
A Role for EZH2 in Silencing of IFN- Inducible MHC2TA Transcription in Uveal Melanoma
Upload
independentCategory
view
0download
0
of July 8, 2016.This information is current as
with Antigen Processing-1 Promoter ActivitySTAT1 to Regulate Transporter Associated IFN Regulatory Factor-2 Cooperates with
Serge Fichelson, Agnès Buzyn and Isabelle Dusanter-FourtMarie-Christine Rouyez, Marta Lestingi, Martine Charon,
http://www.jimmunol.org/content/174/7/3948doi: 10.4049/jimmunol.174.7.3948
2005; 174:3948-3958; ;J Immunol
Referenceshttp://www.jimmunol.org/content/174/7/3948.full#ref-list-1
, 32 of which you can access for free at: cites 56 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2005 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on July 8, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on July 8, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

IFN Regulatory Factor-2 Cooperates with STAT1 toRegulate Transporter Associated with Antigen Processing-1Promoter Activity1
Marie-Christine Rouyez,* Marta Lestingi,* Martine Charon,* Serge Fichelson,* Agnes Buzyn,†
and Isabelle Dusanter-Fourt2*
Class I MHC complexes (MHCI) are essential in mediating immune response. The transport of antigenic peptides (TAP) to MHCI
and the stable expression of MHCI on the cell surface require the presence of a dedicated TAP. In this study we report that IFN-�and thrombopoietin (TPO) strongly increase TAP1 protein expression in megakaryocytes, followed by an enhanced expression ofMHCI on the cell surface. This expression parallels the enhanced TAP1 promoter activity and TAP1 mRNA expression, which areindependent of protein synthesis. We also show that this cytokine-dependent expression of TAP1 transcripts depends on STAT1and IFN regulatory factor-2 (IRF-2), but not on IRF-1, and provide evidence that IRF-2 constitutively binds to the TAP1 genepromoter and enhances TAP1 promoter activity. We show that IRF-2 forms a complex with STAT1 and the cytokine-responsiveregion of the TAP1 promoter in any TPO or IFN-� target cells tested. Interaction of IRF-2 and STAT1 on the promoter dependson the DNA-binding domain of IRF-2. Overall, our data indicate that TPO and IFN-� activate the expression of TAP1 via a newmechanism that involves functional cooperation between STAT1 and IRF-2 on the TAP1 promoter. The Journal of Immunology,2005, 174: 3948–3958.
C lass I MHC complex (MHCI)3 and class II MHC mole-
cules are membrane-bound glycoproteins that fulfill keyroles in the immune response by their ability to present
antigenic peptides to T lymphocytes (1). MHCI molecules arecomposed of an H chain and an L chain (�2-microglobulin) and areubiquitously expressed. They are essential in mediating immuneresponses against viruses and tumor cells. The class I binding pep-tides are generally derived from proteins synthesized within thecytoplasm of the cell. They are produced by the multicatalyticprotease complex (proteasome) whose cleavage specificity can bealtered by the IFN-inducible subunits low molecular weightpolypeptide (LMP)2 and LMP7 (2). They are transported into thelumen of the endoplasmic reticulum, where class I MHC mole-cules fold and mature, by a dedicated transporter of antigenic pep-tides (TAP; a complex of TAP1 and TAP2), which associates withMHCI H chains (3). The TAP1 and TAP2 heterodimers as well asthe peptides provided by the complex of TAP are important for theproper folding, maturation, and efficient cell surface expression ofclass I MHC molecules (4, 5). Cells that lack TAP1 or TAP2 have
selective inability to present intracellular Ags on MHCIs on thecell surface (6, 7).
Down-regulation of genes associated with peptide productionand transport as well as cell surface expression of class I com-plexes are common in mouse and human tumors, allowing tumorcells to evade antitumor-associated Ag immunity (8, 9). In con-trast, proinflammatory cytokines, such as IFNs and TNF, inducethe coordinate expression of structural class I chains and TAP,providing efficient maturation and surface expression of the classI molecules. Transcriptional regulation of MHCI and TAP geneshas been analyzed in detail in epithelial and immune cells. TheTAP1 gene is regulated by a promoter region that it shares with theLMP2 gene. The region between the two genes, which are tran-scribed in opposite directions, is only 593 bp (10). Within thisregulatory region, five cis-acting elements have been mapped andstudied. These are SP-1 and NF-�B binding sites and a combina-tion of two overlapping IFN regulatory factor elements (or IFNconsensus sequences (ICS)) and an IFN-�-activated sequence(GAS) (10–13). ICSs are the target sites for transcription factors ofthe IFN regulatory factor (IRF) family that have been implicated inantiviral defense, cell growth, and immune regulation (14). Ninemembers of the family have been identified. All IRFs share sig-nificant homology within their N-terminal DNA-binding domain(DBD), which determines a characteristic DNA binding selectivityfor GAAA elements (15, 16). GASs, which exhibit the canonicalTTCnnnGAA motif, are target sites for transcription factors of theSTAT family that are involved in a variety of biological processes.Tyrosine phosphorylation of the STATs renders them competentfor dimerization, nuclear translocation, and DNA binding. Mostactivities of IFN-� result from the STAT1-mediated transcriptionalresponse. The NF-�B site of the TAP1 promoter has been shownto be important for the induction of promoter activity after stim-ulation with TNF-� (10, 11). Data regarding regulation by IFN-�have been conflicting; some reports concluded that the binding of
Departments of *Hematology and †Immunology, Institut Cochin, Paris, France
Received for publication June 25, 2004. Accepted for publication December 13, 2004.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported in part by l’Association de Recherche contre le Cancer (toI.D.-F).2 Address correspondence and reprint requests to Dr. Isabelle Dusanter-Fourt, De-partment d’hematologie, Institut Cochin, Institut National de la Sante et de la Re-cherche Medicale, Unite 567, Unite Mixte de Recherche 8104, Universite Paris 5,Maternite Port Royal, 123 boulevard Port Royal, 75014 Paris, France. E-mail address:[email protected] Abbreviations used in this paper: MHCI, class I MHC complex; TAP, transport ofantigenic peptide; DBD, DNA-binding domain; EPO, erythropoietin; GAS, IFN-�-activated sequence; ICS, IFN consensus sequence; IRF, IFN regulatory factor; LMP,low molecular weight polypeptide; MEF, murine embryonic fibroblast; TPO,thrombopoietin.
The Journal of Immunology
Copyright © 2005 by The American Association of Immunologists, Inc. 0022-1767/05/$02.00
by guest on July 8, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

STAT1 to the TAP1-GAS site was essential, whereas others pro-vided evidence that IRF-1 bound to the ICS was fundamental forthe IFN-� response (11, 17).
Thrombopoietin (TPO) is the primary physiological regulator ofplatelet production; it is a lineage dominant factor affectingmegakaryocytopoiesis (18). TPO also acts during early hemopoi-esis, regulating hemopoietic stem cell production and function(19), and retroviral transduction of a truncated and constitutivelyactivated TPO-receptor, v-mpl, led to a myeloproliferative syn-drome in infected mice (20). TPO is known to favor themegakaryocytic maturation of two human multipotent cell lines,UT7 and F36P, after introduction of an exogenous TPO receptor,as we and others demonstrated (21, 22). With the purpose of iden-tifying genes regulated by TPO, we have set up a subtractive li-brary approach and screened for cDNA selectively expressed uponTPO treatment of UT7-mpl cells. Among the clones selected, weidentified the cDNA coding for TAP1. In the present paper weshow that megakaryocytes do express important amounts of TAP1.We analyzed and compared the regulation of expression of theTAP1 gene after TPO or IFN-� treatment. Our results indicate thatboth cytokines activate the expression of TAP1 mRNA by acti-vating TAP1 promoter through a new mechanism that involvesboth the presence of DNA-bound IRF-2 and activation of STAT1.
Materials and MethodsReagents and Abs
The TPO mimetic peptide GW395058 (23) was synthesized by GenosysBiotechnologies. Human rIFN-� and erythropoietin (EPO) were obtainedfrom Roche; rTPO was provided by Amgen. The following Abs were pur-chased: anti-IRF-1 and anti-IRF-2 Abs from Santa Cruz Biotechnology;polyclonal anti-STAT1 and monoclonal anti-STAT3 Abs from BD Trans-duction Laboratories; anti-phosphospecific STAT1(Tyr701) Abs from NewEngland Biolabs; and anti-LMP2 Abs from Affiniti Research Products.Purified mAb 148.3 directed against TAP1 was provided by Dr. P. M. VanEndert (Institute National de la Sante et de la Recherche Medicale, Paris,France) and Dr. R. Tampe (Institute of Biochemistry, Biozentrum Frank-furt, Johann Wolfgang Goethe-University, Frankfurt/Main, Germany) (24).The anti-HLA-A2 BB7.2 and anti-MHC class I W6-32 Abs were purifiedfrom hybridoma culture medium.
