
Role of the conserved arginine 507 and glutamate 727 residues
Upload
independentCategory
view
1download
0
10.1128/MCB.23.11.3918-3928.2003.
2003, 23(11):3918. DOI:Mol. Cell. Biol. Martha K. CathcartDavid Anthony, David A. Frank, Gerald M. Feldman and Bo Xu, Ashish Bhattacharjee, Biswajit Roy, Hong-Min Xu, Stat3Serine 727 Phosphorylation of Stat1 and Mitogen-Activated Protein Kinase-MediatedGene Expression Requires p38 Interleukin-13 Induction of 15-Lipoxygenase
http://mcb.asm.org/content/23/11/3918Updated information and services can be found at:
These include:
REFERENCEShttp://mcb.asm.org/content/23/11/3918#ref-list-1at:
This article cites 63 articles, 42 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on October 24, 2014 by guest
http://mcb.asm
.org/D
ownloaded from
on O
ctober 24, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

MOLECULAR AND CELLULAR BIOLOGY, June 2003, p. 3918–3928 Vol. 23, No. 110270-7306/03/$08.00�0 DOI: 10.1128/MCB.23.11.3918–3928.2003Copyright © 2003, American Society for Microbiology. All Rights Reserved.
Interleukin-13 Induction of 15-Lipoxygenase Gene ExpressionRequires p38 Mitogen-Activated Protein Kinase-Mediated
Serine 727 Phosphorylation of Stat1 and Stat3Bo Xu,1 Ashish Bhattacharjee,1 Biswajit Roy,1 Hong-Min Xu,1 David Anthony,1
David A. Frank,2 Gerald M. Feldman,3 and Martha K. Cathcart1*Department of Cell Biology, Lerner Research Institute, Cleveland Clinic Foundation, Cleveland, Ohio 441951; Department of
Adult Oncology, Dana-Farber Cancer Institute, Harvard Medical School, Boston, Massachusetts 021152; andDivision of Monoclonal Antibodies, Office of Therapeutics, Research and Review, Center for Biologics
Evaluation and Research, Food and Drug Administration, Bethesda, Maryland 208923
Received 2 October 2002/Returned for modification 19 November 2002/Accepted 12 March 2003
Interleukin-13 (IL-13) is a cytokine secreted by Th2 lymphocytes that is capable of inducing expression of15-lipoxygenase (15-LO) in primary human monocytes. We recently demonstrated that induction of 15-LOrequires the activation of Jak2 and Tyk2 kinases and Stats 1, 3, 5, and 6. Since IL-13-induced 15-LO expressionwas inhibited by H7 (a serine-threonine kinase inhibitor), we predicted that Stat serine phosphorylation mayalso be crucial for 15-LO expression. In this study, we present evidence indicating that IL-13-induced 15-LOmRNA expression was detectable as early as 1 h by real-time reverse transcription-PCR. We found that IL-13induced a time-dependent serine phosphorylation of both Stat1 and Stat3, detectable at 15 min after IL-13treatment. In addition, the activation of p38 mitogen-activated protein kinase (MAPK) was detected in atime-dependent fashion, with peak phosphorylation at 15 min after IL-13 treatment. SB202190, a p38 MAPK-specific inhibitor, markedly inhibited IL-13-induced Stat1 and Stat3 serine phosphorylation as well as DNAbinding. Furthermore, treatment of cells with Stat1 or Stat3 decoys significantly impaired IL-13-induced 15-LOexpression. Taken together, our results provide the first evidence that IL-13 induces p38 MAPK phosphory-lation/activation, which regulates Stat1 and Stat3 serine 727 phosphorylation. Both of these events areimportant steps in IL-13-induced 15-LO expression in human monocytes.
Monocytes are one of the unique cell types that can respondto the T lymphocyte-derived cytokines interleukin (IL)-4 andIL-13 (14). One of the novel proteins induced upon monocyteexposure to IL-4 and IL-13 is the lipid-oxidizing enzyme called15-lipoxygenase (15-LO) (11, 35, 44). 15-LO is expressed andenzymatically active in human atherosclerotic lesions (8).Through specific lipid oxidation, it generates a series of pro-and anti-inflammatory molecules, termed HPODEs/HODEs,which have been extracted from atherosclerotic lesions and arepotent mediators of inflammatory responses (4, 16, 22). Thisenzyme is believed to be important for the pathogenesis ofatherosclerosis as well as for generating potent inflammatorymediators.
Previously, our group showed the involvement of the Jak/Stat pathway in 15-LO induction in IL-13-treated humanmonocytes. Our studies demonstrated that activation of Jak2and Tyk2 kinases was required for IL-13-induced 15-LO pro-tein expression (44). Our recent studies have defined the func-tional IL-13 receptor complex, association of the Jaks with thereceptor constituents, and the tyrosine phosphorylation of spe-cific Stat molecules, Stat1, Stat3, Stat5, and Stat6, in responseto IL-13 (43). These studies established a novel and selectivesignal transduction pathway from the receptor to the nucleus inhuman monocytes.
Tyrosine phosphorylation of Stat proteins by specific Jakkinases facilitates the dimerization of Stat molecules by bind-ing the SH2 domain of one Stat molecule to the phosphotyro-sine of another Stat (50). The dimerized Stat complex is thentranslocated to the nucleus, binds DNA, and regulates theexpression of the corresponding target gene (63). In additionto tyrosine phosphorylation, serine phosphorylation of the Statmolecules is necessary for optimal transcriptional activity buthas no influence on either dimer formation or nuclear trans-location of the Stat complex (57, 58, 62). Recent reports sug-gest that the serine phosphorylation of Stat molecules [e.g.,Stat1� (Ser727), Stat3 (Ser727), Stat5A (Ser 725), and Stat5B(Ser730)] is mediated by different kinases (1–3, 9, 19, 20, 26, 28,29, 34, 46, 48, 56, 60).
A group of serine-threonine kinases, including ERK1 and -2,p38 mitogen-activated protein kinase (MAPK), and c-Jun N-terminal kinases (JNKs), all components of distinct but evolu-tionally conserved MAPK signaling cascade, and protein ki-nase C� have been reported to be involved in the serinephosphorylation of Stat1 and Stat3 (3, 9, 19, 20, 26, 28, 29, 34,46, 48, 56). The best-known and best-characterized candidatesof the serine-threonine kinases are the ERKs, characterized bythe p42 and p44 MAP kinases ERK1 and ERK2 (10). ERK1and -2 are activated in response to a wide variety of growthfactors and mitogens (7). JNKs are related to the ERKs but areregulated differently. Usually JNKs are activated in response tostress or cytokines (13, 31, 52). The p38 MAPK family is alsoactivated in response to osmotic stress, cytokines, or phorbolesters (21, 55, 61). Upon activation, the MAP kinases phos-
* Corresponding author. Mailing address: Department of Cell Biol-ogy, Lerner Research Institute, Cleveland Clinic Foundation, 9500Euclid Ave., Cleveland, OH 44195. Phone: (216) 444-5222. Fax: (216)444-9404. E-mail: [email protected].
3918
on October 24, 2014 by guest
http://mcb.asm
.org/D
ownloaded from

phorylate and activate transcription factors, including the Stats.The activation and tyrosine/threonine phosphorylation of p38MAPK are induced in response to several hematopoieticgrowth factors, including IL-3 and granulocyte-macrophagecolony-stimulating factor, as well as physical and chemicalstresses (17).
Previous studies suggested the involvement of p38 MAPK inStat1 serine phosphorylation and transcriptional activation in-duced by alpha interferon (IFN-�) and IFN-� (19). In addi-tion, p38 MAPK was reported to play an important role inregulating Stat1 and Stat3 serine phosphorylation in responseto the combination of IL-2 and IL-12 in T cells (20). Here, wereport for the first time that in primary human monocytes,IL-13 induces activation of p38 MAPK, which in turn regulatesthe serine 727 phosphorylation of both Stat1 and Stat3. Fur-thermore, our results demonstrate that activation of p38MAPK and subsequent phosphorylation of 727 serine residueson Stat1 and Stat3 molecules are critical in IL-13-induced15-LO expression in human monocytes.
MATERIALS AND METHODS
Reagents. Recombinant human IL-13 was purchased from Biosource Interna-tional (Camarillo, Calif.). Antibody against rabbit reticulocyte 15-LO, cross-reacting with human 15-LO, was raised in sheep and was kindly provided byJoseph Cornicelli, Parke-Davis, or purchased from Cayman Chemical (AnnArbor, Mich.). Antibodies specific for Stat1 phosphorylated on Ser-727 (P-Ser-Stat1) and for Stat3 phosphorylated on Ser-727 (P-Ser-Stat3) were generated inrabbits and characterized previously (18). Antiphosphotyrosine-Stat antibodiesraised against Stat1(Y701) and Stat3(Y705) were obtained from Upstate Bio-technology and were used to detect tyrosine-phosphorylated Stat proteins onWestern blots. Each of the Stat antibodies used was non-cross-reactive with otherStat molecules and was good for detecting proteins on Western blots. Antibodiesto phospho-p38 (active p38) and to total p38 MAPK were purchased from CellSignaling Technology (Beverly, Mass.). Actinomycin D was obtained from Sigma(St. Louis, Mo.). p38 MAPK inhibitors SB202190 and SB203580 and the inactiveanalogue SB202474 were purchased from Calbiochem (La Jolla, Calif.). H7 andits structural analogue HA1004 were purchased from Biomol (Plymouth Meet-ing, Pa.). All the pharmacological inhibitors were dissolved in dimethyl sulfoxideand stored at �20°C as 10 mM stock solutions.
Isolation of human monocytes. Human peripheral blood monocytes wereisolated from heparinized whole blood as described earlier (44). Adherent cellswere released from the flask with 5 mM EDTA and plated in six-well cell cultureplates (Nunclon, Roskilde, Denmark). The cell preparations usually had morethan 95% monocytes and were maintained in Dulbecco’s modified Eagle’s me-dium containing 10% bovine calf serum at 37°C in the presence of 10% CO2.
