
Identification and functional expression of a second human ��-galactoside...
12
Identification and functional expression of a second human b-galactoside a2,6-sialyltransferase, ST6Gal II Marie-Ange Krzewinski-Recchi 1 , Sylvain Julien 1 , Sylvie Juliant 2 , Me ´ lanie Teintenier-Lelie ` vre 1 , Be ´ ne ´ dicte Samyn-Petit 1 , Maria-Dolores Montiel 1 , Anne-Marie Mir 1 , Martine Cerutti 2 , Anne Harduin-Lepers 1 and Philippe Delannoy 1 1 Unite´ de Glycobiologie Structurale et Fonctionnelle, UMR CNRS – USTL 8576, Universite´ des Sciences et Technologies de Lille, F-59655 Villeneuve d’Ascq, France; 2 Station de Recherche de Pathologie Compare ´e, UMR CNRS – INRA 5087, F-30380 Saint Christol-lez-Ale`s, France BLAST analysis of the human and mouse genome sequence databases using the sequence of the human CMP-sialic acid: b-galactoside a-2,6-sialyltransferase cDNA (hST6Gal I, EC2.4.99.1) as a probe allowed us to identify a putative sialyltransferase gene on chromosome 2. The sequence of the corresponding cDNA was also found as an expressed sequence tag of human brain. This gene contained a 1590 bp open reading frame divided in five exons and the deduced amino-acid sequence didn’t correspond to any sialyltransferase already known in other species. Multiple sequence alignment and subsequent phylogenic analysis showed that this new enzyme belonged to the ST6Gal subfamily and shared 48% identity with hST6Gal-I. Consequently, we named this new sialyltransferase ST6Gal II. A construction in pFlag vector transfected in COS-7 cells gave raise to a soluble active form of ST6Gal II. Enzymatic assays indicate that the best acceptor substrate of ST6Gal II was the free disaccharide Galb1–4GlcNAc structure whereas ST6Gal I preferred Galb1–4GlcNAc-R disaccharide sequence linked to a protein. The a2,6-linkage was confirmed by the increase of Sambucus nigra agglutinin- lectin binding to the cell surface of CHO transfected with the cDNA encoding ST6Gal II and by specific sialidases treatment. In addition, the ST6Gal II gene showed a very tissue specific pattern of expression because it was found essentially in brain whereas ST6Gal I gene is ubiquitously expressed. Keywords: human; b-galactoside a2,6-sialyltransferase; molecular cloning. Sialylated sugar chains are present at the cell surface of various animal species. Due to their position, they are thought to serve important roles in a large variety of biological functions such as cell–cell and cell–substrate interactions, bacterial and virus adhesion, and protein targeting [1,2]. Sialylated glycoconjugates exhibit remark- ably diverse structures [3–5] and their expression has been shown to change during development [6], differentiation, disease and oncogenic transformation [7]. In mammals, sialic acids are found at the nonreducing terminal position of glycoconjugates sugar chains, a2,3- or a2,6-linked to a b-D-galactopyranosyl (Gal) residue, or a2,6-linked to a b-D-N-acetylgalactosaminyl (GalNAc) or a b-D-N-acetyl- glucosaminyl (GlcNAc) residue. Sialic acids are also found a2,8-linked to sialic acid residues in gangliosides and in polysialic acid, a linear a2,8-homopolymer observed on several glycoproteins including the neural cell adhesion molecule N-CAM [8]. In addition, a2,6-linked sialic acid is also present in free oligosaccharides such as 6¢-sialyllactose from human milk [9], monosialylganglioside of the human meconium [10]; Neu5Aca2–6GalNAcb1–4GlcNAc-R sequence has been described as a terminal sequence of the N-glycans of pituitary hormones [11]. The biosynthesis of sialylated oligosaccharides is cata- lysed by a family of enzymes named sialyltransferases [3,12]. These enzymes are a subset of the glycosyltransferases family (family 29 in the CAZy database [13]) that use CMP- Neu5Ac as the activated sugar donor to catalyse the transfer of sialic acid residues to the terminal position of oligosac- charide chains of glycolipids and glycoproteins. Sialyltrans- ferases are a family of type II membrane-bound glycoproteins with a short NH 2 -terminal cytoplasmic tail, a 16–20 amino acid signal anchor domain that is involved with retention of the protein in the Golgi apparatus, a stem region, highly variable in length (from 20 amino acids to 200 amino acids), ending with a large COOH-terminal catalytic domain that resides in the Golgi lumen [14]. The catalytic domain contains three highly conserved amino-acid sequences termed sialylmotifs L (large), S (small), and VS (very small). Correspondence to M.-A. Krzewinski-Recchi, Unite´ de Glycobiologie Structurale et Fonctionnelle, UMR CNRS no. 8576, Laboratoire de Chimie Biologique, Universite´ des Sciences et Technologies de Lille, F-59655 Villeneuve d’Ascq, France. Fax: + 33 320 43 65 55, Tel.: + 33 320 43 69 23, E-mail: [email protected] Abbreviations: EST, expressed sequence tag; Gal, b-D-galactopyrano- syl residue; GalNAc, b-D-N-acetylgalactosaminyl; N-CAM, neural cell adhesion molecule; EGT, ecdysone-S-glycosyltransferase. Enzyme: CMP-sialic acid:b-galactoside a-2,6-sialyltransferase cDNA (hST6Gal I, EC2.4.99.1). Note: nucleotide sequence data are available in the DDBJ/EMBL/ GenBank databases under the accession number AJ512141. (Received 5 November 2002, revised 7 January 2003, accepted 10 January 2003) Eur. J. Biochem. 270, 950–961 (2003) ȑ FEBS 2003 doi:10.1046/j.1432-1033.2003.03458.x
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Identification and functional expression of a second human ��-galactoside...

Identification and functional expression of a second humanb-galactoside a2,6-sialyltransferase, ST6Gal II
Marie-Ange Krzewinski-Recchi1, Sylvain Julien1, Sylvie Juliant2, Melanie Teintenier-Lelievre1,Benedicte Samyn-Petit1, Maria-Dolores Montiel1, Anne-Marie Mir1, Martine Cerutti2,Anne Harduin-Lepers1 and Philippe Delannoy1
1Unite de Glycobiologie Structurale et Fonctionnelle, UMR CNRS – USTL 8576, Universite des Sciences et Technologies de Lille,
F-59655 Villeneuve d’Ascq, France; 2Station de Recherche de Pathologie Comparee, UMR CNRS – INRA 5087,
F-30380 Saint Christol-lez-Ales, France
BLAST analysis of the human and mouse genome sequencedatabases using the sequence of the human CMP-sialicacid:b-galactoside a-2,6-sialyltransferase cDNA (hST6Gal I,EC2.4.99.1) as a probe allowed us to identify a putativesialyltransferase gene on chromosome 2. The sequence ofthe corresponding cDNA was also found as an expressedsequence tag of human brain. This gene contained a1590 bp open reading frame divided in five exons and thededuced amino-acid sequence didn’t correspond to anysialyltransferase already known in other species. Multiplesequence alignment and subsequent phylogenic analysisshowed that this new enzyme belonged to the ST6Galsubfamily and shared 48% identity with hST6Gal-I.Consequently, we named this new sialyltransferaseST6Gal II. A construction in pFlag vector transfected in
COS-7 cells gave raise to a soluble active form of ST6Gal II.Enzymatic assays indicate that the best acceptor substrate ofST6Gal II was the free disaccharide Galb1–4GlcNAcstructure whereas ST6Gal I preferred Galb1–4GlcNAc-Rdisaccharide sequence linked to a protein. The a2,6-linkagewas confirmed by the increase of Sambucus nigra agglutinin-lectin binding to the cell surface of CHO transfected withthe cDNA encoding ST6Gal II and by specific sialidasestreatment. In addition, the ST6Gal II gene showed a verytissue specific pattern of expression because it was foundessentially in brain whereas ST6Gal I gene is ubiquitouslyexpressed.
Keywords: human; b-galactoside a2,6-sialyltransferase;molecular cloning.
Sialylated sugar chains are present at the cell surface ofvarious animal species. Due to their position, they arethought to serve important roles in a large variety ofbiological functions such as cell–cell and cell–substrateinteractions, bacterial and virus adhesion, and proteintargeting [1,2]. Sialylated glycoconjugates exhibit remark-ably diverse structures [3–5] and their expression has beenshown to change during development [6], differentiation,disease and oncogenic transformation [7]. In mammals,sialic acids are found at the nonreducing terminal positionof glycoconjugates sugar chains, a2,3- or a2,6-linked to a
b-D-galactopyranosyl (Gal) residue, or a2,6-linked to ab-D-N-acetylgalactosaminyl (GalNAc) or a b-D-N-acetyl-glucosaminyl (GlcNAc) residue. Sialic acids are also founda2,8-linked to sialic acid residues in gangliosides and inpolysialic acid, a linear a2,8-homopolymer observed onseveral glycoproteins including the neural cell adhesionmolecule N-CAM [8]. In addition, a2,6-linked sialic acid isalso present in free oligosaccharides such as 6¢-sialyllactosefrom human milk [9], monosialylganglioside of the humanmeconium [10]; Neu5Aca2–6GalNAcb1–4GlcNAc-Rsequence has been described as a terminal sequence of theN-glycans of pituitary hormones [11].
The biosynthesis of sialylated oligosaccharides is cata-lysed by a family of enzymes named sialyltransferases [3,12].These enzymes are a subset of the glycosyltransferasesfamily (family 29 in the CAZy database [13]) that use CMP-Neu5Ac as the activated sugar donor to catalyse the transferof sialic acid residues to the terminal position of oligosac-charide chains of glycolipids and glycoproteins. Sialyltrans-ferases are a family of type IImembrane-bound glycoproteinswith a short NH2-terminal cytoplasmic tail, a 16–20 aminoacid signal anchor domain that is involved with retention ofthe protein in the Golgi apparatus, a stem region, highlyvariable in length (from 20 amino acids to 200 amino acids),ending with a large COOH-terminal catalytic domain thatresides in the Golgi lumen [14]. The catalytic domaincontains three highly conserved amino-acid sequencestermed sialylmotifs L (large), S (small), and VS (very small).
