
Tau-like Proteins Associated with Centrosomes in Cultured Cells
10
EXPERIMENTAL CELL RESEARCH 229, 378–387 (1996) ARTICLE NO. 0383 Tau-like Proteins Associated with Centrosomes in Cultured Cells DANIEL CROSS,* , ²LUCI ´ A TAPIA,* , ²JORGE GARRIDO,² , ‡ AND RICARDO B. MACCIONI* , ² ,1 *Department of Biology, Faculty of Sciences, University of Chile, Las Palmeras 3425, N ˜ un ˜ oa, Santiago, Chile; ²International Center for Cancer and Developmental Biology (ICC) Casilla 70111, Santiago 7, Chile; and ‡Department of Cellular and Molecular Biology, Catholic University of Chile, Casilla 114-D, Santiago, Chile INTRODUCTION The subcellular association of tau-like proteins with Microtubule-associated proteins (MAPs), including centrosomes in cultured cell lines and its effects in the high-molecular-weight components and tau, copur- nucleating microtubule assembly were analyzed using ify with tubulin through cycles of assembly and disas- biochemical and immunocytochemical approaches. sembly. There is increasing evidence that these pro- Tau proteins, major components of microtubules, ap- teins stabilize assembled microtubules in the cell and pear to be tightly associated with actin filaments in a that they appear to play a major regulatory role in variety of cell lines, while in pathological conditions tubulin polymerization into microtubules [1 – 4]. In ad- of neurons, they are part of paired helical filaments dition, current evidence obtained using viscosimetry, found in Alzheimer’s disease. Different studies suggest photobleaching, cosedimentation analysis, and immu- that, in addition to tau interactions with the compo- nofluorescence suggests that the linkages of actin mi- nents of the cytoskeletal network, tau polypeptides ap- crofilaments with the microtubule system are mediated pear to be associated with highly structured cellular by microtubule-associated proteins [5 – 7]. It has also elements, in both interphase and mitotic cells. An been shown that MAPs interact with actin in vitro [5, in-depth analysis of tau subcellular distribution us- 8], while a differential role of tau isoforms in their in- ing different polyclonal and monoclonal antibodies teractions with the distinct cytoskeletal filaments has showed colocalization of tau-like components with centrosomes in interphase cells of the human Huh-7 been recently demonstrated [9]. Despite the experi- hepatoma, in SW-13 adenocarcinoma, and in normal mental data that point to the roles of MAPs in linking human fibroblasts. Tau associated with centrosomes filamentous components within the cytoskeletal struc- in mitotic Huh-7 cells was also identified. However, ture, the detailed molecular aspects of in vivo functions antibodies against the tau binding repeats did not of MAPs, and specifically tau, are not well understood. stain centrosomes. A set of different tau isoforms was Tau has been widely described as an axonal protein also identified by Western blot analysis on isolated distributed mainly in the cytoplasm and associated centrosomal preparations from Huh-7 cells, obtained with microtubules [10, 11] that contributes to maintain by differential centrifugation through sucrose gradi- the neuronal architecture [4, 12 – 14]. However, current ents. Microtubule nucleation in vitro over isolated information points to the presence of tau-like proteins 2 centrosomes was inhibited by both the polyclonal anti- in a variety of cell lines, thus suggesting a widespread body against native tau and an antibody to the N-ter- distribution of tau components in cells [7]. Further- minal tau sequence, as revealed by immunofluores- more, tau components have been partially purified cence analysis and assembly kinetics experiments. The from nonneuronal cell sources [7, 9]. The tau-like pro- antibody TRS1.2 against the fragment containing the teins appear to colocalize with stress fibers in a variety first binding repeat on tau did not affect nucleation. of cultured cell lines [7, 14] and have been also identi- These studies allowed us to characterize tau associa- fied in close association with the microtubule cytoskele- tion with the isolated centrosomal preparation and its ton [15]. Most recently, cell microinjection experiments involvement in microtubule assembly nucleated over of labeled tau allowed us to visualize a distribution of centrosomes, thus suggesting possible structural and these proteins between the nucleus and the cytoplasm functional roles for these interactions. q 1996 Academic and suggested their colocalization with centrosomes in Press, Inc. specific cell types [16]. Studies also detected tau in the 2 Nonstandard terminology: Tau-like proteins is used to refer to a set of polypeptides from nonneuronal sources that share common 1 To whom correspondence and reprint requests should be ad- features with brain tau, such as their thermal resistance, perchloric acid solubility, and common epitopes. dressed. Fax: (562) 218-6245. 378 0014-4827/96 $18.00 Copyright q 1996 by Academic Press, Inc. All rights of reproduction in any form reserved.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Tau-like Proteins Associated with Centrosomes in Cultured Cells

EXPERIMENTAL CELL RESEARCH 229, 378–387 (1996)ARTICLE NO. 0383
Tau-like Proteins Associated with Centrosomes in Cultured Cells
DANIEL CROSS,*,† LUCIA TAPIA,*,† JORGE GARRIDO,†,‡ AND RICARDO B. MACCIONI*,†,1
*Department of Biology, Faculty of Sciences, University of Chile, Las Palmeras 3425, Nunoa, Santiago, Chile; †International Center forCancer and Developmental Biology (ICC) Casilla 70111, Santiago 7, Chile; and ‡Department of Cellular and Molecular Biology, Catholic
University of Chile, Casilla 114-D, Santiago, Chile
INTRODUCTION
The subcellular association of tau-like proteins withMicrotubule-associated proteins (MAPs), includingcentrosomes in cultured cell lines and its effects in