RNA extraction and real-time RT-PCR. Monocytes were plated in six-wellculture plates at 5 � 106 cells/well in 2 ml of medium. Two hours after plating,cells were treated with 500 pM recombinant IL-13 for different times, as indi-cated. In some other experiments, monocytes were either treated with inhibitorsfor 30 min and then with IL-13 for various times or transfected with Stat1 andStat3 decoys for 24 h before IL-13 was added. After each treatment, cells werecollected and washed with phosphate-buffered saline (PBS). Total cellular RNAwas extracted with the RNeasy minikit from Qiagen (Valencia, Calif.). Onemicrogram of total RNA from different treatments was reverse transcribed withthe TaqMan reverse transcription kit (PE Applied Biosystem, Foster City,Calif.). The cDNAs were then subjected to real-time reverse transcription (RT)-PCR with Sybr Green PCR core reagents. The PCRs were carried out in an ABIPrism 7700 sequence detector (PE Applied Biosystems), with the thermal cycleconditions suggested by the manufacturer. The PCR primers for 15-LO wereselected from the regions displaying minimal sequence homology to human 5-and 12-LO and spanned two introns. The sequences of the 15-LO primers were5�-GCTGGAAGGATCTAGATGACT-3� and 5�-TGGCTACAGAGAATGACGTTG-3�. The amplicon for 15-LO was 294 bp. Data were acquired and analyzedwith the sequence detector 7700 software.
Detection of Stat1 and Stat3 serine phosphorylation and p38 MAPK phos-phorylation. Monocytes were plated at 2 � 106/ml in six-well plates and treatedwith inhibitors (30 min) when required and then treated with IL-13 (500 pM) fordifferent times as indicated. All cells were treated with 100 �M sodium or-
thovanadate solution 15 min before lysis (43). The cells were then washed threetimes with PBS to remove traces of Dulbecco’s modified Eagle’s medium–10%bovine calf serum before lysis. Whole-cell extracts and nuclear extracts wereprepared according to previously published protocols (42, 47) with the exceptionthat 0.5 M NaCl was used in the whole-cell lysis buffer (43). After the proteinconcentration was determined with the Bio-Rad protein assay kit, lysates wereloaded on a sodium dodecyl sulfate–8% polyacrylamide gel electrophoresis(SDS-8% PAGE) gel (10 to 75 �g/well). The proteins were transferred to apolyvinylidene difluoride membrane, blocked with 5% nonfat milk in PBS with0.1% Tween 20, and the membranes were probed with anti-phosphoserine-Stat1(Ser 727), anti-phosphoserine-Stat3 (Ser 727), or anti-phospho-p38 MAPK an-tibody overnight (diluted 1:1,000 in 3% bovine serum albumin in PBS with 0.1%Tween 20). A horseradish peroxidase-labeled secondary antibody (diluted1:1,000 in 3% bovine serum albumin in PBS with 0.1% Tween 20) was thenadded for 1 h, and the hybridization signal was detected with an ECL kit (Pierce).In several experiments, immunoblots were stripped and reprobed to assess equalloading according to the previously published protocol (43).
15-LO protein induction and detection. IL-13 (500 pM) was added to six-wellplates containing 5 � 106 cells/well treated with inhibitors when required andthen washed three times with PBS to remove the traces of Dulbecco’s modifiedEagle’s medium–10% bovine calf serum. The cells were lysed, and the 15-LOprotein was detected on Western blots following the previously described pro-tocol (44). Quantification of Western blot results was conducted with the soft-ware program NIH Image.
Nuclear protein extraction and electrophoretic mobility shift assay. To assessthe role of p38 MAPK in IL-13-induced Stat1 and Stat3 DNA binding activity, anelectrophoretic mobility shift assay was performed with nuclear extracts andspecific Stat1 and Stat3 probes (Santa Cruz Biotechnology Inc.). Monocytes weretreated with 5 �M SB202190 for 30 min, followed by the addition of IL-13 foranother 30 min. Nuclear proteins were extracted, and the electrophoretic mo-bility shift assay was performed following the protocol described earlier (43).
Transfection of Stat1 and Stat3 decoy and mismatched oligodeoxyribonucle-otides into monocytes. The phosphorothioated oligodeoxyribonucleotides usedfor Stat1 and Stat3 decoys were purchased from Sigma-Genosys Biotechnologies,Inc. (The Woodlands, Tex.). These decoys were used in previous studies andshown to provide specific inhibition of Stat1 and Stat3 activities (24). The se-quences for the decoys were 5�-ATATTCCTGTAAGTG-3� and 3�-TATAAGGACATTCAC-5� for Stat1; 5�-ATATTGGAGTAAGTG-3� and 3�-TATAACCTCATTCAC-5� for the mismatched Stat1 decoy; 5�-GATCCTTCTGGGAATTCCTAGATC-3� and 3�CTAGGAAGACCCTTAAGGATCTAG-5� for the Stat3 decoy;and 5�-GATCCTTCTGGGCCGTCCTAGATC-3� and 3�-CTAGGAAGACCCGGCAGGATCTAG-5� for the mismatched Stat3 decoy. The consensus sequencesare in italics and correspond to the high-affinity Ly-6E gamma interferon-acti-vated site (GAS) (27) and the acute-phase response element in the rat �2-macroglobulin gene (6), respectively.
The single-stranded oligodeoxyribonucleotides were annealed by incubation at65°C for 10 min and then slowly being cooled to room temperature for 2 h.Double-stranded decoys were transfected into monocytes at a final concentrationof 2 �M by using Superfect (Qiagen) following the manufacturer’s instruction.After 24 h of transfection, cells were treated with IL-13 for either 4 h for thereal-time RT-PCR analysis or for 24 h for Western blot analysis to assess 15-LOmRNA or protein expression.
RESULTS
Induction of 15-LO expression in IL-13-treated monocytes.Earlier reports indicated that IL-4 and IL-13 can uniquelyinduce 15-LO expression and activity in human peripheralblood monocytes following 36 to 72 h of treatment (11, 35, 44).With a more quantitative approach, real-time RT-PCR, weperformed a time-dependent analysis of IL-13 induction of15-LO mRNA expression to examine this induction in greaterdetail. The results of representative experiments are illustratedin Fig. 1. 15-LO mRNA levels were detected in untreatedmonocytes and increased continuously with time of exposureto IL-13 up to 24 h (Fig. 1A and C). The induction of 15-LOmRNA was detected as early as 1 h of IL-13 treatment. Theinduction of the mRNA level was profound, with a remarkable4,000-fold induction (212) by 24 h. The mRNA levels of
VOL. 23, 2003 IL-13 INDUCTION OF 15-LO GENE EXPRESSION 3919
on October 24, 2014 by guest
http://mcb.asm
.org/D
ownloaded from
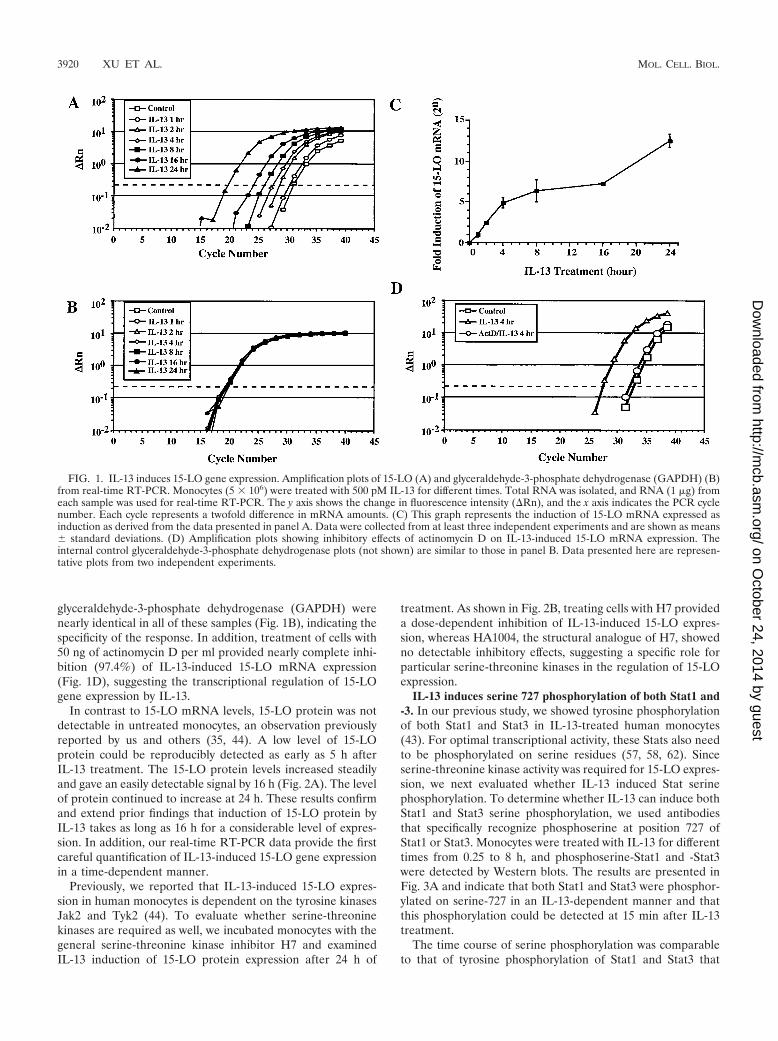
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) werenearly identical in all of these samples (Fig. 1B), indicating thespecificity of the response. In addition, treatment of cells with50 ng of actinomycin D per ml provided nearly complete inhi-bition (97.4%) of IL-13-induced 15-LO mRNA expression(Fig. 1D), suggesting the transcriptional regulation of 15-LOgene expression by IL-13.
In contrast to 15-LO mRNA levels, 15-LO protein was notdetectable in untreated monocytes, an observation previouslyreported by us and others (35, 44). A low level of 15-LOprotein could be reproducibly detected as early as 5 h afterIL-13 treatment. The 15-LO protein levels increased steadilyand gave an easily detectable signal by 16 h (Fig. 2A). The levelof protein continued to increase at 24 h. These results confirmand extend prior findings that induction of 15-LO protein byIL-13 takes as long as 16 h for a considerable level of expres-sion. In addition, our real-time RT-PCR data provide the firstcareful quantification of IL-13-induced 15-LO gene expressionin a time-dependent manner.
