
Encoding of Time-varying Stimuli in Populations of Cultured Neurons
15
Biol Cybern (2006) 94: 335–349 DOI 10.1007/s00422-006-0051-2 ORIGINAL PAPER Laura Cozzi · Paolo D’Angelo · Vittorio Sanguineti Encoding of time-varying stimuli in populations of cultured neurons Received: 4 August 2005 / Accepted: 21 December 2005 / Published online: 15 February 2006 © Springer-Verlag 2006 Abstract We wondered whether random populations of dissociated cultured cortical neurons, despite of their lack of structure and/or regional specialization, are capable of mod- ulating their neural activity as the effect of a time-varying stimulation – a simulated ‘sensory’ afference. More specifi- cally, we used localized low-frequency, non-periodic trains of stimuli to simulate sensory afferences, and asked how much information about the original trains of stimuli could be ex- tracted from the neural activity recorded at the different sites. Furthermore, motivated by the results of studies performed both in vivo and in vitro on different preparations, which sug- gested that isolated spikes and bursts may play different roles in coding time-varying signals, we explored the amount of such ‘sensory’ information that could be associated to these different firing modes. Finally, we asked whether and how such ‘sensory’ information is transferred from the sites of stimulation (i.e., the ‘sensory’ areas), to the other regions of the neural populations. To do this we applied stimulus reconstruction techniques and information theoretic concepts that are typically used to investigate neural coding in sensory systems. Our main results are that (1) slow variations of the rate of stimulation are coded into isolated spikes and in the time of occurrence of bursts (but not in the bursts’ temporal struc- ture); (2) increasing the rate of stimulation has the effect of increasing the proportion of isolated spikes in the average evoked response and their importance in coding for the stim- uli; and, (3) the ability to recover the time course of the pattern of stimulation is strongly related to the degree of functional connectivity between stimulation and recording sites. These observations parallel similar findings in intact ner- vous systems regarding the complementary roles of bursts and tonic spikes in encoding sensory information. L. Cozzi (B ) · P. D’Angelo · V. Sanguineti Department of Informatics, Systems and Telematics, University of Genova, Via Opera Pia 13, 16145 Genova, Italy E-mail: [email protected] Tel.: +39-010-3536550 Fax:+39-010-3532154 Our results also have interesting implications in the field of neuro-robotic interfaces. In fact, the ability of populations of neurons to code information is a prerequisite for obtaining hybrid systems, in which neuronal populations are used to control external devices. 1 Introduction High level functions in vertebrates, like learning, memory and processing of information emerge from the collective activity of large groups of neurons. These functions are pres- ent in different organisms, regardless of their complexity and anatomical structure. Dissociated cortical neurons, cultured on micro-electrode arrays (MEAs), are a useful model system for studying basic and universal mechanisms underlying high-level cognitive functions and for investigating whether simple computa- tional properties are maintained at lower levels of organi- zation (Martinez-Conde et al. 2002). They can be kept in healthy conditions for a long time, several months (Potter and DeMarse 2001). Their morphological and physiologi- cal properties closely resemble those of the tissue of origin (Kriegstein and Dichter 1983). In fact, the distribution of types of cells in culture is similar to their in vivo counterparts (Huettner and Baughman 1986). Also, from the functional point of view, neuron cultures display activity-dependent, path-specific synaptic modifications (Bi and Poo 1999; Jimbo et al. 1999) that closely resemble the long-term potentiation and depression phenomena found in acute preparations and in vivo. However, the planar substrate provided by the MEA induces a mono-layer of cells, and synaptic connections tend to grow randomly. Developing cultures of dissociated cortical neurons are characterized by spontaneous activity which synchronizes across the whole culture into regular, periodic firing patterns (bursts), with long, totally quiescent intervals (Maeda et al. 1995; van Pelt et al. 2004). After 3–4 weeks in culture, bursts become more spatially confined, and the neural activity loses
Transcript of Encoding of Time-varying Stimuli in Populations of Cultured Neurons

Biol Cybern (2006) 94: 335–349DOI 10.1007/s00422-006-0051-2
ORIGINAL PAPER
Laura Cozzi · Paolo D’Angelo · Vittorio Sanguineti
Encoding of time-varying stimuli in populationsof cultured neurons
Received: 4 August 2005 / Accepted: 21 December 2005 / Published online: 15 February 2006© Springer-Verlag 2006
Abstract We wondered whether random populations ofdissociated cultured cortical neurons, despite of their lack ofstructure and/or regional specialization, are capable of mod-ulating their neural activity as the effect of a time-varyingstimulation – a simulated ‘sensory’ afference. More specifi-cally, we used localized low-frequency, non-periodic trains ofstimuli to simulate sensory afferences, and asked how muchinformation about the original trains of stimuli could be ex-tracted from the neural activity recorded at the different sites.Furthermore, motivated by the results of studies performedboth in vivo and in vitro on different preparations, which sug-gested that isolated spikes and bursts may play different rolesin coding time-varying signals, we explored the amount ofsuch ‘sensory’ information that could be associated to thesedifferent firing modes. Finally, we asked whether and howsuch ‘sensory’ information is transferred from the sites ofstimulation (i.e., the ‘sensory’ areas), to the other regions ofthe neural populations.
To do this we applied stimulus reconstruction techniquesand information theoretic concepts that are typically used toinvestigate neural coding in sensory systems.
Our main results are that (1) slow variations of the rateof stimulation are coded into isolated spikes and in the timeof occurrence of bursts (but not in the bursts’ temporal struc-ture); (2) increasing the rate of stimulation has the effect ofincreasing the proportion of isolated spikes in the averageevoked response and their importance in coding for the stim-uli; and, (3) the ability to recover the time course of the patternof stimulation is strongly related to the degree of functionalconnectivity between stimulation and recording sites.
These observations parallel similar findings in intact ner-vous systems regarding the complementary roles of burstsand tonic spikes in encoding sensory information.
L. Cozzi (B) · P. D’Angelo · V. SanguinetiDepartment of Informatics, Systems and Telematics,University of Genova, Via Opera Pia 13,16145 Genova, ItalyE-mail: [email protected].: +39-010-3536550Fax:+39-010-3532154
Our results also have interesting implications in the fieldof neuro-robotic interfaces. In fact, the ability of populationsof neurons to code information is a prerequisite for obtaininghybrid systems, in which neuronal populations are used tocontrol external devices.
1 Introduction
High level functions in vertebrates, like learning, memoryand processing of information emerge from the collectiveactivity of large groups of neurons. These functions are pres-ent in different organisms, regardless of their complexity andanatomical structure.
Dissociated cortical neurons, cultured on micro-electrodearrays (MEAs), are a useful model system for studying basicand universal mechanisms underlying high-level cognitivefunctions and for investigating whether simple computa-tional properties are maintained at lower levels of organi-zation (Martinez-Conde et al. 2002). They can be kept inhealthy conditions for a long time, several months (Potterand DeMarse 2001). Their morphological and physiologi-cal properties closely resemble those of the tissue of origin(Kriegstein and Dichter 1983). In fact, the distribution oftypes of cells in culture is similar to their in vivo counterparts(Huettner and Baughman 1986). Also, from the functionalpoint of view, neuron cultures display activity-dependent,path-specific synaptic modifications (Bi and Poo 1999; Jimboet al. 1999) that closely resemble the long-term potentiationand depression phenomena found in acute preparations andin vivo.
However, the planar substrate provided by the MEAinduces a mono-layer of cells, and synaptic connections tendto grow randomly.
Developing cultures of dissociated cortical neurons arecharacterized by spontaneous activity which synchronizesacross the whole culture into regular, periodic firing patterns(bursts), with long, totally quiescent intervals (Maeda et al.1995; van Pelt et al. 2004). After 3–4 weeks in culture, burstsbecome more spatially confined, and the neural activity loses

336 L. Cozzi et al.
part of its periodic regularity (Maeda et al. 1998). This kindof dynamics persists for the remaining life span, thus rep-resenting a mature state of the network (Marom and Shahaf2002). It has been suggested that the ability of cells to exhibitbursting behavior requires de-inactivation of specific Ca++channels (see Sherman 2001 for a review). However, in cul-tured neurons, a major contribution to the widespread, syn-chronous bursting activity is likely to come from a hyper-excitation through positive feedback (Robinson et al. 1993).Modelling and experimental studies (Latham et al. 2000a,b)have suggested that a necessary condition for network burst-ing is the presence of a small portion of tonically active cells.An increase of such portion would result in a transition to alow-rate firing regime.
Maeda et al. (1995) studied the effect of slow, periodic fo-cal electrical stimulation on cultured neurons, at 1–30 s inter-stimulus intervals (i.e., rates of stimulation of 0.03–1 Hz).They found that up to a certain rate of stimulation, the evokedbursts are strictly time-locked to the stimuli. Wagenaar et al.(2005) suggested that synchronous bursting is the result ofsensory deprivation, and demonstrated that distributed stim-ulation with global rates of up to 50 Hz spread over up to 25sites (i.e., up to 2 Hz per site), is indeed capable of disruptingthe bursting behavior and inducing a switch to a tonic firingregime.