the high-molecular-weight components and tau, copur-nucleating microtubule assembly were analyzed usingify with tubulin through cycles of assembly and disas-biochemical and immunocytochemical approaches.sembly. There is increasing evidence that these pro-Tau proteins, major components of microtubules, ap-teins stabilize assembled microtubules in the cell andpear to be tightly associated with actin filaments in athat they appear to play a major regulatory role invariety of cell lines, while in pathological conditionstubulin polymerization into microtubules [1–4]. In ad-of neurons, they are part of paired helical filamentsdition, current evidence obtained using viscosimetry,found in Alzheimer’s disease. Different studies suggestphotobleaching, cosedimentation analysis, and immu-that, in addition to tau interactions with the compo-nofluorescence suggests that the linkages of actin mi-nents of the cytoskeletal network, tau polypeptides ap-crofilaments with the microtubule system are mediatedpear to be associated with highly structured cellularby microtubule-associated proteins [5–7]. It has alsoelements, in both interphase and mitotic cells. Anbeen shown that MAPs interact with actin in vitro [5,in-depth analysis of tau subcellular distribution us-8], while a differential role of tau isoforms in their in-ing different polyclonal and monoclonal antibodiesteractions with the distinct cytoskeletal filaments hasshowed colocalization of tau-like components with
centrosomes in interphase cells of the human Huh-7 been recently demonstrated [9]. Despite the experi-hepatoma, in SW-13 adenocarcinoma, and in normal mental data that point to the roles of MAPs in linkinghuman fibroblasts. Tau associated with centrosomes filamentous components within the cytoskeletal struc-in mitotic Huh-7 cells was also identified. However, ture, the detailed molecular aspects of in vivo functionsantibodies against the tau binding repeats did not of MAPs, and specifically tau, are not well understood.stain centrosomes. A set of different tau isoforms was Tau has been widely described as an axonal proteinalso identified by Western blot analysis on isolated distributed mainly in the cytoplasm and associatedcentrosomal preparations from Huh-7 cells, obtained with microtubules [10, 11] that contributes to maintainby differential centrifugation through sucrose gradi- the neuronal architecture [4, 12–14]. However, currentents. Microtubule nucleation in vitro over isolated information points to the presence of tau-like proteins2
centrosomes was inhibited by both the polyclonal anti- in a variety of cell lines, thus suggesting a widespreadbody against native tau and an antibody to the N-ter- distribution of tau components in cells [7]. Further-minal tau sequence, as revealed by immunofluores- more, tau components have been partially purifiedcence analysis and assembly kinetics experiments. The from nonneuronal cell sources [7, 9]. The tau-like pro-antibody TRS1.2 against the fragment containing the teins appear to colocalize with stress fibers in a varietyfirst binding repeat on tau did not affect nucleation. of cultured cell lines [7, 14] and have been also identi-These studies allowed us to characterize tau associa- fied in close association with the microtubule cytoskele-tion with the isolated centrosomal preparation and its ton [15]. Most recently, cell microinjection experimentsinvolvement in microtubule assembly nucleated over of labeled tau allowed us to visualize a distribution ofcentrosomes, thus suggesting possible structural and these proteins between the nucleus and the cytoplasmfunctional roles for these interactions. q 1996 Academic and suggested their colocalization with centrosomes inPress, Inc. specific cell types [16]. Studies also detected tau in the
2 Nonstandard terminology: Tau-like proteins is used to refer to aset of polypeptides from nonneuronal sources that share common
1 To whom correspondence and reprint requests should be ad- features with brain tau, such as their thermal resistance, perchloricacid solubility, and common epitopes.dressed. Fax: (562) 218-6245.
3780014-4827/96 $18.00Copyright q 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.
AID ECR 3360 / 6i18$$$181 12-02-96 01:52:56 eca

379TAU-LIKE PROTEINS ASSOCIATED WITH CENTROSOMES
microtubule nucleation are still unknown, and cer-tainly their identification is a critical step to analyzefunctional aspects of centrosomes at the molecularlevel. Recent studies have provided clues on the rolesof CP-60 and DMAP-190 in centrosomes of Drosophilaembryos [23, 24]. On the other hand, it has been re-ported that a protein related to microtubule-associatedprotein MAP-1B is present in centrosomes of mamma-lian cells [25], although no evidence on its activity androles has been provided. The present study is based onthe finding of tau-like components in centrosomes ofa variety of cultured nonneuronal cell lines and theirpossible mechanisms of action at the level of this cellu-lar structure.
Within the context of an analysis of the subcellularorganization of tau isospecies in various cell types, we
FIG. 1. Double immunofluorescence study indicating the colocal-ization of tau with tubulin in centrosomes of the human adenocarci-noma cell line vimentin positive (vim/) SW-13 cells using an anti-b-tubulin monoclonal antibody (A) and a polyclonal antibody againstnative tau (B). The protein antigen reactive with the anti-tubulinantibody was visualized using a rhodamine-conjugated second anti-body, while FITC-conjugated second antibodies were used to revealanti-tau staining. Centrosomes (arrow 1) appear clearly stained withthe anti-tau polyclonal antibody shown in B. Arrows 2 show cyto-plasmic microtubules in which tau colocalization is observed. Bars,10 mm.
cell nucleus, evidencing a discrete staining of con-densed chromosomes by indirect immunofluorescenceanalysis [17, 18]. In addition, another set of evidenceindicates that MAP-2 and tau promote DNA replicationand that tau polypeptides present a high and conservedaffinity for some specific sequences on the DNA struc-ture [19–21]. All these studies lead us to describe tauas a multifunctional set of proteins of major cellular
FIG. 2. Immunofluorescence pattern showing the localization ofimportance.immunoreactive tubulin and tau components with centrosomes inIt has been widely established that centrosomes nu-primary cultures of human fibroblasts. The cells were extracted,cleate microtubules and play an essential role in the fixed, and incubated with an anti-b-tubulin monoclonal antibody (A)
organization of dynamic microtubule arrays in all ani- and a polyclonal serum raised against the native tau protein (B).Bar, 5 mm.mal cells [22, 23]. The proteins that are involved in
AID ECR 3360 / 6i18$$$182 12-02-96 01:52:56 eca

380 CROSS ET AL.
FIG. 3. Single immunofluoresce patterns of several interphase Huh-7 human hepatoma transformed cells showing the centrosomesimmunostained with either the monoclonal anti-b-tubulin antibody (A) or the polyclonal antibody against native tau (B). Arrows indicatethe centrosomal localization. Bar, 10 mm.
