection using anti-CD14-
conjugated magnetic microbeads (Miltenyi, Germany). moDC
were generated by culturing monocytes for 5–6 days in RPMI
1640 (Cambrex, Verviers, Belgium) supplemented with 2 mM
L-glutamine, 100 U/mL penicillin, 100 mg/mL streptomycin, 1%
non-essential amino acids, 1% sodium pyruvate, 50 mM
b2-mercaptoethanol and 10% FBS (Hyclone, Perbio, Erembode-
Eur. J. Immunol. 2009. 39: 2789–2799 Immunity to infection 2795
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

gem, Belgium), plus GM-CSF (500 U/mL) and IL-4 (1000 U/mL).
GM-CSF was obtained from Leucomax (Shering-Plough, France)
and IL-4 was from R&D Systems (Abingdon, UK).
Virus
HCMVTB40/E is an endothelial cell-propagated HCMV strain
isolated from a bone marrow transplant recipient [51]. Human
foreskin fibroblasts HFF-1 (ATCC SCRC-1041, LGC Promochem,
Middlesex, UK) were cultured in DMEM (Cambrex) supplemented
with 100 U/mL penicillin, 100mg/mL streptomycin and 15% FBS.
Cells were grown to about 90% confluence and infected at low
MOI (0.1–1 PFU/cell) of HCMVTB40/E in a low volume of DMEM for
3 h at 371C. Complete DMEM with 15% FBS was then added and
cells cultured until the first cytopathic effects appeared (6–7 days).
Supernatants of infected HFF-1 were collected every 3–4 days,
until the cytopathic effects became advanced. Each supernatant
was centrifuged at 1200 g for 10 min to remove cell debris and the
supernatant was stored at �801C. Concentrated virus stock was
obtained by ultracentrifugation at 80 000 g for 120 min at room
temperature. The pellet containing viral particles from 20–25 mL
supernatant was resuspended in 1 mL RPMI, titrated and frozen at
�801C. Viral titers were determined by immunofluorescence on
MRC-5 cells and varied between 107 to 108 PFU/mL.
Infection of moDC
moDC (5� 105/mL) were either infected with HCMVTB40/E at
MOI between 0.5 and 20 PFU/cell or mock-infected with 0.5 mL
MEM 5% FCS for 3 h at 371C. After removal of the virus or
medium, moDC were washed and cultured in complete RPMI
containing 10% FBS for a total time indicated in the figure
legends.
Flow cytometry and detection of infection
Cells were harvested, washed in PBS supplemented with 0.5%
BSA and labeled for surface molecules for 20 min at 41C with the
following Ab: anti-CD80 PE, anti-CD86 APC, anti-HLA-A, B, C
APC and anti-HLA-DR PerCp (all from BD Biosciences, Erembo-
degem, Belgium). moDC were also stained with corresponding
isotype-control monoclonal Ab. The cells were then fixed in 4%
paraformaldehyde and permeabilized with 0.025% Triton-X100
(20 min each step at room temperature). After wash, cells were
stained intracellularly with anti-HCMV IE protein Ab (Argene,
France). The samples were analyzed on a CyAnTM
ADP flow
cytometry analyzer (DakoCytomation) using Summit Software
(version 4.2).
Cytokines quantification
Supernatants from moDC were collected after 24 h of incubation
and analyzed using specific ELISA for IFN-a, IFN-b and IL-12 p40
(all from Biosource, Nivelles, Belgium) with a detection limit of
19.5 pg/mL, 3.1 U/mL, and 9.8 pg/mL, respectively. IFN-l1 and
CXCL-9 levels were determined using ELISA kits (R&D Systems)
with a detection limit of 62.5 and 7.8 pg/mL, respectively. IL-12
p70 levels were measured using Quantikine HS ELISA (R&D
Systems) with a detection limit of 0.6 pg/mL. Statistical analyses
were performed using the Mann–Whitney test. Differences were
considered statistically significant when p value was less than or
equal to 0.05.
RNA purification and real-time RT-PCR
Cells were lyzed and mRNA was extracted using a MagNA Pure
LC RNA Isolation Kit-High Performance (Roche Diagnostics,
Brussels, Belgium). Reverse transcription and real-time PCR were
then carried out using LightCycler-RNA Master Hybridisation
Probes (one-step procedure). Primer sequences are listed in
Table s4 (Supporting Information).