Correspondence to M.-A. Krzewinski-Recchi, Unite de Glycobiologie
Structurale et Fonctionnelle, UMR CNRS no. 8576, Laboratoire de
Chimie Biologique, Universite des Sciences et Technologies de Lille,
F-59655 Villeneuve d’Ascq, France.
Fax: + 33 320 43 65 55, Tel.: + 33 320 43 69 23,
E-mail: [email protected]
Abbreviations: EST, expressed sequence tag; Gal, b-D-galactopyrano-syl residue; GalNAc, b-D-N-acetylgalactosaminyl; N-CAM, neural
cell adhesion molecule; EGT, ecdysone-S-glycosyltransferase.
Enzyme: CMP-sialic acid:b-galactoside a-2,6-sialyltransferase cDNA
(hST6Gal I, EC2.4.99.1).
Note: nucleotide sequence data are available in the DDBJ/EMBL/
GenBank databases under the accession number AJ512141.
(Received 5 November 2002, revised 7 January 2003,
accepted 10 January 2003)
Eur. J. Biochem. 270, 950–961 (2003) � FEBS 2003 doi:10.1046/j.1432-1033.2003.03458.x

Sialylmotifs L and S are involved in the binding of donorand acceptor substrates, respectively [15,16], whereas thesialylmotif VS is involved in the catalytic process [17].
To date, 19 different sialyltransferases have been identi-fied in mouse and humans but only one of these enzymes,ST6Gal I (CMP-sialic acid:b-galactoside a2,6-sialyltrans-ferase, EC 2.4.99.1) is known to mediate the transfer of asialic acid residue in a2,6-linkage to the galactose residue ofthe type 2 disaccharide (Galb1–4GlcNAc) found as a freedisaccharide or as a terminal N-acetyllactosamine unit ofN- and O-glycans. However, as reviewed previously [3],ST6Gal I has been shown in in vitro assays to have a lowactivity for transferring sialic acid onto other oligosac-charide structures, such as lactose (Galb1–4Glc), type 1disaccharide structure (Galb1–3GlcNAc) [18] or type 2structure GalNAcb1–4GlcNAc [19,20] but not ontotype 3 structure (Galb1–3GalNAc). ST6Gal I has beenpurified to homogeneity from animal livers and hepatomacells (reviewed in [3]) and cDNA has been cloned from ratliver [21], human placenta [22], mouse liver [23], bovinetissues [20] and chick embryo [24]. Several mRNA isoformsare generated from a unique gene encoding ST6Gal Ithrough the use of physically distinct promoters. Thesetranscripts differ only in their 5¢-untranslated region andshare an identical ST6Gal I coding region. These transcriptsare expressed in a tissue-specific manner and contribute tothe regulation of a2,6-sialylation in tissue and cells duringcell differentiation [25,26], inflammation [27] and oncogenictransformation [28,29].
BLAST analysis of the mouse and human genomedatabases allowed us to identify an unknown sialyltrans-ferase gene encoding a second Galb1–4GlcNAc a2,6-sialyltranferase that has been named hST6Gal II. In thisreport, we describe the functional analysis of a recombinanthuman ST6Gal II, which shows slightly different substratespecificity than ST6Gal I. The expression pattern of thegene was also examined in various human tissues and foundto be very restricted, mainly to brain and fetal tissues.
Materials and methods
Materials
CMP-[14C]Neu5Ac (10.7 GBqÆmmol)1), Redivue stabilized[a-32P]dCTP (110 TBqÆmmol)1) and Rediprime II DNAlabelling system were from Amersham Pharmacia Biotech(Little Chalfont, UK). The DyNazyme EXT DNA poly-merase was from Ozyme (Saint Quentin en Yvelines,France). Oligonucleotides were synthesized by Eurogentec(Seraing, Belgium). Dulbecco’s modified Eagle’s medium(DMEM) containing 4.5 gÆL)1 glucose and lacking gluta-mine was from BioWhittaker Europe. Alpha Eagle’sminimal essential medium (aMEM), OPTIMEM, L-gluta-mine and antibiotics used in cell culture were from GibcoBRL (Cergy-Pontoise, France). Fetal bovine serum wasfromD. Dustscher (Issy-les-Moulineaux, France). Lipofect-AMINE PLUS Reagent was from Invitrogen. The block-ing reagent and fluorescein labelled anti-digoxigenin Fabfragments and DOTAP transfection reagent were fromRoche (Meylan, France). a1-Acid glycoprotein, fetuin,Gala-O-pNp, GalNAca-O-pNp, GlcNAca-O-pNp,Galb1–3GalNAca-O-benzyl, Galb1–3GlcNAc, Galb1–
4GlcNAc, the expression vector pFlag-CMV-1, the mono-clonal antibody (mAb) anti-Flag BioM2, alkaline phospha-tase conjugated goat anti-[mouse IgG (Fab specific)] Ig andCMP-Agarose beads were from Sigma (St Louis, MO,USA). Lacto-N-neotetraose and Lacto-N-tetraose were thegenerous gift of G. Strecker and F. Chirat (UMR CNRS8576, Villeneuve d’Ascq, France). Galb1–3GlcNAcb-O-octyl and Galb1–4GlcNAcb-O-octyl were the generous giftof C. Auge (URA CNRS 462, Orsay, France). MTNTM
multiple tissue Northern blot and MTETM multiple tissueexpression arrays were from Clontech (Palo Alto, CA,USA). Sialic Acid Linkage Analysis Kit was from GlykoInc. (Novato, CA, USA). The expressed sequence tag (EST)clone (GenBank/EBI accession number AB058780) was agenerous gift from T. Nagase, KAZUSA DNA ResearchInstitute, Chiba, Japan.
Preparation of asialo-glycoproteins
Fetuin and a1-acid glycoprotein (10 mgÆmL)1) were incu-bated for 1 h in 0.05 M sulfuric acid at 80 �C. The asialo-products were then neutralized, dialyzed and lyophilizedprior to use. The carbohydrate content of asialo-glycopro-teins was analysed by GC-MS [30].
Construction of expression vectors of hST6Gal IIand transfections
A truncated form of hST6Gal II lacking the first 33 aminoacids of the open reading frame, was generated by PCRamplification using a 5¢ primer containing an EcoRI site,5¢-CCGACAGGAATTCCGCTGAGCCTGTACCCAGCTCCC-3¢ (nucleotides 91–127, Fig. 1A) and 3¢ primercontaining a BamHI site, 5¢-ACATTGGATCCCAAGAAACCCTTTTTAAGAGTGTGG-3¢ (nucleotides 1577–1614, Fig. 1A). A full-length open reading frame ofST6Gal II was also prepared by PCR amplification usinga 5¢ primer containing an EcoRI site, 5¢-CCCTCTGA
ATTCAGACACAAGGTGCTGACCGCAGAG-3¢ (nuc-leotide 1–35, Fig. 1A) and the 3¢ primer described above.The 25 lL of PCR mixture consisted of 1 unit ofDyNazyme EXT, 0.3 lM of each primer, 0.2 mM dNTPand 1.5 ng of plasmid DNA. Reactions were run using thefollowing conditions: 1 min at 96 �C, 4 min at 72 �C for 40cycles. Two amplification fragments of 1656 bp and1522 bp, respectively, were obtained and subcloned in theTopo TA cloning vector (Invitrogen, USA). The insertedfragments were cut out by digestion withBamHI andEcoRIand inserted into BamHI and EcoRI sites of the pFlag-CMV-1 expression vector, which contains the preprotrypsinleader sequence. Restriction enzymes digestions and DNAsequencing by Genoscreen (Lille, France) confirmed thecDNA sequence and the insert junctions. The resultingplasmids encoded a soluble fusion protein consisting of theFlag sequence and a truncated form of ST6Gal II (pFlag-sST6Gal-II) or the whole coding region of ST6Gal II(pFlag-wST6Gal-II). COS-7 cells were grown in DMEMwith 4.5 gÆL)1 glucose without glutamine supplementedwith 10% fetal bovine serum, L-glutamine 20 mM, penicillin,streptomycin at 37 �C under 5% CO2.
Twelve micrograms of Qiagen-purified pFlag-sST6Gal-IIor pFlag plasmids were transiently transfected into COS-7
� FEBS 2003 Characterization of the human ST6Gal II (Eur. J. Biochem. 270) 951
952 M.-A. Krzewinski-Recchi et al. (Eur. J. Biochem. 270) � FEBS 2003

cells in a 100-mm diameter dish using LipofectAMINEPLUS reagent, following the manufacturer’s instructions.The medium was harvested 48 h after transfection. Theenzymatic protein expressed in the medium was used as theenzyme source.
Western blot analysis of a soluble ST6Gal II
Nine milliliters of media from COS-7 cells transfected withthe expression plasmid pFlag-sST6Gal II and from mock-transfected cells were concentrated into 2 mL on Macrosep30K and 1.5 mL of this preparation were incubated with150 lL of CMP-Agarose beads (2.8 lmolÆmL)1 CMP).After washing, the supernatants were discarded and beadswere boiled for 5 min in 100 lL SDS/PAGE loading buffer,centrifuged and loaded on a 4–20% gradient polyacryl-amidegel under reducing conditions.AfterWesternblotting,the nitrocellulosemembranewas incubatedwith 10 lgÆmL)1
anti-Flag BioM2 mAb. Alkaline phosphatase-labelled goatanti-(mouse IgG) Ig was used as the second antibody andrevealed by Nitro Blue tetrazolium/5-bromo-4-chloro-3-indolyl-phosphate and X-phosphate staining.