some cases, and incubated with the primary antibodies: polyclonalused polyclonal antibodies to brain tau and site-di-anti-tau, monoclonal anti-b-tubulin, or the site-specific antibodies torected probes bearing specific sequences of tau iso-tau at dilutions between 1:50 and 1:100 for at least 2 h. Secondforms. The studies clearly showed the in vivo distribu- antibodies, anti-rabbit IgG’s used to reveal protein recognized by the
tion of tau-like proteins within the centrosomal struc- primary anti-tubulin antibody or to reveal tau, were used at a 1:500dilution and incubated for the same period. For visualization of im-ture. Colocalization of immunoreactive tau withmunostained material, coverslips were further incubated with FITCtubulin in the centrosomes of both mitotic and in-or rhodamine-conjugated second antibodies for 1 h. Cytoskeletalterphase cells was also observed. It was noteworthystructures and tau distribution were visualized under a Zeiss Axiov-
that antibodies against native tau and site-specific an- ert microscope, equipped with an epifluorescence attachment.tibodies against a peptide bearing the N-terminal tau Antibody production. The following antibodies were used as
probes for studies in this research: (a) Polyclonal antibodies againstsequence inhibited microtubule growth nucleated overthe synthetic peptide resembling the sequences of N-terminal moietycentrosomes.were generously provided by Dr. Jacques Nunez from Inserm,France. (b) Antibodies against native brain tau were obtained by
MATERIALS AND METHODS immunization of rabbits with pure brain tau obtained after cosedi-mentation with microtubules [26], followed by electrophoresis and
Materials. The commercial monoclonal anti-b-tubulin antibody, selection of tau-containing slices from polyacrylamide gels. Whiteclone TUB 2.2, was obtained from Sigma (St. Louis, MO). The anti- New Zealand rabbits were immunized subcutaneously every 20 daysbody to N-terminal tau moiety was supplied by Dr. J. Nunez (Inserm, three times, with 100 mg tau extracted from the acrylamide slices.France). The polyclonal antibody to native tau, the site-specific probe First injection was in complete Freund’s adjuvant followed by twoTRS1.2 against the first tau repeat [26], and the monoclonal anti- more injections in incomplete adjuvant. Two weeks before bleeding,idiotypic antibody MTB6.22 [27] were generated in our laboratory. rabbits were boosted with a dose of pure tau in incomplete adjuvant.Nocodazol, immunological reagents, and secondary antibodies were The antibody titer was determined by ELISA assay. (c) Site-specificpurchased from Sigma. Taxol was provided by the Developmental antibodies to tau, including the polyclonal antibody TRS1.2 obtainedTherapeutic Program, Division of Cancer Treatment, NCI. All other against the first tau repeat and the monoclonal anti-idiotypic anti-reagents were of the highest analytical quality. body MTB6.22, were also used in this study. All the antibodies gener-
Cell culture. Human fibroblasts, hepatoma Huh-7 transformed ated throughout the research program have been widely character-cells, and SW-13 adenocarcinoma cells were cultured in Dulbecco’s ized, as indicated in the respective reports [26–29]. The polyclonalmodified Eagle’s medium supplemented with 10% fetal calf serum antibodies were subjected to immunoabsorption before use, with tauin a 5% CO2 atmosphere. Confluent or subconfluent 100-mm culture in the case of the antibody to native tau or with the peptide of taudishes of the different cell lines were used in these preparations. repeat V187 0 G204 in the case of TRS1.2 antibody.
Differential centrifugation. The cell centrosomes were isolated byImmunofluorescence. Indirect immunofluorescence approacheswere used as previously described [7, 28]. Cells were seeded over differential centrifugation through a 45–70% sucrose gradient fol-
lowing the protocol of Mitchison and Kirschner [30] and using thecoverslips for at least 24 h before double immunostaining. Materialon coverslips was extracted in prewarmed MSB buffer plus 0.5% sequence of steps indicated by Bornen et al. [31] with minor modifi-
cations. The supernatants of Huh-7 hepatoma cells containingTriton X-100 for 40 s. The extracted cells were fixed by immersionin 4% paraformaldehyde for 10 min, or in cold methanol (0207C) in centrosomal structures were overlaid on top of the sucrose gradient,
AID ECR 3360 / 6i18$$$182 12-02-96 01:52:56 eca
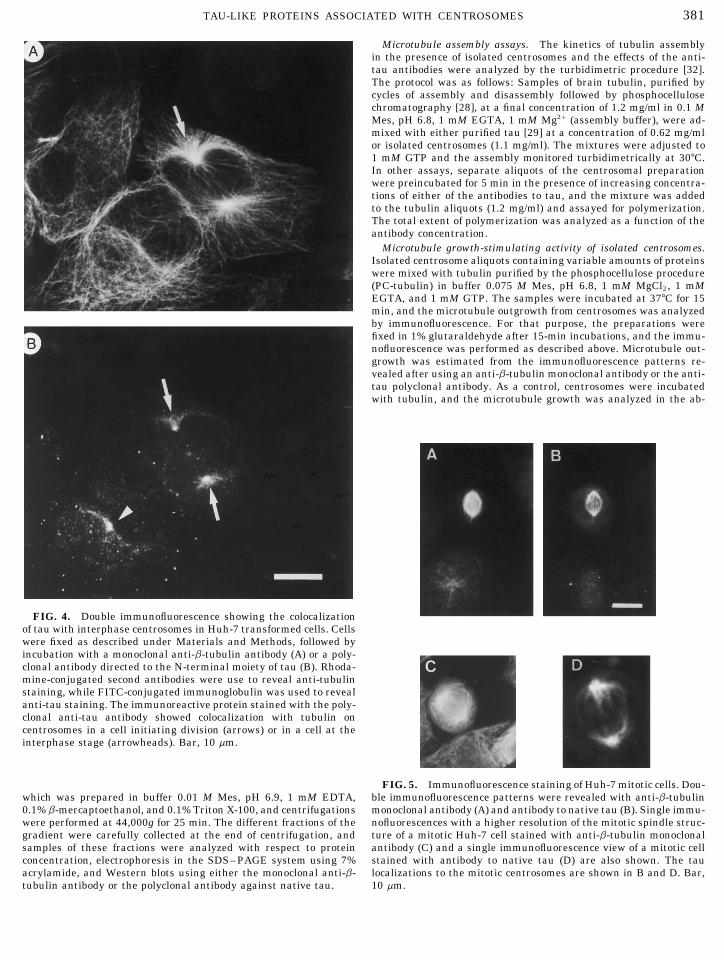
381TAU-LIKE PROTEINS ASSOCIATED WITH CENTROSOMES
Microtubule assembly assays. The kinetics of tubulin assemblyin the presence of isolated centrosomes and the effects of the anti-tau antibodies were analyzed by the turbidimetric procedure [32].The protocol was as follows: Samples of brain tubulin, purified bycycles of assembly and disassembly followed by phosphocellulosechromatography [28], at a final concentration of 1.2 mg/ml in 0.1 MMes, pH 6.8, 1 mM EGTA, 1 mM Mg2/ (assembly buffer), were ad-mixed with either purified tau [29] at a concentration of 0.62 mg/mlor isolated centrosomes (1.1 mg/ml). The mixtures were adjusted to1 mM GTP and the assembly monitored turbidimetrically at 307C.In other assays, separate aliquots of the centrosomal preparationwere preincubated for 5 min in the presence of increasing concentra-tions of either of the antibodies to tau, and the mixture was addedto the tubulin aliquots (1.2 mg/ml) and assayed for polymerization.The total extent of polymerization was analyzed as a function of theantibody concentration.