One major issue in brain functions is how sensory infor-mation is coded into the spike trains generated by primarysensory neurons, and how such information is transferred tohigher-order neurons and to motor areas. A better understand-ing of how the neural activity of non-sensory systems relatesto sensory activity would contribute to unveil the mechanismsof information representation and processing in the nervoussystem.
With this in mind, we asked whether and to what extentpopulations of dissociated cultured neurons, despite their ran-dom connectivity and lack of regional specialization, arecapable of modulating their neural activity as the effect ofa time-varying stimulation – a simulated ‘sensory’ affer-ence. We also asked how such ‘sensory’ information is trans-ferred from the sites of stimulation (i.e., the primary ‘sensory’areas) to the neurons which are further away from them interms of functional connectivity. Like Maeda et al. (1995),we used low-frequency stimulation (up to 2 Hz), but appliednon-periodic trains of stimuli to simulate sensory afferences.
It has been suggested (Bakkum et al. 2004; Martinoiaet al. 2004) that populations of dissociated cultured neuronscan be seen as a technology, which may potentially allowimplementation of bio-artificial evolvable, adaptive informa-tion processing and/or control modules. As part of an effortin this direction (Cozzi et al. 2005), the trains of stimuli wereobtained according to a variety of ‘coding’ rules from sam-ples of the recordings of the infra-red proximity sensors ofa mobile robot during free navigation in its playground. Wethen asked how much information about the original sen-sory recordings could be extracted from the neural activityrecorded at the various sites.
Furthermore, motivated by the results of studiesperformed both in vivo and in vitro on different prepara-tions which suggest that different firing modes (bursts andlow-rate isolated spikes) play different roles in coding time-varying signals (Metzner et al. 1998; Reinagel et al. 1999;Sherman 2001; Martinez-Conde et al. 2002; Krahe and Gab-biani 2004), we explored the amount of ‘sensory information’that could be associated to these different firing modes.
We followed the same approach for the analysis asReinagel et al. (1999) on in vivo recordings from lateralgeniculate nucleus (LGN) cells in anesthetized cats.
2 Materials and methods
2.1 Neural preparation and electrophysiological techniques
We used primary cultures of cortical neurons, extracted fromrat embryos (E18). Cells were plated on planar arrays of60 TiN/SiN electrodes (diameter 30µm, distance 200µm),arranged along a square grid (MEA60, Multichannel Sys-tems, Reutlingen, Germany) and coated with adhesion fac-tors (poly-D-lysine and laminin). When cells adhered to thesubstrate, 1 ml of medium was added in each device. Cellswere incubated with 1% Glutamax, 2% B-27 supplementedNeurobasal medium (Invitrogen), in a humidified atmosphere(5% CO2, 95% air) at 37◦C until the beginning of the exper-imental sessions. 50% of the medium was changed everyweek. MEA dishes were covered with caps to prevent evap-oration.
Neurons rearrange randomly in a bi-dimensional struc-ture and, after having established synaptic interconnections[at about 7 days in vitro (DIVs)], they show spontaneous neu-ral activity that can be modulated by means of electrical stim-ulation (Maeda et al. 1998; Jimbo et al. 2000; Marom andShahaf 2002). Experiments were performed at 18–42 DIVs,when the neuronal network reaches its ‘mature’ state, con-sisting of non-periodic, synchronized, clustered activity withminute-to-minute fluctuations in the probability of firing(Marom and Shahaf 2002). All the recordings were per-formed in Neurobasal medium.
2.2 Stimulation protocol
Sensory information consisted of samples of the activity ofthe proximity sensors of a mobile robot (duration: 300 s),moving inside a circular playground (diameter 0.80 m) thatcontained cylindrical obstacles. The robot (Khepera II, K-team) is equipped with two wheels and eight infrared sensorsthat provide information about the proximity of the robotto obstacles. We set a reactive controller implementing anobstacle avoidance behavior – a Braitenberg’s vehicle, (Brait-enberg 1984). The maximum speed of the robot was set at40 mm/s.

Encoding of time-varying stimuli in populations of cultured neurons 337
The sensory signals recorded from the proximity sensorsof the robot were converted (i.e., coded) into spatio-temporalpatterns of stimuli according to two alternative schemes:
• Intensity coding: the instantaneous rate of stimulation isproportional to sensor activity s(t), and the maximum rateof stimulation, rmax
s , is only attained when the robot hitsan obstacle. More precisely, the time to the next stimulusis proportional to the inverse of sensor activity at time ti :
ISIi = �τi
= ti+1 − ti = 1
rmaxs
· smax − smin
s(ti ) − smin∝ 1
s(ti ). (1)
Higher rates of stimulation allow a better temporal reso-lution in the coding of the sensory signal (Gabbiani et al.1996; Cozzi et al. 2005). To test how sensory coding isaffected by this, we performed experiments respectivelyat maximum stimulation rates of 1 and 2 Hz.
• Event coding: a stimulus is generated only if the deriva-tive of the sensory signal, s(t), is above a fixed threshold,S. In other words, one stimulus is generated wheneverthere is an abrupt increase in the value of s(t).
Figure 1a shows a sample of the sensory signal and illus-trates the patterns of stimulation that result from the threedifferent coding schemes. Figure 1b displays the statisticaldistribution of the values of s(t) and s(t)
∣∣s(t)>S . The infra-
red range sensors of the Khepera robot are highly non-linear.
–5 0 5
0
500
1000
Time [s]
EventInt 1 HzInt 2 Hz
ST
A
c
Sensory signal
[Derivative]+
Event
5 s
Intensity 1 Hz
Intensity 2 Hz
a
0 500 10000
0.2
0.4
0.6
0.8
Encoded signal
p(en
code
d si
gnal
) IntensityEvent
b
Fig. 1 The different coding schemes. a From top to bottom a sampleof the sensory signal; the positive part of the derivative of the sen-sory signal, used in case of ‘event’ coding (a stimulus was generatedwhenever the derivative exceeded the threshold); stimuli obtained fromthe intensity coding scheme (maximum stimulation rates 1 and 2 Hz);stimuli obtained from event coding scheme. b Probability density func-tion of the encoded signal respectively for intensity and event coding. cStimulus-triggered average, i.e., average signal feature that evokes onestimulus, for the three coding types. STAs were obtained as the impulseresponses of the filter that optimally reconstructs the sensory signalfrom the trains of stimuli that result from the three coding schemes (seeSect. 2)
They tend to saturate when there is a reflective surface (i.e.,an obstacle) at a distance of 5 cm or less. However, if thereare no obstacles in close proximity, they are only weakly ac-tive, reflecting the level of ambient light. As a consequence,while the robot moves, the sensory signal s(t) tends to switchbetween two extreme values (i.e., absence or presence ofobstacles). Figure 1b shows the statistical distribution of theencoded signals during a typical robot experiment. The solidline (‘intensity’ coding) has a bi-modal distribution, witha peak at around s = 1, 000 (reflecting sensor saturation,which occurs when the robot comes close to an obstacle),and a Poisson-like component (reflecting ambient light fluc-tuations) at lower values of s. The different coding schemesare completely described by their stimulus-triggered average(see Fig. 1c), i.e., the average sensory signal that elicits astimulus. In particular, in intensity and event coding, stim-uli are evoked by signals that increase more slowly or moreabruptly, respectively.
The coding schemes also differ in that they end up indifferent average rates of stimulation. More specifically, inevent coding, the average rate of stimulation is 0.11 Hz.In intensity coding at 1 and 2 Hz, the rates of stimulationare, respectively, 0.26 and 0.51 Hz.
Individual stimuli consisted of monopolar, biphasic pulses(positive phase first), with a 1.5–2.5 V peak-to-peak ampli-tude, and a total duration of 250µs; these parameters are inthe range of maximally effective stimulation as reported byWagenaar et al. (2004).
2.3 Experimental protocol
The experiments consisted of delivering trains of stimuli toone specific MEA site, and recording the activity from the(up to 59) remaining electrodes. The site of stimulation wasselected randomly, and changed if no observable responsewas detected.
Each experimental session involved a different neuronalculture. A total of eight preparations were tested. The pro-tocol consisted of multiple epochs; each epoch consisted ofa 300 s interval during which stimuli generated according toone of the three coding schemes (intensity 1 Hz, intensity2 Hz, event) were applied to a selected site of stimulation.
For each experiment, epochs were iterated with a differentcoding scheme until there was a detectable evoked response.For each experiment, the type of coding scheme was random-ized over epochs, and subsequent epochs were interleaved byresting periods of 600 s.