Cell culture
Human UT7-mpl cells were maintained in �-MEM supplemented with10% FCS and 2 U of EPO/ml as previously described (22). UT7-mpl cellswere treated for the appropriate time with either 10 U of human IFN-�/ml,10 nM TPO mimetic peptide GW395058, or 100 ng of recombinant cyto-kine/ml (25, 26). Where indicated, UT7-mpl cells were pretreated for 30min in the presence of cycloheximide (10 �g/ml; Sigma-Aldrich) beforeadding TPO or IFN-�. IRF-1/IRF-2 double-knockout murine embryonicfibroblasts (MEF) were provided by Dr. G. Blanck (Department of Bio-chemistry and Molecular Biology, University of South Florida, Tampa, FL)with permission from Dr. T. Taniguchi (Department of Immunology, Uni-versity of Tokyo, Tokyo, Japan). Human K562 cells, HeLa cells, THP-1cells, 293T cells, and IRF-1�/� IRF-2�/� MEF were all maintained in�-MEM supplemented with 10% FCS and treated for the appropriate timewith 10 U of IFN-�/ml. Umbilical cord blood units from normal full-termdeliveries from the Obstetrics Unit of the Hopital Saint-Vincent de Paulwere obtained after informed consent of the mothers. Cord bloods werediluted and submitted to Ficoll density gradient. Low density cells wererecovered, and CD34� cells were separated by positive selection using animmunomagnetic procedure (CD34� isolation kit; Miltenyi Biotec). Cellswere cultured in serum-free IMDM (Invitrogen Life Technologies), in thepresence of 15% of a commercial mixture of BSA, insulin, and transferrin(BIT 9500; StemCell Technologies) supplemented with stem cell factor (25ng/ml; Amgen), and TPO (100 ng/ml) to amplify megakaryocyte progen-itors, as previously described (27). Cells were incubated at 37°C for 10–12days. Blood samples from adults were obtained after informed consent wasprovided by the patients. Cells were submitted to Ficoll density gradient,and low density cells were recovered and diluted in RPMI 1640 mediumsupplemented with 10% serum. Cells (106/cm2) were maintained in cultureflasks for 2 h at 37°C, and nonadherent cells were discarded. After twowashes, a cell population enriched in adherent monocytes was collected.
Plasmid constructs
The luciferase reporter wild-type TAP1-pGL2 encompassing the wholepromoter region between the two initiator codons of LMP-2 and TAP1 aswell as the one including the TAP1 promoter fragment (�210, �1) wereprovided by Dr. D. Johnson (Department of Pathology, Yale UniversitySchool of Medicine, New Haven, CT) (11). An expression plasmid encod-ing full length IRF-2 was kindly provided by Dr. J. Wietzerbin (InstitutCurie, Paris, France). IRF-2 cDNA sequence was subcloned in thepcDNA3 expression vector. Mutated TAP1 promoter reporter constructsand vectors encoding deletion mutants of IRF-2 were obtained by exchang-ing appropriate PCR fragments from the wild-type constructs.
Northern blot
Total RNA was prepared using TRIzol reagent (Invitrogen Life Technol-ogies). RNA (10 �g) was separated onto 1% agarose denaturing gel andtransferred to Hybond-N membranes (Amersham Biosciences). Blots werehybridized with an �-32P-labeled probe corresponding to the human TAP1coding sequence and subsequently to a GAPDH-labeled probe.
Transient transfections
Transient transfections of UT7-mpl cells were performed using 10 �g ofluciferase reporter plasmid and 1 �g of plasmid Rous sarcoma virus-gal-actosidase as an internal control, as previously described (25). After elec-troporation, cells were resuspended in proliferative medium and splitamong three flasks containing TPO (10 nM), IFN-� (10 U/ml), or no cy-tokine. Cells were maintained in culture for 15 h before lysis. Luciferaseactivities were normalized to �-galactosidase activities and expressed asrelative luciferase units. Where indicated, expression vectors coding forfull-length or truncated versions of IRF-2 were transfected in UT7-mplcells by electroporation. Cells were maintained in culture for 18–24 h, thenincubated with IFN-� (10 U/ml) or TPO (10 nM) for 0.5–3 h and imme-diately processed for preparing nuclear extracts.
Transfections into either 293T cells or IRF-1�/�IRF-2�/�MEF (2 � 105
cells/experiment) were performed in triplicate with 200 ng of the wild-typeTAP1-luciferase reporter construct and varying amounts (0–200 ng) ofpcDNA3-IRF-2 expression vector using Lipofectamine (Invitrogen LifeTechnologies) according to the manufacturer’s instructions. Cells weremaintained in culture for 15–18 h, harvested, and processed for luciferaseassays.
Immunoprecipitation and Western blotting
Total cell extracts were prepared by lysing the cells for 30 min at 4°C inNonidet P-40 buffer (50 mM Tris (pH 7.4), 137 mM NaCl, 1 mM EDTA,10% glycerol, 1% Nonidet P-40, 1 mM PMSF, 1 mM sodium orthovana-date, 10 �g/ml leupeptin, 10 �g/ml aprotinin, and 1 �g/ml pepstatin) andsubsequent centrifugation at 20,000 � g for 15 min. Nuclear extracts wereprepared as previously described (25). Conventional Western blot analyseswere conducted with peroxidase-conjugated anti-mouse or anti-rabbit Absusing chemiluminescence as instructed by the manufacturer (AmershamBiosciences).
Flow cytometric analysis
Cells treated, or not, with TPO or IFN-� were washed in PBA (PBS con-taining 0.2% sodium azide and 1% BSA) and preincubated for 30 min at4°C with 1 mg of normal goat aggregated IgG/ml, then incubated withmouse primary mAbs for 30 min at 4°C and further incubated with FITC-conjugated F(ab�)2 of goat anti-mouse IgG (1/100) for another 30 min.Cells were fixed with 1% formaldehyde and analyzed for fluorescence onan ELITE flow cytometer.
EMSAs
For each reaction, nuclear extracts (10 �g) were incubated in bindingbuffer in ice for 30 min with oligonucleotide probes end-labeled with[�-32P]ATP and loaded on a 6% nondenaturing polyacrylamide gel as de-scribed previously (25). Where indicated, the reaction was allowed to pro-ceed in the presence of a 5- to 100-fold molar excess of unlabeled oligo-nucleotides or 1 �g of the indicated Abs. The probes used in the EMSAsare listed in Table I. Nuclear extract preparations were always checked forequivalent nuclear extraction by binding on a consensus octamer probe.
3949The Journal of Immunology
by guest on July 8, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

ResultsTPO and IFN-� induce the expression of functional TAP1 inmegakaryocyte progenitors
The multipotent human hemopoietic UT7-mpl cells express exog-enous TPO-R and require EPO or GM-CSF for their growth andsurvival. Upon TPO addition, UT7-mpl cells reduce their prolif-erative rate and undergo megakaryocytic differentiation (22, 26). Asubtractive cDNA library was constructed from RNAs extractedfrom UT7-mpl cells incubated for 8 h in the presence or the ab-sence of TPO. Among the cDNA selectively expressed, we iden-tified TAP1 as a potential TPO target gene. The expression ofTAP1 proteins was analyzed in UT7-mpl cells after TPO treat-ment. As shown in Fig. 1A, untreated UT7-mpl cells expressedbarely detectable levels of TAP1. TAP1 expression strongly in-creased after 3–6 h of treatment with TPO and reached a plateauat 12–24 h. TAP1 expression similarly increased after IFN-� ad-dition (Fig. 1B), whereas neither EPO nor GM-CSF enhancedTAP1 in UT7-mpl cells (not shown). In parallel, LMP2 levels wereunaffected by the presence of TPO or IFN-� (Fig. 1, A and B, lowerpanels).
We analyzed the expression of TAP1 in normal TPO-responsivecells. Megakaryocyte progenitors were selected and amplifiedfrom purified human cord blood CD34� cells by a 10- to 12-dayex vivo culture in TPO-containing medium, as previously de-scribed (27). Cells were next maintained in TPO-deprived mediumfor 24 h, then stimulated with TPO or IFN-�. As shown in Fig. 1C,TPO and IFN-� enhanced the expression of TAP1 in megakaryo-cytes (upper panel), but none of these cytokines affected the ex-pression of LMP2 (lower panel).
The functionality of TPO-induced TAP1 molecules was as-sessed by testing their capacity to increase cell surface expressionof class I MHC molecules. As shown in Fig. 2, TPO enhanced theexpression of class I MHC molecules on the surface of UT7-mplcells. This effect was rapid, being detectable as early as 4 h afteraddition of the cytokine, in accordance with the kinetics of TAP1expression. It was detected with Abs that were specific for themonomorphic MHCI determinant (W6/32, not shown) as well aswith Abs specific for the A2 determinant expressed by the UT7cells (BB7.2; Fig. 2). Similarly, IFN-� enhanced the expression ofclass I MHC molecules on the surface of UT7-mpl cells, with amagnitude similar to that observed after TPO treatment (left andright panels).
Together, these data indicate that TPO enhances the expressionof functional TAP1 molecules in a rapid and sustained manner,much as IFN-� does in the same cell context. We thus analyzed themechanisms that participated in the TPO/IFN-�-dependent in-crease in TAP1 expression in this cellular context.
TPO and IFN-� activate transcription of TAP1 gene
The kinetics of expression of TAP1 mRNA were studied in UT7-mpl cells. Both TPO and IFN-� induced a similar and rapid accu-mulation of TAP1 mRNA (Fig. 3, lanes 1–7). This accumulationpeaked at 6–10 h, then slowly decreased (data not shown). Suchkinetics totally agreed with previous reports on TAP1 mRNA lev-els in IFN-�-stimulated epithelial and endothelial cells (11, 28). Todecipher the mechanism of TAP1 mRNA induction, we firsttreated cells in the presence of a protein synthesis inhibitor andtested the consequence of such a treatment on TAP1 mRNA ex-pression. We observed that TPO and IFN-� both enhanced theexpression of TAP1 transcripts regardless of the presence of theprotein synthesis inhibitor cycloheximide (lanes 8–15), at a con-centration of cycloheximide that totally blocked translation inUT7-mpl cells (see the expression of IRF-1, Fig. 7A). This resultindicates that TAP1 mRNA induction by the two cytokines doesnot require ongoing protein synthesis.
The activity of the TAP1 gene promoter was then assessed bytransient transfection assays, followed by stimulation with TPO orIFN-�. The 593-bp intergenic sequence between the LMP2 and
Table I. Sequences of the oligonucleotides used in EMSAa
Sequences
GAS GCTTTCCCCTAAATGICS12 GGCCGCTTTCGTTTCGCTTTCCICS2-GAS CGAATTTCGCTTTCCCCTAAATGICS12-GAS GGCCGCTTTCGATTTCGCTTTCCCCTAAATG
a Consensus GAS and ICS are underlined.
