
Identification and characterization of a matrix metalloproteinase ( Pta1-MMP ) expressed during...
16
Planta (2009) 230:339–354 DOI 10.1007/s00425-009-0949-8 123 ORIGINAL ARTICLE IdentiWcation and characterization of a matrix metalloproteinase (Pta1-MMP) expressed during Loblolly pine (Pinus taeda) seed development, germination completion, and early seedling establishment Supriya M. Ratnaparkhe · E. M. Ulrika Egertsdotter · Barry S. Flinn Received: 7 March 2009 / Accepted: 6 May 2009 / Published online: 24 May 2009 © Springer-Verlag 2009 Abstract Extracellular matrix (ECM) modiWcations occur during plant growth, development, and in response to environmental stimuli. Key modulators of ECM modiWca- tion in vertebrates, the extracellular matrix metalloprotein- ases (MMPs), have also been described in a few plants. Here, we report the identiWcation of Loblolly pine (Pinus taeda) Pta1-MMP and its characterization during seed development and germination. Pta1-MMP protein has the structural characteristics of other plant MMPs, the recombi- nant protein exhibits Zn 2+ -dependent protease activity, and is inhibited by EDTA and the active site-binding hydroxa- mate inhibitor GM6001. The Pta1-MMP gene is expressed in both embryo and megagametophyte, with transcript lev- els increasing in both during the period from proembryo to early cotyledonary stage, then declining during late embryogenesis and maturation drying. Protein extracts exhibited similar developmental-stage MMP-like activity. Seed germination was stimulated by GA 3 and inhibited by ABA, and the timing of germination completion was mir- rored by the presence of MMP-like protease activity in both water- and GA 3 -imbibed embryos. Pta1-MMP gene tran- script levels increased in association with radicle protrusion for both GA 3 - and water-treated embryos, in agreement with MMP-like activity. In contrast, by 11 days after imbi- bition, Pta1-MMP gene transcripts in ABA-treated embryos were at levels similar to the other treatments, although MMP-like activity was not observed. The applica- tion of GM6001 during Loblolly pine seed germination inhibited radicle protrusion. Our results suggest that MMP activity may be involved in ECM modiWcation, facilitating the cell division and expansion required during seed devel- opment, germination completion, and subsequent seedling establishment. Keywords Pinus taeda L. · Embryo development · Germination · Extracellular matrix · Proteolysis · Matrix metalloproteinase Abbreviations ECM Extracellular matrix MMP Matrix metalloproteinase GA 3 Gibberellic acid 3 ABA Abscisic acid AGP Arabinogalactan protein HRGP Hydroxyproline-rich glycoprotein TIMP Tissue inhibitor of matrix metalloproteinase DE Dry embryo DM Dry megagametophyte DAI Days after imbibition DMSO Dimethyl sulfoxide E. M. Ulrika Egertsdotter and Barry S. Flinn contributed equally to project development, support, and supervision. Electronic supplementary material The online version of this article (doi:10.1007/s00425-009-0949-8) contains supplementary material, which is available to authorized users. S. M. Ratnaparkhe · E. M. U. Egertsdotter · B. S. Flinn Department of Forestry, Virginia Polytechnic Institute and State University, Blacksburg, VA 24061-0002, USA S. M. Ratnaparkhe · B. S. Flinn (&) Institute for Sustainable and Renewable Resources, Institute for Advanced Learning and Research, 150 Slayton Avenue, Danville, VA 24540, USA e-mail: barry.X[email protected] E. M. U. Egertsdotter School of Biology, Georgia Institute of Technology, 500 Tenth Street, NW, Atlanta, GA 30332-0620, USA
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Identification and characterization of a matrix metalloproteinase ( Pta1-MMP ) expressed during...

Planta (2009) 230:339–354
DOI 10.1007/s00425-009-0949-8ORIGINAL ARTICLE
IdentiWcation and characterization of a matrix metalloproteinase (Pta1-MMP) expressed during Loblolly pine (Pinus taeda) seed development, germination completion, and early seedling establishment
Supriya M. Ratnaparkhe · E. M. Ulrika Egertsdotter · Barry S. Flinn
Received: 7 March 2009 / Accepted: 6 May 2009 / Published online: 24 May 2009© Springer-Verlag 2009
Abstract Extracellular matrix (ECM) modiWcationsoccur during plant growth, development, and in response toenvironmental stimuli. Key modulators of ECM modiWca-tion in vertebrates, the extracellular matrix metalloprotein-ases (MMPs), have also been described in a few plants.Here, we report the identiWcation of Loblolly pine (Pinustaeda) Pta1-MMP and its characterization during seeddevelopment and germination. Pta1-MMP protein has thestructural characteristics of other plant MMPs, the recombi-nant protein exhibits Zn2+-dependent protease activity, andis inhibited by EDTA and the active site-binding hydroxa-mate inhibitor GM6001. The Pta1-MMP gene is expressedin both embryo and megagametophyte, with transcript lev-els increasing in both during the period from proembryo toearly cotyledonary stage, then declining during late
embryogenesis and maturation drying. Protein extractsexhibited similar developmental-stage MMP-like activity.Seed germination was stimulated by GA3 and inhibited byABA, and the timing of germination completion was mir-rored by the presence of MMP-like protease activity in bothwater- and GA3-imbibed embryos. Pta1-MMP gene tran-script levels increased in association with radicle protrusionfor both GA3- and water-treated embryos, in agreementwith MMP-like activity. In contrast, by 11 days after imbi-bition, Pta1-MMP gene transcripts in ABA-treatedembryos were at levels similar to the other treatments,although MMP-like activity was not observed. The applica-tion of GM6001 during Loblolly pine seed germinationinhibited radicle protrusion. Our results suggest that MMPactivity may be involved in ECM modiWcation, facilitatingthe cell division and expansion required during seed devel-opment, germination completion, and subsequent seedlingestablishment.
Keywords Pinus taeda L. · Embryo development · Germination · Extracellular matrix · Proteolysis · Matrix metalloproteinase
AbbreviationsECM Extracellular matrixMMP Matrix metalloproteinaseGA3 Gibberellic acid 3ABA Abscisic acidAGP Arabinogalactan proteinHRGP Hydroxyproline-rich glycoproteinTIMP Tissue inhibitor of matrix metalloproteinaseDE Dry embryoDM Dry megagametophyteDAI Days after imbibitionDMSO Dimethyl sulfoxide
E. M. Ulrika Egertsdotter and Barry S. Flinn contributed equally to project development, support, and supervision.
Electronic supplementary material The online version of this article (doi:10.1007/s00425-009-0949-8) contains supplementary material, which is available to authorized users.
S. M. Ratnaparkhe · E. M. U. Egertsdotter · B. S. FlinnDepartment of Forestry, Virginia Polytechnic Institute and State University, Blacksburg, VA 24061-0002, USA
S. M. Ratnaparkhe · B. S. Flinn (&)Institute for Sustainable and Renewable Resources, Institute for Advanced Learning and Research, 150 Slayton Avenue, Danville, VA 24540, USAe-mail: [email protected]
E. M. U. EgertsdotterSchool of Biology, Georgia Institute of Technology, 500 Tenth Street, NW, Atlanta, GA 30332-0620, USA
123

340 Planta (2009) 230:339–354
Introduction
Conifers form an important part of the forest economy inmany parts of the world. It is believed that the introductionof clonal forestry and plantation growth of conifers willincrease productivity and allow the forestry sector to reapthe beneWts. At present, the only clonal propagation methodsuitable for the large scale production of superior conifersis through somatic embryogenesis (Sutton 2002). However,somatic embryogenesis is not attainable with every geno-type, indicating a genetic component to the ability torespond positively in vitro (Park et al. 2006). One promis-ing strategy to alleviate this blockage in recalcitrant geno-types and improve somatic embryogenesis is to identify andcharacterize the genes involved in embryo developmentand determine their relationship to the physiological eventsof successful embryogenesis.
Embryogenesis involves a number of physiological andbiochemical processes in both symplastic and apoplasticenvironments. The extracellular matrix (ECM), consistingof the cell wall and surrounding environment, plays animportant role in developmental processes and can beexpected to change during cell division and cell expansion,key components of growth and development. The ECMconsists of a wide range of molecules and can include poly-saccharides and fatty acid esters, as well as protein compo-nents such as hydroxyproline-rich glycoproteins (HRGPs),proline-rich proteins, glycine-rich proteins, and variousreceptor proteins. Many extracellular proteins are expressedcell-speciWcally and developmentally, and their importancein development has been shown (reviewed by Brownlee2002).
Microscopic studies have shown that the plant ECMchanges in appearance during various stages of embryodevelopment (Briggs 1996; Konieczny et al. 2005). Extra-cellular molecules can modulate embryo development(Egertsdotter 1996; Mo et al. 1996), and extracellular mole-cule modiWcations are believed to be involved in the gener-ation of signaling substances regulating developmentalprocesses (Egertsdotter and von Arnold 1995; Malinowskiand Filipecki 2002).
These results suggest that a study of ECM molecules andECM remodeling would enhance our understanding ofembryogenesis. Proteins represent a minor component ofthe ECM, with many (like glycine-rich proteins andHRGPs) playing a structural role. In addition to secreted,intact proteins, several examples exist in which the proteo-lytic modiWcation of extracellular proteins is required fornormal developmental processes (Taylor et al. 1997;Berger and Altmann 2000; Li et al. 2001; Vieira et al. 2001;Hamilton et al. 2003; Xia et al. 2004; Rooney et al. 2005).
Key proteolytic regulators of the mammalian ECM arethe matrix metalloproteinases (MMPs), which play signiW-
cant roles in mammalian biological processes (Sternlichtand Werb 2001; Visse and Nagase 2003). In contrast tomammalian systems, plant MMPs have been studied in rel-atively few plants (Arabidopsis thaliana [Arabidopsis],Medicago truncatula [Medicago], Glycine max [Soybean],and Cucumis sativa [Cucumber]), with little known of theirroles and mechanisms of action in plant development.MMPs are a family of calcium-dependent, zinc-containingendopeptidases (Bode et al. 1999), secreted in a latent form,which require activation before they are functional. EachMMP consists of a speciWc domain sequence with severaldomain motifs, including the signal peptide, the propeptidedomain, the catalytic domain, and the C-terminal hemo-pexin-like domain. However, several MMPs have addi-tional domains such as a transmembrane or cytoplasmicdomain (Woessner and Nagase 2000).