Confocal microscopy
CHO cells were grown in aMEM with glutamax, supple-mented with 10% fetal bovine serum, penicillin andstreptomycin at 34 �C under 5% CO2. CHO and COS-7cells were transiently transfected with the pFlag plasmid orpFlag plasmid containing the full-length ST6Gal II cDNA(pFlag-wST6Gal-II). Briefly, 15 000 cells were seeded oneight chamber slides (LAB-TEK Nalgen Nunc Interna-tional) and cultured in standard conditions until mid-confluence. Then cells were transfected with 0.25 lg ofpurified plasmid per well in 200 lL OPTIMEM, using theLipofectamine Reagent Plus kit. After transfection, cellswere cultured for 24 h in fresh medium containing fetalbovine serum. Cells were then fixed for 30 min at 4 �C with4% paraformaldehyde and quenched 30 min with 50 mM
NH4Cl in phosphate buffered saline (NaCl/Pi). Cells werethen saturated for 30 min at 4 �C with 2% polyvinylpyr-olidone in Tris buffered saline (NaCl/Tris) and incubatedwith digoxigenin-labeled Sambucus nigra agglutinin(10 lgÆmL)1). After washing with NaCl/Tris, the cells werefurther saturated for 30 min with the blocking reagent.
S. nigra agglutinin-DIG was revealed using anti-digoxige-nin-fluorescein Fab fragments diluted 1 : 100 in NaCl/Tris,1% BSA. Laser confocal microscopy analysis was per-formed using a Zeiss-instrument (Model LSM 510).
Construction of recombinant baculovirusesand production of soluble ST6Gal I and ST3Gal III in Sf9
In order to express soluble andHis6-tagged enzymatic formsof human ST6Gal I (GenBank accession number X17247)and rat ST3Gal III (clone ST3N-1 [31]), the 5¢ end of thesegenes were modified. The cytoplasmic tail and the trans-membrane domain were deleted and the signal peptidesequence of a viral gene ecdysone-S-glycosyltransferase(EGT) was inserted [32]. For this purpose, a 247 bp PCRfragment corresponding to the hST6Gal I amino acids52–122 was amplified using a pflag/hST6Gal I plasmid,generated by PCR from HepG2 cDNA library (C. Baisezand A. Harduin-Lepers, unpublished data), as the templateand two specific primers For 6I 5¢-CGATGAATTC
GTTAACGCTCATCACCATCACCATCACGGGAAATTGGCCATGGGGT-3¢ containing a HpaI site and Back6I 5¢-CGATGGTACCGTACTTGTTCATGCTTAGG-3¢and subcloned into pUC19 for further sequencing (Euro-gentec, Belgium). This plasmid was then digested withAvrIIand BamHI and ligated to the remaining 909 bp AvrII–BamHI fragment containing the 3¢ end of the gene purifiedfrom the original construct pflag/hST6Gal I. This construc-tion named pUC 6hisST6Gal I contained an additionalHpaI site, the last codon of the EGT signal peptide sequenceand six histidine codons. The modified hST6Gal I sequencewas then excised as an 1104 bpHpaI–BamHI fragment andinserted into the HpaI–BglII sites of pUC-PSEGT. ThepUC-PSEGT plasmid was generated by inserting a 78 bpfragment encoding the signal peptide sequence of EGT gene[32]. This 78 bp fragment was obtained after the annealingof the two following synthetic oligonucleotides For EGT 5¢-GATCCGCCACCATGACCATCTTATGTTGGCTCGCTCTCCTGAGCACACTCACAGCTGTTAACGCTGACATCA-3¢ and Back EGT 5¢-GATCTGATGTCAGCGTTAACAGCTGTGAGTGTGCTCAGGAGAGCGAGCCAACATAAGATGGTCATGGTGGCG-3¢. (In orderto avoid homologous recombination between the two EGTsequences, codons were degenerate.) For rST3Gal III, a108 bp DNA fragment containing a HpaI site, the lastcodon of the EGT signal peptide sequence, six histidinecodons and 19 codons corresponding to amino-acidresidues 34–52 was reconstituted using a set of nineoverlapping synthetic oligonucleotides and four uniquerestriction sites AvrII, MunI, HindIII and SacI. The DNAfragment was subcloned in a pUC plasmid and sequenced.The reconstituted fragment was HpaI–SacI cut and clonedin HpaI–SacI sites of the pUC-PSEGT plasmid describedabove. The resulting construct was digest with AvrII–MunIto receive the 1900 bp AvrII–EcoRI fragment preparedfrom pBS SK ST3Gal III (clone ST3N-1 [31]). The plasmidpUC PS6HisST3Gal III was obtained. The full lengthmodified hST6Gal I gene was then excised after digestionwith BamHI and HindIII and inserted at the BglII–HindIIIsites of the p119 transfer vector designed for recombinationinto the p10 locus of the baculovirus giving rise to the p119-PS ST6Gal I construct [33]. The full-length modified
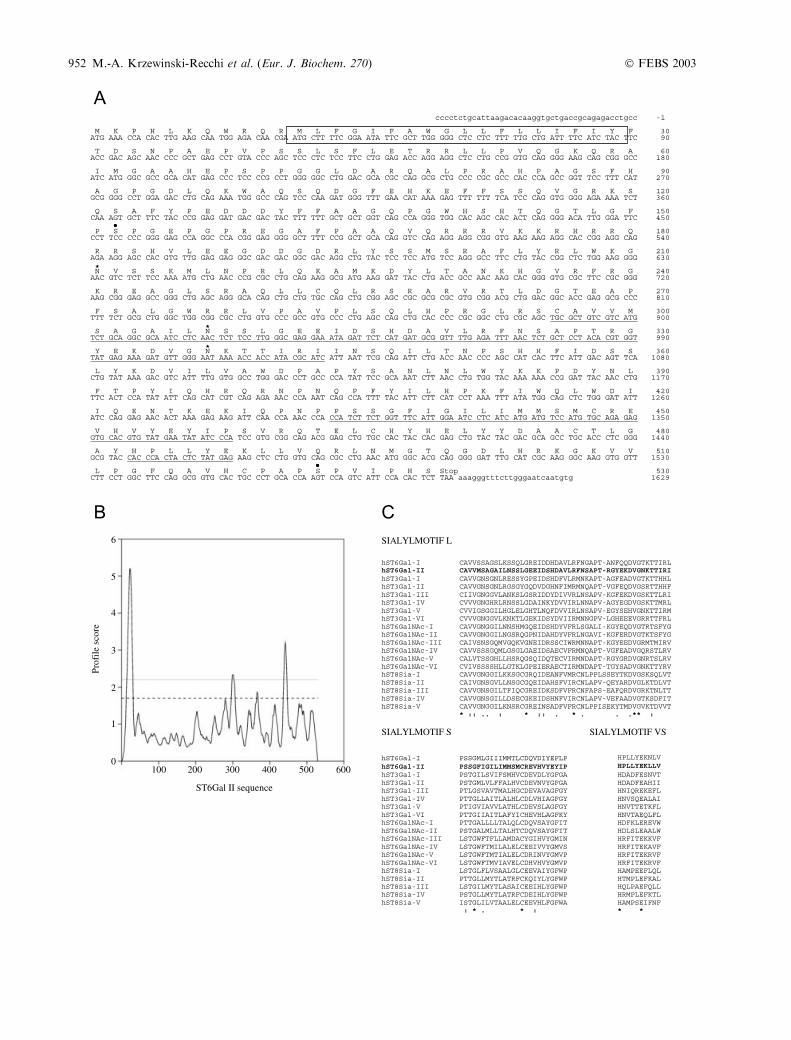
Fig. 1. Nucleotide and predicted amino acid sequence (A) and hydro-
pathy profile (B) of human ST6Gal II, and (C) comparison of the sia-
lylmotifs L, S and VS of hST6Gal II with those of previously cloned
sialyltransferases. (A) Numbering of the cDNA begins with the initi-
ation codon. The amino-acid sequence is shown in single letter code.
The putative N-terminal transmembrane domain is boxed. Putative
N-glycosylation site (N-X-S/T) are marked with asterisks (*) and
O-glycosylation sites (NetOGlyc 2.0) with back dots (d). Sialylmotifs
L, S and VS are underlined. (B) The prediction of transmembrane
region has been determined by the Dense Alignment Surface method
according to Cserzo et al. (1997) [49]. The portions (positive numbers)
above the horizontal dotted line correspond to hydrophobic regions.
(C) The sialyltransferases protein sequences were aligned using the
CLUSTALW algorithm. Amino acid identities are marked with asterisks
and dots indicate a position that is well conserved.
� FEBS 2003 Characterization of the human ST6Gal II (Eur. J. Biochem. 270) 953

rST3Gal III gene was excised with BamHI andHindIII andinserted at the BglII–HindIII site of the p119 transfer vectorgiving the p119-PS ST3Gal III construct. Sf9 cells (ATCCCRL1711) were cotransfected by lipofection [34] usingDOTAP with the transfer vectors and purified viral DNA.The recombinant baculoviruses were plaque purified andviral clones were tested for sialyltransferase activity asdescribed below.
Sialyltransferase assays
Sialyltransferase assays were performed as described previ-ously [30,35–36]. In brief, enzyme activity was measured in0.1 M cacodylate buffer pH 6.2, 10 mM MnCl2, 0.2%Triton CF-54, 50 lM CMP-[14C]Neu5Ac (1.85 KBq), withone of the acceptor substrate (2 mgÆmL)1 for glycoproteinor 1 mM for arylglycosides and oligosaccharides) and 23 lLof the enzyme source in a final volume of 50 lL. Thereactions were performed at 37 �C for 4 h. Reactionproducts were separated from CMP-[14C]Neu5Ac depend-ing on the acceptor substrate. For glycoproteins, thereaction was terminated either by precipitation and filtra-tion as previously described [35] or by SDS/PAGE. AfterWestern blotting, the radioactive products were detectedand quantified by radio-imaging using a PersonalMolecularImager FX (Bio-Rad, France). Quantification was per-formed within the linear range of standard radioactivity.For arylglycosides, the reaction was stopped with theaddition of 1 mL H2O and products were isolated byhydrophobic chromatography on C18 SepPak cartridges(Millipore Corp., Milford, MA, USA). For free oligosac-charides, the reaction mixture was heated at 100 �C for5 min, centrifuged and subjected to a paper descendingchromatography (Whatman 3) in the following solvent:pyridine/ethyl acetate/acetic acid/H2O (5 : 5 : 1 : 3, v/v/v/v)and the radioactive products were detected and quantifiedby radio-imaging. Under these conditions, the productformation from the individual acceptor substrates waslinear up to 8 h.