2.4 Data analysis
2.4.1 Spike detection
The raw electrophysiological signals were sampled at 10 kHz,and the individual spikes were detected through a threshold-ing algorithm over a 4-ms time window (Chiappalone et al.

338 L. Cozzi et al.
2003; Novellino et al. 2003). Signals recorded on each chan-nel reflect the activity of 1–2 neurons; however, we did not usespike sorting to distinguish the contributions of the differentneurons.
2.4.2 Evoked response
To investigate the neural activity evoked by stimulation, foreach epoch and for each recording site we measured thepost-stimulus time histogram (PSTH), which represents theimpulse response of each site of the neural preparation toelectrical stimulation. The PSTHs were calculated by taking500-ms time windows from the recordings that follow eachstimulus. We then counted the number of spikes occurring in-side each 4-ms bin, and divided this measure by the numberof stimuli and by the bin size so that the PSTH is expressed asinstantaneous firing rate. In these cultured cortical neurons,typical PSTHs have an ‘early’ (less than 25 ms) and a ‘late’(25–500 ms) component (Jimbo et al. 2000). To quantify theshape of the PSTH, we computed the following indicators bydefining 0–32 and 32–500 ms as the ‘early’ and ‘late’ inter-vals respectively:
1. Latency (s) of the peak of the early and late component.2. Magnitude (spikes/s) of the peak of the early and late
component.
2.4.3 Stimulus reconstruction
To assess the extent to which neural activity encodes time-varying sensory activity, for each recording site we estimatedthe sensory input from the observed spike train through anoptimal linear filtering approach (Gabbiani and Koch 1996;Rieke et al. 1997).
More specifically, we looked for the linear filter with im-pulse response h(·) that optimally estimates the sensory sig-nal s(t) from the train of stimuli, y(t):
s(t) = sest(t) + n(t) =+∞∫
∞h(τ )y(t − τ)dτ + n(t), (2)
where sest(t) is the portion of s(t) reconstructed by the filter,and n(t) is the residual – i.e., the unmodeled portion of s(t).The optimal h(·) minimizes the variance of n(t). Its Fou-rier transform (i.e., the transfer function) is given by (Wiener1949):
H(ω) = Sys(ω)
Syy(ω), (3)
where Sys(ω) , is the cross-spectrum between y(t) and s(t)and Syy(ω) is the power spectrum of y(t).
We then took the fraction of the variance of s(t) explainedby its estimate, sest(t), as a measure of the reconstruction per-formance, i.e., how well one specific recording site encodesthe sensory signal. Hereafter we indicate this quantity as thecoding fraction (CF) of that specific recording site (Gabbiani1996; Gabbiani and Koch 1996). To understand how recon-struction performance – and therefore coding – changes at
different frequencies, we also compared the power spectra ofthe sensory signal and of its reconstruction.
All coding schemes imply a loss of information, whichdepends on the statistics and power spectrum of the sensorysignal, the specific coding scheme and the maximum rate ofstimulation (Gabbiani and Koch 1996). As a reference, wecomputed the fraction of the variance of the sensory signalthat can be reconstructed from the stimuli. This representsan upper bound for the coding fraction for a specific codingscheme and rate of stimulation. We found, in particular, astimulus coding fraction of 65, 77 and 88% for, respectively,event and intensity coding (the latter, at 1 and 2 Hz).
2.4.4 Information transmission and coding efficiency
To estimate the coding efficiency (in an information theoreticsense) of our artificial sensory system, we obtained an upperbound for the entropy rate of the observed spike train from theestimated probability density function, P(τ ), of their inter-spike intervals (ISI) (Bialek et al. 1991; Rieke et al. 1997;Reinagel et al. 1999). Under the assumption that subsequentISIs are generated independently, i.e., that the spike train isa renewal process, the entropy rate can be estimated as:
Rcapacity ≤ −ry
∑
P(τ ) · log2 P(τ ) (bits/s) (4)
where the sum is taken over all the ISI values, and ry is themean firing rate (MFR) [spikes/s]. This quantity specifies theinformation capacity of the neural channel.
We then estimated the amount of information (in a unittime interval) that the spike train y(t) conveys about the sen-sory input s(t), as the mutual information of s and y (Gab-biani and Koch 1996; Rieke et al. 1997). A lower bound forthe mutual information (Gabbiani and Koch 1996) can becomputed as:
Rinf = Is,y ≥+ fc∫
− fc
log2[SNR( f )]d f (bits/s) (5)
where fc denotes the bandwidth of the sensory stimulus.From the power spectrum of the sensory signals, we usedfc = 0.25 Hz.
The signal-to-noise ratio (SNR), where noise was definedas in Eq. (2), was evaluated from the results of the optimallinear reconstruction.
We took Rinf as an estimate of the rate of informationtransmitted by the spike train about the stimulus. The rateof transmitted information represents the information that iseffectively transmitted through the communication channel,while the entropy of the spike train is a measure of the amountof information that could in principle be conveyed by thespike train. Therefore, it is interesting to assess the potentialthat the communication channel does actually exploit, i.e.,its coding efficiency. This is defined as the ratio of mutualinformation and the channel capacity (Reinagel et al. 1999):
ε = Rinf
Rcapacity(6)

Encoding of time-varying stimuli in populations of cultured neurons 339
0 0.5 10
0.5
1
ISI – pre [s]
ISI –
pos
t [s]
Burst Event
Raw data
Spike train
Bursts
0.2 s
a b
Fig. 2 Definition of bursts and isolated spikes. a Scatter plot of pre- and post-ISI, for one typical experiment and recording site. The clusters ofpoints that are parallel to the coordinate axes denote beginning and end of bursts. b Classification into isolated spikes and bursts. Spikes lyingless than 100 ms apart were classified as belonging to bursts. Burst events are the initial spikes in each burst
It should be noted that this is a lower bound of the actual cod-ing efficiency, as Rinf and Rcapacity are the lower and upperbound estimates, respectively.
2.4.5 Burst detection
The spontaneous activity of cultured neurons is dominatedby bursts. To study whether this is still the case in evoked re-sponses, we distinguished between bursts and isolated spikes.In particular, we defined as ‘isolated’ the spikes that are atleast 100 ms away (before and after) from other adjacentspikes. All other spikes were considered as parts of bursts.According to this criterion, we did not require a minimumnumber of spikes to form a burst: therefore, “bursts” mightexist, formed by any number of spikes (down to 2).
The choice of the above threshold for the classificationof bursts versus isolate spikes was suggested by scatter plotsof the joint inter-spike interval (JISI) (See Fig. 2). For eachspike, a single data point is plotted representing the preced-ing ISI as a function of its subsequent ISI. The two clus-ters that are closer to the coordinate axes correspond to theinitial and final spikes in a burst. The 100 ms threshold for theburst detection algorithm (corresponding to the dashed linesin Fig. 2a) allows confining the clusters to the shaded area.Based on this criterion, from each spike train we obtained fivedifferent time series for analysis: (1) all spikes (the originalspike train itself, A), (2) bursts (spikes belonging to bursts,B), (3) burst events (the initial spikes of each burst, BE), (4)isolated spikes (spikes not belonging to bursts, I), and (5)isolated spikes + burst events (the union of burst events andisolated spikes, BE+I). Figure 2b shows an example of theapplication of the algorithm.
2.4.6 Statistical analysis
We investigated whether and how the individual firing modesdiffer in their ability to code sensory information. For this pur-pose, for the intensity coding epochs, we applied an ANO-VA to each of the estimated CFs, with two factors: (1) firingmode (all, bursts, burst event, isolated, burst event +isolated);and (2) maximum rate of stimulation (1 and 2 Hz). For event
coding, the ANOVA only had one factor (firing mode). Weused a random effect model (random intercept) to properlyaccount for the correlation among different sites and differ-ent epochs of the same experiment. We also compared thecoding performances for each pair of firing modes.
We then explored the relationship between functionalconnectivity and coding performance. For this purpose, weapplied a 2-way ANOVA to the estimated CF (factors: spiketype and the descriptive parameters of the impulse response).
3 Results
Shape and magnitude of the estimated impulse response(PSTH) varied in different preparations and at differentrecording sites within the same preparation (see Fig. 3 foran example).
Shape and magnitude of the PSTHs provide indicationson the functional connectivity between neurons placed in theproximity of the stimulation site, and those lying around eachrecording site. In particular, we assumed that sharp, earlypeaks indicate more direct synaptic pathways, and thereforestronger functional connectivity. In contrast, late componentsreflect longer and/or more indirect pathways (see Fig. 3b).Such late components display greater spatio-temporal fluc-tuations. This is in accordance with previous findings withthis type of preparation (Jimbo et al. 2000; Marom and Shahaf2002).