RNA preparation and Affymetrix GeneChiphybridization
Adult and neonatal moDC (three donors in each group) were
either mock-infected or infected with HCMV at an MOI
of 10 PFU/cell for 6 and 16 h. Total RNA was isolated using
TRIzol reagent (Invitrogen, Merelbeke, Belgium) and RNeasy
Micro kit (Qiagen, The Netherlands). Samples were checked
for integrity and concentration using the RNA 6000 Pico
LabChip kit on the Agilent Bioanalyzer 2100 (Agilent Technolo-
gies, Palo Alto, CA, USA). All RNA used met the quality
criteria defined by Agilent and were processed according to
Affymetrix’s (Santa Clara, CA, USA) instructions. Briefly, probes
were generated using the Affymetrix GeneChip Expression
30 Amplification Two-cycle Target Labeling and Controls Reagents
kit . Two rounds of T7 polymerase-based linear RNA amplification
were performed by reverse transcription of RNA using
a T7-(dT) 24 primer and a random hexamer primer, as
described in the expression technical manual from Affymetrix
(http://www.affymetrix.com). Then, biotinylated cRNA was
synthesized from the double stranded cDNA using T7 RNA
polymerase and a biotin-conjugated pseudo-uridine containing
nucleotide mixture provided in the kit. Prior to hybridization, the
cRNA was purified and fragmented. About 10mg from each
experimental sample along with Affymetrix eukaryotic hybridiza-
tion controls were hybridized for 16 h to Human U 133 Plus 2.0
genome arrays, containing 54 676 transcripts. Arrays were washed
and stained with streptavidin-phycoerythrin (Molecular Probes,
Eugene, OR, USA), biotinylated anti-streptavidin (Vector Labora-
tories, Burlingame, CA, USA). Arrays were then scanned with an
Affymetrix GeneChips Scanner 3000 at 570 nm. The Affymetrix
eukaryotic hybridization control kit and Poly-A RNA control kit
were used to ensure efficiency of hybridization and cRNA
amplification. All cRNA were synthesized at the same time.
Eur. J. Immunol. 2009. 39: 2789–2799Joelle Renneson et al.2796
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Microarray data analysis
Each array images (‘‘DAT’’ files) were visually screened to
discount for signal artifacts, scratches or debris and each
of the Genes Chips (‘‘rpt’’ files) was checked for the control
of hybridization defined by Affymetrix GeneChip Manual
(like the uniformity of the signal, the good position of
the grid, the scale factor, the background, the percent of
presence, the house keeping genes and the spikes control).
The data were first analyzed using Affymetrix Microarray
Suite 5.0 software. Various Bioconductor packages for R
(www.bioconductor.org) were used for data analysis. In brief,
Simpleaffy package was used to pre-process the raw data for
background correction, normalization and probe summarization
by the RMA method [52, 53]. Differentially expressed genes
were defined using a statistical tool called LIMMA package,
which is part of the Bioconductor statistical analysis software
[54]. Three statistical approaches, all part of BioConductor
software, were used to identify differentially expressed genes:
(i) significance of analysis for microarrays [55], (ii) a paired t test
controlling the false discovery rate using Benjamini and
Hochberg correction [56], and (iii) the moderated t test in linear
models for microarray data [57] also encompassing the Benja-
mini and Hochberg correction. Data were first filtered based
on ‘‘MA plot’’, where M 5 log2 (fluorescent dye intensity of
infected DC/uninfected DC) and A 5 log2 Ofluorescent dye
intensity of (infected DC)� (uninfected DC), as described by
Yang et al. for cDNA microarray [58]. In the first comparison
between infected and uninfected DC, a gene was called
differentially expressed if M value was more than 2 or less than
�2 and p value was less than or equal to 0.001. In the second
comparison between differentially regulated genes in adult
and neonatal DC, a gene was called differentially regulated
if M value was more than 1 or less than �1 and p value was
less than or equal to 0.05. From the previous gene lists, we
analyzed whether some ontologies or pathways are linked with
those lists. We scanned the Gene Ontology database as well as the
pathways from KEGG. We have used the same methodology
developed by Sorin Draghici for his Onto-Express algorithm
(http://vortex.cs.wayne.edu/ontoexpress/), which calculates
that an ontology or a pathway is statistically associated with a
list of genes according to the number of genes present on the
GeneChips, the number of genes associated with the ontology or
pathway and the number of genes associated with this ontology
or pathways from our gene list. From this information, the
algorithm calculates a p-value that will be corrected for multiple
testing to provide the information that the ontology or pathway
founded is not due to chance.