Microtubule growth-stimulating activity of isolated centrosomes.Isolated centrosome aliquots containing variable amounts of proteinswere mixed with tubulin purified by the phosphocellulose procedure(PC-tubulin) in buffer 0.075 M Mes, pH 6.8, 1 mM MgCl2, 1 mMEGTA, and 1 mM GTP. The samples were incubated at 377C for 15min, and the microtubule outgrowth from centrosomes was analyzedby immunofluorescence. For that purpose, the preparations werefixed in 1% glutaraldehyde after 15-min incubations, and the immu-nofluorescence was performed as described above. Microtubule out-growth was estimated from the immunofluorescence patterns re-vealed after using an anti-b-tubulin monoclonal antibody or the anti-tau polyclonal antibody. As a control, centrosomes were incubatedwith tubulin, and the microtubule growth was analyzed in the ab-
FIG. 4. Double immunofluorescence showing the colocalizationof tau with interphase centrosomes in Huh-7 transformed cells. Cellswere fixed as described under Materials and Methods, followed byincubation with a monoclonal anti-b-tubulin antibody (A) or a poly-clonal antibody directed to the N-terminal moiety of tau (B). Rhoda-mine-conjugated second antibodies were use to reveal anti-tubulinstaining, while FITC-conjugated immunoglobulin was used to revealanti-tau staining. The immunoreactive protein stained with the poly-clonal anti-tau antibody showed colocalization with tubulin oncentrosomes in a cell initiating division (arrows) or in a cell at theinterphase stage (arrowheads). Bar, 10 mm.
FIG. 5. Immunofluorescence staining of Huh-7 mitotic cells. Dou-ble immunofluorescence patterns were revealed with anti-b-tubulinwhich was prepared in buffer 0.01 M Mes, pH 6.9, 1 mM EDTA,
0.1% b-mercaptoethanol, and 0.1% Triton X-100, and centrifugations monoclonal antibody (A) and antibody to native tau (B). Single immu-nofluorescences with a higher resolution of the mitotic spindle struc-were performed at 44,000g for 25 min. The different fractions of the
gradient were carefully collected at the end of centrifugation, and ture of a mitotic Huh-7 cell stained with anti-b-tubulin monoclonalantibody (C) and a single immunofluorescence view of a mitotic cellsamples of these fractions were analyzed with respect to protein
concentration, electrophoresis in the SDS–PAGE system using 7% stained with antibody to native tau (D) are also shown. The taulocalizations to the mitotic centrosomes are shown in B and D. Bar,acrylamide, and Western blots using either the monoclonal anti-b-
tubulin antibody or the polyclonal antibody against native tau. 10 mm.
AID ECR 3360 / 6i18$$$182 12-02-96 01:52:56 eca

382 CROSS ET AL.
FIG. 6. (A) Isolation of centrosomes from Huh-7 cells, essentially on the basis of the procedure described by Mitchison and Kirschner[30], considering the sequence of steps by Bornen et al. [31]. Centrosomes were separated by high-speed centrifugation using a 45–70%sucrose gradient, and the protein concentration is represented by the histograms for each fraction of the gradient. (B) The electrophoreticprofiles for the gradient fractions are shown after staining with Coomassie blue. Molecular weight standards are represented at the left(ST): myosin (205 kDa), b-galactosidase (116 kDa), phosphorylase b (97 kDa), serum albumin (66 kDa), and ovalbumin (45 kDa). (C) Samplesof 20 mg from gradient fractions were also assayed by immunoblots. Centrosomal tau-like protein isospecies were identified by the Westernblot analysis using the antibody to native tau, mainly in fractions 2 through 5. Only samples 2 through 6, corresponding to those wherecentrosomes sedimented, are shown. A Western blot of a control sample of bovine brain tau is shown. In the same set of fractions, tubulinwas also identified using an anti-b-tubulin monoclonal antibody (right). The immunoblot of brain tubulin is also shown. Other details areindicated under Materials and Methods.
sence of added GTP. In another set of experiments, centrosomal sam- staining. For immunoblots, proteins were transferred from the poly-ples were preincubated for 5 min with 20–50 mg of one of the different acrylamide gel to nitrocellulose filters (0.2 mm) in a semi-dry transferantibodies to tau, the tubulin was added, and the microtubule out- chamber at 5.5 mA/cm2 per gel for 35 min in 50 mM Tris, 0.38 Mgrowth was analyzed by immunofluorescence. glycine, 0.1% SDS, and 20% methanol [33]. Transferred proteins
were visualized by Ponceau’s staining, and subsequently nitrocellu-Protein electrophoresis and immunoblots. Tau-like proteins andlose filters were blocked with 0.5% Tween 20 in phosphate-bufferedcytoskeletal proteins were electrophoresed on 7.5% SDS–polyacryl-
amide gels. The protein on gels was visualized by Coomassie blue saline for at least 30 min at room temperature. Nitrocellulose filters
AID ECR 3360 / 6i18$$$182 12-02-96 01:52:56 eca
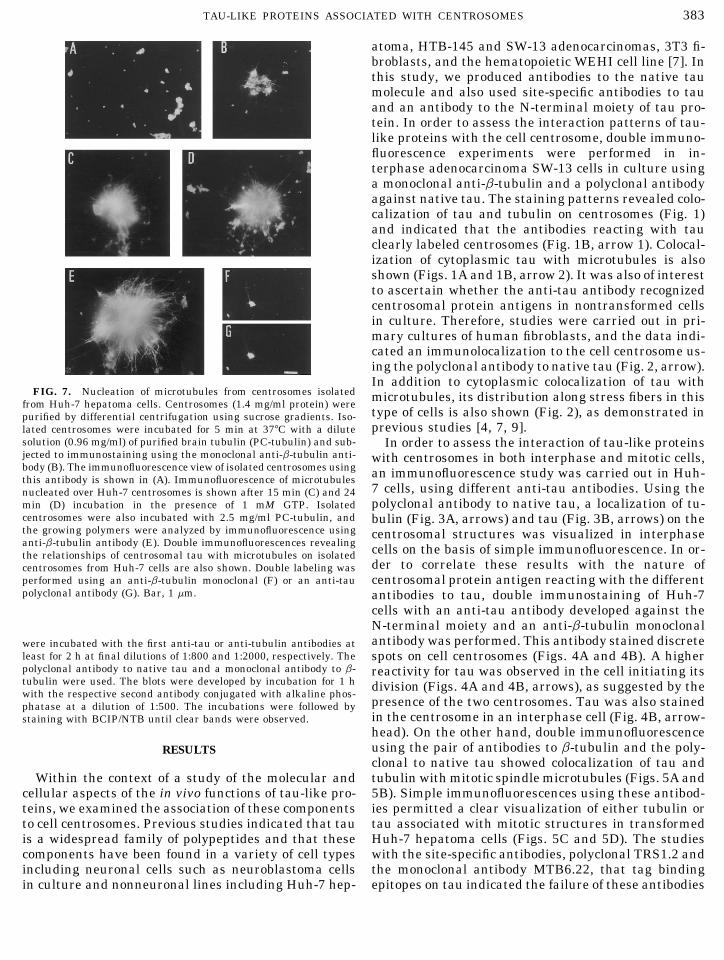
383TAU-LIKE PROTEINS ASSOCIATED WITH CENTROSOMES
atoma, HTB-145 and SW-13 adenocarcinomas, 3T3 fi-broblasts, and the hematopoietic WEHI cell line [7]. Inthis study, we produced antibodies to the native taumolecule and also used site-specific antibodies to tauand an antibody to the N-terminal moiety of tau pro-tein. In order to assess the interaction patterns of tau-like proteins with the cell centrosome, double immuno-fluorescence experiments were performed in in-terphase adenocarcinoma SW-13 cells in culture usinga monoclonal anti-b-tubulin and a polyclonal antibodyagainst native tau. The staining patterns revealed colo-calization of tau and tubulin on centrosomes (Fig. 1)and indicated that the antibodies reacting with tauclearly labeled centrosomes (Fig. 1B, arrow 1). Colocal-ization of cytoplasmic tau with microtubules is alsoshown (Figs. 1A and 1B, arrow 2). It was also of interestto ascertain whether the anti-tau antibody recognizedcentrosomal protein antigens in nontransformed cellsin culture. Therefore, studies were carried out in pri-mary cultures of human fibroblasts, and the data indi-cated an immunolocalization to the cell centrosome us-ing the polyclonal antibody to native tau (Fig. 2, arrow).In addition to cytoplasmic colocalization of tau with