Previously, we reported that IL-13-induced 15-LO expres-sion in human monocytes is dependent on the tyrosine kinasesJak2 and Tyk2 (44). To evaluate whether serine-threoninekinases are required as well, we incubated monocytes with thegeneral serine-threonine kinase inhibitor H7 and examinedIL-13 induction of 15-LO protein expression after 24 h of
treatment. As shown in Fig. 2B, treating cells with H7 provideda dose-dependent inhibition of IL-13-induced 15-LO expres-sion, whereas HA1004, the structural analogue of H7, showedno detectable inhibitory effects, suggesting a specific role forparticular serine-threonine kinases in the regulation of 15-LOexpression.
IL-13 induces serine 727 phosphorylation of both Stat1 and-3. In our previous study, we showed tyrosine phosphorylationof both Stat1 and Stat3 in IL-13-treated human monocytes(43). For optimal transcriptional activity, these Stats also needto be phosphorylated on serine residues (57, 58, 62). Sinceserine-threonine kinase activity was required for 15-LO expres-sion, we next evaluated whether IL-13 induced Stat serinephosphorylation. To determine whether IL-13 can induce bothStat1 and Stat3 serine phosphorylation, we used antibodiesthat specifically recognize phosphoserine at position 727 ofStat1 or Stat3. Monocytes were treated with IL-13 for differenttimes from 0.25 to 8 h, and phosphoserine-Stat1 and -Stat3were detected by Western blots. The results are presented inFig. 3A and indicate that both Stat1 and Stat3 were phosphor-ylated on serine-727 in an IL-13-dependent manner and thatthis phosphorylation could be detected at 15 min after IL-13treatment.
The time course of serine phosphorylation was comparableto that of tyrosine phosphorylation of Stat1 and Stat3 that
FIG. 1. IL-13 induces 15-LO gene expression. Amplification plots of 15-LO (A) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (B)from real-time RT-PCR. Monocytes (5 � 106) were treated with 500 pM IL-13 for different times. Total RNA was isolated, and RNA (1 �g) fromeach sample was used for real-time RT-PCR. The y axis shows the change in fluorescence intensity (Rn), and the x axis indicates the PCR cyclenumber. Each cycle represents a twofold difference in mRNA amounts. (C) This graph represents the induction of 15-LO mRNA expressed asinduction as derived from the data presented in panel A. Data were collected from at least three independent experiments and are shown as means� standard deviations. (D) Amplification plots showing inhibitory effects of actinomycin D on IL-13-induced 15-LO mRNA expression. Theinternal control glyceraldehyde-3-phosphate dehydrogenase plots (not shown) are similar to those in panel B. Data presented here are represen-tative plots from two independent experiments.
3920 XU ET AL. MOL. CELL. BIOL.
on October 24, 2014 by guest
http://mcb.asm
.org/D
ownloaded from
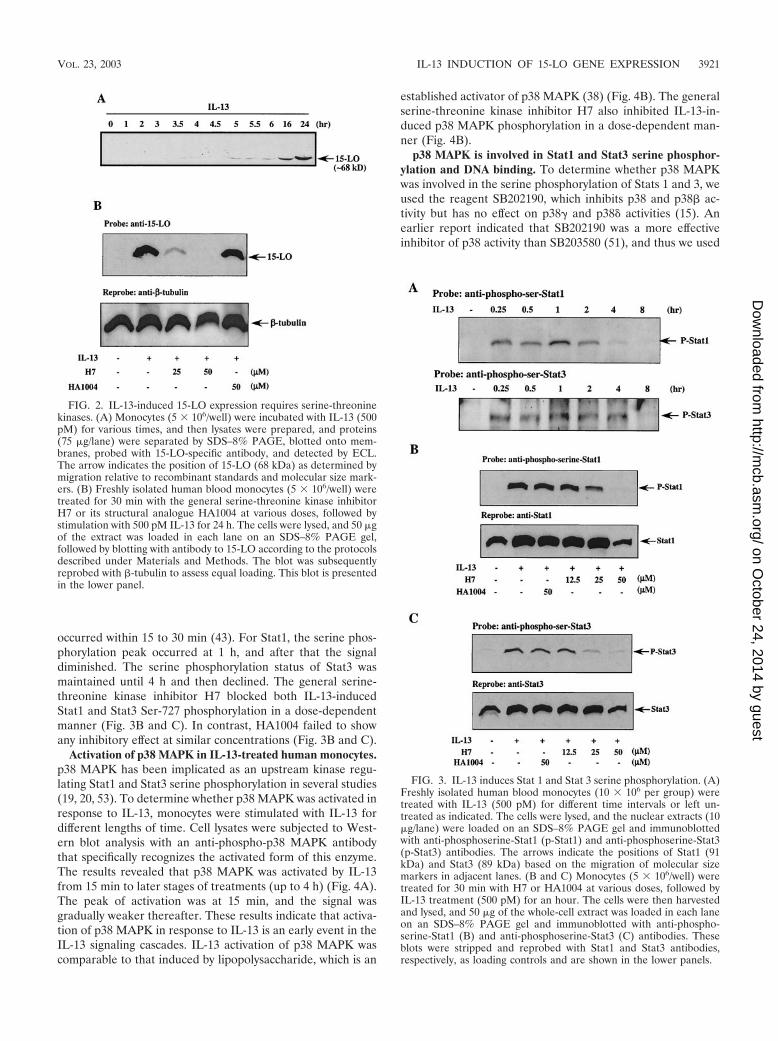
occurred within 15 to 30 min (43). For Stat1, the serine phos-phorylation peak occurred at 1 h, and after that the signaldiminished. The serine phosphorylation status of Stat3 wasmaintained until 4 h and then declined. The general serine-threonine kinase inhibitor H7 blocked both IL-13-inducedStat1 and Stat3 Ser-727 phosphorylation in a dose-dependentmanner (Fig. 3B and C). In contrast, HA1004 failed to showany inhibitory effect at similar concentrations (Fig. 3B and C).
Activation of p38 MAPK in IL-13-treated human monocytes.p38 MAPK has been implicated as an upstream kinase regu-lating Stat1 and Stat3 serine phosphorylation in several studies(19, 20, 53). To determine whether p38 MAPK was activated inresponse to IL-13, monocytes were stimulated with IL-13 fordifferent lengths of time. Cell lysates were subjected to West-ern blot analysis with an anti-phospho-p38 MAPK antibodythat specifically recognizes the activated form of this enzyme.The results revealed that p38 MAPK was activated by IL-13from 15 min to later stages of treatments (up to 4 h) (Fig. 4A).The peak of activation was at 15 min, and the signal wasgradually weaker thereafter. These results indicate that activa-tion of p38 MAPK in response to IL-13 is an early event in theIL-13 signaling cascades. IL-13 activation of p38 MAPK wascomparable to that induced by lipopolysaccharide, which is an
established activator of p38 MAPK (38) (Fig. 4B). The generalserine-threonine kinase inhibitor H7 also inhibited IL-13-in-duced p38 MAPK phosphorylation in a dose-dependent man-ner (Fig. 4B).
p38 MAPK is involved in Stat1 and Stat3 serine phosphor-ylation and DNA binding. To determine whether p38 MAPKwas involved in the serine phosphorylation of Stats 1 and 3, weused the reagent SB202190, which inhibits p38 and p38� ac-tivity but has no effect on p38� and p38� activities (15). Anearlier report indicated that SB202190 was a more effectiveinhibitor of p38 activity than SB203580 (51), and thus we used
FIG. 2. IL-13-induced 15-LO expression requires serine-threoninekinases. (A) Monocytes (5 � 106/well) were incubated with IL-13 (500pM) for various times, and then lysates were prepared, and proteins(75 �g/lane) were separated by SDS–8% PAGE, blotted onto mem-branes, probed with 15-LO-specific antibody, and detected by ECL.The arrow indicates the position of 15-LO (68 kDa) as determined bymigration relative to recombinant standards and molecular size mark-ers. (B) Freshly isolated human blood monocytes (5 � 106/well) weretreated for 30 min with the general serine-threonine kinase inhibitorH7 or its structural analogue HA1004 at various doses, followed bystimulation with 500 pM IL-13 for 24 h. The cells were lysed, and 50 �gof the extract was loaded in each lane on an SDS–8% PAGE gel,followed by blotting with antibody to 15-LO according to the protocolsdescribed under Materials and Methods. The blot was subsequentlyreprobed with �-tubulin to assess equal loading. This blot is presentedin the lower panel.
FIG. 3. IL-13 induces Stat 1 and Stat 3 serine phosphorylation. (A)Freshly isolated human blood monocytes (10 � 106 per group) weretreated with IL-13 (500 pM) for different time intervals or left un-treated as indicated. The cells were lysed, and the nuclear extracts (10�g/lane) were loaded on an SDS–8% PAGE gel and immunoblottedwith anti-phosphoserine-Stat1 (p-Stat1) and anti-phosphoserine-Stat3(p-Stat3) antibodies. The arrows indicate the positions of Stat1 (91kDa) and Stat3 (89 kDa) based on the migration of molecular sizemarkers in adjacent lanes. (B and C) Monocytes (5 � 106/well) weretreated for 30 min with H7 or HA1004 at various doses, followed byIL-13 treatment (500 pM) for an hour. The cells were then harvestedand lysed, and 50 �g of the whole-cell extract was loaded in each laneon an SDS–8% PAGE gel and immunoblotted with anti-phospho-serine-Stat1 (B) and anti-phosphoserine-Stat3 (C) antibodies. Theseblots were stripped and reprobed with Stat1 and Stat3 antibodies,respectively, as loading controls and are shown in the lower panels.
VOL. 23, 2003 IL-13 INDUCTION OF 15-LO GENE EXPRESSION 3921
on October 24, 2014 by guest
http://mcb.asm
.org/D
ownloaded from

SB202190. The results of a representative experiment areshown in Fig. 5. Monocytes were treated with SB202190 for 30min prior to the addition of IL-13, either treated or not withIL-13 for an hour, and harvested. The serine phosphorylationstatus of both Stat1 and Stat3 was determined by Western blotswith phosphoserine-Stat-specific antibodies. The results, pre-sented in Fig. 5A, indicate that SB202190 inhibited Stat1 serinephosphorylation in a dose-dependent manner. The degree ofinhibition was nearly complete ( 85%) at a 5 �M concentra-tion of SB202190.