Acknowledgements: This work was supported by a grant from
the Fonds National de la Recherche Scientifique (FNRS, Belgium)
and the Televie program. The Institute for Medical Immunology
is supported by GlaxoSmithKline Biologicals and the government
of the Walloon Region. Stanislas Goriely is a postdoctoral
research fellow at the FNRS, Belgium. Arnaud Marchant is a
senior research associate at the FNRS, Belgium. Arnaud Marchant
and Michel Goldman have served as consultants and received
financial research support from GlaxoSmithKline Biologicals.
Conflict of interest: The Institute for Medical Immunology is
supported by GlaxoSmithKline Biologicals. Arnaud Marchant and
Michel Goldman have served as consultants and received finan-
cial research support from GlaxoSmithKline Biologicals. The
authors declare no other financial or commercial conflict of
interest.
References
1 Wilson, C. B. and Lewis, D. B., Basis and implications of selectively
diminished cytokine production in neonatal susceptibility to infection.
Rev. Infect. Dis. 1990. 12:S410–S420.
2 Marodi, L., Innate cellular immune responses in newborns. Clin. Immunol.
2006. 118: 137–144.
3 Adkins, B., Leclerc, C. and Marshall-Clarke, S., Neonatal adaptive
immunity comes of age. Nat. Rev. Immunol. 2004. 4: 553–564.
4 Stagno, S., Pass, R. F., Cloud, G., Britt, W. J., Henderson, R. E., Walton, P. D.,
Veren, D. A. et al., Primary cytomegalovirus infection in pregnancy.
Incidence, transmission to fetus, and clinical outcome. J. Am. Med. Assoc.
1986. 256: 1904–1908.
5 Gandhi, M. K. and Khanna, R., Human cytomegalovirus: clinical aspects,
immune regulation, and emerging treatments. Lancet Infect. Dis. 2004. 4:
725–738.
6 Kenneson, A. and Cannon, M. J., Review and meta-analysis of the
epidemiology of congenital cytomegalovirus (CMV) infection. Rev. Med.
Virol. 2007. 17: 253–276.
7 Marchant, A., Appay, V., Van Der, S. M., Dulphy, N., Liesnard, C., Kidd, M.,
Kaye, S. et al., Mature CD8(1) T lymphocyte response to viral infection
during fetal life. J. Clin. Invest. 2003. 111: 1747–1755.
8 Tu, W., Chen, S., Sharp, M., Dekker, C., Manganello, A. M., Tongson, E. C.,
Maecker, H. T. et al., Persistent and selective deficiency of CD41T cell
immunity to cytomegalovirus in immunocompetent young children.
J. Immunol. 2004. 172: 3260–3267.
9 Miles, D. J., Sande, M., Kaye, S., Crozier, S., Ojuola, O., Palmero, M. S.,
Sanneh, M. et al., CD4(1) T cell responses to cytomegalovirus in early life:
a prospective birth cohort study. J. Infect. Dis. 2008. 197: 658–662.
10 Pass, R. F., Stagno, S., Britt, W. J. and Alford, C. A., Specific cell-mediated
immunity and the natural history of congenital infection with cytome-
galovirus. J. Infect. Dis. 1983. 148: 953–961.
11 Pollara, G., Kwan, A., Newton, P. J., Handley, M. E., Chain, B. M. and Katz,
D. R., Dendritic cells in viral pathogenesis: protective or defective? Int.
J. Exp. Pathol. 2005. 86: 187–204.
12 Cella, M., Scheidegger, D., Palmer-Lehmann, K., Lane, P., Lanzavecchia,
A. and Alber, G., Ligation of CD40 on dendritic cells triggers production of
high levels of interleukin-12 and enhances T cell stimulatory capacity:
T-T help via APC activation. J Exp. Med. 1996. 184: 747–752.
13 Watford, W. T., Moriguchi, M., Morinobu, A. and O’Shea, J. J., The biology
of IL-12: coordinating innate and adaptive immune responses. Cytokine
Growth Factor Rev. 2003. 14: 361–368.