3.1 Coding of time-varying sensory signals
We examined the intensity coding experiments in terms of theperformance in reconstructing the ‘sensory’ input from theobserved spike train. Figure 4a shows, for a typical exper-iment, the ‘sensory’ signals and their reconstruction. Forreference, the figure also reports signal reconstruction fromthe train of stimuli that were delivered to the preparation.The latter estimate can be considered as an upper bound forthe reconstruction performance determined by the specific

340 L. Cozzi et al.
0
150
0
150
0
150
0
150
0
150
0
150
0 0.50 0.50 0.50 0.50 0.50 0.5
Time [s]
PS
TH
[spk
/s]
a
0 0.1 0.2 0.3 0.40.40
50
100
Time [s]
PS
TH
[spk
/s]
b
Fig. 3 Impulse responses [post-stimulus time histograms (PSTH)] of the neural preparation to an electrical stimulus. a PSTHs for each recordingsite, arranged according the actual topography of the Micro-electrode array electrodes, in a typical experiment. The evoked response changesin both shape and magnitude. Blank squares indicate sites that are not responding; the X indicates the site of stimulation. Bin size: 8 ms. bMagnification and zoom of the PSTH for electrode 58: the dotted line discriminates between the early and the late response (time = 32 ms)
coding scheme and the specific sensory signal. The recon-struction performance from the spike trains is far less accurate(maximum 34% – intensity coding, and 38% – event coding).Moreover, it varies greatly at different sites (Fig. 4b), and isreasonably accurate (CF ≥ 30%) only in a small fraction ofthe recording sites (Fig. 4c).
A typical impulse response of the reconstruction filtersthat allows optimal estimation of the sensory signals fromthe spike trains is displayed in Fig. 4d. This response canbe interpreted as the spike-triggered average stimulus (STA)(Rieke et al. 1997), i.e., the average sensory signal that, onaverage, elicits a spike. At least on the sites displaying goodreconstruction performance, a spike is elicited by a gradualincrease (in few cases, a decrease) of the sensory signal. Thisbehavior is qualitatively similar in different preparations, andis essentially a consequence of the intensity coding scheme(see Fig. 1c).
3.2 Coding of sensory signals by different firing modes
To investigate the contribution of the different typologies ofspikes to sensory coding, we separately analyzed the recon-struction performance obtained from each specific spike type.
For a typical experiment, Fig. 5a shows the results of thereconstruction from: (1) all the recorded spikes; (2) the ‘burst’spikes i.e., the spikes that belong to bursts, (3) the ‘burstevents’, i.e., the initial spikes in each burst (4) the ‘isolated’spikes, i.e., those that are not part of bursts, and (5) theisolated spikes and the burst events, taken together. To com-pare the coding performance obtained for the different firing
modes, for each experiment and epoch we focused on therecording sites which displayed a reasonably accurate recon-struction performance (CF ≥ 30%) for at least one spike type.
Statistical analysis revealed that different firing modesdiffer significantly in their coding performances (F(4, 652) =115.70, p < 0.0001). The maximum rate of stimulation alsosignificantly affects the CF (F(1, 84) = 5.12, p = 0.026).Significant interactions are also present (F(4, 652) = 21.29,p < 0.0001).
Figure 5b suggests that burst events and isolated spikesresult in a better coding performance. In fact, burst events andisolated spikes, overall, significantly outperform the recon-struction performance from all spikes and from bursts(p < 0.0001, in both cases). Moreover, the difference be-tween isolated spikes and burst event is not statistically sig-nificant, but isolated spikes and burst events outperform bothof them, when taken separately (p < 0.0001).
Figure 6 displays the STA for isolated spikes and burstevents (the burst-triggered average stimulus, i.e., the averagesensory signal that evokes one burst), for different prepara-tions and recording sites. Their shapes are quite similar, thussuggesting that burst events and isolated spikes code are sim-ilar features of the signal.
To further investigate this, we compared the power spectraof the actual sensory signal, and of its linear reconstructions(see Fig. 6b). In general, only the lower frequency compo-nents of the sensory signal can be reconstructed from theneural activity, and there are no obvious differences in theportions of spectrum reconstructed from isolated spikes andburst events. However, a look at the reconstructed signal inFig. 5a suggests that, when the sensory signal has two peaks,

Encoding of time-varying stimuli in populations of cultured neurons 341
5 s
Sensorysignal
Fromstimuli
Fromspikes
77%
34%
a
10
20
30
40b
0 10 20 30 4010
–3
10–2
10–1
100
EventInt 1 HzInt 2 Hz
CF (%)
p(C
F)
c
–10 –5 0 5
CF = 33%
SpikesStimulus
Time [s] S
TA
d
Fig. 4 Reconstruction performance from all spikes (intensity coding, maximum stimulation rate: 1 Hz). a From top to bottom sensory signal, itsreconstruction from the pattern of stimulation, and its reconstruction from one recording site. The numbers on the right indicate the correspondingcoding fractions CFs. b Variability of the CFs at different recording sites, for one experiment. The stimulation site is indicated with the X . cstatistical distribution of the CF values, for the different coding schemes. d Spike-triggered average stimuli (STA) for intensity coding (1 Hz).The STAs were computed as the impulse responses of the optimal reconstruction filters; the figure shows the STAs respectively obtained fromthe pattern of stimuli and from the spike train on one recording site, after normalization
one is coded into burst events, and the other into isolatedspikes. One possible explanation is that when the neuronsrespond with a burst, for some time the network is unable togenerate additional bursts. This is indeed the case, as shownin the data sample of Fig. 6c.
Opitz et al. (2002) found that the duration of an evokedburst increases with the time between stimulus and the previ-ous spontaneous burst. In other words, burst duration codesfor the time from the previous inter-stimulus interval.
We asked whether this is still the case with sustained,time-varying stimulation. Only occasionally did we find aclear relation between the number of spikes per burst and thetime from the previously evoked burst (see Fig. 6d); how-ever, the finding was not consistent in all preparations (seeFig. 6e).
This may explain the observation that burst events codefor the sensory signal more reliably than entire bursts. Infact, if all bursts had exactly the same duration or number ofspikes, bursts and burst events would be expected to resultin an almost identical reconstruction performance as burstsdo not carry additional information about the stimulus. How-ever, in linear reconstruction techniques, each spike codes forthe same stimulus feature, and more spikes would predict astronger stimulus. Therefore, if burst duration or number ofspikes per burst were proportional to the instantaneous rate
of stimulation (i.e., inversely proportional to the time fromthe previous stimulus), reconstruction based on bursts wouldhave resulted in a better performance in comparison to burstevents. Such types of burst-based neural codes have beensuggested by (Kepecs et al. 2002; Kepecs and Lisman 2003)as emergent biophysical properties of model pyramidal neu-rons. However, with a neural code in which burst duration isproportional to the time from the previous stimulus, as foundby Opitz et al. (2002), a stronger sensory signal would resultin less spikes per burst. Therefore, the performance of lin-ear reconstruction from all spikes in a burst would have beenworse than that from burst events. Performance would havebeen even worse if burst structure had no relationship at allwith the sensory signal, as we found here.
3.3 Effect of the rate of stimulation
For isolated spikes, the coding performance was significantlygreater for the 2 Hz rate compared to 1 Hz (p < 0.0001).A similar effect was observed for burst events and isolatedspikes, taken together. Instead, the rate of stimulation does notseem to significantly affect the reconstruction performance ofburst events. In contrast, for all the spikes and the burst spikes,the coding performance decreases when the maximum rate of

342 L. Cozzi et al.
Sensorysignal
Stimuli 88%
All spikes 5%
Bursts 3%
BurstEvents 45%
Isolated 52%
Isolated + Burst Events
72%
5 s
a
A B BE I BE+I0
10
20
30
40
Ave
rage
CF
(%
)
b
Fig. 5 a Reconstruction performance from different firing modes, forone sample experimental session (intensity coding, maximum stimula-tion rate: 2 Hz). From top to bottom sensory signals, their reconstruc-tion from: the stimuli; all the spikes (A); bursts (B); burst events (BE);isolated spikes (I); and isolated spikes and burst events (BE+I). Thenumbers on the right indicate the respective CFs (%). b Summary ofreconstruction performances per firing mode, for all experiments
stimulation increases (p = 0.0051 and p = 0.0026 respec-tively) (see Fig. 7a, b).
We then asked whether there are substantial differencesin the average impulse responses that result from 1 and 2 Hzstimulation. In particular, we assessed the ratio between theaverage number of evoked isolated spikes and the averagenumber of evoked ‘events’ (i.e., either isolated spikes orburst events). We found that doubling the rate of stimulationhas the effect of greatly increasing the relative proportion ofevoked isolated spikes (from 31.1 to 49.7%, p < 0.0001) (seeFig. 7c).
3.4 Coding of isolated ‘events’
An entirely different situation was found for event coding(see Fig. 8). In this case, a stimulus is delivered to the neuro-nal population when there is an abrupt increase of the sensorysignal, and linear reconstruction simply aims at recoveringthe original pattern of stimulation from the neural activity.
