
Immunomodulation by dietary vitamin C and glucan in rainbow trout ( Oncorhynchus mykiss

Aquatic Toxicology 73 (2005) 115–127
Identification of a multixenobiotic resistance mechanism inprimary cultured epidermal cells fromOncorhynchus mykissand
the effects of environmental complex mixtures on its activity
Sharon Nı Shuilleabhaina,∗, Maria Davorena, Carmel Mothersillb, David Sheehanc,Mark G.J. Hartlc, Michael Kilemadec, Nora M. O’Brienc, John O’Halloranc,
Frank N.A.M. Van Peltc, Fiona M. Lynga
a Radiation and Environmental Science Centre, FOCAS Institute, Dublin Institute of Technology, Kevin Street, Dublin 8, Irelandb McMaster University, Hamilton, Ont., Canada
c Environmental Research Institute, University College, Cork, Ireland
Received 18 November 2004; received in revised form 9 March 2005; accepted 12 March 2005
Abstract
Multixenobiotic resistance (MXR) is a mechanism analogous to the mammalian multidrug resistance (MDR) phenotype,whereby, simultaneous resistance is conferred against the intracellular accumulation of structurally and functionally diverse,natural, endogenous and environmental toxicants. Expression of P-glycoproteins (P-gp), ATP-dependent transporters encoded for
al teleostsh species
rainbowof fish.ompounduorescentl analysis,e presenceevaluate
ctivity asted in theronmental
opy
by the mdr1 gene that have been implicated in this xenobiotic efflux mechanism, have previously been detected in normtissues involved in a secretory, absorption or a barrier function. The presence of these proteins in the epidermis of fihas not to our knowledge previously been investigated. In the present study, primary cultures of epidermis from thetroutOncorhynchus mykisswere employed to investigate whether an MXR mechanism is functional in the epidermisThe efflux of the fluorescent mdr1 substrate rhodamine 123 from the cells was significantly inhibited by verapamil, a cknown to interfere with P-gp mediated transport. The cultured epidermal cells were also observed to accumulate this fldye in a verapamil sensitive manner, thus indicating the presence of an mdr1-like mechanism. Immunocytochemicausing a monoclonal antibody (JSB1) directed against a conserved cytoplasmic P-gp epitope, also demonstrated thof P-gp-like proteins. Sediment elutriate extracts were employed as models of environmental complex mixtures tothe potential of the epidermal cultures to discriminate between samples of varying contaminant burden using MXR aan endpoint. The induction of P-gp expression was found to be in accordance with the level of contamination detecsediments from which the elutriates were extracted. The findings of the functional study also demonstrated that envipollutants, which interfere with P-gp function, could be identified using this model.© 2005 Elsevier B.V. All rights reserved.
Keywords:P-glycoprotein; Verapamil; Rhodamine 123; Chemosensitisers; Immunocytochemistry; Confocal laser scanning microsc
∗ Corresponding author. Tel.: +353 1 4027979; fax: +353 1 4027904.E-mail addresses:[email protected] (S. Nı Shuilleabhain), [email protected] (M. Davoren).
0166-445X/$ – see front matter © 2005 Elsevier B.V. All rights reserved.doi:10.1016/j.aquatox.2005.03.014

116 S. Nı Shuilleabhain et al. / Aquatic Toxicology 73 (2005) 115–127
1. Introduction
P-glycoproteins (P-gps), function in the ATP-dependent transport of moderately hydrophobiccompounds (Bard, 2000). These high molecular weightproteins are encoded for by multidrug resistance(MDR) genes (Bard, 2000; van Tellingen, 2001). Theover expression of P-gps, encoded for by the mdr1gene, is attributed to the phenomenon of mammalianmulti drug resistance, whereby, simultaneous resis-tance is conferred against the intracellular accumula-tion of structurally and functionally diverse cytotoxicdrugs (Juliano and Ling, 1976). A similar phenotype,known as multixenobiotic resistance (MXR), is in-duced in many aquatic organisms (e.g. mussels, oysters,clams, sponges and fish) exposed to toxins of natu-ral and anthropogenic origin (Kurelec, 1992; Bard etal., 2002; Smital and Sauerborn, 2002; Smital et al.,2003; Luedeking and Koehler, 2004). Expression ofmultixenobiotic resistance (MXR) proteins may be anecotoxicologically important characteristic, as it couldcritically influence the susceptibility of aquatic organ-isms to pollutants (Smital and Kurelec, 1998; Minieret al., 1999; Bard, 2000).
The tissue distribution of P-gps is indicative of theirfunction in accelerating excretion or preventing theuptake of xenobiotics. In addition to their occurrence intumour cells (Kohler et al., 1998; Cooper et al., 1999),expression of P-gps associated with an mdr1-likemechanism have been detected in normal teleost tissuest rrierf tine( t al.,1 02T thee ted.
lti-f ithta antsa ppeat ymea eta on-t ub-s st oft cen-
tration of the substance and the species of fish (Nicholset al., 1996).
Many researchers have demonstrated the value ofemploying in vitro primary cell cultures (hepatocytes)as a tool to evaluate the functional activity of the MXRmechanism in fish (Strum et al., 2001a,b; Tutundjian etal., 2002). These cultures maintain many of the in vivophysiological capabilities of the tissue from which theyare derived, while possessing the benefits of in vitro testsystems (Freshney, 1994). An in vitro primary culturesystem of rainbow trout (Oncorhynchus mykiss) skindeveloped byMothersill et al. (1995), was employed toinvestigate whether an MXR mechanism is functionalin the epidermis of fish. This explant culture systemis morphologically differentiated and contains the twomajor cell types found in salmonid epidermis; sialy-ated glycoprotein secreting goblet cells, and malphi-gian cells. Previous studies have shown the effects ofecotoxicants detected in the primary epidermal culturescomplement in vivo observations (Lyons-Alcantara etal., 1996; Kilemade et al., 2002), in addition to demon-strating the sensitivity of this in vitro system to eval-uate the effects of contaminants at the molecular andcellular level (Lyons-Alcantara et al., 1998; Dowlingand Mothersill, 1999, 2001; Kilemade and Mothersill,2000, 2001; Lyng et al., 2004).
The competitive inhibition of P-gp substrates,such as the non-toxic fluorescent dye rhodamine 123(Rh123) is considered as evidence of the functional ac-tivity of an MXR mechanism in cells (Toomey andE al.,2 i-c iumc peti-tm reso thee uno-c
XRa thisp in di-r nte l.,1 eo itroc ntals n the
hat are involved in a secretory, absorption or baunction such as the liver, kidneys, gills and intesHemmer et al., 1995; Schramm et al., 1995; Bard e998; Doi et al., 2000; Smital and Sauerborn, 20).o our knowledge, the expression of P-gps inpidermis of fish has not been previously investiga
Teleost epidermis is a metabolically active, muunctional tissue that is in constant direct contact whe aqueous environment (Bullock et al., 1978). It actss a physical barrier to both pathogens and toxicnd has been shown to secrete compounds that a
o have specific defence functions, such as lysoznd complement (Harrell et al., 1976; Hjelmelandl., 1983). In addition, it has been demonstrated to c
ribute substantially to the exchange of lipophilic stances (Nichols et al., 1996). Absorption of toxicanthrough the epidermis may range from 1.6 to 50%otal absorption depending on the nature and con
r
pel, 1993; Minier and Moore, 1996; Strum et001a,b). In this study, we employed confocal mroscopy to visualise the ability of verapamil, a calchannel blocker frequently used as a model comive inhibitor (Ford and Hait, 1990; Germann, 1996), toodulate the transport of Rh123 in the in vitro cultuf trout epidermal cells. The expression of P-gp inpidermal cultures was also examined using immytochemical techniques.
Several studies have proposed the potential of Mctivity as a biomarker of contaminant exposure, ashenotype has been demonstrated to be inducedect proportion to the level of pollution in the ambienvironment (Smital and Kurelec, 1998; Minier et a999; Albertus and Laine, 2001). The second objectivf this study was to evaluate the potential of the in vulture system to discriminate between environmeamples of varying contaminant burden, based o

S. Nı Shuilleabhain et al. / Aquatic Toxicology 73 (2005) 115–127 117
modulating effects of these samples on the expressionand function of the MXR mechanism. Sediment elu-triate extracts were employed as model complex mix-tures, as sediment is a known repository for persistentanthropogenic contaminants.