Eur. J. Immunol. 2009. 39: 2789–2799 Immunity to infection 2797
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

14 Trinchieri, G., Interleukin-12 and the regulation of innate resistance and
adaptive immunity. Nat. Rev. Immunol. 2003. 3: 133–146.
15 Grandvaux, N., tenOever, B. R., Servant, M. J. and Hiscott, J., The
interferon antiviral response: from viral invasion to evasion. Curr. Opin.
Infect. Dis. 2002. 15: 259–267.
16 Boxel-Dezaire, A. H., Rani, M. R. and Stark, G. R., Complex modulation of
cell type-specific signaling in response to type I interferons. Immunity
2006. 25: 361–372.
17 Meager, A., Visvalingam, K., Dilger, P., Bryan, D. and Wadhwa, M.,
Biological activity of interleukins-28 and -29: comparison with type I
interferons. Cytokine 2005. 31: 109–118.
18 Kotenko, S. V., Gallagher, G., Baurin, V. V., Lewis-Antes, A., Shen, M.,
Shah, N. K., Langer, J. A. et al., IFN-lambdas mediate antiviral protection
through a distinct class II cytokine receptor complex. Nat. Immunol. 2003.
4: 69–77.
19 Sheppard, P., Kindsvogel, W., Xu, W., Henderson, K., Schlutsmeyer, S.,
Whitmore, T. E., Kuestner, R. et al., IL-28, IL-29 and their class II cytokine
receptor IL-28R. Nat. Immunol. 2003. 4: 63–68.
20 Aksoy, E., Albarani, V., Nguyen, M., Laes, J. F., Ruelle, J. L., De Wit, D.,
Willems, F. et al., Interferon regulatory factor 3-dependent responses to
lipopolysaccharide are selectively blunted in cord blood cells. Blood 2007.
109: 2887–2893.
21 Goriely, S., Vincart, B., Stordeur, P., Vekemans, J., Willems, F., Goldman, M.
and De Wit, D., Deficient IL-12(p35) gene expression by dendritic cells
derived from neonatal monocytes. J Immunol. 2001. 166: 2141–2146.
22 Velilla, P. A., Rugeles, M. T. and Chougnet, C. A., Defective antigen-
presenting cell function in human neonates. Clin. Immunol. 2006. 121:
251–259.
23 Moutaftsi, M., Mehl, A. M., Borysiewicz, L. K. and Tabi, Z., Human
cytomegalovirus inhibits maturation and impairs function of monocyte-
derived dendritic cells. Blood 2002. 99: 2913–2921.
24 Riegler, S., Hebart, H., Einsele, H., Brossart, P., Jahn, G. and Sinzger, C.,
Monocyte-derived dendritic cells are permissive to the complete replica-
tive cycle of human cytomegalovirus. J. Gen. Virol. 2000. 81: 393–399.
25 Cao, W. and Liu, Y. J., Innate immune functions of plasmacytoid dendritic
cells. Curr. Opin. Immunol. 2007. 19: 24–30.
26 Diebold, S. S., Montoya, M., Unger, H., Alexopoulou, L., Roy, P., Haswell,
L. E., Al Shamkhani, A. et al., Viral infection switches non-plasmacytoid
dendritic cells into high interferon producers. Nature 2003. 424:
324–328.
27 Osterlund, P., Veckman, V., Siren, J., Klucher, K. M., Hiscott, J.,
Matikainen, S. and Julkunen, I., Gene expression and antiviral activity
of alpha/beta interferons and interleukin-29 in virus-infected human
myeloid dendritic cells. J. Virol. 2005. 79: 9608–9617.
28 Libraty, D. H., Pichyangkul, S., Ajariyakhajorn, C., Endy, T.P. and Ennis,
F. A., Human dendritic cells are activated by dengue virus infection:
enhancement by gamma interferon and implications for disease
pathogenesis. J. Virol. 2001. 75: 3501–3508.
29 Kvale, E. O., Dalgaard, J., Lund-Johansen, F., Rollag, H., Farkas, L.,
Midtvedt, K., Jahnsen, F. L. et al., CD11c1 dendritic cells and plasmacytoid
DCs are activated by human cytomegalovirus and retain efficient T cell-
stimulatory capability upon infection. Blood 2006. 107: 2022–2029.