FIG. 7. Nucleation of microtubules from centrosomes isolated microtubules, its distribution along stress fibers in thisfrom Huh-7 hepatoma cells. Centrosomes (1.4 mg/ml protein) were
type of cells is also shown (Fig. 2), as demonstrated inpurified by differential centrifugation using sucrose gradients. Iso-previous studies [4, 7, 9].lated centrosomes were incubated for 5 min at 377C with a dilute
solution (0.96 mg/ml) of purified brain tubulin (PC-tubulin) and sub- In order to assess the interaction of tau-like proteinsjected to immunostaining using the monoclonal anti-b-tubulin anti- with centrosomes in both interphase and mitotic cells,body (B). The immunofluorescence view of isolated centrosomes using an immunofluorescence study was carried out in Huh-this antibody is shown in (A). Immunofluorescence of microtubules
7 cells, using different anti-tau antibodies. Using thenucleated over Huh-7 centrosomes is shown after 15 min (C) and 24polyclonal antibody to native tau, a localization of tu-min (D) incubation in the presence of 1 mM GTP. Isolated
centrosomes were also incubated with 2.5 mg/ml PC-tubulin, and bulin (Fig. 3A, arrows) and tau (Fig. 3B, arrows) on thethe growing polymers were analyzed by immunofluorescence using centrosomal structures was visualized in interphaseanti-b-tubulin antibody (E). Double immunofluorescences revealing cells on the basis of simple immunofluorescence. In or-the relationships of centrosomal tau with microtubules on isolated
der to correlate these results with the nature ofcentrosomes from Huh-7 cells are also shown. Double labeling wasperformed using an anti-b-tubulin monoclonal (F) or an anti-tau centrosomal protein antigen reacting with the differentpolyclonal antibody (G). Bar, 1 mm. antibodies to tau, double immunostaining of Huh-7
cells with an anti-tau antibody developed against theN-terminal moiety and an anti-b-tubulin monoclonalantibody was performed. This antibody stained discretewere incubated with the first anti-tau or anti-tubulin antibodies at
least for 2 h at final dilutions of 1:800 and 1:2000, respectively. The spots on cell centrosomes (Figs. 4A and 4B). A higherpolyclonal antibody to native tau and a monoclonal antibody to b- reactivity for tau was observed in the cell initiating itstubulin were used. The blots were developed by incubation for 1 h division (Figs. 4A and 4B, arrows), as suggested by thewith the respective second antibody conjugated with alkaline phos-
presence of the two centrosomes. Tau was also stainedphatase at a dilution of 1:500. The incubations were followed byin the centrosome in an interphase cell (Fig. 4B, arrow-staining with BCIP/NTB until clear bands were observed.head). On the other hand, double immunofluorescenceusing the pair of antibodies to b-tubulin and the poly-RESULTSclonal to native tau showed colocalization of tau andtubulin with mitotic spindle microtubules (Figs. 5A andWithin the context of a study of the molecular and
cellular aspects of the in vivo functions of tau-like pro- 5B). Simple immunofluorescences using these antibod-ies permitted a clear visualization of either tubulin orteins, we examined the association of these components
to cell centrosomes. Previous studies indicated that tau tau associated with mitotic structures in transformedHuh-7 hepatoma cells (Figs. 5C and 5D). The studiesis a widespread family of polypeptides and that these
components have been found in a variety of cell types with the site-specific antibodies, polyclonal TRS1.2 andthe monoclonal antibody MTB6.22, that tag bindingincluding neuronal cells such as neuroblastoma cells
in culture and nonneuronal lines including Huh-7 hep- epitopes on tau indicated the failure of these antibodies
AID ECR 3360 / 6i18$$$182 12-02-96 01:52:56 eca

384 CROSS ET AL.
FIG. 8. Effects of the polyclonal antibody against native tau, the polyclonal antibody to the N-terminal tau moiety, and the site-specificantibody TRS1.2 on the microtubule assembly nucleated over centrosomes from Huh-7 cells. Isolated centrosomes were stained with theanti-b-tubulin monoclonal antibody. A control sample of isolated centrosomes was preincubated for 3 min with the preimmune rabbit serum,then added to 1.6 mg/ml PC-tubulin, and the resulting assembly product was analyzed by immunofluorescence after 15 min incubation at377C (A). Another sample was preincubated with 20 mg of the antibody to native tau for 3 min, added to 1.6 mg/ml PC-tubulin, and incubatedat 377C for 15 min, and the resulting product was analyzed by immunofluorescence (B). Samples were also preincubated with 20 mg ofeither the antibody against the N-terminal tau moiety (C) or the antibody TRS1.2 against the first tau repeat (D), and the resulting productswere analyzed by immunofluorescence. The effects of antibodies to native tau and to the N-terminal tau domain in blocking microtubulenucleation over centrosomes were observed. Bar, 1 mm. Other details are under Materials and Methods.
to label tau on centrosomes in either interphase or mi- ties in fractions 2 and 5 might be a result of the factthat, in addition to tau association with centrosomes,totic cells.