2. Materials and methods
2.1. Primary epidermal cell culture
Healthy juvenile rainbow trout (O. mykiss) wereobtained from a commercial fish farm in CountyTipperary, Ireland. Epidermal cell outgrowths wereinitiated using a modification of the primary explantculture technique developed byMothersill et al. (1995).Briefly, the trout were killed using an overdose ofthe anaesthetic 4-ethyl-aminobenzoate (BDH Labora-tory Supplies, Poole, UK). A section of skin belowthe dorsal fin was removed and immersed in RPMI-1640 complete growth medium (Gibco Biocult, Pais-ley, UK) supplemented with 12% fetal calf serum,8% horse serum, 0.05 IU/mL human recombinant in-sulin, 1�g/mL hydrocortisone, 40 IU/mL penicillin,40�g/mL streptomycin solution, 2 mMl-glutamineand 15 mM HEPES. All supplements were purchasedfrom Gibco Biocult (Paisley, UK), except human re-combinant insulin and hydrocortisone, which were pur-chased from Novo Nordisk (Copenhagen) and Sigma(Dublin), respectively. The epidermis was asepticallyd cuti r-m cmt mLo bea ons nc,D resw ot-t rew
2e
2ed
a a,I e ef-
flux of rhodamine 123 (Rh123) (Molecular Probes, TheNetherlands), in the in vitro cultures of trout epidermalcells. Rh123 is a fluorescent substrate of mammalianmdr1-type P-gps, which preferentially accumulates inthe mitochondria of cells (Chieli et al., 1994).
2.2.1.1. Efflux assay.Following 96 h in culture,growth medium was removed and the outgrowths fromthe epidermal explants were rinsed with phosphatebuffered saline containing 1 mM CaCl2 and MgCl2(Ca2+/Mg2+ buffer). Cultures were incubated for 2 hin Clonetics keratinocyte growth medium (KGM) (Clo-netics Corporation, USA) containing 5�M Rh123. Thecultures were rinsed with Ca2+/Mg2+ buffer and incu-bated for a further 30 min to 4 h in KGM alone or incombination with 10, 20 or 50�M verapamil to de-termine the optimal concentration of verapamil thatwould inhibit Rh123 efflux. The cultures were subse-quently rinsed with Ca2+/Mg2+ buffer and fluorescencewas measured using a Zeiss LSM Meta 510 confocalmicroscope (laser excitation 488 nm, bandpass (BP)emission filter 505–550 nm). Eight areas of the cultureoutgrowth were examined at random, in three replicatecultures, and repeated in at least two independent ex-periments. Fluorescence measurements were correctedfor cell number by dividing the fluorescence intensitymeasured by the number of cells in the field of viewand expressed as a percentage of control values.
2.2.1.2. Accumulation assay.Growth medium was re-m antswc h inK nw ntlyr asm ores-c mbera
22 -p cul-t GMaR laro a-l ced
issected free of muscle and fibrous tissue andnto approximately 2 mm2 pieces. Individual epide
al sections were placed mucosal side up in 242
issue culture flasks (Nunc, Denmark) containing 2f complete growth medium. Explant cultures tonalysed using confocal microscopy were platedterile glass coverslips in 6-well microplates (Nuenmark) containing 1.5 mL growth medium. Cultuere maintained in a refrigerated incubator (Leec, N
ingham, UK) at 18◦C under a normoxic atmospheithout media change.
.2. In vitro determination of mdr1-like activity inpidermal cells
.2.1. Confocal microscopyFunctioning of mdr1-like activity was determin
s the ability ofR(+) verapamil hydrochloride (Sigmreland) to enhance the accumulation and inhibit th
oved and the outgrowths from the epidermal explere rinsed with Ca2+/Mg2+ buffer following 96 h inulture. The cultures were incubated from 1 to 4GM containing 5�M Rh123 alone or in additioith 10�M verapamil. The cultures were subseque
insed with Ca2+/Mg2+ buffer and fluorescence weasured, as described for the efflux assay. Flu
ence measurements were corrected for cell nund expressed as a percentage of control values.
.2.2. Immunocytochemical analysis
.2.2.1. Exposure conditions.To investigate the exression of P-gp-like proteins in primary epidermal
ures, the explant outgrowths were incubated in Klone (control cultures) or in KGM containing 5�Mh123 for 4 h following 96 h in culture. The celluutgrowths were fixed in 10% neutral buffered form
in for 1 h. The formalin was subsequently repla

118 S. Nı Shuilleabhain et al. / Aquatic Toxicology 73 (2005) 115–127
with phosphate buffered saline (PBS) and the cultureswere stored at 4◦C. Immunocytochemical analysis wasconducted within 24 h of fixation.
2.2.2.2. Immunocytochemistry.Monoclonal mouseP-gp antibody clone JSB1 (NOVO Castra, UK) wasused to detect and quantify P-gp labelling in the rain-bow trout epidermal cultures. This antibody identifiesa highly conserved cytoplasmic epitope of the humanplasma membrane associated with a 170–180 kDaglycoprotein, the expression of which is stronglycorrelated with the degree of MDR (Scheper et al.,1988). Paraffin embedded sections of normal humanliver obtained from the Cell Pathology Laboratory,Dublin Institute of Technology, was employed as apositive control. The avidin–biotin complex (ABC)method of immunoperoxidase staining was employedto visualise P-gp expression. Immunostaining wasperformed using a mouse monoclonal Vectastain ABCkit (Vectastain Corporation Burlingame, USA), whichis based upon the standard indirect immunoperoxidasetechnique. Culture flasks containing the formalin fixedcultures were cracked open using a pair of pliers toallow in situ processing of the cellular outgrowth.The cultures were rinsed with PBS and subsequentlyincubated with 3% hydrogen peroxide (H2O2) inmethanol for 5 min to quench endogenous peroxidaseenzyme activity. Dilute normal horse blocking serumwas applied to the cultures for 20 min. Excess serumwas blotted from the cultures prior to a 1 h incubationw asd nota erer edb ul-t stainA ishp inga for1 ,3d a,D tlyc HL ing ).
2 ti-fi age
Analyser (Leica Microsystems Holdings, Ernst-Leitz-Strasse, Wetzlar, Germany), by counting the number ofimmunopositive cells along two transects perpendicu-lar to each other through the centre of the outgrowth.This ensured that any differences between the cells atthe centre and the edge regions of the outgrowth couldbe accounted for. The immunocytochemical stainingof the cells was classified as intensely positive, weaklypositive or negative, relative to control treatment cellsand is expressed as a percentage of the total numberof cells quantified (over 3000 cells examined per treat-ment).
2.3. In vitro characterisation of mdr1-like activityfollowing exposure to sediment elutriate samples
2.3.1. Site selection and sediment samplingThree sampling sites from around the Irish coast
were selected on the basis of previously publishedreports that demonstrated them to be representativeof a range of contaminant burdens (Marine Institute,1999; Byrne and O’Halloran, 2000). Ballymacoda Es-tuary, Co. Cork (7◦55′W, 51◦52′N) was chosen asa relatively uncontaminated site while the DouglasEstuary, Co., Cork (8◦23′W, 51◦52′N) and the EastWall site in the Liffey Estuary, Co., Dublin (06◦07′W,53◦20′N) are moderately and highly contaminated, re-spectively. At low tide oxic surface sediment (top 2 cm)was sampled from random areas each measuring ap-proximately 300 cm2. Sub-samples were compositedap r nol ricanS
2s
ctedt eno n ofa ysi-c odaae theE dy.T de-t ovend ic
ith the primary antibody (P-gp JSB1), which wiluted 1:60 in PBS. The primary antibody waspplied to negative control slides. The cultures winsed with PBS followed by the application of dilutiotinylated secondary antibody for 30 min. The c
ures were subsequently incubated with the VectaBC reagent (avidin DH:biotinylated horseraderoxidase H complex) for a further 30 min. Followfinal PBS rinse, the cultures were incubated
0 min in the dark with the peroxidase substrate 3′-iaminobenzidine tetrahydrochloride (DAB) (Sigmublin). Cultures were rinsed in tap water, lighounterstained with Harris’s haematoxylin (BDaboratory Supplies, Poole, UK) and mountedlycerol jelly (BDH Laboratory Supplies, Poole, UK
.2.2.3. Quantification.P-gp expression was quaned in the epidermal cultures using a Leica Im
nd homogenised thoroughly and stored at 4◦C inolyethylene bags. Whole sediment was stored fo
onger than 2 weeks as recommended by the Ameociety for Testing and Materials (ASTM, 2004).