The Wrst report of plant MMP activity was in soybeanleaves, and termed Azocollase (Ragster and Chrispeels1979) which was later puriWed and characterized as SMEP1(Graham et al. 1991). Since then, only a few plant MMPshave been studied in Arabidopsis (Maidment et al. 1999;Golldack et al. 2002), Cucumber (Delorme et al. 2000),Medicago (Combier et al. 2007), and Soybean (Pak et al.1997; Liu et al. 2001). All of the plant MMPs reported sofar share common structural features with animal MMPs,except that they lack a hemopexin domain. It has beenspeculated that plant MMPs are involved in remodeling ofthe plant ECM in association with plant growth, develop-ment, plant–microbe interactions, and programmed celldeath (Pak et al. 1997; Maidment et al. 1999; Liu et al.2001; Delorme et al. 2000; Combier et al. 2007). Geneves-tigator analysis of publicly available microarray data hasindicated that 2 of the 5 Arabidopsis MMP genes (At1-MMP and At4-MMP) are highly expressed during seed/sil-ique development (Flinn 2008), suggesting some role forthese extracellular proteases in seed development.
Plant morphogenesis and growth occur via localized celldivisions and targeted cell enlargement, processes whichrequire numerous changes to the ECM. Therefore, embryo-genesis, germination, and seedling growth involve speciWcECM polysaccharide and protein component changes (Yeoet al. 1998; Shevell et al. 2000; Van Hengel et al. 2001;Dyachok et al. 2002; Ramírez et al. 2004; Leboeuf et al. 2005;Cannon et al. 2008). As proteins are ECM structural compo-nents and structural modiWers, as well as sensing and signal-ing conduits from the cell wall to the cytoplasm, proteolyticmodiWers of these proteins have the potential to impact ECMintegrity, function, and developmental Wdelity. Plant MMPsrepresent potential modiWers of this protein component, andsince non-plant MMPs have shown the ability to degradeplant cell wall proteins (Dow et al. 1998; Ringli et al. 2001),they may contribute to the ECM changes required for suc-cessful embryogenesis, germination, and growth.
123

Planta (2009) 230:339–354 341
Our lab is interested in the application of somaticembryogenesis for Loblolly pine clonal propagation andgenetic modiWcation. In an eVort to understand and enhancethe eYciency of the embryogenic and resulting plantletdevelopment process, we have targeted ECM modiWcation,due to its potential to modulate the cell wall formation andexpansion required for growth and development, as well asits potential involvement in embryogenic signal transduc-tion. As MMPs are known modulators of the mammalianECM, and there is some suggestive gene expression evi-dence for MMP involvement in Arabidopsis embryo devel-opment and germination (Flinn 2008), we initiated studieson Loblolly pine MMP expression during embryogenesis,germination, and early seedling establishment.
Here, we report on the identiWcation and characterizationof an MMP gene (Pta1-MMP) from Loblolly pine (Pinustaeda) during seed development, germination completion,and early seedling establishment. We found that the expres-sion of Pta1-MMP and MMP-like activity was enhancedsigniWcantly in the embryo and megagametophyte duringthe period corresponding to early to mid-stages of zygoticembryo development, as well as in the embryo during radi-cle protrusion. A recombinant protein was generated andthe proteolytic activity characterized. The activity wasfound to be inhibited by general inhibitors of MMPs. Thisis the Wrst report of a gymnosperm MMP and its involve-
ment in seed development, germination completion, andsubsequent seedling establishment.
Materials and methods
Database search
A BLAST search was performed to Wnd homology betweenavailable plant MMP sequences and the pine ESTsequences available on http://www.fungen.org. Several ESTclones were identiWed (STRR1_76_H11.b1—shoot tipcDNA library, RTCNT1_5_D08_g1—root cDNA library,STRS1_39_H03.g1—stem cDNA library), and alignedusing ClustalW (Supplemental Fig. 1). PCR primers weredesigned against various regions (Table 1; SupplementalFig. 1) with the help of the IDT PrimerQuest (http://www.idtdna.com/Scitools/Applications/Primerquest/) pro-gram and were obtained from Integrated DNA Technolo-gies (San Diego, CA).
Plant material and pre-treatments
Two sets of plant material were used in this study. In theWrst set, cones were collected from July 2008 to August2008, from the Virginia Tech Reynold’s Homestead Weld
Table 1 Primers used for isolation and characterization of Pta1-MMP
AmpliWcation use Primer Sequence Annealing temperature (°C)
PCR product size (bp)
RT-PCR PtMMP1 5� CATACCAGAAAAACTTCGGGCTCA 3� 55 507
PtMMP2 5� GGCGATGGACTCCAGGTCGA 3�
GenomeWalker Primer 1Reverse primer
5� AAGAACGTGTACAGACCACGTCCCCC 3� Gradient from 56 to 63
978
Primer 2Nested reverse primer
5� GAGGAATAATCGGAGCTCTCGGTGA 3�
GSP1 5� GGCGGAGCTCCCGTTGATGATGTCCTCGC 3� 60.3 952
GSP2 5� CCGCGGCACCATGAGCTGCGAAATCGTGTC 3�
Library screen Conserved region
Forward 5� CGGGCTCAACGTTACCGGGGTCCTGGACG 3� 65 491
Reverse 5� GGCGATGGACTCCAGGTCGATGGCT 3�
3� UTR
Forward 5� CGCGAACCAGAAAAGTCGAC 3� 55 223
Reverse 5� GGTAAAGTAAGGGGAAAGTGCAG 3�
Recombinant protein production
Forward 5� AACCCCCATATGAGTTCAGGACTGACG 3� 58 931
Reverse 5� CCCCGGATCCTCACAGAATTAATGGCA 3�
Real-time PCR Gene speciWc
Forward 5� TGACGTGGATGGCGTCCAGTATTT 3� 58 132
Reverse 5� AGAATTGGGAGCAAAGAAGCTCCA 3�
Endogenous control
Tubulin forward 5� ACAGCTGATCTCGGGCAAAGAAGA 3�
Tubulin reverse 5� TGCAGTTATCGGCTAGCTTCCTCA 3� 119
123

342 Planta (2009) 230:339–354
site. Cones were stored at 4°C until tissue dissection, whichwas done within a week of receipt. Seeds were collectedfrom the cones, with embryo and megagametophyteremoval by dissection. Embryos were classiWed accordingto their developmental stage based on morphological crite-ria (Buchholz and Stiemert 1945) and stored at ¡80°C untilrequired. Immature zygotic embryos were separated atdiVerent stages of development representing proembryos(Stage 1), globular embryos (Stage 2), early cotyledonaryembryos with small cotyledonary nubs (Stage 3), larger cot-yledonary embryos in which the cotyledons have over-grown the shoot apex (Stage 4), late cotyledonary embryosduring the maturation drying stage (Stage 5), and mature,dry seed embryos (DE). Megagametophytes were collectedat the same time from seeds at the developmental stagesdescribed earlier, as well as mature, dry seed megagameto-phytes (DM).
The second set of plant material comprised P. taedamature dry seeds (4 years old) obtained from the VirginiaDepartment of Forestry. These were soaked for 72 h indarkness in pre-treatment growth regulator solutions[gibberellic acid 3 (GA3), BA, NAA, 2,4-D or abscisicacid (ABA)] at various molar concentrations (10¡4, 10¡5,10¡6, and 10¡7), or in water alone (control). The seedswere removed from the pre-treatment solutions by Wltra-tion and transferred onto wetted Wlter paper (with wateror the appropriate growth regulator solution). Fifty seedswere placed on each plate in triplicate, in order to deter-mine the impact of each growth regulator solution oncompletion of germination. After determining the appro-priate growth regulator concentrations to use, 120 seedswere used for each optimum treatment and treated asmentioned earlier. The same number of seeds was har-vested from each treatment and at each of the time points:day 0, day 1, day 2, day 3 and when the radicle emerged,and the zygotic embryos were separated from the mega-gametophyte. The experiment was carried out threetimes.
Reverse transcription PCR
Total RNA was isolated (RNAeasy Kit—Qiagen) fromembryo and megagametophytes of mature pine seeds, vari-ous seedling tissues (cotyledons, hypocotyls, and radicles),and also from mature tree needles. Each RNA sample(2.5 �g) was converted to cDNA (SMART™ PCR cDNASynthesis Kit—Clontech) and PCR performed with theabove-mentioned cDNA samples as templates and primersdesigned against conserved regions (Table 1, SupplementalFig. 1—primers I and II) using a MyCycler (Bio-Rad) andthe following conditions: cycle 1 (1£) 94°C for 2 min;cycle 2 (35£) 94°C for 45 s, an annealing temperature gra-dient ranging from 58 to 65°C for 45 s, 72°C for 1 min; and
cycle 3 (1£) 72°C for 1 min. Products were gel-puriWed,cloned into pGEM-T Easy (Promega) and sequenced on aBeckman CEQ8800.
Cloning of a full-length seed expressed MMP
To conWrm seed-expression of the EST-derived sequence, acDNA library was prepared from mature zygotic embryos(CloneMiner cDNA Library Construction Kit—Stratagene)and screened for the presence of a MMP clone by PCRreactions using primers designed in various regions of theEST assembly, including the 3� UTR (Table 1, Supplemen-tal Fig. 1—primers III and IV). PCR conditions used were:cycle 1 (1£) 94°C for 2 min; cycle 2 (35£) 94°C for 45 s,58°C for 45 s, 72°C for 1 min; cycle 3 (1£) 72°C for 5 min.The PCR products were gel-puriWed, cloned into pGEM-TEasy and sequenced. As a full-length cDNA clone was notobtained, genomic DNA was extracted from mature needles(DNAeasy—Qiagen) and used with the GenomeWalkerUniversal Kit (Clontech). Gene-speciWc primers (GSPs)were designed (Table 1, Supplemental Fig. 1—primers Vand VI) in order to amplify the 5� end. Primary and nestedPCR reactions were performed with these GSPs and theadaptor sequence primers. The PCR conditions were as fol-lows: cycle 1 (7£) 94°C for 25 s, then 72°C for 3 min;cycle 2 (32£) 94°C for 25 s, then 67°C for 3 min; cycle 3(1£) 67°C for 7 min. The PCR products were cloned intopGEM-T Easy and sequenced as above. Following theidentiWcation of the 5� end of the MMP sequence, the com-plete full-length open reading frame was ampliWed fromgenomic DNA using primers designed to 5� UTR and 3�
UTR sequences (Table 1). The correct sized product wasobtained, digested with EcoRV (New England Biolabs) torelease the predicted fragment sizes, and the fragmentssequenced for conWrmation of identity. The genomic DNAsequence containing the complete ORF was deposited inGenbank (Accession no. FJ389668). A phylogenetic treewas generated with PAUP (Version 4.0), using the pineamino acid sequence information, as well as selected plantMMP fragments (Flinn 2008) and Human MMP-7. All ana-lyzed fragments were of similar size and spanned the con-served region from the cysteine switch to Met-turn motif.Bootstrap values were calculated as percentages.