For kinetic analysis, incubations were performed asdescribed above using various concentrations of acceptorsubstrates: 0–500 lM of CMP-Neu5Ac, 0–500 lM ofGalb1–4GlcNAcb-O-octyl, or 0–5 mgÆmL)1 of asialo-a1-acid glycoprotein. Kinetic parameters were determined byLineweaver–Burk plots and the Km for asialo-a1-acidglycoprotein is expressed in mM relative to the 18 molterminal Gal residues per mol of human of asialo-a1-acidglycoprotein [37].
Linkage analysis by sialidase digestion
For linkage analysis, asialo-a1-acid glycoprotein sialylatedeither with the soluble ST6Gal II or, for the control, withsoluble ST6Gal I and ST3Gal III, was precipitated withethanol and air dried, dissolved in water and then digestedwith specific sialidase (sialic acid linkage analysis kit, GlycoInc., USA): NANase I (specific for a2,3-linked sialic acid,0.5 mUÆlL)1), NANase II (specific for a2,3 and a2,6-linkedsialic acid, 1 mUÆlL)1), or NANase III (specific for a2,3,a2,6 and a2,8/9-linked sialic acid, 0.5 mUÆlL)1) at 37 �C for1 h. Digested materials were then analysed by SDS/PAGEandthe radioactiveproductswereanalysedby radio-imaging.
Multiple tissue expression array and northern analysis
An EcoRI–BamHI 1.6 kb fragment of the human ST6GalII cDNA and a 1.8 kb human b-actin cDNA (Clontech)used as a positive control for Northern were labelled with[a-32P]dCTP by random priming using the Rediprime IIDNA labelling system. The human multiple tissues arraymembrane and Northern blot were probed according to themanufacturer’s instructions and analysed by radio-imaging.
Results
Identification and isolation of human ST6Gal II cDNA
Similarity searches using the tBLASTn algorithm in thehuman expressed sequence tag (EST), high throughputgenomic sequences (HTGs) and human genomic sequencesdivisions of the GenBankTM/EBI databases at the NationalCenter for Biotechnology Information allowed us toidentify nucleotide sequences with significant similaritiesto hST6Gal I (X17247 [22]). These sequences (GenBankaccession numbers AB058780, BC008660, AA385852;EST clones and AC108049, AC016994, AC005040 andNT_005429; genomic clones) were subsequently used toreconstitute a nucleotide sequence potentially encoding asialyltransferase as yet not described. Clone AB058780represented a full-length cDNA sequence already clonedfrom hippocampus [38] whereas EST clones BC008660and AA385852, found in ovary adenocarcinoma and inthyroid, respectively, represented truncated nucleotidesequences. Oligonucleotides were designed and partialcDNA sequence (nucleotides 940–1629) was obtained byRT-PCR using neuroblastoma cells NSK total RNAs astemplate (data not shown). This nucleotide sequence(GenBank accession number AJ512141) was subclonedand sequenced and found to be 100% identical to theAB058780 corresponding sequence. However, we failed toamplify the corresponding full-length open reading frameas one fragment and thus we further worked with theclone AB058780 kindly provided by T. Nagase, KazusaDNA Research Institute (Kisarazu, Chiba, Japan). Asshown in Fig. 1A, the nucleotide sequence contains anopen reading frame of 1586 bp encoding a putative 529amino-acid polypeptide with three putative N-glycosyla-tion sites and two putative O-glycosylation sites. Hydro-pathy profile analysis of the predicted protein (Fig. 1B)suggests that it has the structural organization of amembrane-bound type II glycoprotein, which is com-monly described for Golgi glycosyltransferases. Thispolypeptide shows a short cytosolic region of 11 amino-acid residues, a single hydrophobic segment of 20 amino-acid residues and a large luminal catalytic domain (498amino-acid residues). Comparison of the primary structureof this new sialyltransferase with that of the 18 othercloned human sialyltransferases indicates that there aresignificant similarities in the three sialyltransferases con-served regions named sialylmotifs L, S and VS (Fig. 1C).In particular, this protein shares with hST6Gal I acommon motif YEXXP in the sialylmotif S where theglutamic acid residue (E) is present only in these twoproteins. This analysis strongly suggests that this proteinrepresents a new sialyltransferase and since this polypep-
954 M.-A. Krzewinski-Recchi et al. (Eur. J. Biochem. 270) � FEBS 2003
tide shows 48% overall identity with the human ST6Gal I,we have named it hST6Gal II.
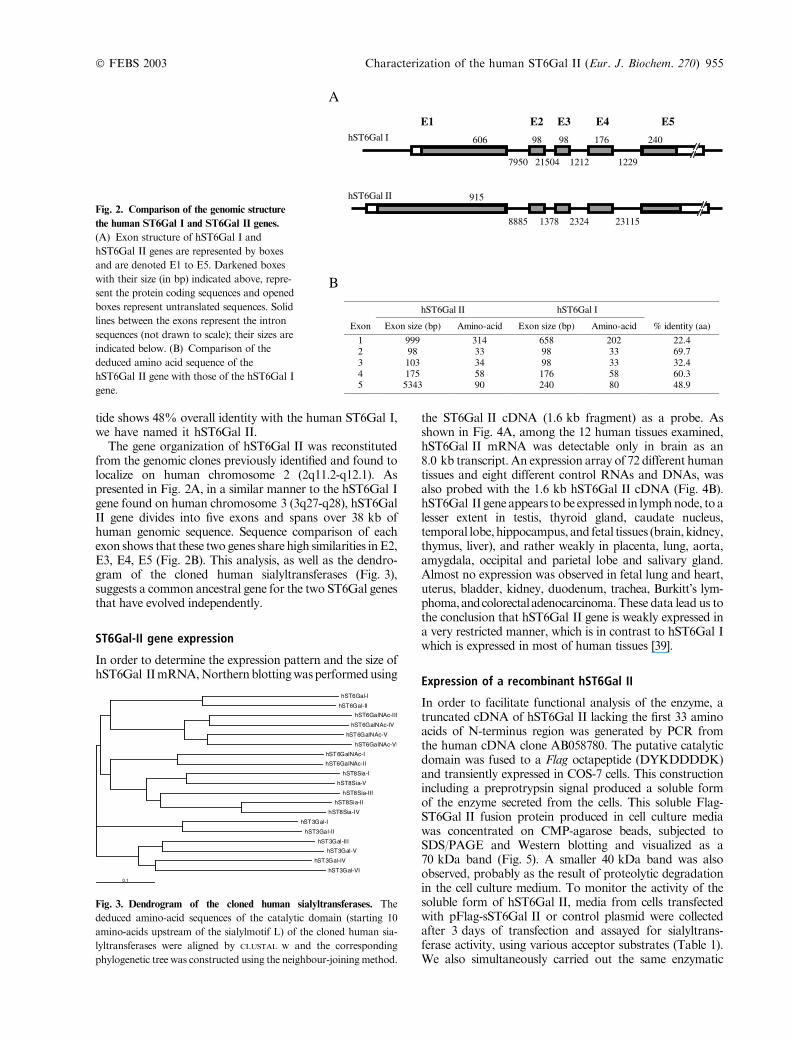
The gene organization of hST6Gal II was reconstitutedfrom the genomic clones previously identified and found tolocalize on human chromosome 2 (2q11.2-q12.1). Aspresented in Fig. 2A, in a similar manner to the hST6Gal Igene found on human chromosome 3 (3q27-q28), hST6GalII gene divides into five exons and spans over 38 kb ofhuman genomic sequence. Sequence comparison of eachexon shows that these two genes share high similarities inE2,E3, E4, E5 (Fig. 2B). This analysis, as well as the dendro-gram of the cloned human sialyltransferases (Fig. 3),suggests a common ancestral gene for the two ST6Gal genesthat have evolved independently.
ST6Gal-II gene expression
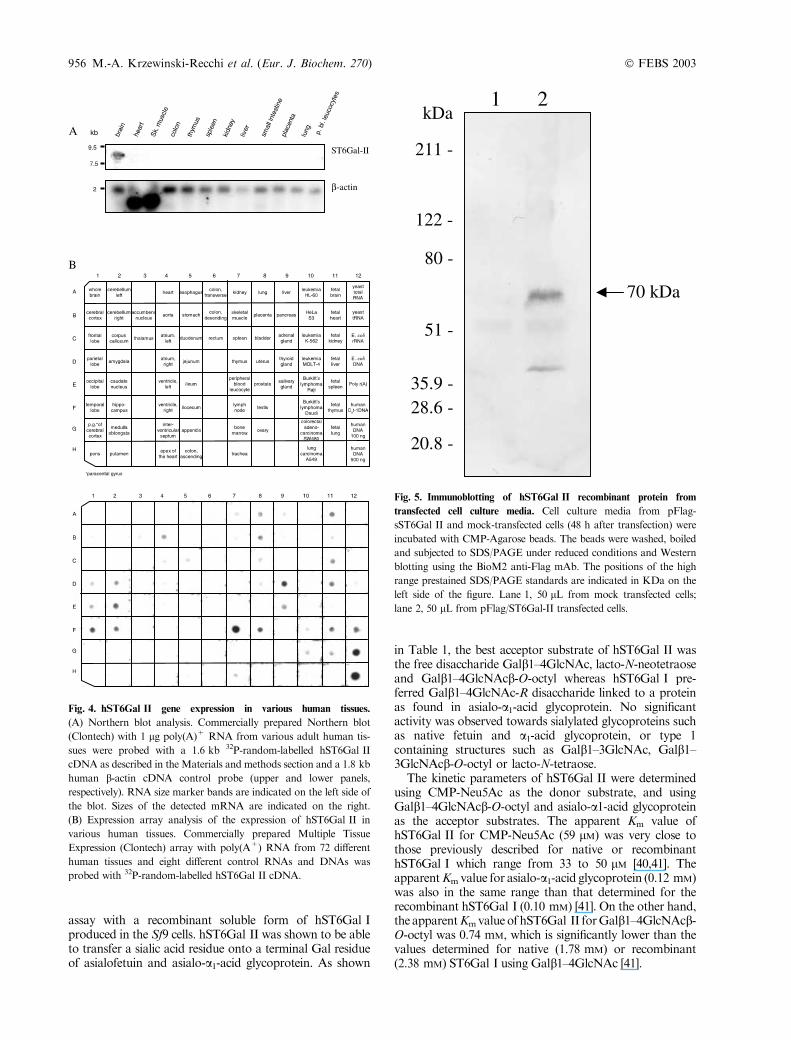
In order to determine the expression pattern and the size ofhST6Gal IImRNA,Northernblottingwasperformedusing
the ST6Gal II cDNA (1.6 kb fragment) as a probe. Asshown in Fig. 4A, among the 12 human tissues examined,hST6Gal II mRNA was detectable only in brain as an8.0 kb transcript. An expression array of 72 different humantissues and eight different control RNAs and DNAs, wasalso probed with the 1.6 kb hST6Gal II cDNA (Fig. 4B).hST6Gal II gene appears tobe expressed in lymphnode, to alesser extent in testis, thyroid gland, caudate nucleus,temporal lobe, hippocampus, and fetal tissues (brain, kidney,thymus, liver), and rather weakly in placenta, lung, aorta,amygdala, occipital and parietal lobe and salivary gland.Almost no expression was observed in fetal lung and heart,uterus, bladder, kidney, duodenum, trachea, Burkitt’s lym-phoma,andcolorectaladenocarcinoma.These data lead us tothe conclusion that hST6Gal II gene is weakly expressed ina very restricted manner, which is in contrast to hST6Gal Iwhich is expressed in most of human tissues [39].