30 De Wit, D., Olislagers, V., Goriely, S., Vermeulen, F., Wagner, H.,
Goldman, M. and Willems, F., Blood plasmacytoid dendritic cell
responses to CpG oligodeoxynucleotides are impaired in human
newborns. Blood 2004. 103: 1030–1032.
31 Gold, M. C., Donnelly, E., Cook, M. S., Leclair, C. M. and Lewinsohn, D. A.,
Purified neonatal plasmacytoid dendritic cells overcome intrinsic
maturation defect with TLR agonist stimulation. Pediatr. Res. 2006. 60:
34–37.
32 Danis, B., George, T. C., Goriely, S., Dutta, B., Renneson, J., Gatto, L.,
Fitzgerald-Bocarsly, P. et al., Interferon regulatory factor 7-mediated
responses are defective in cord blood plasmacytoid dendritic cells. Eur.
J. Immunol. 2008. 38: 507–517.
33 Pichlmair, A., Schulz, O., Tan, C. P., Naslund, T. I., Liljestrom, P.,
Weber, F. and Reis e Sousa, C., RIG-I-mediated antiviral responses
to single-stranded RNA bearing 50-phosphates. Science 2006. 314:
997–1001.
34 Coccia, E. M., Severa, M., Giacomini, E., Monneron, D., Remoli, M. E.,
Julkunen, I., Cella, M. et al., Viral infection and Toll-like receptor agonists
induce a differential expression of type I and lambda interferons in
human plasmacytoid and monocyte-derived dendritic cells. Eur.
J. Immunol. 2004. 34: 796–805.
35 Boehme, K. W., Guerrero, M. and Compton, T., Human cytomegalovirus
envelope glycoproteins B and H are necessary for TLR2 activation in
permissive cells. J. Immunol. 2006. 177: 7094–7102.
36 Kawai, T. and Akira, S., Innate immune recognition of viral infection. Nat.
Immunol. 2006. 7: 131–137.
37 Honda, K., Takaoka, A. and Taniguchi, T., Type I interferon [corrected]
gene induction by the interferon regulatory factor family of transcription
factors. Immunity 2006. 25: 349–360.
38 Sugiyama, S., Nakabayashi, K., Baba, I., Sasazuki, T. and Shirasawa, S.,
Role of epiregulin in peptidoglycan-induced proinflammatory cytokine
production by antigen presenting cells. Biochem. Biophys. Res. Commun.
2005. 337: 271–274.
39 Liao, B., Hu, Y., Herrick, D.J. and Brewer, G., The RNA-binding protein
IMP-3 is a translational activator of insulin-like growth factor II leader-3
mRNA during proliferation of human K562 leukemia cells. J. Biol. Chem.
2005. 280: 18517–18524.
40 Hansenne, I., Renard-Charlet, C., Greimers, R. and Geenen, V., Dendritic
cell differentiation and immune tolerance to insulin-related peptides in
Igf2-deficient mice. J. Immunol. 2006. 176: 4651–4657.
41 Browne, E. P., Wing, B., Coleman, D. and Shenk, T., Altered cellular
mRNA levels in human cytomegalovirus-infected fibroblasts: viral
block to the accumulation of antiviral mRNAs. J Virol. 2001. 75:
12319–12330.
42 Simmen, K. A., Singh, J., Luukkonen, B. G., Lopper, M., Bittner, A., Miller,
N. E., Jackson, M. R. et al., Global modulation of cellular transcription by
human cytomegalovirus is initiated by viral glycoprotein B. Proc. Natl.
Acad. Sci. USA 2001. 98: 7140–7145.
43 Cella, M., Salio, M., Sakakibara, Y., Langen, H., Julkunen, I. and
Lanzavecchia, A., Maturation, activation, and protection of dendritic
cells induced by double-stranded RNA. J. Exp. Med. 1999. 189: 821–829.
44 Carter, L. L. and Murphy, K. M., Lineage-specific requirement for signal
transducer and activator of transcription (Stat)4 in interferon gamma
production from CD4(1) versus CD8(1) T cells. J. Exp. Med. 1999. 189:
1355–1360.