Expression and puriWcation of Pta1-MMP
A portion of Pta1-MMP was ampliWed by PCR (Pfu poly-merase—Stratagene) from P. taeda embryo cDNA usingprimers (Table 1) designed to amplify a 303 amino acidlength region beginning 66 amino acids upstream of thecysteine switch motif region all the way to the stop codon.The primers also added a 5� BamHI site and a 3� Nde1 siteto facilitate cloning into pET-15b (Novagen) using standard
123

Planta (2009) 230:339–354 343
techniques, and the reading frame veriWed by sequencing.The plasmid was then transformed into Escherichia coliBL21(DE3) (Novagen) for expression. Expression was per-formed overnight in 200 ml LB containing 100 mg/mlampicillin and 1 mM IPTG at 37°C. The cell pellet wasground in mortar and pestle with liquid N2 and resuspendedin TALON bead 1£ equilibrium/wash buVer (Clontech)containing 6 M guanidine HCl. Insoluble material was pel-leted by centrifugation (10,000£g) at 4°C for 20 min. Thesupernatant was collected and dialyzed overnight against6 M urea. An aliquot of 20 �l from the sample was ana-lyzed by SDS-PAGE to conWrm expression of desired pro-tein. The His-tagged recombinant protein was aYnitypuriWed using TALON beads (Clontech). Refolding of thepuriWed protein was carried out by serial dialysis withdecreasing concentrations of urea (3, 1.5, and 0.3 M) in10 mM Tris–HCl (7.5), and Wnally refolded overnight bydialysis against 150 ml of 20 mM Tris–H2SO4 pH 7.5,5 mM CaSO4, 100 mM Na2SO4, 1 mM ZnSO4, 10% glyc-erol, 0.05% Brij 35, 0.02% NaN3, and 1% Protease Inhibi-tor Cocktail Mix III (Calbiochem) at 6°C with stirring,followed by aliquoting, freezing in liquid N2, and storage at¡80°C. Protein quantity was estimated by the modiWedmethod of Ghosh et al. (1988). PuriWed fractions were ana-lyzed by SDS-PAGE, such that in addition to the urea-denatured and 6°C refolded samples, 6°C refolded sampleswere also incubated overnight at 37°C to allow for com-plete autocatalytic activation to occur. For N-terminalsequencing, the Wnal refolded protein sample (o/n at 37°C)was separated by SDS-PAGE and blotted onto PVDF mem-brane (Pierce) using a 10 mM MES (pH 6) + 20% methanoltransfer buVer. Following transfer, the membrane waswashed several times with distilled water, stained (0.02%Coomassie Brilliant blue in 40% methanol, 5% acetic acid)for 30 s, destained (40% methanol, 5% acetic acid) for1 min, then given several Wnal distilled water rinses. Aftermembrane drying, the protein band was excised and sent tothe Tufts University Core Facility (http://www.tucf.com/hplc-ms-f.html) for N-terminal sequencing using an ABI494 Protein Sequencer.
Expression studies
Total RNA was extracted from pooled zygotic embryos andmegagametophytes (RNAeasy—Qiagen), and DNAse-treated. Each RNA sample (2.5 �g) was converted to cDNAusing iScript cDNA Synthesis Kit (Bio-Rad) and relativeMMP transcript abundance analyzed by real-time PCRusing primers designed against the sequence proximal tothe 3� UTR (Table 1, Supplemental Fig. 1—primers VIIand VIII) and SYBR Green detection (iScript One-StepRT-PCR Kit with SYBR Green—Bio-Rad). All PCR prod-ucts were sequenced to conWrm identity. The reactions were
carried out using a Bio-Rad iQ5 Multicolor Real TimeDetection system using the default program. Tubulin wasused as endogenous control for normalization.
Protease activity assays
Protein was extracted from the above-mentioned plantmaterials using Tris–HCl buVer (50 mM Tris–HCl, pH 7.5,50 mM NaCl, 5 mM CaCl2, 100 �M ZnCl2, 0.025% Brij35, 0.02% NaN3) and stored at ¡20°C until used. Proteinquantity was estimated by the modiWed method of Ghoshet al. (1988). Degradation of myelin basic protein (MBP)was assayed using bovine MBP (Sigma) at a Wnal concen-tration of 0.25 mg/ml in 100 mM Tris–HCl pH 7.5, 5 mMCaCl2, 0.05% Brij 35, 0.02% NaN3 and 1% Protease Inhib-itor Cocktail Mix III (Calbiochem) A total of 1.5 �g of eachprotein extract was incubated with MBP in a 15 �l reactionvolume for 20 h at 37°C, and products were analyzed bySDS-PAGE to visualize MBP degradation, using 12% sep-arating/5% stacking gels with Bio-Rad reagents and proto-cols. The gels were stained with Imperial protein stain(Pierce) for 45 min and then destained overnight. Inhibitionof protease activity was determined by addition of the gen-eral MMP inhibitor GM6001 at a concentration of 500 �M.Dimethyl sulfoxide (DMSO) was used as the solvent con-trol for GM6001 at the same level (20% v/v) as used in theGM6001 assays. Similar assays were performed to test theactivity of recombinant Pta1-MMP (rPta1-MMP) by incu-bating MBP with rPta1-MMP (Wnal concentration 128 nM)instead of total protein extract. Inhibition of protease activ-ity was determined by the addition of EDTA at a concentra-tion ranging from 100 �M to 1 mM and GM6001 at aconcentration ranging from 50 to 500 �M.
Inhibition of MMP activity in germinating seeds by imbibing them in GM6001
Mature Loblolly pine seeds were treated with various con-centrations of GM6001 (5, 50, and 500 �M). Water andDMSO (20% Wnal concentration) were used for controltreatments. The seeds were soaked in treatment solutionsfor 72 h in darkness and then placed on wet Wlter paper sat-urated with treatment solution (an equal volume of DMSOwas used for each treatment and the volume was adjustedaccording to the highest concentration of GM6001). Radi-cle protrusion from seeds was monitored daily. Seedlinglengths from water-imbibed, DMSO-imbibed, 5 �MGM6001- and 500 �M GM6001-imbibed treatments weremeasured on the 19th day. Embryos were collected from 20randomly selected seeds for each of the treatments, and pro-tein extracts prepared as described earlier. MBP degrada-tion assays were performed as described earlier using theseprotein extracts.
123

344 Planta (2009) 230:339–354
Results
IdentiWcation and cloning of a full-length Loblolly pine MMP sequence
In an attempt to identify conifer MMPs, publicly availableESTs were screened using the region spanning the cysteineswitch and catalytic domains identiWed in other plantMMPs. Several clones were identiWed in the pine EST data-base (http://www.fungen.org). The STRR1_76_H11.b1(stem cDNA library), RTCNT1_5_D08_g1 (root cDNAlibrary), and STRS1_39_H03.g1 (shoot tip cDNA library)sequences generated an assembly (Supplemental Fig. 1),which when translated displayed the two domainsdescribed earlier. To gauge the expression of this MMP invarious Loblolly pine tissues, PCR primers were designedto amplify the region between the cysteine switch and cata-lytic domain (Table 1, Supplemental Fig. 1—primers I andII). The correct fragment was observed in seed tissues(whole embryo, embryonic cotyledons, embryonic hypo-cotyls, megagametophyte), and in needles (Fig. 1). How-ever, the fragment was not observed in radicle tissue underthe conditions used. Sequencing of this fragment conWrmedit as identical to the earlier described ESTs. As the initialPCR primers were directed against conserved domains,additional primers were developed against the 3� UTRregion of the EST assembly and part of the ORF (Table 1,Supplemental Fig. 1—primers III and IV), and used toamplify the correct fragment from a whole seed cDNAlibrary, conWrming that this particular MMP was expressedin seed tissues (data not shown).
In order to isolate a full-length coding sequence, primersdesigned near the 5� end of the available sequence wereused to isolate a full-length MMP sequence from genomicDNA using GenomeWalker technology (Table 1, Supple-mental Fig. 1—primers V and VI). This allowed the identi-Wcation of several hundred additional nucleotides,
containing a putative start codon and 5� UTR. Using this 5�
UTR sequence and the EST 3� UTR sequence, primerswere designed (Table 1) which ampliWed a fragment desig-nated Pta1-MMP, containing an ORF encoding a 364amino acid protein, with no introns and the characteristicsof an MMP (Fig. 2). Analysis of the amino acid translationrevealed a putative signal peptide at the N terminus, inagreement with the postulated role as a secreted, extracellu-lar protein, as well as a characteristic cysteine switch
Fig. 1 RT-PCR products obtained after 36 cycles from Wrst strand cD-NAs of various pine tissues, using pine MMP primers (directed againstthe cysteine switch and the catalytic domain Zn2+-binding region) ortubulin primers
Fig. 2 Nucleotide sequence and amino acid translation of the openreading frame for Pta1-MMP. The predicted signal peptide (under-line), cysteine switch PRCGXXD motif (double underline), structuralZn2+/calcium-binding site (black highlights) catalytic Zn2+-bindingHEXGHXXGXXH sequence (dashed underline), and the plant-spe-ciWc DLESV/IA sequence (italics and wavy underline) are indicated.All N residues (grey highlights) represent potential glycosylation sites
123

Planta (2009) 230:339–354 345
PRCGXXD motif, a structural Zn2+/Ca2+-binding site (non-catalytic), and a catalytic domain Zn2+-binding motif (HEI/LGHXLGXGH), followed by the conserved methionineresidue of the Met turn. In addition, Pta1-MMP containedthe conserved DLESV/IA sequence of unknown function,which is unique to plants. Further software analyses of thepredicted Pta1-MMP protein suggested the presence of sixpotential N-linked glycosylation sites. Analysis of the full-length Pta1-MMP protein sequence using several trans-membrane spanning domain prediction programs yieldedconXicting results. Three programs (Localizome, http://localodom.kobic.re.kr/LocaloDom/index.htm; TMHMM2.0, http://www.cbs.dtu.dk/services/TMHMM/; and Pre-dictProtein, http://www.predictprotein.org/main.php) indi-cated no C-terminal transmembrane domain, while two(TMpred, http://www.ch.embnet.org/software/TMPRED_form.html; TopPred, http://mobyle.pasteur.fr/cgi-bin/portal.py?form=toppred) did. Hence, it is unclear if Pta1-MMP contains a C-terminal transmembrane domain. Phy-logenetic tree analysis using the region from the cysteineswitch to the catalytic domain zinc-binding domain ofseveral predicted plant MMPs indicated that Pta1-MMP
clusters nearest a predicted rice MMP (Accession no.AAK55453), in a clade also housing two grape MMPs(Accession nos. CAN66084 and CAN66782), as well asAt4-MMP and At1-MMP (Fig. 3).