In order to define more precisely the association of it also binds to centrosomal structures containing re-maining fragments of microtubules. It is interesting totau with centrosomes at the molecular level, centroso-
mal structures were isolated on the basis of differential point out that some of the brain tau isospecies exhibitedthe same migration as those of centrosomal tau fromcentrifugation on sucrose gradients, and the centroso-
mal proteins were assayed by electrophoresis and cultured cells. Other isospecies present in brain tauwere not detected in the centrosomal preparation.Western blot analyses. Figure 6 shows the distribution
patterns of the different fractions, including those con- As another approach to analyze cellular tau-like pro-teins on centrosomes, isolated centrosomal structurestaining the centrosomal proteins, along the 45–70%
discontinuous sucrose gradient (Fig. 6A). The electro- were used as in vitro nucleation centers for the out-growth of microtubules. Isolated centrosomes (Fig. 7A)phoretic pattern of different samples obtained from the
gradient (Fig. 6B), and the immunoblot analysis of indi- were incubated with increasing concentrations of PC-tubulin (Figs. 7B–7E), and the assembled microtu-vidual samples from the gradient, using either a poly-
clonal antibody against native tau (Fig. 6C, left) or an bules were analyzed by immunofluorescence. The asso-ciation of tau with nucleating microtubules was alsoanti-tubulin monoclonal antibody (Fig. 6C, right), are
also shown. This experiment indicates that tau-like corroborated by a double immunofluorescence usingthe pair anti-b-tubulin monoclonal antibody and theisospecies were highly enriched in fractions 2 through
5, e.g., fractions from 60 to 70% sucrose in the discon- antibody to native tau (Figs. 7F and 7G).In a parallel study, centrosomes were also preincu-tinuous gradient as revealed by Western blots (Fig. 6C).
These fractions, displaying the highest reactivity with bated with the polyclonal antibody to native tau, andthis preparation was added to purified PC-tubulin tothe anti-tau antibody, also showed reactivity with the
monoclonal anti-b-tubulin antibody (Fig. 6C, right). Ac- assess the effects of the antibody on microtubule re-growth from centrosomes. A significant decrease in thetually, fractions 2 and 3 showed the highest reactivity
with the anti-tubulin antibody. Thus, a coexistence of number and length of microtubules was observed aftertreatment with this anti-tau antibody (Fig. 8B) com-tau-like components and tubulin was evidenced in this
set of fractions. The apparently bimodal distribution of pared with that in centrosomes allowed to nucleate mi-crotubules without the anti-tau antibody (Figs. 7B–immunoreactive tau suggested by the higher reactivi-
AID ECR 3360 / 6i18$$$183 12-02-96 01:52:56 eca
385TAU-LIKE PROTEINS ASSOCIATED WITH CENTROSOMES
N-terminal tau domain and the site-specific TRS1.2 an-tibody [26] to the tau binding repeat. It was observedthat the polyclonal antibody to N-terminal moietyalso blocked microtubule assembly nucleated overcentrosomes (Fig. 8C) compared with controls in thepresence of nonimmune serum (Fig. 8A). However, noeffect was detected in the presence of an equivalentconcentration of the site-specific TRS1.2 antibody totau (Fig. 8D).
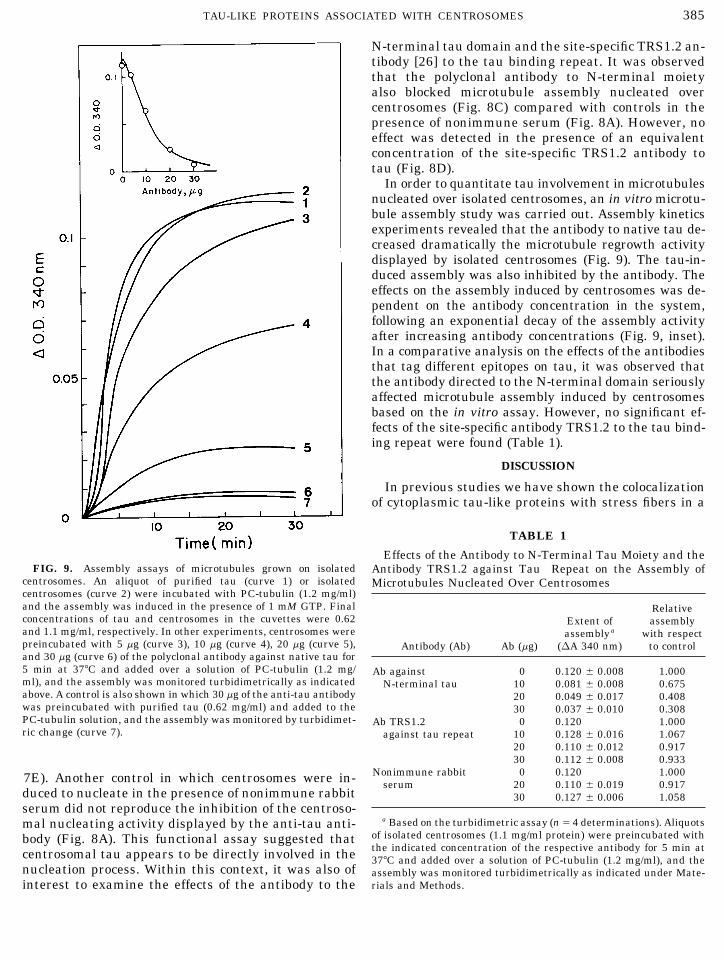
In order to quantitate tau involvement in microtubulesnucleated over isolated centrosomes, an in vitro microtu-bule assembly study was carried out. Assembly kineticsexperiments revealed that the antibody to native tau de-creased dramatically the microtubule regrowth activitydisplayed by isolated centrosomes (Fig. 9). The tau-in-duced assembly was also inhibited by the antibody. Theeffects on the assembly induced by centrosomes was de-pendent on the antibody concentration in the system,following an exponential decay of the assembly activityafter increasing antibody concentrations (Fig. 9, inset).In a comparative analysis on the effects of the antibodiesthat tag different epitopes on tau, it was observed thatthe antibody directed to the N-terminal domain seriouslyaffected microtubule assembly induced by centrosomesbased on the in vitro assay. However, no significant ef-fects of the site-specific antibody TRS1.2 to the tau bind-ing repeat were found (Table 1).