.3.2. Physical and chemical characterisation ofediments
Comprehensive chemical analyses were conduo provide an indication of the contaminant burdf the sediments and to assist in the interpretationy observed alterations in mdr1-like activity. Phal and chemical characterisation of the Ballymacnd Douglas sediments were conducted byKilemadet al. (2004). Characterisation of sediments fromast Wall site was performed as part of this stuhe percentage dry weight of the sediments was
ermined using sub-samples of each sediment byrying at 105◦C to a constant weight. Total organ

S. Nı Shuilleabhain et al. / Aquatic Toxicology 73 (2005) 115–127 119
carbon (TOC) was determined by ALcontrol laborato-ries (Chester, UK) using the combustion method, whichconformed to ISO 17025. The accuracy of this methodwas ensured using a certified reference material (CRM)(Alpha Resources Inc. CSN standard AR873). The ac-curacy was further assessed by the inclusion of a testmaterial (obtained from the QUASIMEME project of-fice), which was found to be within acceptable lim-its. Sediments were wet sieved using the gravimetricmethod (Loring and Rantala, 1992), to obtain the fine(<63�m) sediment fraction on which all chemical anal-yses were conducted. Heavy metal analyses (Pb, Cd,Cu, Zn) were conducted by the Marine Institute (Ab-botstown, Dublin, Ireland). Sediments were acid di-gested using reverse aqua regia (3:1 (v/v) HNO3:HCl)and hydrofluoric acid. Lead, cadmium, and copperconcentrations were determined using Graphite Fur-nace Atomic Absorption Spectrometry with Zeemanbackground correction (Varian SpectrAA 220Z). Zincconcentrations were determined using Flame AtomicAbsorption Spectroscopy (Varian SpectrAA 20 Plus).The East Wall sediment was sent to RIVO, Nether-lands Institute for Fisheries Research, The Netherlands(EN/ISO biphenyls (PCBs) 17025, STERLAB) for theanalyses of polycyclic aromatic hydrocarbons (PAHs),polychlorinated and organochlorine pesticides (OCPs).Table 1 summarises the methodology employed forthe extraction, separation, detection and quantificationof each class of organic contaminant. A comprehen-sive analytical quality assurance program, involvingt h asb dingC
2o
iona
Engler, 1974; USEPA, 1977). Deionised water wasadded to 10 g sub-samples of sediment in a 1:4 (w/v)ratio based on the sediment dry weight. The slurrywas shaken at 240 rpm for 1 h and then centrifugedat 1200×g for 30 min at 4◦C. The supernatant wascollected as elutriate and conductivity, salinity and pHwere measured following filtration through a 0.2�mfilter (Nalgene, NY, USA). The elutriate was stored inprimed glass bottles with minimal headspace at 4◦C inthe dark prior to toxicity testing within 24 h of prepa-ration.
Powdered media (RPMI-1640, Gibco Biocult, Pais-ley, UK) was reconstituted with elutriate extracts toallow the whole elutriate samples to be exposed tothe primary epidermal cultures. Following reconstitu-tion, the samples were sterile filtered and supplementedas outlined for the complete growth medium. Serumwas not added to the elutriate samples employed inthe confocal microscopy studies. Control medium wasreconstituted with deionised water and supplementedaccordingly. The osmolality of all test concentrationswas measured using an Osmomat 030 cryoscopic os-mometer (Gonotec, Berlin, Germany). To account forthe potential effects of sample osmolality on experi-mental findings, an osmolality control correspondingto the final osmolality of the individual elutriate sam-ples was prepared by reconstituting powdered mediumusing a balanced salt solution (Machii and Wada, 1989).
2.3.4. Confocal microscopyter-
f eso ed,e ithC dw sedt ra es,
TA etectio ll sediment
C Se tection
P VydaP CP CDO CP CD
he routine testing of quality control samples, suclanks, replicates and reference materials (incluRMs), underpinned all chemical testing.
.3.3. Preparation of elutriate extracts andsmolality controls
Elutriate extracts were prepared with modificatccording to the Standard Elutriate Test (Keely and
able 1n overview of the analytical methodologies employed for the d
ompounds Extraction procedure
AHs Soxhlet,n-hexane/tolueneCBs Soxhlet, hexane/acetoneCPs Soxhlet, hexane/acetone
a High pressure liquid chromatography.b Gas chromatography–electron capture detection.
The ability of the sediment elutriate extracts to inere with mdr1-like activity in the epidermal culturfO.mykisswas investigated. As previously describxplants that were in culture for 96 h were rinsed wa2+/Mg2+ buffer. Cultures were initially incubateith Rh123 for 2 h. They were subsequently rin
horoughly with Ca2+/Mg2+ buffer and incubated fofurther 2 h with the individual sediment elutriat
n and quantification of the organic contaminants in the East Wa
paration procedure—chromatograph columns De
c HPLCa
-Sil 8, CP-Sil 19 GC-Eb
-Sil 8, CP-Sil 19 GC-E
120 S. Nı Shuilleabhain et al. / Aquatic Toxicology 73 (2005) 115–127
osmolality controls or medium controls prior to mea-surement of Rh123 fluorescence. Eight areas of theculture outgrowth were examined at random, in threereplicate cultures, which was repeated in at least twoindependent experiments. The measurements were cor-rected for cell number and the readings from the elu-triate and osmolality exposed cultures were expressedas a percentage of the medium control values.
2.3.5. ImmunocytochemistryThe primary epidermal cell outgrowths were incu-
bated with the individual elutriate extracts, osmolalitycontrol or control medium for a 48 h period following96 h in culture. They were fixed in 10% neutral bufferedformalin for 1 h, which was subsequently replaced withPBS. Immunocytochemical analysis was conductedwithin 24 h of fixation as previously described. Carewas taken to avoid quantifying of apoptotic or necroticcells which may have been falsely positive due to non-specific staining.
2.4. Statistical analysis
Data were expressed as the arithmetic mean of trip-licate samples which were repeated in at least two in-dependent experiments± standard error of the mean(S.E.M.). Coefficient of variation (CV) for control datawas calculated to ascertain reproducibility when appro-priate. Statistical analysis on the proportion data setswas conducted using Chi-squared analysis on the nu-m e in-tw alv us-i ).T Br g-n
3
3ca
n ofR h of
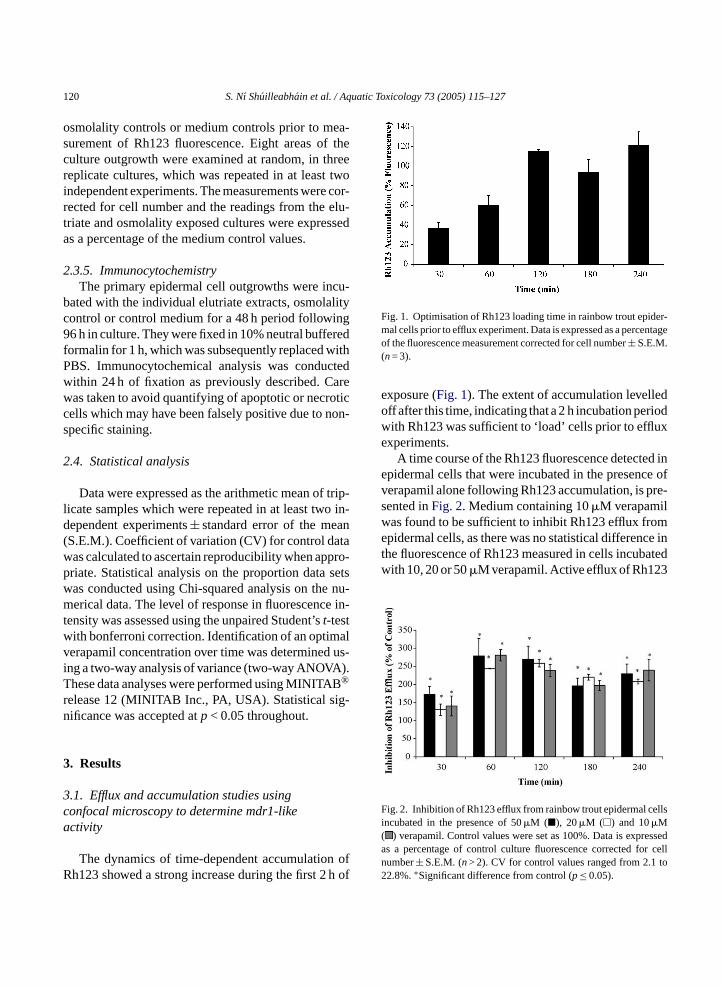
Fig. 1. Optimisation of Rh123 loading time in rainbow trout epider-mal cells prior to efflux experiment. Data is expressed as a percentageof the fluorescence measurement corrected for cell number± S.E.M.(n= 3).
exposure (Fig. 1). The extent of accumulation levelledoff after this time, indicating that a 2 h incubation periodwith Rh123 was sufficient to ‘load’ cells prior to effluxexperiments.