This is clearly reflected in the STAs, which present a sharppeak (Fig. 8b).
The shape of the STA is less variable across recordingsites than in the case of intensity coding, suggesting that theoriginal pattern of stimulation is more easily identifiable thanthe time courses of the instantaneous rate of stimulation. Withevent coding, the reconstruction performances obtained fromall the spikes are remarkably greater than with intensity cod-ing (compare Figs 5b and 8c). The difference is even morerelevant in relative terms. In fact, this coding scheme has thelargest loss of information in going from sensory signal tostimuli (CF = 65%, see sect. 2).
The performance of event coding also differs from inten-sity coding in the relative contributions of each firing mode(see Fig. 8a, c). The CF is still highly affected by the firingmode (F(4, 328) = 363.13, p = 0.0001), but all spikes andburst spikes are significantly better than all the other groups(p < 0.0001 and p < 0.0001, in both cases).
3.5 Information rate and coding efficiency
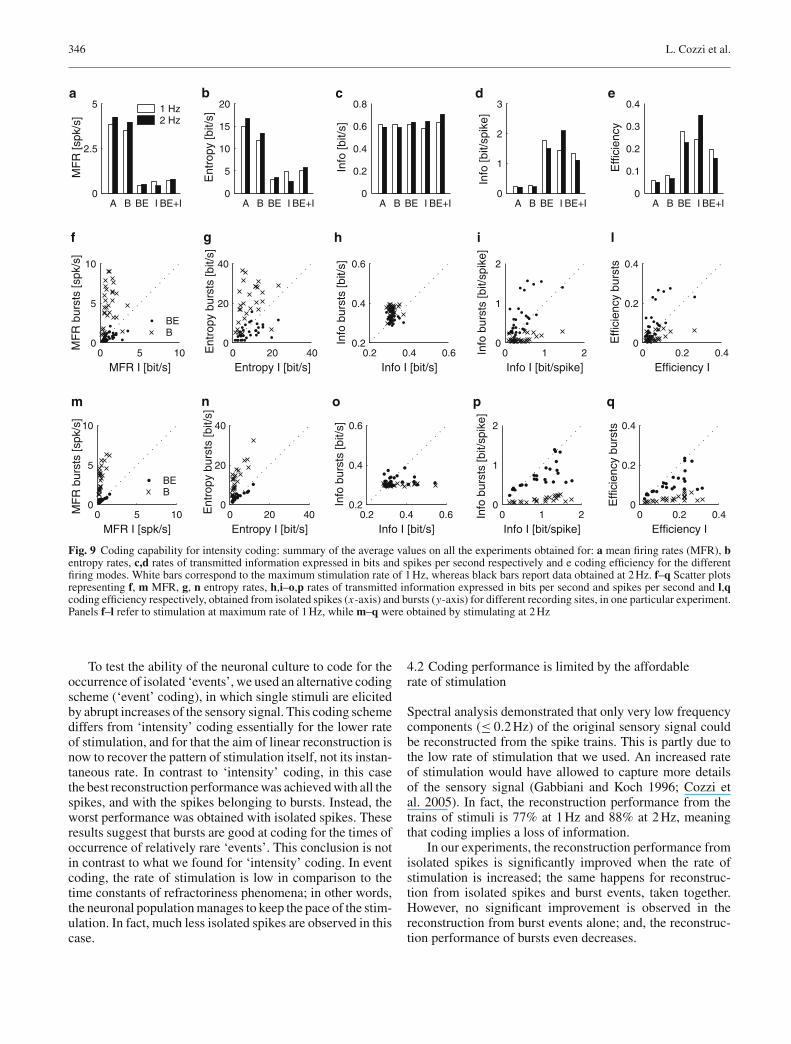
Figure 9 shows the results of the information-theoretic analy-sis for the intensity coding scheme. We restricted the analysisto spike trains which contain at least 60 spikes (correspondingto a MFR of at least 0.2 spikes/s).
Figure 9c reports the average information rates for eachspike type and for the different rates of stimulation. Theseresults are quite similar for the different firing modes, par-ticularly at the maximum stimulation rate of 1 Hz (0.62 bits/sfor all the spikes, bursts and burst events, 0.58 bits/s for iso-lated spikes and 0.63 bits/s for isolated spikes together withburst events). However, when the stimulation rate increases,burst events, isolated spikes and their union slightly outper-form the whole spike train and bursts (0.59, 0.59, 0.64, 0.64and 0.71 bits/s for A, B, BE, I, and BE+I, respectively.). Thedifference in the information content of the firing modes iseven more remarkable if we consider that the correspond-ing spike trains are formed by appreciably different numbersof events as apparent from the MFR displayed in Fig. 9a.Moreover, for each recording site not all the spike typessatisfied the MFR threshold criterion; therefore, only someof them were considered in this analysis; this is why theMFR of the isolated spikes and the burst events taken to-gether differs from the sum of the values obtained taking themseparately.
We then looked at the information per spike. Fig. 9dshows that regardless of the maximum rate of stimulation onaverage burst events and isolated spikes, both taken together(1.32–1.11 bits/spike at 1 and 2 Hz respectively) and sepa-rately (1.77–1.50 bits/spike and 1.42–2.10 bits/spike respec-tively), carry 5–10 times more information than all spikesand bursts (0.23–0.20 and 0.25–0.21 bits/spike respectively).These results basically confirm the findings of linear recon-struction analysis. However, they also suggest that burst eventsand isolated spikes are the most informative spike types atboth 1 and 2 Hz respectively (see Fig. 9d).

Encoding of time-varying stimuli in populations of cultured neurons 343
0.05 0.1 0.2 0.3 0.4 0.50
2
4
6
8
10
12
14
16
18x 10
7
SensorsBEIBE+I
Frequency [Hz]
PS
D (
sens
ory
sign
al)
b
–200
0
200
400
ST
A
–15 –10 –5 0 5 –200
0
200
400
Time [s] –15 –10 –5 0 5
IsolatedBurst events
a
0 5 10 150
10
20
30
40
50
Inter–Stimulus Interval [s]
# sp
ikes
per
bur
st
d
0 5 10 150
10
20
30
40
50
Inter–Stimulus Interval [s]
# sp
ikes
per
bur
st
e
15 15.1 15.2 15.3 15.4 –300
–200
–100
0
100
200
300
Ext
race
llula
r po
tent
ial [
µV]
Time [s]
17.1 17.2 17.3 17.4 17.5
c
Fig. 6 Isolated spikes and burst events code for similar features of the instantaneous rate of stimulation. a STA (solid line) and burst-triggeredaverage (dotted line) stimuli for a selection of recording sites. b Power spectral densities of the original sensory signal, and of its reconstructionfrom the different spike types. c Raw electrophysiological signal showing consequent stimuli evoking alternatively a burst and an isolated spike;insets: magnification of the early (0–32 ms) response (y range = 300µV). d, e Number of spikes belonging to a burst (including the limit case ofone single spike, i.e., an isolated spike) as a function of the temporal distance between the stimulus triggering that burst and the preceding pulse,for one single recording site during an epoch of an experiment. In some cases (d) short inter-stimulus intervals seem to evoke shorter bursts, whilein other cases (e) the relationship is not so clear
We also compared the information content of isolatedspikes and bursts, the latter considered either as unitary eventsor with their inner structure (see Fig. 9f). The information
rates (in bits/s) for the different recording sites in one par-ticular preparation tend to cluster around the diagonal, thussuggesting that the information contents of bursts and isolated

344 L. Cozzi et al.
A B BE I BE+I0
10
20
30
40
50A
vera
ge C
F (
%)
1 Hz2 Hz
a
0.05 0.1 0.2 0.3 0.40.50
5
10
15
x 107
Frequency [Hz]
PS
D (
sens
ory
sign
al)
Sensors1 Hz2 Hz
b
1 20
10
20
30
40
50
Isol
ated
/Tot
al e
vent
s (%
)
Max stimulation rate [Hz]
c
Fig. 7 Effect of an increased rate of stimulation. a Coding performanceof the different firing modes for intensity coding at 1 and 2 Hz. The anal-ysis was performed on the sites for which at least one of the CFs was≥ 30%. b Power spectrum of the sensory signal and its reconstructionfrom intensity coding at 1 and 2 Hz, respectively for the same exper-iment and site of recording, using burst events together with isolatedspikes. c Proportion of evoked isolated spikes over the average numberof evoked events for rates of stimulation of 1 and 2 Hz
spikes are approximately equivalent (see Fig. 9h). However,when the rate of stimulation is increased from 1 to 2 Hz,the information carried by isolated spikes is comparativelygreater than that carried by bursts and burst events (Fig. 9o).This is confirmed by the comparison of the normalized infor-mation rates (bits/spike) (see Figs. 9i, p). While at 1 Hz thedata points mostly lie above the diagonal, meaning that burstevents carry more information than isolated spikes, the oppo-site occurs at 2 Hz.