DISCUSSION
In previous studies we have shown the colocalizationof cytoplasmic tau-like proteins with stress fibers in a
TABLE 1
Effects of the Antibody to N-Terminal Tau Moiety and theFIG. 9. Assembly assays of microtubules grown on isolated Antibody TRS1.2 against Tau Repeat on the Assembly of
centrosomes. An aliquot of purified tau (curve 1) or isolated Microtubules Nucleated Over Centrosomescentrosomes (curve 2) were incubated with PC-tubulin (1.2 mg/ml)and the assembly was induced in the presence of 1 mM GTP. Final Relativeconcentrations of tau and centrosomes in the cuvettes were 0.62 Extent of assemblyand 1.1 mg/ml, respectively. In other experiments, centrosomes were assemblya with respectpreincubated with 5 mg (curve 3), 10 mg (curve 4), 20 mg (curve 5), Antibody (Ab) Ab (mg) (DA 340 nm) to controland 30 mg (curve 6) of the polyclonal antibody against native tau for5 min at 377C and added over a solution of PC-tubulin (1.2 mg/ Ab against 0 0.120 { 0.008 1.000ml), and the assembly was monitored turbidimetrically as indicated N-terminal tau 10 0.081 { 0.008 0.675above. A control is also shown in which 30 mg of the anti-tau antibody 20 0.049 { 0.017 0.408was preincubated with purified tau (0.62 mg/ml) and added to the 30 0.037 { 0.010 0.308PC-tubulin solution, and the assembly was monitored by turbidimet- Ab TRS1.2 0 0.120 1.000ric change (curve 7). against tau repeat 10 0.128 { 0.016 1.067
20 0.110 { 0.012 0.91730 0.112 { 0.008 0.933
Nonimmune rabbit 0 0.120 1.0007E). Another control in which centrosomes were in- serum 20 0.110 { 0.019 0.917duced to nucleate in the presence of nonimmune rabbit 30 0.127 { 0.006 1.058serum did not reproduce the inhibition of the centroso-
a Based on the turbidimetric assay (nÅ 4 determinations). Aliquotsmal nucleating activity displayed by the anti-tau anti-of isolated centrosomes (1.1 mg/ml protein) were preincubated withbody (Fig. 8A). This functional assay suggested thatthe indicated concentration of the respective antibody for 5 min atcentrosomal tau appears to be directly involved in the 377C and added over a solution of PC-tubulin (1.2 mg/ml), and the
nucleation process. Within this context, it was also of assembly was monitored turbidimetrically as indicated under Mate-rials and Methods.interest to examine the effects of the antibody to the
AID ECR 3360 / 6i18$$$183 12-02-96 01:52:56 eca

386 CROSS ET AL.
variety of nonneuronal cells in culture [7, 14]. Further- The studies on the effects of anti-tau antibodies onmicrotubules grown from isolated centrosomes, alongmore, there is evidence on the association of tau poly-
peptides with vimentin intermediate filaments in ade- with the analysis on the inhibitory effects of these anti-bodies on the assembly kinetics, point to a functionalnocarcinoma SW-13 cells [34]. In the present report,
we have developed an integrated analysis of the subcel- role of tau in promoting microtubule outgrowth fromcentrosomes. The antibody to native tau blocked micro-lular association of tau-like proteins with centrosomes
in cultured cells. The analysis carried out in this study tubule assembly nucleated over centrosomes in a con-centration-dependent fashion (Fig. 9, inset), an effectallowed us to confirm the multifunctional nature of tau-
like proteins [17] and their widespread subcellular dis- that could be mediated by the interaction of the anti-body with tau on centrosomes as the anchorage site,tribution as evidenced by their association with the
centrosomes. The overall study on tau cellular organi- thus preventing its activity in the growth and stabiliza-tion of nucleating polymers. Steric effects could be in-zation suggests a major structural role for tau and tau-
like components, allowing linkages of cytoskeletal ele- volved in the inhibitory action by the antibody. A posi-tive control showed that the antibody also blocked thements on the one hand [4, 7] and possibly mediating
interactions at the level of the centrosomes on the effect of pure tau on microtubule assembly. It is worth-while to point out that, in control experiments usingother. These observations were corroborated in a set of
different cell lines and in both interphase and mitotic nonimmune serum, no inhibition on microtubulegrowth or on the assembly kinetics was observed.cells.
Current research points to tau as a protein involved These studies suggest that tau association with do-mains on the centrosomal structure is functionally in-in regulatory roles within the cells [4]. At the cyto-
plasmic level, there is increasing evidence that tau volved in nucleation phenomena and that anti-tau anti-bodies preclude the possibility to exert microtubule nu-modulates interactions between microtubules and
stress fibers [7]. According to the present data, tau- cleation activity. Our studies also indicate that theepitopes on the centrosomal tau recognized by the anti-like proteins appear to be involved in the microtubule
nucleation capacity of centrosomes. On the other hand, tau probes are accessible to the antibody reaction. Onthe other hand, the antibody directed to the N-terminalat the level of the cell nucleus, tau appears to modulate
DNA synthesis in vitro, a finding which could be related moiety on tau also recognized tau epitopes on thecentrosome, and this antibody affected microtubule re-to its association with nuclear components [19, 20].
The present studies show that tau is structurally growth from isolated centrosomes in a similar fashionto the antibody to native tau. At the functional level,associated with cellular centrosomes. Tau appears to
colocalize with assembled tubulin in both systems: in studies indicated that the site-directed antibodyTRS1.2 [26] failed to block microtubule nucleation asin vitro preparations of isolated organelles as well as
in cellular centrosomes. The coexistence of tau-like spe- monitored by immunofluorescence and assembly assay(Table 1). This is an interesting observation, consider-cies and tubulin in the sucrose gradient fractions con-
taining isolated centrosomes, together with the set of ing that a MAP-reacting anti-idiotypic monoclonalprobe directed against binding repeats is able to inhibitimmunofluorescence and functional studies, indicates
that the material isolated from the gradients corre- MAP-induced tubulin assembly in solution [27, 28].This situation is different from that of centrosomal tau,sponded to centrosomes. These studies are in agree-
ment with previous reports suggesting the involvement which may exhibit interactions with other proteins ofthis organelle. These observations were correlated withof tau proteins at the level of the centrosome organiza-
tion [35]. More interestingly, the present studies on the immunolocalization findings, in which the antibodyTRS1.2 and the monoclonal MTB6.22 [27], which recog-microtubule nucleation over isolated centrosomes
based on the immunofluorescence analysis (Fig. 8) and nize tau repetitive sequences, did not stain tau oncentrosomes, possibly because binding epitopes wereassembly assays (Fig. 9) of the effects of anti-tau anti-
bodies provide indications of the possible roles of this hidden or involved in tau interactions with tubulin orother centrosomal proteins under the conditions ofprotein in the microtubule outgrowth from these or-
ganizing centers in interphase as well as in mitotic these experiments.The question of whether tau associates with g-tu-cells. In this context, interesting observations on cell-
cycle-dependent relocalizations have been observed bulin on the centrosomal surface has not been eluci-dated yet. The present studies open new avenues towith the Drosophila embryonary MAP-like proteins
DMAP-190 and CP-60 [23, 24] and also with the novel unravel the functional organization of tau polypeptidesand provide data on their association with domains onmicrotubule-interacting regulatory protein Mip-90 in
HeLa cells and in human fibroblasts [36]. However, the the centrosomal structure. These findings indicate thatthese polypeptides could play a critical role in modulat-precise redistributions that tau-like peptides experi-
ence during the cell cycle are not clear at the present ing microtubule nucleation and growth from these or-ganizing centers in the cell. Future studies should pro-stage of our investigations.