A time course of the Rh123 fluorescence detected inepidermal cells that were incubated in the presence ofverapamil alone following Rh123 accumulation, is pre-sented inFig. 2. Medium containing 10�M verapamilwas found to be sufficient to inhibit Rh123 efflux fromepidermal cells, as there was no statistical difference inthe fluorescence of Rh123 measured in cells incubatedwith 10, 20 or 50�M verapamil. Active efflux of Rh123
Fig. 2. Inhibition of Rh123 efflux from rainbow trout epidermal cellsincubated in the presence of 50�M (�), 20�M (�) and 10�M( ) verapamil. Control values were set as 100%. Data is expressedas a percentage of control culture fluorescence corrected for celln to2
erical data. The level of response in fluorescencensity was assessed using the unpaired Student’st-testith bonferroni correction. Identification of an optimerapamil concentration over time was determinedng a two-way analysis of variance (two-way ANOVAhese data analyses were performed using MINITA®
elease 12 (MINITAB Inc., PA, USA). Statistical siificance was accepted atp< 0.05 throughout.
. Results
.1. Efflux and accumulation studies usingonfocal microscopy to determine mdr1-likectivity
The dynamics of time-dependent accumulatioh123 showed a strong increase during the first 2
umber± S.E.M. (n> 2). CV for control values ranged from 2.12.8%.∗Significant difference from control (p≤ 0.05).
S. Nı Shuilleabhain et al. / Aquatic Toxicology 73 (2005) 115–127 121
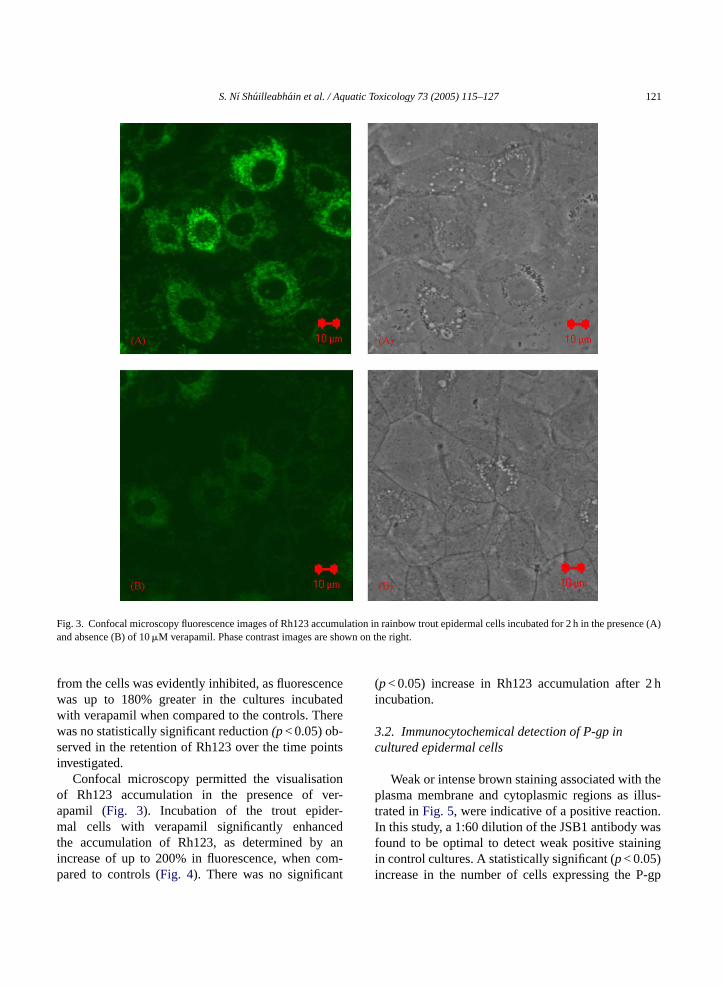
Fig. 3. Confocal microscopy fluorescence images of Rh123 accumulation in rainbow trout epidermal cells incubated for 2 h in the presence (A)and absence (B) of 10�M verapamil. Phase contrast images are shown on the right.
from the cells was evidently inhibited, as fluorescencewas up to 180% greater in the cultures incubatedwith verapamil when compared to the controls. Therewas no statistically significant reduction(p< 0.05) ob-served in the retention of Rh123 over the time pointsinvestigated.
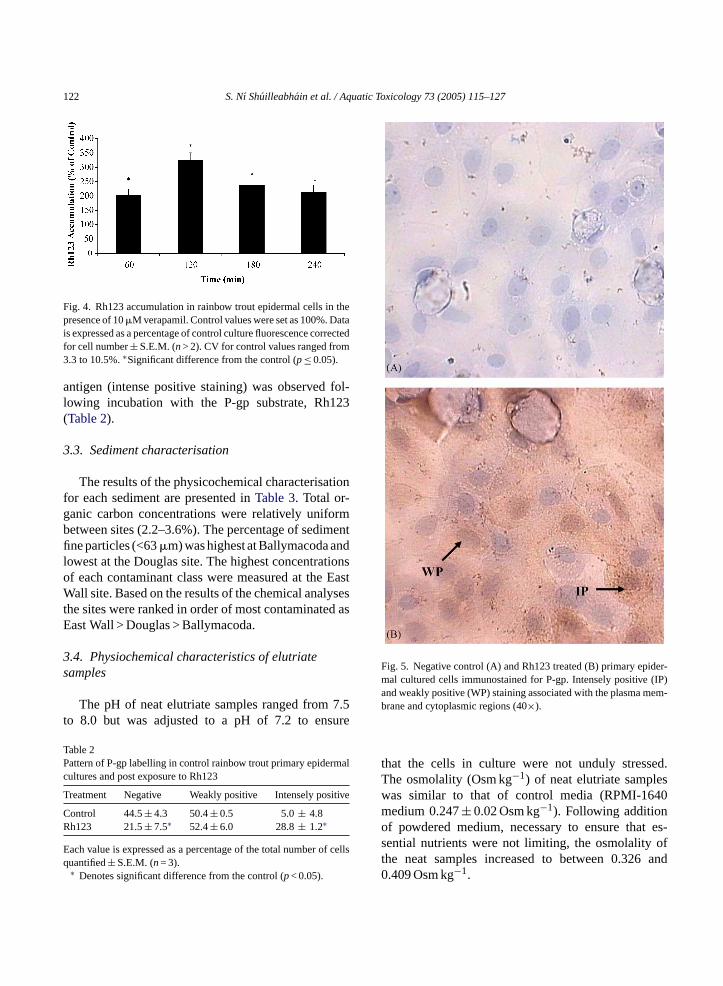
Confocal microscopy permitted the visualisationof Rh123 accumulation in the presence of ver-apamil (Fig. 3). Incubation of the trout epider-mal cells with verapamil significantly enhancedthe accumulation of Rh123, as determined by anincrease of up to 200% in fluorescence, when com-pared to controls (Fig. 4). There was no significant
(p< 0.05) increase in Rh123 accumulation after 2 hincubation.