We conducted similar experiments in which we studied the
effect of SB202190 on Stat3 serine phosphorylation. The cellswere treated with SB202190 (30 min before IL-13 addition),then similarly treated with IL-13 for an hour, and harvested.Subsequently, phosphoserine-Stat3 was detected on Westernblots, as shown on Fig. 5B. The results demonstrate thatSB202190 (5 �M) had an effect on Stat3 serine phosphoryla-tion ( 90% inhibition) similar to that exhibited in the case ofStat1 serine phosphorylation in Fig. 5A.
To evaluate whether SB202190 had any effects on Stat1 andStat3 tyrosine phosphorylation, the same cell lysates were usedfor Western blot analysis. The phosphorylation status of Stat
FIG. 4. IL-13 induces phosphorylation of p38 MAPK. (A) Monocytes were left untreated or treated with IL-13 (500 pM) for different timesas indicated. The cell lysates (75 �g/lane) were separated by SDS-PAGE and immunoblotted with anti-phospho-p38 antibody. The blot wassubsequently stripped and reprobed with anti-p38 antibody to assess equal loading of protein in each lane. The arrow indicates the position ofphospho-p38 MAPK and total p38 MAPK. The results are representative of at least three similar experiments. The ratio of activated p38 to totalp38 MAPK from one of the representative data, shown in arbitrary units, was obtained with NIH Image software and is presented in the lowerpanel. (B) Monocytes (5 � 106/well) were treated for 30 min with H7 or lipopolysaccharide at various doses, followed by IL-13 stimulation (500pM) for 15 min. Whole-cell lysates (50 �g/lane) were separated by SDS–8% PAGE and immunoblotted with anti-phospho-p38 MAPK antibody.The blot was stripped and reprobed with anti-p38 MAPK antibody as a loading control and is shown in the lower panel.
3922 XU ET AL. MOL. CELL. BIOL.
on October 24, 2014 by guest
http://mcb.asm
.org/D
ownloaded from
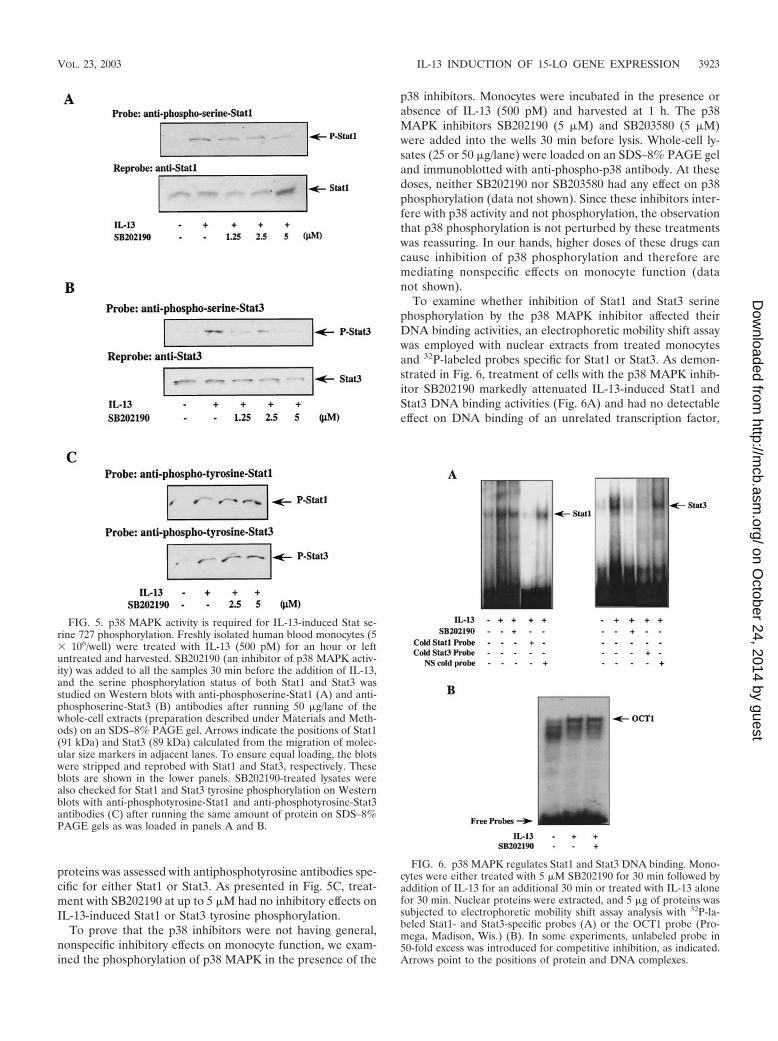
proteins was assessed with antiphosphotyrosine antibodies spe-cific for either Stat1 or Stat3. As presented in Fig. 5C, treat-ment with SB202190 at up to 5 �M had no inhibitory effects onIL-13-induced Stat1 or Stat3 tyrosine phosphorylation.
To prove that the p38 inhibitors were not having general,nonspecific inhibitory effects on monocyte function, we exam-ined the phosphorylation of p38 MAPK in the presence of the
p38 inhibitors. Monocytes were incubated in the presence orabsence of IL-13 (500 pM) and harvested at 1 h. The p38MAPK inhibitors SB202190 (5 �M) and SB203580 (5 �M)were added into the wells 30 min before lysis. Whole-cell ly-sates (25 or 50 �g/lane) were loaded on an SDS–8% PAGE geland immunoblotted with anti-phospho-p38 antibody. At thesedoses, neither SB202190 nor SB203580 had any effect on p38phosphorylation (data not shown). Since these inhibitors inter-fere with p38 activity and not phosphorylation, the observationthat p38 phosphorylation is not perturbed by these treatmentswas reassuring. In our hands, higher doses of these drugs cancause inhibition of p38 phosphorylation and therefore aremediating nonspecific effects on monocyte function (datanot shown).
To examine whether inhibition of Stat1 and Stat3 serinephosphorylation by the p38 MAPK inhibitor affected theirDNA binding activities, an electrophoretic mobility shift assaywas employed with nuclear extracts from treated monocytesand 32P-labeled probes specific for Stat1 or Stat3. As demon-strated in Fig. 6, treatment of cells with the p38 MAPK inhib-itor SB202190 markedly attenuated IL-13-induced Stat1 andStat3 DNA binding activities (Fig. 6A) and had no detectableeffect on DNA binding of an unrelated transcription factor,
FIG. 5. p38 MAPK activity is required for IL-13-induced Stat se-rine 727 phosphorylation. Freshly isolated human blood monocytes (5� 106/well) were treated with IL-13 (500 pM) for an hour or leftuntreated and harvested. SB202190 (an inhibitor of p38 MAPK activ-ity) was added to all the samples 30 min before the addition of IL-13,and the serine phosphorylation status of both Stat1 and Stat3 wasstudied on Western blots with anti-phosphoserine-Stat1 (A) and anti-phosphoserine-Stat3 (B) antibodies after running 50 �g/lane of thewhole-cell extracts (preparation described under Materials and Meth-ods) on an SDS–8% PAGE gel. Arrows indicate the positions of Stat1(91 kDa) and Stat3 (89 kDa) calculated from the migration of molec-ular size markers in adjacent lanes. To ensure equal loading, the blotswere stripped and reprobed with Stat1 and Stat3, respectively. Theseblots are shown in the lower panels. SB202190-treated lysates werealso checked for Stat1 and Stat3 tyrosine phosphorylation on Westernblots with anti-phosphotyrosine-Stat1 and anti-phosphotyrosine-Stat3antibodies (C) after running the same amount of protein on SDS–8%PAGE gels as was loaded in panels A and B.
FIG. 6. p38 MAPK regulates Stat1 and Stat3 DNA binding. Mono-cytes were either treated with 5 �M SB202190 for 30 min followed byaddition of IL-13 for an additional 30 min or treated with IL-13 alonefor 30 min. Nuclear proteins were extracted, and 5 �g of proteins wassubjected to electrophoretic mobility shift assay analysis with 32P-la-beled Stat1- and Stat3-specific probes (A) or the OCT1 probe (Pro-mega, Madison, Wis.) (B). In some experiments, unlabeled probe in50-fold excess was introduced for competitive inhibition, as indicated.Arrows point to the positions of protein and DNA complexes.
VOL. 23, 2003 IL-13 INDUCTION OF 15-LO GENE EXPRESSION 3923
on October 24, 2014 by guest
http://mcb.asm
.org/D
ownloaded from
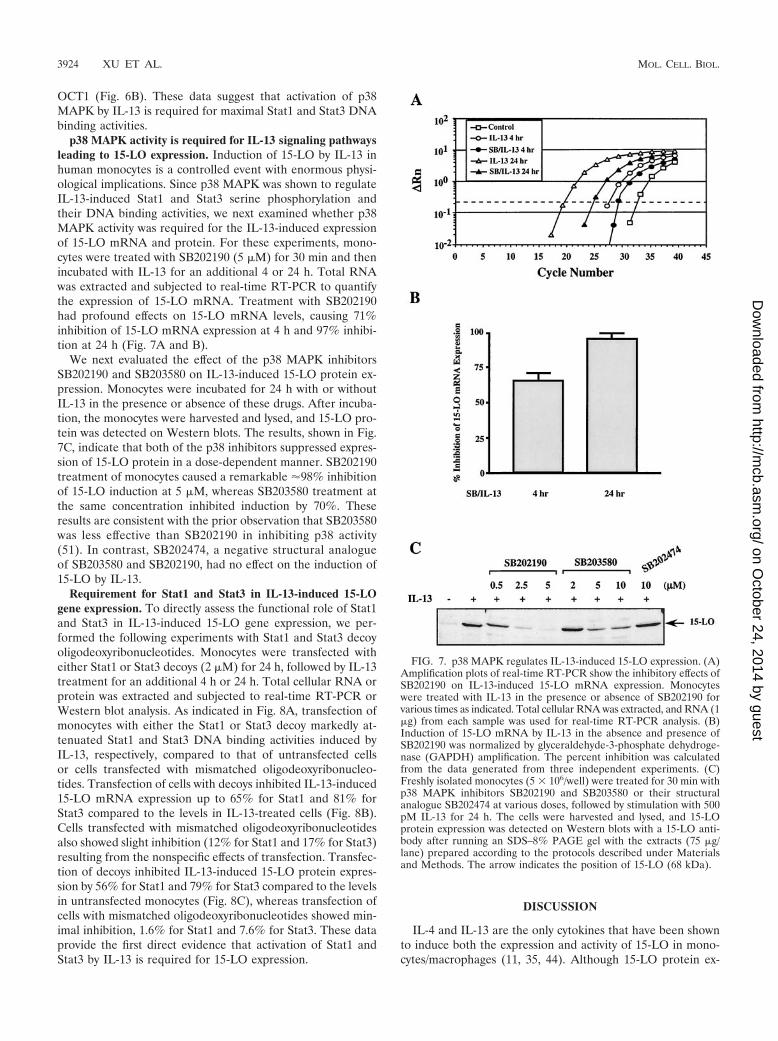
OCT1 (Fig. 6B). These data suggest that activation of p38MAPK by IL-13 is required for maximal Stat1 and Stat3 DNAbinding activities.