FIGURE 1. TPO and IFN-� induce the expression of TAP1 inmegakaryocytes. UT7-mpl cells were treated with TPO (A) or IFN-� (B)for the indicated time, and total cell extracts were prepared. Proteins (100�g) were analyzed by Western blotting with either anti-TAP1 or anti-LMP2 Abs, as indicated on the right. C, Human megakaryocyte progeni-tors were selected and amplified from purified cord blood CD34� by a10-day ex vivo culture in TPO-containing medium. Cells were then de-prived of TPO for 1 day, stimulated for 15 h in the presence of TPO (10nM) or IFN-� (100 U/ml), and lysed. Proteins (100 �g) were analyzed byWestern blotting using either anti-TAP1 or anti-LMP2 Abs as indicated.
FIGURE 2. TPO and IFN-� enhance the expression of MHCI proteinson the cell surface. UT7-mpl cells were treated for the indicated time withTPO or IFN-�. Cell surface expression of the MHCI-A2 determinant com-plex was analyzed by flow cytometry using the specific mAb BB7.2.
3950 IRF-2 COOPERATES WITH STAT1 TO REGULATE TAP1
by guest on July 8, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

TAP1 coding sequences was subcloned into the promoterlesspGL2 plasmid (Fig. 4A). As shown in Fig. 4B, the activity of theTAP1 promoter was enhanced 9- to 10-fold after TPO (o) orIFN-� (f) treatment, indicating that both cytokines transcription-ally regulated the expression of TAP1. A deletion mutant was con-structed encompassing only the first 210 bp upstream of the TAP1translation initiation codon, because this region is sufficient to pro-mote IFN-�- and TNF-�-induced TAP1 expression in HeLa cells(11). This deletion mutant was still cytokine-responsive and me-diated the TPO and IFN-� responses of the TAP1 promoter in theUT7-mpl cells (Fig. 4B).
The cytokine-responsive region of the TAP1 promoter was pre-viously reported to contain five cis-acting elements (Fig. 4A) (11).Three regularly spaced and adjacent TTTC motifs were present inthe overlapping ICSs and GAS, the last was included within the 5�half of GAS. Each of these repeats may potentially bind IRFs, as
deduced from crystallographic studies of IRF-DNA complexes(15, 16). We performed site-directed mutagenesis to generate con-structs where IRF or STAT binding sequences were selectivelydisrupted without altering the functionality of the neighboring sites(see Fig. 4). One mutant, thereafter designated mICS12, disruptedboth the first and second TTTC motifs present in the overlappingICS1-ICS2 sequence and could no longer bind IRF-binding pro-teins (data not shown); another one, thereafter designated mGAS,disrupted only the 3� end, but not the 5� end, of GAS that over-lapped with the third IRF binding core motif and could no longerbind STAT (data not shown). Transfection of these mutant pro-moters into UT7-mpl cells showed that loss of either GAS or ICSindividually impaired the ability of the promoter to respond toIFN-� treatment (Fig. 4B, f). Mutating either GAS or ICS alsoimpaired the ability of the promoter to respond to TPO (o). Eachof these two mutations strongly inhibited the IFN-� response,whereas they had only partial effects on the TPO response. Thesedata indicate that GAS and ICS12 both participate in TPO- andIFN-�-dependent TAP1 gene promoter activation, whereas an ad-ditional pathway(s) also contributes to TPO-dependent activationof this promoter.
Identification of nuclear factors that bind to TAP1 promoter
The ICS and GAS promoter elements are known to bind IRFs andSTATs, respectively. TPO can activate STAT1, STAT3, andSTAT5A/B depending on cell contexts (29), whereas STAT1 is themajor STAT activated by IFN-� in IFN-�-responsive cells. In theUT7-mpl cells, TPO induced a sustained activation of STAT1(Fig. 5A) and STAT3 (data not shown), in addition to the previ-ously reported activation of STAT5 (25, 30). Western blot analysis
FIGURE 3. TPO and IFN-� induce a similar accumulation of TAP1mRNA in the presence and the absence of protein synthesis. UT7-mpl cellswere incubated for 30 min at 37°C in the absence (lanes 1–7) or the pres-ence (lanes 8–15) of cycloheximide and further treated for the indicatedtime with TPO or IFN-�. Total RNA were isolated and analyzed for thepresence of TAP1 or GAPDH mRNA by Northern blot.
FIGURE 4. TPO and IFN-� activate the TAP1 pro-moter. A, Schematic representation of the intergenic se-quence between the human TAP1 and LMP2 transcrip-tion units. The IRF core binding motifs, TTTC, areindicated in bold. The ICS1, ICS2, and GAS sequencesare boxed. Mutations that have been introduced into thevarious motifs are indicated at the bottom. B, UT7-mplcells were transiently transfected with the indicated pro-moter luciferase reporters along with control �-galac-tosidase expression vector. After transfection, cellswere treated with medium (�), TPO (o), or IFN-� (f)for 18 h, and luciferase and �-galactosidase activitieswere determined. Results are expressed as luciferaseactivity relative to �-galactosidase activity (upperpanel). Alternatively, promoter activity was determinedby dividing the relative luciferase activity of cells trans-fected with the mutated constructs by that of the cellstransfected with the wild-type (wt) TAP1 promoter con-struct (lower panel). All plasmids were transfected aminimum of five times in separate experiments. Thelocations of the mutated nucleotides in the cis-actingelements are indicated within the schematic boxes.
3951The Journal of Immunology
by guest on July 8, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

using Abs directed against the active tyrosine-phosphorylated formof STAT1 also indicated that TPO and IFN-� induced activation ofSTAT1 with similar kinetics (Fig. 5A). We used EMSAs to deter-mine which of the TPO-activated STAT factors actually bound tothe TAP1-GAS regulatory sequence. After stimulation with TPO,STAT1 (Fig. 5B, lanes 2 and 3), but not STAT3 or STAT5A/B(not shown), strongly bound to the TAP1-GAS motif. A similarstrong binding of STAT1 was observed with nuclear extracts fromUT7-mpl cells stimulated with IFN-� (data not shown).
We next analyzed the expression of IRFs by Western blotting.Untreated UT7-mpl cells weakly expressed IRF-1 and highly ex-pressed IRF-2 (Fig. 5A, lane 5). TPO and IFN-� strongly enhancedthe expression of IRF-1 (Fig. 5A, middle panel), whereas the ex-pression of IRF-2 remained unchanged (lower panel). EMSAswere performed to identify the IRFs that bind to the TAP1-ICSs.Because three adjacent TTTC/GAAA core elements lie in this se-quence that may each bind IRFs (see Fig. 4A), we used two dif-ferent oligonucleotides as probes, one encompassing ICS1 andICS2 sequences (designated ICS12) and another encompassingICS2 and GAS (designated ICS2-GAS; see Table I). The ICS12probe was active in binding IRF factors; in the presence of un-treated cell extracts, a large and diffuse band formed with the probe(Fig. 5B, lane 4, see area square bracketed) that could be resolvedin two discrete bands with a longer time of electrophoresis (Fig.5C, lane 1). Upon TPO or IFN-� addition, the complexes increased(Fig. 5B, lanes 4–6; Fig. 5C, lanes 1–3; data not shown). Super-shift analysis using Abs to each of the IRF factors indicated thatIRF-1 and IRF-2 were both present in the large complex observedupon cytokine treatment (Fig. 5B, lanes 7 and 8, see the decreasearea bracketed and apparition of supershifted complexes). IRF-2was the only factor initially bound to the ICS12 probe in cytokine-untreated cell extracts (data not shown). In accordance with itsenhanced expression upon cytokine treatment, binding of IRF-1 tothe probe increased with time, as revealed by supershift assays
with anti-IRF-2 Abs (Fig. 5C, lanes 4–6). We did not detect otherIRF members bound to the probe (data not shown). We next usedthe ICS2-GAS as a probe and remarkably observed no binding ofIRFs to this probe with either untreated or cytokine-treated (TPOor IFN-�) cell extracts even after prolonged cytokine treatments(Fig. 5B, lanes 10–15, see area bracketed; data not shown). In-stead, only STAT1 bound to the probe in a rapid and sustainedmanner, as characterized by supershift assay (lane 17). These dataindicate that ICS2-GAS is not a functional IRF-binding elementdespite the presence of two TTTC core elements, but is an activeSTAT1 binding site.
Overall, this first analysis indicates that cytokine treatment in-duces the recruitment of STAT1 to GAS and IRF-1 to ICS12 se-quences, whereas IRF-2 binds to ICS12 regardless of cytokinetreatment.