3.2. Immunocytochemical detection of P-gp incultured epidermal cells
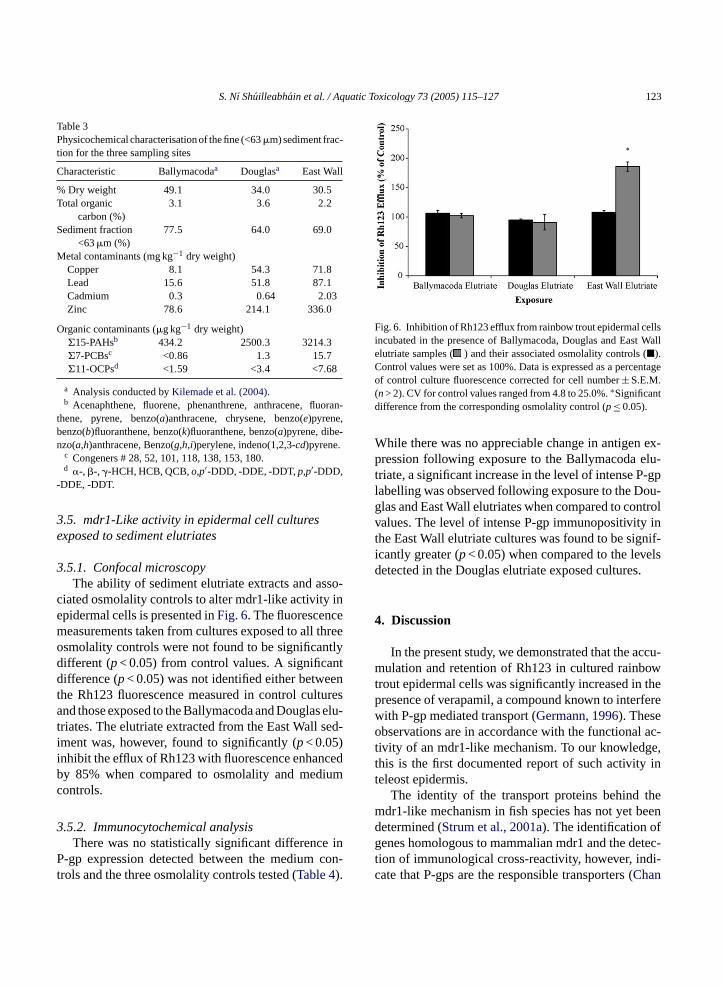
Weak or intense brown staining associated with theplasma membrane and cytoplasmic regions as illus-trated inFig. 5, were indicative of a positive reaction.In this study, a 1:60 dilution of the JSB1 antibody wasfound to be optimal to detect weak positive stainingin control cultures. A statistically significant (p< 0.05)increase in the number of cells expressing the P-gp
122 S. Nı Shuilleabhain et al. / Aquatic Toxicology 73 (2005) 115–127
Fig. 4. Rh123 accumulation in rainbow trout epidermal cells in thepresence of 10�M verapamil. Control values were set as 100%. Datais expressed as a percentage of control culture fluorescence correctedfor cell number± S.E.M. (n> 2). CV for control values ranged from3.3 to 10.5%.∗Significant difference from the control (p≤ 0.05).
antigen (intense positive staining) was observed fol-lowing incubation with the P-gp substrate, Rh123(Table 2).
3.3. Sediment characterisation
The results of the physicochemical characterisationfor each sediment are presented inTable 3. Total or-ganic carbon concentrations were relatively uniformbetween sites (2.2–3.6%). The percentage of sedimentfine particles (<63�m) was highest at Ballymacoda andlowest at the Douglas site. The highest concentrationsof each contaminant class were measured at the EastWall site. Based on the results of the chemical analysesthe sites were ranked in order of most contaminated asEast Wall > Douglas > Ballymacoda.
3.4. Physiochemical characteristics of elutriatesamples
The pH of neat elutriate samples ranged from 7.5to 8.0 but was adjusted to a pH of 7.2 to ensure
Table 2Pattern of P-gp labelling in control rainbow trout primary epidermalcultures and post exposure to Rh123
Treatment Negative Weakly positive Intensely positive
Control 44.5± 4.3 50.4± 0.5 5.0± 4.8Rh123 21.5± 7.5∗ 52.4± 6.0 28.8± 1.2∗
Each value is expressed as a percentage of the total number of cellsq
Fig. 5. Negative control (A) and Rh123 treated (B) primary epider-mal cultured cells immunostained for P-gp. Intensely positive (IP)and weakly positive (WP) staining associated with the plasma mem-brane and cytoplasmic regions (40×).
that the cells in culture were not unduly stressed.The osmolality (Osm kg−1) of neat elutriate sampleswas similar to that of control media (RPMI-1640medium 0.247± 0.02 Osm kg−1). Following additionof powdered medium, necessary to ensure that es-sential nutrients were not limiting, the osmolality ofthe neat samples increased to between 0.326 and0.409 Osm kg−1.
uantified± S.E.M. (n= 3).∗ Denotes significant difference from the control (p< 0.05).
S. Nı Shuilleabhain et al. / Aquatic Toxicology 73 (2005) 115–127 123
Table 3Physicochemical characterisation of the fine (<63�m) sediment frac-tion for the three sampling sites
Characteristic Ballymacodaa Douglasa East Wall
% Dry weight 49.1 34.0 30.5Total organic
carbon (%)3.1 3.6 2.2
Sediment fraction<63�m (%)
77.5 64.0 69.0
Metal contaminants (mg kg−1 dry weight)Copper 8.1 54.3 71.8Lead 15.6 51.8 87.1Cadmium 0.3 0.64 2.03Zinc 78.6 214.1 336.0
Organic contaminants (�g kg−1 dry weight)�15-PAHsb 434.2 2500.3 3214.3�7-PCBsc <0.86 1.3 15.7�11-OCPsd <1.59 <3.4 <7.68
a Analysis conducted byKilemade et al. (2004).b Acenaphthene, fluorene, phenanthrene, anthracene, fluoran-
thene, pyrene, benzo(a)anthracene, chrysene, benzo(e)pyrene,benzo(b)fluoranthene, benzo(k)fluoranthene, benzo(a)pyrene, dibe-nzo(a,h)anthracene, Benzo(g,h,i)perylene, indeno(1,2,3-cd)pyrene.
c Congeners # 28, 52, 101, 118, 138, 153, 180.d �-, �-, �-HCH, HCB, QCB,o,p′-DDD, -DDE, -DDT,p,p′-DDD,
-DDE, -DDT.
3.5. mdr1-Like activity in epidermal cell culturesexposed to sediment elutriates
3.5.1. Confocal microscopyThe ability of sediment elutriate extracts and asso-
ciated osmolality controls to alter mdr1-like activity inepidermal cells is presented inFig. 6. The fluorescencemeasurements taken from cultures exposed to all threeosmolality controls were not found to be significantlydifferent (p< 0.05) from control values. A significantdifference (p< 0.05) was not identified either betweenthe Rh123 fluorescence measured in control culturesand those exposed to the Ballymacoda and Douglas elu-triates. The elutriate extracted from the East Wall sed-iment was, however, found to significantly (p< 0.05)inhibit the efflux of Rh123 with fluorescence enhancedby 85% when compared to osmolality and mediumcontrols.
3.5.2. Immunocytochemical analysisThere was no statistically significant difference in
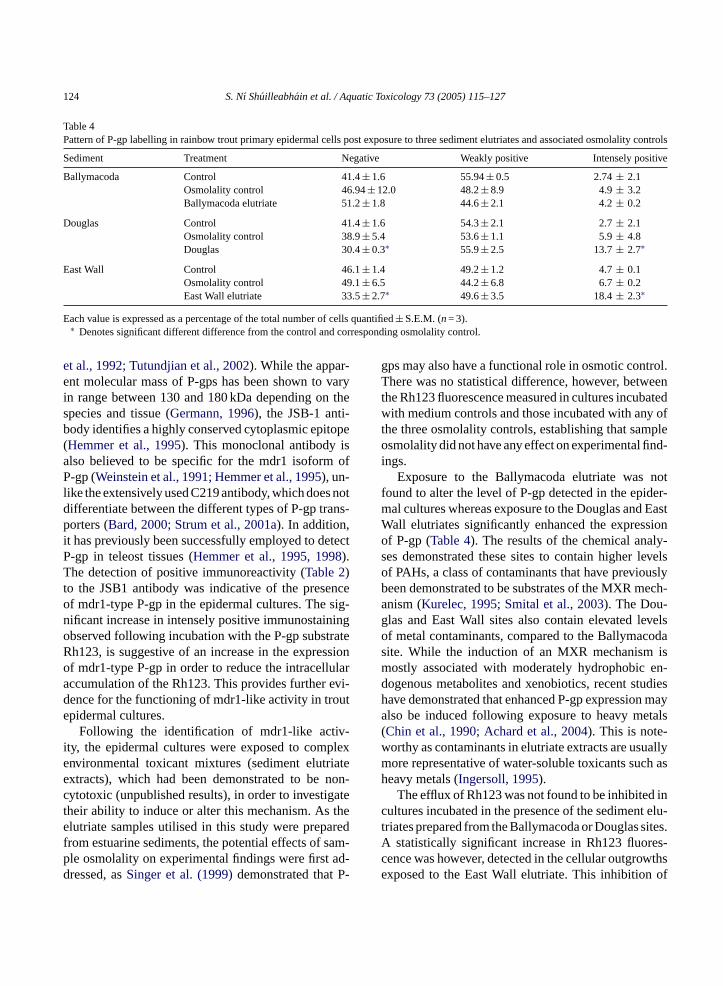
P-gp expression detected between the medium con-trols and the three osmolality controls tested (Table 4).