Recombinant Pta1-MMP expression and protein characterization
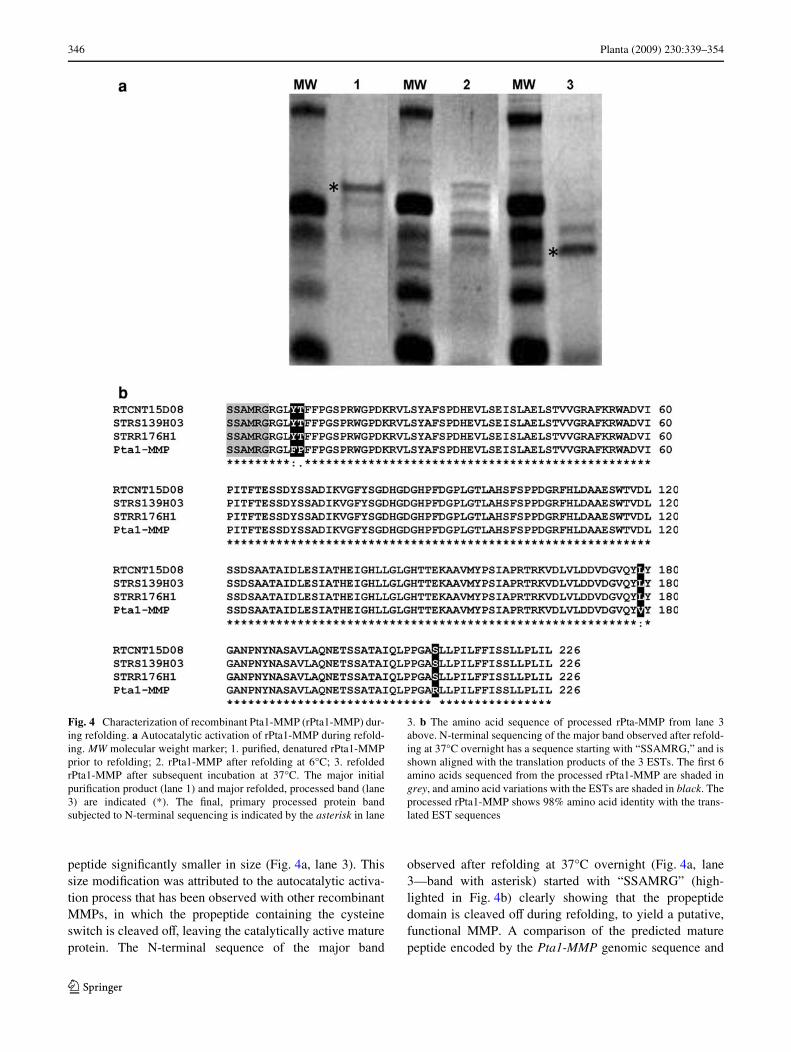
A 303 amino acid fragment, spanning the region from 66amino acids upstream of the cysteine switch to the terminalamino acid, was cloned in frame into pET-15b andexpressed as a His-tagged protein in bacteria. This regionwas chosen so as to contain the majority of the predictedprotein, with the propeptide domain and catalytic domain,but lacking the extreme N-terminal region, which includesthe predicted signal peptide. This recombinant Pta1-MMPprotein (rPta1-MMP) was isolated under denaturing condi-tions, yielding a protein which migrated at approximately43 kDa (Fig. 4a, lane 1) under the SDS-PAGE conditionsused. During refolding in the cold and subsequent incuba-tion at 37°C, several processing intermediates wereobserved (Fig. 4a, lane 2), eventually yielding a mature
Fig. 3 Phylogenetic tree of selected plant MMP protein fragmentsand human MMP-7, generated using PAUP version 4.0. All analyzedfragments are of similar size and span the conserved region from thecysteine switch to Met-turn motif (Using data from Flinn 2008). Boot-
strap values shown as percentages. Genbank Accession numbers (pro-tein) are listed if available, and if not, the Genbank Accession numbers(nucleotide) used for the translation are provided
123
346 Planta (2009) 230:339–354
peptide signiWcantly smaller in size (Fig. 4a, lane 3). Thissize modiWcation was attributed to the autocatalytic activa-tion process that has been observed with other recombinantMMPs, in which the propeptide containing the cysteineswitch is cleaved oV, leaving the catalytically active matureprotein. The N-terminal sequence of the major band
observed after refolding at 37°C overnight (Fig. 4a, lane3—band with asterisk) started with “SSAMRG” (high-lighted in Fig. 4b) clearly showing that the propeptidedomain is cleaved oV during refolding, to yield a putative,functional MMP. A comparison of the predicted maturepeptide encoded by the Pta1-MMP genomic sequence and
Fig. 4 Characterization of recombinant Pta1-MMP (rPta1-MMP) dur-ing refolding. a Autocatalytic activation of rPta1-MMP during refold-ing. MW molecular weight marker; 1. puriWed, denatured rPta1-MMPprior to refolding; 2. rPta1-MMP after refolding at 6°C; 3. refoldedrPta1-MMP after subsequent incubation at 37°C. The major initialpuriWcation product (lane 1) and major refolded, processed band (lane3) are indicated (*). The Wnal, primary processed protein bandsubjected to N-terminal sequencing is indicated by the asterisk in lane
3. b The amino acid sequence of processed rPta-MMP from lane 3above. N-terminal sequencing of the major band observed after refold-ing at 37°C overnight has a sequence starting with “SSAMRG,” and isshown aligned with the translation products of the 3 ESTs. The Wrst 6amino acids sequenced from the processed rPta1-MMP are shaded ingrey, and amino acid variations with the ESTs are shaded in black. Theprocessed rPta1-MMP shows 98% amino acid identity with the trans-lated EST sequences
123
Planta (2009) 230:339–354 347
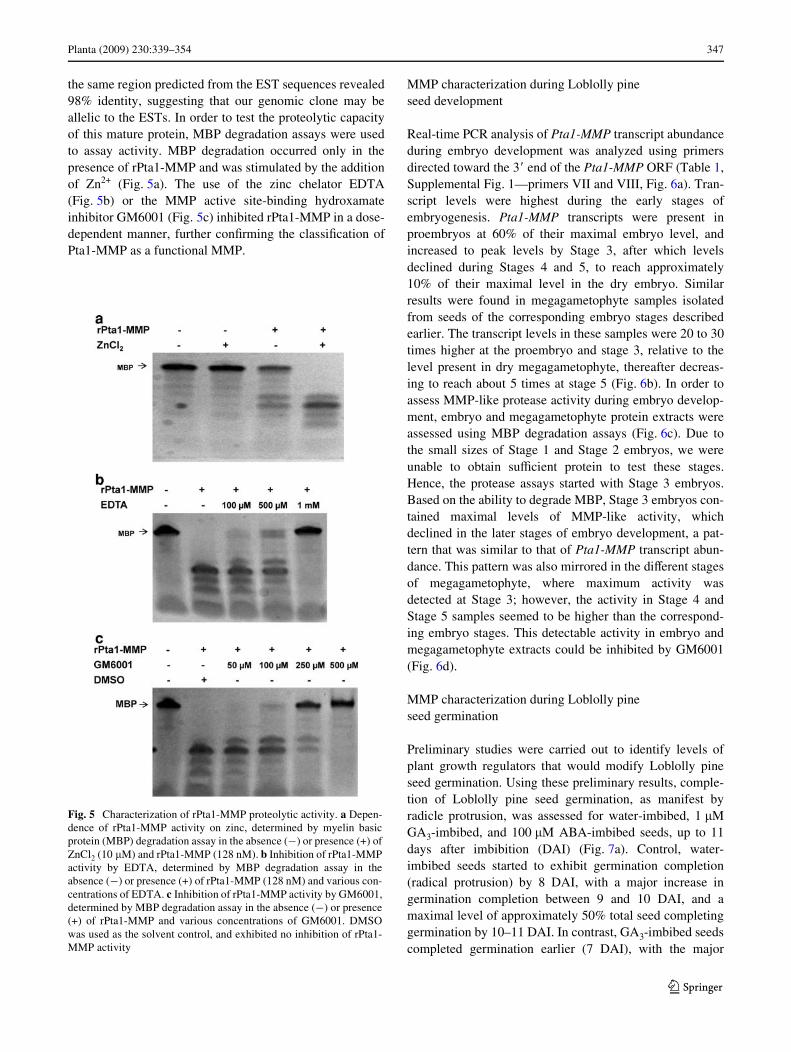
the same region predicted from the EST sequences revealed98% identity, suggesting that our genomic clone may beallelic to the ESTs. In order to test the proteolytic capacityof this mature protein, MBP degradation assays were usedto assay activity. MBP degradation occurred only in thepresence of rPta1-MMP and was stimulated by the additionof Zn2+ (Fig. 5a). The use of the zinc chelator EDTA(Fig. 5b) or the MMP active site-binding hydroxamateinhibitor GM6001 (Fig. 5c) inhibited rPta1-MMP in a dose-dependent manner, further conWrming the classiWcation ofPta1-MMP as a functional MMP.