p38 MAPK activity is required for IL-13 signaling pathwaysleading to 15-LO expression. Induction of 15-LO by IL-13 inhuman monocytes is a controlled event with enormous physi-ological implications. Since p38 MAPK was shown to regulateIL-13-induced Stat1 and Stat3 serine phosphorylation andtheir DNA binding activities, we next examined whether p38MAPK activity was required for the IL-13-induced expressionof 15-LO mRNA and protein. For these experiments, mono-cytes were treated with SB202190 (5 �M) for 30 min and thenincubated with IL-13 for an additional 4 or 24 h. Total RNAwas extracted and subjected to real-time RT-PCR to quantifythe expression of 15-LO mRNA. Treatment with SB202190had profound effects on 15-LO mRNA levels, causing 71%inhibition of 15-LO mRNA expression at 4 h and 97% inhibi-tion at 24 h (Fig. 7A and B).
We next evaluated the effect of the p38 MAPK inhibitorsSB202190 and SB203580 on IL-13-induced 15-LO protein ex-pression. Monocytes were incubated for 24 h with or withoutIL-13 in the presence or absence of these drugs. After incuba-tion, the monocytes were harvested and lysed, and 15-LO pro-tein was detected on Western blots. The results, shown in Fig.7C, indicate that both of the p38 inhibitors suppressed expres-sion of 15-LO protein in a dose-dependent manner. SB202190treatment of monocytes caused a remarkable 98% inhibitionof 15-LO induction at 5 �M, whereas SB203580 treatment atthe same concentration inhibited induction by 70%. Theseresults are consistent with the prior observation that SB203580was less effective than SB202190 in inhibiting p38 activity(51). In contrast, SB202474, a negative structural analogueof SB203580 and SB202190, had no effect on the induction of15-LO by IL-13.
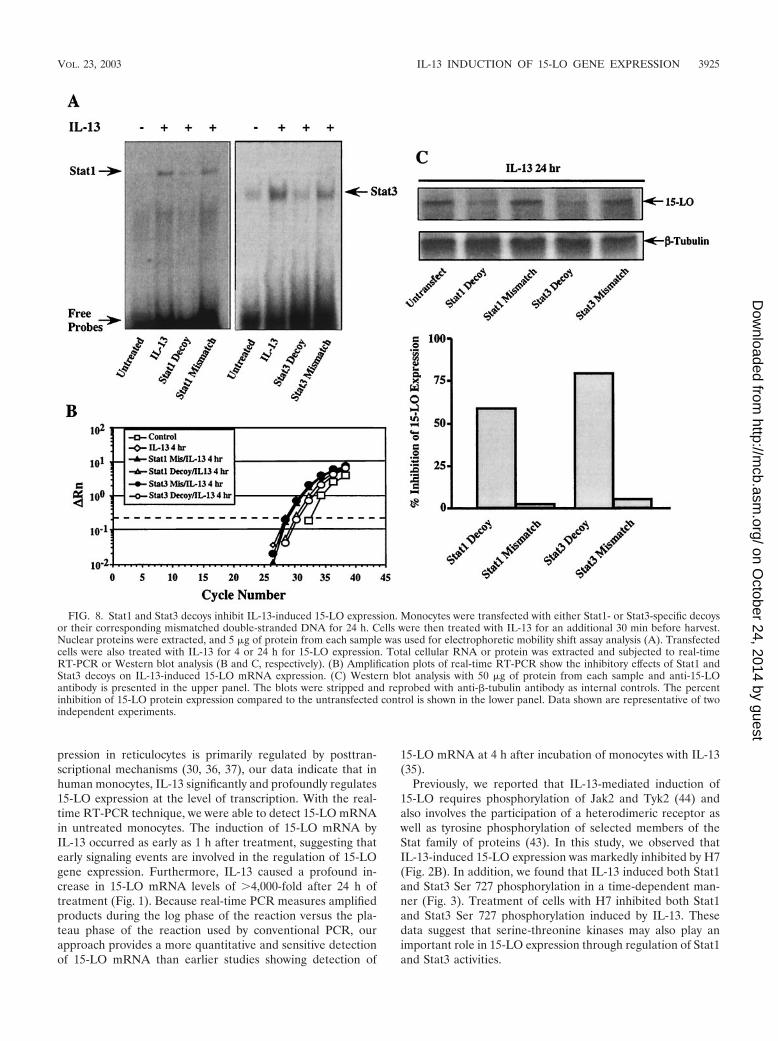
Requirement for Stat1 and Stat3 in IL-13-induced 15-LOgene expression. To directly assess the functional role of Stat1and Stat3 in IL-13-induced 15-LO gene expression, we per-formed the following experiments with Stat1 and Stat3 decoyoligodeoxyribonucleotides. Monocytes were transfected witheither Stat1 or Stat3 decoys (2 �M) for 24 h, followed by IL-13treatment for an additional 4 h or 24 h. Total cellular RNA orprotein was extracted and subjected to real-time RT-PCR orWestern blot analysis. As indicated in Fig. 8A, transfection ofmonocytes with either the Stat1 or Stat3 decoy markedly at-tenuated Stat1 and Stat3 DNA binding activities induced byIL-13, respectively, compared to that of untransfected cellsor cells transfected with mismatched oligodeoxyribonucleo-tides. Transfection of cells with decoys inhibited IL-13-induced15-LO mRNA expression up to 65% for Stat1 and 81% forStat3 compared to the levels in IL-13-treated cells (Fig. 8B).Cells transfected with mismatched oligodeoxyribonucleotidesalso showed slight inhibition (12% for Stat1 and 17% for Stat3)resulting from the nonspecific effects of transfection. Transfec-tion of decoys inhibited IL-13-induced 15-LO protein expres-sion by 56% for Stat1 and 79% for Stat3 compared to the levelsin untransfected monocytes (Fig. 8C), whereas transfection ofcells with mismatched oligodeoxyribonucleotides showed min-imal inhibition, 1.6% for Stat1 and 7.6% for Stat3. These dataprovide the first direct evidence that activation of Stat1 andStat3 by IL-13 is required for 15-LO expression.
DISCUSSION
IL-4 and IL-13 are the only cytokines that have been shownto induce both the expression and activity of 15-LO in mono-cytes/macrophages (11, 35, 44). Although 15-LO protein ex-
FIG. 7. p38 MAPK regulates IL-13-induced 15-LO expression. (A)Amplification plots of real-time RT-PCR show the inhibitory effects ofSB202190 on IL-13-induced 15-LO mRNA expression. Monocyteswere treated with IL-13 in the presence or absence of SB202190 forvarious times as indicated. Total cellular RNA was extracted, and RNA (1�g) from each sample was used for real-time RT-PCR analysis. (B)Induction of 15-LO mRNA by IL-13 in the absence and presence ofSB202190 was normalized by glyceraldehyde-3-phosphate dehydroge-nase (GAPDH) amplification. The percent inhibition was calculatedfrom the data generated from three independent experiments. (C)Freshly isolated monocytes (5 � 106/well) were treated for 30 min withp38 MAPK inhibitors SB202190 and SB203580 or their structuralanalogue SB202474 at various doses, followed by stimulation with 500pM IL-13 for 24 h. The cells were harvested and lysed, and 15-LOprotein expression was detected on Western blots with a 15-LO anti-body after running an SDS–8% PAGE gel with the extracts (75 �g/lane) prepared according to the protocols described under Materialsand Methods. The arrow indicates the position of 15-LO (68 kDa).
3924 XU ET AL. MOL. CELL. BIOL.
on October 24, 2014 by guest
http://mcb.asm
.org/D
ownloaded from
pression in reticulocytes is primarily regulated by posttran-scriptional mechanisms (30, 36, 37), our data indicate that inhuman monocytes, IL-13 significantly and profoundly regulates15-LO expression at the level of transcription. With the real-time RT-PCR technique, we were able to detect 15-LO mRNAin untreated monocytes. The induction of 15-LO mRNA byIL-13 occurred as early as 1 h after treatment, suggesting thatearly signaling events are involved in the regulation of 15-LOgene expression. Furthermore, IL-13 caused a profound in-crease in 15-LO mRNA levels of 4,000-fold after 24 h oftreatment (Fig. 1). Because real-time PCR measures amplifiedproducts during the log phase of the reaction versus the pla-teau phase of the reaction used by conventional PCR, ourapproach provides a more quantitative and sensitive detectionof 15-LO mRNA than earlier studies showing detection of
15-LO mRNA at 4 h after incubation of monocytes with IL-13(35).