Activated STAT1 forms a new complex with IRF-2 and thecytokine-responsive element of TAP1
The complex structure of the cytokine-responsive region of TAP1raised the possibility of a cooperative contribution of the ICSs withthe overlapping GAS. We therefore performed EMSA with a probeencompassing the entire ICS1-ICS2-GAS region, hereafter re-ferred to as ICS12-GAS. Similar to what we observed with ICS12(Fig. 5B, lanes 4–9), a large and diffuse complex bound to theICS12-GAS probe in nuclear extracts from untreated cells (Fig.5B, lane 18, see square bracket) that rapidly increased upon TPOtreatment (Fig. 5B, lane 19). However, two new complexes alsorapidly appeared after a few minutes of stimulation with TPO (Fig.5B, lane 19, and Fig. 6), and were maintained over 3–24 h ofstimulation with no change in their respective proportions (Fig. 6B,lanes 2–6, and data not shown). These two complexes appearedwith the same kinetics. They were similarly induced after IFN-�treatment (data not shown). The lower complex (Fig. 5B, whitearrowhead) migrated exactly as the inducible STAT1 dimer that
FIGURE 5. TPO and IFN-� activate STAT1 and in-duce the expression of IRF-1. A, Western blot analysisof nuclear cell extracts from cells treated for the indi-cated time with TPO or IFN-� was performed usinganti-tyrosine phosphorylated STAT1 (STAT1-PY), anti-IRF-1, or anti-IRF-2 Abs, as indicated on the left. B,EMSAs were performed with probes corresponding toGAS (lanes 1–3), ICS12 (lanes 4–9), ICS2-GAS (lanes10–17), or ICS12-GAS (lanes 18–20) sequences fromthe TAP1 promoter (see schemes on the top of eachpanel) and nuclear extracts from UT7-mpl cells incu-bated with TPO for various periods of time. Supershiftassays were performed using anti-STAT1 (�-S1; lanes3, 9, and 17), anti-IRF-1 (�-I1; lanes 7 and 20), or anti-IRF-2 (�-I2; lane 8) Abs. Alternatively, EMSA wasperformed in the presence of a 100-fold excess of coldICS2-GAS oligonucleotide (ON; lane 16). To better vi-sualize protein-DNA complexes, free probe was run outof the gel. All EMSAs were performed under similarconditions, and gels were run for identical time. Squarebrackets point to monomeric IRF-containing com-plexes. Stars point to supershifted complexes. C,EMSA was similarly performed with extracts fromcells treated for 0–1 h with TPO in the absence (lanes1–3) or the presence of anti-IRF-2 Abs (lanes 4–6)using the ICS12 probe. Gel was subjected to a pro-longed electrophoretic time and exposed to autoradiog-raphy. Only the area where the major monomeric IRF-containing complexes were visible (square bracket) isshown. ns, nonspecific band.
3952 IRF-2 COOPERATES WITH STAT1 TO REGULATE TAP1
by guest on July 8, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
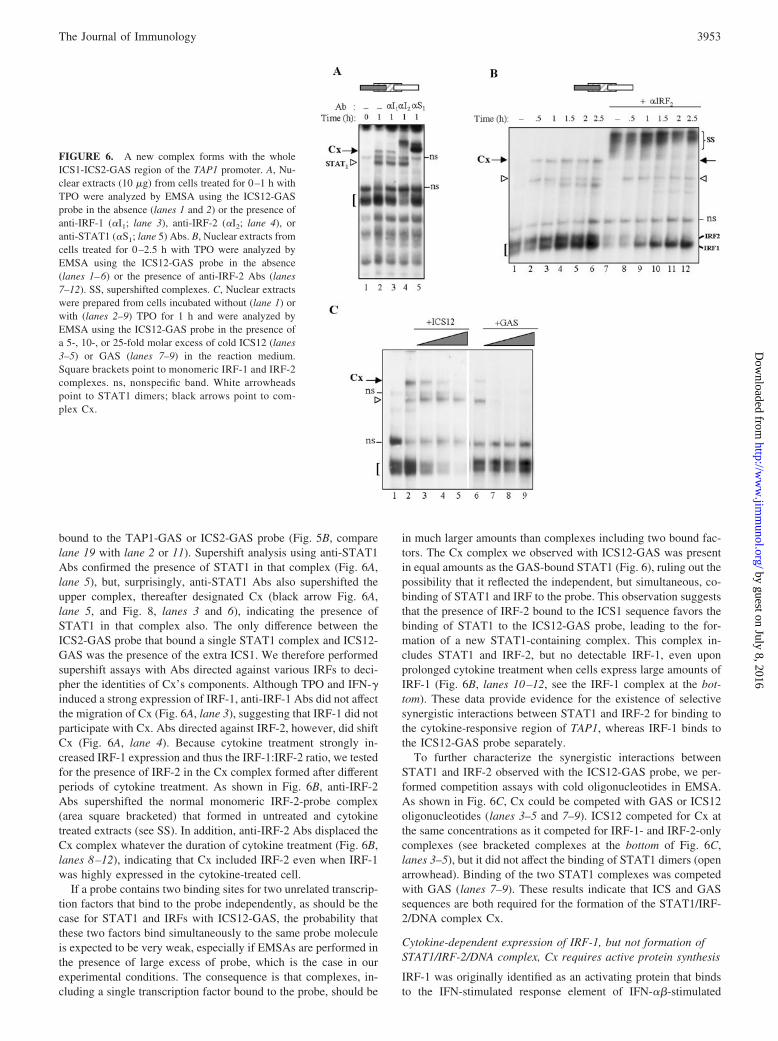
bound to the TAP1-GAS or ICS2-GAS probe (Fig. 5B, comparelane 19 with lane 2 or 11). Supershift analysis using anti-STAT1Abs confirmed the presence of STAT1 in that complex (Fig. 6A,lane 5), but, surprisingly, anti-STAT1 Abs also supershifted theupper complex, thereafter designated Cx (black arrow Fig. 6A,lane 5, and Fig. 8, lanes 3 and 6), indicating the presence ofSTAT1 in that complex also. The only difference between theICS2-GAS probe that bound a single STAT1 complex and ICS12-GAS was the presence of the extra ICS1. We therefore performedsupershift assays with Abs directed against various IRFs to deci-pher the identities of Cx’s components. Although TPO and IFN-�induced a strong expression of IRF-1, anti-IRF-1 Abs did not affectthe migration of Cx (Fig. 6A, lane 3), suggesting that IRF-1 did notparticipate with Cx. Abs directed against IRF-2, however, did shiftCx (Fig. 6A, lane 4). Because cytokine treatment strongly in-creased IRF-1 expression and thus the IRF-1:IRF-2 ratio, we testedfor the presence of IRF-2 in the Cx complex formed after differentperiods of cytokine treatment. As shown in Fig. 6B, anti-IRF-2Abs supershifted the normal monomeric IRF-2-probe complex(area square bracketed) that formed in untreated and cytokinetreated extracts (see SS). In addition, anti-IRF-2 Abs displaced theCx complex whatever the duration of cytokine treatment (Fig. 6B,lanes 8–12), indicating that Cx included IRF-2 even when IRF-1was highly expressed in the cytokine-treated cell.
If a probe contains two binding sites for two unrelated transcrip-tion factors that bind to the probe independently, as should be thecase for STAT1 and IRFs with ICS12-GAS, the probability thatthese two factors bind simultaneously to the same probe moleculeis expected to be very weak, especially if EMSAs are performed inthe presence of large excess of probe, which is the case in ourexperimental conditions. The consequence is that complexes, in-cluding a single transcription factor bound to the probe, should be
in much larger amounts than complexes including two bound fac-tors. The Cx complex we observed with ICS12-GAS was presentin equal amounts as the GAS-bound STAT1 (Fig. 6), ruling out thepossibility that it reflected the independent, but simultaneous, co-binding of STAT1 and IRF to the probe. This observation suggeststhat the presence of IRF-2 bound to the ICS1 sequence favors thebinding of STAT1 to the ICS12-GAS probe, leading to the for-mation of a new STAT1-containing complex. This complex in-cludes STAT1 and IRF-2, but no detectable IRF-1, even uponprolonged cytokine treatment when cells express large amounts ofIRF-1 (Fig. 6B, lanes 10–12, see the IRF-1 complex at the bot-tom). These data provide evidence for the existence of selectivesynergistic interactions between STAT1 and IRF-2 for binding tothe cytokine-responsive region of TAP1, whereas IRF-1 binds tothe ICS12-GAS probe separately.
To further characterize the synergistic interactions betweenSTAT1 and IRF-2 observed with the ICS12-GAS probe, we per-formed competition assays with cold oligonucleotides in EMSA.As shown in Fig. 6C, Cx could be competed with GAS or ICS12oligonucleotides (lanes 3–5 and 7–9). ICS12 competed for Cx atthe same concentrations as it competed for IRF-1- and IRF-2-onlycomplexes (see bracketed complexes at the bottom of Fig. 6C,lanes 3–5), but it did not affect the binding of STAT1 dimers (openarrowhead). Binding of the two STAT1 complexes was competedwith GAS (lanes 7–9). These results indicate that ICS and GASsequences are both required for the formation of the STAT1/IRF-2/DNA complex Cx.
Cytokine-dependent expression of IRF-1, but not formation ofSTAT1/IRF-2/DNA complex, Cx requires active protein synthesis
IRF-1 was originally identified as an activating protein that bindsto the IFN-stimulated response element of IFN-��-stimulated
FIGURE 6. A new complex forms with the wholeICS1-ICS2-GAS region of the TAP1 promoter. A, Nu-clear extracts (10 �g) from cells treated for 0–1 h withTPO were analyzed by EMSA using the ICS12-GASprobe in the absence (lanes 1 and 2) or the presence ofanti-IRF-1 (�I1; lane 3), anti-IRF-2 (�I2; lane 4), oranti-STAT1 (�S1; lane 5) Abs. B, Nuclear extracts fromcells treated for 0–2.5 h with TPO were analyzed byEMSA using the ICS12-GAS probe in the absence(lanes 1–6) or the presence of anti-IRF-2 Abs (lanes7–12). SS, supershifted complexes. C, Nuclear extractswere prepared from cells incubated without (lane 1) orwith (lanes 2–9) TPO for 1 h and were analyzed byEMSA using the ICS12-GAS probe in the presence ofa 5-, 10-, or 25-fold molar excess of cold ICS12 (lanes3–5) or GAS (lanes 7–9) in the reaction medium.Square brackets point to monomeric IRF-1 and IRF-2complexes. ns, nonspecific band. White arrowheadspoint to STAT1 dimers; black arrows point to com-plex Cx.