MMP characterization during Loblolly pine seed development
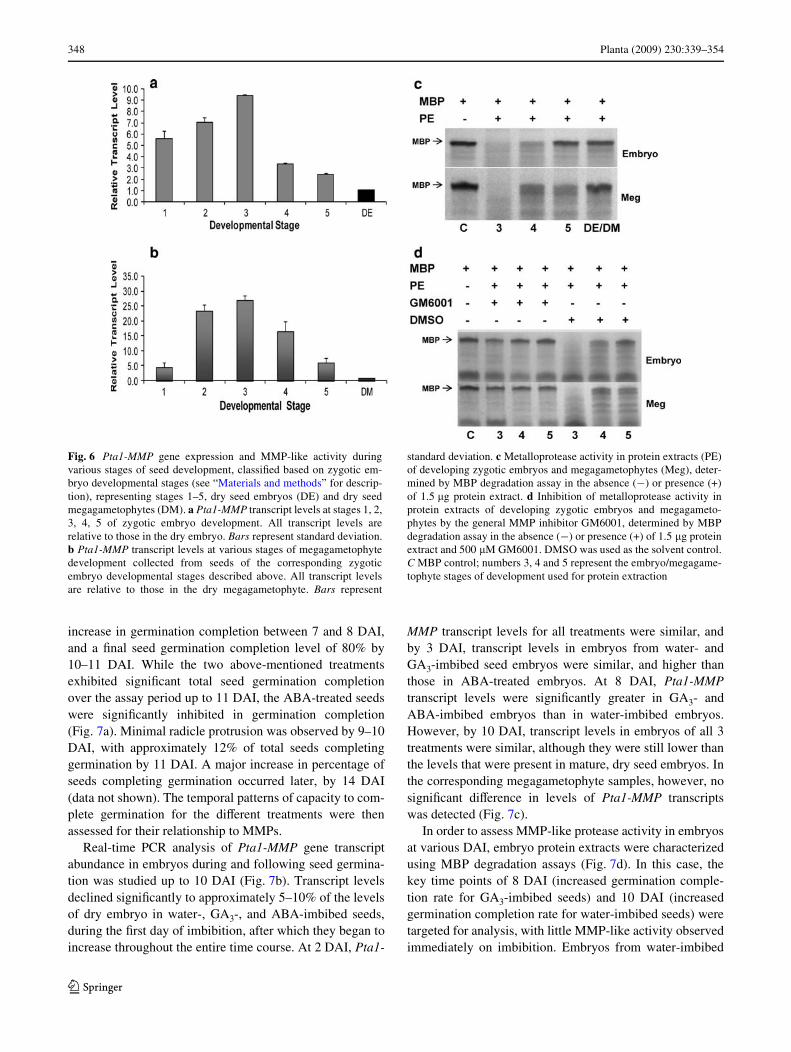
Real-time PCR analysis of Pta1-MMP transcript abundanceduring embryo development was analyzed using primersdirected toward the 3� end of the Pta1-MMP ORF (Table 1,Supplemental Fig. 1—primers VII and VIII, Fig. 6a). Tran-script levels were highest during the early stages ofembryogenesis. Pta1-MMP transcripts were present inproembryos at 60% of their maximal embryo level, andincreased to peak levels by Stage 3, after which levelsdeclined during Stages 4 and 5, to reach approximately10% of their maximal level in the dry embryo. Similarresults were found in megagametophyte samples isolatedfrom seeds of the corresponding embryo stages describedearlier. The transcript levels in these samples were 20 to 30times higher at the proembryo and stage 3, relative to thelevel present in dry megagametophyte, thereafter decreas-ing to reach about 5 times at stage 5 (Fig. 6b). In order toassess MMP-like protease activity during embryo develop-ment, embryo and megagametophyte protein extracts wereassessed using MBP degradation assays (Fig. 6c). Due tothe small sizes of Stage 1 and Stage 2 embryos, we wereunable to obtain suYcient protein to test these stages.Hence, the protease assays started with Stage 3 embryos.Based on the ability to degrade MBP, Stage 3 embryos con-tained maximal levels of MMP-like activity, whichdeclined in the later stages of embryo development, a pat-tern that was similar to that of Pta1-MMP transcript abun-dance. This pattern was also mirrored in the diVerent stagesof megagametophyte, where maximum activity wasdetected at Stage 3; however, the activity in Stage 4 andStage 5 samples seemed to be higher than the correspond-ing embryo stages. This detectable activity in embryo andmegagametophyte extracts could be inhibited by GM6001(Fig. 6d).
MMP characterization during Loblolly pine seed germination
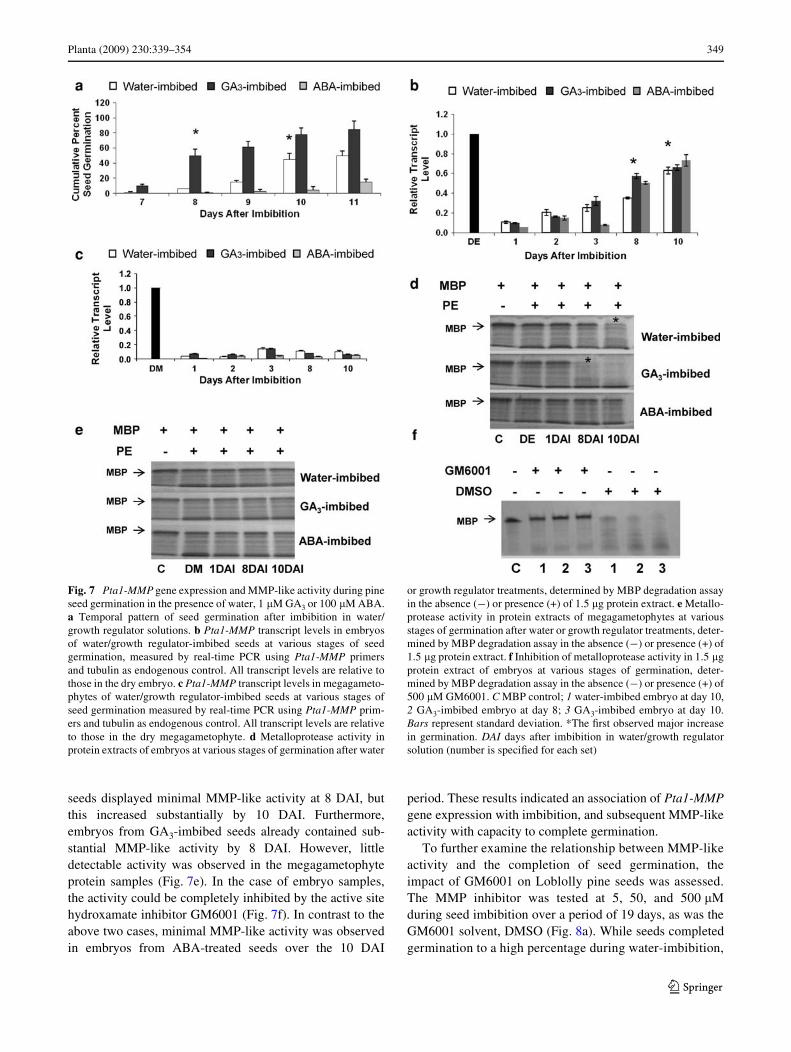
Preliminary studies were carried out to identify levels ofplant growth regulators that would modify Loblolly pineseed germination. Using these preliminary results, comple-tion of Loblolly pine seed germination, as manifest byradicle protrusion, was assessed for water-imbibed, 1 �MGA3-imbibed, and 100 �M ABA-imbibed seeds, up to 11days after imbibition (DAI) (Fig. 7a). Control, water-imbibed seeds started to exhibit germination completion(radical protrusion) by 8 DAI, with a major increase ingermination completion between 9 and 10 DAI, and amaximal level of approximately 50% total seed completinggermination by 10–11 DAI. In contrast, GA3-imbibed seedscompleted germination earlier (7 DAI), with the major
Fig. 5 Characterization of rPta1-MMP proteolytic activity. a Depen-dence of rPta1-MMP activity on zinc, determined by myelin basicprotein (MBP) degradation assay in the absence (¡) or presence (+) ofZnCl2 (10 �M) and rPta1-MMP (128 nM). b Inhibition of rPta1-MMPactivity by EDTA, determined by MBP degradation assay in theabsence (¡) or presence (+) of rPta1-MMP (128 nM) and various con-centrations of EDTA. c Inhibition of rPta1-MMP activity by GM6001,determined by MBP degradation assay in the absence (¡) or presence(+) of rPta1-MMP and various concentrations of GM6001. DMSOwas used as the solvent control, and exhibited no inhibition of rPta1-MMP activity
123
348 Planta (2009) 230:339–354
increase in germination completion between 7 and 8 DAI,and a Wnal seed germination completion level of 80% by10–11 DAI. While the two above-mentioned treatmentsexhibited signiWcant total seed germination completionover the assay period up to 11 DAI, the ABA-treated seedswere signiWcantly inhibited in germination completion(Fig. 7a). Minimal radicle protrusion was observed by 9–10DAI, with approximately 12% of total seeds completinggermination by 11 DAI. A major increase in percentage ofseeds completing germination occurred later, by 14 DAI(data not shown). The temporal patterns of capacity to com-plete germination for the diVerent treatments were thenassessed for their relationship to MMPs.
Real-time PCR analysis of Pta1-MMP gene transcriptabundance in embryos during and following seed germina-tion was studied up to 10 DAI (Fig. 7b). Transcript levelsdeclined signiWcantly to approximately 5–10% of the levelsof dry embryo in water-, GA3-, and ABA-imbibed seeds,during the Wrst day of imbibition, after which they began toincrease throughout the entire time course. At 2 DAI, Pta1-
MMP transcript levels for all treatments were similar, andby 3 DAI, transcript levels in embryos from water- andGA3-imbibed seed embryos were similar, and higher thanthose in ABA-treated embryos. At 8 DAI, Pta1-MMPtranscript levels were signiWcantly greater in GA3- andABA-imbibed embryos than in water-imbibed embryos.However, by 10 DAI, transcript levels in embryos of all 3treatments were similar, although they were still lower thanthe levels that were present in mature, dry seed embryos. Inthe corresponding megagametophyte samples, however, nosigniWcant diVerence in levels of Pta1-MMP transcriptswas detected (Fig. 7c).
In order to assess MMP-like protease activity in embryosat various DAI, embryo protein extracts were characterizedusing MBP degradation assays (Fig. 7d). In this case, thekey time points of 8 DAI (increased germination comple-tion rate for GA3-imbibed seeds) and 10 DAI (increasedgermination completion rate for water-imbibed seeds) weretargeted for analysis, with little MMP-like activity observedimmediately on imbibition. Embryos from water-imbibed
Fig. 6 Pta1-MMP gene expression and MMP-like activity duringvarious stages of seed development, classiWed based on zygotic em-bryo developmental stages (see “Materials and methods” for descrip-tion), representing stages 1–5, dry seed embryos (DE) and dry seedmegagametophytes (DM). a Pta1-MMP transcript levels at stages 1, 2,3, 4, 5 of zygotic embryo development. All transcript levels arerelative to those in the dry embryo. Bars represent standard deviation.b Pta1-MMP transcript levels at various stages of megagametophytedevelopment collected from seeds of the corresponding zygoticembryo developmental stages described above. All transcript levelsare relative to those in the dry megagametophyte. Bars represent
standard deviation. c Metalloprotease activity in protein extracts (PE)of developing zygotic embryos and megagametophytes (Meg), deter-mined by MBP degradation assay in the absence (¡) or presence (+)of 1.5 �g protein extract. d Inhibition of metalloprotease activity inprotein extracts of developing zygotic embryos and megagameto-phytes by the general MMP inhibitor GM6001, determined by MBPdegradation assay in the absence (¡) or presence (+) of 1.5 �g proteinextract and 500 �M GM6001. DMSO was used as the solvent control.C MBP control; numbers 3, 4 and 5 represent the embryo/megagame-tophyte stages of development used for protein extraction
123
Planta (2009) 230:339–354 349
seeds displayed minimal MMP-like activity at 8 DAI, butthis increased substantially by 10 DAI. Furthermore,embryos from GA3-imbibed seeds already contained sub-stantial MMP-like activity by 8 DAI. However, littledetectable activity was observed in the megagametophyteprotein samples (Fig. 7e). In the case of embryo samples,the activity could be completely inhibited by the active sitehydroxamate inhibitor GM6001 (Fig. 7f). In contrast to theabove two cases, minimal MMP-like activity was observedin embryos from ABA-treated seeds over the 10 DAI
period. These results indicated an association of Pta1-MMPgene expression with imbibition, and subsequent MMP-likeactivity with capacity to complete germination.