Fig. 6. Inhibition of Rh123 efflux from rainbow trout epidermal cellsincubated in the presence of Ballymacoda, Douglas and East Wallelutriate samples ( ) and their associated osmolality controls (�).Control values were set as 100%. Data is expressed as a percentageof control culture fluorescence corrected for cell number± S.E.M.(n> 2). CV for control values ranged from 4.8 to 25.0%.∗Significantdifference from the corresponding osmolality control (p≤ 0.05).
While there was no appreciable change in antigen ex-pression following exposure to the Ballymacoda elu-triate, a significant increase in the level of intense P-gplabelling was observed following exposure to the Dou-glas and East Wall elutriates when compared to controlvalues. The level of intense P-gp immunopositivity inthe East Wall elutriate cultures was found to be signif-icantly greater (p< 0.05) when compared to the levelsdetected in the Douglas elutriate exposed cultures.
4. Discussion
In the present study, we demonstrated that the accu-mulation and retention of Rh123 in cultured rainbowtrout epidermal cells was significantly increased in thepresence of verapamil, a compound known to interferewith P-gp mediated transport (Germann, 1996). Theseobservations are in accordance with the functional ac-tivity of an mdr1-like mechanism. To our knowledge,this is the first documented report of such activity inteleost epidermis.
The identity of the transport proteins behind themdr1-like mechanism in fish species has not yet beendetermined (Strum et al., 2001a). The identification ofgenes homologous to mammalian mdr1 and the detec-tion of immunological cross-reactivity, however, indi-cate that P-gps are the responsible transporters (Chan
124 S. Nı Shuilleabhain et al. / Aquatic Toxicology 73 (2005) 115–127
Table 4Pattern of P-gp labelling in rainbow trout primary epidermal cells post exposure to three sediment elutriates and associated osmolality controls
Sediment Treatment Negative Weakly positive Intensely positive
Ballymacoda Control 41.4± 1.6 55.94± 0.5 2.74± 2.1Osmolality control 46.94± 12.0 48.2± 8.9 4.9± 3.2Ballymacoda elutriate 51.2± 1.8 44.6± 2.1 4.2± 0.2
Douglas Control 41.4± 1.6 54.3± 2.1 2.7± 2.1Osmolality control 38.9± 5.4 53.6± 1.1 5.9± 4.8Douglas 30.4± 0.3∗ 55.9± 2.5 13.7± 2.7∗
East Wall Control 46.1± 1.4 49.2± 1.2 4.7± 0.1Osmolality control 49.1± 6.5 44.2± 6.8 6.7± 0.2East Wall elutriate 33.5± 2.7∗ 49.6± 3.5 18.4± 2.3∗
Each value is expressed as a percentage of the total number of cells quantified± S.E.M. (n= 3).∗ Denotes significant different difference from the control and corresponding osmolality control.
et al., 1992; Tutundjian et al., 2002). While the appar-ent molecular mass of P-gps has been shown to varyin range between 130 and 180 kDa depending on thespecies and tissue (Germann, 1996), the JSB-1 anti-body identifies a highly conserved cytoplasmic epitope(Hemmer et al., 1995). This monoclonal antibody isalso believed to be specific for the mdr1 isoform ofP-gp (Weinstein et al., 1991; Hemmer et al., 1995), un-like the extensively used C219 antibody, which does notdifferentiate between the different types of P-gp trans-porters (Bard, 2000; Strum et al., 2001a). In addition,it has previously been successfully employed to detectP-gp in teleost tissues (Hemmer et al., 1995, 1998).The detection of positive immunoreactivity (Table 2)to the JSB1 antibody was indicative of the presenceof mdr1-type P-gp in the epidermal cultures. The sig-nificant increase in intensely positive immunostainingobserved following incubation with the P-gp substrateRh123, is suggestive of an increase in the expressionof mdr1-type P-gp in order to reduce the intracellularaccumulation of the Rh123. This provides further evi-dence for the functioning of mdr1-like activity in troutepidermal cultures.
Following the identification of mdr1-like activ-ity, the epidermal cultures were exposed to complexenvironmental toxicant mixtures (sediment elutriateextracts), which had been demonstrated to be non-cytotoxic (unpublished results), in order to investigatetheir ability to induce or alter this mechanism. As theelutriate samples utilised in this study were preparedf am-p ad-d -
gps may also have a functional role in osmotic control.There was no statistical difference, however, betweenthe Rh123 fluorescence measured in cultures incubatedwith medium controls and those incubated with any ofthe three osmolality controls, establishing that sampleosmolality did not have any effect on experimental find-ings.
Exposure to the Ballymacoda elutriate was notfound to alter the level of P-gp detected in the epider-mal cultures whereas exposure to the Douglas and EastWall elutriates significantly enhanced the expressionof P-gp (Table 4). The results of the chemical analy-ses demonstrated these sites to contain higher levelsof PAHs, a class of contaminants that have previouslybeen demonstrated to be substrates of the MXR mech-anism (Kurelec, 1995; Smital et al., 2003). The Dou-glas and East Wall sites also contain elevated levelsof metal contaminants, compared to the Ballymacodasite. While the induction of an MXR mechanism ismostly associated with moderately hydrophobic en-dogenous metabolites and xenobiotics, recent studieshave demonstrated that enhanced P-gp expression mayalso be induced following exposure to heavy metals(Chin et al., 1990; Achard et al., 2004). This is note-worthy as contaminants in elutriate extracts are usuallymore representative of water-soluble toxicants such asheavy metals (Ingersoll, 1995).
The efflux of Rh123 was not found to be inhibited incultures incubated in the presence of the sediment elu-triates prepared from the Ballymacoda or Douglas sites.A es-c wthse of
rom estuarine sediments, the potential effects of sle osmolality on experimental findings were firstressed, asSinger et al. (1999)demonstrated that P
statistically significant increase in Rh123 fluorence was however, detected in the cellular outgroxposed to the East Wall elutriate. This inhibition

S. Nı Shuilleabhain et al. / Aquatic Toxicology 73 (2005) 115–127 125
Rh123 efflux indicates the presence of contaminantsknown as chemosensitisers, in the East Wall elutri-ates. These inhibitors are capable of disrupting P-gpmediated transport by a number of mechanisms. Forexample, chemosensitisers that are P-gp substrates,such as verapamil, competitively inhibit the efflux ofnon-modulating substrates such as Rh123. These sub-strates tend to be more lipophilic and traverse bilayerstoo quickly to measure (Eytan et al., 1996) as op-posed to non-modulating substrates, which diffuse rela-tively slowly across membranes (Sharom, 1997). Somechemosensitising agents influence P-gp indirectly bydisrupting transport, e.g. ATPase inhibitors and mem-brane fluidisers (Sharom, 1997), thus leading to anincreased accumulation of MXR substrates. Hydropho-bic pesticides, such as pentachlorophenol and dacthalhave been shown to have significant inhibitory effectson MXR activity as they bind to P-gp but are not trans-ported (Cornwall et al., 1995; Bain and LeBlanc, 1996).Kurelec et al. (1998)also demonstrated that innocuousmoderately lipophilic substances known to be presentin household waste etc. functioned as chemosensitisersas they saturated the MXR mechanism when present athigh concentrations. As the expression of mdr1-type P-gp was enhanced following exposure to the East Wallelutriate (as indicated by the immunocytochemical re-sults), the inhibition of Rh123 efflux is most likelydue to competitive inhibition with chemosensitiser(s)which are also a P-gp substrate.
The presence of chemosensitising agents in thee s in-h re-s taint inn fromt ousa rchi edi-a envi-r icalc d.
ityi icals thee re-s ctiono tam-i lso
demonstrated that environmental pollutants, which in-terfere with P-gp function could be identified using thismodel. While subsequent in vivo validation studies arerequired, the presence of an inducible P-gp phenotypeand an mdr1-like efflux mechanism in the epidermalcultures indicate that this in vitro model could proveto be a valuable tool in the hazard assessment of en-vironmental contaminants. In addition, the transfer ofthis culture technique to other teleosts could facilitatethe investigation of functional activity and expressionof the MXR mechanism in the epidermis of differentpiscine species.
In conclusion, this study demonstrates the presenceof an mdr1-like mechanism in trout epidermal cells,which can be induced/modulated by environmentalcontaminants. Subsequent studies should address therelevance of P-gp mediated transport in teleost epider-mis as a defence against the deleterious effects of tox-icants.