Expression of a recombinant hST6Gal II
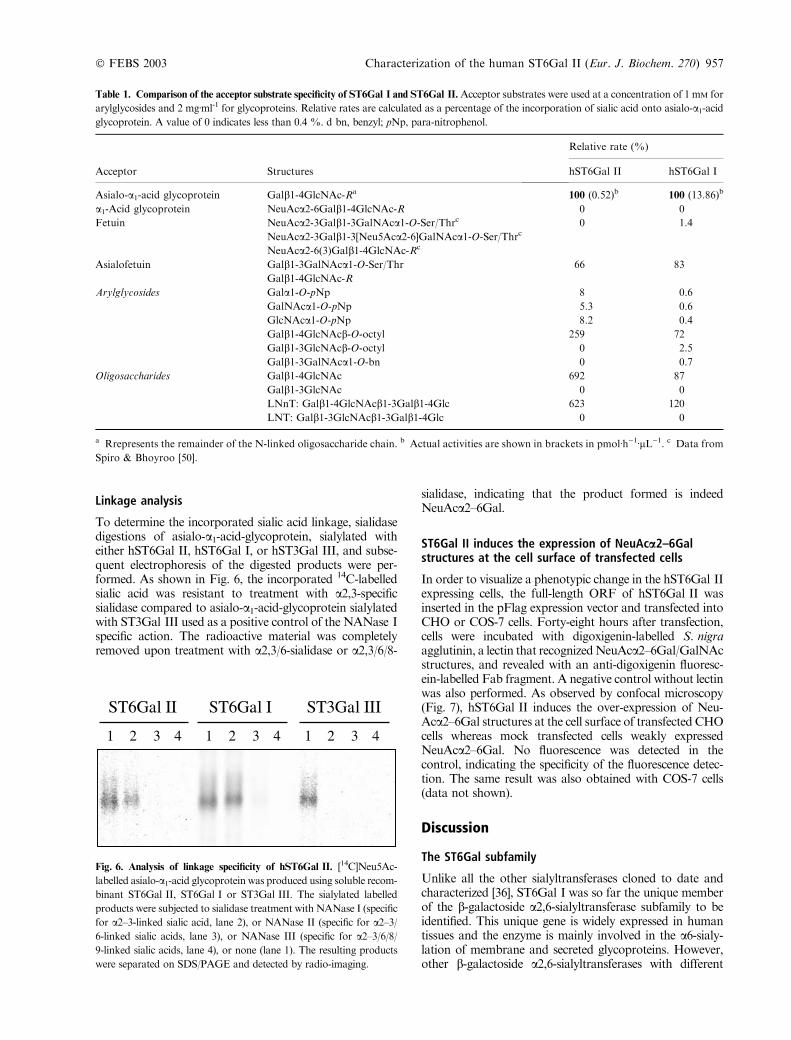
In order to facilitate functional analysis of the enzyme, atruncated cDNA of hST6Gal II lacking the first 33 aminoacids of N-terminus region was generated by PCR fromthe human cDNA clone AB058780. The putative catalyticdomain was fused to a Flag octapeptide (DYKDDDDK)and transiently expressed in COS-7 cells. This constructionincluding a preprotrypsin signal produced a soluble formof the enzyme secreted from the cells. This soluble Flag-ST6Gal II fusion protein produced in cell culture mediawas concentrated on CMP-agarose beads, subjected toSDS/PAGE and Western blotting and visualized as a70 kDa band (Fig. 5). A smaller 40 kDa band was alsoobserved, probably as the result of proteolytic degradationin the cell culture medium. To monitor the activity of thesoluble form of hST6Gal II, media from cells transfectedwith pFlag-sST6Gal II or control plasmid were collectedafter 3 days of transfection and assayed for sialyltrans-ferase activity, using various acceptor substrates (Table 1).We also simultaneously carried out the same enzymatic
Fig. 3. Dendrogram of the cloned human sialyltransferases. The
deduced amino-acid sequences of the catalytic domain (starting 10
amino-acids upstream of the sialylmotif L) of the cloned human sia-
lyltransferases were aligned by CLUSTAL W and the corresponding
phylogenetic tree was constructed using the neighbour-joiningmethod.
Fig. 2. Comparison of the genomic structure
the human ST6Gal I and ST6Gal II genes.
(A) Exon structure of hST6Gal I and
hST6Gal II genes are represented by boxes
and are denoted E1 to E5. Darkened boxes
with their size (in bp) indicated above, repre-
sent the protein coding sequences and opened
boxes represent untranslated sequences. Solid
lines between the exons represent the intron
sequences (not drawn to scale); their sizes are
indicated below. (B) Comparison of the
deduced amino acid sequence of the
hST6Gal II gene with those of the hST6Gal I
gene.
� FEBS 2003 Characterization of the human ST6Gal II (Eur. J. Biochem. 270) 955
assay with a recombinant soluble form of hST6Gal Iproduced in the Sf9 cells. hST6Gal II was shown to be ableto transfer a sialic acid residue onto a terminal Gal residueof asialofetuin and asialo-a1-acid glycoprotein. As shown
in Table 1, the best acceptor substrate of hST6Gal II wasthe free disaccharide Galb1–4GlcNAc, lacto-N-neotetraoseand Galb1–4GlcNAcb-O-octyl whereas hST6Gal I pre-ferred Galb1–4GlcNAc-R disaccharide linked to a proteinas found in asialo-a1-acid glycoprotein. No significantactivity was observed towards sialylated glycoproteins suchas native fetuin and a1-acid glycoprotein, or type 1containing structures such as Galb1–3GlcNAc, Galb1–3GlcNAcb-O-octyl or lacto-N-tetraose.
The kinetic parameters of hST6Gal II were determinedusing CMP-Neu5Ac as the donor substrate, and usingGalb1–4GlcNAcb-O-octyl and asialo-a1-acid glycoproteinas the acceptor substrates. The apparent Km value ofhST6Gal II for CMP-Neu5Ac (59 lM) was very close tothose previously described for native or recombinanthST6Gal I which range from 33 to 50 lM [40,41]. TheapparentKmvalue for asialo-a1-acid glycoprotein (0.12 mM)was also in the same range than that determined for therecombinant hST6Gal I (0.10 mM) [41]. On the other hand,the apparentKmvalueof hST6Gal II forGalb1–4GlcNAcb-O-octyl was 0.74 mM, which is significantly lower than thevalues determined for native (1.78 mM) or recombinant(2.38 mM) ST6Gal I using Galb1–4GlcNAc [41].
Fig. 4. hST6Gal II gene expression in various human tissues.
(A) Northern blot analysis. Commercially prepared Northern blot
(Clontech) with 1 lg poly(A)+ RNA from various adult human tis-
sues were probed with a 1.6 kb 32P-random-labelled hST6Gal II
cDNA as described in the Materials and methods section and a 1.8 kb
human b-actin cDNA control probe (upper and lower panels,
respectively). RNA size marker bands are indicated on the left side of
the blot. Sizes of the detected mRNA are indicated on the right.
(B) Expression array analysis of the expression of hST6Gal II in
various human tissues. Commercially prepared Multiple Tissue
Expression (Clontech) array with poly(A+) RNA from 72 different
human tissues and eight different control RNAs and DNAs was
probed with 32P-random-labelled hST6Gal II cDNA.
Fig. 5. Immunoblotting of hST6Gal II recombinant protein from
transfected cell culture media. Cell culture media from pFlag-
sST6Gal II and mock-transfected cells (48 h after transfection) were
incubated with CMP-Agarose beads. The beads were washed, boiled
and subjected to SDS/PAGE under reduced conditions and Western
blotting using the BioM2 anti-Flag mAb. The positions of the high
range prestained SDS/PAGE standards are indicated in KDa on the
left side of the figure. Lane 1, 50 lL from mock transfected cells;
lane 2, 50 lL from pFlag/ST6Gal-II transfected cells.
956 M.-A. Krzewinski-Recchi et al. (Eur. J. Biochem. 270) � FEBS 2003
Linkage analysis
To determine the incorporated sialic acid linkage, sialidasedigestions of asialo-a1-acid-glycoprotein, sialylated witheither hST6Gal II, hST6Gal I, or hST3Gal III, and subse-quent electrophoresis of the digested products were per-formed. As shown in Fig. 6, the incorporated 14C-labelledsialic acid was resistant to treatment with a2,3-specificsialidase compared to asialo-a1-acid-glycoprotein sialylatedwith ST3Gal III used as a positive control of the NANase Ispecific action. The radioactive material was completelyremoved upon treatment with a2,3/6-sialidase or a2,3/6/8-
sialidase, indicating that the product formed is indeedNeuAca2–6Gal.