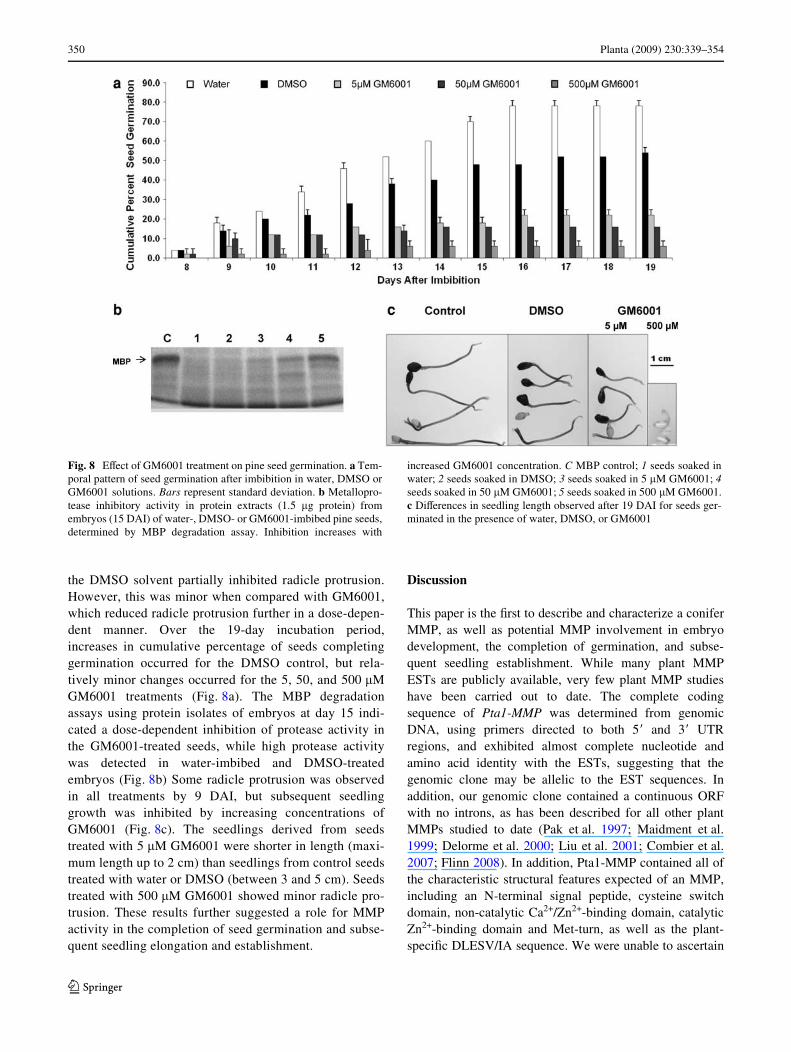
To further examine the relationship between MMP-likeactivity and the completion of seed germination, theimpact of GM6001 on Loblolly pine seeds was assessed.The MMP inhibitor was tested at 5, 50, and 500 �Mduring seed imbibition over a period of 19 days, as was theGM6001 solvent, DMSO (Fig. 8a). While seeds completedgermination to a high percentage during water-imbibition,
Fig. 7 Pta1-MMP gene expression and MMP-like activity during pineseed germination in the presence of water, 1 �M GA3 or 100 �M ABA.a Temporal pattern of seed germination after imbibition in water/growth regulator solutions. b Pta1-MMP transcript levels in embryosof water/growth regulator-imbibed seeds at various stages of seedgermination, measured by real-time PCR using Pta1-MMP primersand tubulin as endogenous control. All transcript levels are relative tothose in the dry embryo. c Pta1-MMP transcript levels in megagameto-phytes of water/growth regulator-imbibed seeds at various stages ofseed germination measured by real-time PCR using Pta1-MMP prim-ers and tubulin as endogenous control. All transcript levels are relativeto those in the dry megagametophyte. d Metalloprotease activity inprotein extracts of embryos at various stages of germination after water
or growth regulator treatments, determined by MBP degradation assayin the absence (¡) or presence (+) of 1.5 �g protein extract. e Metallo-protease activity in protein extracts of megagametophytes at variousstages of germination after water or growth regulator treatments, deter-mined by MBP degradation assay in the absence (¡) or presence (+) of1.5 �g protein extract. f Inhibition of metalloprotease activity in 1.5 �gprotein extract of embryos at various stages of germination, deter-mined by MBP degradation assay in the absence (¡) or presence (+) of500 �M GM6001. C MBP control; 1 water-imbibed embryo at day 10,2 GA3-imbibed embryo at day 8; 3 GA3-imbibed embryo at day 10.Bars represent standard deviation. *The Wrst observed major increasein germination. DAI days after imbibition in water/growth regulatorsolution (number is speciWed for each set)
123
350 Planta (2009) 230:339–354
the DMSO solvent partially inhibited radicle protrusion.However, this was minor when compared with GM6001,which reduced radicle protrusion further in a dose-depen-dent manner. Over the 19-day incubation period,increases in cumulative percentage of seeds completinggermination occurred for the DMSO control, but rela-tively minor changes occurred for the 5, 50, and 500 �MGM6001 treatments (Fig. 8a). The MBP degradationassays using protein isolates of embryos at day 15 indi-cated a dose-dependent inhibition of protease activity inthe GM6001-treated seeds, while high protease activitywas detected in water-imbibed and DMSO-treatedembryos (Fig. 8b) Some radicle protrusion was observedin all treatments by 9 DAI, but subsequent seedlinggrowth was inhibited by increasing concentrations ofGM6001 (Fig. 8c). The seedlings derived from seedstreated with 5 �M GM6001 were shorter in length (maxi-mum length up to 2 cm) than seedlings from control seedstreated with water or DMSO (between 3 and 5 cm). Seedstreated with 500 �M GM6001 showed minor radicle pro-trusion. These results further suggested a role for MMPactivity in the completion of seed germination and subse-quent seedling elongation and establishment.
Discussion
This paper is the Wrst to describe and characterize a coniferMMP, as well as potential MMP involvement in embryodevelopment, the completion of germination, and subse-quent seedling establishment. While many plant MMPESTs are publicly available, very few plant MMP studieshave been carried out to date. The complete codingsequence of Pta1-MMP was determined from genomicDNA, using primers directed to both 5� and 3� UTRregions, and exhibited almost complete nucleotide andamino acid identity with the ESTs, suggesting that thegenomic clone may be allelic to the EST sequences. Inaddition, our genomic clone contained a continuous ORFwith no introns, as has been described for all other plantMMPs studied to date (Pak et al. 1997; Maidment et al.1999; Delorme et al. 2000; Liu et al. 2001; Combier et al.2007; Flinn 2008). In addition, Pta1-MMP contained all ofthe characteristic structural features expected of an MMP,including an N-terminal signal peptide, cysteine switchdomain, non-catalytic Ca2+/Zn2+-binding domain, catalyticZn2+-binding domain and Met-turn, as well as the plant-speciWc DLESV/IA sequence. We were unable to ascertain
Fig. 8 EVect of GM6001 treatment on pine seed germination. a Tem-poral pattern of seed germination after imbibition in water, DMSO orGM6001 solutions. Bars represent standard deviation. b Metallopro-tease inhibitory activity in protein extracts (1.5 �g protein) fromembryos (15 DAI) of water-, DMSO- or GM6001-imbibed pine seeds,determined by MBP degradation assay. Inhibition increases with
increased GM6001 concentration. C MBP control; 1 seeds soaked inwater; 2 seeds soaked in DMSO; 3 seeds soaked in 5 �M GM6001; 4seeds soaked in 50 �M GM6001; 5 seeds soaked in 500 �M GM6001.c DiVerences in seedling length observed after 19 DAI for seeds ger-minated in the presence of water, DMSO, or GM6001
123

Planta (2009) 230:339–354 351
if Pta1-MMP contained a predicted C-terminal membrane-spanning domain. Various plant MMPs have been identiWedand predicted to be free, or cell surface-attached through aGPI-anchor or C-terminal membrane spanning domain(Flinn 2008). To date, there has been no demonstration ofextracellular localization or membrane attachment for anyplant MMP. Using phylogenetic tree analysis of proteinsequences, Pta1-MMP clustered with Arabidopsis At1-MMP and At4-MMP, whose genes are maximally expressedduring seed/silique development (At1-MMP, At4-MMP) andsubsequent germination (At4-MMP) (Flinn 2008), suggest-ing that Pta1-MMP is an ortholog to one of these.
The expression and puriWcation of rPta1-MMP in bacte-ria under denaturing conditions yielded a protein whichunderwent a size reduction during enzyme refolding. AllMMPs are expected to be synthesized as inactivezymogens, requiring activation through the removal of theprodomain containing the cysteine switch. Activation ofAt1-MMP has been achieved through the use of the MMPactivator APMA (Maidment et al. 1999), although MMPautocatalytic activation has also been observed (Delormeet al. 2000). In this study, rPta1-MMP underwent suspectedautocatalytic activation during refolding. Protease activityof rPta1-MMP was conWrmed by the ability to degradeMBP, as was the requirement of Zn2+ for enzyme activity.Furthermore, the MMP active site-binding inhibitorGM6001 was able to inhibit rPta1-MMP activity. Othercharacterized plant MMPs have displayed the requirementfor zinc ions, as well as inhibition by zinc chelators andhyroxamate compounds (Graham et al. 1991; Maidmentet al. 1999; Delorme et al. 2000; Liu et al. 2001). Takentogether, the sequence structural information and biochemi-cal characterization of the recombinant protein point toPta1-MMP as being a true, functional plant MMP.