3953The Journal of Immunology
by guest on July 8, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
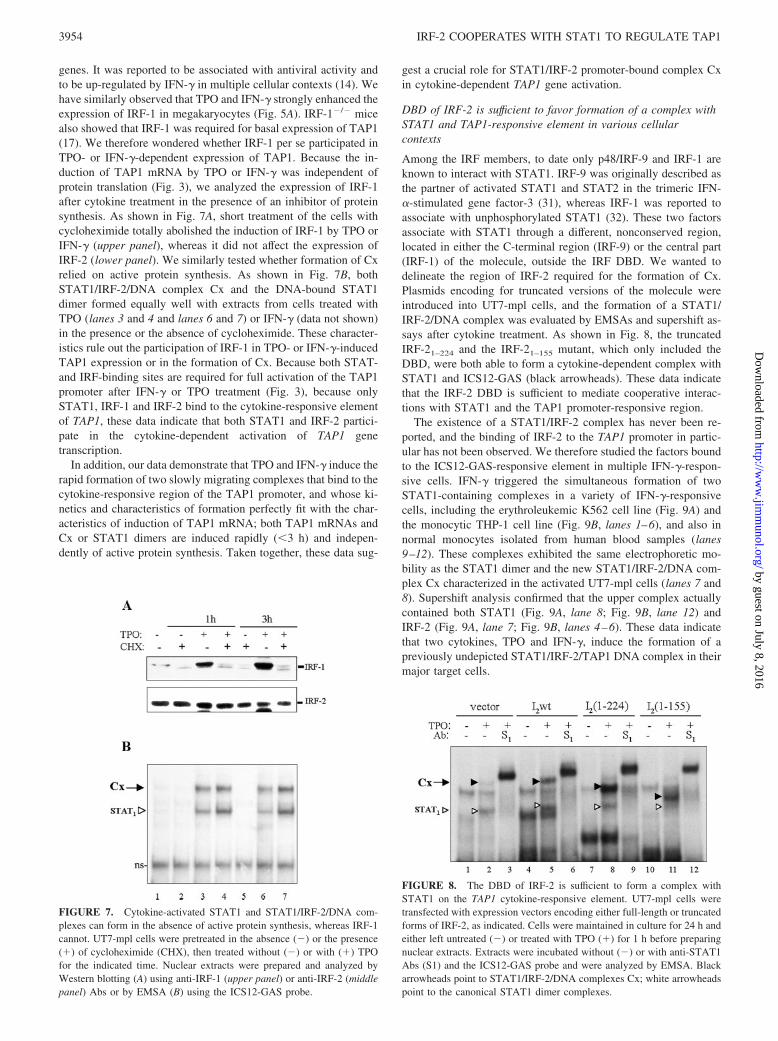
genes. It was reported to be associated with antiviral activity andto be up-regulated by IFN-� in multiple cellular contexts (14). Wehave similarly observed that TPO and IFN-� strongly enhanced theexpression of IRF-1 in megakaryocytes (Fig. 5A). IRF-1�/� micealso showed that IRF-1 was required for basal expression of TAP1(17). We therefore wondered whether IRF-1 per se participated inTPO- or IFN-�-dependent expression of TAP1. Because the in-duction of TAP1 mRNA by TPO or IFN-� was independent ofprotein translation (Fig. 3), we analyzed the expression of IRF-1after cytokine treatment in the presence of an inhibitor of proteinsynthesis. As shown in Fig. 7A, short treatment of the cells withcycloheximide totally abolished the induction of IRF-1 by TPO orIFN-� (upper panel), whereas it did not affect the expression ofIRF-2 (lower panel). We similarly tested whether formation of Cxrelied on active protein synthesis. As shown in Fig. 7B, bothSTAT1/IRF-2/DNA complex Cx and the DNA-bound STAT1dimer formed equally well with extracts from cells treated withTPO (lanes 3 and 4 and lanes 6 and 7) or IFN-� (data not shown)in the presence or the absence of cycloheximide. These character-istics rule out the participation of IRF-1 in TPO- or IFN-�-inducedTAP1 expression or in the formation of Cx. Because both STAT-and IRF-binding sites are required for full activation of the TAP1promoter after IFN-� or TPO treatment (Fig. 3), because onlySTAT1, IRF-1 and IRF-2 bind to the cytokine-responsive elementof TAP1, these data indicate that both STAT1 and IRF-2 partici-pate in the cytokine-dependent activation of TAP1 genetranscription.
In addition, our data demonstrate that TPO and IFN-� induce therapid formation of two slowly migrating complexes that bind to thecytokine-responsive region of the TAP1 promoter, and whose ki-netics and characteristics of formation perfectly fit with the char-acteristics of induction of TAP1 mRNA; both TAP1 mRNAs andCx or STAT1 dimers are induced rapidly (�3 h) and indepen-dently of active protein synthesis. Taken together, these data sug-
gest a crucial role for STAT1/IRF-2 promoter-bound complex Cxin cytokine-dependent TAP1 gene activation.
DBD of IRF-2 is sufficient to favor formation of a complex withSTAT1 and TAP1-responsive element in various cellularcontexts
Among the IRF members, to date only p48/IRF-9 and IRF-1 areknown to interact with STAT1. IRF-9 was originally described asthe partner of activated STAT1 and STAT2 in the trimeric IFN-�-stimulated gene factor-3 (31), whereas IRF-1 was reported toassociate with unphosphorylated STAT1 (32). These two factorsassociate with STAT1 through a different, nonconserved region,located in either the C-terminal region (IRF-9) or the central part(IRF-1) of the molecule, outside the IRF DBD. We wanted todelineate the region of IRF-2 required for the formation of Cx.Plasmids encoding for truncated versions of the molecule wereintroduced into UT7-mpl cells, and the formation of a STAT1/IRF-2/DNA complex was evaluated by EMSAs and supershift as-says after cytokine treatment. As shown in Fig. 8, the truncatedIRF-21–224 and the IRF-21–155 mutant, which only included theDBD, were both able to form a cytokine-dependent complex withSTAT1 and ICS12-GAS (black arrowheads). These data indicatethat the IRF-2 DBD is sufficient to mediate cooperative interac-tions with STAT1 and the TAP1 promoter-responsive region.
The existence of a STAT1/IRF-2 complex has never been re-ported, and the binding of IRF-2 to the TAP1 promoter in partic-ular has not been observed. We therefore studied the factors boundto the ICS12-GAS-responsive element in multiple IFN-�-respon-sive cells. IFN-� triggered the simultaneous formation of twoSTAT1-containing complexes in a variety of IFN-�-responsivecells, including the erythroleukemic K562 cell line (Fig. 9A) andthe monocytic THP-1 cell line (Fig. 9B, lanes 1–6), and also innormal monocytes isolated from human blood samples (lanes9–12). These complexes exhibited the same electrophoretic mo-bility as the STAT1 dimer and the new STAT1/IRF-2/DNA com-plex Cx characterized in the activated UT7-mpl cells (lanes 7 and8). Supershift analysis confirmed that the upper complex actuallycontained both STAT1 (Fig. 9A, lane 8; Fig. 9B, lane 12) andIRF-2 (Fig. 9A, lane 7; Fig. 9B, lanes 4–6). These data indicatethat two cytokines, TPO and IFN-�, induce the formation of apreviously undepicted STAT1/IRF-2/TAP1 DNA complex in theirmajor target cells.
FIGURE 7. Cytokine-activated STAT1 and STAT1/IRF-2/DNA com-plexes can form in the absence of active protein synthesis, whereas IRF-1cannot. UT7-mpl cells were pretreated in the absence (�) or the presence(�) of cycloheximide (CHX), then treated without (�) or with (�) TPOfor the indicated time. Nuclear extracts were prepared and analyzed byWestern blotting (A) using anti-IRF-1 (upper panel) or anti-IRF-2 (middlepanel) Abs or by EMSA (B) using the ICS12-GAS probe.
FIGURE 8. The DBD of IRF-2 is sufficient to form a complex withSTAT1 on the TAP1 cytokine-responsive element. UT7-mpl cells weretransfected with expression vectors encoding either full-length or truncatedforms of IRF-2, as indicated. Cells were maintained in culture for 24 h andeither left untreated (�) or treated with TPO (�) for 1 h before preparingnuclear extracts. Extracts were incubated without (�) or with anti-STAT1Abs (S1) and the ICS12-GAS probe and were analyzed by EMSA. Blackarrowheads point to STAT1/IRF-2/DNA complexes Cx; white arrowheadspoint to the canonical STAT1 dimer complexes.
3954 IRF-2 COOPERATES WITH STAT1 TO REGULATE TAP1
by guest on July 8, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

In the course of these studies we observed that human embry-onic kidney 293T cells expressed undetectable levels of activeIRF-1 or IRF-2 by EMSA (see area bracketed, Fig. 9B, lanes 13and 14). IFN-� only triggered the binding of STAT1 dimers to theICS12-GAS probe, but the STAT1/IRF-2 complex Cx was notdetected (lane 14). We introduced an expression vector for IRF-2and treated the 293T cells with IFN-�. In these transfected cells,IFN-� triggered the formation of two DNA-bound complexes sim-ilar to those observed in UT7-mpl cells (lanes 15 and 16), whichboth included STAT1 (data not shown). This indicates that the solepresence of IRF-2 allows the formation of a STAT1/IRF-2-con-taining complex with the TAP1 cytokine-responsive element. Wealso introduced the vectors encoding the same truncated forms ofIRF-2 as described above (Fig. 8) and stimulated the cells withIFN-�. We similarly observed that the IRF-2 DBD is sufficient formediating cooperative interactions with IFN-�-activated STAT1and the TAP1 promoter-responsive region in this cellular context(data not shown).
Overall, these data indicate that the sole DBD of IRF-2 cancooperate with STAT1 for binding to the cytokine-responsive re-gion of TAP1 after stimulation with two different cytokines (TPOand IFN-�) in two different cellular contexts (hemopoietic andepithelial).