ST6Gal II induces the expression of NeuAca2–6Galstructures at the cell surface of transfected cells
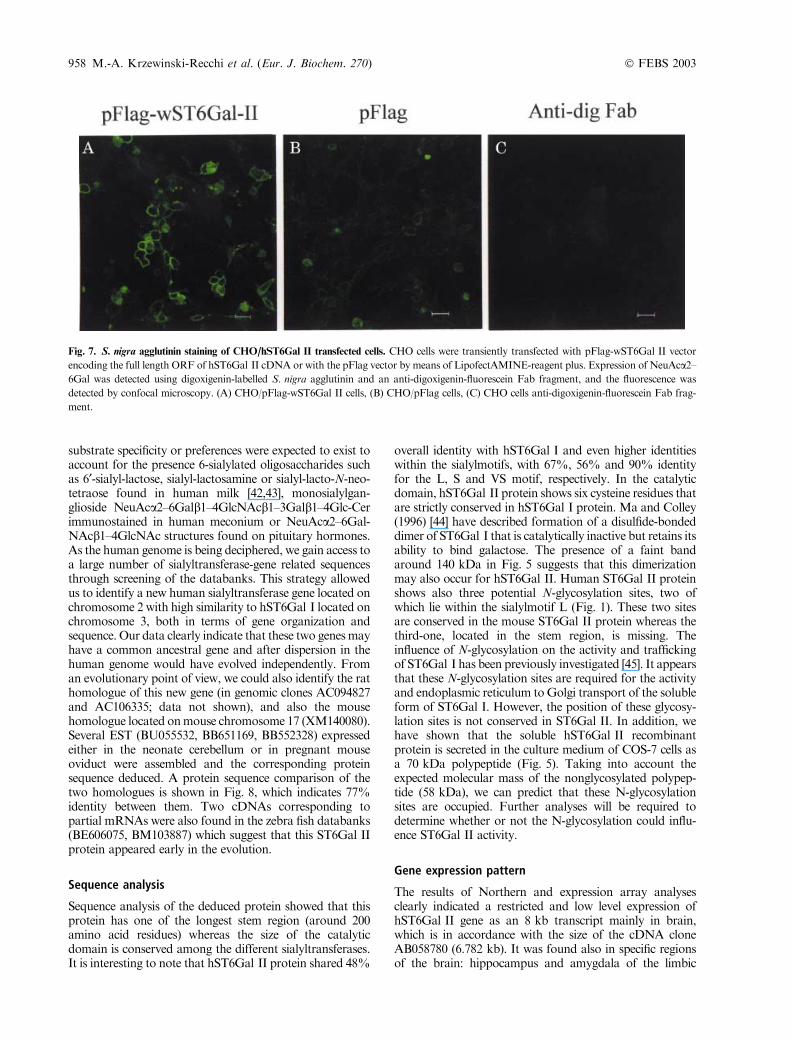
In order to visualize a phenotypic change in the hST6Gal IIexpressing cells, the full-length ORF of hST6Gal II wasinserted in the pFlag expression vector and transfected intoCHO or COS-7 cells. Forty-eight hours after transfection,cells were incubated with digoxigenin-labelled S. nigraagglutinin, a lectin that recognizedNeuAca2–6Gal/GalNAcstructures, and revealed with an anti-digoxigenin fluoresc-ein-labelled Fab fragment. A negative control without lectinwas also performed. As observed by confocal microscopy(Fig. 7), hST6Gal II induces the over-expression of Neu-Aca2–6Gal structures at the cell surface of transfected CHOcells whereas mock transfected cells weakly expressedNeuAca2–6Gal. No fluorescence was detected in thecontrol, indicating the specificity of the fluorescence detec-tion. The same result was also obtained with COS-7 cells(data not shown).
Discussion
The ST6Gal subfamily
Unlike all the other sialyltransferases cloned to date andcharacterized [36], ST6Gal I was so far the unique memberof the b-galactoside a2,6-sialyltransferase subfamily to beidentified. This unique gene is widely expressed in humantissues and the enzyme is mainly involved in the a6-sialy-lation of membrane and secreted glycoproteins. However,other b-galactoside a2,6-sialyltransferases with different
Fig. 6. Analysis of linkage specificity of hST6Gal II. [14C]Neu5Ac-
labelled asialo-a1-acid glycoprotein was produced using soluble recom-
binant ST6Gal II, ST6Gal I or ST3Gal III. The sialylated labelled
products were subjected to sialidase treatment with NANase I (specific
for a2–3-linked sialic acid, lane 2), or NANase II (specific for a2–3/6-linked sialic acids, lane 3), or NANase III (specific for a2–3/6/8/9-linked sialic acids, lane 4), or none (lane 1). The resulting products
were separated on SDS/PAGE and detected by radio-imaging.
Table 1. Comparison of the acceptor substrate specificity of ST6Gal I and ST6Gal II. Acceptor substrates were used at a concentration of 1 mM for
arylglycosides and 2 mgÆml-1 for glycoproteins. Relative rates are calculated as a percentage of the incorporation of sialic acid onto asialo-a1-acidglycoprotein. A value of 0 indicates less than 0.4 %. d bn, benzyl; pNp, para-nitrophenol.
Acceptor Structures
Relative rate (%)
hST6Gal II hST6Gal I
Asialo-a1-acid glycoprotein Galb1-4GlcNAc-Ra 100 (0.52)b 100 (13.86)b
a1-Acid glycoprotein NeuAca2-6Galb1-4GlcNAc-R 0 0
Fetuin NeuAca2-3Galb1-3GalNAca1-O-Ser/Thrc 0 1.4
NeuAca2-3Galb1-3[Neu5Aca2-6]GalNAca1-O-Ser/Thrc
NeuAca2-6(3)Galb1-4GlcNAc-Rc
Asialofetuin Galb1-3GalNAca1-O-Ser/Thr 66 83
Galb1-4GlcNAc-R
Arylglycosides Gala1-O-pNp 8 0.6
GalNAca1-O-pNp 5.3 0.6
GlcNAca1-O-pNp 8.2 0.4
Galb1-4GlcNAcb-O-octyl 259 72
Galb1-3GlcNAcb-O-octyl 0 2.5
Galb1-3GalNAca1-O-bn 0 0.7
Oligosaccharides Galb1-4GlcNAc 692 87
Galb1-3GlcNAc 0 0
LNnT: Galb1-4GlcNAcb1-3Galb1-4Glc 623 120
LNT: Galb1-3GlcNAcb1-3Galb1-4Glc 0 0
a Rrepresents the remainder of the N-linked oligosaccharide chain. b Actual activities are shown in brackets in pmolÆh)1ÆlL)1. c Data from
Spiro & Bhoyroo [50].
� FEBS 2003 Characterization of the human ST6Gal II (Eur. J. Biochem. 270) 957
substrate specificity or preferences were expected to exist toaccount for the presence 6-sialylated oligosaccharides suchas 6¢-sialyl-lactose, sialyl-lactosamine or sialyl-lacto-N-neo-tetraose found in human milk [42,43], monosialylgan-glioside NeuAca2–6Galb1–4GlcNAcb1–3Galb1–4Glc-Cerimmunostained in human meconium or NeuAca2–6Gal-NAcb1–4GlcNAc structures found on pituitary hormones.As the human genome is being deciphered, we gain access toa large number of sialyltransferase-gene related sequencesthrough screening of the databanks. This strategy allowedus to identify a new human sialyltransferase gene located onchromosome 2 with high similarity to hST6Gal I located onchromosome 3, both in terms of gene organization andsequence. Our data clearly indicate that these two genesmayhave a common ancestral gene and after dispersion in thehuman genome would have evolved independently. Froman evolutionary point of view, we could also identify the rathomologue of this new gene (in genomic clones AC094827and AC106335; data not shown), and also the mousehomologue located onmouse chromosome 17 (XM140080).Several EST (BU055532, BB651169, BB552328) expressedeither in the neonate cerebellum or in pregnant mouseoviduct were assembled and the corresponding proteinsequence deduced. A protein sequence comparison of thetwo homologues is shown in Fig. 8, which indicates 77%identity between them. Two cDNAs corresponding topartial mRNAs were also found in the zebra fish databanks(BE606075, BM103887) which suggest that this ST6Gal IIprotein appeared early in the evolution.
Sequence analysis
Sequence analysis of the deduced protein showed that thisprotein has one of the longest stem region (around 200amino acid residues) whereas the size of the catalyticdomain is conserved among the different sialyltransferases.It is interesting to note that hST6Gal II protein shared 48%
overall identity with hST6Gal I and even higher identitieswithin the sialylmotifs, with 67%, 56% and 90% identityfor the L, S and VS motif, respectively. In the catalyticdomain, hST6Gal II protein shows six cysteine residues thatare strictly conserved in hST6Gal I protein. Ma and Colley(1996) [44] have described formation of a disulfide-bondeddimer of ST6Gal I that is catalytically inactive but retains itsability to bind galactose. The presence of a faint bandaround 140 kDa in Fig. 5 suggests that this dimerizationmay also occur for hST6Gal II. Human ST6Gal II proteinshows also three potential N-glycosylation sites, two ofwhich lie within the sialylmotif L (Fig. 1). These two sitesare conserved in the mouse ST6Gal II protein whereas thethird-one, located in the stem region, is missing. Theinfluence of N-glycosylation on the activity and traffickingof ST6Gal I has been previously investigated [45]. It appearsthat these N-glycosylation sites are required for the activityand endoplasmic reticulum toGolgi transport of the solubleform of ST6Gal I. However, the position of these glycosy-lation sites is not conserved in ST6Gal II. In addition, wehave shown that the soluble hST6Gal II recombinantprotein is secreted in the culture medium of COS-7 cells asa 70 kDa polypeptide (Fig. 5). Taking into account theexpected molecular mass of the nonglycosylated polypep-tide (58 kDa), we can predict that these N-glycosylationsites are occupied. Further analyses will be required todetermine whether or not the N-glycosylation could influ-ence ST6Gal II activity.