Previously, we reported that IL-13-mediated induction of15-LO requires phosphorylation of Jak2 and Tyk2 (44) andalso involves the participation of a heterodimeric receptor aswell as tyrosine phosphorylation of selected members of theStat family of proteins (43). In this study, we observed thatIL-13-induced 15-LO expression was markedly inhibited by H7(Fig. 2B). In addition, we found that IL-13 induced both Stat1and Stat3 Ser 727 phosphorylation in a time-dependent man-ner (Fig. 3). Treatment of cells with H7 inhibited both Stat1and Stat3 Ser 727 phosphorylation induced by IL-13. Thesedata suggest that serine-threonine kinases may also play animportant role in 15-LO expression through regulation of Stat1and Stat3 activities.
FIG. 8. Stat1 and Stat3 decoys inhibit IL-13-induced 15-LO expression. Monocytes were transfected with either Stat1- or Stat3-specific decoysor their corresponding mismatched double-stranded DNA for 24 h. Cells were then treated with IL-13 for an additional 30 min before harvest.Nuclear proteins were extracted, and 5 �g of protein from each sample was used for electrophoretic mobility shift assay analysis (A). Transfectedcells were also treated with IL-13 for 4 or 24 h for 15-LO expression. Total cellular RNA or protein was extracted and subjected to real-timeRT-PCR or Western blot analysis (B and C, respectively). (B) Amplification plots of real-time RT-PCR show the inhibitory effects of Stat1 andStat3 decoys on IL-13-induced 15-LO mRNA expression. (C) Western blot analysis with 50 �g of protein from each sample and anti-15-LOantibody is presented in the upper panel. The blots were stripped and reprobed with anti-�-tubulin antibody as internal controls. The percentinhibition of 15-LO protein expression compared to the untransfected control is shown in the lower panel. Data shown are representative of twoindependent experiments.
VOL. 23, 2003 IL-13 INDUCTION OF 15-LO GENE EXPRESSION 3925
on October 24, 2014 by guest
http://mcb.asm
.org/D
ownloaded from

While the Jak kinases are involved in tyrosine phosphoryla-tion of Stat proteins, a variety of serine-threonine kinases,mostly the components of the MAPK signaling cascade, havebeen implicated in Ser phosphorylation of Stats. In humanmonocytes, we showed that p38 MAPK was phosphorylated/activated by IL-13 as early as 15 min of treatment and that H7inhibited p38 MAPK activation (Fig. 4), indicating that p38MAPK may be one of the key components involved in the earlysignaling events induced by IL-13. Although recent studiesreported that IL-4 induces p38 MAPK phosphorylation/activa-tion in dermal fibroblasts (25), our results provide the firstevidence demonstrating that IL-13 can induce p38 MAPK ac-tivation in human monocytes.
Activation of p38 MAPK by IL-13 was involved in the phos-phorylation of Stat1 and Stat3 on serine 727. Previous studieshave indicated that p38 MAPK may be involved in Stat serinephosphorylation and transcriptional activation in different cellsystems. In Ewing’s sarcoma cells, IFN-�-induced Stat1 Ser727 phosphorylation was observed with concomitant activationof p38 MAPK (46). In T cells, Gollob et al. (20) showed thatthe functional synergy between IL-12 and IL-2 was dependentupon the activation of p38 MAPK, which correlated with Statserine phosphorylation. They proposed that this Ser 727 Statphosphorylation induced by IL-12 and IL-2 might be mediatedby p38 MAPK. Earlier, Goh et al. (19) suggested that p38MAPK may regulate Stat1 serine phosphorylation and tran-scriptional activity in response to IFN-� and IFN-� in HeLa S3cells. While our results are in good agreement with the worksby Goh et al. (19) and Gollob et al. (20), it is worth noting thatin other systems, p38 MAPK regulates IFN-�-dependent genetranscription without affecting Stat1 and Stat3 serine phos-phorylation, whereas protein kinase C� mediates Ser 727 phos-phorylation on Stat1 (54, 56). These discrepant findings likelyresult from differences in cell-specific signaling pathways usedby various stimuli. More studies are needed to sort out thevarious signal transduction cascades mediated by different cy-tokines.
In addition to its effects on Stat1 and Stat3 serine phosphor-ylation, treatment with SB202190 also markedly reduced Stat1and Stat3 DNA binding activities (Fig. 6). These findings arenot likely due to nonspecific effects of the inhibitor becauseWestern blot analysis with the same cell lysates showed thattreatment of cells with SB202190 did not alter IL-13-inducedStat1 and Stat3 tyrosine phosphorylation (Fig. 5C). Previousstudies with various cell lines indicated that while Ser 727phosphorylation of Stat1 and Stat3 is important for maximaltranscriptional activation, this modification appeared to haveminimal effects on their DNA binding activities (13, 20). Theresults from our study and others (5), however, suggest that inprimary human monocytes, p38 MAPK activity is required forboth Stat1 and Stat3 Ser 727 phosphorylation and their DNAbinding activities. These findings may reflect the difference insignaling pathways between primary cells and cell lines. In celllines, there may be multiple signaling pathways mediating Statserine phosphorylation, whereas primary monocytes may onlyuse selected pathways for their specifically committed biolog-ical functions. Multiple signaling pathways in cell lines may beinvolved in serine phosphorylation in addition to Ser 727 ofStat proteins or other targets that may facilitate Stat DNAbinding activity. Therefore, inhibition of Stat Ser 727 phos-
phorylation or introduction of point-mutants of Stat Ser 727into cell lines may not affect Stat DNA binding activities ifother signaling cascades are able to compensate for the defect.The precise mechanisms of how Ser 727 phosphorylation af-fects Stat1 and Stat3 DNA binding in monocytes remains to beclarified.
The composition of the IL-13 receptor complex variesamong various cell types. Differential signaling pathways in-duced in response to IL-13 and IL-4 have been reported in celllines of various origins with varying expression of receptorcomponents and related signaling components. It appears thatwhen IL-2 receptor �C is part of the receptor complex andbecomes phosphorylated upon exposure to IL-4, Jak1 and Jak3are tyrosine phosphorylated in response to IL-4 (32, 45). Incontrast, Jak2 and Tyk2 have been shown to be phosphorylatedin cells where IL-2 receptor �C is not phosphorylated uponIL-13 exposure (for example, in monocytes) (43, 44). Earlierreports indicated Jak2 phosphorylation by IL-4 and IL-13 incells not expressing �C or �C that is not associated with IL-4receptor � in forming the receptor complex (33, 39). A signif-icant role of Jak2 leading to activation of p38 MAPK (64) andthe requirement for Jak2, Ras, and Raf for activation of ERK/MAPK (59) have also been described. Thus, the possibility ofinvolvement of MAPK in the IL-13 signaling pathway seemedreasonable in human monocytes. In this study, we furtherproved that activation of p38 MAPK is required for IL-13-induced 15-LO expression. With pharmacological inhibitorsspecific to p38 MAPK, we found that the induction of 15-LOmRNA and protein by IL-13 was substantially blocked, withcompatible degrees of inhibition (Fig. 7). Although we cannotrule out some contribution of translational regulation, our datastrongly suggest that IL-13-induced 15-LO expression is pri-marily regulated at the transcriptional level in human mono-cytes.
Recently, two reports suggested that p38 MAPK is requiredfor transcriptional responses of Stat target genes indepen-dently of Stat serine phosphorylation (40, 41). These findingsmay raise an alternative interpretation of our current results;however, there are fundamental differences between our studyand theirs. First, p38 MAPK is not activated by IFN-� (41) orby IL-4 alone (40) in the cell lines used in these reports,whereas p38 MAPK is immediately activated upon IL-13 stim-ulation in human monocytes (Fig. 4). Second, IFN-�-inducedStat1 Ser 727 phosphorylation is mediated through kinasesother than p38 MAPK (41) and Stat6 is not a direct substratefor p38 MAPK (40). In human monocytes, however, p38MAPK is required for IL-13-induced Stat1/Stat3 Ser 727 phos-phorylation and their DNA binding activities. These data againreflect the unique signaling pathways used by IL-13 in primaryhuman monocytes.
Transcription factor decoy technology has recently been de-veloped to inhibit the expression of several important geneswith synthetic double-stranded oligodeoxyribonucleotidescontaining the consensus binding sequence of a transcrip-tion factor. Previous studies reported that Stat1 and Stat3 de-coy treatment significantly inhibited the advanced glycationend product-induced DNA binding activities of Stat1 and Stat3and cellular mitogenesis in NRK-49F cells (24). The results ofour decoy experiments indicated that both Stat1 and Stat3 aremediated in IL-13-induced 15-LO expression (Fig. 8). Al-
3926 XU ET AL. MOL. CELL. BIOL.
on October 24, 2014 by guest
http://mcb.asm
.org/D
ownloaded from

though the majority of attention has focused on the role ofStat6 in 15-LO expression (12, 23, 30, 49), our data providenovel evidence that Stat1 and Stat3 are also crucial players inthe regulation of 15-LO expression.
Future studies pursuing IL-13 signal transduction pathwaysand the regulation of 15-LO expression in primary monocytesare somewhat constrained in this cell type. Since primarymonocytes are essentially nontransfectable by cDNA, tradi-tional approaches of expressing mutant Stats or dominant neg-ative/constitutively active p38 are not feasible. Existing mono-cytic cell lines unfortunately do not respond to IL-13 in termsof induction of 15-LO expression. The only cell line that hasbeen documented to possess this unique response is A549, anepithelial lung carcinoma. We found, however, that the IL-13-mediated signaling pathways differ between primary humanmonocytes and A549 cells because IL-13-induced 15-LO ex-pression is insensitive to SB202190 in A549 cells (B. Xu andM. K. Cathcart, unpublished observations), whereas SB202190markedly inhibits IL-13-induced 15-LO expression in mono-cytes (Fig. 7). Although murine bone marrow-derived macro-phages do not express 15-LO upon exposure to murine IL-13(A. Bhattacharjee and M. K. Cathcart, unpublished observa-tions), we are currently investigating whether murine bloodmonocytes respond similarly to human monocytes. This wouldoffer additional tools for evaluating in greater detail the mech-anisms involved in IL-13 induction of 15-LO as well as allow usto evaluate the contributions of p38 MAPK and Stats in 15-LOexpression and pathogenesis of atherosclerosis in vivo.