The coding efficiency of the different firing modes can becompared looking at Fig. 9e. Burst events and isolated spikes(both considered separately or together) provide a higher effi-ciency in coding sensory information than all spikes andbursts. More specifically, we concluded that all spikes andbursts are the least efficient; in contrast, burst events andisolated spikes are the most efficient at both 1 and 2 Hz respec-tively (see Figs. 9e, l, q).
Moreover (see Fig. 9h), since the majority of the pointsin the scatter plots are under the diagonal, isolated spikes aremore efficient in coding information than burst events. Andburst events are more informative than when considering theirwhole structure.
As regards event coding, the main results are summarizedin Fig. 10. The MFR (Fig. 10a) and entropy rates (Fig. 10b)are similar to intensity coding, but the difference between allspikes and bursts with respect to the other spike trains is lessevident. Instead, the rate of transmitted information (in bits/s)is slightly greater for all spikes and bursts (Fig. 10c, h); theopposite occurs for the information per spike (Fig. 10d, i).In terms of coding efficiency, all the spike types are roughlyequivalent with the partial exception of all spikes that resultin lower values (Figs. 10e, l).
3.6 Coding performance and functional connectivity
To assess whether and to what extent the coding performanceis related to features of the observed evoked response, weexamined the relationship between the CF and the descriptiveindicators that we defined for the PSTH. More specifically,we computed the correlation coefficients between the CFsand different parameters of the PSTH (early and late peakamplitude and latency) for each firing mode (see Fig. 11a).
The indicator that correlates most to the CF is the ampli-tude of the early peak of the PSTH, which can be taken as ameasure of functional connectivity between stimulation andrecording sites. More precisely, statistical analysis showedthat the early peak of PSTH does not significantly affect theCF, but there are significant interactions with the firing mode(F(4, 651) = 174.97, p < 0.0001). A closer look showedthat functional connectivity is significantly related to the cod-ing capability for burst events and isolated spikes, both takentogether and separately (p < 0.0001 in all cases).
This observation is illustrated in Fig. 11b, whichshows the relationship between connectivity and coding per-formance for all the recording sites in one epoch of a spe-cific experiment: the scatter plots represent CFs as a functionof PSTH early peak, for each recording electrode. The sitesthat resulted in a good coding performance also have a morereliable evoked response (i.e., a sharp early portion of thePSTH), i.e., a stronger functional connectivity. The trend isparticularly evident for burst events, isolated spikes and theirunion.
Figure 11a also shows that the reconstruction perfor-mance of burst events correlates with the ‘late’ portion of theimpulse response, which reflects long-range, poly-synaptic

Encoding of time-varying stimuli in populations of cultured neurons 345
Sensory signal
Stimuli 65%
All 38%
Bursts 38%
Burst Events 19%
Isolated 1%Isolated + Burst Events
16%
5 s
a
–10 –5 0 5–5
0
5
10
15
Time [s]
ST
A
AllBursts
b
A B BE I BE+I0
10
20
30
40
Ave
rage
CF
(%
)
c
Fig. 8 Coding of isolated events. a From top to bottom sensory signal, its reconstruction from the pattern of stimulation, and its reconstructionfrom one recording site. The numbers on the right indicate the corresponding CFs. b STAs for event coding, for all spikes and for burst spikes. cSummary of the coding performance for the different firing modes. Numbers indicate the respective CFs (%)
connections. This finding suggests an additional differencebetween isolated spikes and bursts: the former cannot prop-agate much further away from the ‘sensory’ regions; in con-trast, bursts contribute to stimulus reconstruction even afterthey have propagated through multiple synapses and over thewhole population.
4 Discussion
We investigated whether and how, in a random population ofcultured cortical neurons, the neural activity can be modu-lated by focal, sustained time-varying stimulation. The exacttiming of the stimuli was determined according to variouscoding schemes and average rates of stimulation, reflectingthe activity of an artificial sensor – the infra-red proximitysensor of a small mobile robot recorded while performingan obstacle avoidance task. In all the experiments, we couldidentify a fraction of the recording electrodes in which a sub-stantial fraction of the sensory signal could be reconstructedfrom the spike trains (CF ≥30%).
4.1 Do bursts play a role in coding for sensory information?
We found that slow variations in the timing of the stimuliare more reliably coded into isolated spikes or burst events.
Moreover, isolated spikes and burst events appear to betriggered by similar stimulus features, but reconstruction per-formance is greatly improved if isolated spikes and burstevents are taken together.
This finding is consistent with the observation (Maeda etal. 1995) of a refractory period following each burst, whenstimulation is in the 5–10 s range (i.e., 0.1–0.5 Hz). This islikely due to inactivation of the inward voltage-dependentionic currents by the long-lasting intracellular plateau poten-tials that occur during bursts, which prevent new burst gen-eration (Robinson et al. 1993). By stimulating at a 1 Hz rate,Maeda et al. (1995) observed absolute refractory periods (noresponse), and relative refractory periods (reduced numberof spikes in the response, associated with faster-decaying,smaller-amplitude synaptic currents).
Similarly, Opitz et al. (2002) found that the duration ofan evoked burst increases with the time between stimulusand the previous spontaneous burst. In other words, burstduration codes for the time from the previous inter-stimulusinterval. This type of neural code would be incompatible withthat based on the timing of evoked events (i.e., either isolatedspikes or burst events), which implies that higher rates ofstimulation result in higher firing rates. In fact, in our exper-iments we found no clear relation between stimulus featuresand burst structure. This would explain why linear recon-struction based on burst events outperforms that which isbased on the whole bursts (see Sect. 3).
346 L. Cozzi et al.
A B BE I BE+I0
2.5
5
MF
R [s
pk/s
]
A B BE I BE+I0
0.2
0.4
0.6
0.8
Info
[bit/
s]
A B BE I BE+I0
5
10
15
20
Ent
ropy
[bit/
s]A B BE I BE+I
0
1
2
3
Info
[bit/
spik
e]
A B BE I BE+I0
0.1
0.2
0.3
0.4
Effi
cien
cy
1 Hz2 Hz
a b c d e
0 5 100
5
10
MFR I [bit/s]
MF
R b
urst
s [s
pk/s
]
0 20 400
20
40
Entropy I [bit/s]
Ent
ropy
bur
sts
[bit/
s]
0.2 0.4 0.60.2
0.4
0.6
Info I [bit/s]
Info
bur
sts
[bit/
s]0 1 2
0
1
2
Info I [bit/spike]
Info
bur
sts
[bit/
spik
e]
0 0.2 0.40
0.2
0.4
Efficiency I
Effi
cien
cy b
urst
s
BEB
f g h i l
0 5 100
5
10
MFR I [spk/s]
MF
R b
urst
s [s
pk/s
]
0 20 400
20
40
Entropy I [bit/s]
Ent
ropy
bur
sts
[bit/
s]
0.2 0.4 0.60.2
0.4
0.6
Info I [bit/s]
Info
bur
sts
[bit/
s]
0 1 20
1
2
Info I [bit/spike]
Info
bur
sts
[bit/
spik
e]
0 0.2 0.40
0.2
0.4
Efficiency I
Effi
cien
cy b
urst
s
BEB
m n o p q
Fig. 9 Coding capability for intensity coding: summary of the average values on all the experiments obtained for: a mean firing rates (MFR), bentropy rates, c,d rates of transmitted information expressed in bits and spikes per second respectively and e coding efficiency for the differentfiring modes. White bars correspond to the maximum stimulation rate of 1 Hz, whereas black bars report data obtained at 2 Hz. f–q Scatter plotsrepresenting f, m MFR, g, n entropy rates, h,i–o,p rates of transmitted information expressed in bits per second and spikes per second and l,qcoding efficiency respectively, obtained from isolated spikes (x-axis) and bursts (y-axis) for different recording sites, in one particular experiment.Panels f–l refer to stimulation at maximum rate of 1 Hz, while m–q were obtained by stimulating at 2 Hz
To test the ability of the neuronal culture to code for theoccurrence of isolated ‘events’, we used an alternative codingscheme (‘event’ coding), in which single stimuli are elicitedby abrupt increases of the sensory signal. This coding schemediffers from ‘intensity’ coding essentially for the lower rateof stimulation, and for that the aim of linear reconstruction isnow to recover the pattern of stimulation itself, not its instan-taneous rate. In contrast to ‘intensity’ coding, in this casethe best reconstruction performance was achieved with all thespikes, and with the spikes belonging to bursts. Instead, theworst performance was obtained with isolated spikes. Theseresults suggest that bursts are good at coding for the times ofoccurrence of relatively rare ‘events’. This conclusion is notin contrast to what we found for ‘intensity’ coding. In eventcoding, the rate of stimulation is low in comparison to thetime constants of refractoriness phenomena; in other words,the neuronal population manages to keep the pace of the stim-ulation. In fact, much less isolated spikes are observed in thiscase.