Gene expression pattern
The results of Northern and expression array analysesclearly indicated a restricted and low level expression ofhST6Gal II gene as an 8 kb transcript mainly in brain,which is in accordance with the size of the cDNA cloneAB058780 (6.782 kb). It was found also in specific regionsof the brain: hippocampus and amygdala of the limbic
Fig. 7. S. nigra agglutinin staining of CHO/hST6Gal II transfected cells. CHO cells were transiently transfected with pFlag-wST6Gal II vector
encoding the full length ORF of hST6Gal II cDNA or with the pFlag vector by means of LipofectAMINE-reagent plus. Expression of NeuAca2–6Gal was detected using digoxigenin-labelled S. nigra agglutinin and an anti-digoxigenin-fluorescein Fab fragment, and the fluorescence was
detected by confocal microscopy. (A) CHO/pFlag-wST6Gal II cells, (B) CHO/pFlag cells, (C) CHO cells anti-digoxigenin-fluorescein Fab frag-
ment.
958 M.-A. Krzewinski-Recchi et al. (Eur. J. Biochem. 270) � FEBS 2003

system, caudate nucleus and the cerebral cortex temporallobe. Very low levels of mRNAwere detected also in lymphnode, testis, thyroid gland and fetal tissues. This low andrestricted level of expression is in agreement with the littlenumber of ST6Gal II EST in databanks. This is in contrastto the expression pattern of the ST6Gal I gene, which isabundantly expressed in almost all human tissues examined,including fetal tissues but with the notable exceptions oftestis and brain [39] where it is expressed at lower levels. Onecan postulate that expression of ST6Gal II in these twotissues could compensate for the lower expression ofST6Gal I or be responsible for the specific synthesis ofa6-sialylated glycoconjugates. We also identified twoupstream untranslated exons, exon A (119 bp) found in
the ESTBE613250 and located 42 941 bp upstream the firstcoding exon, and exon B (62 bp) found in the mRNAsequence AB058780 and located 42 058 bp upstream thefirst coding exon (data not shown). The hST6Gal II genewould thus drive the expression of at least two individualmRNAs through the use of two distinct promoters. Verypreliminary analysis of the two upstream genomic sequencesidentified in the databanks indicated the presence potentialtrans-acting factors binding sites such as SP1 and TBPbinding sites found upstream exon B. This finding wouldargue for a ubiquitous and low-level expression of exon Bcontaining transcripts. On the other hand, the presence ofCREB and SREBP binding sites upstream exon Awould indicate a specific expression of exon A containing
Fig. 8. Comparison of the putative amino-acid sequence of hST6Gal II and mST6Gal II. The two vertical dots indicate identical amino acid residues
and single dots indicate similar amino-acid residues. The underlined amino acid residues indicate the sialylmotifs L, S and VS. Putative conserved
N-glycosylation site (N-X-S/T) are marked with asterisks (*).
� FEBS 2003 Characterization of the human ST6Gal II (Eur. J. Biochem. 270) 959

transcripts in brain and fetal tissues. Future transcriptionalregulation studies will help elucidate the biological signifi-cance of each transcript.
Expression and specific activity of hST6Gal II
The acceptor substrate specificity of ST6Gal II was deter-mined using a Flag-tagged soluble form of the enzymemissing the first 33 amino-acids. The results of theenzymatic assays clearly suggested a narrow activity ofhST6Gal II in transferring sialic acid residues in the6-position of the Gal residue of the Galb1–4GlcNAcdisaccharide, exclusively. Although active with asialo-glycoproteins, hST6Gal II showed a higher activity towardsarylglycosides and above all onto free oligosaccharides. Inparticular, the Km value determined using Galb1–4Glc-NAcb-O-octyl (0.74 mM) was significantly lower than thevalue determined for native (1.78 mM) or recombinant(2.38 mM) ST6Gal I [41]. These results raised the questionof the biological significance of the presence of a secondGalb1–4GlcNAc a2,6-sialyltransferase preferentially actingonto free oligosaccharides since hST6Gal I showed alsoactivities towards these substrates (Table 1). The release offree 6-sialylated oligosaccharides in human milk is knownfor a long time (reviewed in [46]) and previous reportsdescribed elevated mammary gland sialyltransferase activityaccompanying lactation [47]. More recently, Dalziel et al.(2001) [48] described an ST6Gal I mRNA inductionmediated by recruitment of a novel 5¢ untranslated exonprobably driven by a lactogenic promoter. Our futurestudies will aim to refine ST6Gal II gene expression invarious free 6-sialylated oligosaccharides producing tissuessuch as lactating mammary gland but also to refineST6Gal II enzymatic activity towards substrate acceptorssuch as GalNAcb1–4GlcNAc or glycolipids.
In order to shed light on the biological function ofST6Gal II, in vivo activity of ST6Gal II was assessedthrough transient transfection of CHO cells and COS-7cells with the full-length ST6Gal II cDNA. Significantchanges in the sialylation pattern of cell surface glycocon-jugates were observed through the use ofS. nigra agglutinin.Western blot analysis of ST6Gal II transfected CHO cellshas also shown that only a small number of glycoproteinsexhibit a modified sialylation profile (data not shown). Thisobservation lent support to the conclusion that thisb-galactoside a2,6-sialyltransferase generated NeuAca2–6Galb1–4GlcNAc epitopes and may have very specificglycoproteins or glycolipids substrate acceptors, which ourfuture studies will aim to determine. The presence of thisnew b-galactoside a2,6-sialyltransferase ST6Gal II will shednew light onto previous studies conducted with ST6Gal I.
Acknowledgements
The authors are very thankful to Dr Claudine Auge (URACNRS 462,
Orsay, France), Drs Gerard Strecker and Frederic Chirat (UMR
CNRS-USTL 8576, Villeneuve d’Ascq, France) for the generous gift of
acceptor substrates and to Prof. J. C. Paulson for the kind gift of
ST3N-1 clone. TheUniversity of Sciences and Technologies of Lille, the
European Carbohydrate Research Network GlycoTrain, the CNRS
Reseau >G3�, and the Association pour la Recherche sur le Cancer
(grant no. 5469) have supported this work.
References
1. Varki, A. (1993) Biological roles of oligosaccharides: all of the
theories are correct. Glycobiology 3, 97–130.
2. Kelm, S. & Schauer, R. (1997) Sialic acids in molecular and cel-
lular interactions. Int. Rev. Cytol. 175, 137–240.
3. Harduin-Lepers, A., Recchi, M.A. & Delannoy, P. (1995) 1994,
the year of sialyltransferases. Glycobiology 5, 5741–5758.
4. Varki, A. (1994) Selectin ligands. Proc. Natl Acad. Sci. USA 91,
7390–7397.
5. Schauer, R. (2000) Biochemistry of sialic acid diversity. In
Carbohydrates in Chemistry and Biology Part II, Biology of the
Saccharides, V (Ernst, B., Hart, G.W. & Sinay, P., eds), Vol. 3,
pp. 227–243. Wiley-VCH, Wienhein, Germany.
6. Vallejo, V., Reyes-Leyva, J., Hernandez, J., Ramirez, H., Delan-
noy, P. & Zenteno, E. (2000) Differential expression of sialic acid
on porcine organs during the maturation process.Comp. Biochem.
Phys. B126, 415–424.
7. Hakomori, S.I. (1991) Possible functions of tumor-associated
carbohydrate antigens. Curr. Opin. Immunol. 3, 646–653.
8. Finne, J., Finne, U., Deagostini-Bazin, H. & Goridis, C. (1983)
Occurrence of a2–8 linked polysialosyl units in a neural cell
adhesion molecule. Biochem. Biophys. Res. Commun. 112, 482–
487.
9. Gyorgy, P., Jeanloz, R.W., von Nicolai, H. & Zilliken, F. (1974)
Undialyzable growth factors for Lactobacillus bifidus var. penn-
sylvanicus. Protective effect of sialic acid bound to glycoproteins
and oligosaccharides against bacterial degradation. Eur. J. Bio-
chem. 43, 29–33.
10. Prieto, P.A. & Smith, D.F. (1985) A new ganglioside in human
meconium detected by antiserum against the human milk sialy-
loligosaccharide, LS-tetrasaccharide b. Arch. Biochem. Biophys.
241, 281–289.
11. Green, E.D. & Baenziger, J.U. (1988) Asparagine-linked oligo-
saccharides on lutropin, follitropin, and thyrotropin. I. Structural
elucidation of the sulfated and sialylated oligosaccharides on
bovine, ovine, and human pituitary glycoprotein hormones. J.
Biol. Chem. 263, 25–35.
12. Harduin-Lepers, A., Vallejo-Ruiz, V., Krzewinski-Recchi, M.A.,
Samyn-Petit, B., Julien, S. & Delannoy, P. (2001) The human
sialyltransferase family. Biochimie 83, 727–737.
13. Coutinho, P.M. & Henrissat, B. (1999) Carbohydrate-active
enzymes: an integrated database approach. In Recent Advances in
Carbohydrate Bioengineering (Gilbert, H.J., Davies, G., Henrissat,
B. & Svensson, B., eds), pp. 3–12. TheRoyal Society of Chemistry,
Cambridge, UK.
14. Paulson, J.C., Weinstein, J. & Schauer, A. (1989) Tissue-specific
expression of sialyltransferases. J. Biol. Chem. 264, 10931–10934.
15. Datta, A.K. & Paulson, J.C. (1995) The sialyltransferase �sialyl-motif� participates in binding the donor substrate CMP-NeuAc.
J. Biol. Chem. 270, 1497–1500.
16. Datta, A.K., Sinha, A. & Paulson, J.C. (1998) Mutation of the
sialyltransferase S-sialylmotif alters the kinetics of the donor and
acceptor substrates. J. Biol. Chem. 273, 9608–9614.