In summary, we report here the transcriptional regulation of15-LO by IL-13 and the involvement of p38 MAPK in theIL-13 signaling pathway of human monocytes. Our results in-dicate that IL-13 activates p38 MAPK in a time-dependentmanner and that this activation has a role in serine phosphor-ylation of Stats 1 and 3 on serine 727 as well as 15-LO expres-sion. Finally, with the Stat1 and Stat3 decoys, our results sug-gest a direct link between Stat activation and IL-13-induced15-LO expression in human monocytes. These data confirmthat the JAK-STAT and MAPK signaling pathways interact atthe level of Stat proteins and that this interaction is necessaryfor regulating 15-LO expression in IL-13-stimulated primaryhuman monocytes. The results of this study provide new in-sights into how 15-LO expression is regulated by inflammatorycytokines in human monocytes and will help us to better un-derstand the role of 15-LO and cytokines in the pathogenesisof asthma and atherosclerosis.
ACKNOWLEDGMENTS
B. Xu and A. Bhattacharjee contributed equally to this study andshare first authorship.
These studies were supported by NIH grant HL51068 awarded toM.K.C.
REFERENCES
1. Abe, K., M. Hirai, K. Mizuno, N. Higashi, T. Sekimoto, T. Miki, T. Hirano,and K. Nakajima. 2001. The YXXQ motif in gp 130 is crucial for STAT3phosphorylation at Ser727 through an H7-sensitive kinase pathway. Onco-gene 20:3464–3474.
2. Beadling, C., J. Ng, J. W. Babbage, and D. A. Cantrell. 1996. Interleukin-2activation of STAT5 requires the convergent action of tyrosine kinases anda serine/threonine kinase pathway distinct from the Raf1/ERK2 MAP kinasepathway. EMBO J. 15:1902–1913.
3. Bode, J. G., P. Gatsios, S. Ludwig, U. R. Rapp, D. Haussinger, P. C. Hein-rich, and L. Graeve. 1999. The mitogen-activated protein (MAP) kinase p38
and its upstream activator MAP kinase kinase 6 are involved in the activationof signal transducer and activator of transcription by hyperosmolarity.J. Biol. Chem. 274:30222–30227.
4. Brooks, C. J., G. Steel, J. D. Gilbert, and W. A. Harland. 1971. Lipids ofhuman atheroma. 4. Characterisation of a new group of polar sterol estersfrom human atherosclerotic plaques. Atherosclerosis 13:223–237.
5. Burysek, L., T. Syrovets, and T. Simmet. 2002. The serine protease plasmintriggers expression of MCP-1 and CD40 in human primary monocytes viaactivation of p38 MAPK and janus kinase (JAK)/STAT signaling pathways.J. Biol. Chem. 277:33509–33517.
6. Campbell, G. S., D. J. Meyer, R. Raz, D. E. Levy, J. Schwartz, and C.Carter-Su. 1995. Activation of acute phase response factor (APRF)/Stat3transcription factor by growth hormone. J. Biol. Chem. 270:3974–3979.
7. Campbell, J. S., R. Seger, J. D. Graves, L. M. Graves, A. M. Jensen, andE. G. Krebs. 1994. The Map kinase cascade. Raven Press, Baltimore, Md.
8. Cathcart, M. K., and V. A. Folcik. 2000. Lipoxygenases and atherosclerosis:protection versus pathogenesis. Free Radic. Biol. Med. 28:1726–1734.
9. Chung, J., E. Uchida, T. C. Grammer, and J. Blenis. 1997. STAT3 serinephosphorylation by ERK-dependent and -independent pathways negativelymodulates its tyrosine phosphorylation. Mol. Cell. Biol. 17:6508–6516.
10. Cobb, M. H., and E. J. Goldsmith. 1995. How MAP kinases are regulated.J. Biol. Chem. 270:14843–14846.
11. Conrad, D. J., H. Kuhn, M. Mulkins, E. Highland, and E. Sigal. 1992.Specific inflammatory cytokines regulate the expression of human monocyte15-lipoxygenase. Proc. Natl. Acad. Sci. USA 89:217–221.
12. Conrad, D. J., and M. Lu. 2000. Regulation of human 12/15-lipoxygenase byStat6-dependent transcription. Am. J. Respir. Cell Mol. Biol. 22:226–234.
13. Derijard, B., M. Hibi, I. H. Wu, T. Barrett, B. Su, T. Deng, M. Karin, andR. J. Davis. 1994. JNK1: a protein kinase stimulated by UV light and Ha-Rasthat binds and phosphorylates the c-Jun activation domain. Cell 76:1025–1037.
14. de Waal Malefyt, R., C. G. Figdor, R. Huijbens, S. Mohan-Peterson, B.Bennett, J. Culpepper, W. Dang, G. Zurawski, and J. E. de Vries. 1993.Effects of IL-13 on phenotype, cytokine production, and cytotoxic function ofhuman monocytes. Comparison with IL-4 and modulation by IFN-gamma orIL-10. J. Immunol. 151:6370–6381.
15. Enslen, H., J. Raingeaud, and R. J. Davis. 1998. Selective activation of p38mitogen-activated protein (MAP) kinase isoforms by the MAP kinase ki-nases MKK3 and MKK6. J. Biol. Chem. 273:1741–1748.
16. Folcik, V. A., R. A. Nivar-Aristy, L. P. Krajewski, and M. K. Cathcart. 1995.Lipoxygenase contributes to the oxidation of lipids in human atheroscleroticplaques. J. Clin. Investig. 96:504–510.
17. Foltz, I. N., J. C. Lee, P. R. Young, and J. W. Schrader. 1997. Hemopoieticgrowth factors with the exception of interleukin-4 activate the p38 mitogen-activated protein kinase pathway. J. Biol. Chem. 272:3296–3301.
18. Frank, D. A., S. Mahajan, and J. Ritz. 1997. B lymphocytes from patientswith chronic lymphocytic leukemia contain signal transducer and activator oftranscription (STAT) 1 and STAT3 constitutively phosphorylated on serineresidues. J. Clin. Investig. 100:3140–3148.
19. Goh, K. C., S. J. Haque, and B. R. Williams. 1999. p38 MAP kinase isrequired for STAT1 serine phosphorylation and transcriptional activationinduced by interferons. EMBO J. 18:5601–5608.
20. Gollob, J. A., C. P. Schnipper, E. A. Murphy, J. Ritz, and D. A. Frank. 1999.The functional synergy between IL-12 and IL-2 involves p38 mitogen-acti-vated protein kinase and is associated with the augmentation of STAT serinephosphorylation. J. Immunol. 162:4472–4481.
21. Han, J., J. D. Lee, L. Bibbs, and R. J. Ulevitch. 1994. A MAP kinase targetedby endotoxin and hyperosmolarity in mammalian cells. Science 265:808–811.
22. Harland, W. A., J. D. Gilbert, G. Steel, and C. J. Brooks. 1971. Lipids ofhuman atheroma. 5. The occurrence of a new group of polar sterol esters invarious stages of human atherosclerosis. Atherosclerosis 13:239–246.
23. Heydeck, D., L. Thomas, K. Schnurr, F. Trebus, W. E. Thierfelder, J. N. Ihle,and H. Kuhn. 1998. Interleukin-4 and -13 induce upregulation of the murinemacrophage 12/15-lipoxygenase activity: evidence for the involvement oftranscription factor STAT6. Blood 92:2503–2510.
24. Huang, J. S., J. Y. Guh, W. C. Hung, M. L. Yang, Y. H. Lai, H. C. Chen, andL. Y. Chuang. 1999. Role of the Janus kinase (JAK)/signal transducters andactivators of transcription (STAT) cascade in advanced glycation end-prod-uct-induced cellular mitogenesis in NRK-49F cells. Biochem. J. 342:231–238.
25. Ihn, H., K. Yamane, Y. Asano, M. Kubo, and K. Tamaki. 2002. IL-4 up-regulates the expression of tissue inhibitor of metalloproteinase-2 in dermalfibroblasts via the p38 mitogen-activated protein kinase dependent pathway.J. Immunol. 168:1895–1902.
26. Jain, N., T. Zhang, W. H. Kee, W. Li, and X. Cao. 1999. Protein kinase Cdelta associates with and phosphorylates Stat3 in an interleukin-6-dependentmanner. J. Biol. Chem. 274:24392–24400.
27. Khan, K. D., K. Shuai, G. Lindwall, S. E. Maher, J. E. Darnell, Jr., and A. L.Bothwell. 1993. Induction of the Ly-6A/E gene by interferon alpha/beta andgamma requires a DNA element to which a tyrosine-phosphorylated 91-kDaprotein binds. Proc. Natl. Acad. Sci. USA 90:6806–6810.
28. Kovarik, P., M. Mangold, K. Ramsauer, H. Heidari, R. Steinborn, A. Zotter,D. E. Levy, M. Muller, and T. Decker. 2001. Specificity of signaling by STAT1
VOL. 23, 2003 IL-13 INDUCTION OF 15-LO GENE EXPRESSION 3927
on October 24, 2014 by guest
http://mcb.asm
.org/D
ownloaded from

depends on SH2 and C-terminal domains that regulate Ser727 phosphory-lation, differentially affecting specific target gene expression. EMBO J. 20:91–100.
29. Kovarik, P., D. Stoiber, P. A. Eyers, R. Menghini, A. Neininger, M. Gaestel,P. Cohen, and T. Decker. 1999. Stress-induced phosphorylation of STAT1 atSer727 requires p38 mitogen-activated protein kinase whereas IFN-gammauses a different signaling pathway. Proc. Natl. Acad. Sci. USA 96:13956–13961.
30. Kuhn, H., D. Heydeck, R. Brinckman, and F. Trebus. 1999. Regulation ofcellular 15-lipoxygenase activity on pretranslational, translational, and post-translational levels. Lipids 34:S273–279.
31. Kyriakis, J. M., P. Banerjee, E. Nikolakaki, T. Dai, E. A. Rubie, M. F.Ahmad, J. Avruch, and J. R. Woodgett. 1994. The stress-activated proteinkinase subfamily of c-Jun kinases. Nature 369:156–160.