IRF-2 activates TAP1 gene promoter
IRF-2 has never been reported to regulate TAP1 expression. More-over, IRF-2 was originally described as a transcriptional repressorthat antagonized IRF-1 activity by competing for binding to the
ICS of IFNs and IFN-inducible genes (14). However, recent evi-dence indicates that IRF-2 can act as a dual function transcriptionfactor, because it also activates the transcription of some genes(33, 34). Therefore, to ascertain the role of IRF-2 in the regulationof TAP1 promoter activity, we evaluated the biological contribu-tion of IRF-2 to the activity of the TAP1 promoter in two cellularcontexts. We performed transient transfection assays with theTAP1 promoter reporter construct in the presence of an IRF-2expression vector, both introduced either in 293T cells, which ex-press undetectable amounts of IRF-1/2-binding activity (see above,Fig. 9B), or in MEF from IRF-1�/� IRF-2�/� double-knockoutmice devoid of IRF-1 and IRF-2 proteins. As shown in Fig. 10,IRF-2 increased the activity of the TAP1 gene promoter in a dose-dependent manner in both 293T cells and IRF-1�/�IRF-2�/�
MEF. These data indicate that IRF-2 positively regulates TAP1promoter activity.
DiscussionOur results first show that TPO activates the expression of func-tional TAP1 in TPO-responsive hemopoietic cells, with the con-sequence of enhanced expression of class I MHC on the cell sur-face. This expression of TAP1 and class I MHC molecules was ofthe same magnitude as that triggered by IFN-� in the same cells.These observations raise the possibility that TPO-responsive cellssuch as megakaryocytes directly participate in the immune re-sponse. Although the main functional role attributed to plateletswas hemostasis and maintenance of the integrity of blood vesselwall, it has become apparent that platelets also play a prominent
FIGURE 9. DNA-bound IRF-2 and IFN-�-activatedSTAT1 form a complex with the TAP1 cytokine-re-sponsive element in a variety of cellular contexts. K562(A) or THP-1 (B, lanes 1–6), UT7-mpl cells (B, lanes 7and 8), and peripheral monocytes (B, lanes 9–12) wereincubated with IFN-� as indicated on the top of eachpanel. Nuclear extracts were prepared and analyzed byEMSA in the absence or the presence of the indicatedAbs, using the ICS12-GAS probe. Alternatively, 293Tcells were transfected with empty vector (B, lanes 13and 14) or IRF-2 expression vector (lanes 15 and 16)and maintained in culture for 24 h. Where indicated (1),IFN-� was added for 1 h before the end of the culture.Nuclear extracts were prepared and analyzed by EMSAusing the ICS12-GAS probe. Black arrowheads showthe STAT1/IRF-2/DNA complexes (Cx) formed witheach type of extracts; empty arrowheads show the ca-nonical STAT1 dimer. Monomeric IRF-1 or IRF-2complexes are indicated by square brackets.
3955The Journal of Immunology
by guest on July 8, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

role in modulating inflammation. Upon activation, platelets releasechemokines and cytokines and, upon binding to endothelial cells,induce the production of a broad array of inflammatory molecules(35–38). Their capacity to function during host defense againstinfection has recently received attention (37). Platelets expressHLA molecules. Taking into account platelet number, it was esti-mated that approximately two-thirds of the HLA molecules fromblood were carried by platelets. The majority of these HLA mol-ecules were reported to be of endogenous origin, and it was gen-erally assumed that this synthesis occurred at the megakaryocytestage (39). Moreover, activated platelets can bind and phagocytosevarious infectious microorganisms, and engulfing vacuoles werereported to fuse with �-granules that contain microbicidal proteins(40–42). These interactions can induce degradation of pathogens.Moreover, viral Ags subsequently appeared on the surface of thecells (38). Platelet concentrates were also shown to have a viru-cidal effect, and they released potent antimicrobial peptides thatkilled pathogens (37, 41). All these properties are expected formegakaryocytes, which can be activated and are known to containmultivesicular bodies and secretory granules and to degranulateexactly as platelets do, and are known to be a direct target of HIVinfection (40, 43, 44). These processes may favor the expression ofantigenic peptides. In addition, platelets were recently shown toactively participate in innate and adaptive immunity via theirCD154/CD40L that induced dendritic cell maturation, B cell iso-type switching, and enhanced CD8� T cell response both in vitroand in vivo (45). Moreover, depletion of platelets in normal miceresults in decreased Ag-specific Ab production (45). High expres-sion of TAP1 and class I MHC in megakaryocytes or platelets maythus reflect an additional role of this cell lineage in host defense.
Our study searched for the mechanism(s) responsible for theenhanced expression of TAP1 observed upon IFN-� or TPO treat-ment of megakaryocytic cells. It provides evidence that these twocytokines activate the expression of TAP1 through a novel mech-anism which involves STAT1 and the un-expected participation ofIRF-2. Numerous complementary observations support this con-clusion. First, TPO and IFN-� activate the expression of TAP1 atthe transcriptional level and the enhanced TAP1 promoter activityinduced by the two cytokines depends on the presence of both afunctional GAS and an active IRF-binding site (ICS12). Second,this ICS12 only binds IRF-1 and IRF-2, but no other members ofthe IRF family in vitro, therefore suggesting that IRF-1, IRF-2, orboth regulate TAP1 promoter activity after cytokine treatment.
Third, the accumulation of TAP1 mRNA induced by IFN-�/TPOdoes not require the presence of an active translation machinery,whereas IFN-�- and TPO-dependent expression of IRF-1 requiresactive protein synthesis, thereby ruling out the participation ofIRF-1 in the cytokine-dependent activation of TAP1. Fourth, TPOand IFN-� rapidly activate STAT1. In accordance with the kineticsof TAP1 mRNA induction, cytokine-activated STAT1 forms twotypes of complexes with the cytokine-responsive element of theTAP1 promoter in vitro: 1) a STAT1 dimer, which is the solecomplex bound to the isolated TAP1-GAS element; and 2) a novelSTAT1/IRF-2/DNA complex that is recognized by both anti-STAT1 and anti-IRF-2 Abs and whose formation follows the samekinetics as STAT1 dimers, thus indicating cooperative binding ofIRF-2 and STAT1 to the cytokine-responsive region. Fifth, IRF-2per se can transactivate the TAP1 gene. Finally, we show thatDNA-bound IRF-2 cooperates for the binding of TPO/IFN-�-ac-tivated STAT1 to the TAP1-responsive element in vitro in all theTPO/IFN-�-responsive cell types we have tested, including circu-lating monocytes, the major normal IFN-� target cells, suggestingthat IRF-2 activates the TAP1 gene promoter after TPO or IFN-�in all their responsive cells.
Previous studies of IRF-1-deficient mice have emphasized thecrucial role of IRF-1 for the basal expression of TAP1 (17). How-ever, IFN-�-dependent expression of TAP1 was unaffected inthese IRF-1�/� animals. Our data extend these preliminary obser-vations and rule out the participation of IRF-1 in IFN-�-dependentexpression of TAP1, because that expression is independent ofprotein synthesis. They, moreover, show that IRF-2 participates inTAP1 gene expression as a basal transactivator, and also contrib-utes selectively to the extra cytokine-dependent activation of theTAP1 gene because both GAS and ICS are required to activateTAP1 promoter. Furthermore, IRF-2 also cooperates for the re-cruitment of active STAT1 to the promoter.
Although the specific binding of STAT1 to TAP1-GAS and thatof IRF-1 to ICS have been observed in multiple cellular contexts,the nuclear factors bound to the entire ICS12-GAS region havenever been analyzed. Because of the presence of overlapping ICSand GAS elements within the TAP1 promoter, steric clashes, syn-ergies, or changes in selectivity for DNA binding between tran-scription factors were expected. We demonstrate that the ICS12sequence when linked to GAS allows the formation of a new com-plex composed of STAT1 and IRF-2, with no detectable IRF-1.This result was not expected, because TPO and IFN-� trigger therapid synthesis of large amounts of IRF-1, which binds to ICS12or ICS12-GAS elements individually, as detected in EMSA. It thusindicates that IRF-2 contains a specific element that allows selec-tive cooperative binding of STAT1 and IRF-2 on the TAP1promoter.
The reason for this cooperation is presently unknown, but wepropose three hypotheses. First, IRF-2 may directly interact withSTAT1. STAT1 has already been reported to interact with IRF-1,but this interaction involves the rather specific central region ofIRF-1, outside the DBD. Remarkably, this interaction involves un-phosphorylated STAT1, and IFN-� does not affect this basal in-teraction, suggesting that IRF-1 cannot form a more stable com-plex with a cytokine-activated STAT1 (32). IRF-9 was alsooriginally described as the partner of activated STAT1 and STAT2in the trimeric IFN-�-stimulated gene factor-3 complex (31), butthis factor associates with STAT1 through a different, noncon-served region, located in the C-terminal region of the molecule.We now show that the sole DBD of IRF-2 is sufficient to allowcooperative binding of STAT1 to the TAP1 promoter. This domainis highly conserved among IRF members and exhibits a high ho-mology (84%) with IRF-1, making the presence of a selective and
FIGURE 10. IRF-2 activates the TAP1 promoter. 293T cells (left panel)or IRF-1�/�IRF-2�/� MEF (right panel) were transiently transfected withthe wild-type TAP1 promoter luciferase reporter (200 ng) along with theindicated quantity of a vector encoding wild-type IRF-2. Cells were main-tained for 18 h in culture before measuring luciferase activity. All plasmidswere transfected a minimum of five times in separate experiments. Thefigure shows one representative experiment with each cell type.