The expression proWling of Pta1-MMP during Loblollypine seed embryo development indicated high levels ofexpression during the early to mid-cotyledonary stages ofembryo development, associated with cell division andexpansion, with transcript levels declining during the laterstages of embryo maturation and drying. Additionally,MMP-like activity paralleled Pta1-MMP transcript abun-dance, being high in early cotyledonary embryos anddeclining during the Wnal maturation stages and transitionto dry seed. A similar pattern was observed in the corre-sponding megagametophytes. To date, no published studieshave characterized MMP expression and activity in depthduring plant embryogenesis. Two of the 5 ArabidopsisMMP genes (At1-MMP and At4-MMP) are expressed atmaximal or near maximal levels during seed/silique devel-opment (Flinn 2008), suggesting potential involvement inembryogenesis. We have conWrmed the expression of oneLoblolly pine MMP during embryogenesis, as well asMMP-like activity. As we could not deduce what contribu-
tion Pta1-MMP made to the total activity detected in thisstudy, the embryo extract may well reXect the activity ofmore than one Pta-MMP, plus other metalloproteases. It isexpected that modiWcations to the ECM will be required ascell division and cell expansion during embryo patterningand growth occur. Changes in the appearance of the ECMhave been observed microscopically during embryo devel-opment (Briggs 1996; Konieczny et al. 2005), and variousextracellular plant proteases, such as the cysteine proteaseDEK1 (Lid et al. 2002) and the substilisin-like serine prote-ase ALE1 (Tanaka et al. 2001), are important for normalembryo and seed development. It is probable that additionalproteolytic modiWcation of the ECM through MMP activityis also required for various aspects of seed morphogenesisand growth.
In addition to seed development, MMP involvement wasalso indicated during the completion of Loblolly pine seedgermination. We observed that in mature seeds, the tran-script levels of Pta1-MMP decreased initially as the seedswere imbibed in water/hormone solution, probably owingto degradation of extant mRNA. However, the transcriptlevels and MMP-like activity increased upon completion ofgermination in embryos of water- and GA3-imbibed seedssuch that embryos from GA3-imbibed seed embryos dis-played a somewhat earlier transcript increase and a signiW-cantly earlier increase in MMP-like activity compared toembryos from water-imbibed seed. Interestingly, ABA-imbibed seed embryos, which were inhibited in germina-tion, showed similar Pta1-MMP transcript level increasesduring imbibition, but not the corresponding increase inMMP-like activity. It could be argued that since the majorrate of radicle protrusion in ABA-imbibed seeds is delayedby only 2–3 days as compared to water-imbibed seeds,there is an accumulation of MMP transcripts as radicle pro-trusion approaches. However, there is still a correspondinglack of MMP-like activity. It could be that Pta1-MMP, andother potential Pta-MMPs, may be under some form oftranslational or post-translational regulation in the ABA-imbibed seed embryos, blocking the conversion of Pta-MMP transcripts into functional protease activity. Whetherthis reXects inhibition of Pta-MMP mRNA translation intoprotein, or inhibition of Pta-MMP activation, or some othermechanism, remains to be determined. However, this isspeculative and requires further experimental analysis. Insupport of this view, it is known that various responses toABA are mediated by translational or post-translationalprotein modiWcations (Morcillo et al. 2001; Chibani et al.2006; Miura et al. 2007).
Additional evidence for MMP involvement in comple-tion of Loblolly pine seed germination was provided by theuse of the general MMP inhibitor GM6001. We observed asigniWcant decrease in the radicle protrusion from seedstreated with GM6001 at various concentrations. Interest-
123

352 Planta (2009) 230:339–354
ingly, for those seeds that were capable of completing ger-mination on GM6001, there was also a reduction inseedling lengths. A portion of the inhibition of both radicleprotrusion and seedling elongation could be attributed toDMSO, but the decrease in GM6001-treated seeds wasmuch more signiWcant than that in DMSO controls anddose-dependent, suggesting an impact based on MMPactivity inhibition in planta. While hydroxamate inhibitorshave been used for several plant MMP protein characteriza-tion studies (Maidment et al. 1999; Delorme et al. 2000),this is the Wrst report that we are aware of utilizing them inplant bioassays.
Seed germination commences with the uptake of waterby imbibition of the dry seed, followed by embryo expan-sion, culminating in the rupture of the covering layers andemergence of the radicle, which is generally considered asthe completion of the germination process (Bewley 1997).Radicle protrusion at the completion of seed germinationdepends on embryo growth driven by water uptake. Variousextracellular proteins have been shown to be involved inECM modiWcation and cell wall disassembly during andimmediately following germination, including expansins(Chen and Bradford 2000), polygalacturonases (Sitrit et al.1999), rhamnogalacturonases (Veronesi et al. 2007) andxyloglucan endotransglycosylases (Chen et al. 2002). Inaddition, rapid extracellular proteolytic events can beexpected to take place leading up to the completion of ger-mination, contributing to cell wall loosening and remodel-ing, relieving the tissue constraints and allowing radicleprotrusion. The Cardosin extracellular aspartic proteaseshave already been implicated in such a process (Pereiraet al. 2008), and this current study also implicates MMPinvolvement in the ECM modiWcation required for germi-nation.
In the absence of knowledge regarding plant MMP sub-strates, it is diYcult to pinpoint the function of Pta1-MMP,and Pta-MMP activity overall, in embryo development.Several non-plant MMPs have the ability to degrade theplant ECM. A secreted metalloprotease from the plantpathogen Xanthomonas campestris degraded the turnip pro-line-rich glycoprotein gp120 and the hydroxyproline-richprotein gpS-3, as well as extensin from potato and tomato(Dow et al. 1998). Furthermore, mammalian collagenasehas been shown to degrade the glycine-rich cell wall pro-tein, GRP1.8 (Ringli et al. 2001). In addition, plants haveseveral types of proteins that are similar to vertebrate cellsurface proteins that are known to be MMP substrates(Cauwe et al. 2007). These include plant proteins similar tovitronectin (Sanders et al. 1991), Wbronectin (Pellenc et al.2004), and integrin (Katembe et al. 1997). Interestingly,RGDS tetrapeptides, which are known to bind to integrins,can modify somatic embryo development when applied tocarrot cultures (Blackman et al. 2001), implicating integrin-
like molecules in the regulation of embryo development.Whether the above types of proteins represent plant MMPsubstrates remains to be addressed. It is tempting to specu-late that Pta1-MMP could be involved in the generation ofcertain signaling molecules by the cleavage of cell surfacepeptides, and/or it could be responsible for the cleavage ofECM proteins required for cell wall stabilization, facilitat-ing cell expansion and growth, or the easy diVusion ofextraneous molecules. A more systematic research directedtoward identiWcation of plant MMP substrates would helpelaborate the exact function of these proteases in plants.EVorts are underway in our lab to identify the natural sub-strates of these proteases.
The purpose of our study was to assess the role of MMPsin embryo development and germination, as these are twoimportant developmental processes in the plant life cycle,and an in-depth understanding of these processes may leadto the development of improved somatic embryogenesisprotocols. The importance of MMPs in mammalian biolog-ical processes, including embryogenesis, tissue remodeling,wound healing, angiogenesis and a variety of diseases(Sternlicht and Werb 2001; Visse and Nagase 2003) is wellknown. However, little is known about their role in plantdevelopment in general, and embryo development in partic-ular. The results of this study indicate a role for MMPs dur-ing the morphogenesis and expansion phases of seeddevelopment, and the expansion phase of germination andearly seedling growth.
Acknowledgments This work represents a portion of S. Ratnapar-khe’s doctoral dissertation. Many thanks to Dr. Amy Brunner andDr. Ian Clark for their comments during the course of this study, KellyMerricks for her help in gene cloning, and Dr. Jerzy Nowak for hiscomments on the article. This work was funded through the “HighValue Horticulture and Forestry – Virginia” Special Grants (ProjectNo. 2003-38891-02112) and HATCH funds (Project No. VA-135816)from the United States Department of Agriculture—CSREES, andoperating funds from the Commonwealth of Virginia to the Institutefor Advanced Learning and Research.