4.2 Coding performance is limited by the affordablerate of stimulation
Spectral analysis demonstrated that only very low frequencycomponents (≤ 0.2 Hz) of the original sensory signal couldbe reconstructed from the spike trains. This is partly due tothe low rate of stimulation that we used. An increased rateof stimulation would have allowed to capture more detailsof the sensory signal (Gabbiani and Koch 1996; Cozzi etal. 2005). In fact, the reconstruction performance from thetrains of stimuli is 77% at 1 Hz and 88% at 2 Hz, meaningthat coding implies a loss of information.
In our experiments, the reconstruction performance fromisolated spikes is significantly improved when the rate ofstimulation is increased; the same happens for reconstruc-tion from isolated spikes and burst events, taken together.However, no significant improvement is observed in thereconstruction from burst events alone; and, the reconstruc-tion performance of bursts even decreases.

Encoding of time-varying stimuli in populations of cultured neurons 347
A B BE I BE+I0
2.5
5
MF
R [s
pk/s
]
A B BE I BE+I0
0.2
0.4
0.6
0.8
Info
[bit/
s]
A B BE I BE+I0
5
10
15
20
Ent
ropy
[bit/
s]A B BE I BE+I
0
1
2
3
Info
[bit/
spik
e]
A B BE I BE+I0
0.1
0.2
0.3
0.4
Effi
cien
cy
a b c d e
0 5 100
5
10
MFR I [spk/s]
MF
R b
urst
s [s
pk/s
]
0 20 400
20
40
Entropy I [bit/s]
Ent
ropy
bur
sts
[bit/
s]
0.2 0.4 0.60.2
0.4
0.6
Info I [bit/s]
Info
bur
sts
[bit/
s]0 1 2
0
1
2
Info I [bit/spike]
Info
bur
sts
[bit/
spik
e]
0 0.2 0.40
0.2
0.4
Efficiency I
Effi
cien
cy b
urst
s
BEB
f g h i l
Fig. 10 Coding capability for event coding. a–e Summary of the average values on all the experiments, obtained for: (a) MFR, (b) entropy rates,(c,d) rates of transmitted information expressed in bits per second and spikes per second and (e) coding efficiency respectively, for the differentfiring modes. f–l Scatter plots representing (f) MFR, (g) entropy rates, (h,i) rates of transmitted information expressed in bits per second andspikes per second and (l) coding efficiency respectively, obtained from isolated spikes (x-axis) and bursts (y-axis) for different recording sites, inone particular experiment
LAT. AMPL. LAT. AMPL.0
0.1
0.2
0.3
0.4
0.5
Early peak Late peakPSTH shape
Cor
rela
tion
with
CF
BEII+BE
a
0 100 2000
50
100
CF
(%
)
A
0 100 200
B
0 100 200
PSTH early peak [spk/s]
BE
0 100 2000
50
100 I
0 100 200
BE+I
b
Fig. 11 Coding performance depends on functional connectivity. a Average correlation coefficients between various indicators of PSTH shape(the peak amplitude and latency of the early and late component of the evoked response respectively) and the corresponding CF, for burst events,isolated spikes and burst events together with isolated spikes. b Scatter plots representing the dependence of the CF on the corresponding mag-nitude of the early PSTH peak (i.e., the indicator that best correlates with the CF), for one typical experimental session. Each point represents adifferent recording electrode. Each plot refers to a different firing mode. A trend between CF and PSTH early peak can be identified
We also found that an increased rate of stimulation resultsin a significant increase in the ratio of evoked isolated spikesover the total number of evoked events. In other words, athigher rates of stimulation, stimuli are more likely to evokeisolated spikes than bursts.
This result is consistent with the already mentioned find-ings of Maeda et al. (1995) that when increasing the rate ofstimulation the evoked response is more often made of fewisolated spikes; and with those of Wagenaar et al. (2005), whofound that sustained stimulation seems to induce a change inthe dynamic regime, resulting in the disruption of bursts.
In our experiments, we used low stimulus frequencies(up to 2 Hz), similar to that used by Maeda et al. (1995). Theprevious considerations would suggest that coding accuracy
should benefit from increasing the stimulation rate. How-ever, this is the maximum that these preparations can affordwith this type of electrodes without inducing desensitization(Wagenaar et al. 2004). This limitation imposes an upperbound on the coding performance, in case of focal stimu-lation from a single site. Higher rates of stimulation (up to20 Hz) have been applied in some studies (Wagenaar et al.2005), by distributing stimuli over multiple electrodes.
4.3 The ability to code sensory information is relatedto functional connectivity
We found a clear relationship between coding performanceand the shape and magnitude of the impulse response. More

348 L. Cozzi et al.
specifically, in the recording sites that code sensory infor-mation more accurately – i.e., those from which the sensorysignal can be reconstructed more reliably – the ‘early’ portionof the response to electrical stimulation tends to be greater.In fact, a significant correlation was observed between theCF for isolated spikes and burst events and the magnitude ofthe ‘early’ portion of the evoked response.
As the latter can be taken as a measure of functional con-nectivity between stimulation and recording sites, this findingsuggests that functional connectivity is a major determinantof the ability to ‘code’ sensory information by isolated spikes,burst events and their combination.
However, the reconstruction performance of burst eventsalso correlates to the ‘late’ portion of the impulse response,which reflects long-range, poly-synaptic connections. Thisfinding suggests an additional difference between isolatedspikes and bursts: the former cannot propagate much furtheraway from the ‘sensory’ regions; in contrast, bursts may bemore reliable at propagating through multiple synapses andover the whole population. This is consistent with the sug-gestion of Lisman (1997) that bursts may play an importantrole in information processing, as they are transmitted morereliably through some synapses that filter out isolated spikes.
4.4 Relation to in vivo studies
There is a substantial body of research on the role of burstsin coding for sensory information in an intact nervous sys-tem. The electro-sensory system of the weakly electric fishand the relay neurons of the mammalian thalamus have beenused as model systems – see Krahe et al. (2002) for a review– to characterize the roles of the different types of neurons,and the relative contributions of bursting and non-burstingfiring modes to the encoding of sensory signals.
Overall, these findings suggest that bursts code specific,behaviorally relevant events (Gabbiani et al. 1996; Metzneret al. 1998; Krahe et al. 2002). In particular, Metzner et al.(1998) showed that in pyramidal cells of the electro-sensorysystem of the weak electric fish, spikes as parts of burstscarried the most reliable information about the presence ofup- and down-strokes in the stimulus waveform. In the samepreparation, Oswald et al. (2004) found that bursts and iso-lated spikes represent different frequency components of thesensory signal. More precisely, they showed that within asingle spike train, bursts are biased toward low-frequencyevents, while isolated spikes simultaneously code for the en-tire frequency range.
In experiments performed on the dorsal LGN cells ofanesthetized cats (Reinagel et al. 1999), linear stimulusreconstruction methods were applied to both bursts and tonicspikes to assess whether sensory signals are coded in theprecise time structure of spikes within bursts. The main find-ings were that there are no relevant differences between theaverage sensory stimuli which optimally trigger isolated andburst spikes. Moreover, isolated spikes carried about twiceas much information per action potential as bursts, but bursts
as unitary events encoded about three times more informa-tion per event than tonic spikes. Along the same line, LGNbursts in awake monkeys were shown to be correlated moreclosely than isolated spikes with preceding microsaccades(Martinez-Conde et al. 2002).
The notion that the biophysical mechanisms of spike gen-eration enable individual neurons to encode different stim-ulus features into distinct spike patterns was also confirmedby modeling studies (Kepecs et al. 2002; Kepecs and Lisman2003), which suggested that bursts code for positive slopesof the stimulus, and burst duration codes for the magnitudeof such slope. Moreover, Sherman (2001) proposed that inthalamic relay cells, feedback from the cortex could switchbetween two coding ‘states’, i.e., signal detection and stim-ulus estimation.
We may ask how relevant are our results to these in vivoobservations. Cultured neurons are fundamentally differentfrom in vivo preparations in terms of anatomy and synapticconnections. Moreover, in our experiments, we used focal,low-frequency stimulation which is clearly not a realistic sen-sory input, and the rate of stimulation (maximum of 2 Hz inour experiments) is low in comparison to the typical firingrates found in primary sensory areas. Finally, while in intactnervous systems, sensory afferences distribute over extendedportions of it, and different neurons possibly code for differ-ent stimulus features, in cultured neurons the connectivity israndom and there is no functional specialization.
Nevertheless, the results presented here are qualitativelysimilar to the in vivo findings of Oswald et al. (2004) in thatbursts code for relatively rare events, whereas isolated spikescode for slow variations of the rate of stimulation.