3956 IRF-2 COOPERATES WITH STAT1 TO REGULATE TAP1
by guest on July 8, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

restricted association of IRF-2 DBD with activated STAT1 quiteunexpected. All our attempts to detect possible association ofSTAT1 with IRF-2 in solution failed, suggesting that free IRF-2may be unable to interact with activated STAT1. These data led usto favor a second hypothesis. IRF-2 is known to induce particularlystrong DNA bending, which is responsible for efficient interactionsof DNA-bound IRF-2 with a second IRF-2 molecule (16) or withIRF-8/ICSBP (46). Such bending of DNA sequences was observedwith the sole DBD of IRF-2, whereas the highly homologous DBDof IRF-1 was without effect (15, 16). We thus propose that bindingof IRF-2 to the TAP1 gene induces a selective DNA conformationthat enhances the recruitment of STAT1 to neighboring DNA se-quences. Third, and not mutually exclusive with the second hy-pothesis, the binding of IRF-2 to DNA may modify IRF-2 con-formation in such a way that it exposes a STAT1 interactiondomain, or even an interaction domain for a partner (transcriptionregulator, coactivators, etc.) shared with the activated STAT1 thatwould thus, indirectly, favor the association of STAT1 to DNA-bound IRF-2.
IRF-2 was originally described as an antagonist of IRF-1-me-diated regulation of IFN-inducible genes by competition for bind-ing at the same DNA locus. IRF-1 was shown to induce the ex-pression of IFN-�� and to have antioncogenic activity, whereasIRF-2 repressed these actions (14). In addition to its repressiveeffects on IFN-related genes, IRF-2 can activate the transcriptionof a set of genes, such as histone H4 and VCAM-1 genes (33, 47,48), whereas IRF-1 cannot. IRF-2 also synergizes with IRF-1 inactivating two well-known IFN-� target genes, CIITA and guany-late-binding protein (34, 49). Surprisingly, IRF-1- and IRF-2-de-ficient mice were shown to have remarkably similar phenotypes interms of their defect in Th1 differentiation, NK cell development,and cytokine secretion profiles, suggesting that IRF-2, like IRF-1,mediates part of its effect through direct transcriptional activationand confirming a specific role for IRF-2 in the immune system (50,51). Moreover, recent studies have highlighted a role for IRFs inhemopoietic differentiation and shown that IRF-1 favors granulo-cytic differentiation, whereas IRF-2 drives megakaryocytic differ-entiation (52). Our results demonstrate that IRF-2 can transactivatea gene whose product directly participates in immune recognition.IRF-2 has long been described to possess both a transcriptionalrepression domain and a latent activation domain (53), consistentwith the negative and positive regulation of gene expression bythis factor. IRF-2 was recently shown to interact with histoneacetylases, such as p300 and pCAF, originally known as globalcoactivators and members of the transcription machinery and to behighly acetylated compared with IRF-1 (54, 55). Moreover, a newbromo domain protein that exhibits similarities with transcriptionalcoactivators was identified that interacts with IRF-2 (56). Some ofthese partners also interact with STAT1 (p300/CBP) and may pro-vide the active link responsible for DNA-binding cooperation be-tween STAT1 and prebound IRF-2. All these partners may partic-ipate in the IRF-2-dependent activation of the TAP1 promoter.
Overall, our data provide the first evidence that the cytokineTPO, as the inflammatory cytokine IFN-�, can enhance the ex-pression of TAP1 in megakaryocytes. It, moreover, demonstratesthat both IRF-2 and STAT1 play active roles in TPO- and IFN-�-mediated TAP1 promoter activation. We propose that constitutivebinding of IRF-2 to the TAP1 promoter enhances the selectiverecruitment of activated STAT1 and favors subsequent functionaland positive associations with the basal transcription machinery.
AcknowledgmentsWe thank Dr. D. Johnson (New Haven, CT) for providing the wild-typeTAP1 promoter construct, and Dr. P. M. van Endert (Paris, France) and
Dr. R. Tampe (Frankfurt/Main, Germany) for providing anti-TAP1 Abs;and Drs. L. Benit, V. Mignotte, and P.-H. Romeo for helpful comments onthe manuscript.
DisclosuresThe authors have no financial conflict of interest.
References1. Yewdell, J. W., and J. R. Bennink. 1992. Cell biology of antigen processing and
presentation to major histocompatibility complex class I molecule-restricted Tlymphocytes. Adv. Immunol. 52:1.
2. Groettrup, M., A. Soza, U. Kuckelkorn, and P. M. Kloetzel. 1996. Peptide antigenproduction by the proteasome: complexity provides efficiency. Immunol. Today17:429.
3. Neefjes, J. J., F. Momburg, and G. J. Hammerling. 1993. Selective and ATP-dependent translocation of peptides by the MHC-encoded transporter. Science261:769.
4. Owen, B. A., and L. R. Pease. 1999. TAP association influences the conformationof nascent MHC class I molecules. J. Immunol. 162:4677.
5. Johnson, D. R., and B. Mook-Kanamori. 2000. Dependence of elevated humanleukocyte antigen class I molecule expression on increased heavy chain, lightchain (�2-microglobulin), transporter associated with antigen processing, tapasin,and peptide. J. Biol. Chem. 275:16643.
6. Spies, T., and R. DeMars. 1991. Restored expression of major histocompatibilityclass I molecules by gene transfer of a putative peptide transporter. Nature351:323.
7. de la Salle, H., D. Hanau, D. Fricker, A. Urlacher, A. Kelly, J. Salamero,S. H. Powis, L. Donato, H. Bausinger, M. Laforet, et al. 1994. Homozygoushuman TAP peptide transporter mutation in HLA class I deficiency. Science265:237.
8. Blanchet, O., J. F. Bourge, H. Zinszner, A. Israel, P. Kourilsky, J. Dausset,L. Degos, and P. Paul. 1992. Altered binding of regulatory factors to HLA classI enhancer sequence in human tumor cell lines lacking class I antigen expression.Proc. Natl. Acad. Sci. USA 89:3488.
9. Rotem-Yehudar, R., M. Groettrup, A. Soza, P. M. Kloetzel, and R. Ehrlich. 1996.LMP-associated proteolytic activities and TAP-dependent peptide transport forclass 1 MHC molecules are suppressed in cell lines transformed by the highlyoncogenic adenovirus 12. J. Exp. Med. 183:499.
10. Wright, K. L., L. C. White, A. Kelly, S. Beck, J. Trowsdale, and J. P. Ting. 1995.Coordinate regulation of the human TAP1 and LMP2 genes from a shared bidi-rectional promoter. J. Exp. Med. 181:1459.
11. Min, W., J. S. Pober, and D. R. Johnson. 1996. Kinetically coordinated inductionof TAP1 and HLA class I by IFN-�: the rapid induction of TAP1 by IFN-� ismediated by Stat1�. J. Immunol. 156:3174.
12. Cramer, L. A., S. L. Nelson, and M. J. Klemsz. 2000. Synergistic induction of theTap-1 gene by IFN-� and lipopolysaccharide in macrophages is regulated bySTAT1. J. Immunol. 165:3190.
13. Chatterjee-Kishore, M., R. Kishore, D. J. Hicklin, F. M. Marincola, andS. Ferrone. 1998. Different requirements for signal transducer and activator oftranscription 1� and interferon regulatory factor 1 in the regulation of low mo-lecular mass polypeptide 2 and transporter associated with antigen processing 1gene expression. J. Biol. Chem. 273:16177.
14. Taniguchi, T., K. Ogasawara, A. Takaoka, and N. Tanaka. 2001. Irf family oftranscription factors as regulators of host defense. Annu. Rev. Immunol. 19:623.
15. Escalante, C. R., J. Yie, D. Thanos, and A. K. Aggarwal. 1998. Structure of IRF-1with bound DNA reveals determinants of interferon regulation. Nature 391:103.
16. Fujii, Y., T. Shimizu, M. Kusumoto, Y. Kyogoku, T. Taniguchi, andT. Hakoshima. 1999. Crystal structure of an IRF-DNA complex reveals novelDNA recognition and cooperative binding to a tandem repeat of core sequences.EMBO J. 18:5028.
17. White, L. C., K. L. Wright, N. J. Felix, H. Ruffner, L. F. Reis, R. Pine, andJ. P. Ting. 1996. Regulation of LMP2 and TAP1 genes by IRF-1 explains thepaucity of CD8� T cells in IRF-1�/� mice. Immunity 5:365.
18. Kaushansky, K. 1999. The enigmatic megakaryocyte gradually reveals its secrets.BioEssays 21:353.
19. Kaushansky, K. 1999. Thrombopoietin and hematopoietic stem cell development.Ann. NY Acad. Sci. 872:314.
20. Souyri, M., I. Vigon, J. F. Penciolelli, J. M. Heard, P. Tambourin, andF. Wendling. 1990. A putative truncated cytokine receptor gene transduced by themyeloproliferative leukemia virus immortalizes hematopoietic progenitors. Cell63:1137.
21. Matsumura, I., K. Nakajima, H. Wakao, S. Hattori, K. Hashimoto, H. Sugahara,T. Kato, H. Miyazaki, T. Hirano, and Y. Kanakura. 1998. Involvement of pro-longed ras activation in thrombopoietin-induced megakaryocytic differentiationof a human factor-dependent hematopoietic cell line. Mol. Cell. Biol. 18:4282.
22. Porteu, F., M. C. Rouyez, L. Cocault, L. Benit, M. Charon, F. Picard,S. Gisselbrecht, M. Souyri, and I. Dusanter-Fourt. 1996. Functional regions of themouse thrombopoietin receptor cytoplasmic domain: evidence for a critical re-gion which is involved in differentiation and can be complemented by erythro-poietin. Mol. Cell. Biol. 16:2473.