45 Way, S. S., Havenar-Daughton, C., Kolumam, G. A., Orgun, N. N. and
Murali-Krishna, K., IL-12 and Type-I IFN synergize for IFN-gamma
production by CD4 T cells, whereas neither are required for IFN-gamma
production by CD8 T cells after Listeria monocytogenes infection.
J. Immunol. 2007. 178: 4498–4505.
46 Marchant, A. and Goldman, M., T cell-mediated immune responses in
human newborns: ready to learn? Clin. Exp. Immunol. 2005. 141: 10–18.
47 Biron, C. A., Interferons alpha and beta as immune regulators--a new
look. Immunity 2001. 14: 661–664.
Eur. J. Immunol. 2009. 39: 2789–2799Joelle Renneson et al.2798
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

48 Stetson, D. B. and Medzhitov, R., Type I interferons in host defense.
Immunity 2006. 25: 373–381.
49 Kolumam, G. A., Thomas, S., Thompson, L. J., Sprent, J. and Murali-
Krishna, K., Type I interferons act directly on CD8 T cells to allow clonal
expansion and memory formation in response to viral infection. J Exp.
Med. 2005. 202: 637–650.
50 Cousens, L. P., Peterson, R., Hsu, S., Dorner, A., Altman, J. D., Ahmed, R.
and Biron, C. A., Two roads diverged: interferon alpha/beta- and
interleukin 12-mediated pathways in promoting T cell interferon gamma
responses during viral infection. J. Exp. Med. 1999. 189: 1315–1328.
51 Sinzger, C., Schmidt, K., Knapp, J., Kahl, M., Beck, R., Waldman, J., Hebart,
H. et al., Modification of human cytomegalovirus tropism through
propagation in vitro is associated with changes in the viral genome.
J. Gen. Virol. 1999. 80: 2867–2877.
52 Bolstad, B. M., Irizarry, R. A., Astrand, M. and Speed, T. P., A comparison
of normalization methods for high density oligonucleotide array data
based on variance and bias. Bioinformatics 2003. 19: 185–193.
53 Irizarry, R. A., Hobbs, B., Collin, F., Beazer-Barclay, Y. D., Antonellis, K. J.,
Scherf, U. and Speed, T. P., Exploration, normalization, and summaries of
high density oligonucleotide array probe level data. Biostatistics 2003. 4:
249–264.
54 Gentleman, R. C., Carey, V. J., Bates, D. M., Bolstad, B., Dettling, M.,
Dudoit, S., Ellis, B. et al., Bioconductor: open software development for
computational biology and bioinformatics. Genome Biol. 2004. 5: R80.
55 Tusher, V. G., Tibshirani, R. and Chu, G., Significance analysis of
microarrays applied to the ionizing radiation response. Proc. Natl. Acad.
Sci. USA 2001. 98: 5116–5121.
56 Benjamini, Y. and Hochberg, Y., Controlling the false discovery rate: a
practical and powerful approach to multiple testing. J. R. Stat. Soc. B 1995.
57: 289–300.
57 Smyth, G. K., Linear models and empirical bayes methods for assessing
differential expression in microarray experiments. Stat. Appl. Genet. Mol.
Biol. 2004. 3: Article 3.
58 Yang, Y. H., Dudoit, S., Luu, P., Lin, D.M., Peng, V., Ngai, J. and Speed,
T. P., Normalization for cDNA microarray data: a robust composite
method addressing single and multiple slide systematic variation. Nucleic
Acids Res. 2002. 30: e15.
Abbreviations: HCMV: human CMV � IMP-3: IGF-II mRNA-binding
protein 3 � IRF: IFN-regulatory factor � ISG: IFN-stimulated genes �moDC: monocyte-derived DC � PFU: plaque forming unit
Full correspondence: Dr. Arnaud Marchant, Institute for Medical
Immunology, Universite Libre de Bruxelles, Rue Adrienne Bolland 8,
B-6041 Charleroi, Belgium
Fax: 132-2-650-9563
e-mail: [email protected]
Supporting Information for this article is available at
www.wiley-vch.de/contents/jc_2040/2009/39414_s.pdf
Received: 12/3/2009
Revised: 2/7/2009
Accepted: 17/7/2009
Eur. J. Immunol. 2009. 39: 2789–2799 Immunity to infection 2799
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
Copyright © 2022 FDOKUMEN




