32. Miyazaki, T., A. Kawahara, H. Fujii, Y. Nakagawa, Y. Minami, Z. J. Liu, I.Oishi, O. Silvennoinen, B. A. Witthuhn, J. N. Ihle, and et al. 1994. Func-tional activation of Jak1 and Jak3 by selective association with IL-2 receptorsubunits. Science 266:1045–1047.
33. Murata, T., P. D. Noguchi, and R. K. Puri. 1996. IL-13 induces phosphor-ylation and activation of JAK2 Janus kinase in human colon carcinoma celllines: similarities between IL-4 and IL-13 signaling. J. Immunol. 156:2972–2978.
34. Nakagami, H., R. Morishita, K. Yamamoto, Y. Taniyama, M. Aoki, K.Matsumoto, T. Nakamura, Y. Kaneda, M. Horiuchi, and T. Ogihara. 2001.Mitogenic and antiapoptotic actions of hepatocyte growth factor throughERK, STAT3, and AKT in endothelial cells. Hypertension 37:581–586.
35. Nassar, G. M., J. D. Morrow, L. J. Roberts 2nd, F. G. Lakkis, and K. F. Badr.1994. Induction of 15-lipoxygenase by interleukin-13 in human blood mono-cytes. J. Biol. Chem. 269:27631–27634.
36. Ostareck, D. H., A. Ostareck-Lederer, M. Wilm, B. J. Thiele, M. Mann, andM. W. Hentze. 1997. mRNA silencing in erythroid differentiation: hnRNP Kand hnRNP E1 regulate 15-lipoxygenase translation from the 3� end. Cell89:597–606.
37. Ostareck-Lederer, A., D. H. Ostareck, N. Standart, and B. J. Thiele. 1994.Translation of 15-lipoxygenase mRNA is inhibited by a protein that binds toa repeated sequence in the 3� untranslated region. EMBO J. 13:1476–1481.
38. Paccani, S. R., M. Boncristiano, C. Ulivieri, M. M. D’Elios, G. Del Prete, andC. T. Baldari. 2002. Nonsteroidal anti-inflammatory drugs suppress T-cellactivation by inhibiting p38 MAPK induction. J. Biol. Chem. 277:1509–1513.
39. Palmer-Crocker, R. L., C. C. Hughes, and J. S. Pober. 1996. IL-4 and IL-13activate the JAK2 tyrosine kinase and Stat6 in cultured human vascularendothelial cells through a common pathway that does not involve thegamma c chain. J. Clin. Investig. 98:604–609.
40. Pesu, M., S. Aittomaki, K. Takaluoma, A. Lagerstedt, and O. Silvennoinen.2002. p38 Mitogen-activated protein kinase regulates interleukin-4-inducedgene expression by stimulating STAT6-mediated transcription. J. Biol.Chem. 277:38254–38261.
41. Ramsauer, K., I. Sadzak, A. Porras, A. Pilz, A. R. Nebreda, T. Decker, andP. Kovarik. 2002. p38 MAPK enhances STAT1-dependent transcriptionindependently of Ser-727 phosphorylation. Proc. Natl. Acad. Sci. USA 99:12859–12864.
42. Rosen, R. L., K. D. Winestock, G. Chen, X. Liu, L. Hennighausen, and D. S.Finbloom. 1996. Granulocyte-macrophage colony-stimulating factor prefer-entially activates the 94-kDa STAT5A and an 80-kDa STAT5A isoform inhuman peripheral blood monocytes. Blood 88:1206–1214.
43. Roy, B., A. Bhattacharjee, B. Xu, D. Ford, A. L. Maizel, and M. K. Cathcart.2002. IL-13 signal transduction in human monocytes: phosphorylation ofreceptor components, association with Jaks, and phosphorylation/activationof Stats. J. Leukoc. Biol. 72:580–589.
44. Roy, B., and M. K. Cathcart. 1998. Induction of 15-lipoxygenase expressionby IL-13 requires tyrosine phosphorylation of Jak2 and Tyk2 in humanmonocytes. J. Biol. Chem. 273:32023–32029.
45. Russell, S. M., J. A. Johnston, M. Noguchi, M. Kawamura, C. M. Bacon, M.Friedmann, M. Berg, D. W. McVicar, B. A. Witthuhn, O. Silvennoinen, et al.1994. Interaction of IL-2R beta and gamma c chains with Jak1 and Jak3:implications for XSCID and XCID. Science 266:1042–1045.
46. Sanceau, J., J. Hiscott, O. Delattre, and J. Wietzerbin. 2000. IFN-betainduces serine phosphorylation of Stat-1 in Ewing’s sarcoma cells and me-
diates apoptosis via induction of IRF-1 and activation of caspase-7. Onco-gene 19:3372–3383.
47. Schreiber, E., P. Matthias, M. M. Muller, and W. Schaffner. 1989. Rapiddetection of octamer binding proteins with ’mini-extracts’, prepared from asmall number of cells. Nucleic Acids Res. 17:6419.
48. Schuringa, J. J., L. V. Dekker, E. Vellenga, and W. Kruijer. 2001. Sequentialactivation of Rac-1, SEK-1/MKK-4, and protein kinase Cdelta is required forinterleukin-6-induced STAT3 Ser-727 phosphorylation and transactivation.J. Biol. Chem. 276:27709–27715.
49. Shankaranarayanan, P., P. Chaitidis, H. Kuhn, and S. Nigam. 2001. Acet-ylation by histone acetyltransferase CREB-binding protein/p300 of STAT6 isrequired for transcriptional activation of the 15-lipoxygenase-1 gene. J. Biol.Chem. 276:42753–42760.
50. Shuai, K., C. M. Horvath, L. H. Huang, S. A. Qureshi, D. Cowburn, and J. E.Darnell, Jr. 1994. Interferon activation of the transcription factor Stat91involves dimerization through SH2-phosphotyrosyl peptide interactions. Cell76:821–828.
51. Singh, R. P., P. Dhawan, C. Golden, G. S. Kapoor, and K. D. Mehta. 1999.One-way cross-talk between p38(MAPK) and p42/44(MAPK). Inhibition ofp38(MAPK) induces low density lipoprotein receptor expression throughactivation of the p42/44(MAPK) cascade. J. Biol. Chem. 274:19593–19600.
52. Su, B., E. Jacinto, M. Hibi, T. Kallunki, M. Karin, and Y. Ben-Neriah. 1994.JNK is involved in signal integration during costimulation of T lymphocytes.Cell 77:727–736.
53. Turkson, J., T. Bowman, J. Adnane, Y. Zhang, J. Y. Djeu, M. Sekharam,D. A. Frank, L. B. Holzman, J. Wu, S. Sebti, and R. Jove. 1999. Requirementfor Ras/Rac1-mediated p38 and c-Jun N-terminal kinase signaling in Stat3transcriptional activity induced by the Src oncoprotein. Mol. Cell. Biol.19:7519–7528.
54. Uddin, S., F. Lekmine, N. Sharma, B. Majchrzak, I. Mayer, P. R. Young,G. M. Bokoch, E. N. Fish, and L. C. Platanias. 2000. The Rac1/p38 mitogen-activated protein kinase pathway is required for interferon alpha-dependenttranscriptional activation but not serine phosphorylation of Stat proteins.J. Biol. Chem. 275:27634–27640.
55. Uddin, S., B. Majchrzak, J. Woodson, P. Arunkumar, Y. Alsayed, R. Pine,P. R. Young, E. N. Fish, and L. C. Platanias. 1999. Activation of the p38mitogen-activated protein kinase by type I interferons. J. Biol. Chem. 274:30127–30131.
56. Uddin, S., A. Sassano, D. K. Deb, A. Verma, B. Majchrzak, A. Rahman, A. B.Malik, E. N. Fish, and L. C. Platanias. 2002. Protein kinase C-delta (proteinkinase C-delta) is activated by type I interferons and mediates phosphory-lation of Stat1 on serine 727. J. Biol. Chem. 277:14408–14416.
57. Wen, Z., and J. E. Darnell, Jr. 1997. Mapping of Stat3 serine phosphoryla-tion to a single residue (727) and evidence that serine phosphorylation hasno influence on DNA binding of Stat1 and Stat3. Nucleic Acids Res. 25:2062–2067.
58. Wen, Z., Z. Zhong, and J. E. Darnell, Jr. 1995. Maximal activation oftranscription by Stat1 and Stat3 requires both tyrosine and serine phosphor-ylation. Cell 82:241–250.
59. Winston, L. A., and T. Hunter. 1995. JAK2, Ras, and Raf are required foractivation of extracellular signal-regulated kinase/mitogen-activated proteinkinase by growth hormone. J. Biol. Chem. 270:30837–30840.
60. Yokogami, K., S. Wakisaka, J. Avruch, and S. A. Reeves. 2000. Serinephosphorylation and maximal activation of STAT3 during CNTF signaling ismediated by the rapamycin target mTOR. Curr. Biol. 10:47–50.
61. Zauberman, A., D. Zipori, M. Krupsky, and R. Ben-Levy. 1999. Stress acti-vated protein kinase p38 is involved in IL-6 induced transcriptional activa-tion of STAT3. Oncogene 18:3886–3893.
62. Zhang, X., J. Blenis, H. C. Li, C. Schindler, and S. Chen-Kiang. 1995.Requirement of serine phosphorylation for formation of STAT-promotercomplexes. Science 267:1990–1994.
63. Zhong, Z., Z. Wen, and J. E. Darnell, Jr. 1994. Stat3: a STAT family memberactivated by tyrosine phosphorylation in response to epidermal growth factorand interleukin-6. Science 264:95–98.
64. Zhu, T., and P. E. Lobie. 2000. Janus kinase 2-dependent activation of p38mitogen-activated protein kinase by growth hormone. Resultant transcrip-tional activation of ATF-2 and CHOP, cytoskeletal reorganization and mi-togenesis. J. Biol. Chem. 275:2103–2114.
3928 XU ET AL. MOL. CELL. BIOL.
on October 24, 2014 by guest
http://mcb.asm
.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