23. Cwirla, S. E., P. Balasubramanian, D. J. Duffin, C. R. Wagstrom, C. M. Gates,S. C. Singer, A. M. Davis, R. L. Tansik, L. C. Mattheakis, C. M. Boytos, et al.1997. Peptide agonist of the thrombopoietin receptor as potent as the naturalcytokine. Science 276:1696.
3957The Journal of Immunology
by guest on July 8, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

24. van Endert, P. M., R. Tampe, T. H. Meyer, R. Tisch, J. F. Bach, andH. O. McDevitt. 1994. A sequential model for peptide binding and transport bythe transporters associated with antigen processing. Immunity 1:491.
25. Magne, S., S. Caron, M. Charon, M. C. Rouyez, and I. Dusanter-Fourt. 2003.STAT5 and Oct-1 form a stable complex that modulates cyclin D1 expression.Mol. Cell. Biol. 23:8934.
26. Caron, S., M. Charon, E. Cramer, N. Sonenberg, and I. Dusanter-Fourt. 2004.Selective modification of eukaryotic initiation factor 4F (eIF4F) at the onset ofcell differentiation: recruitment of eIF4GII and long-lasting phosphorylation ofeIF4E. Mol. Cell. Biol. 24:4920.
27. Fichelson, S., J. M. Freyssinier, F. Picard, M. Fontenay-Roupie, M. Guesnu,M. Cherai, S. Gisselbrecht, and F. Porteu. 1999. Megakaryocyte growth anddevelopment factor-induced proliferation and differentiation are regulated by themitogen-activated protein kinase pathway in primitive cord blood hematopoieticprogenitors. Blood 94:1601.
28. Ma, W., P. J. Lehner, P. Cresswell, J. S. Pober, and D. R. Johnson. 1997. Inter-feron-� rapidly increases peptide transporter (TAP) subunit expression and pep-tide transport capacity in endothelial cells. J. Biol. Chem. 272:16585.
29. Kaushansky, K. 1999. Thrombopoietin: from theory to reality. Thromb. Haemost.82:312.
30. Pallard, C., F. Gouilleux, L. Benit, L. Cocault, M. Souyri, D. Levy, B. Groner,S. Gisselbrecht, and I. Dusanter-Fourt. 1995. Thrombopoietin activates a STAT5-like factor in hematopoietic cells. EMBO J. 14:2847.
31. Horvath, C. M., G. R. Stark, I. M. Kerr, and J. E. Darnell, Jr. 1996. Interactionsbetween STAT and non-STAT proteins in the interferon-stimulated gene factor 3transcription complex. Mol. Cell. Biol. 16:6957.
32. Chatterjee-Kishore, M., K. L. Wright, J. P. Ting, and G. R. Stark. 2000. HowStat1 mediates constitutive gene expression: a complex of unphosphorylatedStat1 and IRF1 supports transcription of the LMP2 gene. EMBO J. 19:4111.
33. Vaughan, P. S., F. Aziz, A. J. van Wijnen, S. Wu, H. Harada, T. Taniguchi,K. J. Soprano, J. L. Stein, and G. S. Stein. 1995. Activation of a cell-cycle-regulated histone gene by the oncogenic transcription factor IRF-2. Nature377:362.
34. Xi, H., D. D. Eason, D. Ghosh, S. Dovhey, K. L. Wright, and G. Blanck. 1999.Co-occupancy of the interferon regulatory element of the class II transactivator(CIITA) type IV promoter by interferon regulatory factors 1 and 2. Oncogene18:5889.
35. Majka, M., T. Rozmyslowicz, B. Lee, S. L. Murphy, Z. Pietrzkowski,G. N. Gaulton, L. Silberstein, and M. Z. Ratajczak. 1999. Bone marrow CD34�
cells and megakaryoblasts secrete �-chemokines that block infection of hemato-poietic cells by M-tropic R5 HIV. J. Clin. Invest. 104:1739.
36. Lindemann, S., N. D. Tolley, D. A. Dixon, T. M. McIntyre, S. M. Prescott,G. A. Zimmerman, and A. S. Weyrich. 2001. Activated platelets mediate inflam-matory signaling by regulated interleukin 1� synthesis. J. Cell Biol. 154:485.
37. Klinger, M. H., and W. Jelkmann. 2002. Role of blood platelets in infection andinflammation. J. Interferon Cytokine Res. 22:913.
38. Yeaman, M. R. 1997. The role of platelets in antimicrobial host defense. Clin.Infect. Dis. 25:951.
39. Gouttefangeas, C., M. Diehl, W. Keilholz, R. F. Hornlein, S. Stevanovic, andH. G. Rammensee. 2000. Thrombocyte HLA molecules retain nonrenewable en-dogenous peptides of megakaryocyte lineage and do not stimulate direct allocy-totoxicity in vitro. Blood 95:3168.
40. Youssefian, T., A. Drouin, J. M. Masse, J. Guichard, and E. M. Cramer. 2002.Host defense role of platelets: engulfment of HIV and Staphylococcus aureusoccurs in a specific subcellular compartment and is enhanced by platelet activa-tion. Blood 99:4021.
41. Maurice, A., M. Marchand-Arvier, D. Edert, A. Le Faou, G. Gondrexon, andC. Vigneron. 2002. The virucidal effect of platelet concentrates: preliminarystudy and first conclusions. Platelets 13:219.
42. Krijgsveld, J., S. A. Zaat, J. Meeldijk, P. A. van Veelen, G. Fang, B. Poolman,E. Brandt, J. E. Ehlert, A. J. Kuijpers, G. H. Engbers, et al. 2000. Thrombocidins,microbicidal proteins from human blood platelets, are C-terminal deletion prod-ucts of CXC chemokines. J. Biol. Chem. 275:20374.
43. Louache, F., A. Bettaieb, A. Henri, E. Oksenhendler, J. P. Farcet, P. Bierling,M. Seligmann, and W. Vainchenker. 1991. Infection of megakaryocytes by hu-man immunodeficiency virus in seropositive patients with immune thrombocy-topenic purpura. Blood 78:1697.
44. Cramer, E. M., J. M. Masse, J. P. Caen, I. Garcia, J. Breton-Gorius, N. Debili, andW. Vainchenker. 1993. Effect of thrombin on maturing human megakaryocytes.Am. J. Pathol. 143:1498.
45. Elzey, B. D., J. Tian, R. J. Jensen, A. K. Swanson, J. R. Lees, S. R. Lentz,C. S. Stein, B. Nieswandt, Y. Wang, B. L. Davidson, et al. 2003. Platelet-medi-ated modulation of adaptive immunity: a communication link between innate andadaptive immune compartments. Immunity 19:9.
46. Meraro, D., S. Hashmueli, B. Koren, A. Azriel, A. Oumard, S. Kirchhoff,H. Hauser, S. Nagulapalli, M. L. Atchison, and B. Z. Levi. 1999. Protein-proteinand DNA-protein interactions affect the activity of lymphoid-specific IFN regu-latory factors. J. Immunol. 163:6468.
47. Vaughan, P. S., C. M. van der Meijden, F. Aziz, H. Harada, T. Taniguchi,A. J. van Wijnen, J. L. Stein, and G. S. Stein. 1998. Cell cycle regulation ofhistone H4 gene transcription requires the oncogenic factor IRF-2. J. Biol. Chem.273:194.
48. Jesse, T. L., R. LaChance, M. F. Iademarco, and D. C. Dean. 1998. Interferonregulatory factor-2 is a transcriptional activator in muscle where it regulatesexpression of vascular cell adhesion molecule-1. J. Cell Biol. 140:1265.
49. Xi, H., B. Goodwin, A. T. Shepherd, and G. Blanck. 2001. Impaired class IItransactivator expression in mice lacking interferon regulatory factor-2. Onco-gene 20:4219.
50. Matsuyama, T., T. Kimura, M. Kitagawa, K. Pfeffer, T. Kawakami, N. Watanabe,T. M. Kundig, R. Amakawa, K. Kishihara, A. Wakeham, et al. 1993. Targeteddisruption of IRF-1 or IRF-2 results in abnormal type I IFN gene induction andaberrant lymphocyte development. Cell 75:83.
51. Lohoff, M., G. S. Duncan, D. Ferrick, H. W. Mittrucker, S. Bischof, S. Prechtl,M. Rollinghoff, E. Schmitt, A. Pahl, and T. W. Mak. 2000. Deficiency in thetranscription factor interferon regulatory factor (IRF)-2 leads to severely com-promised development of natural killer and T helper type 1 cells. J. Exp. Med.192:325.
52. Stellacci, E., U. Testa, E. Petrucci, E. Benedetti, R. Orsatti, T. Feccia,M. Stafsnes, E. M. Coccia, G. Marziali, and A. Battistini. 2004. Interferon reg-ulatory factor-2 drives megakaryocytic differentiation. Biochem. J. 377:367.
53. Yamamoto, H., M. S. Lamphier, T. Fujita, T. Taniguchi, and H. Harada. 1994.The oncogenic transcription factor IRF-2 possesses a transcriptional repressionand a latent activation domain. Oncogene 9:1423.
54. Masumi, A., Y. Yamakawa, H. Fukazawa, K. Ozato, and K. Komuro. 2003.Interferon regulatory factor-2 regulates cell growth through its acetylation.J. Biol. Chem. 278:25401.
55. Masumi, A., and K. Ozato. 2001. Coactivator p300 acetylates the interferon reg-ulatory factor-2 in U937 cells following phorbol ester treatment. J. Biol. Chem.276:20973.
56. Staal, A., J. M. Enserink, J. L. Stein, G. S. Stein, and A. J. van Wijnen. 2000.Molecular characterization of celtix-1, a bromodomain protein interacting withthe transcription factor interferon regulatory factor 2. J. Cell. Physiol. 185:269.
3958 IRF-2 COOPERATES WITH STAT1 TO REGULATE TAP1
by guest on July 8, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