AID ECR 3360 / 6i18$$$183 12-02-96 01:52:56 eca

387TAU-LIKE PROTEINS ASSOCIATED WITH CENTROSOMES
15. Himler, A., Dreschel, D., Kirschner, M. W., and Martin, D. W.vide clues on whether there are specific sets of tau(1989) Mol. Cell. Biol. 9, 1381–1388.components involved in centrosomal associations or
16. Lu, Q., and Wood, J. G. (1993) Cell Motil. Cytoskel. 25, 190–whether the same cytoplasmic tau peptides play this200.important cellular role.
17. Wang, Y., Loomis, P. A., Zinkowski, R. P., and Binder, L. I.(1993) J. Cell Biol. 121, 257–267.
This research was supported by Grants 195-0696 from Fondecyt, 18. Loomis, P. A., Howard, T. H., Castleberry, R. P., and Binder,Chile, and The Council for Tobacco Research, U.S.A. (to R.B.M.). We L. I. (1990) Proc. Natl. Acad. Sci. USA 87, 8422–8426.are grateful to the Division of Cancer Treatment of the National19. Shioda, M., Murofushi, H., Murakami-Murofushi, K., and Sa-Cancer Institute, NIH, U.S.A. for the supply of taxol and to Dr. J.
kai, H. (1989) Biochem. Biophys. Res. Commun. 159, 834–841.Nunez, Inserm, France, for the generous supply of the antibody to20. Shioda, M., Okuhara, K., Murofushi, H., Mori, A., Sakai, H.,the N-terminal tau domain. We also thank Mrs. Isabel A. Maccioni
Murakami-Murofushi, K., Susuki, M., and Yoshida, S. (1991)for help in the preparation of the manuscript.Biochemistry 30, 11403–11412.
21. Solomon, F., Magendantz, M., and Salzman, A. (1979) Cell 18,REFERENCES431–438.
22. Karsenti, E., and Maro, B. (1986) Trends Biochem. Sci. 11, 460–1. Olmsted, J. B. (1986) Annu. Rev. Cell Biol. 2, 421–457.463.2. Maccioni, R. B., and Arechaga, J. (1987) The Cytoskeleton in
23. Kellog, D. R., Oegema, K., Raff, J., Schneider, K., and Alberts,Cell Differentiation and Development, ICSU Press-IRL, OxfordB. M. (1995) Mol. Biol. Cell 6, 1673–1684.Univ. Press, London.
24. Kellog, D. R., Field, C., and Alberts, B. M. (1989) J. Cell Biol.3. Kanai, Y., Takemura, R., Oshima, T., Mori, H., Ihara, Y., Ma-109, 1173–1184.sashi, Y., Masaki, T., and Hirokawa, N. (1989) J. Cell Biol. 109,
25. Dominguez, J., Buendia, B., Lopez-Otin, C., Antony, C., and1173–1184.Avila, J. (1994) J. Cell Sci. 107, 601–611.4. Maccioni, R. B., and Cambiazo, V. (1995) Physiol. Rev. 75, 835–
26. Cambiazo, V., Gonzalez, M., and Maccioni, R. B. (1995) J. Neu-864.rochem. 64, 1288–1297.5. Griffith, L. M., and Pollard, T. D. (1982) J. Cell Biol. 78, 958–
27. Cross, D., FarıB as, G., Dominguez, J., Avila, J., and Maccioni,965.R. B. (1994) Mol. Cell. Biochem. 132, 81–90.6. Sattilaro, R. F. (1986) Biochemistry 25, 2003–2009.
28. Vera, J. C., Rivas, C., and Maccioni, R. B. (1988) Proc. Natl.7. Cross, D., Vial, C., and Maccioni, R. B. (1993) J. Cell Sci. 105,Acad. Sci. USA 85, 6763–6767.51–60.
29. Farias, G. A., Vial, C., and Maccioni, R. B. (1992) Mol. Cell.8. Moraga, D., Nunez, P., Garrido, and Maccioni, R. B. (1993) J.Biochem. 112, 81–88.Neurochem. 61, 979–986.
30. Mitchison, T., and Kirschner, M. W. (1984) Nature London 312,9. HenrıB quez, J. P., Cross, D., Vial, C., and Maccioni, R. B. (1995)232–237.Cell Biochem. Funct. 13, 239–250.
31. Bornen, M., Paintrand, M., Berges, J., Marty, M. C., and Karse-10. Binder, L. I., Frankfurter, A., and Rebhun, L. (1985) J. Cellnti, E. (1987) Cell Motil. Cytoskel. 8, 238–249.Biol. 101, 1371–1378.
32. Gaskin, F., Cantor, C. R., and Shelanki, M. L. (1975) Ann. N. Y.11. Drubin, D., and Kirschner, M. W. (1986) J. Cell Biol. 103, 2739–Acad. Sci. 253, 133–146.2746.
33. Towbin, H., Stahelin, T., and Gordon, J. (1979) Proc. Natl. Acad.12. Caceres, A., and Kosik, K. S. (1990) Nature 343, 461–463.Sci. USA 76, 4350–4354.13. Knops, J., Kosik, K. S., Lee, G., Pardee, J. D., Cohen-Gould, L.,
34. Capote, C., and Maccioni, R. B. (1994) Notic. Biol. 2, 64–65.and McConlogue, L. (1991) J. Cell Biol. 114, 725–733.35. Brandt, R., and Lee, G. (1993) J. Neurochem. 61, 997–1005.14. Maccioni, R. B., Vial, C., Cross, D., and Gonzalez, M. (1992) in
Progress in Cell Biology (J. Becerra, J. Perez, and P. Fernandez, 36. Gonzalez, M., Cambiazo, V., and Maccioni, R. B. (1995) Eur. J.Cell Biol. 67, 158–169.Eds.), p. 153–159, Servicio Editorial Universidad de Malaga.
Received January 16, 1996Revised version received September 17, 1996
AID ECR 3360 / 6i18$$$184 12-02-96 01:52:56 eca




