However, in intact nervous systems, bursts are evoked byparticular stimulus features reflected in specific pre-synapticpatterns of activity. In contrast, in our preparation, focal, low-frequency stimulation is likely to be perceived by individualneurons as an abrupt increase in the membrane current, a con-dition which would optimally elicit a burst. In other words,bursts would be, and in fact are, the main modes of responseto focal stimulation. However, when the inter-stimulus inter-val is decreased, the global level of excitation increases;bursts tend to become shorter, and isolated evoked spikesalso appear. Therefore, in cultured neurons, the different fir-ing modes do not reflect different features of the input signalbut rather the current level of global excitation.
In spite of these differences, our experiments suggest thatmany of the in vivo findings are very general properties ofneuronal population, irrespective of their micro-architectureand/or anatomical organization.
4.5 Neural populations as computing devices
It has been suggested that cultured neurons may be relevantin investigating the structure and the emergence of distrib-uted representations of sensorimotor information in an intactbrain. In spite of the absence of the structured morphologyand regional specialization found in an intact nervous system,

Encoding of time-varying stimuli in populations of cultured neurons 349
which are shaped by evolution to process sensory informa-tion, our results show that cultured neurons maintain somecapability to encode time-varying stimuli. This is consistentwith the view of random populations of cultured neurons asgeneral ‘computing’ devices. In fact, in a number of experi-ments, neuronal populations in vitro have been connected toeither a ‘virtual’, computer-simulated environment (DeMarseet al. 2001), or to a robotic body (Reger et al. 2000; Martinoiaet al. 2004).
If the goal is to establish an input/output mapping throughthe neural preparation, our results would point to (1) the needto look at events (burst events or isolated spikes), and (2) theneed to carefully select recording sites that are reliably func-tionally connected to the site of stimulation.
Acknowledgements This work is partly supported by the EU grantIST-2001-33564 (Neurobit), and by a grant from the Italian Ministry ofUniversity, Education and Research. The authors wish to thank Dr. M.Tedesco for cell culture preparation, and Prof. S. Martinoia for usefuldiscussions.
References
Bakkum DJ, Shkolnik AC, Ben-Ary G, Gamblen P, DeMarse TB, PotterSM (2004) Removing some ’A’ from AI: embodied cultured net-works. Embodied Artif Intell 3139:130–145
Bi G, Poo M (1999) Distributed synaptic modification in neural net-works induced by patterned stimulation. Nature 401:792–796
Bialek W, Rieke F, de Ruyter van Steveninck RR, Warland D (1991)Reading a neural code. Science 252:1854–1857
Braitenberg V (1984) Vehicles, experiments in synthetic psychology.MIT Press, Cambridge, Mass
Chiappalone M, Vato A, Tedesco MB, Marcoli M, Davide F, MartinoiaS (2003) Networks of neurons coupled to microelectrode arrays: aneuronal sensory system for pharmacological applications. BiosensBioelectron 18:627–634
Cozzi L, D‘Angelo P, Chiappalone M, Ide AN, Novellino A, Marti-noia S, Sanguineti V (2005) Coding and decoding of information ina bi-directional neural interface. Neurocomputing 65(66):783–792
DeMarse TB, Wagenaar DA, Blau AW, Potter SM (2001) The neural-ly controlled animat: biological brains acting with simulated bodies.Auton Robots 11:305–310
Gabbiani F (1996) Coding of time-varying signals in spike trains oflinear and half-wave rectifying neurons. Netw Comput Neural Syst7:61–85
Gabbiani F, Koch C (1996) Coding of time-varying signals in spiketrains of integrate-and-fire neurons with random threshold. NeuralComput 8:44–66
Gabbiani F, Metzner W, Wessel R, Koch C (1996) From stimulus encod-ing to feature extraction in weakly electric fish. Nature 384:564–567
Huettner JE, Baughman RW (1986) Primary culture of identified neu-rons from the visual cortex of postnatal rats. J Neurosci 6:3044–3060
Jimbo Y, Tateno T, Robinson HP (1999) Simultaneous induction ofpathway-specific potentiation and depression in networks of corticalneurons. Biophys J 76:670–678
Jimbo Y, Kawana A, Parodi P, Torre V (2000) The dynamics of a neu-ronal culture of dissociated cortical neurons of neonatal rats. BiolCybern 83:1–20
Kepecs A, Lisman J (2003) Information encoding and computation withspikes and bursts. Network 14:103–118
Kepecs A, Wang XJ, Lisman J (2002) Bursting neurons signal inputslope. J Neurosci 22:9053–9062
Krahe R, Gabbiani F (2004) Burst firing in sensory systems. Nat RevNeurosci 5:13–23
Krahe R, Kreiman G, Gabbiani F, Koch C, Metzner W (2002) Stimu-lus encoding and feature extraction by multiple sensory neurons. JNeurosci 22:2374–2382
Kriegstein AR, Dichter MA (1983) Morphological classification of ratcortical neurons in cell culture. J Neurosci 3:1634–1647
Latham PE, Richmond BJ, Nelson PG, Nirenberg S (2000a) Intrinsicdynamics in neuronal networks. I. Theory. J Neurophysiol 83:808–827
Latham PE, Richmond BJ, Nirenberg S, Nelson PG (2000b) Intrin-sic dynamics in neuronal networks. II. Experiment. J Neurophysiol83:828–835
Lisman JE (1997) Bursts as a unit of neural information: making unre-liable synapses reliable. Trends Neurosci 20:38–43
Maeda E, Robinson HP, Kawana A (1995) The mechanisms of genera-tion and propagation of synchronized bursting in developing networksof cortical neurons. J Neurosci 15:6834–6845
Maeda E, Kuroda Y, Robinson HP, Kawana A (1998) Modification ofparallel activity elicited by propagating bursts in developing networksof rat cortical neurones. Eur J Neurosci 10:488–496
Marom S, Shahaf G (2002) Development, learning and memory in largerandom networks of cortical neurons: lessons beyond anatomy. Q RevBiophys 35:63–87
Martinez-Conde S, Macknik SL, Hubel DH (2002) The function ofbursts of spikes during visual fixation in the awake primate lateralgeniculate nucleus and primary visual cortex. Proc Natl Acad SciUSA 99:13920–13925
Martinoia S, Sanguineti V, Cozzi L, Berdondini L, van Pelt J, Tomas J,Le Masson G, Davide F (2004) Towards an embodied in vitro elec-trophysiology: the NeuroBIT project. Neurocomputing 58(60):1065–1072
Metzner W, Koch C, Wessel R, Gabbiani F (1998) Feature extractionby burst-like spike patterns in multiple sensory maps. J Neurosci18:2283–2300
Novellino A, Chiappalone M, Vato A, Bove M, Tedesco MT, MartinoiaS (2003) Behaviors from an electrically stimulated spinal cord neu-ronal network cultured on microelectrode arrays. Neurocomputing52(54C):661–669
Opitz T, De Lima AD, Voigt T (2002) Spontaneous development of syn-chronous oscillatory activity during maturation of cortical networksin vitro. J Neurophysiol 88:2196–2206
Oswald AM, Chacron MJ, Doiron B, Bastian J, Maler L (2004) Parallelprocessing of sensory input by bursts and isolated spikes. J Neurosci24:4351–4362
Potter SM, DeMarse TB (2001) A new approach to neural cell culturefor long-term studies. J Neurosci Methods 110:17–24
Reger BD, Fleming KM, Sanguineti V, Alford S, Mussa-Ivaldi FA(2000) Connecting brains to robots: an artificial body for studyingthe computational properties of neural tissues. Artif Life 6:307–324
Reinagel P, Godwin D, Sherman SM, Koch C (1999) Encoding of visualinformation by LGN bursts. J Neurophysiol 81:2558–2569
Rieke F, Warland D, van Steveninck RdR, Bialek W (1997) Spikes:exploring the neural code. The MIT Press, Cambridge, Massachu-setts
Robinson HP, Kawahara M, Jimbo Y, Torimitsu K, Kuroda Y, KawanaA (1993) Periodic synchronized bursting and intracellular calciumtransients elicited by low magnesium in cultured cortical neurons. JNeurophysiol 70:1606–1616
Sherman SM (2001) Tonic and burst firing: dual modes of thalamocor-tical relay. Trends Neurosci 24:122–126
van Pelt J, Wolters PS, Corner MA, Rutten WL, Ramakers GJ (2004)Long-term characterization of firing dynamics of spontaneous burstsin cultured neural networks. IEEE Trans Biomed Eng 51:2051–2062
Wagenaar DA, Pine J, Potter SM (2004) Effective parameters for stimu-lation of dissociated cultures using multi-electrode arrays. J NeurosciMethods 138:27–37
Wagenaar DA, Madhavan R, Pine J, Potter SM (2005) Controlling burst-ing in cortical cultures with closed-loop multi-electrode stimulation.J Neurosci 25:680–688
Wiener N (1949) Extrapolation, interpolation and smoothing of timeseries. Wiley, New York




