Acknowledgements
We gratefully acknowledge Linda Tyrell of the Ma-rine Institute, Abbotstown, Dublin for conducting themetal analysis on the sediment samples. Sincere grati-tude also to Dr. Helen Lambkin of the Cell Pathologylaboratory, Dublin Institute of Technology for the pro-vision of paraffin embedded sections. This research wasfunded by The Higher Education Authority under theP y-c tute,U
R
A 004.there.
A orterviron-227.
A ationAn-sting
B rsetein.
nvironment may be of ecotoxicological concern aibition of P-gps by environmental pollutants mayult in an organism’s increased susceptibility to ceroxicants as the functioning of an MXR mechanismormal tissues is a general defence to protect cells
he intracellular accumulation of natural, endogennd environmental toxins. However, further resea
s needed to elucidate the significance of P-gp mted transport as a protective mechanism againstonmental toxicants in fish before the ecotoxicologonsequences of such inhibition can be addresse
In terms of the potential use of mdr1-like activn primary epidermal cultures as an ecotoxicologcreening tool, the results of this study found thatxpression of P-gp-like proteins are in line with theults of the chemical analyses, as the greatest induf these proteins was associated with the most con
nated site. The findings of the functional study a
rogram of Research in Third Level Institutions (Cle 2) as part of the Environmental Research Instiniversity College Cork.
eferences
chard, M., Baudrimont, M., Boudou, A., Bourdineaud, J.P., 2Induction of a multixenobiotic resistance protein (MXR) inAsiatic clamCorbicula flumineaafter heavy metals exposuAquat. Toxicol. 67, 347–357.
lbertus, J.A., Laine, R.O., 2001. Enhanced xenobiotic transpexpression in normal teleost hepatocytes: response to enmental and chemotherapeutic toxins. J. Exp. Biol. 204, 217–
STM, 1994. Standard guide for collection storage, characterisand manipulation of sediment for toxicological testing. In:nual Book of Standards, E1391-94. American Society for Teand Materials, Philadelphia, USA.
ain, L.J., LeBlanc, G.A., 1996. Interaction of structurally divepesticides with the human MDR1 gene product P-glycoproToxicol. Appl. Pharmacol. 141, 288–298.

126 S. Nı Shuilleabhain et al. / Aquatic Toxicology 73 (2005) 115–127
Bard, S.M., Bello, S.M., Hahn, M.E., Stegeman, J.J., 2002. Ex-pression of P-glycoprotein in killifish (Fundulus heterocli-tus) exposed to environmental xenobiotics. Aquat. Toxicol. 59,237–251.
Bard, S.M., 2000. Multixenobiotic resistance as a cellular defencemechanism in aquatic organisms. Aquat. Toxicol. 48, 357–389.
Bard, S.M., Woodin, B., Stegeman, J.J., 1998. Induction of the mul-tixenobiotic resistance transporter and cytochrome P450 1A inintertidal fish exposed to environmental contaminants. Toxicol.Sci. 42, 16.
Bullock, A.M., Marks, R., Roberts, R.J., 1978. The cell kinetics ofteleost fish epidermis: mitotic activity of the normal epidermis atvarying temperatures in plaice (Pleuronectes platessa). J. Zool.(Lond.) 194, 423–428.
Byrne, P.A., O’ Halloran, J., 2000. Acute and sublethal toxicity ofestuarine sediments to the manila clamTapes semidecussatus.Environ. Toxicol. 15, 456–468.
Chan, K.M., Davies, P.L., Childs, S., Veinot, L., Ling, V., 1992.P-glycoprotein genes in the winter flounderPleuronectes amer-icanus: isolation of two types of genomic clones carrying 3′ ter-minal exons. Biochim. Biophys. Acta 1171, 65–72.
Chieli, E., Santoni-Rugui, E., Cervelli, F., Sabbatini, A., Petrine,M., Romiti, N., Paolicchi, A., Rongiani, R., 1994. Differentialmodulation of P-glycoprotein expression by dexamethasone and3-methylcholanthrene in rat hepatocyte primary cultures. Car-cinogenesis 15, 335–341.
Chin, H.V., Tanaka, S., Darlington, G.B., Pastan, I., Gottesman,M.M., Roninson, I.B., 1990. Heat shock and arsenite increaseexpression of the multidrug resistance (MDR1) gene in humanrenal carcinoma cells. J. Biol. Chem. 265, 221–226.
Cooper, P.S., Vogelbein, W.K., Van Veld, P.A., 1999. Altered expres-sion of the xenobiotic transporter P-glycoprotein in liver and livertumours of mummichog (Fundulus heteroclitus) from a creosote-contaminated environment. Biomarkers 4 (1), 48–58.
Cornwall, R., Holland Toomey, B., Bard, S., Bacon, C., Jar-lti-
tes.
D ul-: El-ro-t Re-Vir-32.
D alicity. 18
D in-l in9.
E le ofe and
F mul-
Freshney, I., 1994. Culture of Animal Cells: A Manual of BasicTechniques, 3rd ed. Alan R. Liss, Inc., New York.
Germann, U.A., 1996. P-glycoprotein—a mediator of multidrug re-sistance in tumour cells. Eur. J. Cancer A 32, 927–944.
Harrell, L.W., Etlinger, H.M., Hodgins, H.O., 1976. Humoral factorsimportant in resistance of salmonid fish to bacterial disease. II.Anti-Vibrio anguillarum activity in mucus and observations oncomplement. Aquaculture 7, 363–370.
Hemmer, M.J., Courtney, L.A., Benson, W.H., 1998. Comparisonof three histological fixatives on the immunoreactivity of mam-malian P-glycoprotein antibodies in the sheepshead minnowCyprinodon variegates. J. Exp. Zool. 281, 251–259.
Hemmer, M.J., Courtney, L.A., Ortego, L.S., 1995. Immunohis-tochemical detection of P-glycoprotein in teleost tissues usingmammalian polyclonal and monoclonal antibodies. J. Exp. Zool.272, 69–77.
Hjelmeland, K., Christie, M., Raa, J., 1983. Skin mucus proteasefrom rainbow troutSalmogardineriRichardson and its biologicalsignificance. J. Fish. Biol. 23, 13–22.
Ingersoll, C.G., 1995. Sediment tests in fundamentals of aquatic tox-icology. In: Rand, G.M. (Ed.), Effects, Environmental Fate, andRisk Assessment. Taylor and Francis, New York, pp. 231–255.
Juliano, R.L., Ling, V., 1976. A surface glycoprotein modulating drugpermeability in Chinese hamster ovary cell mutants. Biochim.Biophys. Acta 455, 152–162.
Keely, J.E., Engler, R.M., 1974. Dredged material research program.Miscellaneous Paper D-74-14, US, Army Engineer WaterwaysExperiment Station, Vicksburg, Mississippi.
Kilemade, M., Hartl, M.G.J., Sheehan, D., Mothersill, C., van Pelt,F.N.A.M., O’Brien, N.M., O’Halloran, J., 2004. An assessmentof the pollutant status of surficial sediment in Cork Harbour inthe South East of Ireland with particular reference to polycyclicaromatic hydrocarbons. Mar. Pollut. Bull. 49, 1084–1096.
Kilemade, M., Lyons-Alcantara, M., Rose, T., Fitzgerald, R., Mother-sill, C., 2002. Rainbow trout primary epidermal cell proliferation
ure
K vels2,4-nvi-
K thepi-
099.K
tfishon inar.
K ofs the419.