17. Geremia, R.A., Harduin-Lepers, A. & Delannoy, P. (1997) Iden-
tification of two novel conserved amino acid residues in eukaryotic
sialyltransferases; implications for their mechanism of action.
Glycobiology 7, 5–7.
18. Paulson, J.C., Rearick, J.I. & Hill, R.L. (1977) Enzymatic prop-
erties of b-D-galactoside a2 leads to 6 sialytransferase from bovine
colostrum. J. Biol. Chem. 252, 2363–2371.
19. Nemansky,M. &Van den Eijnden, D.H. (1992) Bovine colostrum
CMP-NeuAc: Gal b (1–4) GlcNAc-R. a(2–6)-sialyltransferase is
involved in the synthesis of the terminal NeuAca (2–6) GalNAcb(1–4) GlcNAc sequence occurring on N-linked glycans of bovine
milk glycoproteins. Biochem. J. 287, 311–316.
960 M.-A. Krzewinski-Recchi et al. (Eur. J. Biochem. 270) � FEBS 2003

20. Mercier, D., Wierinckx, A., Oulmouden, A., Gallet, P.F., Palcic,
M.M., Harduin-Lepers, A., Delannoy, P., Petit, J.M., Leveziel, H.
& Julien, R. (1999)Molecular cloning, expression and exon/intron
organization of the bovine b-galactoside a2,6-sialyltransferasegene. Glycobiology 9, 851–863.
21. Weinstein, J., Lee, E.U., McEntee, K., Lai, P.H. & Paulson, J.C.
(1987) Primary structure of b-galactoside a2,6-sialyltransferase.Conversion of membrane-bound enzyme to soluble forms by
cleavage of the NH2-terminal signal anchor. J. Biol. Chem. 262,
17735–17743.
22. Grundmann, U., Nerlich, C., Rein, T. & Zettlmeissl, G. (1990)
Complete cDNA sequence encoding human b-galactoside a-2,6-sialyltransferase. Nucleic Acids Res. 18, 667.
23. Hamamoto, T., Kawasaki, M., Kurosawa, N., Nakaoka, T., Lee,
Y.C. & Tsuji, S. (1993) Two step single primer mediated poly-
merase chain reaction. Application to cloning of putative mouse,
b-galactoside a2,6-sialyltransferase cDNA. Bioorg. Med. Chem. 1,
141–145.
24. Kurosawa, N., Kawasaki, M., Hamamoto, T., Nakaoka, T., Lee,
Y.C., Arita, M. & Tsuji, S. (1994) Molecular cloning and
expression of chick embryo Gal b1,4GlcNAc a2,6-sialyltrans-ferase. Comparison with themammalian enzyme.Eur. J. Biochem.
219, 375–381.
25. Lo, N.W. & Lau, J.T. (1996) Transcription of the b-galactosidea2,6-sialyltransferase gene in B-lymphocytes is directed by a sep-
arate and distinct promoter. Glycobiology 6, 271–279.
26. Taniguchi, A., Hasegawa, Y., Higai, K. & Matsumoto, K. (2000)
Transcriptional regulation of human b-galactoside a2,6-sialyl-transferase (hST6Gal I) gene during differentiation of the HL-60
cell line. Glycobiology 10, 623–628.
27. Dalziel, M., Lemaire, S., Ewing, J., Kobayashi, L. & Lau, J.T.
(1999) Hepatic acute phase induction of murine b-galactoside a2,6sialyltransferase (ST6Gal I) is IL-6 dependent and mediated by
elevation of exon H-containing class of transcripts. Glycobiology
9, 1003–1008.
28. Lo, N.W., Dennis, J.W. & Lau, J.T. (1999) Overexpression of the
a2,6-sialyltransferase, ST6Gal I, in a low metastatic variant of a
murine lymphoblastoid cell line is associated with appearance of a
unique ST6Gal I mRNA. Biochem. Biophys. Res. Commun. 264,
619–621.
29. Dall’Olio, F., Chiricolo, M., Ceccarelli, C., Minni, F., Marrano,
D. & Santini, D. (2000) b-galactoside a2,6 sialyltransferase in
human colon cancer: contribution of multiple transcripts to
regulation of enzyme activity and reactivity with Sambucus nigra
agglutinin. Int. J. Cancer 88, 58–65.
30. Samyn-Petit, B., Krzewinski-Recchi, M.-A., Steelant, W.F.A.,
Delannoy, P. &Harduin-Lepers, A. (2000)Molecular cloning and
functional expression of human ST6GalNAc II. Molecular
expression in various human cultured cells.Biochim. Biophys. Acta
1474, 201–211.
31. Wen, D.X., Livingston, B.D., Medzihradszky, K.F., Kelm, S.,
Burlingame, A.L. & Paulson, J.C. (1992) Primary structure of
Galb1,3(4) GlcNAc a2,3-sialyltransferase determined by mass
spectrometry sequence analysis and molecular cloning. Evidence
for a protein motif in the sialyltransferase gene family. J. Biol.
Chem. 267, 21011–21019.
32. O’Reilly, D.R. & Miller, L.K. (1989) A baculovirus blocks insect
molting by producing ecdysteroid UDP-glucosyl transferase.
Science 245, 1110–1112.
33. Misse, D., Cerutti, M., Schmidt, I., Jansen, A., Devauchelle, G.,
Jansen, F. & Veas, F. (1998) Dissociation of the CD4 and CXCR4
binding properties of human immunodeficiency virus type 1 gp120
by deletion of the first putative alpha-helical conserved structure.
J. Virol. 72, 7280–7288.
34. Felgner, P.L. & Ringold, G.M. (1989) Cationic liposome-medi-
ated transfection. Nature 337, 387–388.
35. Giordanengo, V., Bannwarth, S., Laffont, C., Van Miegem, V.,
Harduin-Lepers, A., Delannoy, P. & Lefebvre, J.-C. (1997)
Cloning and expression of cDNA for a human Gal (b1–3) Gal-
NAc a2,3-sialyltransferase from the CEM T-cell line. Eur J. Bio-
chem. 247, 558–566.
36. Harduin-Lepers, A., Stokes, D.C., Steelant, W.F., Samyn-Petit,
B., Krzewinski-Recchi, M.-A., Vallejo-Ruiz, V., Zanetta, J.-P.,
Auge, C. & Delannoy, P. (2000) Cloning, expression and gene
organization of a human Neu5Aca2–3Galb1–3GalNAc a2,6-sialyltransferase: hST6GalNAc IV. Biochem. J. 352, 37–48.
37. Weinstein, J., de Souza-e., -Silva, U. & Paulson, J.C. (1982) Sia-
lylation of glycoprotein oligosaccharides N-linked to asparagine.
Enzymatic characterization of a Galb1–3 (4) GlcNAc a2,3-sial-yltransferase and a Galb1–4GlcNAc a2,6-sialyltransferase from
rat liver. J. Biol. Chem. 257, 13845–13853.
38. Nagase, T., Nakayama, M., Nakajima, D., Kikuno, R. & Ohara,
O. (2001) Prediction of the coding sequences of unidentified
human genes. The complete sequences of 100 new cDNA clones
from brainwhich code for large proteins in vitro.DNARes. 8, 85–95.
39. Kitagawa, H. & Paulson, J.C. (1994) Differential expression of five
sialyltransferase genes in human tissues. J. Biol. Chem. 269, 17872–
17878.
40. Gross, H.J., Rose, U., Krause, J.M., Paulson, J.C., Schmid, K.,
Feeney, R.E. & Brossmer, R. (1989) Transfer of synthetic sialic
acid analogues to N- andO-linked glycoprotein glycans using four
different mammalian sialyltransferases. Biochemistry 28, 7386–
7392.
41. Williams, M.A., Kitagawa, H., Datta, A.K., Paulson, J.C. &
Jamieson, J.C. (1995) Large-scale expression of recombinant sia-
lyltransferases and comparison of their kinetic properties with
native enzymes. Glycoconj. J. 12, 755–761.
42. Nakano, T., Sugawara, M. & Kawakami, H. (2001) Sialic acid in
human milk: composition and functions. Acta Paediatr. Taiwan
42, 11–17.
43. Smith, D.F. & Ginsburg, V. (1980) Antibodies against sialyl-
oligosaccharides coupled to protein. J. Biol. Chem. 255, 55–59.
44. Ma, J. & Colley, K.J. (1996) A disulfide-bonded dimer of the
Golgi b-galactoside a2,6-sialyltransferase is catalytically inactive
yet still retains the ability to bind galactose. J. Biol. Chem. 271,
7758–7766.
45. Chen, C. & Colley, K.J. (2000) Minimal structural and glycosy-
lation requirements for ST6Gal I activity and trafficking. Glyco-
biology 10, 531–583.
46. Rivero-Urgell, M. & Santamaria-Orleans, A. (2001) Oligo-
saccharides: application in infant food. Early Hum. Dev. 65, S43–
S52.
47. Bushway, A.A., Park, C.S. & Keenan, T.W. (1979) Effect of
pregnancy and lactation on glycosyltransferase activities of rat
mammary gland. Int. J. Biochem. 10, 147–154.
48. Dalziel, M., Huang, R.Y., Dall’Olio, F., Morris, J.R., Taylor-
Papadimitriou, J. & Lau, J.T. (2001) Mouse ST6Gal sialyl-
transferase gene expression during mammary gland lactation.
Glycobiology 11, 407–412.
49. Cserzo, M., Wallin, E., Simon, I., von Heijne, G. & Elofsson, A.
(1997) Prediction of transmembrane alpha-helices in prokaryotic
membrane proteins: the dense alignment surface method. Protein
Eng. 10, 673–676.
50. Spiro, R.G. & Bhoyroo, V.D. (1974) Structure of the O-glycosi-
dically linked carbohydrate units of fetuin. J. Biol. Chem. 249,
5703–5717.
� FEBS 2003 Characterization of the human ST6Gal II (Eur. J. Biochem. 270) 961





![[Second Edition,.]](https://static.fdokumen.com/doc/165x107/6322fad1887d24588e04752c/second-edition.jpg)