References
Berger D, Altmann T (2000) A substilisin-like serine protease involvedin the regulation of stomatal density and distribution in Arabidop-sis thaliana. Genes Dev 14:1119–1131. doi:10.1101/gad.14.9.1119
Bewley JD (1997) Seed germination and dormancy. Plant Cell 9:1055–1066
Blackman SA, Miedema M, Yeung EC, Staves MP (2001) EVect of thetetrapeptide RGDS on somatic embryogenesis in Daucus carota.Physiol Plant 112:567–571. doi:10.1034/j.1399-3054.2001.1120415.x
Bode W, Fernandez-Catalan C, Grams F, Gomis-Ruth FX, Nagase H,Tschesche H et al (1999) Insights into MMP-TIMP interactions.Ann N Y Acad Sci 878:73–91
Briggs CL (1996) An ultrastructural study of the embryo/endosperminterface in the developing seeds of Solanum nigrum L. zygote to
123

Planta (2009) 230:339–354 353
mid torpedo stage. Ann Bot 78:295–304. doi:10.1006/anbo.1996.0124
Brownlee C (2002) Role of extracellular matrix in cell-cell signaling:paracrine paradigms. Curr Opin Plant Biol 5:396–401.doi:10.1016/S1369-5266(02)00286-8
Buchholz JT, Stiemert ML (1945) Development of seeds and embryosin Pinus ponderosa, with special reference to seed size. Trans IllAcad Sci 38:27–50
Cannon MC, Terneus K, Hall Q, Tan L, Wang Y, Wegenhart BL, ChenL, Lamport DTA, Chen Y, Kieliszewski MJ (2008) Self-assemblyof the plant cell wall requires an extensin scaVold. Proc Natl AcadSci USA 105:2226–2231. doi:10.1073/pnas.0711980105
Cauwe B, Van den Steen PE, Opdenakker G (2007) The biochemical,biological, and pathological kaleidoscope of cell surface sub-strates processed by matrix metalloproteinases. Crit Rev BiochemMol Biol 42:113–185. doi:10.1080/10409230701340019
Chen F, Bradford KJ (2000) Expression of an expansin is associatedwith endosperm weakening during tomato seed germination.Plant Physiol 124:1265–1274
Chen F, Nonogaki H, Bradford KJ (2002) A gibberellin-regulated xy-loglucan endotransglycosylase gene is expressed in the endo-sperm cap during tomato seed germination. J Exp Bot 53:215–223
Chibani K, Ali-Rachedi S, Job C, Job D, Jullien M, Grappin P (2006)Proteomic analysis of seed dormancy in Arabidopsis. Plant Phys-iol 142:1493–1510. doi:10.1104/pp.106.087452
Combier J-P, Vernié T, de Billy F, Yahyaoui FE, Mathis R, Gamas P(2007) The MtMMPL1 early nodulin is a novel member of thematrix metalloendoproteinase family with a role in Medicagotruncatula infection by Sinorhizobium meliloti. Plant Physiol144:703–716. doi:10.1104/pp.106.092585
Delorme VGR, McCabe PF, Kim DJ, Leaver CJ (2000) A matrixmetalloproteinase gene is expressed at the boundary of senes-cence and programmed cell death in cucumber. Plant Physiol123:917–928
Dow JM, Davies HA, Daniels MJ (1998) A metalloprotease from Xan-thomonas campestris that speciWcally degrades proline/hydrox-yproline-rich glycoproteins of the plant extracellular matrix. MolPlant Microbe Interact 11:1085–1093. doi:10.1094/MPMI.1998.11.11.1085
Dyachok JV, Wiweger M, Kenne L, von Arnold S (2002) EndogenousNod-factor-like signal molecules promote early somatic embryodevelopment in Norway spruce. Plant Physiol 128:523–533.doi:10.1104/pp.010547
Egertsdotter U (1996) Regulation of somatic embryo development inNorway spruce (Picea abies). Dissertation submitted to Depart-ment of Forest Genetics, Swedish University of Agricultural Sci-ences, Uppsala, Sweden
Egertsdotter U, von Arnold S (1995) Importance of arabinogalactanproteins for the development of somatic embryos of Norwayspruce (Picea abies). Physiol Plant 93:334–345. doi:10.1111/j.1399-3054.1995.tb01412.x
Flinn B (2008) Plant extracellular matrix metalloproteinases. FunctPlant Biol 35:1183–1193. doi:10.1071/FP08182
Ghosh S, Gepstein S, Heikkila J, DumbroV EB (1988) Use of a scan-ning densitometer or an ELISA plate reader for measurement ofnanogram amounts of protein in crude extracts from biologicaltissues. Anal Biochem 169:227–233
Golldack D, Popova OV, Dietz KJ (2002) Mutation of the matrixmetalloproteinase At2-MMP inhibits growth and causes lateXowering and early senescence in Arabidopsis. J Biol Chem277:5541–5547. doi:10.1074/jbc.M106197200
Graham JS, Xiong J, Gillikin JW (1991) PuriWcation and developmen-tal analysis of a metalloendoproteinase from the leaves of Glycinemax. Plant Physiol 97:786–792
Hamilton JMU, Simpson DJ, Hyman SC, Ndimba BK, Slabas AR(2003) Ara12 subtilisin-like protease from Arabidopsis thaliana:
puriWcation, substrate speciWcity and tissue localization. BiochemJ 370:57–67. doi:10.1042/BJ20021125
Katembe WJ, Swatzell LJ, MakaroV CA, Kiss JZ (1997) Immunolocal-ization of integrin-like proteins in Arabidopsis and Chara. Phys-iol Plant 99:7–14. doi:10.1111/j.1399-3054.1997.tb03424.x
Konieczny R, Bohdanowicz J, Czaplicki AZ, Przywara L (2005)Extracellular matrix surface network during plant regeneration inwheat anther culture. Plant Cell Tissue Organ Cult 83:201–208.doi:10.1007/s11240-005-5771-9
Leboeuf E, Guillon F, Thoiron S, Lahaye M (2005) Biochemical andimmunohistochemical analysis of pectic polysaccharides I thecell walls of Arabidopsis mutant QUASIMODO 1 suspension-cultured cells: implications for cell adhesion. J Exp Bot 56:3171–3182. doi:10.1093/jxb/eri314
Li J, Lease KA, Tax FE, Walker JC (2001) BRS1, a serine carboxypep-tidase, regulates BRI1 signaling in Arabidopsis thaliana. ProcNatl Acad Sci USA 98:5916–5921. doi:10.1073/pnas.091065998
Lid SE, Gruis D, Jung R, Lorentzen JA, Ananiev E, Chamberlin M,Niu X, Meeley R, Nichols S, Olsen OA (2002) The defective ker-nel1 (dek1) gene required for aluerone cell development in endo-sperm of maize grains encodes a membrane protein of the calpaingene superfamily. Proc Natl Acad Sci USA 99:5460–5465.doi:10.1073/pnas.042098799
Liu Y, Dammann C, Bhattacharyya MK (2001) The matrix metallo-proteinase gene GmMMP2 is activated in response to pathogenicinfections in soybean. Plant Physiol 127:1788–1797.doi:10.1104/pp.127.4.1788
Maidment JM, Moore D, Murphy GP, Murphy G, Clark IM (1999)Matrix metalloproteinase homologues from Arabidopsis thali-ana—expression and activity. J Biol Chem 274:34706–34710.doi:10.1074/jbc.274.49.34706
Malinowski R, Filipecki M (2002) The role of cell wall in plantembryogenesis. Cell Mol Biol Lett 7:1137–1151
Miura K, Jin JB, Hasegawa PM (2007) Sumoylation, a post-transla-tional regulatory process in plants. Curr Opin Plant Biol 10:495–502. doi:10.1016/j.pbi.2007.07.002
Mo LH, Egertsdotter U, von Arnold S (1996) Secretion of speciWcextracellular proteins by somatic embryos of Picea abies isdependent on embryo morphology. Ann Bot 77:143–152.doi:10.1006/anbo.1996.0016
Morcillo F, Hartmann C, Duval Y, Tregear JW (2001) Regulation of7S globulin gene expression in zygotic and somatic embryos ofoil palm. Physiol Plant 112:233–243. doi:10.1034/j.1399-3054.2001.1120212.x
Pak JH, Liu CY, Huangpu J, Graham JS (1997) Construction and char-acterization of the soybean leaf metalloproteinase cDNA. FEBSLett 404:283–288. doi:10.1016/S0014-5793(97)00141-5
Park YS, Lelu-Walter MA, Harvengt L, Trontin JF, MacEacheron I,Klimaszewska K, Bonga JM (2006) Initiation of somatic embryo-genesis in Pinus banksiana, P. strobus, P. pinaster, and P. sylves-tris at three laboratories in Canada and France. Plant Cell TissueOrgan Cult 86:87–101. doi:10.1007/s11240-006-9101-7
Pellenc D, Schmitt E, Gallet O (2004) PuriWcation of a plant cell wallWbronectin-like adhesion protein involved in plant response to saltstress. Protein Expr Purif 34:208–214. doi:10.1016/j.pep.2003.11.011
Pereira CS, Da Costa DS, Pereira S, De Moura Nogueira F, Albuquer-que PM, Teixeira J, Faro C, Pissarra J (2008) Cardosins in post-embryonic development of cardoon: towards an elucidation of thebiological function of plant aspartic proteinases. Protoplasma232:203–213. doi:10.1007/s00709-008-0288-9
Ragster LV, Chrispeels MJ (1979) Azocoll-digesting proteinases insoybean leaves: characteristics and changes during leaf matura-tion and senescence. Plant Physiol 64:857–862
Ramírez C, Testillano PS, Pintos B, Moreno-Risueño MA, Bueno MA,Risueño MC (2004) Changes in pectins and MAPKs related to
123

354 Planta (2009) 230:339–354
cell development during early microspore embryogenesis inQuercus suber L. Eur J Cell Biol 83:213–225. doi:10.1078/0171-9335-00368
Ringli C, Keller B, Ryser U (2001) Glycine-rich proteins as structuralcomponents of plant cell walls. Cell Mol Life Sci 58:1430–1441.doi:10.1007/PL00000786
Rooney HCE, Van’t Klooster JW, Van Der Hoorn RAL, JoostenMHAJ, Jones JDG, De Wit PJGM (2005) Cladosporium Avr2inhibits tomato Rcr3 protease required for Cf-2-dependent diseaseresistance. Science 308:1783–1786. doi:10.1126/science.1111404
Sanders LC, Wang C-S, Walling LL, Lord EM (1991) A homolog ofthe substrate adhesion molecule vitronectin occurs in four speciesof Xowering plants. Plant Cell 3:629–635
Shevell DE, Kunkel T, Chua N-H (2000) Cell wall alterations in theArabidopsis emb30 mutant. Plant Cell 12:2047–2059
Sitrit Y, HadWeld KA, Bennett AB, Bradford KJ, Downie AB (1999)Expression of a polygalacturonase associated with tomato seed ger-mination. Plant Physiol 121:419–428. doi:10.1104/pp.121.2.419
Sternlicht MD, Werb Z (2001) How matrix metalloproteinases regu-late cell behavior. Annu Rev Cell Dev Biol 17:463–516.doi:10.1146/annurev.cellbio.17.1.463
Sutton B (2002) Commercial delivery of genetic improvement to coni-fer plantations using somatic embryogenesis. Ann For Sci59:657–661. doi:10.1051/forest:2002052
Tanaka H, Onouchi H, Kondo M, Hara-Nishimura I, Nishimura M,Machida C, Machida Y (2001) A subtilisin-like serine protease isrequired for epidermal surface formation in Arabidopsis embryosand juvenile plants. Development 128:4681–4689
Taylor AA, Horsch A, Rzepczyk A, Hasenkampf CA, Riggs CD (1997)Maturation and secretion of a serine proteinase is associated with
events of late microsporogenesis. Plant J 12:1261–1271.doi:10.1046/j.1365-313x.1997.12061261.x
Van Hengel AJ, Tadesse Z, Immerzeel P, Schols H, van Kammen A,de Vries SC (2001) N-acetylglucosamine and glucosamine-con-taining arabinogalactan proteins control somatic embryogenesis.Plant Physiol 125:1880–1890
Veronesi C, Bonnin E, Calvez S, Thalouarn P, Simier P (2007) Activ-ity of secreted cell wall-modifying enzymes and expression ofperoxidase-encoding gene following germination of Orobancheramosa. Biol Plantarum 51:391–394. doi:10.1007/s10535-007-0084-y
Vieira M, Pissarra J, Veríssimo P, Castanheira P, Costa Y, Pires E,Faro C (2001) Molecular cloning and characterization of cDNAencoding cardosin B, an aspartic proteinase accumulating extra-cellularly in the transmitting tissue of Cynara cardunculus L.Plant Mol Biol 45:529–539. doi:10.1023/A:1010675015318
Visse R, Nagase H (2003) Matrix metalloproteinases and tissue inhib-itors of metalloproteinases: structure, function, and biochemistry.Circ Res 92:827–839. doi:10.1161/01.RES.0000070112.80711.3D
Woessner FJ, Nagase H (2000) Matrix metalloproteinases and TIMPs.Oxford University Press, Oxford
Xia Y, Suzuki H, Borevitz J, Blount J, Guo Z, Patel K, Sixon RA,Lamb C (2004) An extracellular aspartic protease functions inArabidopsis disease resistance signaling. EMBO J 23:980–988.doi:10.1038/sj.emboj.7600086
Yeo U-D, Kohmura H, Nakagawa N, Sakurai N (1998) Quantitativeand qualitative changes of cell wall polysaccharides duringsomatic embryogenesis and plantlet development of Asparagus(Asparagus oYcinalis L.). Plant Cell Physiol 39:607–614
123




