K ch-Total
K m in
man, W.M., Epel, D., 1995. Characterisation of muxenobiotic/multidrug transport in the gills of the musselMytiluscalifornianus and identification of environmental substraAquat. Toxicol. 31, 277–296.
oi, A.M., Kleinow, K.M., Venugopal, C.S., Holmes, E., 2000. Mtixenobiotic resistance mechanism in the catfish intestine. Inskus, A.A., Vogelbein, W.K., McLaughlin, S.M., Kane, A.S., Pceedings of the 10th International Symposium on Pollutansponses in Marine Organisms (PRIMO 10). Williamsburg,ginia, USA. 25–29 April 1999. Mar. Environ. Res. 50 (1–5) 3
owling, K., Mothersill, C., 1999. Use of rainbow trout epidermcell cultures as an alternative to immortalised cell lines in toxassessment: a study with nonoxynol. Environ. Toxicol. Chem(12), 2846–2850.
owling, K., Mothersill, C., 2001. The further development of rabow trout primary epithelial cell cultures as a diagnostic tooecotoxicological assessment. Aquat. Toxicol. 19, 2093–209
ytan, C.D., Regev, R., Oren, G., Assaraf, Y.G., 1996. The ropassive transbilayer drug movement in multidrug resistancits modulation. J. Biol. Chem. 271, 12897–12902.
ord, J.M., Hait, W.N., 1990. Pharmacology of drugs that altertidrug resistance in cancer. Pharmacol. Rev. 8, 137–171.
as an indicator of aquatic toxicity: an in vivo/in vitro exposcomparison. Aquat. Toxicol. 60, 43–59.
ilemade, M., Mothersill, C., 2001. Heat shock protein 70 lein rainbow trout primary epidermal cultures in response todichloroaniline exposure: a novel in vitro aquatic marker. Eron. Toxicol. 16, 253–259.
ilemade, M., Mothersill, C., 2000. An in vitro assessment oftoxicity of 2,4-dichloroanaline using rainbow trout primary edermal cell cultures. Environ. Toxicol. Chem. 19 (8), 2093–2
ohler, A., Lauritzen, B., Bahns, S., George, S.G., Forlin, L., VanNo-orden, C.J.F., 1998. Clonal adaptation of cancer cells in flaliver to environmental contamination by changes in expressiP-gp related MXR, CYP450 GST-A and G6PDH activity. MEnviron. Res. 46, 191–195.
urelec, B., Britvic, S., Pivcecic, B., Smital, T., 1998. Fragilitymultixenobiotic resistance in aquatic organisms enhancecomplexity of risk assessment. Mar. Environ. Res. 46, 415–
urelec, B., 1995. Inhibition of multixenobiotic resistance meanism in aquatic organisms: ecotoxic consequences. Sci.Environ. 171, 197–204.
urelec, B., 1992. The multixenobiotic resistance mechanisaquatic organisms. Crit. Rev. Toxicol. 22 (1), 23–43.

S. Nı Shuilleabhain et al. / Aquatic Toxicology 73 (2005) 115–127 127
Loring, D.H., Rantala, R.T.T., 1992. Manual for the geochemicalanalyses of marine sediments and suspended particulate matter.Earth Sci. Rev. 32, 235–283.
Luedeking, A., Koehler, A., 2004. Regulation of expression of mul-tixenobiotic resistance (MXR) genes by environmental factors inthe blue musselMytilus edulis. Aquat. Toxicol. 68, 1–10.
Lyng, F.M., Lyons-Alcantara, M., Olwell, P., Nı Shuilleabhain, S.,Seymour, C., Cottell, D., Mothersill, C., 2004. Ionizing radiationinduces a stress response in primary cultures of rainbow troutskin. Radic. Res. 162, 226–232.
Lyons-Alcantara, M., Mooney, R., Lyng, F., Cottell, D., Mothersill,C., 1998. The effects of cadmium exposure on the cytology andfunction of primary cultures from rainbow trout. Cell Biochem.Funct. 16, 1–13.
Lyons-Alcantara, M., Tarazona, J.V., Mothersill, C., 1996. The differ-ential effects of cadmium exposure on the growth and survival ofprimary and established cells from fish and mammals. Cell Biol.Toxicol. 12, 29–38.
Machii, A., Wada, K.T., 1989. Some marine invertebrates tissue cul-ture. In: Mitsuhashi, J. (Ed.), Invertebrate Cell System Applica-tions, vol. 2. CRC Press, Boca Raton, FL.
Marine Institute, 1999. Ireland’s Marine and Coastal Areas and Ad-jacent Seas: An Environmental Assessment. Department of theMarine and Natural Resources, Dublin, Ireland.
Minier, C., Eufemia, N., Epel, D., 1999. The multi-xenobiotic re-sistance phenotype as a tool to biomonitor the environment.Biomarkers 4 (6), 442–454.
Minier, C., Moore, M.N., 1996. Induction of multixenobiotic re-sistance in mussel blood cells. Mar. Environ. Res. 42 (1–4),389–392.
Mothersill, C., Lyng, F., Lyons, M., Cottell, D., 1995. Growth anddifferentiation of epidermal cells from the rainbow trout estab-lished as explants and maintained in various media. J. Fish Biol.46, 1011–1025.
Nichols, J.W., McKim, J.M., Lien, G.J., Hoffman, A.D., Bertelsen,eticam.
S nder-M.,.M.,
epi-nce.
S P-orin
analogue by teleost renal proximal tubules. Am. J. Physiol. (Re-nal Fluid Electrolyte Physiology 37) 268, F46–F52.
Sharom, F.J., 1997. The P-glycoprotein efflux pump: how does ittransport drugs? J. Membrane Biol. 160, 161–175.
Singer, T.D., Tucker, S.J., Marshall, W.S., Higgins, C.F., 1999. A di-vergent CFTR homologue: highly regulated salt transport in theeuryhaline teleost Rheteroclitus. Am. J. Physiol. (Cell Physiol-ogy 43) 274, C715–C723.
Smital, T., Sauerborn, R., Hackenberger, B.K., 2003. Inducibility ofthe P-glycoprotein transport activity in the marine musselMytilusgalloprovincialisand the freshwater musselDressiena polymor-pha. Aquat. Toxicol. 65, 443–465.
Smital, T., Sauerborn, R., 2002. Measurement of the activity of mul-tixenobiotic resistance mechanism in the common carpCyprinuscarpio. Mar. Environ. Res. 54, 449–453.
Smital, T., Kurelec, B., 1998. The chemosensitizers of multixenobi-otic resistance mechanism in aquatic invertebrates: a new classof pollutants. Mutat. Res. 399, 43–53.
Strum, A., Cravedi, J.P., Segner, H., 2001a. Prochloraz and nonylphe-nol diethoxylate inhibit an mdr1-like activity in vitro, but do notalter hepatic levels of P-glycoprotein in trout exposed in vivo.Aquat. Toxicol. 53, 215–228.
Strum, A., Ziemann, C., Hirsch-Ernst, K.I., Segner, H., 2001b. Ex-pression and functional activity of P-glycoprotein in culturedhepatocytes fromOncorhynchus mykiss. Am. J. Physiol. Regul.Integr. Comp. Physiol. 281, 1119–1126.
Toomey, B.H., Epel, D., 1993. Multixenobiotic resistance inUre-chis caupoembryos: protection from environmental toxins. Biol.Bull. 185, 355–364.
Tutundjian, R., Cachot, J., Leboulenger, F., Minier, C., 2002. Ge-netic and immunological characterisation of a multixenobioticresistance system in the turbot (Scophthalmus maximus). Comp.Biochem. Physiol. B 132, 463–471.
USEPA, 1977. Dredged material sample collection and preparation.Appendix B. In: Ecological evaluation of proposed discharge
ippi,nicalcts
tion.v g P-
W kens,t (P-R.S.C.V.
S.L., Elonen, C.M., 1996. A physiologically based toxicokinmodel of dermal absorption of organic chemicals in fish. FundAppl. Toxicol. 31, 229–242.
cheper, R.J., Bulte, J.W.M., Brakkee, J.G.P., Quak, J.J., Vaschoot, E., Anderschoot, E., Balm, A.J.M., Meijer, C.J.L.Broxterman, H.J., Kuiper, C.M., Lankelma, J., Pinedo, H1988. Monoclonal antibody JSB1 detects a highly conservedtope on the P-glycoprotein associated with multidrug resistaInt. J. Can. 42 (3), 389–394.
chramm, U., Fricker, G., Wenger, R., Miller, D.S., 1995.glycoprotein-mediated secretion of a fluorescent cyclosp
of dredged material into ocean waters. Vicksburg, MississEnvironmental Protection Agency/Corps of Engineers TechCommittee for Dredged and Fill Material. Environmental EffeLaboratory, U.S. Army Engineer Waterways Experiment Sta
an Tellingen, O., 2001. The importance of drug-transportinglycoproteins in toxicology. Toxicol. Lett. 120, 31–41.
einstein, R.S., Grogan, T.M., Duszak, J.R., Jakate, S.M., KlusL.F., Coon, J.S., 1991. Multidrug resistance gene producglycoprotein) in normal tissue and tumors. In: Weinstein,(Ed.), Advances in Pathology and Laboratory Medicine.Mosby Co., New York, pp. 207–234.
Copyright © 2022 FDOKUMEN




















